
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 17 June 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.916701
This article is part of the Research TopicMetabolic Conditions of Chronic Inflammatory DiseasesView all 5 articles
 Lisa Schielke*
Lisa Schielke* Nick Zimmermann
Nick Zimmermann Sarah HobelsbergerJulian SteiningerAnne StrunkKristin BlauJessica HernandezStephan KünzelRobert ZiegenbalgSarah RösingStefan BeissertSusanne Abraham
Sarah HobelsbergerJulian SteiningerAnne StrunkKristin BlauJessica HernandezStephan KünzelRobert ZiegenbalgSarah RösingStefan BeissertSusanne Abraham Claudia Günther*
Claudia Günther*Psoriasis is frequently associated with the metabolic syndrome and occurs more often in obese individuals. In order to understand innate immune mechanisms mediating this inflammatory pattern we investigated expression of the chemokine and lipid scavenger receptor CXCL16 in patients with psoriasis and associated comorbidities. CXCL16 expression was enhanced on all monocyte subsets in psoriatic patients compared with healthy controls and positively correlated with psoriasis activity and severity index, body mass index and the risk for cardiovascular disease indicated by PROCAM score. The intensity of CXCL16 expression on monocytes further correlated with their capability to phagocytose oxidized LDL indicating the possibility to transform into foam cells in atherosclerotic plaques. Patients with psoriasis and atherosclerosis or obesity displayed elevated numbers of innate lymphoid cells in blood with specific increase of the IFN-γ or IL-17 producing ILC1 and ILC3 subpopulations. The expression of the CXCL16 receptor, CXCR6, was increased in ILCs and co-expressed with CCR6 but not CCR7 indicating their migratory potential to psoriatic skin or adipose tissue that is characterized by strong CXCL16 and CCL20 expression. This hypothesis was supported by the finding that the percentage of CXCR6 expressing ILCs was alleviated in blood of psoriatic patients. Together these data link a strong expression of CXCL16 to metabolic syndrome in psoriasis and indicate a possible link to ILC activation and tissue distribution in obese psoriatic patients. These data contribute to the understanding of the complex interaction of innate immunity and metabolic state in psoriasis.
Psoriasis vulgaris is a chronic inflammatory disease affecting approximately 2-3% of the population with pathognomonic chronic scaling erythematous plaques at the skin (1). However, psoriasis does not only involve the skin but is now regarded as systemic disease because psoriasis patients have an increased prevalence of metabolic syndrome (2–5). In 20% to 50% psoriasis is associated with hyperlipidemia, obesity, hypertension or peripheral insulin resistance (6–8) causing an elevated cardiovascular risk (CVR) and premature mortality (4, 9–11). Obesity has also been described as an independent risk factor for the development of psoriasis (12). Consequently, reasons for the association between metabolic disorders and psoriasis vulgaris are currently explored. In particular, the innate immune system is suggested to promote metabolic disorders and initiate cutaneous manifestations of psoriasis vulgaris. Innate lymphoid cells (ILCs) represent an important component of the innate immune system. They arise from a common lymphoid progenitor and are located in mucosal barriers and several tissues, including the intestine, lung, skin and adipose tissue (13–15). ILCs are divided into three different subsets – ILC1, ILC2, and ILC3 – according to their dependence on distinct lineage-determining transcription factors and cytokine secretion profiles (16). Group 1 ILC include the natural killer cells (NK) and the non-killer-cell ILC1. They are defined by their capability to produce amounts of IFN-γ but also TNF-α, IL-1β and IL-6 (17, 18). Group 2 ILCs are able to produce TH2 cell associated cytokines, including IL-4, IL-5, IL-9 and IL-13 (19). The Group 3 ILC are competent producers of the TH17 cell-associate cytokines IL-17 and IL-22 and have been described in psoriatic skin (15, 20, 21). Circulating and adipose tissue resident ILC1 have been more frequently detected in obese individuals resulting in aggravating morbidities like insulin resistance and metabolic disorders (22–24). In addition, Pantelyushin et al. demonstrated that mice lacking NK cells and ILCs did not develop psoriasiform plaques after topically administered imiquimod in contrast to normal mice, indicating a substantial impact of ILCs in the skin inflammation of psoriasis vulgaris (25). The role of human ILCs in obese individuals concerning the initiation and aggravation of psoriasis vulgaris is not well understood.
Mobilization from bone marrow and distribution of ILCs in different organs is orchestrated by chemokines. Innate lymphoid cell precursors can be recruited from bone marrow due to expression of CXCR6. The corresponding chemokine CXCL16 is circulating in blood and upregulated in psoriatic skin (26). The chemokine is unique as it is one of only two chemokines that is expressed as a membrane bound protein exerting its chemotactic function after enzymatic cleavage (27–29). We have shown that early innate immune stimulation by TLR ligands can induce CXCL16 expression in keratinocytes and monocytes (30). CXCL16 contributes to psoriatic inflammation by mediating recruitment of neutrophils and CXCR6+ CD8+ T cells into human skin (26). Interestingly, membrane bound CXCL16 on peripheral monocytes also acts as a scavenger receptor for oxidized low-density lipoproteins (oxLDL) (31). The phagocytosis and accumulation of lipids in monocytes or macrophages can accelerate the conversion into foam cells and promote lipid plaques in arterial vessels (32–35), that suggests a link between cutaneous inflammation and associated comorbidities in psoriasis.
Therefore, we investigated the expression of CXCL16 on proinflammatory monocytes and its role in lipid uptake in psoriasis associated atherosclerosis and metabolic syndrome.
To investigate the role of the innate immune system in metabolic inflammation in psoriasis we examined CXCL16 expression on monocytes in 49 patients with psoriasis (mean age 57 ± 16.8 years, 33% female; mean PASI 18 ± 9.3) and correlated these levels with the extend of skin involvement and associated comorbidities. Hypertension and obesity (BMI ≥ 30 kg/m2) were present in 74% and 53% of the patients, respectively (Table 1). 37% of the patients presented with peripheral insulin resistance (pre-existing diabetes treated by oral antidiabetic drugs or insulin or elevated HbA1c), 43% had a hyperlipidemia (high serum level of cholesterol, HDL, LDL or triglyceride) and 47% were diagnosed with steatosis or atherosclerosis. 13 patients met all criteria for metabolic syndrome (27%) including insulin resistance, hyperlipidemia, obesity, and hypertension. One study participant aged 20 years had no other disorders besides psoriasis vulgaris.
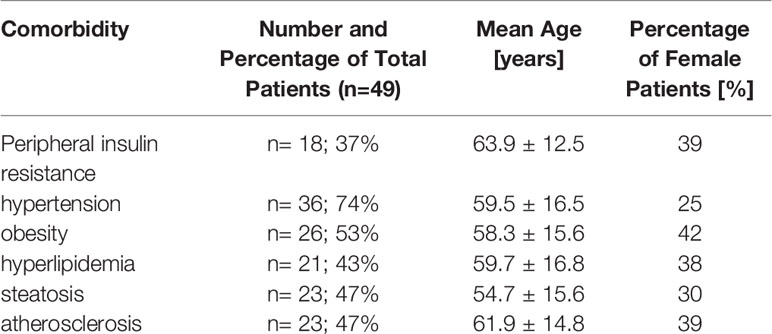
Table 1 Metabolic comorbidities in the investigated patients.
We found an enhanced expression of CXCL16 on all three monocyte subpopulations divided into classical monocytes with high expression of CD14 and low CD16 (CD14++CD16-), intermediate monocytes with expression of both antigens (CD14++CD16+) and the non-classical monocytes characterized by high amounts of CD16 and low CD14 on their surface (CD14low CD16++) (36) compared with healthy individuals (Figure 1A). In particular, the expression of CXCL16 on classical monocytes was increased threefold compared with controls (mean MFI CXCL16 healthy: 8.6; mean MFI CXCL16 psoriasis: 23.9). The highest expression of membrane bound CXCL16 was detected on intermediate monocytes of psoriatic patients (mean MFI CXCL16: 46.9) (Figure 1A). Interestingly, the intensity of CXCL16 expression on all monocytes and the monocytes subsets correlated significantly with the skin involvement scored by psoriasis activity and severity index (PASI) (Figures 1B, C). Obese patients with psoriasis showed a more severe clinical skin activity as well as a higher cardiovascular risk compared with non-obese patients (Figure 1D).
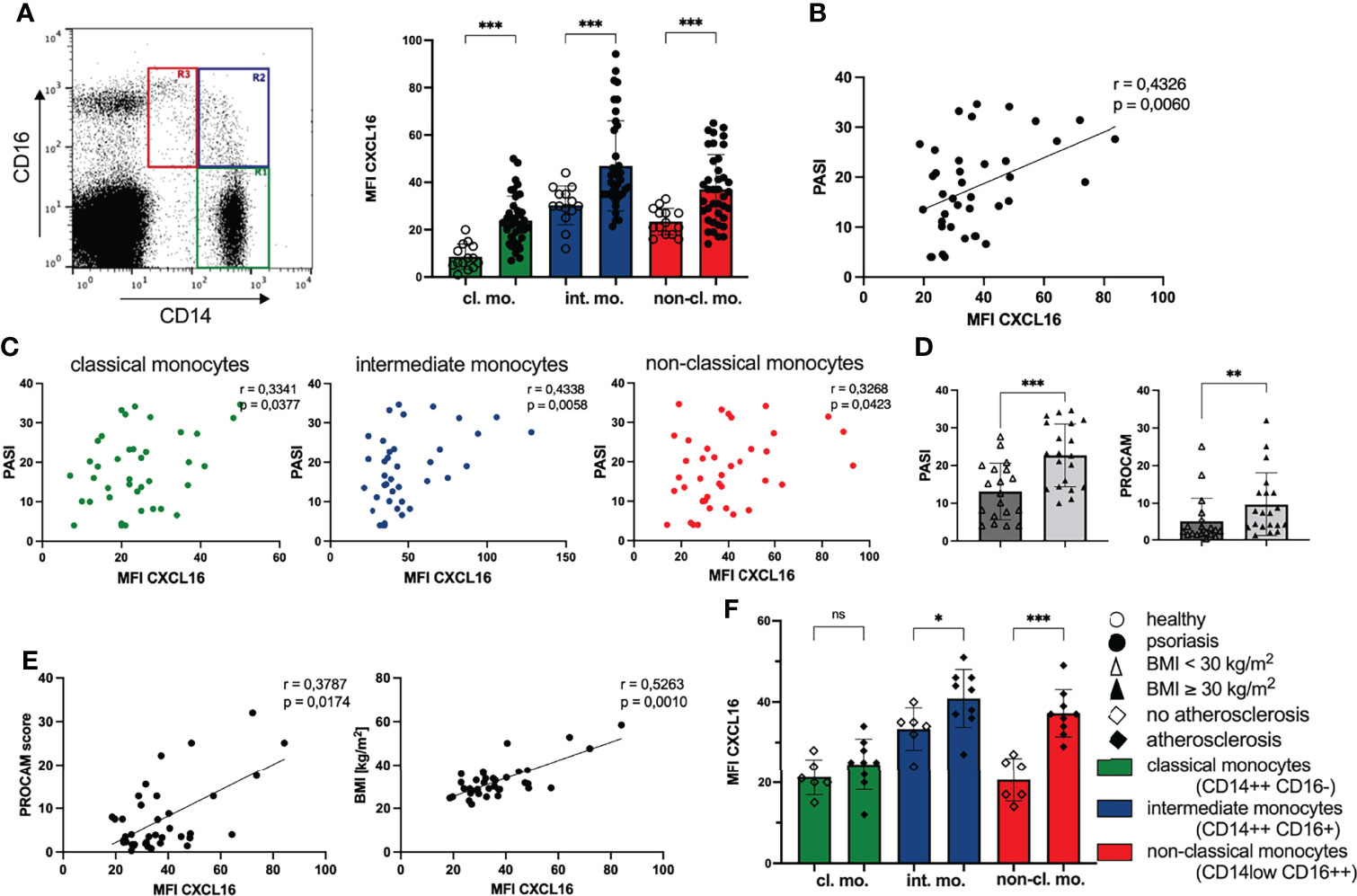
Figure 1 Expression of CXCL16 on monocytes is associated with enhanced clinical disease activity and cardiovascular risk. (A) Flow cytometry analysis of CXCL16 expression on monocyte subpopulations defined by gating for CD14 and CD16 expression in PBMC: classical monocytes (CD14++ CD16-, R1 green), intermediate monocytes (CD14++ CD16+, R2 blue) and non-classical monocytes (CD14low CD16++, R3, red), mean and SD of healthy controls (n= 14; unfilled dots) and patients with psoriasis vulgaris (n= 39; filled dots), Welch’s t test. (B) Correlation of CXCL16 expression on all monocytes of psoriatic patients (n=39) and clinical disease activity (PASI), Pearson’s correlation. (C) Correlation for each monocyte subset defined by color indicated in (A) and PASI score, Pearson’s correlation. (D) Mean and SD of PASI (unpaired t test) and PROCAM (Mann-Whitney test) of normal-weight (unfilled triangles; n=19) and obese psoriatic patients (filled triangles; n=20). (E) Correlation of chemokine expression with cardiovascular risk (PROCAM score); and body-mass-index (BMI). Each dot represents an individual patient, Pearson correlation. (F) CXCL16 expression on monocytes in psoriatic patients without atherosclerosis (n= 6; unfilled trapezoid) or with atherosclerosis (n= 9; filled trapezoid), mean and SD, unpaired t test. *p < 0.05; **p < 0.01; ***< 0.001. ns, not significant.
Furthermore, the expression of CXCL16 on peripheral monocytes of psoriatic patients correlated significantly with PROCAM (Prospective Cardiovascular Münster Study) score (Figure 1E) that indicates patients’ risk for developing cardiac diseases, such as myocardial infarction or cardiac death in the next 10 years (37). Extending this analysis, we searched for further correlations of chemokine expression with parameters potentiating the cardiovascular risk. We found a positive correlation of CXCL16 expression on monocytes and the body-mass-index in 39 psoriatic patients (Figure 1E). In addition, psoriatic patients with diagnosed atherosclerosis by medical imaging (transabdominal ultrasound or computed axial tomography or thoracal X-ray) displayed higher amounts of CXCL16 on their peripheral blood monocytes than psoriatic patients without atherosclerosis (Figure 1F). The clearest difference between these two groups was found in the expression of CXCL16 on non-classical monocytes with a nearly twice elevated chemokine expression in patients with psoriasis and atherosclerosis (MFI CXCL16 no atherosclerosis: 20.6; MFI CXCL16 atherosclerosis: 37.2). However, we did not observe a correlation between CXCL16 expression on monocytes and serum glucose, HbA1c or serum lipids (data not shown).
The membrane bound chemokine CXCL16 on monocytes is also a scavenger receptor for oxLDL (31). This promoted us to analyze the phagocytosis of oxLDL by CXCL16 on monocytes. We incubated PBMC with red fluorescing oxLDL for 3 hours and explored the fluorescence intensity of membrane bound CXCL16 on monocytes and intracellular oxLDL (Figure 2A). We found a positive correlation of the expressed membrane bound chemokine CXCL16 on the cell surface of monocytes and their uptake of oxLDL indicating that monocytes with elevated levels of CXCL16 on their surface accumulated phagocytosed oxLDL (Figure 2B). In line with this correlation, the classical and intermediate monocytes of psoriatic patients took up significantly more oxLDL than the monocytes of healthy individuals (Figure 2C). The mean fluorescence intensity of oxLDL in classical monocytes of psoriatic patients was 304 in comparison to classical monocytes of healthy volunteers with a MFI of 6. Likewise, the MFI of oxLDL in intermediate monocytes was increased threefold in psoriatic patients (MFI oxLDL healthy: 450; MFI oxLDL psoriasis: 1484). Interestingly, intermediate monocytes expressing highest amounts of CXCL16 on their surface also accumulated most intracellular oxLDL and monocytes of obese psoriatic patients accumulated a higher amount of oxLDL compared with normal-weight patients (Figure 2D).
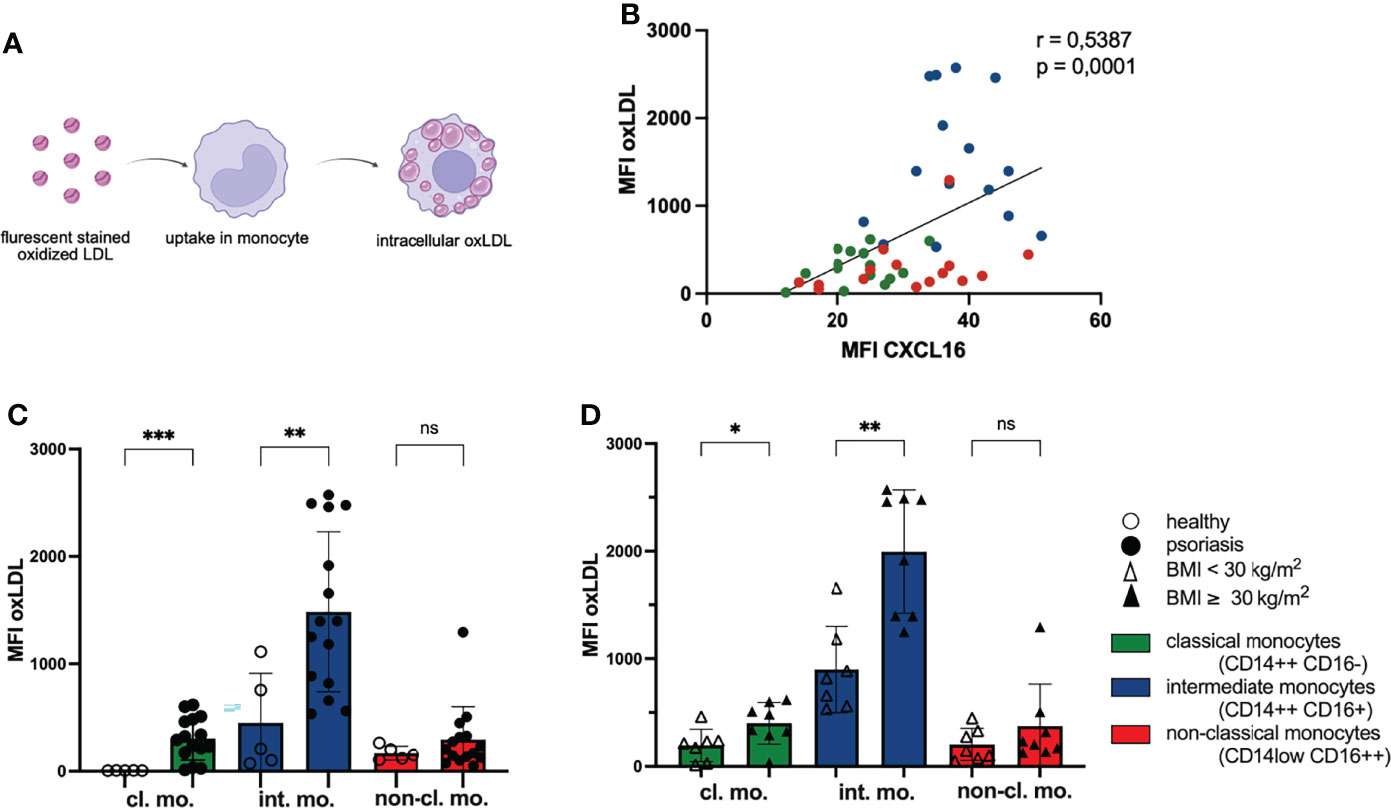
Figure 2 Phagocytosis of oxLDL by monocytes increases with CXCL16 expression. (A) PBMC were incubated with red fluorescing oxLDL for 3 h and subsequently stained for CD14, CD16 and CXCL16 expression. (B) The mean fluorescence intensity (MFI) of CXCL16 and oxLDL is shown. Every psoriasis patient is represented by three dots (n= 15). Colors represent monocyte subsets as defined in Figure 1A, Spearman correlation. (C) oxLDL concentrations in monocytes subsets of healthy controls (unfilled dots; n=5) and patients with psoriasis (filled dots; n=15), mean and SD, unpaired t test. (D) intracellular oxLDL concentrations in monocytes of normal-weight psoriatic patients (unfilled triangles; n=7) and obese patients (filled triangles; n=8), mean and SD, unpaired t test. *p < 0.05; **p < 0.01; ***p < 0.001. ns, not significant.
The uptake of oxLDL by monocytes/macrophages is an important pathogenic step in the development of atherosclerosis and occurs in arterial vessel walls (38). The migration of monocytes into this area might be supported by CXCL16 because the chemokine is also expressed by endothelial cells (34). Staining CXCR6 on monocyte subpopulations revealed highest expression on intermediate monocytes (Supplementary Figure 1). Interestingly, this subgroup had the highest capacity of oxLDL phagocytosis implicating a potential role in atherosclerotic plaque formation.
Obesity and metabolic diseases are associated with a dysbalance in innate lymphoid cells (39) that have also been suggested to play an important role in cutaneous psoriatic inflammation (21, 40–42). To investigate a possible systemic role for ILCs in the metabolic disorder psoriasis we first analyzed the composition of ILCs in the blood of psoriasis patients. Upon multicolor fluorescence staining and gating (Figure 3A) we observed enhanced levels of total ILCs and all three subgroups of ILCs in psoriatic patients compared to healthy controls (Figure 3B) indicating a general activation in patients with psoriasis. Each ILC subset was nearly twofold enhanced in psoriatic patients (mean ILC1 healthy: 42.8/µL and psoriasis: 84.1/µL; mean ILC2 healthy: 9.9/µL and psoriasis: 25.8/µL; mean ILC3 healthy: 21.9/µL and psoriasis: 40.33/µL) and the population of ILC1 was most frequent (Figure 3B).
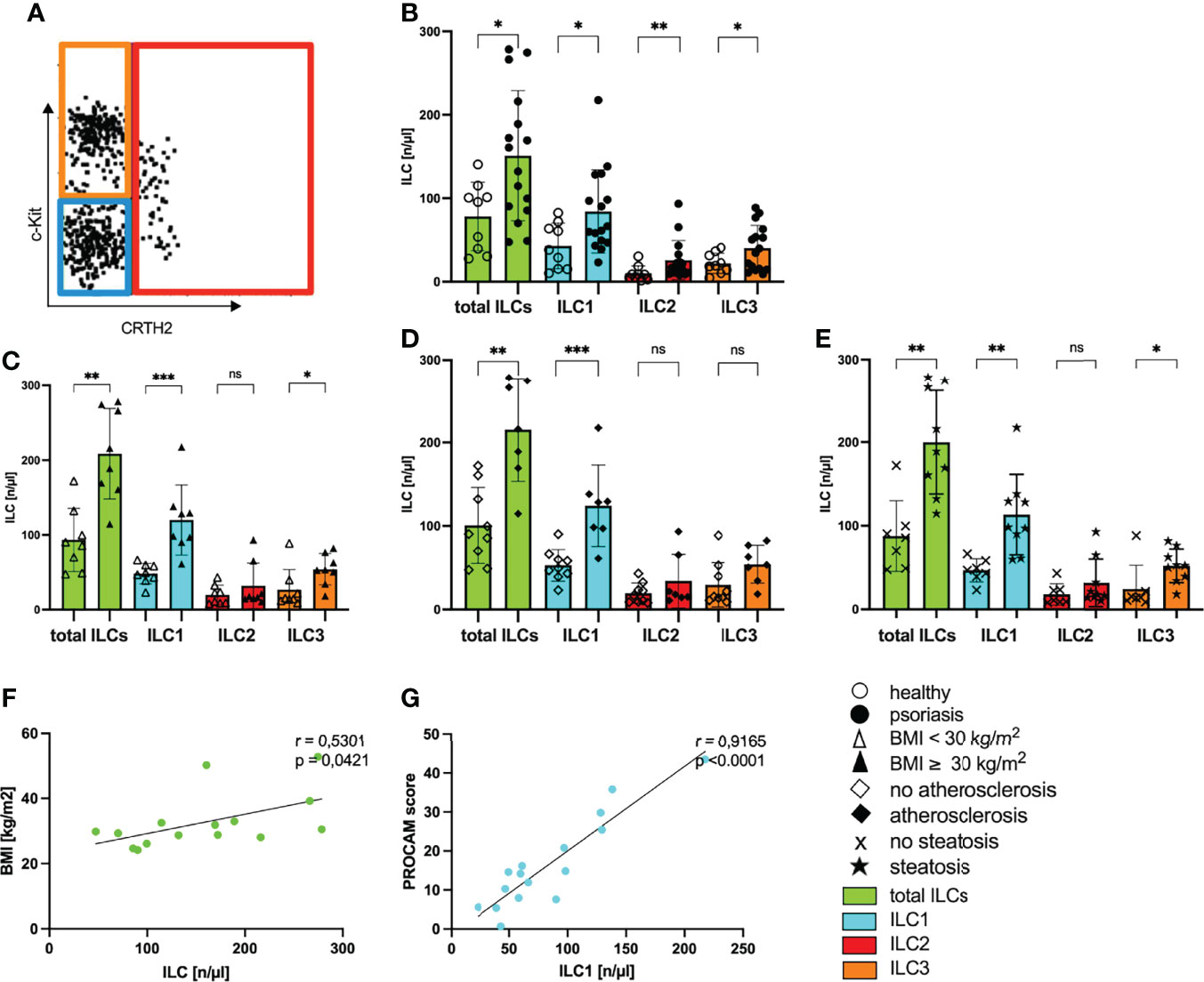
Figure 3 Increased numbers of ILCs in blood of psoriatic patients with metabolic disorders. (A) Gating strategy to define subpopulations of innate lymphoid cells ILC1, ILC2, ILC3. Analysis was performed by flow cytometry by using lineage Cocktail-Pacific Blue (Biolegend), CD11c-BV421, CD34-BV421, CD94-BV421, FcERIa-BV421, CD45-VioGreen, CD117-PE (c-Kit), IL7RA-APC (CD127), CRTH2-APC-Cy7 (CD294). ILCs are pregated as CD45 positive cells that are further defined by negative expression of the lineage cocktail and positive staining for IL7RA (Supplementary Figure 2). The figure is presenting cells covering these criteria and their further subdivision by individual expression of surface antigens CD117 and CRTH2. ILC1 are blue bordered (CD117-, CRTH2-), ILC2 red (CRTH2+) and ILC3 orange (CD117+, CRTH2-). Total ILCs are colored in green. (B) Number of ILCs of healthy controls (unfilled dots) and patients with psoriasis vulgaris (filled dots) in blood, mean and SD, each dot represents an individual patient, Mann-Whitney test. (C) Number of ILCs in blood of normal-weight psoriatic patients (body mass index, BMI<30 kg/m2; unfilled triangle; n=8) and obese patients (BMI ≥30 kg/m2; filled triangle; n=8) are shown, mean and SD, Mann-Whitney test. (D) Number of ILCs in blood of psoriatic patients without atherosclerosis (unfilled trapezoid; n= 9) and with atherosclerosis (filled trapezoid; n=7) are shown, mean and SD, Mann-Whitney test. (E) Number of ILCs of psoriatic patients without hepatic remodeling (cross symbol; n=7) and patients with steatosis hepatis (star; n=9), mean and SD, Mann-Whitney test. (F) Linear correlation of the number of circulating ILC and BMI (n=15), Pearson correlation. (G) Linear correlation of the number of circulating ILC1 and PROCAM score [%] (n=16), Pearson correlation. *p < 0.05; **p < 0.01; ***p < 0.001. ns, not significant.
When we examined the number of ILCs in psoriatic patients with metabolic disorders, we found significantly increased levels of ILCs in obese patients with psoriasis compared with non-obese patients (Figure 3C). Among the subsets, ILC1 were nearly twofold increased in patients with psoriasis and obesity (Figure 3C) indicating that there is a rise in the amount of ILC1 in psoriatic patients with metabolic disorders. In line with these results, the number of total ILCs and ILC1 were also enhanced in patients with additional hepatic steatosis or atherosclerosis (Figures 3D, E). The number of total ILCs correlated positively with the BMI of psoriatic patients (Figure 3F). Underlining the impact of ILC1 in psoriatic patients with metabolic disorders, we found a positive correlation between the number of circulating ILC1 and the PROCAM score measuring the cardiovascular risk (Figure 3G).
ILCs develop from innate lymphoid precursor cells which express high levels of CXCR6 (43), the chemokines receptor for CXCL16. CXCR6 was expressed on a subset of ILC subpopulation with the highest proportion in ILC1 and ILC3 where we observed well-demarcated CXCR6 positive subpopulations among ILC1 and ILC3 (Figure 4A). The intensity of CXCR6 expression on ILCs was enhanced in psoriatic patients (Figure 4B). In addition, mean expression intensity of CXCR6 on ILCs correlated with the PASI score indicating an association of skin involvement with the intensity of CXCR6 expression (Figure 4C). CXCR6 was only found in CCR7 negative ILCs and all of CXCR6+ ILCs co-expressed CCR6 (Figure 4D). This pattern indicated that CXCR6 expressing ILCs might be recruited to peripheral organs and this recruitment might be supported by CCR6. Indeed, when we analyzed the percentage of peripheral ILCs expressing the receptor CXCR6 on their surface, the numbers were reduced in patients with psoriasis (Figure 4E). This finding suggests that CXCR6+ ILCs are not prone to CCL21 or CCL19 mediated migration to lymphoid tissue (44) but rather migrate to peripheral organs with high CXCL16 expression such as the skin (26, 28) or atherosclerotic plaques (35) and that this pathway is active in obese individuals with elevated CXCL16 expression.
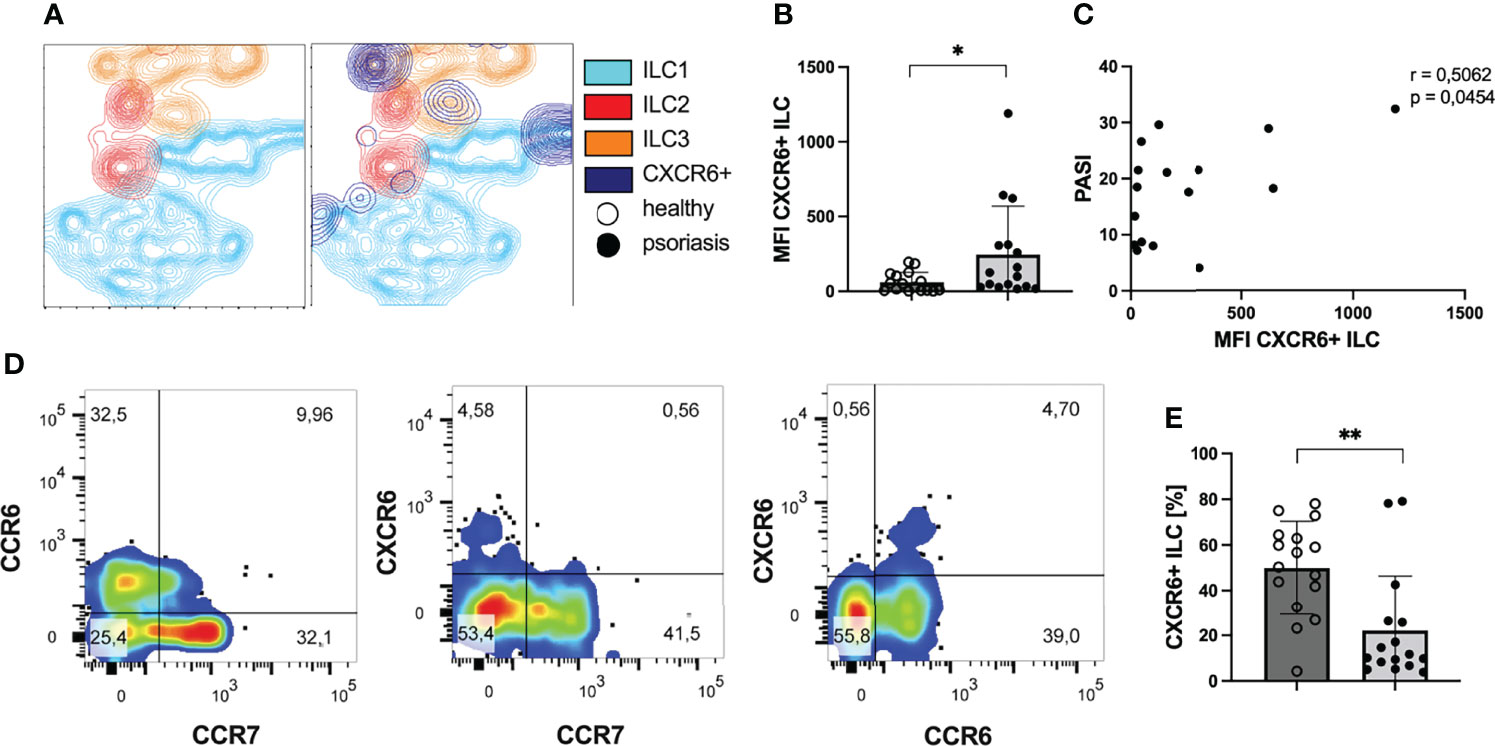
Figure 4 ILCs express CXCR6 in patients with psoriasis. (A) T-SNE contour plot presenting the clustering of ILCs in subgroups ILC1 (blue), ILC2 (red) and ILC3 (orange) and their expression of the chemokine receptor CXCR6 (dark blue). There are subgroups of ILC1 and ILC3 expressing the receptor CXCR6. (B) Flow cytometry analysis of MFI of CXCR6 expression on peripheral ILCs of psoriatic patients (n=16) and healthy volunteers (n=16), mean and SD, Welch t test. (C) Correlation of MFI of the receptor CXCR6 and the clinical skin involvment (n= 16), Pearson correlation. (D) Coexpression of chemokine receptors CXCR6, CCR6 and CCR7 among all ILCs. (E) Mean and SD of percentage of CXCR6 expressing ILCs among all ILCs of healthy volunteers (n=16) and psoriatic patients (n=16), Mann-Whitney test. *p < 0.05; **p < 0.01.
Here we show that the metabolic disease state in psoriasis is associated with an upregulation of the chemokine and lipid receptor CXCL16 on monocytes of psoriatic patients and the activation of ILCs. Expression of CXCL16 was increased on all subsets of monocytes circulating in the blood of psoriatic patients and this expression correlated not only with the PASI score, indicating severity of cutaneous involvement, but also with the amount of adipose tissue, as indicated by an elevated BMI.
It is known that obesity and increased body mass index (BMI) caused by high calorie diet are significant risk factors for the development of psoriasis (45–47). High calorie diet may exacerbate or trigger disease (46). Adipose tissue functions as endocrine organ that plays a key role in lipid and glucose metabolism, insulin-mediated processes and inflammation. The systemic effects are mediated by large abdominal white adipose tissue (48). However, adipose tissue in the skin might also impact the local inflammation (48). Secretion of cytokines such as IFN-γ and TNF-α by adipose tissue macrophages (49) could stimulate the expression of proinflammatory chemokines like CXCL16 in monocytes and keratinocytes in the skin (50). Further studies have shown that a high fat western diet is associated with an increased expression of type I interferon (IFN) and IFN stimulated genes (51). We have previously demonstrated, that type I IFNs are strong stimuli of CXCL16 upregulation in psoriatic dermis and epidermis that could support the cutaneous migration of neutrophils and CD8+ T cells (26, 52). Recently, expression of CXCR6 has been described on slan positive monocytes that correspond to the CD14low CD16++ non-classical monocytes (53). The cells migrated in vitro to CXCL16 which might support their recruitment into the skin of psoriatic patients where they accumulate and contribute to perpetuation of the disease (53–55).
On monocytes, CXCL16 is not only a chemokine but also a scavenger receptor for oxidized lipids (31). Interestingly, a high expression of CXCL16 on monocytes was associated with a stronger uptake of oxLDL which might have implications for the evolution of systemic disease manifestations in psoriasis. Lipid-overloaded monocytes may convert into foam cells in the intima (56) forming lipid plaques and arterial stenosis (32, 34). In addition, patients with metabolic syndrome reveal a higher percentage of circulating CXCR6 expressing platelets and CXCR6 expressing platelet-bound neutrophils, resulting in enhanced CXCR6/CXCL16-dependent adhesion to the dysfunctional arterial endothelium (57). Attached platelets form a white thrombus and present continuously CXCR6 to further circulating CXCL16 positive peripheral blood mononuclear cells (PBMCs) enhancing inflammation (58). Furthermore, endothelial CXCL16 is acting as potent adhesion ligand (58). This could be relevant for platelets, neutrophils and also for the immigration of monocytes that partly express CXCR6. (53). Therefore, chemokine CXLC16 und chemokine receptor CXCR6 are potent adhesion molecules in the atherosclerosis-prone vessel walls and thus may promote the progression of atherosclerosis. In line with this, psoriatic patients with coincident atherosclerosis expressed elevated levels of CXCL16 on monocytes compared with patients without vascular comorbidities. These findings suggest that CXCL16 expression on monocytes contributes to the development of cardiovascular diseases in psoriatic patients, which is further supported by the observed correlation between CXCL16 expression on monocytes and an enhanced PROCAM score.
In addition to the scavenger receptor function of CXCL16 that might result in lipid plaque formation, CXCL16 may induce cellular proliferation of aortic smooth muscle cells and pro-inflammatory cytokines in an NF-kappa B-dependent manner (33) and aggravate inflammation by adhesion of CXCR6 positive immune cells (34).
Among the innate immune cells perpetuating atherosclerosis are ILC1 (59). Here, we demonstrate that ILCs and especially ILC1 cells are more frequent in blood of psoriasis patients and the highest numbers are found in patients with atherosclerosis or obesity. ILC1 are characterized by their capability to produce high amounts of IFN-γ but also TNF-α, IL-1β and IL-6 (17, 18). Beside the role of these cytokines in psoriatic inflammation, ILC1 derived IFN-γ can accelerate atherosclerosis in mice (59). In addition, IFN-γ can affect macrophage homeostasis in the visceral adipose tissue by polarization toward M1 macrophages and the preferential killing of adipose M2 macrophages (22, 60). The proinflammatory M1 macrophages are capable to secrete TNF-α and IL-6 which may lead to insulin resistance (61, 62) and CXCL16 upregulation (30, 34).
Part of ILC1 expressed the chemokine receptor CXCR6 that might be relevant for their recruitment into atherosclerotic plaques, adipose tissue and even the skin. The expression of CXCR6 on ILCs was elevated in psoriasis patients. CXCR6 was expressed on a well described population of CCR7 negative ILCs. As CCR7 is required for guidance to lymphoid tissues by CCL19 and CCL21 (44, 63), the CXCR6 expressing cells might reflect a population on its way to peripheral organs with high CXCL16 expression such as the skin (28), atherosclerotic plaques (35) or the adipose tissue (64). Nearly all CXCR6+ ILCs co-expressed CCR6, the chemokine receptor for CCL20 which is known to induce migration of CCR6 expressing T cells to the skin (65) and is also upregulated in adipose tissue of obese individuals (66). This co-expression might enhance the migratory potential of the cells. The pattern was found in ILC1 and ILC3. The latter are of special importance for the cutaneous inflammation in psoriasis because they are competent producers of the TH17 cell-associated cytokines IL-17 and IL-22 (15, 20).
The finding demonstrating a reduced percentage of CXCR6 expressing ILCs in blood of patients with psoriasis compared with healthy controls might further support the concept of an active recruitment of CXCR6+ cells to peripheral sites. Trapping of ILCs in skin, atherosclerotic plaques or adipose tissue can explain their alleviation in the peripheral blood.
Following the current development in ILC differentiation and clustering that is driven by single cells sequencing a recent publication described CD94 positive cytotoxic ILCs upregulated in Crohn´s disease (67). As Crohn´s disease is associated with psoriasis (68) it would be interesting to investigate the prevalence of this population in psoriasis in a future analysis. Here, CD94+ cells were excluded as part of the NK-cell exclusion strategy following Soare et al. (69).
In conclusion, we showed that CXCL16 expression could be an important link between metabolic syndrome, the elevated cardiovascular risk and skin involvement of psoriasis vulgaris. Obesity was associated with an elevated expression of CXCL16 on monocytes that promoted the phagocytosis of oxLDL by scavenger receptor function of CXCL16. OxLDL loaded monocytes can contribute to the development of atherosclerosis. Secretion of CXCL16 in skin and atherosclerotic plaques might support accumulation of proinflammatory ILCs as innate immune stimulators that promote psoriatic inflammation. Therefore, therapeutic strategies could aim in reducing the expression of CXCL16 in psoriatic patients to prevent systemic manifestations. Collado et al. showed that CXCL16 neutralization by the angiotensin-1-receptor antagonist losartan significantly inhibited cell adhesion to arterial vessels. This effect was mediated by downregulation of CXCL16 expression on endothelial cells and therefore CXCL16 inhibition may positively affect the risk of metabolic disorders in psoriatic patients (57).
Blood samples of 49 patients with psoriasis were collected for further analysis of CXCL16 expression, oxLDL phagocytosis and determination of ILC subsets. The mean age of all patients was 57 ± 16.8 years, 33% female, mean PASI 18 ± 9.3. In most patients diagnosis of psoriasis has been known since 18 ± 16.2 years. In 23 of these patients, clinical data on serum lipids, HbA1c, CRP, blood pressure and medical imaging like transabdominal sonography or X-ray were assessed. The patients did not receive any systemic treatment with immunosuppressive drugs for at least 6 months. Diagnosis of psoriasis was confirmed by clinical and histologic criteria. Blood from age-matched healthy volunteers (mean age 58 ± 17,9 years, 68% female) was used as control. The investigational protocols (EK251062016) were approved by the Ethics Committee of the University Hospital of the Technical University Dresden according to the Declaration of Helsinki.
PBMCs were isolated from heparinized whole blood of patients with psoriasis and healthy controls by density gradient centrifugation using Biocoll separating solution (Biochrom, Berlin, Deutschland). Contaminating serum components were removed by washing the cells with PBS (phosphate buffered saline, containing 2% fetal calf serum, 0.02% NaN3) and cells were incubated with sort buffer (phosphate buffered saline, containing 2% fetal calf serum) for 10 minutes to block unspecific bindings. In order to assess low-density lipoprotein uptake, 1x106 cells/ml or 200.000 cells/200µl were seeded in 96 v-bottom well plate in RPMI-1640 medium (Gibco LifeTechnologies, Carlsbad, CA, containing 10% fetal calf serum, 2 mM L-glutamine, 1% nonessential amino acids, 100 U/ml penicillin, and 100 mg/ml streptomycin). Thereafter, 10µg/ml human Dil-labeled medium oxidized low-density lipoprotein (Dil-oxLDL, Kalen Biomedical, Germantown, USA #770232-9) was added and the cells incubated for at least 3 hours at 37°C. Subsequently, the cells were washed with PBS and cells were stained for CXCL16, CD14 and CD16 expression with fluorescence labeled antibodies (Table 2). The incubation with antibodies was performed for 30 minutes in the dark at 4°C on ice and followed by washing in PBS. Dead cells were excluded by Propidium Iodide (0,5 µg/ml) that was added directly before flow cytometry analysis was performed on a five-laser/18channel system (LSR Fortessa, Becton Dickenson, Franklin Lakes, NJ). Determination of transmembrane CXCL16 and intracellular oxLDL was analyzed with FlowJo software. Gating of cell populations were defined by isotype controls. Monocytes were characterized by expression of their surface antigens CD14 and CD16. Classical monocytes: CD14++CD16-, intermediate monocytes: CD14++CD16+ and non-classical monocytes: CD14low CD16++.

Table 2 Antibodies used for flow cytometry immunofluorescence for PBMC staining.
Whole blood (20ml) from healthy controls and psoriatic patients was collected into EDTA tubes. First, 500 μl of whole blood were put into one tube and mixed with fiuorochrome-labeled antibodies (Table 3) or respective isotype controls following previous reports (69) and incubated for 20 minutes. After washing, erythrocytes (RBCs) were lysed and nucleated cells were fixed using RBC lysis and fixation solution (BioLegend, Fell, Germany). Subsequently, flow cytometry analysis was performed on a five-laser/18channel system (LSR Fortessa, Becton Dickenson, Franklin Lakes, NJ) and analyzed with FlowJo software. ILCs were defined as CD45+, IL7RA+, negative for lineage markers CD3, CD11c, CD14, CD16, CD19, CD20, CD34, CD56, CD94, FcERIa (Supplementary Figure 2). In addition, ILC1 were defined as CRTH2-CD117-fraction, ILC2s as CRTH2+ CD117-, and ILC3s as CRTH2- CD117+ fractions, respectively (Supplementary Figure 2). Absolute numbers of cells were calculated as following: % ILC x absolute lymphocyte count (automated count) = absolute number of ILC per cubic millimeter. For calculation, the absolute lymphocyte count was determined by routine automated complete blood count measurement.
Table 3 Antibodies used for flow cytometry immunofluorescence for ILC staining.
For analysis of multiple chemokine receptor expression on ILCs, staining was performed on PBMC to increase the cell number. PBMC were separated from whole blood by density gradient centrifugation using Biocoll separating solution (Biochrom, Berlin, Deutschland). After isolation, cells were incubated with sort buffer (phosphate buffered saline, containing 2% fetal calf serum) for 10 minutes to block unspecific bindings. Then, cells were fixed in formaldehyde 4% and stained with the antibodies listed in Table 3. Additionally, mouse anti-human IgG against CCR7 -BV650 (BioLegend, #353234) and mouse anti-human IgG against CCR6 – PerCP (BioLegend, #353406) were added. Flow cytometry analysis was performed on a five-laser/18channel system (LSR Fortessa, Becton Dickenson, Franklin Lakes, NJ). Staining was analyzed with FlowJo software.
Data are presented as bars (indicating mean ± standard deviation) or dot plots (indicating the correlation coefficient r and the two-tailed (two-sided) P value performed by correlations). Test for normal distribution was performed by Shapiro-Wilk test. In normally distributed values unpaired t test possibly with Welch’s correction or Pearson correlation were used. For nonparametric tests Mann-Whitney test and Spearman correlation were used. Unless otherwise indicated computation with the help of GraphPad Prism 10 (GraphPad Software, San Diego, CA). In all cases, *P < 0.05 was considered to be statistically significant (**P < 0.01, ***P < 0.001).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by Ethics Committee of the University Hospital of the Technical University Dresden. The patients/participants provided their written informed consent to participate in this study.
CG conceived and directed the project. NZ and SR designed and LS performed experiments. LS and CG wrote the manuscript. SH, JS, AS, SK, and KB were involved in the recruitment and diagnosis of psoriasis patients. All authors listed have made a contribution to the work and approved the submitted version.
This study received funding from Novartis and Pfizer and Deutsche Forschungsgemeinschaft (German Research Foundation), grant TRR237 369799452/404458960 to CG. The funders were not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.916701/full#supplementary-material
1. Di Meglio P, Villanova F, Nestle FO. Psoriasis. Cold Spring Harbor Perspect Med (2014) 4(8):a015354. doi: 10.1101/cshperspect.a015354
2. Cohen AD, Sherf M, Vidavsky L, Vardy DA, Shapiro J, Meyerovitch J. Association Between Psoriasis and the Metabolic Syndrome. A Cross-Sectional Study. Dermatology (2008) 216(2):152–5. doi: 10.1159/000111512
3. Hao Y, Zhu YJ, Zou S, Zhou P, Hu YW, Zhao QX, et al. Metabolic Syndrome and Psoriasis: Mechanisms and Future Directions. Front Immunol (2021) 12:711060. doi: 10.3389/fimmu.2021.711060
4. Mallbris L, Ritchlin CT, Ståhle M. Metabolic Disorders in Patients With Psoriasis and Psoriatic Arthritis. Curr Rheumatol Rep (2006) 8(5):355–63. doi: 10.1007/s11926-006-0065-8
5. Langan SM, Seminara NM, Shin DB, Troxel AB, Kimmel SE, Mehta NN, et al. Prevalence of Metabolic Syndrome in Patients With Psoriasis: A Population-Based Study in the United Kingdom. J Invest Dermatol (2012) 132(3 Pt 1):556–62. doi: 10.1038/jid.2011.365
6. Prodanovich S, Kirsner RS, Kravetz JD, Ma F, Martinez L, Federman DG. Association of Psoriasis With Coronary Artery, Cerebrovascular, and Peripheral Vascular Diseases and Mortality. Arch Dermatol (2009) 145(6):700–3. doi: 10.1001/archdermatol.2009.94
7. Qureshi AA, Choi HK, Setty AR, Curhan GC. Psoriasis and the Risk of Diabetes and Hypertension: A Prospective Study of US Female Nurses. Arch Dermatol (2009) 145(4):379–82. doi: 10.1001/archdermatol.2009.48
8. Sommer DM, Jenisch S, Suchan M, Christophers E, Weichenthal M. Increased Prevalence of the Metabolic Syndrome in Patients With Moderate to Severe Psoriasis. Arch Dermatol Res (2006) 298(7):321–8. doi: 10.1007/s00403-006-0703-z
9. Pearce DJ, Morrison AE, Higgins KB, Crane MM, Balkrishnan R, Fleischer AB Jr., et al. The Comorbid State of Psoriasis Patients in a University Dermatology Practice. J Dermatol Treat (2005) 16(5-6):319–23. doi: 10.1080/09546630500335977
10. Gelfand JM, Neimann AL, Shin DB, Wang X, Margolis DJ, Troxel AB. Risk of Myocardial Infarction in Patients With Psoriasis. Jama (2006) 296(14):1735–41. doi: 10.1001/jama.296.14.1735
11. Gisondi P, Fostini AC, Fossà I, Girolomoni G, Targher G. Psoriasis and the Metabolic Syndrome. Clin Dermatol (2018) 36(1):21–8. doi: 10.1016/j.clindermatol.2017.09.005
12. Kumar S, Han J, Li T, Qureshi AA. Obesity, Waist Circumference, Weight Change and the Risk of Psoriasis in US Women. J Eur Acad Dermatol Venereol (2013) 27(10):1293–8. doi: 10.1111/jdv.12001
13. Cherrier M, Ohnmacht C, Cording S, Eberl G. Development and Function of Intestinal Innate Lymphoid Cells. Curr Opin Immunol (2012) 24(3):277–83. doi: 10.1016/j.coi.2012.03.011
14. Guo L, Junttila IS, Paul WE. Cytokine-Induced Cytokine Production by Conventional and Innate Lymphoid Cells. Trends Immunol (2012) 33(12):598–606. doi: 10.1016/j.it.2012.07.006
15. Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, et al. Innate Lymphoid Cells–a Proposal for Uniform Nomenclature. Nat Rev Immunol (2013) 13(2):145–9. doi: 10.1038/nri3365
16. Kim CH, Hashimoto-Hill S, Kim M. Migration and Tissue Tropism of Innate Lymphoid Cells. Trends Immunol (2016) 37(1):68–79. doi: 10.1016/j.it.2015.11.003
17. Fuchs A, Vermi W, Lee JS, Lonardi S, Gilfillan S, Newberry RD, et al. Intraepithelial Type 1 Innate Lymphoid Cells are a Unique Subset of IL-12- and IL-15-Responsive IFN-γ-Producing Cells. Immunity (2013) 38(4):769–81. doi: 10.1016/j.immuni.2013.02.010
18. Klose CSN, Flach M, Möhle L, Rogell L, Hoyler T, Ebert K, et al. Differentiation of Type 1 ILCs From a Common Progenitor to All Helper-Like Innate Lymphoid Cell Lineages. Cell (2014) 157(2):340–56. doi: 10.1016/j.cell.2014.03.030
19. Rauber S, Luber M, Weber S, Maul L, Soare A, Wohlfahrt T, et al. Resolution of Inflammation by Interleukin-9-Producing Type 2 Innate Lymphoid Cells. Nat Med (2017) 23(8):938–44. doi: 10.1038/nm.4373
20. Diefenbach A, Colonna M, Koyasu S. Development, Differentiation, and Diversity of Innate Lymphoid Cells. Immunity (2014) 41(3):354–65. doi: 10.1016/j.immuni.2014.09.005
21. Teunissen MBM, Munneke JM, Bernink JH, Spuls PI, Res PCM, Te Velde A, et al. Composition of Innate Lymphoid Cell Subsets in the Human Skin: Enrichment of NCR(+) ILC3 in Lesional Skin and Blood of Psoriasis Patients. J Invest Dermatol (2014) 134(9):2351–60. doi: 10.1038/jid.2014.146
22. O'Sullivan TE, Rapp M, Fan X, Weizman OE, Bhardwaj P, Adams NM, et al. Adipose-Resident Group 1 Innate Lymphoid Cells Promote Obesity-Associated Insulin Resistance. Immunity (2016) 45(2):428–41. doi: 10.1016/j.immuni.2016.06.016
23. Rolot M, O'Sullivan TE. Living With Yourself: Innate Lymphoid Cell Immunometabolism. Cells (2020) 9(2):334. doi: 10.3390/cells9020334
24. Wang H, Shen L, Sun X, Liu F, Feng W, Jiang C, et al. Adipose Group 1 Innate Lymphoid Cells Promote Adipose Tissue Fibrosis and Diabetes in Obesity. Nat Commun (2019) 10(1):3254. doi: 10.1038/s41467-019-11270-1
25. Pantelyushin S, Haak S, Ingold B, Kulig P, Heppner FL, Navarini AA, et al. Rorγt+ Innate Lymphocytes and γδ T Cells Initiate Psoriasiform Plaque Formation in Mice. J Clin Invest (2012) 122(6):2252–6. doi: 10.1172/JCI61862
26. Gunther C, Carballido-Perrig N, Kaesler S, Carballido JM, Biedermann T. CXCL16 and CXCR6 are Upregulated in Psoriasis and Mediate Cutaneous Recruitment of Human CD8+ T Cells. J Invest Dermatol (2012) 132(3 Pt 1):626–34. doi: 10.1038/jid.2011.371
27. Matloubian M, David A, Engel S, Ryan JE, Cyster JG. A Transmembrane CXC Chemokine is a Ligand for HIV-Coreceptor Bonzo. Nat Immunol (2000) 1(4):298–304. doi: 10.1038/79738
28. Oh ST, Schramme A, Tilgen W, Gutwein P, Reichrath J. Overexpression of CXCL16 in Lesional Psoriatic Skin. Dermatoendocrinol (2009) 1(2):114–8. doi: 10.4161/derm.1.2.7750
29. Shimaoka T, Kume N, Minami M, Hayashida K, Kataoka H, Kita T, et al. Molecular Cloning of a Novel Scavenger Receptor for Oxidized Low Density Lipoprotein, SR-PSOX, on Macrophages. J Biol Chem (2000) 275(52):40663–6. doi: 10.1074/jbc.C000761200
30. Steffen S, Abraham S, Herbig M, Schmidt F, Blau K, Meisterfeld S, et al. Toll-Like Receptor-Mediated Upregulation of CXCL16 in Psoriasis Orchestrates Neutrophil Activation. J Invest Dermatol (2018) 138(2):344–54. doi: 10.1016/j.jid.2017.08.041
31. Minami M, Kume N, Shimaoka T, Kataoka H, Hayashida K, Akiyama Y, et al. Expression of SR-PSOX, a Novel Cell-Surface Scavenger Receptor for Phosphatidylserine and Oxidized LDL in Human Atherosclerotic Lesions. Arterioscler Thromb Vasc Biol (2001) 21(11):1796–800. doi: 10.1161/hq1001.096652
32. Aslanian AM, Charo IF. Targeted Disruption of the Scavenger Receptor and Chemokine CXCL16 Accelerates Atherosclerosis. Circulation (2006) 114(6):583–90. doi: 10.1161/CIRCULATIONAHA.105.540583
33. Chandrasekar B, Bysani S, Mummidi S. CXCL16 Signals via Gi, Phosphatidylinositol 3-Kinase, Akt, I Kappa B Kinase, and Nuclear Factor-Kappa B and Induces Cell-Cell Adhesion and Aortic Smooth Muscle Cell Proliferation. J Biol Chem (2004) 279(5):3188–96. doi: 10.1074/jbc.M311660200
34. Hofnagel O, Engel T, Severs NJ, Robenek H, Buers I. SR-PSOX at Sites Predisposed to Atherosclerotic Lesion Formation Mediates Monocyte-Endothelial Cell Adhesion. Atherosclerosis (2011) 217(2):371–8. doi: 10.1016/j.atherosclerosis.2011.04.021
35. Wuttge DM, Zhou X, Sheikine Y, Wågsäter D, Stemme V, Hedin U, et al. CXCL16/SR-PSOX is an Interferon-Gamma-Regulated Chemokine and Scavenger Receptor Expressed in Atherosclerotic Lesions. Arterioscler Thromb Vasc Biol (2004) 24(4):750–5. doi: 10.1161/01.ATV.0000124102.11472.36
36. Ziegler-Heitbrock L, Ancuta P, Crowe S, Dalod M, Grau V, Hart DN, et al. Nomenclature of Monocytes and Dendritic Cells in Blood. Blood (2010) 116(16):e74–80. doi: 10.1182/blood-2010-02-258558
37. Assmann G. [Assessment of Cardiovascular Risk–PROCAM and New Algorithms]. BioMed Tech (Berl) (2005) 50(7-8):227–32. doi: 10.1515/BMT.2005.033
38. Rosenfeld ME, Khoo JC, Miller E, Parthasarathy S, Palinski W, Witztum JL. Macrophage-Derived Foam Cells Freshly Isolated From Rabbit Atherosclerotic Lesions Degrade Modified Lipoproteins, Promote Oxidation of Low-Density Lipoproteins, and Contain Oxidation-Specific Lipid-Protein Adducts. J Clin Invest (1991) 87(1):90–9. doi: 10.1172/JCI115006
39. Saetang J, Sangkhathat S. Role of Innate Lymphoid Cells in Obesity and Metabolic Disease (Review). Mol Med Rep (2018) 17(1):1403–12. doi: 10.3892/mmr.2017.8038
40. Brüggen MC, Bauer WM, Reininger B, Clim E, Captarencu C, Steiner GE, et al. In Situ Mapping of Innate Lymphoid Cells in Human Skin: Evidence for Remarkable Differences Between Normal and Inflamed Skin. J Invest Dermatol (2016) 136(12):2396–405. doi: 10.1016/j.jid.2016.07.017
41. Dyring-Andersen B, Geisler C, Agerbeck C, Lauritsen JP, Gúdjonsdottir SD, Skov L, et al. Increased Number and Frequency of Group 3 Innate Lymphoid Cells in Nonlesional Psoriatic Skin. Br J Dermatol (2014) 170(3):609–16. doi: 10.1111/bjd.12658
42. Villanova F, Flutter B, Tosi I, Grys K, Sreeneebus H, Perera GK, et al. Characterization of Innate Lymphoid Cells in Human Skin and Blood Demonstrates Increase of NKp44+ ILC3 in Psoriasis. J Invest Dermatol (2014) 134(4):984–91. doi: 10.1038/jid.2013.477
43. Chea S, Possot C, Perchet T, Petit M, Cumano A, Golub R. CXCR6 Expression Is Important for Retention and Circulation of ILC Precursors. Mediators Inflamm (2015) 2015:368427. doi: 10.1155/2015/368427
44. Hauser MA, Legler DF. Common and Biased Signaling Pathways of the Chemokine Receptor CCR7 Elicited by its Ligands CCL19 and CCL21 in Leukocytes. J Leukoc Biol (2016) 99(6):869–82. doi: 10.1189/jlb.2MR0815-380R
45. Armstrong AW, Harskamp CT, Armstrong EJ. The Association Between Psoriasis and Obesity: A Systematic Review and Meta-Analysis of Observational Studies. Nutr Diabetes (2012) 2(12):e54. doi: 10.1038/nutd.2012.26
46. Barrea L, Nappi F, Di Somma C, Savanelli MC, Falco A, Balato A, et al. Environmental Risk Factors in Psoriasis: The Point of View of the Nutritionist. Int J Environ Res Public Health (2016) 13(5):743. doi: 10.3390/ijerph13070743
47. Fleming P, Kraft J, Gulliver WP, Lynde C. The Relationship of Obesity With the Severity of Psoriasis: A Systematic Review. J Cutan Med Surg (2015) 19(5):450–6. doi: 10.1177/1203475415586332
48. Wong Y, Nakamizo S, Tan KJ, Kabashima K. An Update on the Role of Adipose Tissues in Psoriasis. Front Immunol (2019) 10:1507. doi: 10.3389/fimmu.2019.01507
49. Ożańska A, Szymczak D, Rybka J. Pattern of Human Monocyte Subpopulations in Health and Disease. Scand J Immunol (2020) 92(1):e12883. doi: 10.1111/sji.12883
50. Abel S, Hundhausen C, Mentlein R, Schulte A, Berkhout TA, Broadway N, et al. The Transmembrane CXC-Chemokine Ligand 16 is Induced by IFN-Gamma and TNF-Alpha and Shed by the Activity of the Disintegrin-Like Metalloproteinase ADAM10. J Immunol (2004) 172(10):6362–72. doi: 10.4049/jimmunol.172.10.6362
51. Christ A, Günther P, Lauterbach MAR, Duewell P, Biswas D, Pelka K, et al. Western Diet Triggers NLRP3-Dependent Innate Immune Reprogramming. Cell (2018) 172(1-2):162–75.e14. doi: 10.1016/j.cell.2017.12.013
52. Tohyama M, Sayama K, Komatsuzawa H, Hanakawa Y, Shirakata Y, Dai X, et al. CXCL16 is a Novel Mediator of the Innate Immunity of Epidermal Keratinocytes. Int Immunol (2007) 19(9):1095–102. doi: 10.1093/intimm/dxm083
53. Hamers AAJ, Dinh HQ, Thomas GD, Marcovecchio P, Blatchley A, Nakao CS, et al. Human Monocyte Heterogeneity as Revealed by High-Dimensional Mass Cytometry. Arterioscler Thromb Vasc Biol (2019) 39(1):25–36. doi: 10.1161/ATVBAHA.118.311022
54. Gunther C, Starke J, Zimmermann N, Schakel K. Human 6-Sulfo LacNAc (Slan) Dendritic Cells are a Major Population of Dermal Dendritic Cells in Steady State and Inflammation. Clin Exp Dermatol (2012) 37(2):169–76. doi: 10.1111/j.1365-2230.2011.04213.x
55. Hansel A, Gunther C, Ingwersen J, Starke J, Schmitz M, Bachmann M, et al. Human Slan (6-Sulfo LacNAc) Dendritic Cells are Inflammatory Dermal Dendritic Cells in Psoriasis and Drive Strong TH17/TH1 T-Cell Responses. J Allergy Clin Immunol (2011) 127(3):787–94. doi: 10.1016/j.jaci.2010.12.009
56. Goldstein JL, Ho YK, Basu SK, Brown MS. Binding Site on Macrophages That Mediates Uptake and Degradation of Acetylated Low Density Lipoprotein, Producing Massive Cholesterol Deposition. Proc Natl Acad Sci USA (1979) 76(1):333–7. doi: 10.1073/pnas.76.1.333
57. Collado A, Marques P, Escudero P, Rius C, Domingo E, Martinez-Hervás S, et al. Functional Role of Endothelial CXCL16/CXCR6-Platelet-Leucocyte Axis in Angiotensin II-Associated Metabolic Disorders. Cardiovasc Res (2018) 114(13):1764–75. doi: 10.1093/cvr/cvy135
58. Linke B, Meyer Dos Santos S, Picard-Willems B, Keese M, Harder S, Geisslinger G, et al. CXCL16/CXCR6-Mediated Adhesion of Human Peripheral Blood Mononuclear Cells to Inflamed Endothelium. Cytokine (2019) 122:154081. doi: 10.1016/j.cyto.2017.06.008
59. Wu C, He S, Liu J, Wang B, Lin J, Duan Y, et al. Type 1 Innate Lymphoid Cell Aggravation of Atherosclerosis is Mediated Through TLR4. Scand J Immunol (2018) 87(5):e12661. doi: 10.1111/sji.12661
60. Boulenouar S, Michelet X, Duquette D, Alvarez D, Hogan AE, Dold C, et al. Adipose Type One Innate Lymphoid Cells Regulate Macrophage Homeostasis Through Targeted Cytotoxicity. Immunity (2017) 46(2):273–86. doi: 10.1016/j.immuni.2017.01.008
61. Shoelson SE, Lee J, Goldfine AB. Inflammation and Insulin Resistance. J Clin Invest (2006) 116(7):1793–801. doi: 10.1172/JCI29069
62. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is Associated With Macrophage Accumulation in Adipose Tissue. J Clin Invest (2003) 112(12):1796–808. doi: 10.1172/JCI200319246
63. Förster R, Davalos-Misslitz AC, Rot A. CCR7 and its Ligands: Balancing Immunity and Tolerance. Nat Rev Immunol (2008) 8(5):362–71. doi: 10.1038/nri2297
64. Santiago-Fernández C, Martín-Reyes F, Tome M, Gutierrez-Repiso C, Fernandez-Garcia D, Ocaña-Wilhelmi L, et al. Oxidized LDL Increase the Proinflammatory Profile of Human Visceral Adipocytes Produced by Hypoxia. Biomedicines (2021) 9(11):1715. doi: 10.3390/biomedicines9111715
65. Harper EG, Guo C, Rizzo H, Lillis JV, Kurtz SE, Skorcheva I, et al. Th17 Cytokines Stimulate CCL20 Expression in Keratinocytes In Vitro and In Vivo: Implications for Psoriasis Pathogenesis. J Invest Dermatol (2009) 129(9):2175–83. doi: 10.1038/jid.2009.65
66. El Shamieh S, Stathopoulou M, Bonnefond A, Ndiaye N, Lecoeur C, Meyre D, et al. Obesity Status Modifies the Association Between Rs7556897t>C in the Intergenic Region SLC19A3-CCL20 and Blood Pressure in French Children. Clin Chem Lab Med (CCLM) (2020) 1:1819–27. doi: 10.1515/cclm-2019-0292
67. Krabbendam L, Heesters BA, Kradolfer CMA, Haverkate NJE, Becker MAJ, Buskens CJ, et al. CD127+ CD94+ Innate Lymphoid Cells Expressing Granulysin and Perforin are Expanded in Patients With Crohn’s Disease. Nat Commun (2021) 12(1):5841. doi: 10.1038/s41590-019-0423-0
68. Najarian DJ, Gottlieb AB. Connections Between Psoriasis and Crohn's Disease. J Am Acad Dermatol (2003) 48(6):805–21. doi: 10.1067/mjd.2003.540
Keywords: psoriasis, CXCL16, ILC, metabolic syndrome, obesity, monocytes, CXCR6
Citation: Schielke L, Zimmermann N, Hobelsberger S, Steininger J, Strunk A, Blau K, Hernandez J, Künzel S, Ziegenbalg R, Rösing S, Beissert S, Abraham S and Günther C (2022) Metabolic Syndrome in Psoriasis Is Associated With Upregulation of CXCL16 on Monocytes and a Dysbalance in Innate Lymphoid Cells. Front. Immunol. 13:916701. doi: 10.3389/fimmu.2022.916701
Received: 09 April 2022; Accepted: 19 May 2022;
Published: 17 June 2022.
Edited by:
Anja Saalbach, Leipzig University, GermanyReviewed by:
Christoph Siegfried Niki Klose, Charité Universitätsmedizin Berlin, GermanyCopyright © 2022 Schielke, Zimmermann, Hobelsberger, Steininger, Strunk, Blau, Hernandez, Künzel, Ziegenbalg, Rösing, Beissert, Abraham and Günther. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lisa Schielke, bGlzYS5zY2hpZWxrZUB0dS1kcmVzZGVuLmRl; Claudia Günther, Y2xhdWRpYS5ndWVudGhlckB1a2RkLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.