 Carolin Christina Drost1
Carolin Christina Drost1 Alexandros Rovas1
Alexandros Rovas1 Irina Osiaevi2
Irina Osiaevi2 Matthias Rauen1
Matthias Rauen1 Johan van der Vlag3Baranca Buijsers3Rustem Salmenov3Alexander Lukasz1
Johan van der Vlag3Baranca Buijsers3Rustem Salmenov3Alexander Lukasz1 Hermann Pavenstädt1
Hermann Pavenstädt1 Wolfgang A. Linke4
Wolfgang A. Linke4 Philipp Kümpers1*
Philipp Kümpers1*- 1Department of Medicine D, Division of General Internal and Emergency Medicine, Nephrology, and Rheumatology, University Hospital Münster, Münster, Germany
- 2Department of Medicine A, Division of Hematology, Oncology, Hemostaseology and Pneumology, University Hospital Münster, Münster, Germany
- 3Department of Nephrology, Radboud Institute for Molecular Life Sciences, Radboud University Medical Center, Nijmegen, Netherlands
- 4Institute of Physiology II, University of Münster, Münster, Germany
Coronavirus disease 2019 (COVID-19) is a systemic disease associated with injury (thinning) of the endothelial glycocalyx (eGC), a protective layer on the vascular endothelium. The aim of this translational study was to investigate the role of the eGC-degrading enzyme heparanase (HPSE), which is known to play a central role in the destruction of the eGC in bacterial sepsis. Excess activity of HPSE in plasma from COVID-19 patients correlated with several markers of eGC damage and perfused boundary region (PBR, an inverse estimate of glycocalyx dimensions of vessels with a diameter 4-25 µm). In a series of translational experiments, we demonstrate that the changes in eGC thickness of cultured cells exposed to COVID-19 serum correlated closely with HPSE activity in concordant plasma samples (R = 0.82, P = 0.003). Inhibition of HPSE by a nonanticoagulant heparin fragment prevented eGC injury in response to COVID-19 serum, as shown by atomic force microscopy and immunofluorescence imaging. Our results suggest that the protective effect of heparin in COVID-19 may be due to an eGC-protective off-target effect.
Introduction
Coronavirus disease 2019 (COVID-19) caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is presenting as a systemic disease associated with vascular inflammation and endothelial injury (1–3). Using novel, quantitative sublingual video microscopy, we were able to show that severe damage (thinning) of the endothelial glycocalyx (eGC) predicted 60-day in-hospital mortality in our cohort of COVID-19 patients (4).
The endothelial glycocalyx (eGC) is a delicate gel-like layer coating the luminal surface of the vascular endothelium (5, 6). It is up to 3 µm thick, largely consists of highly sulfated glycosaminoglycans and proteoglycans, and it plays a pivotal role in the maintenance of microcirculatory homeostasis (7, 8). Specifically, the eGC acts as a negatively charged “firewall” to reduce leukocyte-endothelial-interactions (9). Its carbohydrate-rich matrix provides resistance to water permeability and contributes to the proportion of albumin molecules “reflected” back into plasma by the vessel wall (10, 11). Beyond that, the glycocalyx contributes to the regulation of the redox state and is crucially involved in the mediation of shear-induced nitric oxide release as well as physiologic anticoagulation (8, 12, 13).
At least in bacterial sepsis, the final common pathway of eGC damage appears to be remarkably consistent: the activation and release of the heparan sulfate (HS)-degrading enzyme heparanase (HPSE) (14, 15). Besides cleaving HS from the cell surface, HPSE also enhances shedding of transmembrane HS proteoglycan syndecan-1 (Syn-1) by upregulating the expression of matrix metalloproteinase 9 (MMP9), which is a syndecan sheddase. Therefore, HPSE (in)directly contributes to increased HS and Syn-1 plasma levels, which are both markers of endothelial glycocalyx degradation. The aim of this translational proof-of-concept study was to investigate the role of HPSE in COVID-19-induced eGC damage.
Methods
Study Design and Study Population
Sublingual video microscopy and blood sampling were performed in 16 adult PCR-confirmed COVID-19 patients, prospectively enrolled from May to June 2020 at the intensive care units (ICU) of the University Hospital Münster and three local teaching hospitals in a non-consecutive manner. Plasma samples were obtained, centrifuged, and stored at -80°C until analysis. Twelve apparently healthy, age-matched volunteers served as controls (Table 1). Some of the participants were already included in a previous study (4). The study was performed in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the General Medical Council Westfalen-Lippe and the WWU Münster, Germany (file number: amendment to 2016-073-f-S). Written informed consent to participate has been obtained from all individuals.
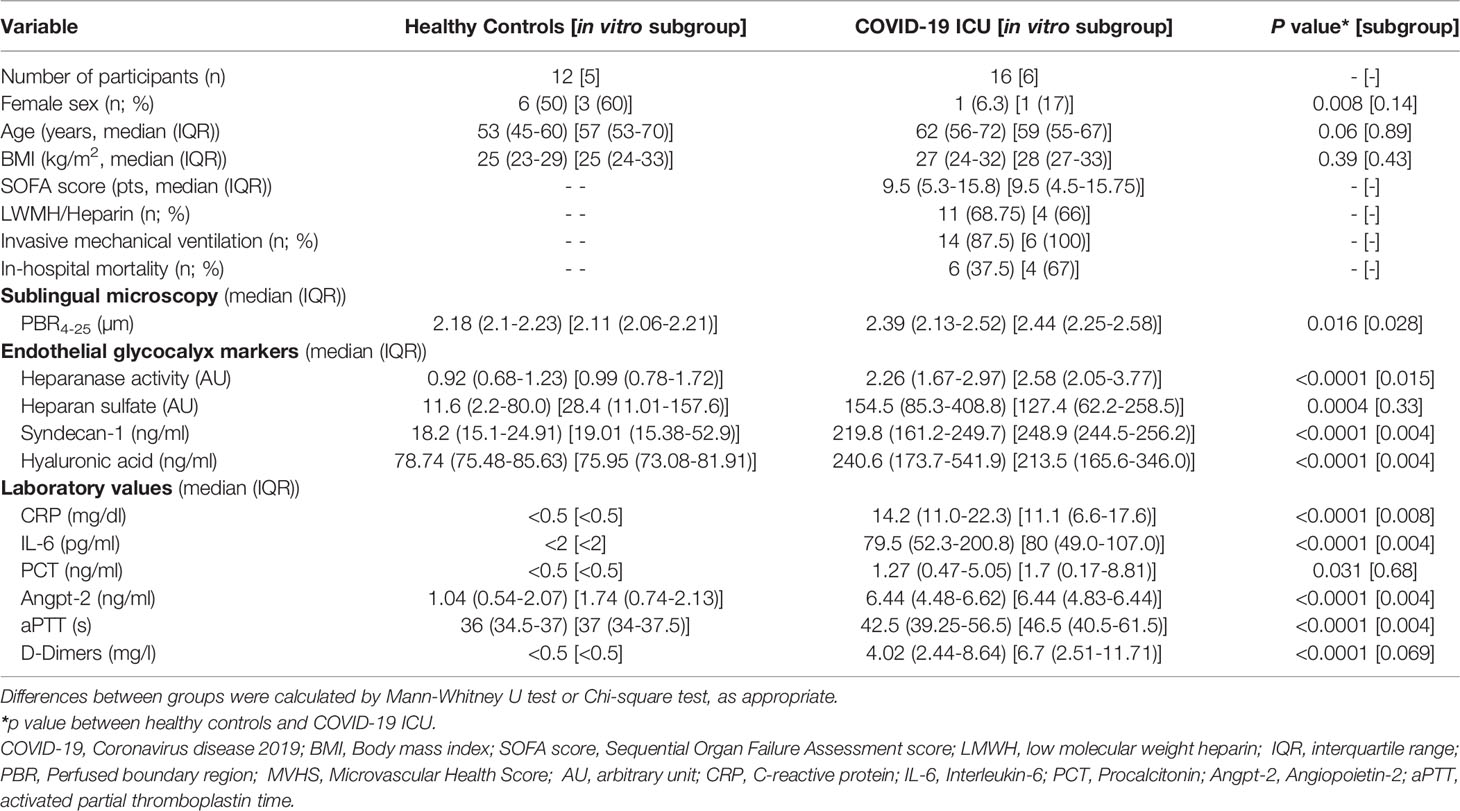
Table 1 Baseline characteristics.
In Vivo Assessment of Sublingual Glycocalyx Dimensions
Real-time intravital microscopy was performed at the bedside with a sidestream dark field (SDF) camera (CapiScope HVCS, KK Technology, Honiton, UK) to visualize the sublingual microvasculature (microvessel diameter 4–25 µm) as reported previously in detail (16, 17). In brief, the SDF camera uses green light emitting stroboscopic diodes (540nm) to detect the hemoglobin of passing red blood cells (RBCs). Image acquisition and analysis was performed by GlycoCheck™ Software (Microvascular Health Solutions Inc., Salt Lake City, UT, USA). It detects the dynamic lateral RBC movement into the glycocalyx, which is expressed as the perfused boundary region (PBR, in µm). An altered or degraded glycocalyx allows more RBCs to penetrate deeply toward the endothelial surface, with a consequent increase in the PBR. In every patient, we performed two complete measurements which were averaged to account for spatial heterogeneity of the sublingual microvasculature.
HPSE Activity and HS Competition Assay
The activity of HPSE in EDTA plasma was determined by a heparanase activity assay (Amsbio, Abingdon, UK, Cat# Ra001-02-K) according to the manufacturer’s instructions but using an in-house developed HPSE buffer (18). The dose dependent inhibition of HPSE with HS isolated from bovine kidney was assessed with an in-house developed HPSE activity assay [11]. HS in EDTA plasma samples was quantified by a previously described HS competition assay. Importantly, this assay is specific to HS, therefore the measurement is not affected by the presence of heparin (18).
Quantification of eGC Components and Angiopoietin-2
Plasma levels of Syndecan-1 (Syn-1; Diaclone, Besançon, France, Cat# 950.640.096), hyaluronic acid (HA; Echelon Biosciences Inc., Salt Lake City, UT, USA, Cat# K-1200-) and Angiopoietin-2 (Angpt-2; R&D Systems, Oxford, United Kingdom, Cat# DANG20) were measured using commercially available enzyme-linked immunosorbent assay (ELISA) kits as described previously (17, 19).
Atomic Force Microscopy and Confocal Immunofluorescence Microscopy
Atomic force microscopy using live-cell nano-indentation technique (AFM; Nanoscope V Multimode AFM, Veeco, Mannheim, Germany) and confocal immunofluorescence microscopy (Leica DMI 6000B-CS/TCS SP8 laser confocal microscope, Leica, Wetzlar, Germany) of HS staining were performed on the human umbilical vein endothelial cell line EA.hy926 essentially as described (Amsbio, Ab Heparan sulfate, Cat# 370255-1, RRID : AB_10891554; Jackson ImmunoResearch Labs, Alexa fluor 488 goat anti-mouse IgG antibody, Cat# 115-545-146, RRID : AB_2307324) (20, 21). Cells were grown in DMEM (Gibco™; Cat# 52100047) supplemented with 10% fetal bovine serum (SigmaAldrich; Cat# S0615-100ML) and 1% penicillin/streptomycin (Biochrom; Cat# A2212) at 37°C in a 5% CO2 enriched environment for at least 3 days until reaching confluence. Intensity analysis was performed with ImageJ software (version 1.51p 22, NIH) as previously described (20, 21). The non-anticoagulant N-desulfated re-N-acetylated heparin (NAH), a very potent and well characterized heparanase inhibitor (12), was obtained from Iduron (Cheshire, United Kingdom, Cat# DSH004/Nac).
Statistical Analysis
Values are presented as absolute values with median and inter-quartile range (IQR). The non-parametric Mann-Whitney U test and the Chi-Square test were used to compare parameters between groups. In vitro data are presented as means and standard error of the mean (SEM) unless otherwise stated. Correlation between PBR and eGC thickness was assessed by Spearman correlation coefficient. To account for both the number of observations from a single experiment and the number of experiments, differences in glycocalyx thickness were tested with a nested analysis of variance and Tukey’s post-hoc test. All tests used were two sided, and statistical significance was set at p<0.05. SPSS (IBM Corporation, Armonk, NY, USA, v.26, RRID : SCR_016479) and GraphPad Prism (GraphPad Prism Software Inc., San Diego, CA, USA, v.8.4.3, RRID : SCR_002798) were used for statistical analyses.
Results
COVID-19 patients at the ICU had a median (IQR) Sequential Organ Failure Assessment (SOFA) score of 9 (5 – 15), were predominantly intubated (14/16, 88%) and showed an in-hospital mortality of 38% (6/16). Compared to healthy subjects, COVID-19 patients showed a significantly higher HPSE activity in plasma (Figure 1A). Accordingly, increased levels of HS (the main substrate of HPSE), HA and Syn-1 demonstrated eGC shedding in COVID-19 (Figure 1B, Table 1). This finding was further confirmed by high PBR values (i.e., a thin eGC) measured in COVID-19 patients (Figure 1C and Table 1). All markers of eGC damage correlated with disease severity, as measured by SOFA score (HPSE: R=0.6, P<0.001; HS: R=0.66, P<0.001; HA: R=0.8, P<0.001; Syn-1: R=0.81, P<0.001; PBR: R=0.38, P<0.05).
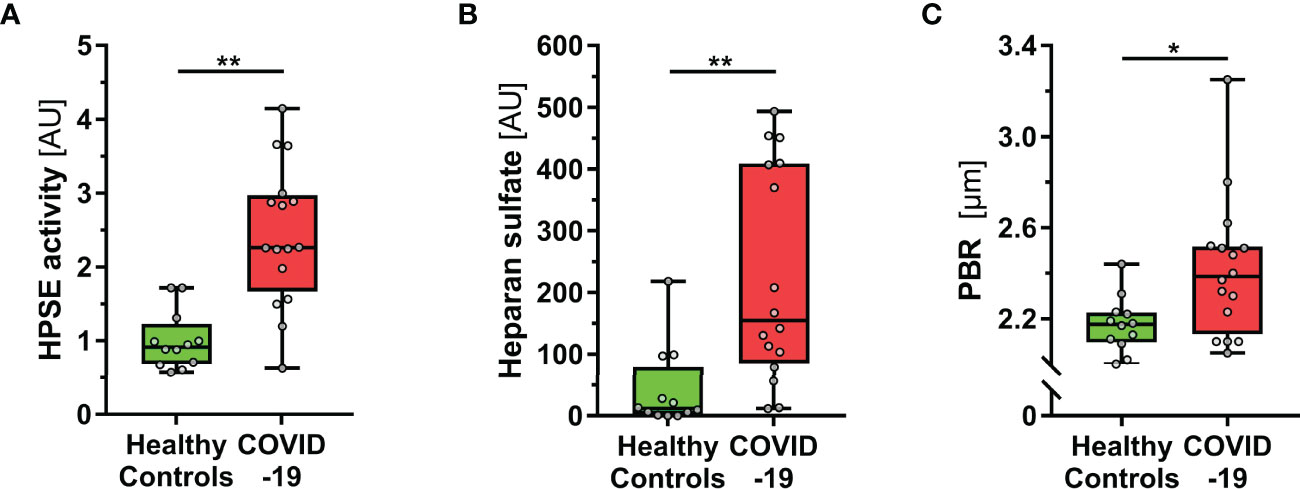
Figure 1 COVID-19 patients show elevated HPSE activity, and damaged eGC in vivo. (A-C) Boxplots showing (A) heparanase (HPSE) activity, (B) heparan sulfate (HS) and (C) perfused boundary region (PBR; an inverse estimate of the sublingual endothelial glycocalyx thickness) in healthy subjects (n = 12) and COVID-19 patients at the ICU (n = 16). Differences between groups were calculated by Mann-Whitney U test. *p < 0.05; **p < 0.001.
To further validate our findings, we exposed endothelial cells (ECs) to randomly selected sterile-filtered sera (5%; diluted in buffer) from 6 COVID-19 patients and 5 healthy subjects for 60 minutes. COVID-19 serum, but not serum from healthy subjects, caused a distinctive decrease of eGC thickness in vitro [-7 (4-14) % vs. -24 (17-28) %, P < 0.01]. The delta changes of eGC thickness measured by AFM in vitro strongly correlated with concordant PBR values (R = -0.66, P = 0.03) obtained by videomicroscopy in vivo (Figure 2A), indicating that the eGC damage seen in COVID-19 can be quantitatively reproduced in cultured ECs.
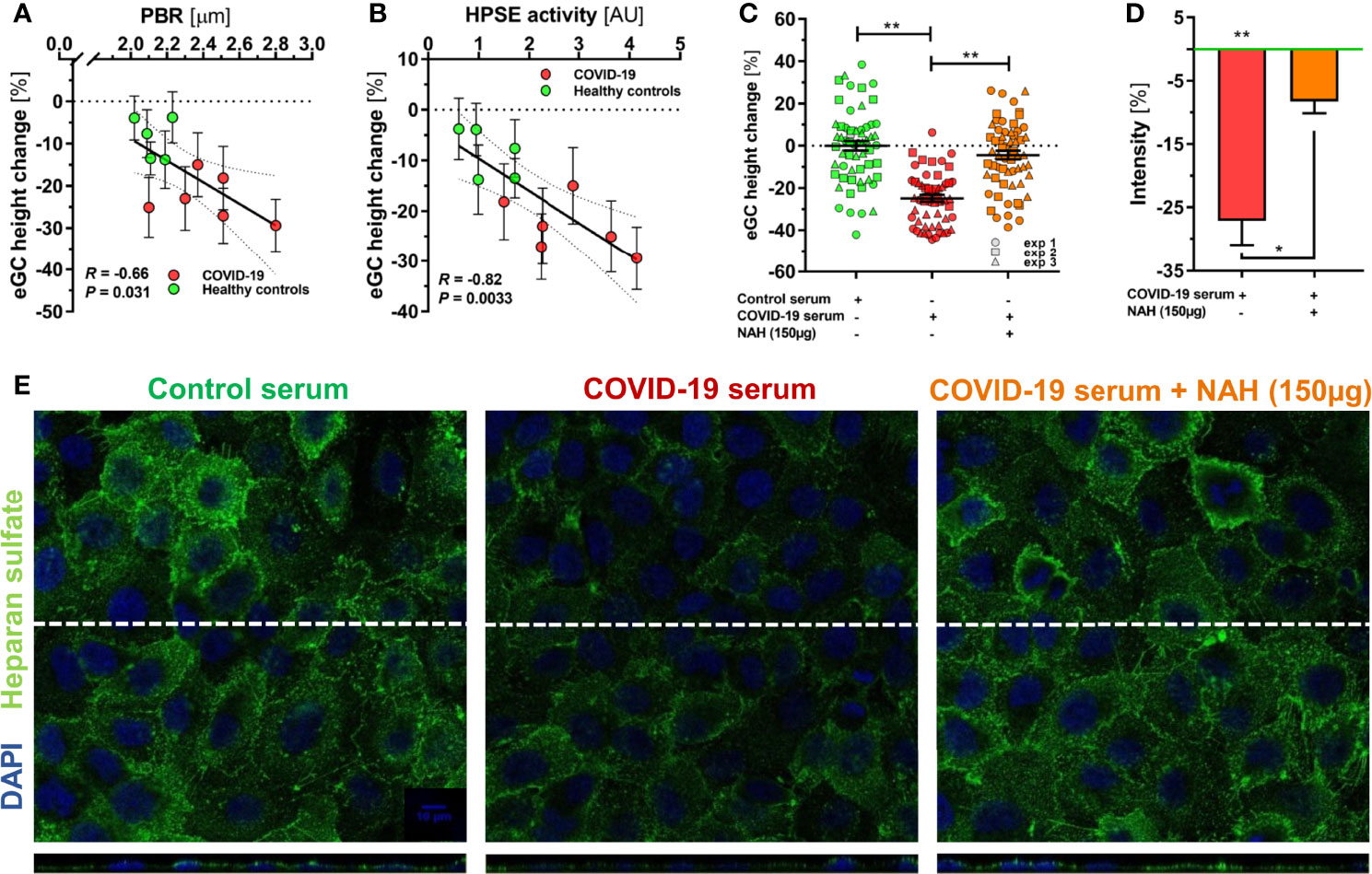
Figure 2 HPSE is a putative mediator of eGC damage in COVID-19. (A-C) Sera from a randomly selected subgroup of 5 healthy controls and 6 COVID-19 patients were sterile-filtered and incubated (5%) on the human umbilical vein endothelial cell line EA.hy926 for 60 min. Endothelial glycocalyx (eGC) thickness was assessed by atomic force microscopy (AFM) using a dedicated nano-indentation protocol. Scatter dot plot showing the association between AFM-derived eGC (in vitro) decline and corresponding (A) PBR-values (in vivo) and (B) HPSE activity for the individuals from the subgroup. Each dot represents the mean ± SEM (standard error of mean) of two independent AFM experiments (consisting of ≥ 4 indentation curves in each of ≥ 8 different cells) for each individual serum. Incubation without human serum served as control. Correlation was assessed by Spearman correlation coefficient. (C) Dot plots from three independent AFM experiments (pooled serum from subgroups) showing values with mean ± SEM. Each dot represents the mean of ≥ 4 indentation curves per cell. Heparanase was blocked by N-desulfated re-N-acetylated heparin (NAH; 150 µg). Differences between groups were calculated with nested ANOVA and Tukey’s post-hoc test. Intensity analysis of heparan sulfate-stained EA.hy926 cells (D) and representative immunofluorescence images (E) after treatment with 5% control serum or COVID-19 serum ± NAH (150 µg) for 60 min. Values are normalized to control serum (zero line) and differences between groups were assessed with nested ANOVA and Tukey’s post-hoc test. Data are presented as mean ± SEM. *p < 0.05; **p < 0.001.
Delta changes of eGC thickness measured by AFM also correlated closely with the respective plasma HPSE activity in the samples (R = 0.82, P = 0.003) (Figure 2B). Competitive inhibition of heparanase by the nonanticoagulant heparin fragment NAH completely prevented the decline in eGC thickness in response to COVID-19 serum (Figure 2C). Similarly, immunofluorescence imaging of the eGC showed a marked decrease in HS positivity and surface coverage in ECs incubated with COVID-19 serum, whereas COVID-19 serum supplemented with NAH caused only a very slight loss in staining intensity and coverage compared to incubation with serum from healthy controls (Figures 2D, E).
In the total cohort, HPSE correlated well with inflammatory mediators, such as C-reactive protein (R = 0.78, P < 0.001), interleukin (IL)-6 (R = 0.63, P < 0.001), and Angpt-2 (R = 0.61, P < 0.001) as well as the eGC marker Syn-1 (R = 0.71, P < 0.001). Surprisingly, however, HPSE activity did not correlate with the PBR (R = 0.11, P = 0.58) or plasma HS (R = 0.34, P = 0.08).
To understand this finding in more detail, we attempted to roughly compensate for the presence of heparin [which dose dependently blocks HPSE (18)] by using activated partial thromboplastin time (aPTT) as an adjustment. The aPTT was significantly prolonged in COVID-19 (Table 1) and correlated positively with HPSE activity (R = 0.58, P < 0.001), suggesting that the true HPSE activity in severely ill COVID-19 patients could be even higher than the values actually measured. Adjusting HPSE activity for aPTT (normalized HPSE/normalized aPTT quotient) indeed revealed a moderate association with the PBR (R = 0.44, P = 0.025).
Finally, we investigated the relationship between HPSE and HS in more detail. Interestingly, separate analysis of the two groups revealed contrasting regression slopes, suggesting that excessive generation of HS fragments in severe COVID-19 (which was not present in controls) may have partially blocked HPSE activity in COVID-19 patients (Figure 3A). Additional in vitro inhibition experiments with HS isolated from bovine kidney confirmed a dose dependent inhibition of HPSE at concentrations between 0 and 6.25 µg/ml; nearly full inhibition of heparanase was reached at a concentration of 6.25 µg/ml HS (Figure 3B).
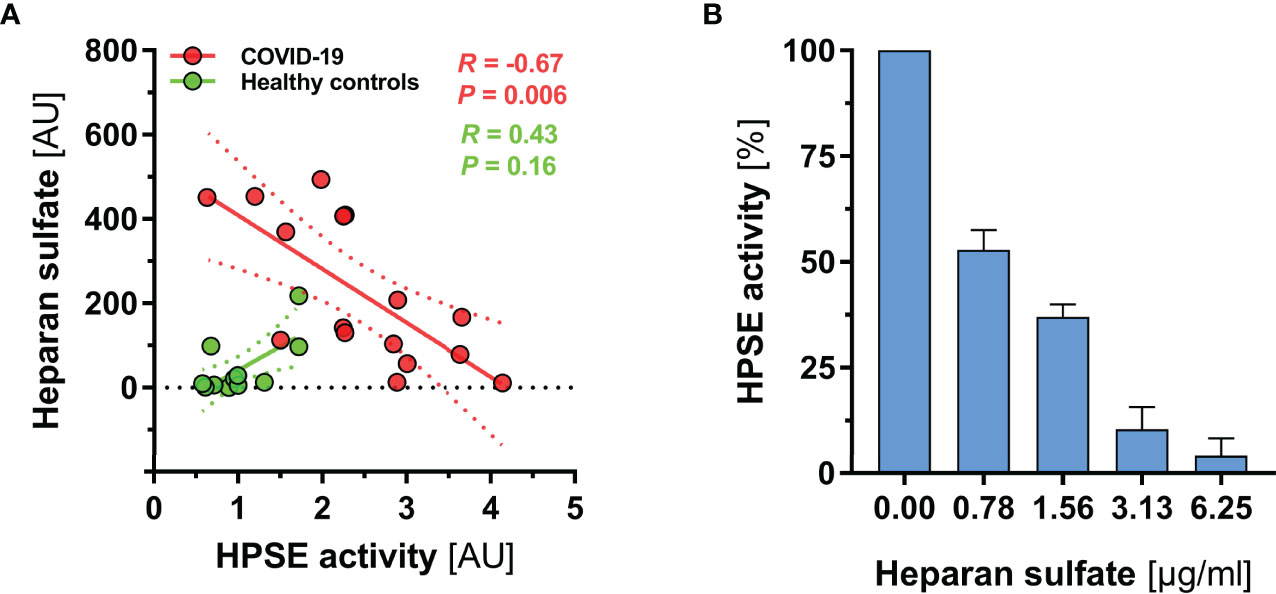
Figure 3 HS fragments in severe COVID-19 may partially block HPSE activity. (A) Scatter dot plot showing regression slopes of HPSE activity vs. HS plasma concentration in healthy subjects (n = 12) and COVID-19 patients (n = 16), respectively. (B) Bar charts showing percentage decrease of HPSE activity with increasing amounts of HS (isolated from bovine kidney) in three independent experiments. For this experiment, recombinant human HPSE was used in a concentration of 150 ng/ml. Data are presented as mean ± SEM.
Discussion
Since the beginning of the COVID-19 pandemic, evidence has emerged that COVID-19 is a vascular rather than purely respiratory illness (1). Here we show that HPSE, the only enzyme in mammals that degrades HS chains from HS proteoglycans of the glycocalyx, plays a key role in mediating eGC damage in COVID-19.
HPSE is predominantly released by vascular endothelium, macrophages, and platelets. Its expression is up-regulated in endothelial cells by several factors, such as reactive oxygen species, inflammatory cytokines, high glucose, and advanced glycosylation products (22–25). Two previous COVID-19 studies measured HPSE antigen levels (26, 27) and two others determined enzymatic HPSE activity (18, 28). All but one study reported significantly elevated HPSE levels in COVID-19 patients. This finding therefore fits well with our previous data showing that sublingual eGC thickness (PBR) appears to be an appropriate risk marker for hospital mortality in COVID-19 (4). The in vitro experiments in our current study show that HPSE is a critical factor for eGC damage, as glycocalyx thinning correlated with HPSE activity and could be completely prevented by HPSE inhibition. The latter finding is even more surprising because the protective effect was achieved despite the presence of numerous cytokines and mediators in the patients’ sera (Table 1). This makes HPSE a promising target for intervention in COVID-19.
The possible involvement of HPSE in COVID-19 pathology was predicted already at the beginning of the pandemic long before initial studies on the subject (22, 29). It was speculated at the time that HPSE might contribute to the removal of HS chains from cell surfaces, facilitating virus release from host cells - a mechanism that has been well described for herpes simplex virus-1 (30, 31). Accordingly, enzymatic removal of HS from the eGC by HPSE is probably not the only underlying mechanism. For example, HPSE can induce matrix metalloproteases, which in turn cut transmembrane heparan sulfate proteoglycans such as syndecan-1 and thus further enhance eGC damage (31). Consistent with this assessment, our patients showed markedly elevated blood levels of hyaluronan and syndecan-1. The finding that HPSE activity in healthy individuals and COVID-19 patients was inversely correlated with HS concentration suggests that a minimum amount of HPSE is required before systemic eGC degradation begins. If so, HPSE would be more likely to be a late marker of disease severity. However, to disrupt the interaction of Sars-CoV-2 with the host cell, early blockade of HPSE could be beneficial.
In bacterial sepsis, the potential of HPSE and eGC as future pharmacological targets has already been highlighted (14, 15, 32). In detail, prevention of eGC damage by inhibition of HPSE significantly abolished vascular hyperpermeability and subsequent lung injury in murine endotoxemia (12). Using AFM we could show earlier, that unfractionated heparin (UFH), which also saturates HPSE, completely abolished the HPSE-induced decline of eGC thickness in freshly isolated rat aorta (20). Furthermore, the heparin fragment NAH prevented thinning of the eGC on human ECs induced by serum from sepsis patients in vitro (21). Based on these data, it would be very reasonable to assume that blocking HPSE may also be an effective approach to protecting eGC in COVID-19. Indeed, by western blotting of EC lysates, Potje et al. showed that low molecular weight heparin (LMWH) reduced the loss of glycocalyx components induced by plasma from COVID-19 patients (26). Our translational study comparing, for the first time, HPSE activity with the corresponding glycocalyx dimensions in vivo and in vitro in matched pairs strongly confirms and extends this exciting finding.
At the clinical level, the administration of heparin and/or LMWH has been shown to provide a significant survival benefit in both entities (33–35). Hence, apart from the anticoagulant effect of heparin the non-anticoagulant and, among these, especially anti-inflammatory properties should not be neglected and deserve closer attention (36). It is conceivable that this protective effect could be partly due to an off-target effect of heparin – i.e., the inhibition of HPSE. Further studies, ideally with accompanying HPSE measurement and sublingual video microscopy, are needed to confirm this assumption. However, the paradoxical decrease in HPSE activity in the COVID-19 patients with the highest HS concentrations certainly complicates any interpretation of HPSE activity in plasma samples. It is interesting to speculate that this could be a feedback loop whose aim is to keep HPSE activity within a certain range and to dampen its deleterious effects on the eGC. However, our data emphasize the importance of analyzing the HPSE in relation to the HS (and also to UFH/LMWH) in future studies. Of note, the investigation of heparins or heparin mimetics in COVID-19 appears particularly promising as these drugs hold the potential to counteract disease onset and progression in various ways. Not only the inhibition of endothelial HPSE, as presented in the current study, and HPSE of viral origin, as mentioned above, emerge as relevant target structures for heparins. Recent studies revealed HS/heparin-related binding sites on the SARS-CoV-2 spike protein and co-binding of HS and angiotensin-converting enzyme 2 seems to be necessary for docking to cellular surfaces. Heparins, on the other hand, were capable to potently block spike protein binding, probably due to blocking these specific binding sites (37). Consequently, not only heparins but also heparan sulfate mimetics – so far especially investigated in context of cancer therapy – come to interest as potential drugs in future COVID-19 treatment studies (38).
As our study is primarily hypothesis-generating, some limitations should be noted. First, the sample size of this proof-of-concept study was rather small. However, our dataset includes a wide range of glycocalyx and inflammatory markers, as well as a novel microscopy method to calculate the PBR in vivo. Although the PBR is not measured directly but estimated based on the radial displacement of red blood cells, this validated method is highly reliable (14, 16, 17, 39). Differences in PBR between patients and controls matched well with values obtained by Stahl et al. in another COVID-19 ICU cohort (27). Second, the use of aPTT as a measure of normalization of HPSE activity is certainly subject to its own variance. It would have been better to use either an inhibitor constant of heparin/LMWH or anti-Xa activity for correction to derive the active HPSE level. As we do not have citrate retain samples, a more accurate correction for heparin/LMWH effect was not possible in this data set. Unfortunately, the type, dose and application route of the heparins used were very heterogeneous, so that we were unable to convert these data into meaningful equivalent concentrations. Third, the number of serum samples used for the in vitro studies is limited because the AFM technique is sophisticated and time-consuming.
In conclusion, our translational study indicates that HPSE is a putative mediator of endothelial glycocalyx damage in COVID-19. Further studies are needed to clarify whether the benefit of heparin administration in COVID-19 is due to an eGC-protective off-target effect.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of the General Medical Council Westfalen-Lippe and the WWU Münster, Germany. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
CD designed and conducted the in vitro experiments, analyzed the data, prepared the figures and contributed to the manuscript; AR and IO enrolled patients, performed sublingual video microscopy, performed ELISA assays and contributed to the manuscript; MR performed AFM experiments and revised the manuscript; BB, RS, and JV performed the HPSE activity and HS competition assays and contributed to the manuscript; AL, HP, and WL advised on the study design, discussed the findings and revised the manuscript; PK had the initial idea, supervised the study, recruited and coordinated participating centers, contributed to the analysis and figures, and drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the German Research Foundation (rotational position of KFO 342 – ZA428/18-1 to CD and AR; KU 2873/3-1 to PK) and the fund Innovative Medical Research of the University of Münster Medical School (I-RO221907 to AR). The funding sources had no role in study design, data collection and analysis, the decision to publish, or preparation of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Jan Sackarnd, Phil-Robin Tepasse, Stephan Braune, Ulrich Göbel, Gerold Thölking and Andreas Gröschel for their help in recruiting patients, Manfred Fobker for assistance in sample processing and Hans Vink for assistance with regard to the Glycocheck® system. We would like to acknowledge the support of the Open Access Publication Fund of the University of Münster.
References
1. Smadja DM, Mentzer SJ, Fontenay M, Laffan MA, Ackermann M, Helms J, et al. Covid-19 Is a Systemic Vascular Hemopathy: Insight for Mechanistic and Clinical Aspects. Angiogenesis (2021) 24(4):755–88. doi: 10.1007/s10456-021-09805-6
2. Pons S, Fodil S, Azoulay E, Zafrani L. The Vascular Endothelium: The Cornerstone of Organ Dysfunction in Severe Sars-Cov-2 Infection. Crit Care (2020) 24(1):353. doi: 10.1186/s13054-020-03062-7
3. Emadi-Baygi M, Ehsanifard M, Afrashtehpour N, Norouzi M, Joz-Abbasalian Z. Corona Virus Disease 2019 (Covid-19) as a System-Level Infectious Disease With Distinct Sex Disparities. Front Immunol (2021) 12:778913. doi: 10.3389/fimmu.2021.778913
4. Rovas A, Osiaevi I, Buscher K, Sackarnd J, Tepasse PR, Fobker M, et al. Microvascular Dysfunction in Covid-19: The Mystic Study. Angiogenesis (2021) 24(1):145–57. doi: 10.1007/s10456-020-09753-7
5. Reitsma S, Slaaf DW, Vink H, van Zandvoort MA, oude Egbrink MG. The Endothelial Glycocalyx: Composition, Functions, and Visualization. Pflugers Arch (2007) 454(3):345–59. doi: 10.1007/s00424-007-0212-8
6. van Haaren PMA, VanBavel E, Vink H, Spaan JAE. Localization of the Permeability Barrier to Solutes in Isolated Arteries by Confocal Microscopy. Am J Physiol Heart Circulatory Physiol (2003) 285(6):H2848–H56. doi: 10.1152/ajpheart.00117.2003
7. Donati A, Damiani E, Domizi R, Romano R, Adrario E, Pelaia P, et al. Alteration of the Sublingual Microvascular Glycocalyx in Critically Ill Patients. Microvasc Res (2013) 90:86–9. doi: 10.1016/j.mvr.2013.08.007
8. Alphonsus CS, Rodseth RN. The Endothelial Glycocalyx: A Review of the Vascular Barrier. Anaesthesia (2014) 69(7):777–84. doi: 10.1111/anae.12661
9. Mulivor AW, Lipowsky HH. Role of Glycocalyx in Leukocyte-Endothelial Cell Adhesion. Am J Physiol Heart Circ Physiol (2002) 283(4)::H1282–91. doi: 10.1152/ajpheart.00117.2002
10. Curry FE, Adamson RH. Endothelial Glycocalyx: Permeability Barrier and Mechanosensor. Ann BioMed Eng (2012) 40(4):828–39. doi: 10.1007/s10439-011-0429-8
11. Salmon AH, Ferguson JK, Burford JL, Gevorgyan H, Nakano D, Harper SJ, et al. Loss of the Endothelial Glycocalyx Links Albuminuria and Vascular Dysfunction. J Am Soc Nephrol (2012) 23(8):1339–50. doi: 10.1681/ASN.2012010017
12. Schmidt EP, Yang Y, Janssen WJ, Gandjeva A, Perez MJ, Barthel L, et al. The Pulmonary Endothelial Glycocalyx Regulates Neutrophil Adhesion and Lung Injury During Experimental Sepsis. Nat Med (2012) 18(8):1217–23. doi: 10.1038/nm.2843
13. Padberg JS, Wiesinger A, di Marco GS, Reuter S, Grabner A, Kentrup D, et al. Damage of the Endothelial Glycocalyx in Chronic Kidney Disease. Atherosclerosis (2014) 234(2):335–43. doi: 10.1016/j.atherosclerosis.2014.03.016
14. Drost CC, Rovas A, Kumpers P. Protection and Rebuilding of the Endothelial Glycocalyx in Sepsis - Science or Fiction? Matrix Biol Plus (2021) 12:100091. doi: 10.1016/j.mbplus.2021.100091
15. Uchimido R, Schmidt EP, Shapiro NI. The Glycocalyx: A Novel Diagnostic and Therapeutic Target in Sepsis. Crit Care (2019) 23(1):16. doi: 10.1186/s13054-018-2292-6
16. Rovas A, Lukasz AH, Vink H, Urban M, Sackarnd J, Pavenstadt H, et al. Bedside Analysis of the Sublingual Microvascular Glycocalyx in the Emergency Room and Intensive Care Unit - The Glyconurse Study. Scand J Trauma Resusc Emerg Med (2018) 26(1):16. doi: 10.1186/s13049-018-0483-4
17. Rovas A, Seidel LM, Vink H, Pohlkotter T, Pavenstadt H, Ertmer C, et al. Association of Sublingual Microcirculation Parameters and Endothelial Glycocalyx Dimensions in Resuscitated Sepsis. Crit Care (2019) 23(1):260. doi: 10.1186/s13054-019-2542-2
18. Buijsers B, Yanginlar C, de Nooijer A, Grondman I, Maciej-Hulme ML, Jonkman I, et al. Increased Plasma Heparanase Activity in Covid-19 Patients. Front Immunol (2020) 11:575047. doi: 10.3389/fimmu.2020.575047
19. Kumpers P, Lukasz A, David S, Horn R, Hafer C, Faulhaber-Walter R, et al. Excess Circulating Angiopoietin-2 Is a Strong Predictor of Mortality in Critically Ill Medical Patients. Crit Care (2008) 12(6):R147. doi: 10.1186/cc7130
20. Wiesinger A, Peters W, Chappell D, Kentrup D, Reuter S, Pavenstadt H, et al. Nanomechanics of the Endothelial Glycocalyx in Experimental Sepsis. PloS One (2013) 8(11):e80905. doi: 10.1371/journal.pone.0080905
21. Drost CC, Rovas A, Kusche-Vihrog K, Van Slyke P, Kim H, Hoang VC, et al. Tie2 Activation Promotes Protection and Reconstitution of the Endothelial Glycocalyx in Human Sepsis. Thromb Haemost (2019) 119(11):1827–38. doi: 10.1055/s-0039-1695768
22. Kinaneh S, Khamaysi I, Karram T, Hamoud S. Heparanase as a Potential Player in Sars-Cov-2 Infection and Induced Coagulopathy. Biosci Rep (2021) 41(7). doi: 10.1042/BSR20210290
23. Pape T, Hunkemoller AM, Kumpers P, Haller H, David S, Stahl K. Targeting the “Sweet Spot” in Septic Shock - a Perspective on the Endothelial Glycocalyx Regulating Proteins Heparanase-1 and -2. Matrix Biol Plus (2021) 12:100095. doi: 10.1016/j.mbplus.2021.100095
24. Masola V, Greco N, Gambaro G, Franchi M, Onisto M. Heparanase as Active Player in Endothelial Glycocalyx Remodeling. Matrix Biol Plus (2022) 13:100097. doi: 10.1016/j.mbplus.2021.100097
25. Vlodavsky I, Ilan N, Sanderson RD. Forty Years of Basic and Translational Heparanase Research. Adv Exp Med Biol (2020) 1221:3–59. doi: 10.1007/978-3-030-34521-1_1
26. Potje SR, Costa TJ, Fraga-Silva TFC, Martins RB, Benatti MN, Almado CEL, et al. Heparin Prevents in Vitro Glycocalyx Shedding Induced by Plasma From Covid-19 Patients. Life Sci (2021) 276:119376. doi: 10.1016/j.lfs.2021.119376
27. Stahl K, Gronski PA, Kiyan Y, Seeliger B, Bertram A, Pape T, et al. Injury to the Endothelial Glycocalyx in Critically Ill Patients With Covid-19. Am J Respir Crit Care Med (2020) 202(8):1178–81. doi: 10.1164/rccm.202007-2676LE
28. Queisser KA, Mellema RA, Middleton EA, Portier I, Manne BK, Denorme F, et al. Covid-19 Generates Hyaluronan Fragments That Directly Induce Endothelial Barrier Dysfunction. JCI Insight (2021) 6(17). doi: 10.1172/jci.insight.147472
29. Tiwari V, Beer JC, Sankaranarayanan NV, Swanson-Mungerson M, Desai UR. Discovering Small-Molecule Therapeutics Against Sars-Cov-2. Drug Discovery Today (2020) 25(8):1535–44. doi: 10.1016/j.drudis.2020.06.017
30. Hadigal SR, Agelidis AM, Karasneh GA, Antoine TE, Yakoub AM, Ramani VC, et al. Heparanase Is a Host Enzyme Required for Herpes Simplex Virus-1 Release From Cells. Nat Commun (2015) 6:6985. doi: 10.1038/ncomms7985
31. Hadigal S, Koganti R, Yadavalli T, Agelidis A, Suryawanshi R, Shukla D. Heparanase-Regulated Syndecan-1 Shedding Facilitates Herpes Simplex Virus 1 Egress. J Virol (2020) 94(6). doi: 10.1128/JVI.01672-19
32. Sullivan RC, Rockstrom MD, Schmidt EP, Hippensteel JA. Endothelial Glycocalyx Degradation During Sepsis: Causes and Consequences. Matrix Biol Plus (2021) 12:100094. doi: 10.1016/j.mbplus.2021.100094
33. Lavinio A, Ercole A, Battaglini D, Magnoni S, Badenes R, Taccone FS, et al. Safety Profile of Enhanced Thromboprophylaxis Strategies for Critically Ill Covid-19 Patients During the First Wave of the Pandemic: Observational Report From 28 European Intensive Care Units. Crit Care (2021) 25(1):155. doi: 10.1186/s13054-021-03543-3
34. Poli D, Antonucci E, Ageno W, Prandoni P, Palareti G, Marcucci R, et al. Low in-Hospital Mortality Rate in Patients With Covid-19 Receiving Thromboprophylaxis: Data From the Multicentre Observational Start-Covid Register. Intern Emerg Med (2022) 17(4):1013–21. doi: 10.1007/s11739-021-02891-w
35. Wang C, Chi C, Guo L, Wang X, Guo L, Sun J, et al. Heparin Therapy Reduces 28-Day Mortality in Adult Severe Sepsis Patients: A Systematic Review and Meta-Analysis. Crit Care (2014) 18(5):563. doi: 10.1186/s13054-014-0563-4
36. Mohamed S, Coombe DR. Heparin Mimetics: Their Therapeutic Potential. Pharmaceut (Basel) (2017) 10(4). doi: 10.3390/ph10040078
37. Clausen TM, Sandoval DR, Spliid CB, Pihl J, Perrett HR, Painter CD, et al. Sars-Cov-2 Infection Depends on Cellular Heparan Sulfate and Ace2. Cell (2020) 183(4):1043–57 e15. doi: 10.1016/j.cell.2020.09.033
38. Schuurs ZP, Hammond E, Elli S, Rudd TR, Mycroft-West CJ, Lima MA, et al. Evidence of a Putative Glycosaminoglycan Binding Site on the Glycosylated Sars-Cov-2 Spike Protein N-Terminal Domain. Comput Struct Biotechnol J (2021) 19:2806–18. doi: 10.1016/j.csbj.2021.05.002
Keywords: COVID-19, heparin, heparanase (HPSE), videomicroscopy, endothelial glycocalyx (EG)
Citation: Drost CC, Rovas A, Osiaevi I, Rauen M, van der Vlag J, Buijsers B, Salmenov R, Lukasz A, Pavenstädt H, Linke WA and Kümpers P (2022) Heparanase Is a Putative Mediator of Endothelial Glycocalyx Damage in COVID-19 – A Proof-of-Concept Study. Front. Immunol. 13:916512. doi: 10.3389/fimmu.2022.916512
Received: 09 April 2022; Accepted: 17 May 2022;
Published: 10 June 2022.
Edited by:
Deirdre R. Coombe, Curtin University, AustraliaReviewed by:
Umesh R. Desai, Virginia Commonwealth University, United StatesTejabhiram Yadavalli, University of Illinois at Chicago, United States
Copyright © 2022 Drost, Rovas, Osiaevi, Rauen, van der Vlag, Buijsers, Salmenov, Lukasz, Pavenstädt, Linke and Kümpers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Philipp Kümpers, cGhpbGlwcC5rdWVtcGVyc0B1a211ZW5zdGVyLmRl