 Ying Meng1*
Ying Meng1* Amber Kautz2
Amber Kautz2- 1School of Nursing, University of Rochester, Rochester, NY, United States
- 2Department of Public Health Sciences, University of Rochester Medical Center, Rochester, NY, United States
Background: Eating behaviors contribute to disproportionate energy intake and are linked to the development of obesity. Animal studies support the role of inflammatory cytokines and chemokines in the regulation of obesity-related eating behaviors and offer a potential target to combat obesity through the modulation of inflammation. However, more complex eating behaviors are present in humans, and their relationships with immune/inflammation markers are unclear. The present study reviewed current literature to synthesize the evidence on the association of immune/inflammation markers with obesity-related eating behaviors in humans.
Methods: A systematic search of three electronic databases yielded 811 articles, of which 11 met the inclusion criteria.
Results: The majority of the included studies (91%) were either case-control or cross-sectional studies. A variety of immune/inflammation markers and obesity-related eating behaviors have been assessed in the chosen studies. Three out of four studies identified a positive relationship between C-reactive protein (CRP)/high-sensitivity CRP and loss of control eating. Other inflammatory markers that potentially have a positive relationship with obesity-related eating behaviors include fractalkine and fibrinogen. Additionally, immune molecules, including interferon gamma (INF-γ), interleukin (IL)-7, IL-10, and α-melanocyte-stimulating hormone-reactive immunoglobulin G (α-MSH/IgG) immune complex, may have negative associations with obesity-related eating behaviors. However, most findings were identified by single studies.
Conclusion: Limited studies have been conducted in humans. Current evidence indicates a potential bi-directional relationship between inflammatory/immune markers and obesity-related eating behaviors. Additional studies with sophisticated research design and comprehensive theoretical models are warranted to further delineate the relationship between immune/inflammation markers and obesity-related eating behaviors.
Introduction
The prevalence of obesity has continued to rise. Over 42% of adults and 19% of children are currently affected by obesity in the United States (1, 2). Eating behaviors, such as eating in the absence of hunger, disinhibited eating, and excessive intake of dietary fat, contribute to disproportionate energy intake and have been linked to the development of obesity (3). Strategies targeting obesity-related eating behaviors may be a key approach to alleviate the obesity epidemic, and comprehensive understanding of the underlying mechanisms is crucial to develop effective interventions.
A growing body of evidence has indicated that immune and inflammatory molecules play a role in the regulation of eating behaviors (4–7). It has long been observed that various disease states can lead to reduced appetite and food intake (8). Several cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1α, IL-1β, IL-6, IL-7, IL-18, have been found to suppress food intake in animals and potentially in humans (4, 7, 9–12). In addition to suppressing appetite, alterations in immunological function may also be involved in the pathogenesis or perpetuation of obesity-related eating behaviors. In rodent models, deficiency in IL-6, IL-1 type I receptor, and IL-18 resulted in hyperphagia and obesity (6, 13, 14). Intracerebroventricular administration of IL-6 and IL-1β reduced sucrose preference and food intake (7, 15). Alleviation of hypothalamic inflammation and deficiency in interferon gamma (IFN-γ) also reduced food intake and diet-induced obesity (16–18). Hence, it is hypothesized that dysregulation of the immune system may initiate or exacerbate appetite and food reward signals and contribute to the development or perpetuation of obesity-related maladaptive eating behaviors.
Previous studies in animals supported the role of cytokines in the regulation of obesity-related eating behaviors (6, 7, 13–18). However, there has been limited research exploring the involvement of immune molecules in modulating obesity-related eating behaviors in humans which are more complex than rodent models (19). Therefore, this review was conducted to synthesize the evidence on the association between immune markers and obesity-related eating behaviors in humans.
Methods
To identify relevant publications, systematic searches were conducted using the following electronic databases: PubMed, Web of Science, and CINAHL. We used a variety of search terms including “eating” or “feeding” combined with “obesity” then crossed with “immune” or “inflammation” or “cytokine”. The searches were limited to “human” studies, and articles were restricted to English language. Primary research studies published through May, 2021 were included if they fulfilled the following criteria: 1) the relationship between immune/inflammatory molecules and eating behaviors related to obesity was examined; 2) to expand the number of articles included, articles comparing anorexia nervosa (AN) with obesity were also included, and AN was considered an eating behavior opposite of those related to obesity; 3) articles examining brain regions related to eating behavior regulation were included, and the brain regions were used as proxies for eating behaviors. Studies were excluded if 1) they discussed the links between inflammatory markers with fasting or intake of single foods, nutrients, or dietary patterns; or 2) they examined local infections, such as gastric infection by Helicobacter pylori; or 3) they assessed leptin instead of inflammatory markers, as leptin is commonly classified as an adipokine; or 4) they were conducted in patients with cancers.
Figure 1 presents the flow of the literature search. A total of 811 articles were screened. Two reviewers (YM and AK) independently screened all titles and abstracts to identify potential articles (n=49). Full texts were extracted and reviewed by the two reviewers. Of these articles, ten articles fulfilled the eligibility criteria. References of the identified articles were further screened. One additional article was identified. A total of 11 articles were included in this review.
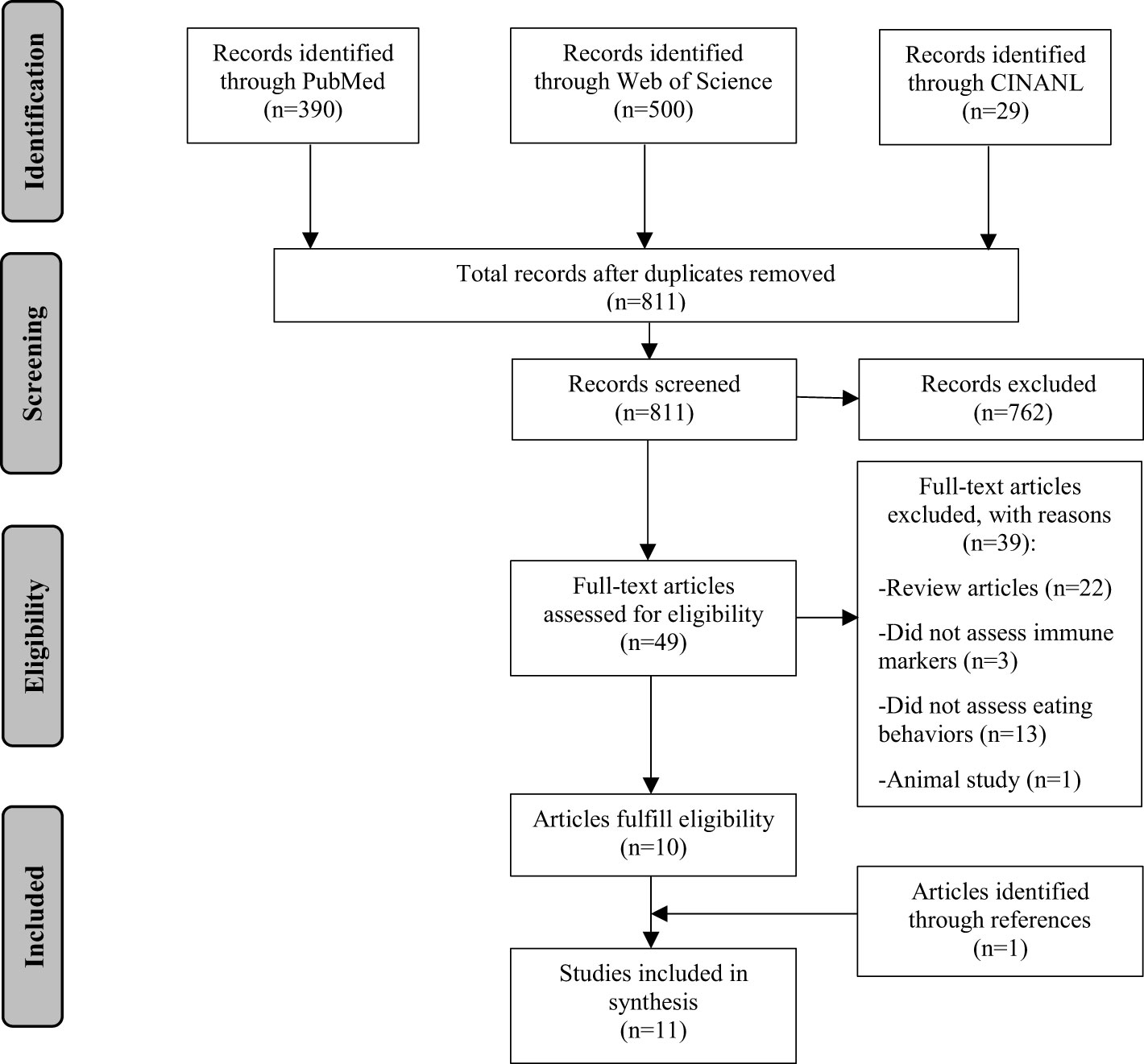
Figure 1 Literature search flow chart.
Results
Study characteristics
Of the eleven included articles (Table 1), five studies assessed a variety of obesity-related eating behaviors, including loss of control eating, hunger, binge eating, and emotional eating. Five studies compared eating disorders with obesity. One study examined brain regions related to the regulation of food intake. A variety of cytokines, chemokines, immunoglobulins and inflammatory markers have been assessed in these studies. With regards to study designs, six studies were case-control studies, four studies were cross-sectional studies, and one study utilized a quasi-experimental design. Six studies were conducted in Europe, three in the USA, one in Brazil, and one in China. The sample sizes ranged from 32 to 194. Five studies included more than or equal to100 participants. Four studies recruited children and adolescents, and the other studies recruited adults or a mixture of adolescents and adults. Five studies only conducted assessments with female participants.
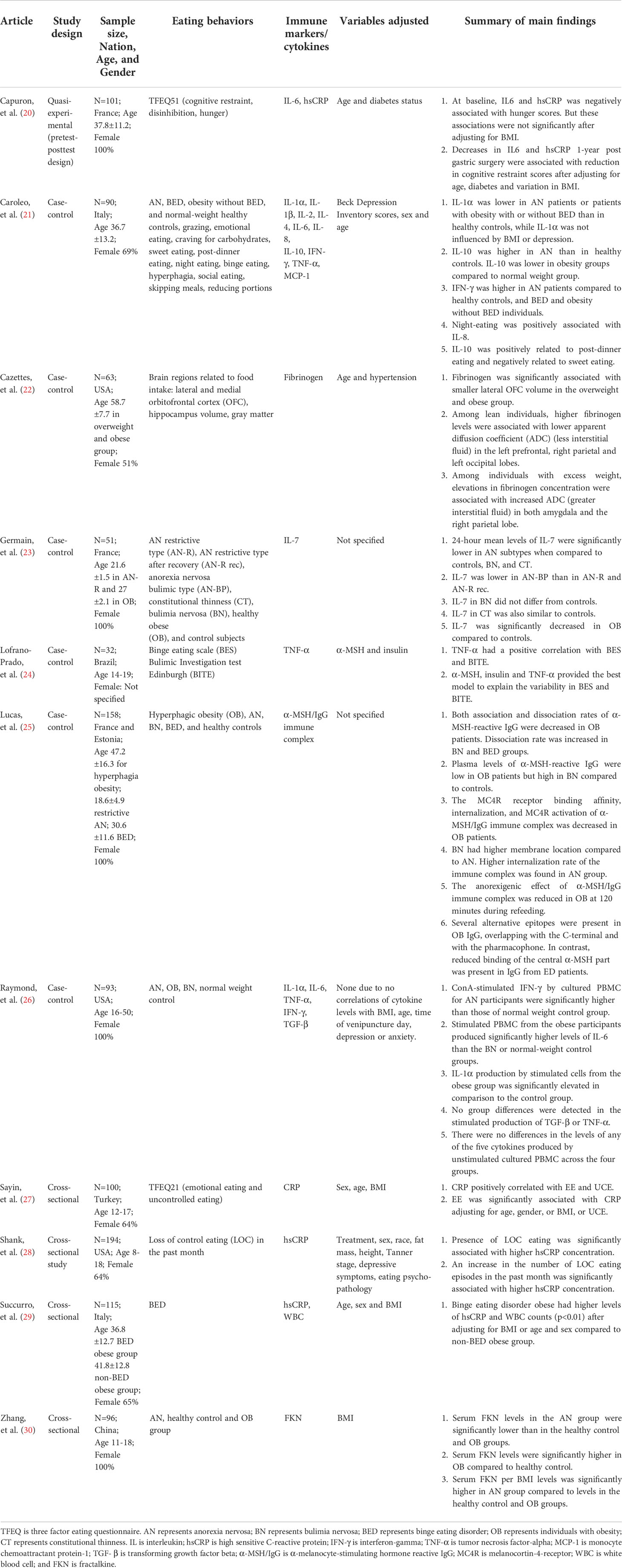
Table 1 Included studies.
Descriptive synthesis of main findings
Four studies examined the relationship between the C-reactive protein (CRP) or high sensitivity-CRP (hsCRP) and eating behaviors, including uncontrolled eating, binge eating, hunger, emotional eating and cognitive restraint eating (Table 2). Two of these studies with relatively large sample sizes reported positive associations of hsCRP with loss of control eating or binge eating after controlling for covariates (28, 29). Particularly, these two studies adjusted for adiposity including fat mass or body mass index (BMI) in their models. Another study also reported a positive correlation between CRP and uncontrolled eating (27). Capuron et al. (20), however, did not find a significant association between hsCRP and disinhibition (20). The eating behavior measurements varied among studies, which may account for the inconsistency of findings. Other than disinhibited eating, Sayin et al. (27) also reported a positive association between CRP and emotional eating after controlling for BMI. Capuron et al. (20) found that reduction in hsCRP one-year post bariatric surgery was associated with decrease in cognitive restraint eating.

Table 2 Results based on individual immune/inflammation markers.
Two studies compared IL-1α levels among participants with eating disorders, obesity and normal weight controls. Caroleo et al. (21) found significant differences among subjects with AN, binge eating disorder (BED), obesity, and normal weight controls (21). Controls had relatively higher serum IL-1α levels, but pair-wise comparisons between groups were not performed. Raymond et al. (26) found that IL-1α production by stimulated peripheral blood mononuclear cell (PBMC) from individuals with obesity was higher relative to normal weight controls, but there were no differences in IL-1α production by unstimulated PBMC among individuals with different conditions (26). INF-γ was also assessed in these two studies. Both studies found higher serum INF-γ or INF-γ production by stimulated PBMC in subjects with AN compared to normal weight controls.
IL-6 was assessed in three studies. Capuron et al. (20) found that a decrease in IL-6 one-year post bariatric surgery was associated with reduction in cognitive restraint eating accounting for variations in BMI. Raymond et al. (26) found that IL-6 production by stimulated PBMC from individuals with obesity was higher than normal weight controls, but no differences were found in IL-6 production by unstimulated PBMC. Caroleo et al. (21), on the other hand, did not identify significant differences in serum IL-6 levels among individuals with eating disorders, obesity, or normal weight.
Tumor necrosis factor (TNF)-α was assessed in three studies (21, 24, 26). Only one study with a relatively smaller sample size (N=32) found positive associations between serum TNF-α levels with binge eating scores and bulimic investigation test Edinburgh scores in a group of adolescents (24). The other two studies (N≥90) did not find significant associations between serum TNF-α levels or production by stimulated PBMC among individuals with eating disorders, obesity, or normal weight.
Other immune/inflammatory markers, including IL-1β, IL-2, IL-4, IL-7, IL-8, IL-10, transforming growth factor beta (TGF-β), monocyte chemoattractant protein 1 (MCP-1), fibrinogen, fractalkine, α-melanocyte-stimulating hormone-reactive immunoglobulin G (α-MSH/IgG) complex, and white blood cell count (WBC), were only assessed in single studies. Twenty-four-hour average plasma IL-7 levels were lower in individuals with obesity compared to those with AN restrictive type, constitutional thinness or normal weight controls (23). Serum IL-8 was positively associated with night time eating (21). IL-10 was found to be lower in individuals with obesity but higher in AN compared to normal weight controls. IL-10 levels were also associated with post-dinner eating and “not having sweet eating” (21). Serum fractalkine levels were higher in adolescent girls with obesity but lower in AN compared to healthy controls (30). Lucas et al. (25) found that plasma levels of α-MSH/IgG immune complex in individuals with obesity were lower than healthy controls (25). The melanocortin-4-receptor (MC4R) binding affinity, internalization and receptor activation of α-MSH/IgG immune complex were also decreased in individuals with obesity. On the contrary, the internalization rate of the immune complex was higher in AN. Higher white blood cell counts were found in obese individuals with BED compared to those without BED (29). Cazettez et al. (22) assessed the associations between fibrinogen and brain regions linked to eating behavior regulation (22). The authors found that higher plasma fibrinogen levels were associated with smaller lateral orbitofrontal cortex (OFC) volume in the overweight and obese group. Among individuals with excess weight, higher fibrinogen levels were linked to greater interstitial fluid in the amygdala and right parietal lobe. Other assessed inflammatory markers, including IL-1β, IL-2, IL-4, TGF-β, and MCP-1, were not significantly associated with obesity-related eating behaviors.
Discussion
To date, only a few studies have focused on the associations between inflammatory/immune molecules and obesity-related eating behaviors. Despite the limited number of studies, a variety of inflammatory/immune markers and eating behaviors have been examined. Yet, the findings have been inconsistent. The inflammatory markers that potentially have a positive relationship with obesity-related eating behaviors include hsCRP, fractalkine, and fibrinogen. Other immune molecules, including INF-γ, IL-7, IL-10 and α-MSH/IgG, may have negative associations with obesity-related eating behaviors. However, these findings were mainly identified by single studies and replications are warranted.
Additionally, the majority of the identified studies were either case-control or cross-sectional studies, which were not capable of clarifying the causality between inflammatory markers and obesity-related eating behaviors. However, current evidence indicates a potential bi-directional relationship. Dysregulation of the immunological molecules may contribute to the development or progression of obesity-related eating behaviors. On the other hand, these eating behaviors may stimulate the production of pro-inflammatory cytokines through imbalanced nutrient intake and/or excess weight gain.
Although the blood-brain barrier prevents large molecules and cells from being freely transported to the central nervous system, studies have indicated that immune cells and cytokines with the facilitation of saturable transporters can pass through the barrier (31, 32). Also, peripheral cytokines may stimulate endothelial cells of the barrier to produce substances, such as prostaglandins, to induce inflammation and activate selective hypothalamic neurons (33, 34). Furthermore, microglia in the central nervous system share immunological functions as mononuclear cells. Both microglia and astrocytes are able to secrete inflammatory cytokines (35, 36). Once immune cells and molecules reach the central nervous system, several mechanisms potentially underlie their role in the regulation of obesity-related eating behaviors (8, 37), including modulation of orexigenic and anorexigenic signals in the hypothalamus, induction of hypothalamic inflammation, and regulation of food reward circuitries (Figure 2). In addition, vagus nerves may partially relay the effect of peripheral immune molecules on the regulation of obesity-related eating behaviors (38).
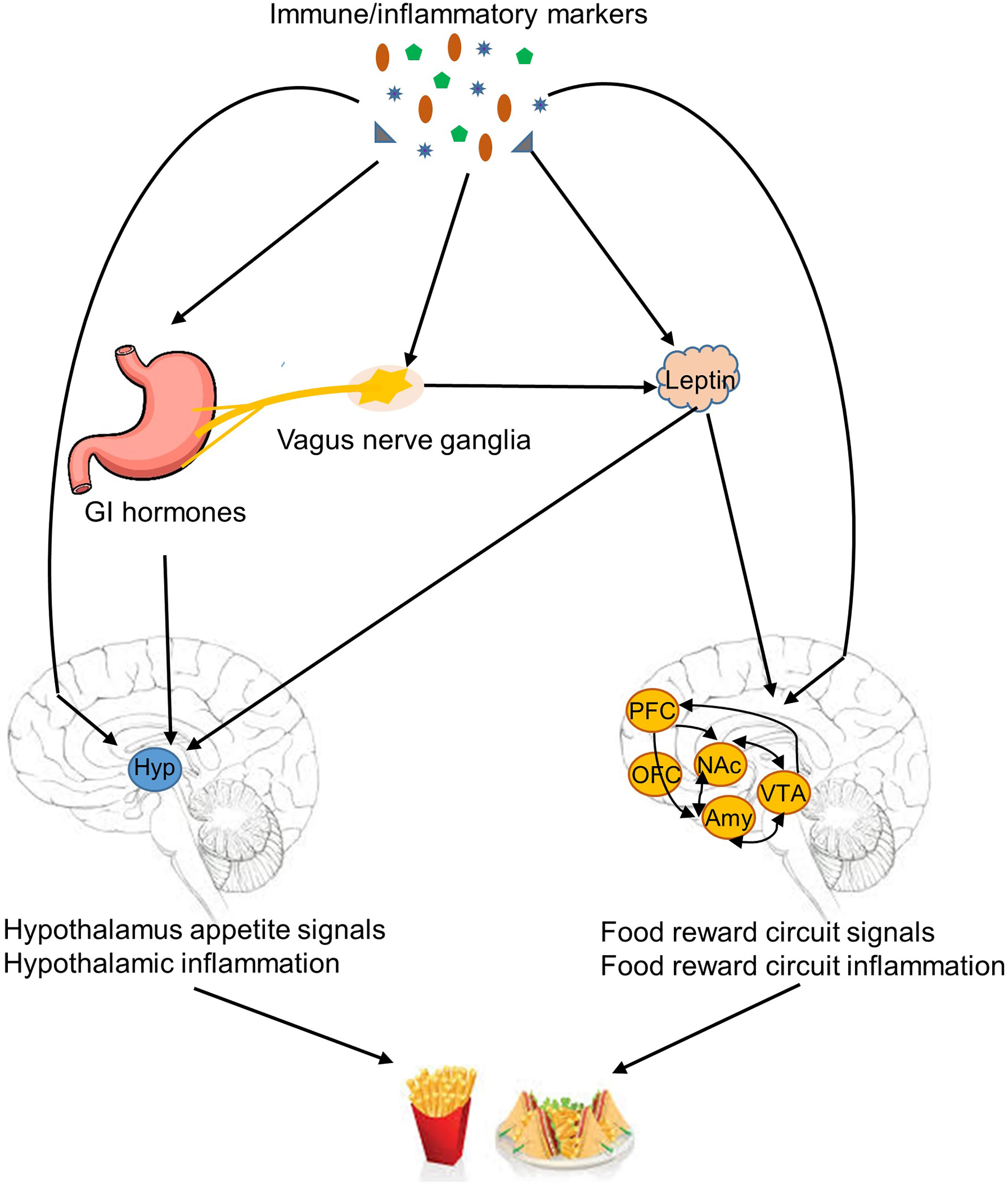
Figure 2 Mechanisms linked immune/inflammatory markers to obesity-related eating behaviors. Immune/inflammatory markers are potentially involved in several mechanisms of regulation of obesity-related eating behaviors, including modulation of orexigenic and anorexigenic signals in the hypothalamus, induction of hypothalamic inflammation, regulation of food reward circuitries, and suppression anorexigenic signals by induction of inflammation in the vagus nerve ganglia. GI, gastrointestinal tract; Hyp, hypothalamus; PFC, prefrontal cortex; OFC, orbitofrontal cortex; NAc, nucleus accumbens; Amy, amygdala; VTA, ventral tegmental area.
Evidence suggests a role of immune molecules in the regulation of orexigenic and anorexigenic signals and subsequent modulation of appetite and satiety in the hypothalamus. Inflammatory cytokines, such as IL-1β, can stimulate leptin production (15, 39). Leptin is a key adipokine that interacts with neurons in the hypothalamus and food reward circuits to modulate food intake (40). IL-6 can enhance central leptin action, increase hypothalamic signal transducer, and reduce food intake (11). Cytokines also interact with other hormones and neuropeptides involved in eating regulation. IL-1β and interferon may reduce circulating appetite-stimulating hormones, such as ghrelin, and increase appetite-suppressing hormones, such as cholecystokinin (CCK) (41–43). Cytokines, such as IL-1β and IL-7, are able to promote the expression of anorexigenic peptides, such as proopiomelanocortin, and inhibit the expression of orexigenic peptides, such as agouti-related peptide and neuropeptide Y (4, 8). Circulating immunoglobulins are also found to bind to α-melanocyte-stimulating hormone (α-MSH) to form immune complexes (α-MSH/IgG) and modulate the activation of MC4R to decrease appetite (25). Furthermore, other than the role in regulating orexigenic and anorexigenic signals, inflammatory cytokines are able to directly activate neurons in key hypothalamic appetite regulatory regions. IL-1 receptors have been found in neurons in the hypothalamic arcuate nucleus (ARC), ventromedial nucleus, paraventricular nucleus, and the lateral hypothalamic area (8). IL-7 has been shown to directly activate and improve survival of the ARC neurons (4).
Another mechanism particularly related to the perpetuation of obesity and obesity-relate eating behaviors is hypothalamic inflammation (44). In animal models, high fat feeding activates cellular inflammation in diverse tissues including the hypothalamus (5, 45). Fractalkine is involved in the recruitment of monocytic cells to the hypothalamus and promotes hypothalamic inflammation induced by high fat diet (5). Hypothalamic inflammation contributes to leptin and insulin resistance (46). Studies have shown that central inhibition of the cellular inflammatory pathway in the hypothalamus can promote leptin and insulin sensitivity, reduce high fat food intake, and consequently protect against high fat food induced obesity (47–49). IL-10, a potent anti-inflammatory cytokine, may decrease hypothalamic inflammation. Controversially, IL-10 deficiency also reduces food intake (50).
In addition to appetite and satiety regulated by hypothalamic nuclei, hedonic eating is modulated by food reward circuitries located in the nucleus accumbens, ventral tegmental area, amygdala, and OFC (40). Decarie-spain et al. (51) found that high saturated fat intake triggers inflammation in the nucleus accumbens in mice and a blockage of cellular inflammation in this brain region suppresses compulsive sucrose seeking behavior (51). The lateral OFC and amygdala are potentially involved in taste and food preference (22, 52), and are important brain regions that participate in food reward (40). Cazette et al. (22) found that elevated levels of fibrinogen, a marker of inflammation, were associated with smaller lateral OFC, and increased interstitial fluid in the amygdala and the right parietal cortex in individuals with excess weight. Therefore, inflammation in food reward circuitries may also play a role in the development or perpetuation of obesity-related eating behaviors.
Other than direct interactions between immune molecules and the central nervous system, the vagus nerve is also involved in connecting peripheral inflammatory cytokines with eating regulation (8). A high fat diet can lead to inflammation in the vagus nerve ganglia, which can attenuate the signals of appetite suppressing hormones, such as leptin and CCK, relayed by the vagus nerve to the central nervous system (38).
In addition to eating regulation by immune molecules, obesity-related eating behaviors potentially affect the serum levels of inflammatory/immune markers through accumulated adipose tissue and food intake. Eating behaviors, such as disinhibited eating, emotional eating, high intake of dietary fat, contribute to excessive calorie intake and subsequently the development of obesity (3). The hyperplasia and hypertrophy of adipose tissue in obesity produces mediators, including adipokines and fatty acids, which trigger the accumulation of macrophages and lymphocytes (53). Cytokines derived from the accumulated immune cells lead to a state of chronic, low-grade systemic inflammation associated with obesity. Therefore, obesity-related eating behaviors may contribute to the systemic inflammation, such as elevated hsCRP, through their impact on adipose tissue (54, 55). Additionally, eating behaviors directly affect dietary intake (56, 57). Diet, such as high intake of dietary fat and low intake of vegetables, and certain dietary patterns, has been linked to increased serum levels of inflammatory markers, such CRP, TNF-α, and IL-6 (58–60). Hence, obesity-related eating behaviors may alter inflammatory status through dietary intake.
Thus far, the majority of evidence on the relationship between immune molecules and obesity-related eating behaviors has been generated by animal studies. More research in humans is desirable to delineate the role of immune/inflammatory molecules in modulating obesity-related eating behaviors and subsequently become potential targets for weight loss interventions. Moving forward, more rigorous research methodologies, such as randomized controlled trials and longitudinal studies, are necessary to delineate the causal relationship between immune/inflammation markers and obesity-related eating behaviors. Additional adjustment of potential confounders will be valuable to further define this relationship (61, 62). For example, systemic inflammation and obesity-related eating behaviors usually concur with obesity. Including or adjusting BMI or fat mass in the analysis is a critical step to elucidate the relationship. Similarly, dietary intake is another potential key mediator. However, none of the included studies considered dietary intake. Future studies with sophisticated research design, comprehensive theoretical models, and robust adjustment of relevant confounders may clarify the relationship between immune/inflammation markers and obesity-related eating behaviors in humans.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
YM designed the study and conducted literature searches. YM and AK screened the identified publications individually. YM summarized previous research studies and wrote the first draft of the manuscript. All authors reviewed and revised the draft and have approved the final manuscript.
Funding
YM is funded by NINR 5 K23 NR019014-02. NINR had no role in the study design, collection, analysis or interpretation of the data, writing the manuscript, or the decision to submit the paper for publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Fryar CD, Carroll MD, Afful J. Prevalence of overweight, obesity, and severe obesity among adults aged 20 and over: United states, 1960–1962 through 2017–2018. NCHS Health E-Stats (2020) 1–7.
2. Hales CM, Carroll MD, Fryar CD, Ogden CL. Prevalence of obesity and severe obesity among adults: United States, 2017-2018. US Department of Health and Human Services, Centers for Disease Control and Prevention (2020) (360):1–8.
3. French SA, Epstein LH, Jeffery RW, Blundell JE, Wardle J. Eating behavior dimensions. associations with energy intake and body weight. a review. Appetite (2012) 59(2):541–9. doi: 10.1016/j.appet.2012.07.001
4. Macia L, Viltart O, Delacre M, Sachot C, Héliot L, Di Santo JP, et al. Interleukin-7, a new cytokine targeting the mouse hypothalamic arcuate nucleus: role in body weight and food intake regulation. PloS One (2010) 5(4):e9953. doi: 10.1371/journal.pone.0009953
5. Morari J, Anhe GF, Nascimento LF, de Moura RF, Razolli D, Solon C, et al. Fractalkine (CX3CL1) is involved in the early activation of hypothalamic inflammation in experimental obesity. Diabetes (2014) 63(11):3770–84. doi: 10.2337/db13-1495
6. Netea MG, Joosten LA, Lewis E, Jensen DR, Voshol PJ, Kullberg BJ, et al. Deficiency of interleukin-18 in mice leads to hyperphagia, obesity and insulin resistance. Nat Med (2006) 12(6):650–6. doi: 10.1038/nm1415
7. Wallenius K, Wallenius V, Sunter D, Dickson SL, Jansson J-O. Intracerebroventricular interleukin-6 treatment decreases body fat in rats. Biochem Biophys Res Commun (2002) 293(1):560–5. doi: 10.1016/S0006-291X(02)00230-9
8. Wong S, Pinkney J. Role of cytokines in regulating feeding behaviour. Curr Drug Targets (2004) 5(3):251–63. doi: 10.2174/1389450043490532
9. Agnello E, Malfi G, Costantino AM, Massarenti P, Pugliese M, Fortunati N, et al. Tumour necrosis factor alpha and oxidative stress as maintaining factors in the evolution of anorexia nervosa. Eat Weight Disord (2012) 17(3):e194–199. doi: 10.1007/bf03325347
10. Loumaye A, Thissen J-P. Biomarkers of cancer cachexia. Clin Biochem (2017) 50(18):1281–8. doi: 10.1016/j.clinbiochem.2017.07.011
11. Sadagurski M, Norquay L, Farhang J, D’Aquino K, Copps K, White MF. Human IL6 enhances leptin action in mice. Diabetologia (2010) 53(3):525–35. doi: 10.1007/s00125-009-1580-8
12. Scheede-Bergdahl C, Watt HL, Trutschnigg B, Kilgour RD, Haggarty A, Lucar E, et al. Is IL-6 the best pro-inflammatory biomarker of clinical outcomes of cancer cachexia? Clin Nutr (2012) 31(1):85–8. doi: 10.1016/j.clnu.2011.07.010
13. García MC, Wernstedt I, Berndtsson A, Enge M, Bell M, Hultgren O, et al. Mature-onset obesity in interleukin-1 receptor I knockout mice. Diabetes (2006) 55(5):1205–13. doi: 10.2337/db05-1304
14. Wallenius V, Wallenius K, Ahrén B, Rudling M, Carlsten H, Dickson SL, et al. Interleukin-6-deficient mice develop mature-onset obesity. Nat Med (2002) v8(1):75–9. doi: 10.1038/nm0102-75
15. Kurosawa N, Shimizu K, Seki K. The development of depression-like behavior is consolidated by IL-6-induced activation of locus coeruleus neurons and IL-1β-induced elevated leptin levels in mice. Psychopharmacology (2016) 233(9):1725–37. doi: 10.1007/s00213-015-4084-x
16. Quarta C, Clemmensen C, Zhu Z, Yang B, Joseph SS, Lutter D, et al. Molecular integration of incretin and glucocorticoid action reverses immunometabolic dysfunction and obesity. Cell Metab (2017) 26(4):620–632.e626. doi: 10.1016/j.cmet.2017.08.023
17. Valdearcos M, Robblee MM, Benjamin DI, Nomura DK, Xu AW, Koliwad SK. Microglia dictate the impact of saturated fat consumption on hypothalamic inflammation and neuronal function. Cell Rep (2014) 9(6):2124–38. doi: 10.1016/j.celrep.2014.11.018
18. Wong N, Fam BC, Cempako GR, Steinberg GR, Walder K, Kay TW, et al. Deficiency in interferon-gamma results in reduced body weight and better glucose tolerance in mice. Endocrinology (2011) 152(10):3690–9. doi: 10.1210/en.2011-0288
19. Ellacott KLJ, Morton GJ, Woods SC, Tso P, Schwartz MW. Assessment of feeding behavior in laboratory mice. Cell Metab (2010) 12(1):10–7. doi: 10.1016/j.cmet.2010.06.001
20. Capuron L, Poitou C, Machaux-Tholliez D, Frochot V, Bouillot JL, Basdevant A, et al. Relationship between adiposity, emotional status and eating behaviour in obese women: role of inflammation. Psychol Med (2011) 41(7):1517–28. doi: 10.1017/s0033291710001984
21. Caroleo M, Carbone EA, Greco M, Corigliano DM, Arcidiacono B, Fazia G, et al. Brain-Behavior-Immune interaction: Serum cytokines and growth factors in patients with eating disorders at extremes of the body mass index (BMI) spectrum. Nutrients (2019) 11(9):1995. doi: 10.3390/nu11091995
22. Cazettes F, Cohen JI, Yau PL, Talbot H, Convit A. Obesity-mediated inflammation may damage the brain circuit that regulates food intake. Brain Res (2011) 1373:101–9. doi: 10.1016/j.brainres.2010.12.008
23. Germain N, Viltart O, Loyens A, Bruchet C, Nadin K, Wolowczuk I, et al. Interleukin-7 plasma levels in human differentiate anorexia nervosa, constitutional thinness and healthy obesity. PloS One (2016) 11(9):e0161890. doi: 10.1371/journal.pone.0161890
24. Lofrano-Prado MC, Prado WL, de Piano A, Tock L, Caranti DA, Nascimento CM, et al. Eating disorders in adolescents: correlations between symptoms and central control of eating behavior. Eat Behav (2011) 12(1):78–82. doi: 10.1016/j.eatbeh.2010.09.008
25. Lucas N, Legrand R, Bôle-Feysot C, Breton J, Coëffier M, Akkermann K, et al. Immunoglobulin G modulation of the melanocortin 4 receptor signaling in obesity and eating disorders. Transl Psychiatry (2019) 9(1):87. doi: 10.1038/s41398-019-0422-9
26. Raymond NC, Dysken M, Bettin K, Eckert ED, Crow SJ, Markus K, et al. Cytokine production in patients with anorexia nervosa, bulimia nervosa, and obesity. Int J Eat Disord (2000) 28(3):293–302. doi: 10.1002/1098-108x(200011)28:3<293::aid-eat6>3.0.co;2-f
27. Sayin FK, Buyukinan M, Arslan D, Deniz CD. Associations between emotional eating and metabolic risk factors at adolescents with obesity. Int J Child Health Nutr (2020) 9(3):93–9. doi: 10.6000/1929-4247.2020.09.03.1
28. Shank LM, Tanofsky-Kraff M, Kelly NR, Schvey NA, Marwitz SE, Mehari RD, et al. Pediatric loss of control eating and high-sensitivity c-reactive protein concentrations. Child Obes (2017) 13(1):1–8. doi: 10.1089/chi.2016.0199
29. Succurro E, Segura-Garcia C, Ruffo M, Caroleo M, Rania M, Aloi M, et al. Obese patients with a binge eating disorder have an unfavorable metabolic and inflammatory profile. Med (Baltimore) (2015) 94(52):e2098. doi: 10.1097/md.0000000000002098
30. Zhang S, Tang H, Gong C, Liu J, Chen J. Assessment of serum CX3CL1/fractalkine level in han Chinese girls with anorexia nervosa and its correlation with nutritional status: a preliminary cross-sectional study. J Investig Med (2017) 65(2):333–7. doi: 10.1136/jim-2016-000209
31. Banks WA. Anorectic effects of circulating cytokines: role of the vascular blood-brain barrier. Nutrition (2001) 17(5):434–7. doi: 10.1016/s0899-9007(01)00507-x
32. Kleine TO, Benes L. Immune surveillance of the human central nervous system (CNS): different migration pathways of immune cells through the blood-brain barrier and blood-cerebrospinal fluid barrier in healthy persons. Cytomet A (2006) 69(3):147–51. doi: 10.1002/cyto.a.20225
33. Bierhaus A, Chen J, Liliensiek B, Nawroth PP. LPS and cytokine-activated endothelium. Semin Thromb Hemost (2000) 26(5):571–87. doi: 10.1055/s-2000-13214
34. Zhang J, Rivest S. A functional analysis of EP4 receptor-expressing neurons in mediating the action of prostaglandin E2 within specific nuclei of the brain in response to circulating interleukin-1beta. J Neurochem (2000) 74(5):2134–45. doi: 10.1046/j.1471-4159.2000.0742134.x
35. Farina C, Aloisi F, Meinl E. Astrocytes are active players in cerebral innate immunity. Trends Immunol (2007) 28(3):138–45. doi: 10.1016/j.it.2007.01.005
36. Harry GJ, Kraft AD. Neuroinflammation and microglia: considerations and approaches for neurotoxicity assessment. Expert Opin Drug Metab Toxicol (2008) 4(10):1265–77. doi: 10.1517/17425255.4.10.1265
37. Borges BC, Elias CF, Elias LL. PI3K signaling: A molecular pathway associated with acute hypophagic response during inflammatory challenges. Mol Cell Endocrinol (2016) 438:36–41. doi: 10.1016/j.mce.2016.07.005
38. Ueno H, Nakazato M. Mechanistic relationship between the vagal afferent pathway, central nervous system and peripheral organs in appetite regulation. J Diabetes Investig (2016) 7(6):812–8. doi: 10.1111/jdi.12492
39. Sarraf P, Frederich RC, Turner EM, Ma G, Jaskowiak NT, Rivet DJ, et al. Multiple cytokines and acute inflammation raise mouse leptin levels: potential role in inflammatory anorexia. J Exp Med (1997) 185(1):171–5. doi: 10.1084/jem.185.1.171
40. Leigh SJ, Morris MJ. The role of reward circuitry and food addiction in the obesity epidemic: An update. Biol Psychol (2018) 131:31–42. doi: 10.1016/j.biopsycho.2016.12.013
41. Kurosawa M, Uvnäs-Moberg K, Miyasaka K, Lundeberg T. Interleukin-1 increases activity of the gastric vagal afferent nerve partly via stimulation of type a CCK receptor in anesthetized rats. J Auton Nerv Syst (1997) 62(1-2):72–8. doi: 10.1016/s0165-1838(96)00111-7
42. Madison LD, Scarlett JM, Levasseur P, Zhu X, Newcomb K, Batra A, et al. Prostacyclin signaling regulates circulating ghrelin during acute inflammation. J Endocrinol (2008) 196(2):263–73. doi: 10.1677/joe-07-0478
43. Stutte S, Ruf J, Kugler I, Ishikawa-Ankerhold H, Parzefall A, Marconi P, et al. Type I interferon mediated induction of somatostatin leads to suppression of ghrelin and appetite thereby promoting viral immunity in mice. Brain Behav Immun (2021) 95:429–43. doi: 10.1016/j.bbi.2021.04.018
44. Wisse BE, Schwartz MW. Does hypothalamic inflammation cause obesity? Cell Metab (2009) 10(4):241–2. doi: 10.1016/j.cmet.2009.09.003
45. De Souza C, Araujo EP, Bordin S, Ashimine R, Zollner RL, Boschero AC, et al. Consumption of a fat-rich diet activates a proinflammatory response and induces insulin resistance in the hypothalamus. Endocrinology (2005) 146(10):4192–9. doi: 10.1210/en.2004-1520
46. Velloso LA. The brain is the conductor: diet-induced inflammation overlapping physiological control of body mass and metabolism. Arq Bras Endocrinol Metabol (2009) 53(2):151–8. doi: 10.1590/s0004-27302009000200006
47. Kleinridders A, Schenten D, Könner AC, Belgardt BF, Mauer J, Okamura T, et al. MyD88 signaling in the CNS is required for development of fatty acid-induced leptin resistance and diet-induced obesity [Article]. Cell Metab (2009) 10(4):249–59. doi: 10.1016/j.cmet.2009.08.013
48. Posey KA, Clegg DJ, Printz RL, Byun J, Morton GJ, Vivekanandan-Giri A, et al. Hypothalamic proinflammatory lipid accumulation, inflammation, and insulin resistance in rats fed a high-fat diet. Am J Physiol Endocrinol Metab (2009) 296(5):E1003–1012. doi: 10.1152/ajpendo.90377.2008
49. Zhang X, Zhang G, Zhang H, Karin M, Bai H, Cai D. Hypothalamic IKKβ/NF-κB and ER stress link overnutrition to energy imbalance and obesity. Cell (2008) 135(1):61–73. doi: 10.1016/j.cell.2008.07.043
50. Gotoh K, Inoue M, Masaki T, Chiba S, Shimasaki T, Ando H, et al. A novel anti-inflammatory role for spleen-derived interleukin-10 in obesity-induced hypothalamic inflammation. J Neurochem (2012) 120(5):752–64. doi: 10.1111/j.1471-4159.2011.07617.x
51. Décarie-Spain L, Sharma S, Hryhorczuk C, Issa-Garcia V, Barker PA, Arbour N, et al. Nucleus accumbens inflammation mediates anxiodepressive behavior and compulsive sucrose seeking elicited by saturated dietary fat. Mol Metab (2018) 10:1–13. doi: 10.1016/j.molmet.2018.01.018
52. Machado CJ, Bachevalier J. Measuring reward assessment in a semi-naturalistic context: the effects of selective amygdala, orbital frontal or hippocampal lesions. Neuroscience (2007) 148(3):599–611. doi: 10.1016/j.neuroscience.2007.06.035
53. Chatzigeorgiou A, Karalis KP, Bornstein SR, Chavakis T. Lymphocytes in obesity-related adipose tissue inflammation. Diabetologia (2012) 55(10):2583–92. doi: 10.1007/s00125-012-2607-0
54. Maki KC, Rains TM, Bell M, Reeves MS, Farmer MV, Yasunaga K. Fat mass, abdominal fat distribution, and c-reactive protein concentrations in overweight and obese men and women. Metab Syndr Relat Disord (2011) 9(4):291–6. doi: 10.1089/met.2010.0132
55. Wen X, Pekkala S, Wang R, Wiklund P, Feng G, Cheng SM, et al. Does systemic low-grade inflammation associate with fat accumulation and distribution? a 7-year follow-up study with peripubertal girls. J Clin Endocrinol Metab (2014) 99(4):1411–9. doi: 10.1210/jc.2013-3267
56. Bui C, Lin LY, Wu CY, Chiu YW, Chiou HY. Association between emotional eating and frequency of unhealthy food consumption among Taiwanese adolescents. Nutrients (2021) 13(8):2739. doi: 10.3390/nu13082739
57. de Lauzon B, Romon M, Deschamps V, Lafay L, Borys JM, Karlsson J, et al. The three-factor eating questionnaire-R18 is able to distinguish among different eating patterns in a general population. J Nutr (2004) 134(9):2372–80. doi: 10.1093/jn/134.9.2372
58. Esposito K, Ciotola M, Giugliano D. Mediterranean Diet, endothelial function and vascular inflammatory markers. Public Health Nutr (2006) 9(8a):1073–6. doi: 10.1017/s1368980007668529
59. Kopf JC, Suhr MJ, Clarke J, Eyun SI, Riethoven JM, Ramer-Tait AE, et al. Role of whole grains versus fruits and vegetables in reducing subclinical inflammation and promoting gastrointestinal health in individuals affected by overweight and obesity: a randomized controlled trial. Nutr J (2018) 17(1):72. doi: 10.1186/s12937-018-0381-7
60. van Dijk SJ, Feskens EJ, Bos MB, Hoelen DW, Heijligenberg R, Bromhaar MG, et al. A saturated fatty acid-rich diet induces an obesity-linked proinflammatory gene expression profile in adipose tissue of subjects at risk of metabolic syndrome. Am J Clin Nutr (2009) 90(6):1656–64. doi: 10.3945/ajcn.2009.27792
61. Koop MA, Lutke Schipholt IJ, Scholten-Peeters GGM, Coppieters MW. Identifying the most important confounders when assessing the association between low-grade systemic inflammation and musculoskeletal pain: A modified Delphi study. Pain Med (2021) 22(11):2661–9. doi: 10.1093/pm/pnab243
Keywords: eating behavior, immune markers, cytokine, obesity, inflammatory marker
Citation: Meng Y and Kautz A (2022) An evidence review of the association of immune and inflammatory markers with obesity-related eating behaviors. Front. Immunol. 13:902114. doi: 10.3389/fimmu.2022.902114
Received: 22 March 2022; Accepted: 27 June 2022;
Published: 15 July 2022.
Edited by:
Mark Andrew Cline, Virginia Tech, United StatesReviewed by:
Aleksandra Klisic, Primary Health Care Center Podgorica, MontenegroChang Cao, University of Illinois at Urbana-Champaign, United States
Copyright © 2022 Meng and Kautz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying Meng, WWluZ19NZW5nQHVybWMucm9jaGVzdGVyLmVkdQ==