
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 30 September 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.901872
 Cristina Segú-Vergés1
Cristina Segú-Vergés1 Silvia Caño2
Silvia Caño2 Elisabeth Calderón-Gómez3Helena Bartra1Teresa Sardon1
Elisabeth Calderón-Gómez3Helena Bartra1Teresa Sardon1 Srini Kaveri4*
Srini Kaveri4* José Terencio2
José Terencio2Intravenous immunoglobulin (IVIg) is used as treatment for several autoimmune and inflammatory conditions, but its specific mechanisms are not fully understood. Herein, we aimed to evaluate, using systems biology and artificial intelligence techniques, the differences in the pathophysiological pathways of autoimmune and inflammatory conditions that show diverse responses to IVIg treatment. We also intended to determine the targets of IVIg involved in the best treatment response of the evaluated diseases. Our selection and classification of diseases was based on a previously published systematic review, and we performed the disease characterization through manual curation of the literature. Furthermore, we undertook the mechanistic evaluation with artificial neural networks and pathway enrichment analyses. A set of 26 diseases was selected, classified, and compared. Our results indicated that diseases clearly benefiting from IVIg treatment were mainly characterized by deregulated processes in B cells and the complement system. Indeed, our results show that proteins related to B-cell and complement system pathways, which are targeted by IVIg, are involved in the clinical response. In addition, targets related to other immune processes may also play an important role in the IVIg response, supporting its wide range of actions through several mechanisms. Although B-cell responses and complement system have a key role in diseases benefiting from IVIg, protein targets involved in such processes are not necessarily the same in those diseases. Therefore, IVIg appeared to have a pleiotropic effect that may involve the collaborative participation of several proteins. This broad spectrum of targets and ‘non-specificity’ of IVIg could be key to its efficacy in very different diseases.
Intravenous immunoglobulin (IVIg) is a preparation of normal human IgG derived from large pools of healthy human plasma (1). IVIg is used as replacement therapy to prevent infections in patients with primary or secondary immune deficiency syndromes and, at high doses, exerts immunomodulatory effects to treat autoimmune and inflammatory disorders (2). Indeed, IVIg has been approved by the European Medicines Agency (EMA) for several autoimmune conditions, such as immune thrombocytopenia (ITP), chronic inflammatory demyelinating polyneuropathy (CIDP), Guillain-Barré syndrome (GBS), Kawasaki disease (KD), and multifocal motor neuropathy (MMN) (3). In addition, the off-label uses of IVIg are growing rapidly, and several other autoimmune and inflammatory conditions have been suggested as potential indications (4). Nonetheless, limited clinical evidence indicates that the successful response to IVIg observed in some autoimmune conditions may not necessarily and systematically be extended to others. For instance, adrenoleukodystrophy and inclusion body myositis do not respond well to this treatment (2, 5, 6).
The beneficial effect of IVIg is explained by mechanisms involving either its F(ab’)2 (7) or Fc domain (8) or both. The immunomodulatory effects of IVIg are complex and intricate; IVIg has been proved to modulate B and T cells, phagocytosis, complement activity, cytokine production, and the properties of dendritic cells (DCs), among many others (1, 9–11). This wide range of cellular and molecular targets can trigger pleiotropic effects in the immune system (1, 9). The specific immune and inflammatory processes modulated by IVIg depend on the pathophysiological process driving each disease (10, 12). Besides, the molecular mechanisms may also depend on the dose of IVIg and the window during the immunopathogenesis when IVIG is administered (13). Although IVIg has been shown to modulate a large number of immunological processes, the complete understanding of the molecular mechanisms defining treatment response remains unclear for some of them.
Systems biology and artificial intelligence tools have been used in the past to unveil the mechanism of action of a variety of drugs for hematological or cardiovascular diseases, among others (14, 15). However, this approach has been rarely used on IVIg, and the few reported examples were centered on the treatment of a specific disease (16–19). Therefore, we considered that the abovementioned tools could be applied in a larger setting to shed some light on the mechanisms of action of IVIg and their diversity depending on the treated disease. By means of system biology and machine learning models based on Artificial Neural Networks (ANNs) (20–22), we evaluated the differences in the pathophysiological pathways of autoimmune and inflammatory conditions that show diversity in responses to treatment with IVIg. Also, we aimed at determining the targets of IVIg involved in the best treatment response of the evaluated diseases.
To compare the heterogeneous efficacy response to IVIg treatment, we resorted to a previously published systematic and evidence-based classification of neurological and autoimmune diseases (2). We further classified the diseases according to their response to IVIg in four clusters, as previously reported (2): ‘definitely beneficial’ (DB), ‘probably beneficial’ (PB), ‘may provide benefit’ (MPB), and ‘unlikely to provide benefit’ (UPB) (Table 1, left column). The level and strength of evidence supported this classification according to: (i) evidence category, in groups Ia, Ib, IIa, IIb, III, and IV (e.g., group Ia referred to evidence obtained from meta-analysis of randomized controlled trials, while group IV to those obtained from expert committee reports, opinions or clinical experience of respected authorities of both), and (ii) strength of recommendation, which estimated the strength of the evidence to assign the condition to an IVIg response cluster, and ranged from A (strongest support) to D (weakest support) (2). For our study, we only included disorders for which the benefit/no benefit of IVIg treatment had been established with enough clinical evidence, i.e. those with a level of evidence Ia, Ib, IIa, or IIb (supported by clinical trials) and a strength of recommendation of A or B (2). We considered ‘IgM anti-myelin–associated glycoprotein (anti-MAG) paraprotein-associated peripheral neuropathy’ and ‘demyelinating neuropathy associated with monoclonal IgM’ as the same disease because of their degree of similarity and lack of differential molecular information available in scientific literature. Therefore, we hereafter referred to these two diseases as ‘IgM anti-MAG paraprotein-associated peripheral neuropathy’. To increase the number of analyzed disorders, we added two other autoimmune diseases with a well-established level of clinical response to IVIg: KD and Crohn´s disease (CD). IVIg has been approved by the US Food and Drug Administration (23) and the EMA (24) to treat KD according to strong evidence supporting its positive response (note that, in the abovementioned systematic review (2), KD was classified as an infection-related disease and not an autoimmune disease). Therefore, we incorporated KD into the DB cluster. On the contrary, unclear clinical evidence on the impact of IVIg on CD prompted us to include it in the UPB cluster in our study. We only reclassified one condition in our study, myasthenia gravis (MG), an approved indication for IVIg by the EMA. MG was moved from the PB to the DB cluster on the basis of the extended clinical use of IVIg in this condition and its positive response according to clinical evidence (25–27).
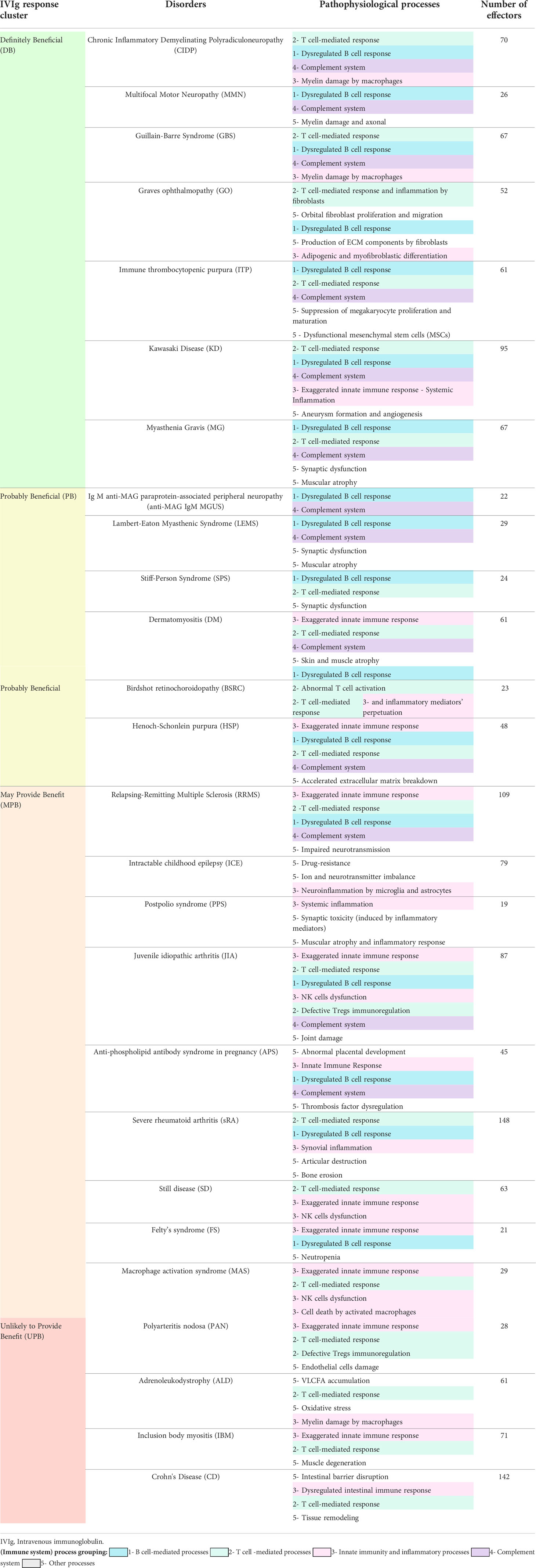
Table 1 Pathophysiological processes and number of effectors of evaluated neuroimmunological and autoimmune diseases according to reported response to IVIg.
We characterized the selected conditions molecularly through manual curation of the current scientific literature, as previously reported (21). First, we searched for reviews on the molecular pathogenesis, pathophysiology, and molecular mechanisms of the conditions in the PubMed database. Second, we retrieved the publications identified in these searches and assessed them at the title and abstract level. If molecular information describing the condition pathophysiology was present, we thoroughly reviewed full texts seeking to identify the main pathophysiological processes involved in the condition. We referred to these processes as ‘pathophysiological processes’. Third, we further characterized each pathophysiological process at the protein level (Supplementary Table S1 in the S1 File). We reviewed the retrieved publications to identify protein/gene candidates to be condition effectors, i.e., proteins whose activity (or lack thereof) was functionally associated with the development of the condition. Finally, if we judged the evidence of the involvement of a candidate gene/protein in the condition not consistent enough for being considered an effector, we performed an additional PubMed search specifically on the protein candidate, including all protein names according to UniProtKB. The specific terms used for searches in the PubMed database and the abovementioned features for each studied disease are listed in Supplementary Table S1 in the S1 File. We performed our searches in June 2020, limited them to 10 years before that date, and expanded them by reviewing relevant references in the results. We only included candidate articles with references more recent than year 2000, with the exception of references for anti-MAG, for which very little molecular information was found with this protocol, and for references for immunoglobulin heavy chain type detected among the autoantibodies in patients of each disease. The list of protein effectors considered for each disease is provided in Supplementary Table S1 in the S1 File.
To characterize the target profile of IVIg, we reviewed the Drugbank (28), Stitch (29), and Supertarget (30) data sources to obtain further information on IVIg targets. In addition, to completely characterize IVIg, we searched in the PubMed database on March 24, 2020, publications from the last ten years regarding known targets and mechanisms of action of IVIg, using the following main keywords in the title and/or abstract: “IVIG”, “Intravenous Immunoglobulin”, “Intravenous Immune Globulin”, “Intravenous Immunoglobulins”, “Intravenous Immune Globulins”, “Molecular”, “Mechanism”, “Pathophysiology”, “Pathogenesis”, “Mode”, “Action”, “Signaling”, “Signalling”, “Expression”, “Activation”, “Inhibition”, “Activity”. All results were evaluated, and reference lists were used to expand the search. Importantly, we considered IVIg targets only those proteins directly blocked or activated by IVIg (direct targets, Table 2). In contrast, we designated as ‘indirect targets’ those genes or proteins that, although lacking a direct interaction with IVIg, were modified by the effect of IVIg over its targets at the downstream level (Table 3). Finally, note that the artificial neuronal network (ANN) analysis performed in this work was focused solely on IVIg direct targets. Once identified, we classified direct targets in functional groups as per their immune-related function.
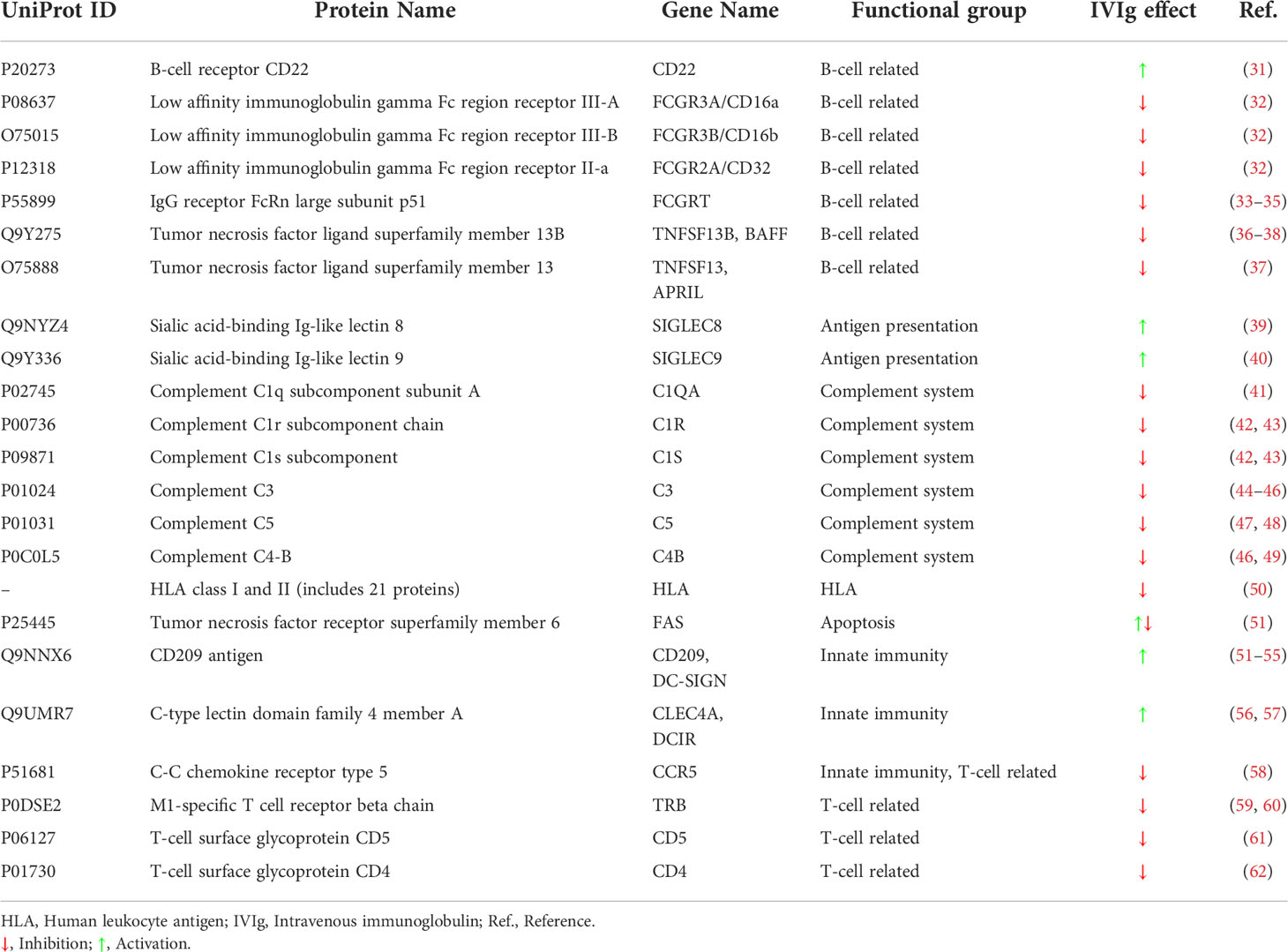
Table 2 List of IVIg protein targets (direct targets) identified by literature search and classified according to their related immunological function.
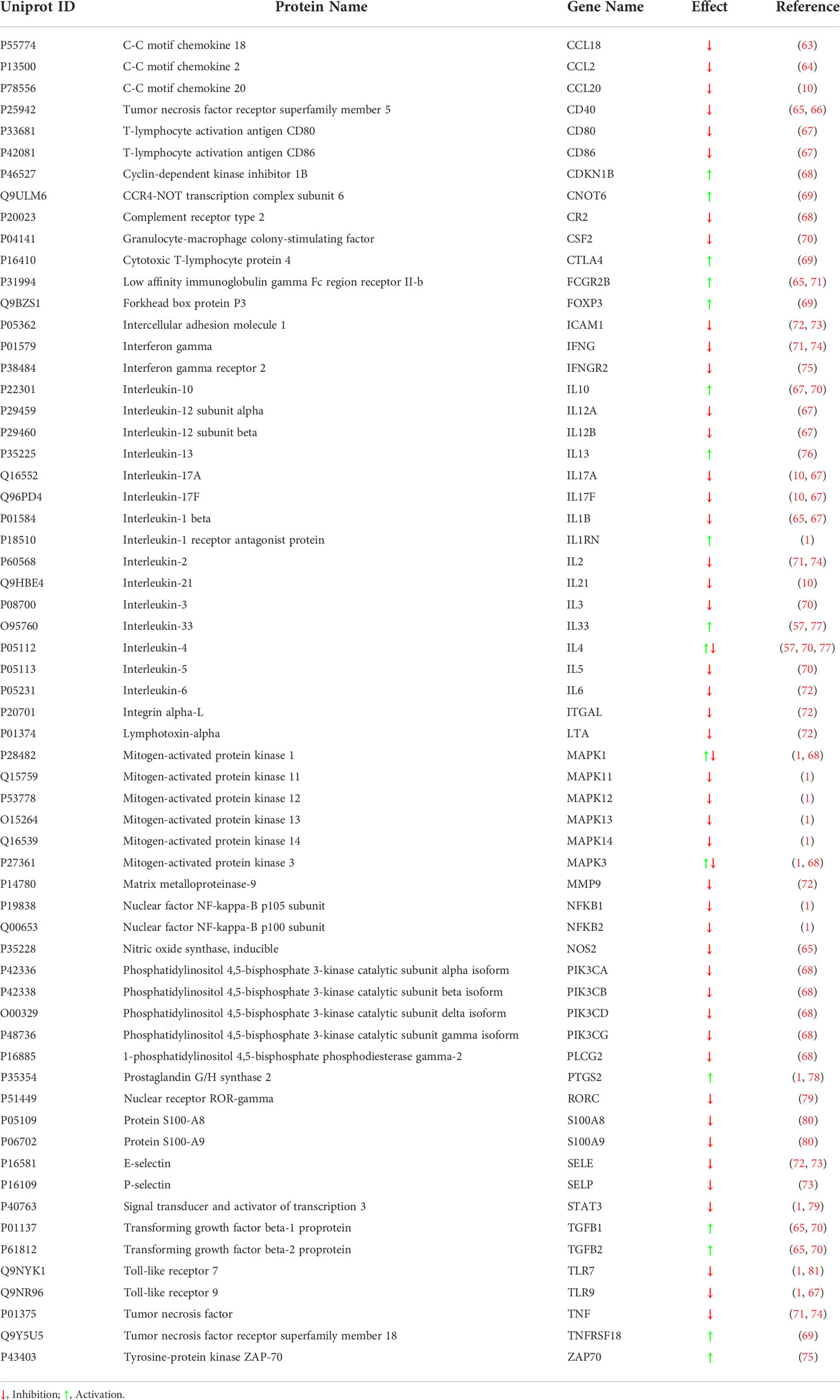
Table 3 Indirect protein targets modulated by IVIg according to the literature.
Therapeutic performance mapping system technology (14), based on systems biology, integrates available protein-protein network information along with physiological and pathophysiological data that allows to create machine learning models based on Artificial Neural Networks (ANNs), which have been previously defined and applied (20–22). ANNs are supervised algorithms that identify relations between proteins (e.g. drug targets) and clinical elements of the network (22, 82, 83) by inferring the probability of the existence of a specific relationship between two or more protein sets. The training set (Supplementary Table S2 in the S2 File) was defined to predict the relationship between the drugs and their clinical conditions, drugs being stratified by the number of their of targets. The learning methodology used consisted in bagging training architecture of stratified ensembles of neural networks as final model. Each neural network model used was a multilayer perceptron (MLP) neural network classifier and they were considered as a weak classifier. These MLP were submitted to randomized initialization, having all of them a hidden layer containing between 7 and 11 nodes. Input database contained pairs of drug targets and biological conditions (drug-indication pairs) obtained from DrugBank and PubChem (84, 85). The final dataset was manually reviewed to avoid redundancies and mistakes that could affect the quality of the training set. The input feature vector is based on topological measures over the protein network from drug targets to condition according to the topological structure of the interconnection graph defined between them. The output was defined by a single node indicating the relation or no relation between the drug targets and the condition. Each training set for each MLP model was selected from a balanced subset of samples randomized by means of a Monte-Carlo cross-validation structure to reduce overfitting in the final model. The process to generate the final model was optimized by Levenberg-Marquardt strategy. A total of 1000 MLPs were trained and the best 75% of them for each stratum are considered in the final model. The implementation code was done in Matlab by using the Neural Network Toolbox, as combined with other Anaxomics Biotech’ developments. The final model performance is based on the capability to predict the pairs drug-indication, being evaluated as the AUC of the model. The final model has 81.77% of correct prediction when drugs tested have all their protein targets in the protein-protein network.
We employed these algorithms to explore the relationships between IVIg targets (either individually or grouped according to their functional group, Table 2) and the molecular definition of each disease (Supplementary Table S1 in the S1 File). ANN analysis provides a score for a target, or group of targets, based on the validations of the prediction capacity of the mathematical models toward the training set (known drugs and diseases, as described in databases). Each score is associated with a p-value that describes the probability that the result is a true positive. Accordingly, and to simplify their interpretation, we divided here the ranking scores in four categories: strong relationship (ANN score >78%, p-value<0.05), medium-strong relationship (ANN score >71%, p-value<0.1), medium relationship (ANN score >38%, p-value<0.25), and low or no relationship (ANN score ≤38%, p-value ≥0.25).
We used the chi-squared test to evaluate independence between the pathophysiological mechanisms involved in each disease and the disease classification based on IVIg response clustering. Also, we calculated inter-cluster disease similarity by direct protein overlap within the list of proteins included in the diseases of each cluster. Then, we performed the arithmetic mean of proportions of protein overlap (O) of each pair of disease clusters (C1, C2) according to equation 1:
We used the InteractiVenn software (86) to create Venn diagrams.
We applied hypergeometric pathway enrichment analysis (87) to determine whether pre-defined protein sets, according to biological reference databases, were represented within groups of protein effectors involved in the IVIg response-based disease clusters. Specifically, we used KEGG (88) and Gene Ontology (89) as reference databases. Results were submitted to Benjamini-Hochberg multi-test correction (90) to control false discovery rate (FDR). We only selected enriched pathways with an FDR q-value<0.05.
A set of 26 autoimmune and inflammatory conditions, with clinical evidence of optimal level to be classified as responders/non-responders to IVIg, was selected. Previously reported IVIg response categories were used (2) (with a few exceptions, see Materials and Methods, Table 1). Once characterized, we compared the diseases at the level of pathophysiological processes to identify those processes more frequently related to each IVIg response cluster. To facilitate this comparison, we grouped them in broad (immune system) processes as follows (Table 1): T cell-mediated response, B cell-mediated response, complement system, innate immunity/inflammation processes, and other (which included disease-specific dysfunctions, such as bone erosion or muscular atrophy). This classification was based on the association of the proteins contained in each characterized pathophysiological process.
Our results suggested that diseases within each IVIg response cluster shared similar pathophysiological processes, especially within the DB and UPB clusters, and that identifiable differences between the clusters could be found. A chi-squared test showed that there was dependency between pathological processes and disease classification based on IVIg response (Figure 1). In particular, diseases assigned to the DB cluster were positively associated with B cell-mediated processes and complement system. On the contrary, innate immunity and inflammatory processes were frequently associated to MPB (statistically significant association with 100% frequency) and UPB (not statistically association significant 100% frequency) clusters (Figure 1).
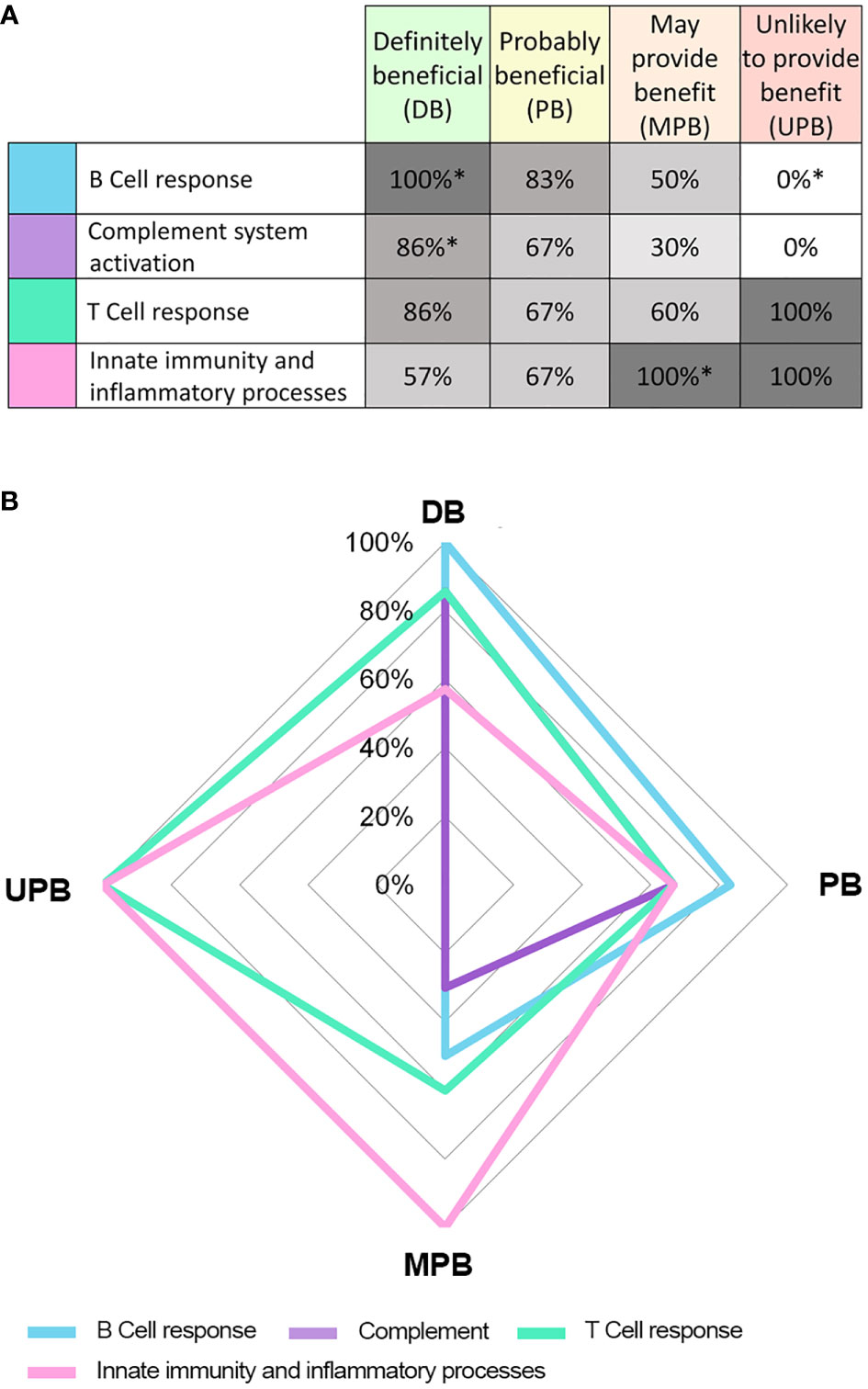
Figure 1 Results from the pathophysiological process grouping independence analysis with respect to different IVIg response clusters. Table (A) and graph (B) show the frequency (%) of conditions in each response cluster associated with each pathophysiological process. * Statistical significant (p< 0.05) result of the chi-squared test indicating dependence between the pathophysiological process group and cluster based in IVIg response.
We evaluated similarity between the disease clusters based on IVIg response with the aim of identifying processes occurring in diseases that clinically benefit from IVIg. When analyzing direct protein overlap (Figure 2A) between the bibliography-based list of effectors (Supplementary Table S1 in the S1 File) among the clusters, the DB and UPB clusters were the more distant. The specific effectors and those shared between the clusters were identified and grouped in three protein sets: proteins present in the DB cluster but not in the UPB cluster (DB w/o UPB), proteins specifically present in DB cluster (only DB), and proteins specifically present in UPB cluster (only-UPB) (Figure 2B). The enrichment analysis over the DB w/o UPB protein set provided 81 pathways enriched (Figure 2B). A detailed analysis of these enriched pathways (Figure 2C) unveiled that, while some of them were very general (e.g., positive regulation of the biological process, regulating signaling pathway), the most specific pathways pointed toward immune-related functions, such as processes of innate immunity response (e.g., leukocyte mediated immunity, phagocytosis), adaptive response (e.g., regulation of lymphocyte activation, adaptive immune response), autoimmunity (e.g., systemic lupus erythematosus, autoimmune thyroid disease), infection, T cell, B cell, and the complement system. Interestingly, there were other pathways enriched, including signaling processes (e.g., transferase activity, kinase activity, adrenergic signaling in cardiomyocytes) specific hormonal pathways (e.g., GnRH signaling pathway, thyroid hormone synthesis), and development pathways (e.g., progesterone-mediated oocyte maturation, long-term potentiation).
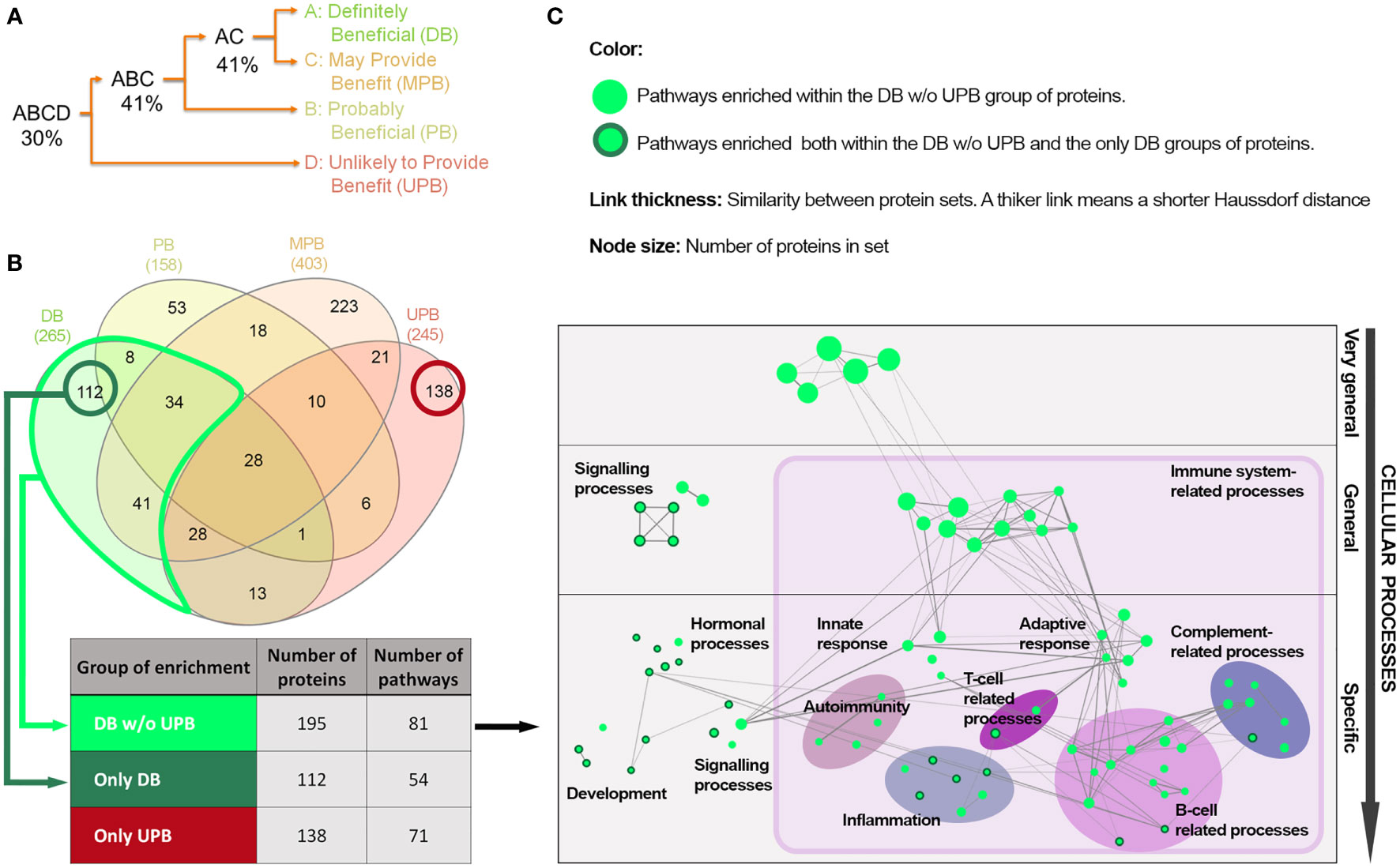
Figure 2 Protein and functional overlap between the IVIg response-based clusters of diseases. (A) Binary tree based on protein overlap among the disease clusters according to IVIg response. Percentages indicate the arithmetic mean of proportions of protein overlap of each pair of disease clusters or group of clusters in the tree branches; (B) Overlap of protein effectors between diseases with different response to IVIg (Venn diagram created with InteractiVenn (27)) and number of enriched pathways in the three protein sets defined according to the response-based clusters of diseases. the sets were defined in terms of number of proteins and enriched pathways according to the hypergeometric method (FDR q-value< 0.05); (C) Network representation of the 81 pathways enriched in the DB w/o UPB set of proteins (see Table 3) considering a Haussdorf distance< 1. IVIg: Intravenous immunoglobulin; only-DB: Protein effectors exclusively present in the IVIg ‘definitely beneficial’ cluster of diseases; DB w/o UPB: Protein effectors present in the IVIg ‘definitely beneficial’ cluster of diseases but not in the IVIg ‘unlikely to provide benefit’ cluster; only-UPB: Protein effectors exclusively present in the IVIg ‘unlikely to provide benefit’ cluster of diseases.
Taking the results of the enrichment analysis of the DB w/o UPB protein set as a reference, the results of the enrichment analysis of the only-DB protein set showed 23 common enriched pathways, while no overlap was found with the only-UPB set (Supplementary Table S3 in the S3 File). The 23 enriched pathways shared between the only-DB and DB w/o UPB sets contained mainly general signaling pathways involved in development (e.g., oocyte meiosis and regulation of dendritic differentiation), hormonal regulation (e.g., endocrine-regulated calcium reabsorption and GnRH signaling pathway), and inflammation (e.g., VEGF and cGMP-PKG signaling pathways), as well as some pathways related to B and T cells (Figure 2C, nodes circled in dark green, and Supplementary Table S3 in the S3 File).
The functional relationship between IVIg targets (Table 2) and the different diseases, defined as the effector proteins previously identified (Supplementary Table S1 in the S1 File), was tested by ANN analysis to measure the mechanistic relationship between them. This approach predicts the possibility that the studied targets modulate the set of proteins involved in the disease pathophysiology. We first tested the functional relationship between the studied conditions and IVIg targets grouped by their immune-related function (Functional group column in Table 2). The results showed that B cell-related and complement system-related targets were more likely associated with IVIg efficacy (Figure 3). To get a closer evaluation of the targets, the analysis was performed for each of them individually, focusing on the targets within the complement system and B cell-related functional groups. The analysis over the individual IVIg targets of the complement system functional group disclosed a stronger association between C3, C4B, and C5 with diseases assigned to the DB cluster than their association with disorders classified in the UPB cluster (Figure 4A). The same evaluation performed on individual targets related to B cells showed that different groups of targets were associated with different diseases (namely, FCGR3A/FCGR3B and TNFSF13/TNFSF13B) (Figure 4B).
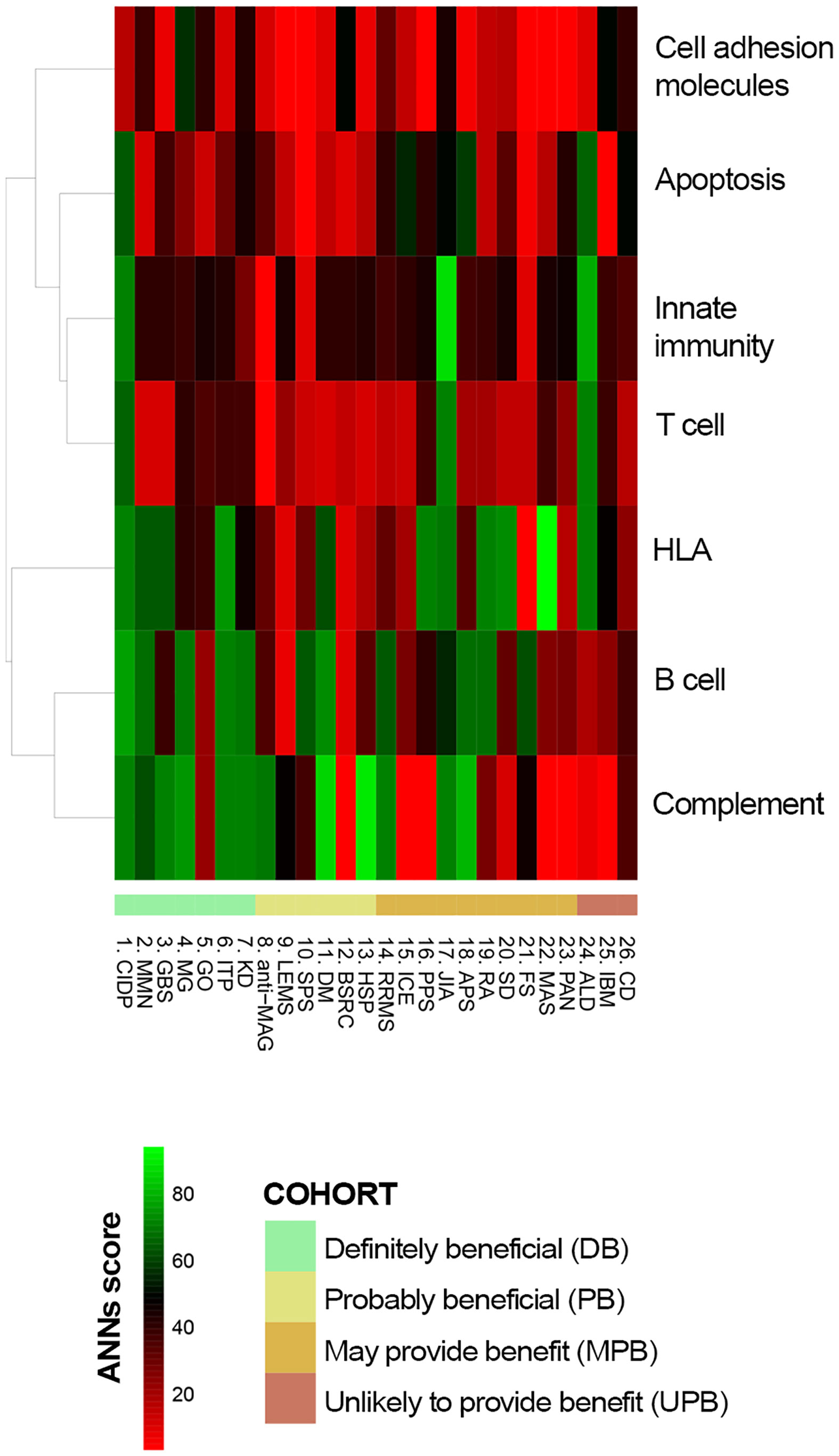
Figure 3 Predicted relationship between each studied immune-related disorder and each pathophysiological process affected by IVIg according to ANN. ANN, Artificial Neural Network; IVIg, Intravenous immunoglobulin.
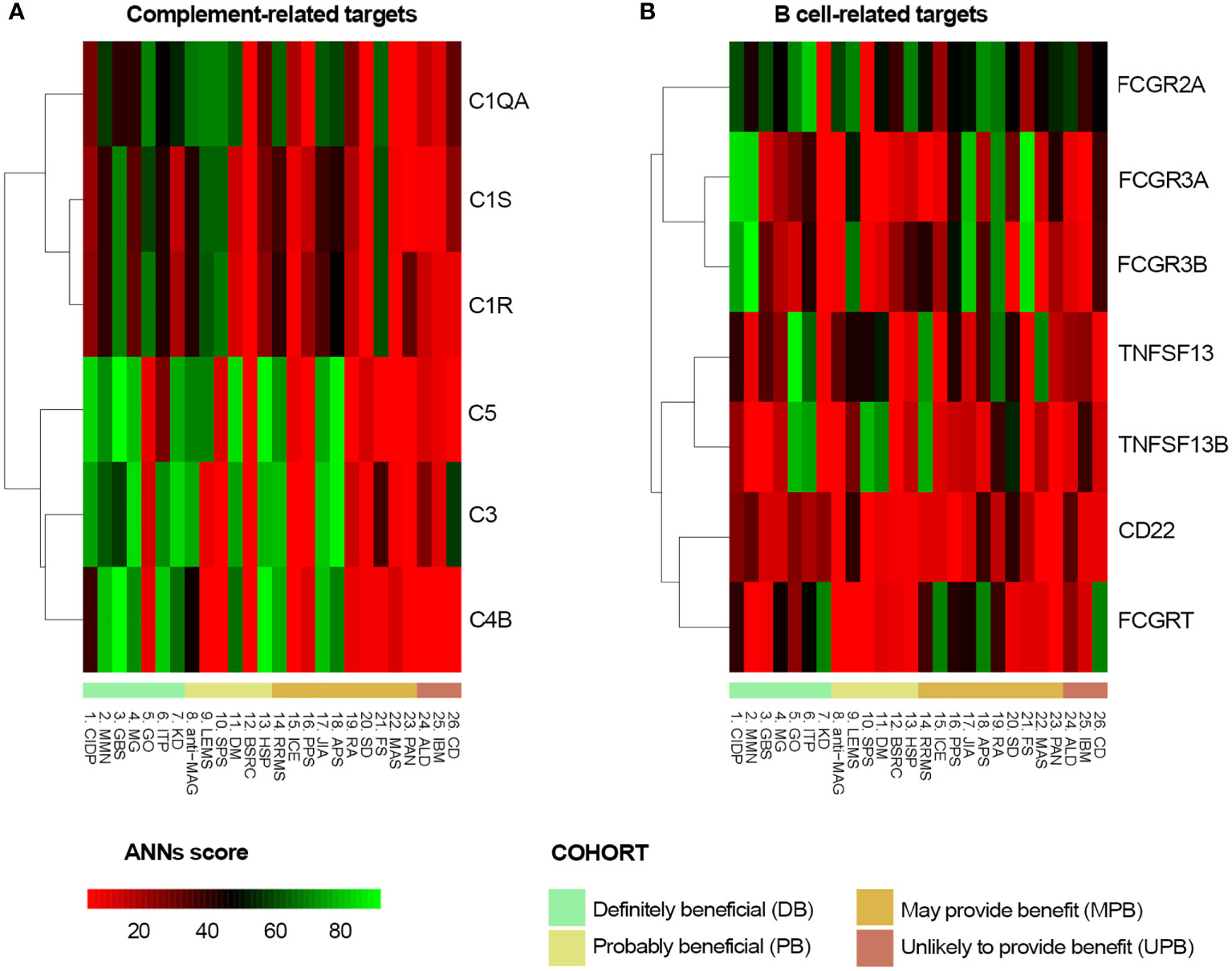
Figure 4 Predicted relationship between each studied disorder and (A) each complement system protein target of IVIg or (B) each B-cell related IVIg protein target according to ANN. ANN, Artificial Neural Network; IVIg, Intravenous immunoglobulin.
The participation in IVIg response of other targets not related to B cell or complement system processes was also explored by ANNs. In this case, the functional relationship between the complete IVIg protein target profile and each disease of the DB cluster was assessed and unveiled a high heterogeneity among diseases (Table 4). Targets associated with HLA (e.g., HLA-DPA1), innate immunity (e.g., CLEC4A), or T-cell related (e.g., CCR5) showed a consistent medium-strong (>71% ANN score) or strong (>78% ANN score) relationship for all diseases (except for MMN) and could be involved in the treatment’s efficacy. Results in Table 4 also reinforced the observation that the contribution of each B cell-related target differed for each condition. The response to IVIg in GBS, MG, and graves ophthalmopathy (GO) could not be attributed to individual B cell-related targets (all of them presented ANN scores below 71%), but complement system-related targets seemed to have a potential individual role (showing at least 71% ANN score; e.g., C5 for all diseases except GO and ITP, and C4B for all diseases as well, except for CIDP, GO, and KD).
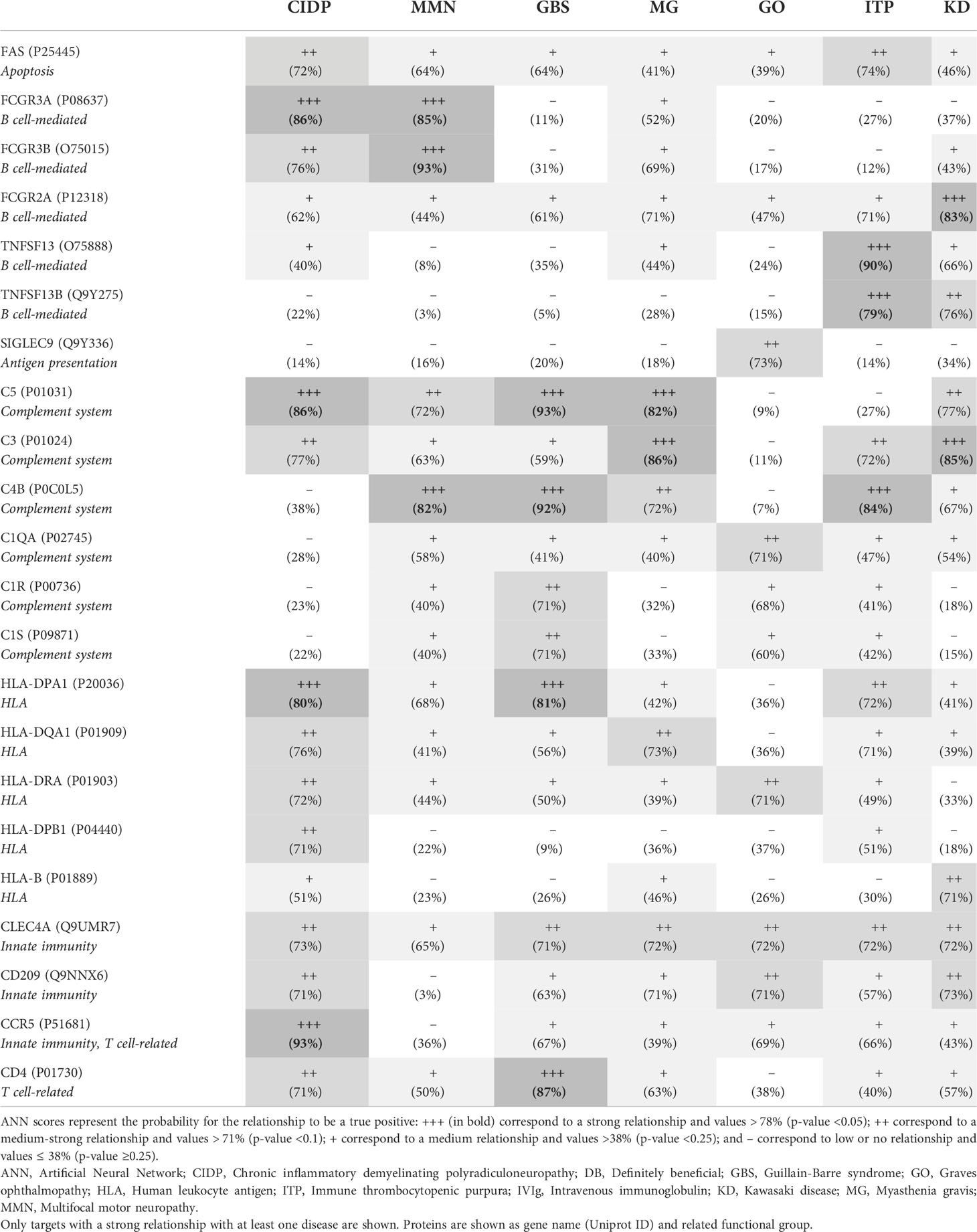
Table 4 Functional relation (ANN score) between IVIg protein targets and diseases from the DB cluster according to ANN analysis.
Our in silico systems biology-based approaches, described here, aimed to explore and compare the pathophysiology of autoimmune diseases with different degrees of clinical response to IVIg treatment. They also addressed IVIg’s mechanisms of action on those disorders responding to IVIg treatment. The results obtained allowed us to differentiate the main pathophysiological processes behind each of the IVIg response-based clusters of diseases analyzed and to suggest which IVIg targets could be involved in the successful response to this treatment.
The classification chosen here as the basis of the study (2) has been repeatedly used in recent publications (91–93), thus, supporting our election and findings. Our results showed that diseases clearly benefiting from IVIg therapy (i.e., belonging to the DB cluster) were mainly characterized by B-cell and complement system-related processes. However, our analyses did not rule out the role of other processes that could be relevant to these diseases. Furthermore, current treatments for the conditions in the DB cluster support the important role of B cell and complement system processes. B cell depletion has been stablished as a relevant target for autoimmunity disorders for long (94). For example, rituximab is a B-cell depleting monoclonal antibody that, despite not being approved for the treatment of neurological disorders, has shown a certain degree of clinical improvement in many diseases included in our DB cluster (i.e., CIDP, MMN, GO, MG) (95, 96). Similarly, eculizumab is a monoclonal antibody that inhibits terminal complement activation by binding to C5 (90); it is approved to treat MG (97) and has also been successfully used in patients with MMN (96). These examples would support the key role of B-cell and/or complement system processes in these conditions, hence, in agreement with our findings.
On the contrary, our results indicated that a minor response to IVIg treatment was likely in pathologies in which B cells were not predominantly involved. It is clear that T cell populations play a critical role in mediating autoimmunity via T cell inflammatory cytokine secretion, their help to B cells via cytokines thus driving the naïve B cells to become autoantibody secreting plasma cells. Interestingly, literature evidence indicates that IVIg modulates T cell-related processes (65, 67) and innate immunity/inflammation (1, 98). Although our results do not support that pathological alterations in these processes determine the level of response to IVIg in a specific disease, they however endorse a role for T cell- and inflammation-related IVIg targets in its therapeutic effect for some of the evaluated diseases.
When evaluating which IVIg targets might have a direct role over B cell and complement system processes through ANN analyses, we found a relevant role for the complement proteins C3, C4B, and C5 in IVIg mechanisms. In fact, it has been described that IVIg directly neutralizes these proteins through its F(ab’)2 region (44), and through the formation of complexes between IVIg and complement components (99). This neutralization mechanism would explain the impact on complement activation in several of the analyzed diseases, which in turn prevents complement-mediated tissue damage (100). Evidence supports the relevance of complement activation inhibition by IVIg in the therapeutic efficacy in autoimmune dermatological conditions, reinforce the relevance of this mechanism within IVIg therapeutic effects (101). In contrast, no common B cell-related targets were found, probably due to the great complexity of B cell pathological role in autoimmune conditions (e.g., autoantibodies production, cytokines release, autoantigens presentation to T cells) (102). Previous reports have proven that IVIg is able to modulate B-cell function and survival (103, 104). In agreement with these facts, our models showed that modulation of B-cell pathways by IVIg might involve different targets for each disease. These results are also in line with previous findings reporting that IVIg contains antibodies against a proliferation-inducing ligand and, more importantly, against B-cell activating factor involved in B-cell survival and with documented deleterious effects in B-cell mediated autoimmune diseases (36, 37). Furthermore, the survival and function of B cells can be significantly impacted by the anti-idiotypic antibodies present in IVIg (103, 105). Also, it has been reported that the anti-CD5 antibodies found in IVIg can inhibit the release of autoantibodies produced by B cells (106).
While not directly addressed in the ANN analysis (which focused on protein targets independently of the cell where they are expressed), evidence points to the role of different cell types in IVIg-mediated immunomodulation. In this sense, DCs can regulate immune responses through interaction with T and B lymphocytes and have been demonstrated to be targets of the immunomodulatory effects of IVIg (107, 108). This has been observed particularly through the inhibition of CD80 and CD86 expression (69) and by promoting a more tolerogenic phenotype (109) that is less competent in driving lymphocyte proliferation (65, 67). Furthermore, IVIg has been found to modulate DCs functions through both FcγR- and non-Fc-receptor-mediated signaling events (110). In contrast, even though DCs regulate B cell function, the effects of IVIg over DCs do not seem to affect B cells directly (111). IVIg has been reported to affect directly and indirectly Th17 and Treg function (69, 79, 112–115), and reciprocally regulate them. Treg function is central in the maintenance of immune tolerance and has been found to be reduced in patients suffering from autoimmune diseases (116). IVIg treatment has been shown to promote development or activation of Treg through diverse mechanisms (113, 117). For example, interaction of IVIg with DCs through binding of sialylated IVIg to C-type lectin receptors that induces inhibitory FcγrIIb expression on DC (56). This renders the DC tolerogenic and leads to a reduced costimulatory molecule expression and proinflammatory cytokine secretion (118), thus favoring Treg function. However, whether Fc-sialylation is critical for IVIG-mediated Treg cell expansion in humans is still unclear (117). It has also been shown that, upon IVIg treatment, DC-SIGN (dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin) signaling is able to expand Treg through prostaglandin E2 secretion in DCs (119, 120). In addition, IVIg can also modulate the production of proinflammatory cytokines by DC, which may play a role in maintaining T-cell tolerance (69). De Groot and coworkers (121) described a DC-dependent mechanism where promiscuous IgG-derived T-cell epitope peptides (Tregitopes) increases pro-tolerogenic cytokine production (IL-10) and converts naïve conventional T cells to T regulatory cells (122). Treg would then inhibit effector Th1, Th2 and Th17 cellular activity in inflammatory microenvironments and secrete anti-inflammatory cytokines (112, 123–125). Overall, the presence of Tregitopes might in part explain the success of IVIg therapy in treating autoimmune diseases (122, 126). IVIg treatment has been reported to affect natural killer cells and subsequently regulate Treg function in KD patients (127). IVIg has been shown to affect T cells chemokine production, as observed by Pigard and colleagues, thus affecting their function and compartmentalization (128). Our results position the modulation of B cells as an outstanding IVIg therapeutic mechanism. In fact, aside from antibody producers, B cells also act as APC and cytokine producing cells (129). This role of B cells as APC in regulating T cell equilibrium, Treg induction and establishment of tolerogenesis must have contributed to these findings (130–132).
Basophils have also been shown to be activated when treated with IVIg in mouse models and human studies (133–136) through indirect mechanisms that our approach was not able to discern. Notwithstanding their importance, the indirect nature of the effect and the uncertainty of the direct molecular target of IVIg over these cell types might have resulted in overlooking these mechanisms in our ANN analysis. However, these mechanisms were not highlighted either as pathologic drivers characterizing the diseases within the DB disease cluster.
In the last decade, research and development have surged on novel molecules as potential therapeutic alternatives to IVIg, aiming to overcome IVIg’s therapeutic limitations (e.g., dependence on the supply of human plasma and the large doses required) (137). These promising new therapies belong to the group of next-generation Fc receptor-targeting biologics (138): (i) Recombinant fragment crystallizable (rFc) multimers, designed to have multiple, organized, and structured IgG-Fc moieties; (ii) Neonatal Fc receptor (FcRn)-targeting therapeutics; and (iii) Fc/Fcγ receptor (FcγR)-targeting therapeutics (137). Importantly, FcRn-targeting compounds should be highlighted since our results did not suggest a major relevance of this IVIg target to achieve a response. The FcRn blocker efgartigimod is currently the compound furthest advanced in clinical trials (139–142), with some of the anti-FcRn monoclonals following not far behind (143, 144), while a modified monomeric recombinant Fc optimized for binding to all FcRns and FcγRs is also under development (145). This search for novel molecules specifically targeting Fc receptors contrasts with the results obtained in our study, which suggest that the ‘non-specificity’ of IVIg and its interaction with a broad spectrum of targets could be key to obtaining a pleiotropic effect and clinical efficacy in very different diseases. Regarding Fc multimers, the activity of these molecules relies not only on the interaction with Fc receptors but also on targeting complement proteins (146). The comparison of the activity of these new drugs with IVIg would be interesting since some of the direct targets of IVIg herein analyzed interact with IgG’s Fab region, such as CD209/DC-SIGN, C3a, and C5a anaphylotoxins (proteolytic degradation products of C3 and C5) (147, 148).
Our study was limited by the intrinsic constraints of systems biology–based modeling approaches, which include issues on information availability about drugs and diseases. First, our results were influenced by the list of diseases included in the study and the IVIg response cluster assigned to each of them. We selected only autoimmune and neuroautoimmune conditions with the strongest clinical evidence (i.e., tested in clinical trials) for their further classification, including only those tested in clinical trials. Thus, some current ‘off-label’ uses of IVIg were not considered here, as is the case of blistering autoimmune diseases (e.g., systematic lupus erythematosus, pemphigus or pemphigoid diseases). IVIg treatment has been shown to benefit patients suffering from these diseases (149–151), and is usually used in a second- or third-line setting for these conditions (77). However, the level of evidence obtained to date, according to the criteria set in the current study, prevented the inclusion of these diseases in the analysis. Of note, many of these conditions are rare and, even if clinical trials had been conducted, the small sample size could have been considered a drawback. This could have precluded their clear classification, thus finishing in the MPB or PB clusters. Consequently, future clinical evidence regarding IVIg use could modify our classification, increasing the pool of available indications in the top and bottom clusters (i.e., DB and UPB). Second, the characterization of the studied diseases may have been incomplete since many of them were rare or complex diseases in which only a few research teams are actively working at the molecular level, potentially leading to biased literature (we found less than 25 effector proteins for several characterized diseases, for instance: Stiff-person syndrome, birdshot retinochoroidopathy, postpolio syndrome, and Felty’s syndrome, see Table 1). This limitation might have affected the characterization and analyses at the protein level, perhaps with a weaker impact on the definition of the pathophysiological processes. Finally, although we reviewed all available information at the time of the study for identification of direct IVIg targets, including FcγR and non-receptor protein targets, the extreme complexity of IVIg, due to its multi-target feature at the molecular and cellular level, could have led us to underestimate the potential impact of IVIg by including an incomplete list of targets because of a dearth of evidence. Indeed, a recent study by Pipi et al. (2021) (101) unraveled potential non-receptor-mediated antioxidative mechanism for high-dose IgG, as neutrophil elastase substrate and through ROS scavenging, potentially involved in the treatment of skin autoimmunity; however, for the latter they did not detail the mechanism nor specified the direct IVIg protein target, which was a requirement for our approach. Also, considering only direct IVIg targets might have oversimplified the model, hindering the detection of effects over immune components indirectly affected by IVIg; and categorizing the identified targets in discrete function compartments, which might have oversimplified the IVIg mechanisms explored in the ANN analysis on overall processes.
However, using systems biology–based modeling approaches minimizes the impact of the potential biases intrinsic to the availability of information. These models compile and reinterpret available biological data to generate new knowledge and hypotheses while reproducing known aspects of the diseases or drugs. Our models were built considering the whole human protein network and a wide range of drug-pathology relationships, not only limited to the studied diseases or inflammatory conditions, presenting cross-validation accuracies above 80%.
Systems biology approaches combined with machine learning are becoming increasingly important for identifying new drug effects and disease mechanisms. Using these techniques, we dedicated our work to compile all available information in order to gain a better understanding of IVIg mode of action. In our analyses, diseases clearly benefiting from IVIg treatment (i.e., the DB cluster) were found to be mainly characterized by deregulated processes in B cells and the complement system. In addition, IVIg targets related to B-cell and complement system pathways seemed to be involved in the clinical response. However, targets related to other immune processes may also play an important role in the IVIg response, supporting its wide range of action through several mechanisms. Besides, although B-cell responses and complement system have a key role in diseases benefiting from IVIg, protein targets involved in such processes are not necessarily the same in those diseases. Our results support further investigations on the role of IVIg in diseases where B cells and the complement system are relevantly involved and for which no evidence has been gathered yet or is inconclusive, for instance, those classified here as PB and MPB. Finally, since the level of relationship varied between proteins included in the DB cluster and different diseases, IVIg appeared to have a pleiotropic effect that may involve the collaborative participation of several targets. Indeed, the weight of each target in treatment efficacy may be different for each condition. Therefore, IVIg’s broad spectrum of targets and ‘non-specificity’ could be key to its efficacy in very different diseases.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
JT, EC-G, SC, and SK have significantly contributed to setting up fundamental questions about IVIg. JT, EC-G, SC, and TS contributed to conceptualize the study. TS and CS-V contributed to study design. EC-G, SC, and TS contributed to supervise and manage the project. CS-V and HB contributed to formal analysis, data curation, and visualization. CS-V contributed to the methodology and investigation. SK contributed to the interpretation of data. CS-V drafted the manuscript and the rest of authors contributed to the critical revision of the text. All authors have given their final approval to the version submitted for publication.
The authors thank Dirk Büscher for his valuable contributions to the conceptualization of the study and Matías Rey (BCN Medical Writing, Spain) for his assistance in writing and copyediting the manuscript.
EC-G, SC, and JT are full time employees at Grifols. CS-V and HB are full time employees at Anaxomics Biotech. TS was a full time-employee at Anaxomics Biotech at the time of the study. SK has received fees for lectures and reviewing research proposals from CSL-Behring and Grifols.
The authors declare that this study received funding from Grifols (including medical writing assistance, and the article processing fees). The funder participated in the original idea, writing of this article and decision to submit it for publication, but was not involved in the study design, collection, analysis and interpretation of data.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.901872/full#supplementary-material
1. Galeotti C, Kaveri SV, Bayry J. IVIG-mediated effector functions in autoimmune and inflammatory diseases. Int Immunol (2017) 29:491–8. doi: 10.1093/intimm/dxx039
2. Perez EE, Orange JS, Bonilla F, Chinen J, Chinn IK, Dorsey M, et al. Update on the use of immunoglobulin in human disease: A review of evidence. J Allergy Clin Immunol (2016) 139:S1–S46. doi: 10.1016/j.jaci.2016.09.023
3. European Medicines Agency. Guideline on core SmPC for human normal immunoglobulin for intravenous administration (IVIg) (2020). Available at: https://www.ema.europa.eu/en/documents/scientific-guideline/draft-guideline-core-smpc-human-normal-immunoglobulin-intravenous-administration-ivig-rev-6_en.pdf.
4. Hartung HP, Mouthon L, Ahmed R, Jordan S, Laupland KB, Jolles S. Clinical applications of intravenous immunoglobulins (IVIg)–beyond immunodeficiencies and neurology. Clin Exp Immunol (2009) 158 Suppl 1:23–33. doi: 10.1111/J.1365-2249.2009.04024.X
5. Dalakas MC, Sonies B, Dambrosia J, Sekul E, Cupler E, Sivakumar K. Treatment of inclusion-body myositis with IVIg: A double-blind, placebo-controlled study. Neurology (1997) 48:712–6. doi: 10.1212/WNL.48.3.712
6. Dalakas MC, Koffman B, Fujii M, Spector S, Sivakumar K, Cupler E. A controlled study of intravenous immunoglobulin combined with prednisone in the treatment of IBM. Neurology (2001) 56:323–7. doi: 10.1212/WNL.56.3.323
7. Bendtzen K, Hansen MB, Ross C, Svenson M. High-avidity autoantibodies to cytokines. Immunol Today (1998) 19:209–11. doi: 10.1016/S0167-5699(98)01252-3
8. Nagelkerke SQ, Kuijpers TW. Immunomodulation by IVIg and the role of fc-gamma receptors: Classic mechanisms of action after all? Front Immunol (2015) 5:674. doi: 10.3389/FIMMU.2014.00674
9. Lünemann JD, Quast I, Dalakas MC. Efficacy of intravenous immunoglobulin in neurological diseases. Neurotherapeutics (2016) 13:34–46. doi: 10.1007/s13311-015-0391-5
10. Chaigne B, Mouthon L. Mechanisms of action of intravenous immunoglobulin. Transfus Apher Sci (2017) 56:45–9. doi: 10.1016/j.transci.2016.12.017
11. Barahona Afonso AF, Pires João CM. The production processes and biological effects of intravenous immunoglobulin. Biomolecules (2016) 6:15. doi: 10.3390/biom6010015
12. Lünemann JD, Nimmerjahn F, Dalakas MC. Intravenous immunoglobulin in neurology-mode of action and clinical efficacy. Nat Rev Neurol (2015) 11:80–9. doi: 10.1038/nrneurol.2014.253
13. Bayry J, Lacroix-Desmazes S, Donkova-Petrini V, Carbonneil C, Misra N, Lepelletier Y, et al. Natural antibodies sustain differentiation and maturation of human dendritic cells. Proc Natl Acad Sci U.S.A. (2004) 101:14210–5. doi: 10.1073/pnas.0402183101
14. Jorba G, Aguirre-Plans J, Junet V, Segú-Vergés C, Ruiz JL, Pujol A, et al. In-silico simulated prototype-patients using TPMS technology to study a potential adverse effect of sacubitril and valsartan. PloS One (2020) 15:e0228926. doi: 10.1371/journal.pone.0228926
15. Lozano ML, Segú-Vergés C, Coma M, Álvarez-Roman MT, González-Porras JR, Gutiérrez L, et al. Elucidating the mechanism of action of the attributed immunomodulatory role of eltrombopag in primary immune thrombocytopenia: An in silico approach. Int J Mol Sci (2021) 22:6907. doi: 10.3390/ijms22136907
16. Rowley AH. Can a systems biology approach unlock the mysteries of Kawasaki disease? Wiley Interdiscip Rev Syst Biol Med (2013) 5:221–9. doi: 10.1002/wsbm.1202
17. Wang T, Liu G, Lin H. A machine learning approach to predict intravenous immunoglobulin resistance in Kawasaki disease patients: A study based on a southeast China population. PloS One (2020) 15:e0237321. doi: 10.1371/journal.pone.0237321
18. Zhang Y, Yu H, Dong R, Ji X, Li F. Application prospect of artificial intelligence in rehabilitation and management of myasthenia gravis. BioMed Res Int (2021), 5592472. doi: 10.1155/2021/5592472
19. Rambabu N, Mathew MJ, Kaveri SV, Bayry J. Boolean analysis of the transcriptomic data to identify novel biomarkers of IVIG response. Autoimmun Rev (2021) 20:102850. doi: 10.1016/j.autrev.2021.102850
20. Artigas L, Coma M, Matos-Filipe P, Aguirre-Plans J, Farrés J, Valls R, et al. In-silico drug repurposing study predicts the combination of pirfenidone and melatonin as a promising candidate therapy to reduce SARS-CoV-2 infection progression and respiratory distress caused by cytokine storm. PloS One (2020) 15:e0240149. doi: 10.1371/journal.pone.0240149
21. Segú-Vergés C, Coma M, Kessel C, Smeets S, Foell D, Aldea A. Application of systems biology-based in silico tools to optimize treatment strategy identification in still’s disease. Arthritis Res Ther (2021) 23:126. doi: 10.1186/s13075-021-02507-w
22. Romeo-Guitart D, Forés J, Herrando-Grabulosa M, Valls R, Leiva-Rodríguez T, Galea E, et al. Neuroprotective drug for nerve trauma revealed using artificial intelligence. Sci Rep (2018) 8:1–15. doi: 10.1038/s41598-018-19767-3
23. U.S. Food and Drug Administration. Immune globulin intravenous (IGIV) indications (2018). Available at: https://www.fda.gov/vaccines-blood-biologics/approved-blood-products/immune-globulin-intravenous-igiv-indications (Accessed December 3, 2021).
24. European Medicines Agency. Guideline on core SmPC for human normal immunoglobulin for intravenous administration (IVIg). EMA/CHMP/BPWP/94038/2007 rev 5 (2018). Available at: https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-core-smpc-human-normal-immunoglobulin-intravenous-administration-ivig-rev-5_en.pdf.
25. Alabdali M, Barnett C, Katzberg H, Breiner A, Bril V. Intravenous immunoglobulin as treatment for myasthenia gravis: Current evidence and outcomes. Expert Rev Clin Immunol (2014) 10:1659–65. doi: 10.1586/1744666X.2014.971757
26. Gajdos P, Tranchant C, Clair B, Bolgert F, Eymard B, Stojkovic T, et al. Treatment of myasthenia gravis exacerbation with intravenous immunoglobulin: A randomized double-blind clinical trial. Arch Neurol (2005) 62:1689–93. doi: 10.1001/archneur.62.11.1689
27. Dalakas MC. Progress in the therapy of myasthenia gravis: Getting closer to effective targeted immunotherapies. Curr Opin Neurol (2020) 33:545–52. doi: 10.1097/WCO.0000000000000858
28. Wishart DS, Feunang YD, Guo AC, Lo EJ, Marcu A, Grant JR, et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res (2018) 46:D1074–82. doi: 10.1093/nar/gkx1037
29. Szklarczyk D, Santos A, Von Mering C, Jensen LJ, Bork P, Kuhn M. STITCH 5: Augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Res (2016) 44:D380–4. doi: 10.1093/nar/gkv1277
30. Hecker N, Ahmed J, Von Eichborn J, Dunkel M, Macha K, Eckert A, et al. SuperTarget goes quantitative: Update on drug-target interactions. Nucleic Acids Res (2012) 40:D1113–7. doi: 10.1093/nar/gkr912
31. Séïté JF, Cornec D, Renaudineau Y, Youinou P, Mageed RA, Hillion S. IVIg modulates BCR signaling through CD22 and promotes apoptosis in mature human b lymphocytes. Blood (2010) 116:1698–704. doi: 10.1182/blood-2009-12-261461
32. Bouhlal H, Martinvalet D, Teillaud JL, Fridman C, Kazatchkine MD, Bayry J, et al. Natural autoantibodies to fcγ receptors in intravenous immunoglobulins. J Clin Immunol (2014) 34 Suppl 1, 4–11. doi: 10.1007/S10875-014-0019-2
33. Hansen RJ, Balthasar JP. Intravenous immunoglobulin mediates an increase in anti-platelet antibody clearance via the FcRn receptor. Thromb Haemost (2002) 88:898–9. doi: 10.1055/s-0037-1613331
34. Dalakas MC, Spaeth PJ. The importance of FcRn in neuro-immunotherapies: From IgG catabolism, FCGRT gene polymorphisms, IVIg dosing and efficiency to specific FcRn inhibitors. Ther Adv Neurol Disord (2021) 14, 1–7. doi: 10.1177/1756286421997381
35. Li N, Zhao M, Hilario-Vargas J, Prisayanh P, Warren S, Diaz LA, et al. Complete FcRn dependence for intravenous ig therapy in autoimmune skin blistering diseases. J Clin Invest (2005) 115:3440–50. doi: 10.1172/JCI24394
36. Le Pottier L, Bendaoud B, Dueymes M, Daridon C, Youinou P, Shoenfeld Y, et al. BAFF, a new target for intravenous immunoglobulin in autoimmunity and cancer. J Clin Immunol (2007) 27:257–65. doi: 10.1007/s10875-007-9082-2
37. Le Pottier L, Sapir T, Bendaoud B, Youinou P, Shoenfeld Y, Pers JO. Intravenous immunoglobulin and cytokines: Focus on tumor necrosis factor family members BAFF and APRIL. Ann N Y Acad Sci (2007) 1110:426–32. doi: 10.1196/annals.1423.044
38. Ritter C, Förster D, Albrecht P, Hartung HP, Kieseier BC, Lehmann HC. IVIG regulates BAFF expression in patients with chronic inflammatory demyelinating polyneuropathy (CIDP). J Neuroimmunol (2014) 274:225–9. doi: 10.1016/j.jneuroim.2014.06.007
39. von Gunten S, Vogel M, Schaub A, Stadler BM, Miescher S, Crocker PR, et al. Intravenous immunoglobulin preparations contain anti–Siglec-8 autoantibodies. J Allergy Clin Immunol (2007) 119:1005–11. doi: 10.1016/J.JACI.2007.01.023
40. Von Gunten S, Schaub A, Vogel M, Stadler BM, Miescher S, Simon HU. Immunologic and functional evidence for anti-Siglec-9 autoantibodies in intravenous immunoglobulin preparations. Blood (2006) 108:4255–9. doi: 10.1182/BLOOD-2006-05-021568
41. Lin RY, Racis SP. In vivo reduction of circulating C1q binding immune complexes by intravenous gammaglobulin administration. Int Arch Allergy Appl Immunol (1986) 79:286–90. doi: 10.1159/000233988
42. Fiane AE, Videm V, Mellbye OJ, Foerster A, Geiran OR, Gladhaug IP, et al. Immunoglobulin prolongs survival of pig kidneys perfused ex vivo with human blood. Scand J Immunol (1998) 47:568–74. doi: 10.1046/J.1365-3083.1998.00341.X
43. Mollnes TE, Høgåsen K, Hoaas BF, Michaelsen TE, Garred P, Harboe M. Inhibition of complement-mediated red cell lysis by immunoglobulins is dependent on the IG isotype and its C1 binding properties. Scand J Immunol (1995) 41:449–56. doi: 10.1111/J.1365-3083.1995.TB03591.X
44. Dalakas MC, Alexopoulos H, Spaeth PJ. Complement in neurological disorders and emerging complement-targeted therapeutics. Nat Rev Neurol (2020) 16:601–17. doi: 10.1038/s41582-020-0400-0
45. Mollnes TE, Høgåsen K, De Carolis C, Vaquero E, Nielsen EW, Fontana L, et al. High-dose intravenous immunoglobulin treatment activates complement. vivo Scand J Immunol (1998) 48:312–7. doi: 10.1046/j.1365-3083.1998.00386.x
46. Frank MM, Basta M, Fries LF. The effects of intravenous immune globulin on complement-dependent immune damage of cells and tissues. Clin Immunol Immunopathol (1992) 62, S82–S86. doi: 10.1016/0090-1229(92)90045-P
47. Esen F, Orhun G, Ozcan PE, Senturk E, Kucukerden M, Giris M, et al. Neuroprotective effects of intravenous immunoglobulin are mediated through inhibition of complement activation and apoptosis in a rat model of sepsis. Intensive Care Med Exp (2017) 5, 1–12. doi: 10.1186/s40635-016-0114-1
48. Konrad S, Baumann U, Schmidt RE, Gessner JE. Intravenous immunoglobulin (IVIG)-mediated neutralisation of C5a: A direct mechanism of IVIG in the maintenance of a high FcγRIIB to FcγRIII expression ratio on macrophages. Br J Haematol (2006) 134:345–7. doi: 10.1111/J.1365-2141.2006.06185.X
49. Basta M, Fries LF, Frank MM. High doses of intravenous ig inhibit in vitro uptake of C4 fragments onto sensitized erythrocytes. Blood (1991) 77:376–80. doi: 10.1182/blood.V77.2.376.376
50. Kaveri S, Vassilev T, Hurez V, Lengagne R, Lefranc C, Cot S, et al. Antibodies to a conserved region of HLA class I molecules, capable of modulating CD8 T cell-mediated function, are present in pooled normal immunoglobulin for therapeutic use. J Clin Invest (1996) 97:865–9. doi: 10.1172/JCI118488
51. Prasad NK, Papoff G, Zeuner A, Bonnin E, Kazatchkine MD, Ruberti G, et al. Therapeutic preparations of normal polyspecific IgG (IVIg) induce apoptosis in human lymphocytes and monocytes: A novel mechanism of action of IVIg involving the fas apoptotic pathway. J Immunol (1998) 161:3781–90.
52. Fiebiger BM, Maamary J, Pincetic A, Ravetch JV. Protection in antibody- and T cell-mediated autoimmune diseases by antiinflammatory IgG fcs requires type II FcRs. Proc Natl Acad Sci U.S.A. (2015) 112:E2385–94. doi: 10.1073/pnas.1505292112
53. Portman MA, Wiener HW, Silva M, Shendre A, Shrestha S. DC-SIGN gene promoter variants and IVIG treatment response in Kawasaki disease. Pediatr Rheumatol (2013) 11:32. doi: 10.1186/1546-0096-11-32
54. Anthony RM, Ravetch JV. A novel role for the IgG fc glycan: The anti-inflammatory activity of sialylated IgG fcs. J Clin Immunol (2010) 30, 9–14. doi: 10.1007/s10875-010-9405-6
55. Yu HR, Chang WP, Wang L, Lin YJ, Di LC, KD Y, et al. DC-SIGN (CD209) promoter -336 A/G (rs4804803) polymorphism associated with susceptibility of Kawasaki disease. Sci World J (2012) 2012, 1–5. doi: 10.1100/2012/634835
56. Massoud AH, Yona M, Xue D, Chouiali F, Alturaihi H, Ablona A, et al. Dendritic cell immunoreceptor: A novel receptor for intravenous immunoglobulin mediates induction of regulatory T cells. J Allergy Clin Immunol (2014) 133, 853–63. doi: 10.1016/j.jaci.2013.09.029
57. Bruckner C, Lehmann C, Dudziak D, Nimmerjahn F. Sweet SIGNs: IgG glycosylation leads the way in IVIG-mediated resolution of inflammation. Int Immunol (2017) 29:499–509. doi: 10.1093/intimm/dxx053
58. Bouhlal H, Hocini H, Quillent-Grégoire C, Donkova V, Rose S, Amara A, et al. Antibodies to c-c chemokine receptor 5 in normal human IgG block infection of macrophages and lymphocytes with primary R5-tropic strains of HIV-1. J Immunol (2001) 166:7606–11. doi: 10.4049/JIMMUNOL.166.12.7606
59. Marchalonis JJ, Kaymaz H, Schluter SF, Yocum DE. Naturally occurring human autoantibodies to defined T-cell receptor and light chain peptides. Adv Exp Med Biol (1994) 347:135–45. doi: 10.1007/978-1-4615-2427-4_14
60. Marchalonis JJ, Kaymaz H, Dedeoglu F, Schluter SF, Yocum DE, Edmundson AB. Human autoantibodies reactive with synthetic autoantigens from T-cell receptor beta chain. Proc Natl Acad Sci U.S.A. (1992) 89:3325–9. doi: 10.1073/PNAS.89.8.3325
61. Vassilev T, Gelin C, Kaveri SV, Zilber MT, Boumsell L, Kazatchkine MD. Antibodies to the CD5 molecule in normal human immunoglobulins for therapeutic use (intravenous immunoglobulins, IVIg). Clin Exp Immunol (1993) 92:369–72. doi: 10.1111/J.1365-2249.1993.TB03407.X
62. Hurez V, Kaveri SV, Mouhoub A, Dietrich G, Mani JC, Klatzmann D, et al. Anti-CD4 activity of normal human immunoglobulin G for therapeutic use. (Intravenous immunoglobulin IVIg) Ther Immunol (1994) 1:269–77.
63. Anh-Tu Hoa S, Hudson M. Critical review of the role of intravenous immunoglobulins in idiopathic inflammatory myopathies. Semin Arthritis Rheum (2017) 46:488–508. doi: 10.1016/j.semarthrit.2016.07.014
64. Corbi AL, Sanchez-Ramon S, Dominguez-Soto A. The potential of intravenous immunoglobulins for cancer therapy: A road that is worth taking? Immunotherapy (2016) 8:601–12. doi: 10.2217/imt.16.9
65. Kuo HC, Hsu YW, Wu MS, Chien SC, Liu SF, Chang WC. Intravenous immunoglobulin, pharmacogenomics, and Kawasaki disease. J Microbiol Immunol Infect (2016) 49:1–7. doi: 10.1016/j.jmii.2014.11.001
66. Imbach P, Lazarus AH, Kuhne T. Intravenous immunoglobulins induce potentially synergistic immunomodulations in autoimmune disorders. Vox Sang (2010) 98:385–94. doi: 10.1111/j.1423-0410.2009.01264.x
67. Chen Y, Wang C, Xu F, Ming F, Zhang H. Efficacy and tolerability of intravenous immunoglobulin and subcutaneous immunoglobulin in neurologic diseases. Clin Ther (2019) 41:2112–36. doi: 10.1016/j.clinthera.2019.07.009
68. Mitrevski M, Marrapodi R, Camponeschi A, Cavaliere FM, Lazzeri C, Todi L, et al. Intravenous immunoglobulin and immunomodulation of b-cell - in vitro and in vivo effects. Front Immunol (2015) 6:4. doi: 10.3389/fimmu.2015.00004
69. Kaufman GN, Massoud AH, Dembele M, Yona M, Piccirillo CA, Mazer BD. Induction of regulatory T cells by intravenous immunoglobulin: A bridge between adaptive and innate immunity. Front Immunol (2015) 6:469. doi: 10.3389/fimmu.2015.00469
70. Thom V, Arumugam TV, Magnus T, Gelderblom M. Therapeutic potential of intravenous immunoglobulin in acute brain injury. Front Immunol (2017) 8:875. doi: 10.3389/fimmu.2017.00875
71. Spurlock NK, Prittie JE. A review of current indications, adverse effects, and administration recommendations for intravenous immunoglobulin. J Vet Emerg Crit Care (San Antonio) (2011) 21:471–83. doi: 10.1111/j.1476-4431.2011.00676.x
72. Forbat E, Ali FR, Al-Niaimi F. Intravenous immunoglobulins in dermatology. part 1: biological mechanisms and methods of administration. Clin Exp Dermatol (2018) 43:513–7. doi: 10.1111/ced.13553
73. Esen F, Ozcan PE, Tuzun E, Boone MD. Mechanisms of action of intravenous immunoglobulin in septic encephalopathy. Rev Neurosci (2018) 29:417–23. doi: 10.1515/revneuro-2017-0065
74. Patwardhan A. The value of intravenous immunoglobulin therapy in idiopathic inflammatory myositis in the current transformed era of biologics. Cureus (2020) 12:e7049. doi: 10.7759/cureus.7049
75. Martinez T, Garcia-Robledo JE, Plata I, Urbano MA, Posso-Osorio I, Rios-Serna LJ, et al. Mechanisms of action and historical facts on the use of intravenous immunoglobulins in systemic lupus erythematosus. Autoimmun Rev (2019) 18:279–86. doi: 10.1016/j.autrev.2018.10.002
76. Kornberg A. Intravenous immunoglobulins in liver transplant patients: Perspectives of clinical immune modulation. World J Hepatol (2015) 7:1494–508. doi: 10.4254/wjh.v7.i11.1494
77. Hoffmann JHO, Enk AH. High-dose intravenous immunoglobulin in skin autoimmune disease. Front Immunol (2019) 10:1090. doi: 10.3389/fimmu.2019.01090
78. Ballow M. Mechanisms of immune regulation by IVIG. Curr Opin Allergy Clin Immunol (2014) 14:509–15. doi: 10.1097/ACI.0000000000000116
79. Kaveri SV. Intravenous immunoglobulin: exploiting the potential of natural antibodies. Autoimmun Rev (2012) 11:792–4. doi: 10.1016/j.autrev.2012.02.006
80. Prabagar MG, Choi HJ, Park JY, Loh S, Kang YS. Intravenous immunoglobulin-mediated immunosuppression and the development of an IVIG substitute. Clin Exp Med (2014) 14:361–73. doi: 10.1007/s10238-013-0255-4
81. Seite JF, Guerrier T, Cornec D, Jamin C, Youinou P, Hillion S. TLR9 responses of b cells are repressed by intravenous immunoglobulin through the recruitment of phosphatase. J Autoimmun (2011) 37:190–7. doi: 10.1016/j.jaut.2011.05.014
82. Bishop C. Pattern recognition and machine learning. 1st ed. New York: Springer-Verlag (2006). 738 p.
83. Pujol A, Mosca R, Farrés J, Aloy P. Unveiling the role of network and systems biology in drug discovery. Trends Pharmacol Sci (2010) 31:115–23. doi: 10.1016/j.tips.2009.11.006
84. Kim S, Chen J, Cheng T, Gindulyte A, He J, He S, et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res (2021) 49:D1388–95. doi: 10.1093/nar/gkaa971
85. Law V, Knox C, Djoumbou Y, Jewison T, Guo AC, Liu Y, et al. DrugBank 4.0: Shedding new light on drug metabolism. Nucleic Acids Res (2014) 42, D1091–97. doi: 10.1093/nar/gkt1068
86. Heberle H, Meirelles GV, da Silva FR, Telles GP, Minghim R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinf (2015) 16:169. doi: 10.1186/s12859-015-0611-3
87. Rivals I, Personnaz L, Taing L, Potier MC. Enrichment or depletion of a GO category within a class of genes: Which test? Bioinformatics (2007) 23:401–7. doi: 10.1093/bioinformatics/btl633
88. Kanehisa M, Goto S, Kawashima S, Nakaya A. The KEGG databases at GenomeNet. Nucleic Acids Res (2002) 30:42–6. doi: 10.1093/nar/30.1.42
89. The Gene Ontology C. The gene ontology resource: 20 years and still GOing strong. Nucleic Acids Res (2019) 47:D330–8. doi: 10.1093/nar/gky1055
90. Benjamini Y, Hochberg Y. Controlling the false discovery rate - a practical and powerful approach to multiple testing. J R Stat Soc Ser B (1995) 57:289–300. doi: 10.2307/2346101
91. Oaklander AL, Lunn MPT, Hughes RA, van Schaik IN, Frost C, Chalk CH. Treatments for chronic inflammatory demyelinating polyradiculoneuropathy (CIDP): An overview of systematic reviews. Cochrane Database Syst Rev (2017) 2017: 1–33. doi: 10.1002/14651858.CD010369.pub2
92. Song F, Al-Samkari H. Management of adult patients with immune thrombocytopenia (Itp): A review on current guidance and experience from clinical practice. J Blood Med (2021) 12:653–64. doi: 10.2147/JBM.S259101
93. Farini A, Villa C, Tripodi L, Legato M, Torrente Y. Role of immunoglobulins in muscular dystrophies and inflammatory myopathies. Front Immunol (2021) 12:666879. doi: 10.3389/fimmu.2021.666879
94. Blank M, Shoenfeld Y. B cell targeted therapy in autoimmunity. J Autoimmun (2007) 28:62–8. doi: 10.1016/j.jaut.2007.02.001
95. Whittam DH, Tallantyre EC, Jolles S, Huda S, Moots RJ, Kim HJ, et al. Rituximab in neurological disease: Principles, evidence and practice. Pract Neurol (2019) 19:5–20. doi: 10.1136/practneurol-2018-001899
96. Eid L, Coste-Verdier V, Longueville E, Ribeiro E, Nicolescu-Catargi B, Korobelnik JF. The effects of rituximab on graves’orbitopathy: A retrospective study of 14 patients. Eur J Ophthalmol (2020) 30:1008–13. doi: 10.1177/1120672119845224
97. Muppidi S, Utsugisawa K, Benatar M, Murai H, Barohn RJ, Illa I, et al. Long-term safety and efficacy of eculizumab in generalized myasthenia gravis. Muscle Nerve (2019) 60:14–24. doi: 10.1002/mus.26447
98. Durandy A, Kaveri SV, Kuijpers TW, Basta M, Miescher S, Ravetch JV, et al. Intravenous immunoglobulins–understanding properties and mechanisms. Clin Exp Immunol (2009) 158 Suppl:2–13. doi: 10.1111/j.1365-2249.2009.04022.x
99. Basta M, Dalakas MC. High-dose intravenous immunoglobulin exerts its beneficial effect in patients with dermatomyositis by blocking endomysial deposition of activated complement fragments. J Clin Invest (1994) 94:1729–35. doi: 10.1172/JCI117520
100. Brennan FH, Kurniawan ND, Vukovic J, Bartlett PF, Käsermann F, Arumugam TV, et al. IVIg attenuates complement and improves spinal cord injury outcomes in mice. Ann Clin Transl Neurol (2016) 3:495. doi: 10.1002/ACN3.318
101. Pipi E, Kasprick A, Iwata H, Goletz S, Hundt JE, Sadeghi H, et al. Multiple modes of action mediate the therapeutic effect of intravenous IgG in experimental epidermolysis bullosa acquisita. J Invest Dermatol (2021) 142: 1552–64. doi: 10.1016/j.jid.2021.08.448
102. Lund FE, Garvy BA, Randall TD, Harris DP. Regulatory roles for cytokine-producing b cells in infection and autoimmune disease. Curr Dir Autoimmun (2005) 8:25–54. doi: 10.1159/000082086
103. Kazatchkine MD, Dietrich G, Hurez V, Ronda N, Bellon B, Rossi F, et al. V Region-mediated selection of autoreactive repertoires by intravenous immunoglobulin (i.v.Ig). Immunol Rev (1994) 139:79–107. doi: 10.1111/j.1600-065X.1994.tb00858.x
104. Séïté JF, Goutsmedt C, Youinou P, Pers JO, Hillion S. Intravenous immunoglobulin induces a functional silencing program similar to anergy in human b cells. J Allergy Clin Immunol (2014) 133:181–188.e9. doi: 10.1016/j.jaci.2013.08.042
105. Dietrich G, Kaveri S-V, Kazatchkine MD. Modulation of autoimmunity by intravenous immune globulin through interaction with the function of the immune/ldiotypic network. Clin immunol Immunopathy (1992) 62:S73–81. doi: 10.1016/0090-1229(92)90044-O
106. Hartung H-P. Advances in the understanding of the mechanism of action of IVIg. J Neurol (2008) 255:3–6. doi: 10.1007/s00415-008-3002-0
107. Misra N, Bayary J, Dasgupta S, Ephrem A, Van Huyen JPD, Delignat S, et al. Intravenous immunoglobulin and dendritic cells. Clin Rev Allergy Immunol (2005) 29:201–5. doi: 10.1385/CRIAI:29:3:201
108. Wang N, Chen Z, Zhang F, Zhang Q, Sun L, Lv H, et al. Intravenous immunoglobulin therapy restores the quantity and phenotype of circulating dendritic cells and CD4+ T cells in children with acute Kawasaki disease. Front Immunol (2022) 13:802690. doi: 10.3389/fimmu.2022.802690
109. Othy S, Bruneval P, Topçu S, Dugail I, Delers F, Lacroix-Desmazes S, et al. Effect of IVIg on human dendritic cell-mediated antigen uptake and presentation: role of lipid accumulation. J Autoimmun (2012) 39:168–72. doi: 10.1016/J.JAUT.2012.05.013
110. Bayry J, Lacroix-Desmazes S, Carbonneil C, Misra N, Donkova V, Pashov A, et al. Inhibition of maturation and function of dendritic cells by intravenous immunoglobulin. Blood (2003) 101:758–65. doi: 10.1182/BLOOD-2002-05-1447
111. Maddur MS, Hegde P, Sharma M, Kaveri SV, Bayry J. B cells are resistant to immunomodulation by “IVIg-educated” dendritic cells. Autoimmun Rev (2011) 11:154–6. doi: 10.1016/j.autrev.2011.08.004
112. Maddur MS, Vani J, Hegde P, Lacroix-Desmazes S, Kaveri SV, Bayry J. Inhibition of differentiation, amplification, and function of human T H17 cells by intravenous immunoglobulin. J Allergy Clin Immunol (2011) 127:823–830.e7. doi: 10.1016/j.jaci.2010.12.1102
113. Bayry J, Mouthon L, Kaveri SV. Intravenous immunoglobulin expands regulatory T cells in autoimmune rheumatic disease. J Rheumatol (2012) 39:450–1. doi: 10.3899/jrheum.111123
114. Ephrem A, Chamat S, Miquel C, Fisson S, Mouthon L, Caligiuri G, et al. Expansion of CD4+CD25+ regulatory T cells by intravenous immunoglobulin: A critical factor in controlling experimental autoimmune encephalomyelitis. Blood (2008) 111:715–22. doi: 10.1182/blood-2007-03-079947
115. Kessel A, Ammuri H, Peri R, Pavlotzky ER, Blank M, Shoenfeld Y, et al. Intravenous immunoglobulin therapy affects T regulatory cells by increasing their suppressive function. J Immunol (2007) 179:5571–5. doi: 10.4049/jimmunol.179.8.5571
116. Goswami TK, Singh M, Dhawan M, Mitra S, Bin ET, AA R, et al. Regulatory T cells (Tregs) and their therapeutic potential against autoimmune disorders–advances and challenges. Hum Vaccines Immunother (2022) 18: 1–16. doi: 10.1080/21645515.2022.2035117
117. Maddur MS, Kaveri SV. Circulating normal IgG as stimulator of regulatory T Cells: Lessons from intravenous immunoglobulin. Trends Immunol (2017) 38:789–92. doi: 10.1016/j.it.2017.08.008
118. Yamazaki S, Dudziak D, Heidkamp GF, Fiorese C, Bonito AJ, Inaba K, et al. CD8 + CD205 + splenic dendritic cells are specialized to induce Foxp3 + regulatory T cells. J Immunol (2008) 181:6923–33. doi: 10.4049/jimmunol.181.10.6923
119. Trinath J, Hegde P, Sharma M, Maddur MS, Rabin M, Vallat JM, et al. Intravenous immunoglobulin expands regulatory T cells via induction of cyclooxygenase-2-dependent prostaglandin E2 in human dendritic cells. Blood (2013) 122:1419–27. doi: 10.1182/blood-2012-11-468264
120. Kalinski P. Regulation of immune responses by prostaglandin e 2. J Immunol (2012) 188:21–8. doi: 10.4049/jimmunol.1101029
121. De Groot AS, Moise L, McMurry JA, Wambre E, Van Overtvelt L, Moingeon P, et al. Activation of natural regulatory T cells by IgG fc-derived peptide “Tregitopes”. Blood (2008) 112:3303–11. doi: 10.1182/blood-2008-02-138073
122. Cousens LP, Najafian N, Mingozzi F, Elyaman W, Mazer B, Moise L, et al. In vitro and in vivo studies of IgG-derived treg epitopes (tregitopes): A promising new tool for tolerance induction and treatment of autoimmunity. J Clin Immunol (2013) 33:43–9. doi: 10.1007/s10875-012-9762-4
123. Andersson U, Björk L, Skansén-Saphir U, Andersson J. Pooled human IgG modulates cytokine production in lymphocytes and monocytes. Immunol Rev (1994) 139:21–42. doi: 10.1111/j.1600-065X.1994.tb00855.x
124. Wei S, Kryczek I, Zou W. Regulatory T-cell compartmentalization and trafficking. Blood (2006) 108:426–31. doi: 10.1182/blood-2006-01-0177
125. Dembele M, Tao S, Massoud AH, Miah SMS, Lelias S, De Groot AS, et al. Tregitopes improve asthma by promoting highly suppressive and antigen-specific tregs. Front Immunol (2021) 12:634509. doi: 10.3389/fimmu.2021.634509
126. Maddur MS, Othy S, Hegde P, Vani J, Lacroix-Desmazes S, Bayry J, et al. Immunomodulation by intravenous immunoglobulin: role of regulatory T cells. J Clin Immunol (2010) 30 Suppl 1:S4–8. doi: 10.1007/s10875-010-9394-5
127. McAlpine SM, Roberts SE, Heath JJ, Käsermann F, Issekutz AC, Issekutz TB, et al. High dose intravenous IgG therapy modulates multiple NK cell and T cell functions in patients with immune dysregulation. Front Immunol (2021) 12:660506. doi: 10.3389/fimmu.2021.660506
128. Pigard N, Elovaara I, Kuusisto H, Paalavuo R, Dastidar P, Zimmermann K, et al. Therapeutic activities of intravenous immunoglobulins in multiple sclerosis involve modulation of chemokine expression. J Neuroimmunol (2009) 209:114–20. doi: 10.1016/j.jneuroim.2009.01.014
129. Fillatreau S. B cells and their cytokine activities implications in human diseases. Clin Immunol (2018) 186:26–31. doi: 10.1016/j.clim.2017.07.020
130. Aubin É, Lemieux R, Bazin R. Indirect inhibition of in vivo and in vitro T-cell responses by intravenous immunoglobulins due to impaired antigen presentation. Blood (2010) 115:1727–34. doi: 10.1182/blood-2009-06-225417
131. Paquin Proulx D, Aubin É, Lemieux R, Bazin R. Inhibition of b cell-mediated antigen presentation by intravenous immunoglobulins (IVIg). Clin Immunol (2010) 135:422–9. doi: 10.1016/j.clim.2010.01.001
132. Séïté JF, Hillion S, Harbonnier T, Pers JO. Review: Intravenous immunoglobulin and b cells: When the product regulates the producer. Arthritis Rheumatol (2015) 67:595–603. doi: 10.1002/art.38910
133. Anthony RM, Kobayashi T, Wermeling F, Ravetch JV. Intravenous gammaglobulin suppresses inflammation through a novel Th2 pathway. Nature (2011) 475:110–3. doi: 10.1038/nature10134
134. Galeotti C, Stephen-Victor E, Karnam A, Das M, Gilardin L, Maddur MS, et al. Intravenous immunoglobulin induces IL-4 in human basophils by signaling through surface-bound IgE. J Allergy Clin Immunol (2019) 144:524–535.e8. doi: 10.1016/j.jaci.2018.10.064
135. Galeotti C, Karnam A, Dimitrov JD, Chevailler A, Kaveri SV, Bayry J. Anti-IgE IgG autoantibodies isolated from therapeutic normal IgG intravenous immunoglobulin induce basophil activation. Cell Mol Immunol (2020) 17:426–9. doi: 10.1038/s41423-019-0334-x
136. Miyake K, Shibata S, Yoshikawa S, Karasuyama H. Basophils and their effector molecules in allergic disorders. Allergy Eur J Allergy Clin Immunol (2021) 76:1693–706. doi: 10.1111/all.14662
137. Zuercher AW, Spirig R, Baz Morelli A, Rowe T, Käsermann F. Next-generation fc receptor–targeting biologics for autoimmune diseases. Autoimmun Rev (2019) 18:102366. doi: 10.1016/j.autrev.2019.102366
138. Zuercher AW, Spirig R, Baz Morelli A, Käsermann F. IVIG in autoimmune disease - potential next generation biologics. Autoimmun Rev (2016) 15:781–5. doi: 10.1016/J.AUTREV.2016.03.018
139. Ulrichts P, Guglietta A, Dreier T, Van Bragt T, Hanssens V, Hofman E, et al. Neonatal fc receptor antagonist efgartigimod safely and sustainably reduces IgGs in humans. J Clin Invest (2018) 128:4372–86. doi: 10.1172/JCI97911
140. Howard JF, Bril V, Burns TM, Mantegazza R, Bilinska M, Szczudlik A, et al. Randomized phase 2 study of FcRn antagonist efgartigimod in generalized myasthenia gravis. Neurology (2019) 92:e2661–73. doi: 10.1212/WNL.0000000000007600
141. Howard JF, Bril V, Vu T, Karam C, Peric S, Margania T, et al. Safety, efficacy, and tolerability of efgartigimod in patients with generalised myasthenia gravis (ADAPT): A multicentre, randomised, placebo-controlled, phase 3 trial. Lancet Neurol (2021) 20:526–36. doi: 10.1016/S1474-4422(21)00159-9
142. Newland AC, Sánchez-González B, Rejtő L, Egyed M, Romanyuk N, Godar M, et al. Phase 2 study of efgartigimod, a novel FcRn antagonist, in adult patients with primary immune thrombocytopenia. Am J Hematol (2020) 95:178–87. doi: 10.1002/ajh.25680
143. Robak T, Kazmierczak M, Jarque I, Musteata V, Trelinski J, Cooper N, et al. Phase 2 multiple-dose study of an FcRn inhibitor, rozanolixizumab, in patients with primary immune thrombocytopenia. Blood Adv (2020) 4:4136–46. doi: 10.1182/bloodadvances.2020002003
144. Bril V, Benatar M, Andersen H, Vissing J, Brock M, Greve B, et al. Efficacy and safety of rozanolixizumab in moderate to severe generalized myasthenia gravis: A phase 2 randomized control trial. Neurology (2021) 96:e853–65. doi: 10.1212/WNL.0000000000011108
145. Monnet C, Jacque E, de Romeuf C, Fontayne A, Abache T, Fournier N, et al. The dual targeting of FcRn and FcγRs via monomeric fc fragments results in strong inhibition of IgG-dependent autoimmune pathologies. Front Immunol (2021) 12:728322. doi: 10.3389/fimmu.2021.728322
146. Fitzpatrick EA, Wang J, Strome SE. Engineering of fc multimers as a protein therapy for autoimmune disease. Front Immunol (2020) 11:496. doi: 10.3389/fimmu.2020.00496
147. Yu X, Vasiljevic S, Mitchell DA, Crispin M, Scanlan CN. Dissecting the molecular mechanism of IVIg therapy: The interaction between serum IgG and DC-SIGN is independent of antibody glycoform or fc domain. J Mol Biol (2013) 425:1253–8. doi: 10.1016/j.jmb.2013.02.006
148. Basta M, Van Goor F, Luccioli S, Billings EM, Vortmeyer AO, Baranyi L, et al. F(ab)’2 -mediated neutralization of C3a and C5a anaphylatoxins: A novel effector function of immunoglobulins. Nat Med (2003) 9:431–8. doi: 10.1038/nm836
149. Amagai M, Ikeda S, Shimizu H, Iizuka H, Hanada K, Aiba S, et al. A randomized double-blind trial of intravenous immunoglobulin for pemphigus. J Am Acad Dermatol (2009) 60:595–603. doi: 10.1016/j.jaad.2008.09.052
150. Iwata H, Vorobyev A, Koga H, Recke A, Zillikens D, Prost-Squarcioni C, et al. Meta-analysis of the clinical and immunopathological characteristics and treatment outcomes in epidermolysis bullosa acquisita patients. Orphanet J Rare Dis (2018) 13: 1–9. doi: 10.1186/s13023-018-0896-1
Keywords: intravenous immunoglobulin, autoimmune diseases, inflammatory diseases, mathematical models, B cells, complement system, immunomodulation, IVIg Immunoglobulins
Citation: Segú-Vergés C, Caño S, Calderón-Gómez E, Bartra H, Sardon T, Kaveri S and Terencio J (2022) Systems biology and artificial intelligence analysis highlights the pleiotropic effect of IVIg therapy in autoimmune diseases with a predominant role on B cells and complement system. Front. Immunol. 13:901872. doi: 10.3389/fimmu.2022.901872
Received: 22 March 2022; Accepted: 31 August 2022;
Published: 30 September 2022.
Edited by:
A. Razzaque Ahmed, Tufts University, United StatesReviewed by:
Ralf J. Ludwig, University of Lübeck, GermanyCopyright © 2022 Segú-Vergés, Caño, Calderón-Gómez, Bartra, Sardon, Kaveri and Terencio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Srini Kaveri, U3JpbmkuS2F2ZXJpQGNyYy5qdXNzaWV1LmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.