 Zeyu Zhang1,2†
Zeyu Zhang1,2† Yang Yu3†
Yang Yu3† Guanxiong Zhu1,2†Liting Zeng1,2†
Guanxiong Zhu1,2†Liting Zeng1,2† Shaofen Xu1,2
Shaofen Xu1,2 Haoyu Cheng1,2Zhaoguang Ouyang1,2Jianwei Chen1,2
Haoyu Cheng1,2Zhaoguang Ouyang1,2Jianwei Chen1,2 Janak L. Pathak1*
Janak L. Pathak1* Lihong Wu1,2*Lina Yu1,2*
Lihong Wu1,2*Lina Yu1,2*- 1Department of Preventive Dentistry, Affiliated Stomatology Hospital of Guangzhou Medical University, Guangdong Engineering Research Center of Oral Restoration and Reconstruction, Guangzhou Key Laboratory of Basic and Applied Research of Oral Regenerative Medicine, Guangzhou, China
- 2School and Hospital of Stomatology, Guangzhou Medical University, Guangzhou, China
- 3Department of Sports and Health, Guangzhou Sport University, Guangzhou, China
Periodontitis is an infectious oral disease, which leads to the destruction of periodontal tissues and tooth loss. Although the treatment of periodontitis has improved recently, the effective treatment of periodontitis and the periodontitis-affected periodontal tissues is still a challenge. Therefore, it is urgent to explore new therapeutic strategies for periodontitis. Natural products show anti-microbial, anti-inflammatory, anti-oxidant and bone protective effects to periodontitis and most of these natural products are safe and cost-effective. Among these, the plant-derived exosome-like nanoparticles (PELNs), a type of natural nanocarriers repleted with lipids, proteins, RNAs, and other active molecules, show the ability to enter mammalian cells and regulate cellular activities. Reports from the literature indicate the great potential of PELNs in the regulation of immune functions, inflammation, microbiome, and tissue regeneration. Moreover, PELNs can also be used as drug carriers to enhance drug stability and cellular uptake in vivo. Since regulation of immune function, inflammation, microbiome, and tissue regeneration are the key phenomena usually targeted during periodontitis treatment, the PELNs hold the promising potential for periodontitis treatment. This review summarizes the recent advances in PELNs-related research that are related to the treatment of periodontitis and regeneration of periodontitis-destructed tissues and the underlying mechanisms. We also discuss the existing challenges and prospects of the application of PELNs-based therapeutic approaches for periodontitis treatment.
Introduction
Periodontitis is a chronic inflammatory disease of periodontal tissue with a prevalence rate 50% (1). Periodontitis leads to chronic pain, gingival swelling, destruction of periodontal ligaments, and loss of alveolar bone and teeth (1). In 2017, severe periodontitis was the sixth most prevalent disease affecting 9.8% (about 796 million) global adult population (2, 3). Periodontitis not only affects oral health but also is linked to various systemic diseases including cardiovascular disease, Alzheimer’s disease, type 2 diabetes mellitus, respiratory tract infection, rheumatoid arthritis, nonalcoholic fatty liver diseases, and certain cancers (4, 5). Therefore, the effective treatment of periodontitis is vital for a healthy life. Conventional treatment approaches to periodontitis including non-surgical, surgical, and adjunctive pharmacological therapy have limitations, such as residual bacterial and calculus in the deep periodontal pocket, limited effect on inflammation regulation, limited periodontal tissue regeneration, and lack of consideration of the effect of systemic diseases (6–8).
In recent years, natural products have attracted more and more attention in the treatment of human diseases (9). It has been widely demonstrated that natural products possess anti-microbial, anti-oxidant, and anti-inflammatory properties and are widely used in the treatment of various diseases including cancer, malaria, and periodontitis (10, 11). In addition, observational studies had shown correlation between the intake of fruits and vegetables with oral health (12). Moreover, Kharaeva et al. indicated that toothpaste containing plant-derived ingredients has an additional therapeutic effect in the treatment and prevention of gingivitis and periodontitis (13). However, shortcomings of natural products such as uncertain stability, limited target specificity, and difficulty in purification limit their clinical applications. Halperin et al. first discovered evidence of the existence of PELNs in carrot cell cultures in 1967 (14). PELNs (50-500 nm in diameter) contain mRNAs, microRNAs (miRNAs), bioactive lipids, and proteins (15, 16). Compared to artificial nanocarriers, PELNs do not exert cytotoxicity on human cells (15). PELNs lipid bilayer contains high contents of glycolipids and phospholipids but lacks cholesterol, which indicates the potential application of PELNs as a tissue targeting drug carrier (16). Recent reports from literature had shown the potential of PELNs to treat various diseases, including inflammatory bowel diseases, lung inflammation, and periodontitis (17–19). The nanoparticle size, lipid membrane, specific targeting, and cargo-carrying capacity of PELNs offer better stability and fewer side effects. Ginger-derived exosome-like nanoparticles (GELNs) had shown an anti-bacterial effect on Porphyromonas gingivalis (P. gingivalis) (19). Although in vitro and animal studies had shown the therapeutic potential of PELNs to treat periodontitis, their clinical application has not been reported yet. This review summarizes the recent research advances in the PELNs related to immune-regulation and periodontitis treatment. We also discuss the shortcomings and prospects of PELNs-based immune-regulation and periodontitis treatment.
PELNs
Extracellular vesicles are a subcellular structure of phospholipid bilayers membrane-enclosed vesicles and contain various cargos, including miRNA, mRNA, DNA, proteins, etc. Both prokaryotic and eukaryotic cells release extracellular vesicles. Extracellular vesicles fall into two broad categories: ectosomes (size: 100 to 500 nm) and exosomes (size: 30 to 150 nm) (20–23). PELNs are a kind of extracellular vesicles ranging in size from 50 to 500 nm derived from plants (15). Besides animal vesicles, PELNs have a complex content of small RNAs, proteins, lipids, and other metabolites. PELNs from various plants and fruits such as ginger, blueberry, and coconut have shown anti-inflammatory properties (24).
Compared to mammalian exosomes, PELNs have unique advantages including undetected by the immune system, higher bioavailability, and innocuity (25). PELNs were observed almost six decades ago but less attention was paid to this field (26). PELNs exhibit better bioavailability compared to miRNAs that are free or associated with proteins (27). PELNs have proven stability in the gastrointestinal tract, and several studies have demonstrated that PELNs can be used for therapeutic application by oral or intranasal administration (28–30). Compared to natural products, PELNs can target specific organs and have higher solubility, higher permeation through barriers, quicker dissolution in blood, and fewer side effects (29). Reports from the literature indicate that plant-derived exosomes have potential application in the treatment of periodontitis through inflammation inhibition or periodontal pathogen inhibition (19, 28). Due to these properties, PELNs show the application prospect for the treatment and prevention of various inflammatory diseases including periodontitis.
Composition of PELNs
PELNs contain various components including proteins, lipids, miRNA, and other active components such as vitamin C (31). Exosomes are derived from animal cells and are typically rich in cholesterol and sphingomyelin but PELNs are rich in phospholipids, including phosphatidic acids, phosphatidylethanolamines, and typical plant lipids (30, 32). Lipids play an important role not only in maintaining the structural stability of exosomes but also in intercellular communication (15). Among these lipids, phosphatidic acids in PELNs can inhibit P. gingivalis growth (19). Proteins are important components of both PELNs and mammalian exosomes, but the levels of proteins in PELNs are lower and the compositions are different (32). In mammalian exosomes, CD9 promotes cell delivery of therapeutic agents through fusing exosomal membranes to target cell membranes and CD47 could escape phagocytosis by releasing special signals and enhancing homogenous endocytosis (33). There are various proteins in PELNs, such as actin, proteolysis, aquaporin, and chloride channels proteins, which are mainly categorized into transmembrane proteins and other plasmalemma-associated proteins (25, 34). Defense proteins in some PELNs such as sunflower seeds can modulate microbiota by affecting fungal growth (35). MicroRNAs are a class of small (17-24 nucleotides) and noncoding RNAs with abilities to inhibit mRNA translation (36, 37). The latest evidence shows that plant miRNA can be absorbed in the intestine and secreted into the circulatory system (38). The previous report had shown that PELNs deliver miRNA to animals, target mammalian mRNA, and have the potential to mediate a specific tissue response (24, 39). In addition, miRNA in PELNs can be taken up by bacteria and alter microbiome composition and host physiology (28, 38, 40). But the functions and mechanisms of miRNAs in PELNs are still unknown. It is unclear how plant-based xenomiRNAs regulate gene expression in humans (41). Besides, some bioactive components such as vitamin C, citrate, 6-gingerol, and 6-shogaol have been found in PELNs (34, 42). Only a handful of studies about the biological function of bioactive components in PELNs had been reported so far. The contents of PELNs are shown in Figure 1.
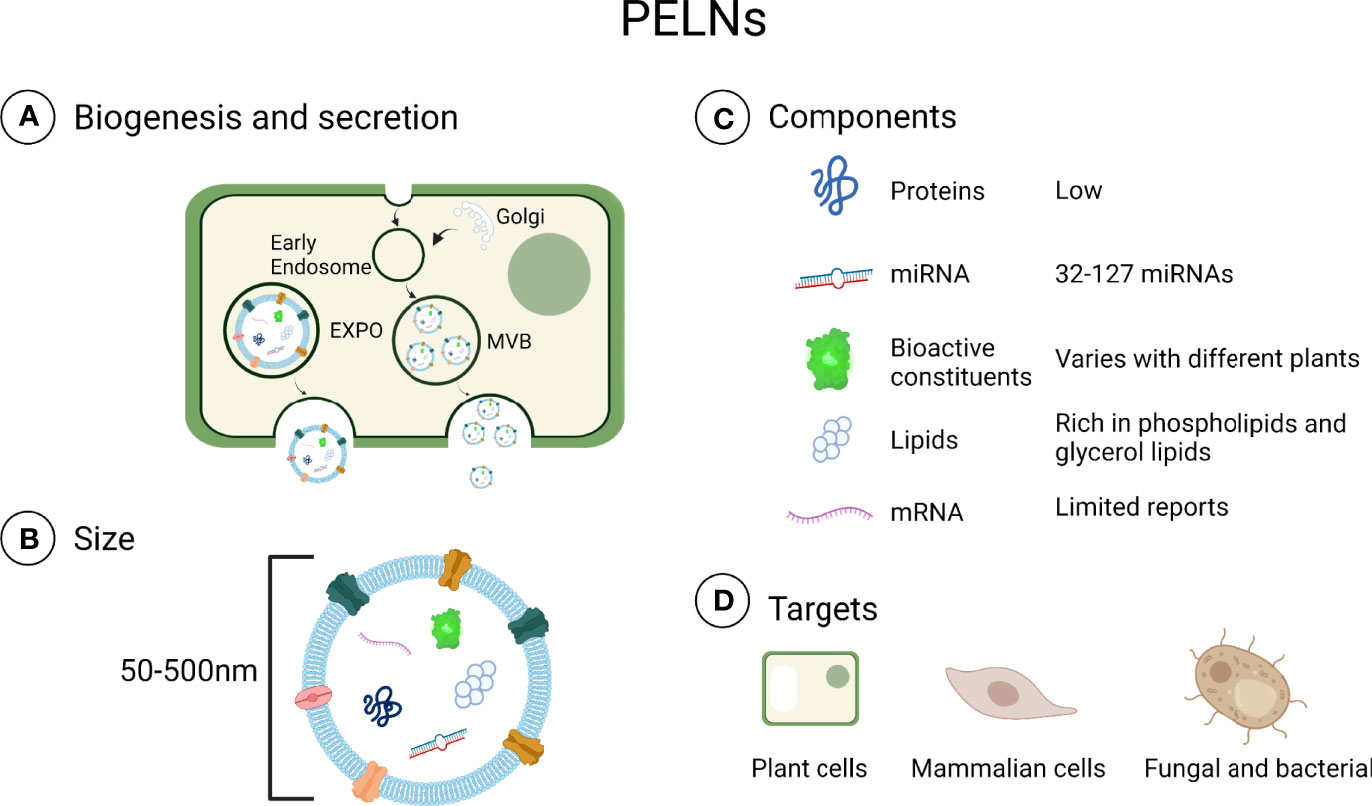
Figure 1 Contents in PELNs. (A) Biogenesis and secretion: Plant cells can secrete PELNs via multivesicular bodies (MVB) and exocyst positive organelles (EXPO) (43, 44). (B) Size: PELNs range in size from 50 to 500 nm. (C) Components: Generally, PELNs contain fewer proteins and miRNAs than exosomes. (D) Targets: PELNs can be internalized in plant cells, mammalian cells, fungi, and bacteria (28, 45–47). Created with BioRender.com.
PELNs Biogenesis and Isolation
The extracellular vesicle formation and secretion require a multi-step cellular process that is well documented in animals. Extracellular vesicles are formed in intracavitary vesicles (ILVs) and multivesicular bodies (MVB). ILVs released into the extracellular space after fusion with the plasma membrane are exosomes (48, 49). However, the process of PELNs release from cell walls is still unknown. PELNs participate in plant-microbe interactions by safely transporting functional molecules including proteins and RNAs (50). The MVB pathway is a key process of PELNs formation (44). The endosomal sorting complex required for transport (ESCRT) binds and sequesters ubiquitinated proteins and sorts these into the ILVs of PELNs (51). However, the ESCRT genes responsible for PELNs have not yet been elucidated. Even though PELNs are secreted by most plant cells, the process of extracellular vesicles passing through the apoplastic space or cell wall is still unclear (52). In addition, exocyst-positive organelle (EXPO), in plant cells, also can expulse PELNs into the apoplast through the fusion of the outer membrane of EXPO with the plasma membrane, but the biological significance of EXPO-mediated PELNs secretion in plants is still undetermined (43).
The isolation of PELNs is mainly based on differential centrifugation. PELNs can be extracted from fruits, roots, stems, and leaves (24, 30, 53). The conventional method is to grind the plants into juice and then strain the juice with a colander. The collected juice goes through differential centrifugation at 3000× g (for 20-30 min) and 10,000× g (for 1 h) (24, 31). This supernatant is then subjected to centrifugation at high speed (100,000–150,000 × g) (29). Because the PELNs yield is usually contaminated by nucleic acids and protein agglomerates after differential ultracentrifugation, sucrose/deuterium oxide gradient ultracentrifugation at ~110,000 × g for 3 h at 4°C is needed for further purification (Figure 2) (25, 54). However, differential centrifugation also has a lot of disadvantages, e.g., low PELNs yields because a large number of nanovesicles are lost during centrifugation, retention of protein aggregates, and disruption of nanovesicles due to high centrifugal forces (55).
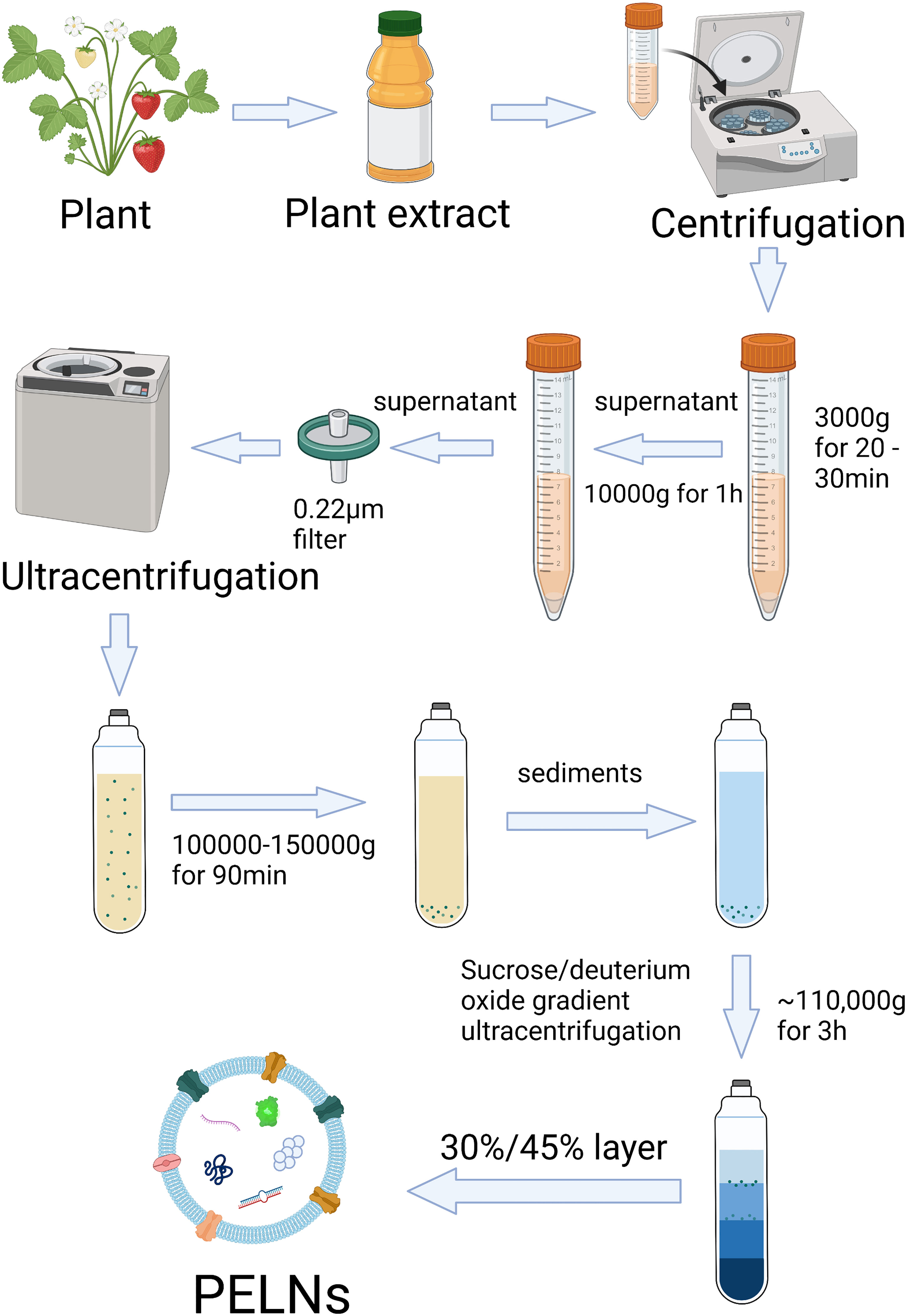
Figure 2 Scheme of isolation and purification of PELNs by differential ultracentrifugation and sucrose gradient ultracentrifugation. Created with BioRender.com.
Polyethylene glycol (PEG)-based precipitation method is another method for PELNs isolation (56). The PEG method is a cost-effective method of PELNs isolation with comparable efficiency to differential ultracentrifugation (57). PEG methods are related to pH. Suresh et al. reported a higher yield of PELNs when PEG precipitation was carried out in pH 4 and 5 (58). Differential ultracentrifugation, PEG, density-gradient ultracentrifugation, gel filtration chromatography, ultrafiltration, immunoaffinity separation, etc. are the methods of PELNs isolation (59). Differential ultracentrifugation is still the “gold standard” due to its wide applicability, large capacity, easy scale-up, and relatively high purity (59). Figure 2 demonstrates the procedure of the commonly used differential ultracentrifugation method of PELNs isolation.
Internalization of PELNs by Mammalian Cells and Mechanisms of Cargo Release
Several studies had reported that PELNs can be internalized by mammalian cells and do not exert cytotoxicity (30, 31, 60). Ju et al. indicated that intestinal stem cells take up grape-derived exosome-like nanoparticles (GrELNs) through micropinocytosis, which can be inhibited by cytochalasin D (30). But the molecular mechanism of micropinocytosis of PELNs in mammalian cells is still unclear. Based on the similarity of components and structure between PELNs and exosomes, it is reasonable to guess that PELNs can be internalized by way of fusion and endocytosis just like mammalian cell exosomes (61, 62). Exosomal membrane proteins play an important role in the uptake of exosomes (61). So further study and analysis of membrane proteins of PELNs are the keys to understanding the specific mechanism of internalization of PELNs in mammalian cells. Besides, mammalian exosomes can induce cellular responses through membrane-bound or soluble signaling, which does not require internalization (63). Whether PELNs can induce cellular responses in a similar way to mammalian exosomes still needs to be further investigated.
The PELNs taken up by cells through fusion can release cargoes into the cytoplasm directly. But the fate of PELNs taken up through endocytosis may be different. Typically, PELNs taken up by cells may transport to the lysosome and lead to degradation-based cargo release (62). Under certain circumstances, cargoes in PELNs can be released into the cytoplasm in different ways including fusion with the lysosome, the disintegration of the early sorting endosome, and fusion with the endoplasmic reticulum and endosomal membrane (64). The possible mechanisms of PELNs internalization in mammalian cells and cargo release are depicted in Figure 3.
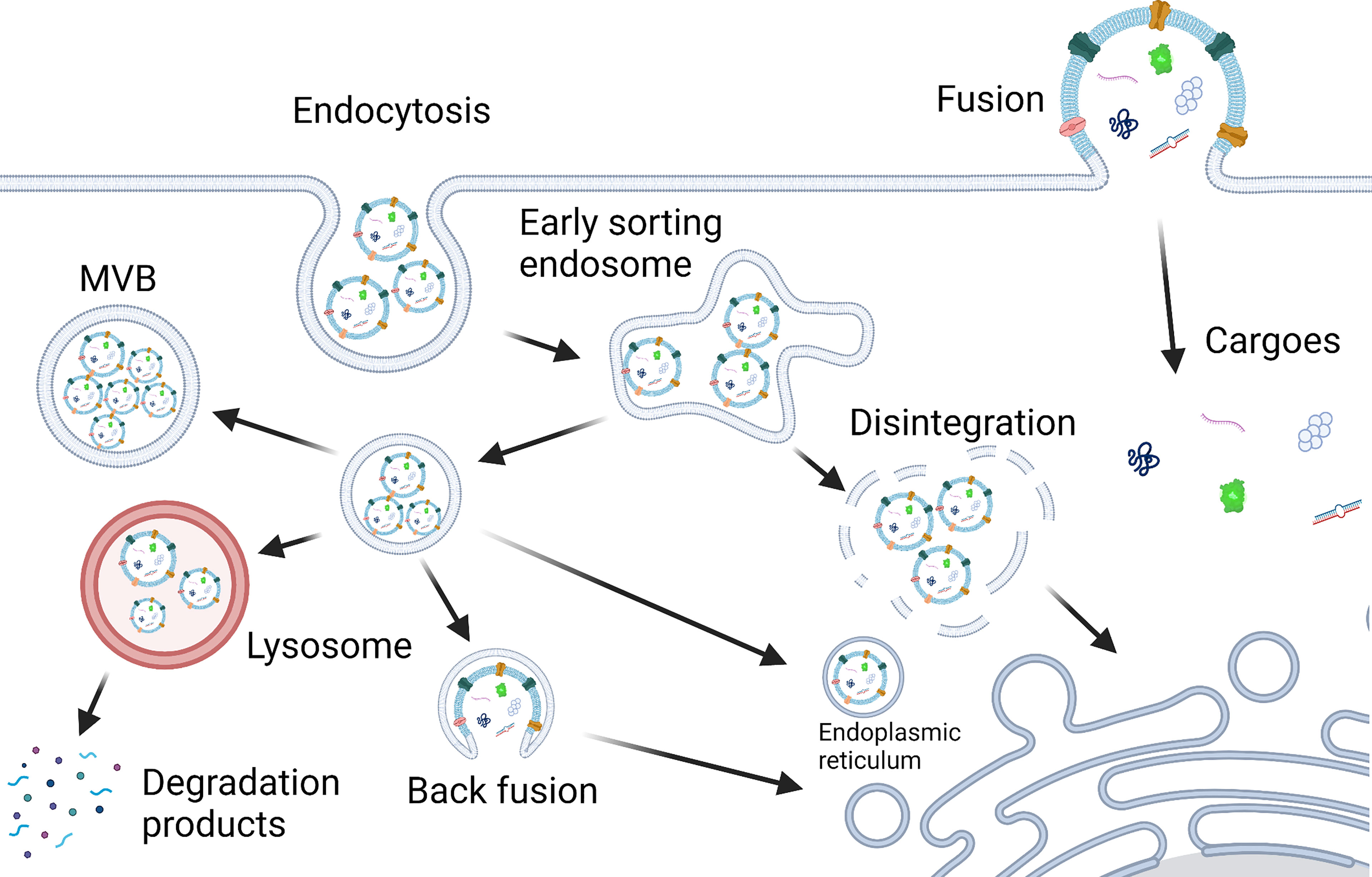
Figure 3 Scheme of PELNs internalization in mammalian cells and cargo release. Mammalian cells take up PELNs possibly through endocytosis and fusion. Cargoes in PELNs can be released into the cytoplasm in different ways, i.e., fusion with the lysosome, the disintegration of the early sorting endosome, and fusion with the endoplasmic reticulum, endosomal membrane and plasma membrane (22, 64). Created with BioRender.com.
PELNs-Mediated Regulation of Human Cells’ Functions
Recently, cross-kingdom regulation of human transcripts by plant miRNAs has been demonstrated (24). PELNs have shown anti-inflammatory properties in human cells. According to a recent report, GELNs show anti-inflammation effects by inhibiting the expression of Nsp12 in lung epithelial cell-exosome-mediated inhibition of macrophagic inflammation (18). Teng et al. reported that GELN RNAs ameliorate mouse colitis by inducing gut probiotic Lactobacillus rhamnosus GG (LGG) indole-3-carboxaldehyde, which promotes the expression of interleukin (IL)-22 in gut lymphocytes of mice through activation of the aryl hydrocarbon receptor (AHR) signaling pathway (28). Exosome-like nanoparticles (ELNs) from blueberry counter the response to tumor necrosis factor (TNF-ɑ)-induced change in gene expression in EA. hy926 cells, pretreatment with blueberry-derived ELNs counters TNF-ɑ-induced reactive oxygen species generation and loss of cell viability and modulates the differential expression of 29 genes induced by TNF-ɑ compared to control (60).
In addition to anti-inflammatory properties, PELNs have shown anti-tumorigenic properties. Berry anthocyanidins-derived ELNs inhibit ovarian cancer cell proliferation in vitro and tumor growth in vivo (65). Citrus-limon juice-derived ELNs inhibit tumor cell growth through a significant downregulation of the Acetyl-CoA Carboxylase 1 (ACACA) (66). Engineered ELNs from Asparagus cochinchinensis show antitumor activity via inducing apoptotic pathways (67).
Treatment of Periodontitis
Biological activities targeted during periodontitis prevention and treatment and periodontal tissue regeneration include inflammation regulation, anti-bacterial, immune-regulation, osteogenesis, periodontal ligament regeneration, and angiogenesis. Scaling and root planing (SRP), the gold standard method for dental plaque removal, has a significant anti-bacterial effect and inhibits the development of periodontal diseases (6, 68). But the complex root anatomy and recolonization of microbiota limits the efficacy of SRP (6–8). Regenerative surgeries have been used in promoting osteogenesis and periodontal ligament regeneration, but the indication and efficacy are limited (69, 70). Although various materials such as autogenous bone, allogeneic bone, and alloplastic substitutes, are used in regenerative surgery, limited materials are currently available with the true periodontal regeneration potential (71, 72).
To overcome the disadvantages of existing therapeutic approaches, adjunctive pharmacological therapies have been used in periodontitis treatment. Antibiotics brought additional benefits as an adjunct in periodontitis treatment, but the risks such as bacterial resistance, hypersensitivity, and superinfection limit the clinical application (73–75). Natural products attract more and more attention in periodontitis treatment due to their therapeutic potential, cost-effectiveness, enough source, and safety. A series of natural products such as honey, propolis, cannabidiol, and green tea show various benefits in periodontitis treatment and prevention including inhibition of periodontal pathogens, anti-inflammation, immunomodulation, and osteogenesis (76–85). But the low stability, uncertain bioavailability, and limited therapeutic effect of natural products limit their clinical application.
The Possible Role of PELNs in the Prevention and Treatment of Periodontitis
PELNs have shown anti-inflammatory, microbiome modulatory, immunomodulatory, and tissue regenerative properties that could be beneficial for the prevention and treatment of periodontitis. To achieve superior therapeutic effects against periodontitis, PELNs can be used in drug delivery systems to increase the bioavailability and biodistribution of the drugs (25). Compared with natural products, PELNs have a broader application range and higher stability. Unlike mammalian exosomes, PELNs can be easily isolated and purified in large quantities, and have better biocompatibility (18). Since PELNs come from plants, there are fewer ethical issues during clinical applications. Therefore, with these properties and advantages, PELNs have shown great value in the prevention and treatment of periodontitis (Table 1).

Table 1 Overview of PELNs biological activities that could be applied in periodontitis treatment.
Immune Regulation and Anti-Inflammation
Macrophages are important parts of the immune system. In periodontitis, macrophages mediate the development and progression of periodontitis through M1 and M2 polarization (97). M1 macrophages produce a series of pro-inflammatory factors including TNF-α and IL-6 to kill bacteria, promote inflammation, and activate osteoclasts that cause absorption of the alveolar ridge. In contrast, M2 macrophages produce anti-inflammatory factors including IL-10 and transforming growth factor (TGF)-β to exert anti-inflammation and angiogenic effects, and activate osteoblasts to restore bone tissue (97–99). A recent study indicated that PELNs can be absorbed by intestinal macrophages and regulate immune response (86). GELNs can be absorbed by macrophages and upregulate the expression of heme oxygenase-1 (HO-1), IL-6, and IL-10. Carrot-derived ELNs induce IL-10 expression in macrophages. Grapefruit, carrot, and ginger-derived ELNs promote activation of nuclear factor (erythroid-derived 2)-like-2 (Nrf2) in macrophages (86). GELNs block the assembly of the NLRP3 inflammasome in macrophages (53). In addition, ginseng-derived ELNs suppress IL-4 and IL-13-induced M2-like polarization of macrophages and increased the secretion of M1-macrophage-associated cytokines including TNF-α, IL-12, and IL-6 (89).
Shreds of literature have reported that ginger, grapes, grapefruit, carrots, and blueberry-derived ELNs have anti-inflammatory effects (18, 60, 86, 100). Mu et al. demonstrated that GELNs induce the expression of heme oxygenase-1 and IL-10 in macrophages, and grapefruit, ginger, and carrot-derived ELNs promote activation of Nrf2 in macrophages (86). GrELNs cause significant induction of intestinal stem cells through the Wnt/β-catenin pathway, which protects mice from dextran sulfate sodium-induced colitis (30). Cabbage and red cabbage-derived ELNs decrease the levels of IL-6, IL-1β, and COX-2 expression in LPS-treated macrophages, showing clear anti-inflammatory effects (55). Orange (Citrus sinensis)-derived ELNs limit inflammation and restore the functional intestinal barrier by altering the expression of HMOX-1, ICAM1, OCLN, CLDN1, and MLCK (91). The anti-inflammatory properties of PELNs could be possibly attributed to the micro-RNA in PELNs. Aquilano et al. reported that mimics of miR159a and miR156c target Tnfrsf1a gene transcript in adipocytes. Nuts (Juglans california, Corylus avellana, and Juglans regia)-derived ELNs containing miR159a and miR156c show anti-inflammatory effects through reducing Tnfrsf1a protein and dampening TNF-α signaling in adipocytes (88). Aly-miR396a-5p present in GELNs inhibits inflammation and cell apoptosis by inhibiting the expression of the Nsp12 viral gene (18). Besides, the natural substance phenolic compounds may also be responsible for the anti-inflammatory effects of PELNs. GrELNs loaded with phenolic compounds inhibit colitis inflammation by decreasing TNF-α and NF-κB (100). Lipids of PELNs also show anti-inflammatory properties. Lipids in Broccoli-derived ELNs inhibit gut inflammation by driving the induction of CD11c tolerogenic dendritic cells (101). A recent report indicated that proteins in PELNs also show anti-inflammatory properties. Mulberry bark-derived ELNs prevent mouse colitis through Mulberry bark ELNs-derived heat stock protein HSP8 induced AhR/COPS8 pathway (17). The phospholipid-based vesicular system enhances the delivery and safety of aceclofenac by topical route (102). As nanoparticles with bilayer phospholipid structure, PELNs can be used as a carrier for anti-inflammatory drug loading. Orally administrated infliximab-loaded GELNs show gastrointestinal stability, colon-targeted delivery, high intestinal epithelium permeability, and better efficacy in colitis than the intravenously administered infliximab (103). PELNs are innocuous and non-immunogenic nanoparticles with higher uptake in human cells compared with other artificial nanoparticles used in drug delivery (15). In addition, natural active components in PELNs could exert a better therapeutic effect than artificially synthesized nanoparticles such as liposomes. Although this field of research has not been explored in-depth, the reports from literature suggest the important role of PELNs in the treatment of inflammatory diseases. Inflammation plays an important role in the tissue destruction of periodontitis. Therefore, PELNs could have the potential to decrease periodontal tissue destruction and slow the rate periodontitis progression through inhibition of inflammation.
Modulation of Microbiota Composition
As a secondary inflammatory response caused by oral microbiome dysbiosis, periodontitis is initiated by the host immune response to changes in the oral microbiome (104). Among the more than 500 bacterial species living in the oral cavity, a bacterial complex called “red complex”, which is made of P. gingivalis, Treponema denticola, and Tannerella forsythia, express various virulence factors, which enable these bacteria to colonize in subgingival space, destroy the host’s defense system, invade periodontal tissue, and promote the host’s immune destruction response (105). Given the role of microbiota in periodontitis, it is crucial to maintain the dynamic equilibrium of oral microecology (106). It is widely accepted that PELNs have a key role in plant-pathogen interactions (107). PELNs have shown regulatory effects not only in immune cells but also in microbiota. PELNs play an important role in immune responses against the fungal pathogen in plants. For instance, Arabidopsis-derived ELNs delivered host sRNA into pathogenic B. cinerea to inhibit its pathogenicity (108). PELNs also inhibit various pathogens such as P. gingivalis and promote the growth of probiotics. Sundaram et al. reported that GELNs inhibit P. gingivalis growth through phosphatidic acid by binding to hemin-binding protein 35 (HBP35) on the surface of P. gingivalis (19). Besides, PELNs show microbiome regulatory properties through the promotion of the growth of probiotics. Lemon-derived ELNs protect LGG from bile possibly through inhibition of the expression of Msps protein, induction of specific bacterial tRNA decay, and inhibition of S.24-7 growth (93). Lemon-derived ELNs manipulate LGG and Streptococcus thermophilus ST-21 to protect mice from Clostridioides difficile infection (94). In addition, plant miRNAs in PELNs modulate microbiota gene expression contributing to the dietary effect on the gut microbiota community’s assembly (109). PELNs-derived small RNAs shape the homeostatic balance between host immunity and gut microbiota and regulate microbiota (28). Teng et al. indicated that GELNs-RNA induces Lactobacillus rhamnosus indole-3-carboxaldehyde to promote the expression of IL-22 through activation of the AHR signaling pathway exerting anti-microbiota immunity and tissue repair (28). With the ability to modulate oral and gut microbiota, it is reasonable to suppose that PELNs have the potential to be used in the treatment of periodontitis through the regulation of the periodontal microbiome.
Periodontal Tissue Regeneration
The progressive loss of periodontal tissues is one of the characterizations of periodontitis, and the unique anatomy and composition of periodontal tissues make periodontal tissue regeneration a complex process (110). The reconstruction of periodontal tissues including cementum, periodontal ligament fibers, and bone remains a major challenge in periodontal treatment (111). Osteogenesis, inflammatory response, angiogenesis, and remodeling play a significant role in periodontal bone regeneration (112). Reports from the literature indicate that mammalian exosomes stimulate both osteogenesis and angiogenesis (113). With component and structure similarity to mammalian exosomes, PELNs could have application potential in tissue regeneration. According to recent research, PELNs have regulatory effects on tissue regeneration. Sahin et al. reported that wheat-derived ELNs promote collagen type I production, proliferation, and migration of fibroblasts (95). Wheat-derived ELNs exert anti-apoptotic activity in human dermal fibroblast, human keratinocyte cell, and human keratinocyte cell. In addition, wheat-derived ELNs induce angiogenesis in human umbilical vein endothelial cells (95). These results suggest the possible periodontal soft tissue regeneration and angiogenesis potential of PELNs. Besides, Syrah GrELNs induce the expression of leucine-rich repeat-containing G-protein-coupled receptor 5 of intestinal stem cells through activating downstream canonical Wnt signals, which is beneficial for the regeneration of intestinal epithelium (30, 114). Lipid contents in GrELNs promote the proliferation of intestinal stem cells (30). Whether PELNs can promote periodontal tissue regeneration by inducing tissue-specific differentiation of precursor cells remains a mystery. Green tea and ginseng-derived ELNs show potential benefits to skin regeneration, through modulating the expression of genes including MMP12, MMP13, HS3ST3A1, FGF12, LOX, VIM, ELOVLs, KRT1, and NOTCH3 in keratinocytes, which are related to skin aging, regeneration, barriers, and moisturizing (96). According to the existing data, we can reasonably guess that PELNs have a potential therapeutic effect on promoting the proliferation of periodontal stem cells and promoting the repair and regeneration of periodontal tissue. The possible applications of PELNs in the treatment of periodontitis are shown in Figure 4.
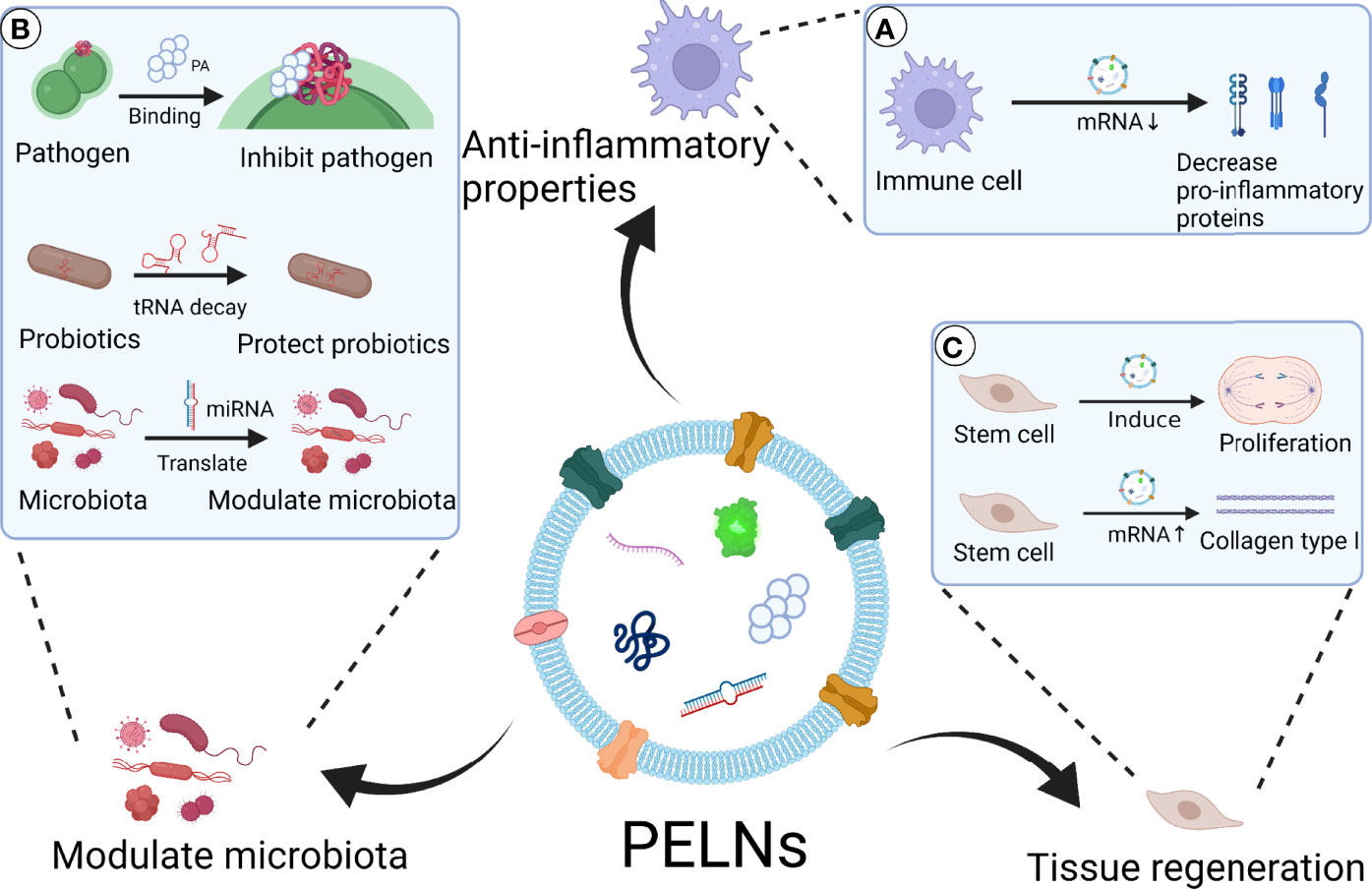
Figure 4 Scheme of PELNs’ possible application in the treatment of periodontitis. PELNs show therapeutic potential for periodontitis via anti-inflammatory effect, microbiota modulation, and tissue regeneration. (A) Anti-inflammatory properties: PELNs inhibit pro-inflammatory protein expression in macrophages, such as IL-1β, IL-6, and TNF-α (55). (B) Modulate microbiota: PELNs inhibit pathogenic bacteria such as P. gingivalis through PA binding to HBP35 (19). PELNs protect probiotics through decay tRNA in probiotics (93). PELNs transform miRNA to microbiota, which has the potential to modulate oral microbiota. (C) Tissue regeneration: PELNs promote the proliferation of stem cells through activation of Wnt signals (30, 114). PENLs promote the production of collagen type I in epithelial cells (95).Created with BioRender.com.
Summary
As a type of plant-derived extracellular nanovesicles, PELNs can be taken into the human circulation from the gut and participate in cross-kingdom communication. The biological properties of PELNs are based on the transmission of miRNAs, proteins, lipids, and other active components. Recent studies indicated that plants communicate with mammalian cells and bacteria through PELNs, and miRNAs in PELNs show the potential to regulate human mRNAs’ activities (24, 28, 86). Increasing evidence indicated that PELNs had great potential in immune regulation and treatment/prevention of periodontitis due to their drug delivery ability and various therapeutic effects including anti-inflammatory, immunomodulatory, microbiota modulatory, and regenerative effects. Besides, compared with conventional chemical drugs and natural products, the special properties including therapeutic and drug delivery ability, stability, and safety enable the PELNs-based therapeutic approaches to break the limitations of the existing periodontitis treatment. Furthermore, compared with mainstream oral hygiene maintenance methods such as chlorhexidine, PELNs are more suitable for daily prevention of periodontitis, such as mouthwash, toothpaste, and chewing gum.
Challenges
Although PELNs show various advantages in the prevention and treatment of periodontitis, there are still some barriers that need to be overcome to explore possibilities for their clinical applications. PELNs as nanocarriers cannot load a high quantity of drugs. To augment cargo loading capability, PELNs amalgamation with artificially synthesized liposomes by the membrane fusion technique may be a feasible approach (25). In addition, the mechanism of PELNs uptake by mammalian cells is also unclear. To further explore the potential of clinical application, it is necessary to determine the mechanisms that facilitate PELNs’ internalization in mammalian cells. The standards of biochemical analysis and biomarker confirmation of PELNs need to be developed for the quality control and engineering process. PELNs derived from the same species of plant may vary from batch to batch. Therefore, it is necessary to develop strategies to minimize the batch-dependent variation of PELNs’ contents.
Prospects
As a cell-free therapy, PELNs are safer than cell therapy and PELNs can target specific cells or organs (115). Compared with other cell-free therapies, such as mammal exosomes, PELNs are easier to extract and have unique advantages including biocompatibility, large-scale production capability, and low immunogenicity (116). PELNs also have the potential to use in oral care products such as buccal tablets for daily prevention of periodontitis. As lipid-based nanoparticles, PELNs can carry and deliver hydrophilic, hydrophobic, and lipophilic drugs (117). It has been reported that Acerola-derived ELNs can encapsulate nucleic acids without the use of special reagents (118, 119). Various methods such as loading by electroporation, saponin membrane permeabilization, and extrusion had been developed drug loading in exosomes, which can be adapted to load desired drugs in PELNs. Besides, PELNs have good stability and can protect the contents from physicochemical damage in vivo. The encapsulated siRNA in PELNs is shown to be stable against physical stimuli including sterilization, homogenization, and sonication (119). These results allow us to hypothesize that PELNs can load anti-inflammatory, anti-microbial drugs or special siRNA for periodontal treatment in the future. In addition, PELNs can be imparted with target specificity through investigating the surface tailoring of PELNs (25). It is possible to modulate the tropisms of PELNs by introducing tissue-specific peptides or proteins onto the surfaces of PELNs (120). Membrane fusion with other extracellular vesicles or cell membranes with special receptors may also be a feasible way to enhance the targeted specificity of PELNs. The combination of PELNs and new clinical techniques of minimally invasive targeted delivery such as convection-enhanced delivery may also extend the application of PELNs by improving the reliability and limiting the confusion of various target bioavailability (121). Wang et al. proposed a new method that GrELNs coated with inflammation-related receptor enriched membranes of activated leukocytes, which showed better target ability to inflammatory tumor tissues (122). Therefore, PELNs have great potential to be developed as a drug delivery system for periodontitis treatment. Potential modifications of PELNs that can improve therapeutic efficacy against periodontitis are listed in Figure 5.
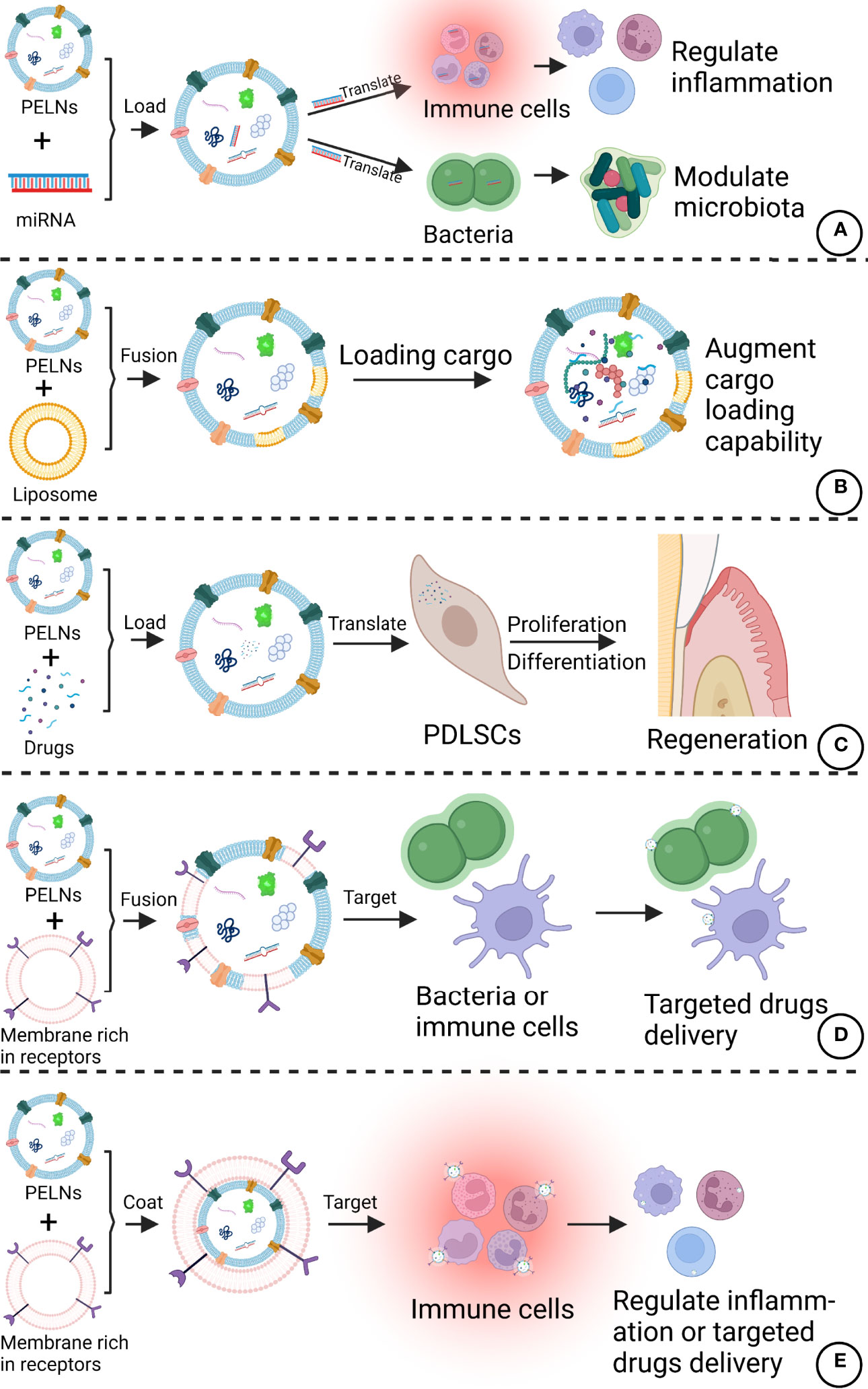
Figure 5 Potential modifications of PELNs that can improve therapeutic efficacy against periodontitis. (A) PELNs can be loaded with specific miRNAs targeting immune cells and bacteria. (B) The amalgamation of liposomes with PELNs could augment the cargo loading capability of PELNs. (C) PELNs can be loaded with the drugs of interest to promote PDLSC functions. (D) PELNs fusion with a membrane rich in special receptors can improve target bacteria or cells in the periodontal region. (E) Coating of membrane rich in special receptors in PELNs can target immune cells. Created with BioRender.com.
Conclusions
In conclusion, PELNs have shown many advantages including biocompatibility, specific cell targeting capability, cost-effectiveness, large-scale production, and drug delivery capability. The biological activities of PELNs including immune-regulation, effect on microbiome homeostasis, inflammation modulation, and tissue regeneration could be applied to the treatment of periodontitis. Although great progress has been obtained in the field of PELNs especially edible PELNs in the last decade, this field is still in its infancy. A lot of challenges should be overcome before the clinical application of PELNs. The relationship between components of natural products and PELNs has not been fully clarified. It is unclear which PELNs have therapeutic effects on periodontitis. The active components and mechanism of PELNs in the treatment of periodontitis are still unknown. In addition, although some PELNs show benefits to tissue regeneration, only one study had been reported so far regarding the therapeutic role of PELNs in periodontal disease. Based on the results from the recent literature, the direct use of PELNs to treat periodontitis or loading drugs and miRNAs, shRNAs, and siRNAs in PELNs to treat periodontitis could be a new era in the prevention and treatment of periodontitis.
Author Contributions
LY, LW, and JP conceived the manuscript. ZZ, YY, GZ, and LZ wrote the original draft. LW, JP, SX, HC, ZO, and JC revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (82150410451) and Guangdong Medical Research Foundation (A2019250): Long-chain noncoding RNA BMP2-1 affects human periodontal ligament stem cells by regulating BMP2.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank our colleagues and collaborators for their invaluable support in the investigation of PELNs.
References
1. Sanz M, Marco Del Castillo A, Jepsen S, Gonzalez-Juanatey JR, D'Aiuto F, Bouchard P, et al. Periodontitis and Cardiovascular Diseases: Consensus Report. J Clin Periodontol (2020) 47(3):268–88.
2. Collaborators GBDOD, Bernabe E, Marcenes W, Hernandez CR, Bailey J, Abreu LG, et al. Global, Regional, and National Levels and Trends in Burden of Oral Conditions From 1990 to 2017: A Systematic Analysis for the Global Burden of Disease 2017 Study. J Dent Res (2020) 99(4):362–73.
3. Kassebaum NJ, Bernabe E, Dahiya M, Bhandari B, Murray CJ, Marcenes W. Global Burden of Severe Periodontitis in 1990-2010: A Systematic Review and Meta-Regression. J Dent Res (2014) 93(11):1045–53.
4. Bui FQ, Almeida-da-Silva CLC, Huynh B, Trinh A, Liu J, Woodward J, et al. Association Between Periodontal Pathogens and Systemic Disease. BioMed J (2019) 42(1):27–35.
5. Hajishengallis G, Chavakis T. Local and Systemic Mechanisms Linking Periodontal Disease and Inflammatory Comorbidities. Nat Rev Immunol (2021) 21(7):426–40.
6. Rosa C, Gomes JML, Moraes SLD, Lemos CAA, da Fonte TP, Limirio J, et al. Use of Chlorhexidine Chip After Scaling and Root Planning on Periodontal Disease: A Systematic Review and Meta-Analysis. Saudi Dent J (2021) 33(1):1–10.
7. Zhao H, Hu J, Zhao L. Adjunctive Subgingival Application of Chlorhexidine Gel in Nonsurgical Periodontal Treatment for Chronic Periodontitis: A Systematic Review and Meta-Analysis. BMC Oral Health (2020) 20(1):34.
8. Hu D, Zhong T, Dai Q. Clinical Efficacy of Probiotics as an Adjunctive Therapy to Scaling and Root Planning in the Management of Periodontitis: A Systematic Review and Meta-Analysis of Randomized Controlled Trails. J Evid Based Dent Pract (2021) 21(2):101547.
9. Yuan H, Ma Q, Ye L, Piao G. The Traditional Medicine and Modern Medicine From Natural Products. Molecules (2016) 21(5):559.
10. Isola G. Current Evidence of Natural Agents in Oral and Periodontal Health. Nutrients (2020) 12(2):585.
11. Thomford NE, Senthebane DA, Rowe A, Munro D, Seele P, Maroyi A, et al. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int J Mol Sci (2018) 19(6):1578.
12. Basu A, Masek E, Ebersole JL. Dietary Polyphenols and Periodontitis-A Mini-Review of Literature. Molecules (2018) 23(7):1786.
13. Kharaeva ZF, Mustafaev MS, Khazhmetov AV, Gazaev IH, Blieva LZ, Steiner L, et al. Anti-Bacterial and Anti-Inflammatory Effects of Toothpaste With Swiss Medicinal Herbs Towards Patients Suffering From Gingivitis and Initial Stage of Periodontitis: From Clinical Efficacy to Mechanisms. Dent J (Basel) (2020) 8(1):10.
14. Halperin W, Jensen WA. Ultrastructural Changes During Growth and Embryogenesis in Carrot Cell Cultures. J Ultrastruct Res (1967) 18(3):428–43.
15. Sarvarian P, Samadi P, Gholipour E, Shams Asenjan K, Hojjat-Farsangi M, Motavalli R, et al. Application of Emerging Plant-Derived Nanoparticles as a Novel Approach for Nano-Drug Delivery Systems. Immunol Invest (2021) 1–21.
16. Yang C, Zhang M, Merlin D. Advances in Plant-Derived Edible Nanoparticle-Based Lipid Nano-Drug Delivery Systems as Therapeutic Nanomedicines. J Mater Chem B (2018) 6(9):1312–21.
17. Sriwastva MK, Deng ZB, Wang B, Teng Y, Kumar A, Sundaram K, et al. Exosome-Like Nanoparticles From Mulberry Bark Prevent DSS-Induced Colitis via the AhR/COPS8 Pathway. EMBO Rep (2022) 23(3):e53365.
18. Teng Y, Xu F, Zhang X, Mu J, Sayed M, Hu X, et al. Plant-Derived Exosomal microRNAs Inhibit Lung Inflammation Induced by Exosomes SARS-CoV-2 Nsp12. Mol Ther (2021) 29(8):2424–40.
19. Sundaram K, Miller DP, Kumar A, Teng Y, Sayed M, Mu J, et al. Plant-Derived Exosomal Nanoparticles Inhibit Pathogenicity of Porphyromonas Gingivalis. iScience (2019) 21:308–27.
20. Shi Q, Huo N, Wang X, Yang S, Wang J, Zhang T. Exosomes From Oral Tissue Stem Cells: Biological Effects and Applications. Cell Biosci (2020) 10:108.
21. Xie C, Ji N, Tang Z, Li J, Chen Q. The Role of Extracellular Vesicles From Different Origin in the Microenvironment of Head and Neck Cancers. Mol Cancer (2019) 18(1):83.
22. Kalluri R, LeBleu VS. The Biology, Function, and Biomedical Applications of Exosomes. Science (2020) 367(6478):eaau6977.
23. Meldolesi J. Exosomes and Ectosomes in Intercellular Communication. Curr Biol (2018) 28(8):R435–R44.
24. Xiao J, Feng S, Wang X, Long K, Luo Y, Wang Y, et al. Identification of Exosome-Like Nanoparticle-Derived microRNAs From 11 Edible Fruits and Vegetables. PeerJ (2018) 6:e5186.
25. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant Exosome-Like Nanovesicles: Emerging Therapeutics and Drug Delivery Nanoplatforms. Mol Ther (2021) 29(1):13–31.
26. Urzi O, Raimondo S, Alessandro R. Extracellular Vesicles From Plants: Current Knowledge and Open Questions. Int J Mol Sci (2021) 22(10):5366.
27. Sundaram GM. Dietary non-Coding RNAs From Plants: Fairy Tale or Treasure? Noncoding RNA Res (2019) 4(2):63–8.
28. Teng Y, Ren Y, Sayed M, Hu X, Lei C, Kumar A, et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe (2018) 24(5):637–52 e8.
29. Di Gioia S, Hossain MN, Conese M. Biological Properties and Therapeutic Effects of Plant-Derived Nanovesicles. Open Med (Wars) (2020) 15(1):1096–122.
30. Ju S, Mu J, Dokland T, Zhuang X, Wang Q, Jiang H, et al. Grape Exosome-Like Nanoparticles Induce Intestinal Stem Cells and Protect Mice From DSS-Induced Colitis. Mol Ther (2013) 21(7):1345–57.
31. Perut F, Roncuzzi L, Avnet S, Massa A, Zini N, Sabbadini S, et al. Strawberry-Derived Exosome-Like Nanoparticles Prevent Oxidative Stress in Human Mesenchymal Stromal Cells. Biomolecules (2021) 11(1):87.
32. Zhang M, Viennois E, Xu C, Merlin D. Plant Derived Edible Nanoparticles as a New Therapeutic Approach Against Diseases. Biomaterials (2016) 4(2):e1134415.
33. Pan S, Pei L, Zhang A, Zhang Y, Zhang C, Huang M, et al. Passion Fruit-Like Exosome-PMA/Au-BSA@Ce6 Nanovehicles for Real-Time Fluorescence Imaging and Enhanced Targeted Photodynamic Therapy With Deep Penetration and Superior Retention Behavior in Tumor. Biomaterials (2020) 230:119606.
34. Zhang M, Viennois E, Prasad M, Zhang Y, Wang L, Zhang Z, et al. Edible Ginger-Derived Nanoparticles: A Novel Therapeutic Approach for the Prevention and Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Biomaterials (2016) 101:321–40.
35. Jimenez-Jimenez S, Hashimoto K, Santana O, Aguirre J, Kuchitsu K, Cardenas L. Emerging Roles of Tetraspanins in Plant Inter-Cellular and Inter-Kingdom Communication. Plant Signal Behav (2019) 14(4):e1581559.
36. Zhang J, Li S, Li L, Li M, Guo C, Yao J, et al. Exosome and Exosomal microRNA: Trafficking, Sorting, and Function. Genomics Proteomics Bioinf (2015) 13(1):17–24.
37. Correia de Sousa M, Gjorgjieva M, Dolicka D, Sobolewski C, Foti M. Deciphering Mirnas' Action Through miRNA Editing. Int J Mol Sci (2019) 20(24):6249.
38. Jia M, He J, Bai W, Lin Q, Deng J, Li W, et al. Cross-Kingdom Regulation by Dietary Plant miRNAs: An Evidence-Based Review With Recent Updates. Food Funct (2021) 12(20):9549–62.
39. Yang J, Hotz T, Broadnax L, Yarmarkovich M, Elbaz-Younes I, Hirschi KD. Anomalous Uptake and Circulatory Characteristics of the Plant-Based Small RNA Mir2911. Sci Rep (2016) 6:26834.
40. Del Pozo-Acebo L, Lopez de Las Hazas MC, Margolles A, Davalos A, Garcia-Ruiz A. Eating microRNAs: Pharmacological Opportunities for Cross-Kingdom Regulation and Implications in Host Gene and Gut Microbiota Modulation. Br J Pharmacol (2021) 178(11):2218–45.
41. Alshehri B. Plant-Derived xenomiRs and Cancer: Cross-Kingdom Gene Regulation. Saudi J Biol Sci (2021) 28(4):2408–22.
42. Akuma P, Okagu OD, Udenigwe CC. Naturally Occurring Exosome Vesicles as Potential Delivery Vehicle for Bioactive Compounds. Front. Sustain. Food Syst (2019) 3:23.
43. Wang J, Ding Y, Wang J, Hillmer S, Miao Y, Lo SW, et al. EXPO, an Exocyst-Positive Organelle Distinct From Multivesicular Endosomes and Autophagosomes, Mediates Cytosol to Cell Wall Exocytosis in Arabidopsis and Tobacco Cells. Plant Cell (2010) 22(12):4009–30.
44. Babst M. MVB Vesicle Formation: ESCRT-Dependent, ESCRT-Independent and Everything in Between. Curr Opin Cell Biol (2011) 23(4):452–7.
45. Liu G, Kang G, Wang S, Huang Y, Cai Q. Extracellular Vesicles: Emerging Players in Plant Defense Against Pathogens. Front Plant Sci (2021) 12:757925.
46. US H, Brotherton D, Inal J. Communication is Key: Extracellular Vesicles as Mediators of Infection and Defence During Host-Microbe Interactions in Animals and Plants. FEMS Microbiol Rev (2022) 46(1):fuab044.
47. He B, Cai Q, Qiao L, Huang CY, Wang S, Miao W, et al. RNA-Binding Proteins Contribute to Small RNA Loading in Plant Extracellular Vesicles. Nat Plants (2021) 7(3):342–52.
48. Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: Biogenesis, Biologic Function and Clinical Potential. Cell Biosci (2019) 9:19.
49. Minciacchi VR, Freeman MR, Di Vizio D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin Cell Dev Biol (2015) 40:41–51.
50. He B, Hamby R, Jin H. Plant Extracellular Vesicles: Trojan Horses of Cross-Kingdom Warfare. FASEB Bioadv (2021) 3(9):657–64.
51. Alfieri M, Leone A, Ambrosone A. Plant-Derived Nano and Microvesicles for Human Health and Therapeutic Potential in Nanomedicine. Pharmaceutics (2021) 13(4):498.
52. Rome S. Biological Properties of Plant-Derived Extracellular Vesicles. Food Funct (2019) 10(2):529–38.
53. Chen X, Zhou Y, Yu J. Exosome-Like Nanoparticles From Ginger Rhizomes Inhibited NLRP3 Inflammasome Activation. Mol Pharm (2019) 16(6):2690–9.
54. Stanly C, Fiume I, Capasso G, Pocsfalvi G. Isolation of Exosome-Like Vesicles From Plants by Ultracentrifugation on Sucrose/Deuterium Oxide (D2O) Density Cushions. Methods Mol Biol (2016) 1459:259–69.
55. You JY, Kang SJ, Rhee WJ. Isolation of Cabbage Exosome-Like Nanovesicles and Investigation of Their Biological Activities in Human Cells. Bioact Mater (2021) 6(12):4321–32.
56. Kalarikkal SP, Sundaram GM. Edible Plant-Derived Exosomal microRNAs: Exploiting a Cross-Kingdom Regulatory Mechanism for Targeting SARS-CoV-2. Toxicol Appl Pharmacol (2021) 414:115425.
57. Yu L, Deng Z, Liu L, Zhang W, Wang C. Plant-Derived Nanovesicles: A Novel Form of Nanomedicine. Front Bioeng Biotechnol (2020) 8:584391.
58. Suresh AP, Kalarikkal SP, Pullareddy B, Sundaram GM. Low pH-Based Method to Increase the Yield of Plant-Derived Nanoparticles From Fresh Ginger Rhizomes. ACS Omega (2021) 6(27):17635–41.
59. Chen C, Wang J, Sun M, Li J, Wang HD. Toward the Next-Generation Phyto-Nanomedicines: Cell-Derived Nanovesicles (CDNs) for Natural Product Delivery. BioMed Pharmacother (2022) 145:112416.
60. De Robertis M, Sarra A, D'Oria V, Mura F, Bordi F, Postorino P, et al. Blueberry-Derived Exosome-Like Nanoparticles Counter the Response to TNF-Alpha-Induced Change on Gene Expression in EA.hy926 Cells. Biomolecules (2020) 10(5):742.
61. Gonda A, Kabagwira J, Senthil GN, Wall NR. Internalization of Exosomes Through Receptor-Mediated Endocytosis. Mol Cancer Res (2019) 17(2):337–47.
62. Wei H, Chen Q, Lin L, Sha C, Li T, Liu Y, et al. Regulation of Exosome Production and Cargo Sorting. Int J Biol Sci (2021) 17(1):163–77.
63. McKelvey KJ, Powell KL, Ashton AW, Morris JM, McCracken SA. Exosomes: Mechanisms of Uptake. J Circ biomark (2015) 4:7.
64. Joshi BS, de Beer MA, Giepmans BNG, Zuhorn IS. Endocytosis of Extracellular Vesicles and Release of Their Cargo From Endosomes. ACS Nano (2020) 14(4):4444–55.
65. Aqil F, Jeyabalan J, Agrawal AK, Kyakulaga AH, Munagala R, Parker L, et al. Exosomal Delivery of Berry Anthocyanidins for the Management of Ovarian Cancer. Food Funct (2017) 8(11):4100–7.
66. Baldini N, Torreggiani E, Roncuzzi L, Perut F, Zini N, Avnet S. Exosome-Like Nanovesicles Isolated From Citrus Limon L. Exert Antioxidative Effect. Curr Pharm Biotechnol (2018) 19(11):877–85.
67. Zhang L, He F, Gao L, Cong M, Sun J, Xu J, et al. Engineering Exosome-Like Nanovesicles Derived From Asparagus Cochinchinensis Can Inhibit the Proliferation of Hepatocellular Carcinoma Cells With Better Safety Profile. Int J Nanomedicine (2021) 16:1575–86.
68. Graziani F, Karapetsa D, Alonso B, Herrera D. Nonsurgical and Surgical Treatment of Periodontitis: How Many Options for One Disease? Periodontol 2000 (2017) 75(1):152–88.
69. Heitz-Mayfield LJ, Lang NP. Surgical and Nonsurgical Periodontal Therapy. Learned unlearned concepts. Periodontol 2000 (2013) 62(1):218–31.
70. Nyman S, Lindhe J, Karring T, Rylander H. New Attachment Following Surgical Treatment of Human Periodontal Disease. J Clin Periodontol (1982) 9(4):290–6.
71. Nibali L, Koidou VP, Nieri M, Barbato L, Pagliaro U, Cairo F. Regenerative Surgery Versus Access Flap for the Treatment of Intra-Bony Periodontal Defects: A Systematic Review and Meta-Analysis. J Clin Periodontol (2020) 47 Suppl 22:320–51.
72. Sculean A, Nikolidakis D, Nikou G, Ivanovic A, Chapple IL, Stavropoulos A. Biomaterials for Promoting Periodontal Regeneration in Human Intrabony Defects: A Systematic Review. Periodontol 2000 (2015) 68(1):182–216.
73. Addy M, Martin MV. Systemic Antimicrobials in the Treatment of Chronic Periodontal Diseases: A Dilemma. Oral Dis (2003) 9 Suppl 1:38–44.
74. Rabelo CC, Feres M, Goncalves C, Figueiredo LC, Faveri M, Tu YK, et al. Systemic Antibiotics in the Treatment of Aggressive Periodontitis. A Systematic Review and a Bayesian Network Meta-Analysis. J Clin Periodontol (2015) 42(7):647–57.
75. Teughels W, Feres M, Oud V, Martin C, Matesanz P, Herrera D. Adjunctive Effect of Systemic Antimicrobials in Periodontitis Therapy: A Systematic Review and Meta-Analysis. J Clin Periodontol (2020) 47 Suppl 22:257–81.
76. Hbibi A, Sikkou K, Khedid K, El Hamzaoui S, Bouziane A, Benazza D. Antimicrobial Activity of Honey in Periodontal Disease: A Systematic Review. J Antimicrob Chemother (2020) 75(4):807–26.
77. Tambur Z, Miljkovic-Selimovic B, Opacic D, Vukovic B, Malesevic A, Ivancajic L, et al. Inhibitory Effects of Propolis and Essential Oils on Oral Bacteria. J Infect Dev Ctries (2021) 15(7):1027–31.
79. Farinon B, Molinari R, Costantini L, Merendino N. The Seed of Industrial Hemp (Cannabis Sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients (2020) 12(7):1935.
80. Stasilowicz A, Tomala A, Podolak I, Cielecka-Piontek J, Cannabis sativa L. As a Natural Drug Meeting the Criteria of a Multitarget Approach to Treatment. Int J Mol Sci (2021) 22(2):778.
81. Napimoga MH, Benatti BB, Lima FO, Alves PM, Campos AC, Pena-Dos-Santos DR, et al. Cannabidiol Decreases Bone Resorption by Inhibiting RANK/RANKL Expression and Pro-Inflammatory Cytokines During Experimental Periodontitis in Rats. Int Immunopharmacol (2009) 9(2):216–22.
82. Lowe H, Toyang N, Steele B, Bryant J, Ngwa W, Nedamat K. The Current and Potential Application of Medicinal Cannabis Products in Dentistry. Dent J (Basel) (2021) 9(9):106.
83. Kaboosaya B, Wulansari LK, Trang Nguyen VN, Kasugai S. Drinking Green Tea Alleviates Alveolar Bone Resorption in Ligature-Induced Periodontitis in Mice. J Oral Biosci (2020) 62(2):162–8.
84. Vargas-Sanchez PK, Pitol DL, de Sousa LG, Beloti MM, Rosa AL, Rossi AC, et al. Green Tea Extract Rich in Epigallocatechin Gallate Impairs Alveolar Bone Loss in Ovariectomized Rats With Experimental Periodontal Disease. Int J Exp Pathol (2020) 101(6):277–88.
85. Sarin S, Marya C, Nagpal R, Oberoi SS, Rekhi A. Preliminary Clinical Evidence of the Antiplaque, Antigingivitis Efficacy of a Mouthwash Containing 2% Green Tea - A Randomised Clinical Trial. Oral Health Prev Dent (2015) 13(3):197–203.
86. Mu J, Zhuang X, Wang Q, Jiang H, Deng ZB, Wang B, et al. Interspecies Communication Between Plant and Mouse Gut Host Cells Through Edible Plant Derived Exosome-Like Nanoparticles. Mol Nutr Food Res (2014) 58(7):1561–73.
87. Zhao WJ, Bian YP, Wang QH, Yin F, Yin L, Zhang YL, et al. Blueberry-Derived Exosomes-Like Nanoparticles Ameliorate Nonalcoholic Fatty Liver Disease by Attenuating Mitochondrial Oxidative Stress. Acta Pharmacol Sin (2022) 43(3):645–58.
88. Aquilano K, Ceci V, Gismondi A, De Stefano S, Iacovelli F, Faraonio R, et al. Adipocyte Metabolism is Improved by TNF Receptor-Targeting Small RNAs Identified From Dried Nuts. Commun Biol (2019) 2:317.
89. Cao M, Yan H, Han X, Weng L, Wei Q, Sun X, et al. Ginseng-Derived Nanoparticles Alter Macrophage Polarization to Inhibit Melanoma Growth. J Immunother Cancer (2019) 7(1):326.
90. Liu B, Li X, Yu H, Shi X, Zhou Y, Alvarez S, et al. Therapeutic Potential of Garlic Chive-Derived Vesicle-Like Nanoparticles in NLRP3 Inflammasome-Mediated Inflammatory Diseases. Theranostics (2021) 11(19):9311–30.
91. Bruno SP, Paolini A, D'Oria V, Sarra A, Sennato S, Bordi F, et al. Extracellular Vesicles Derived From Citrus Sinensis Modulate Inflammatory Genes and Tight Junctions in a Human Model of Intestinal Epithelium. Front Nutr (2021) 8:778998.
92. Zu M, Xie D, Canup BSB, Chen N, Wang Y, Sun R, et al. 'Green' Nanotherapeutics From Tea Leaves for Orally Targeted Prevention and Alleviation of Colon Diseases. Biomaterials (2021) 279:121178.
93. Lei C, Teng Y, He L, Sayed M, Mu J, Xu F, et al. Lemon Exosome-Like Nanoparticles Enhance Stress Survival of Gut Bacteria by RNase P-Mediated Specific tRNA Decay. iScience (2021) 24(6):102511.
94. Lei C, Mu J, Teng Y, He L, Xu F, Zhang X, et al. Lemon Exosome-Like Nanoparticles-Manipulated Probiotics Protect Mice From C. d iff Infection iScience (2020) 23(10):101571.
95. Sahin F, Kocak P, Gunes MY, Ozkan I, Yildirim E, Kala EY. In Vitro Wound Healing Activity of Wheat-Derived Nanovesicles. Appl Biochem Biotechnol (2019) 188(2):381–94.
96. Cho JH, Hong YD, Kim D, Park SJ, Kim JS, Kim H-M, et al. Confirmation of Plant-Derived Exosomes as Bioactive Substances for Skin Application Through Comparative Analysis of Keratinocyte Transcriptome. Appl Biol Chem (2022) 65(1):8.
97. Sun X, Gao J, Meng X, Lu X, Zhang L, Chen R. Polarized Macrophages in Periodontitis: Characteristics, Function, and Molecular Signaling. Front Immunol (2021) 12:763334.
98. Ni C, Zhou J, Kong N, Bian T, Zhang Y, Huang X, et al. Gold Nanoparticles Modulate the Crosstalk Between Macrophages and Periodontal Ligament Cells for Periodontitis Treatment. Biomaterials (2019) 206:115–32.
99. Garaicoa-Pazmino C, Fretwurst T, Squarize CH, Berglundh T, Giannobile WV, Larsson L, et al. Characterization of Macrophage Polarization in Periodontal Disease. J Clin Periodontol (2019) 46(8):830–9.
100. Zhang H, Wang L, Li C, Yu Y, Yi Y, Wang J, et al. Exosome-Induced Regulation in Inflammatory Bowel Disease. Front Immunol (2019) 10:1464.
101. Deng Z, Rong Y, Teng Y, Mu J, Zhuang X, Tseng M, et al. Broccoli-Derived Nanoparticle Inhibits Mouse Colitis by Activating Dendritic Cell AMP-Activated Protein Kinase. Mol Ther (2017) 25(7):1641–54.
102. Sharma G, Goyal H, Thakur K, Raza K, Katare OP. Novel Elastic Membrane Vesicles (EMVs) and Ethosomes-Mediated Effective Topical Delivery of Aceclofenac: A New Therapeutic Approach for Pain and Inflammation. Drug Deliv (2016) 23(8):3135–45.
103. Mao Y, Han M, Chen C, Wang X, Han J, Gao Y, et al. A Biomimetic Nanocomposite Made of a Ginger-Derived Exosome and an Inorganic Framework for High-Performance Delivery of Oral Antibodies. Nanoscale (2021) 13(47):20157–69.
104. Zandbergen D, Slot DE, Niederman R, van der Weijden FA. The Concomitant Administration of Systemic Amoxicillin and Metronidazole Compared to Scaling and Root Planing Alone in Treating Periodontitis: =a Systematic Review=. BMC Oral Health (2016) 16:27.
105. Bodet C, Chandad F, Grenier D. [Pathogenic Potential of Porphyromonas Gingivalis, Treponema Denticola and Tannerella Forsythia, the Red Bacterial Complex Associated With Periodontitis]. Pathol Biol (Paris) (2007) 55(3-4):154–62.
106. Zhang Y, Wang X, Li H, Ni C, Du Z, Yan F. Human Oral Microbiota and its Modulation for Oral Health. BioMed Pharmacother (2018) 99:883–93.
107. Bayat F, Afshar A, Baghban N. Algal Cells-Derived Extracellular Vesicles: A Review With Special Emphasis on Their Antimicrobial Effects. Front Microbiol (2021) 12:785716.
108. Cai Q, Qiao L, Wang M, He B, Lin FM, Palmquist J, et al. Plants Send Small RNAs in Extracellular Vesicles to Fungal Pathogen to Silence Virulence Genes. Science (2018) 360(6393):1126–9.
109. Diez-Sainz E, Lorente-Cebrian S, Aranaz P, Riezu-Boj JI, Martinez JA, Milagro FI. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front Nutr (2021) 8:586564.
110. Han J, Menicanin D, Gronthos S, Bartold PM. Stem Cells, Tissue Engineering and Periodontal Regeneration. Aust Dent J (2014) 59 Suppl 1:117–30.
111. Liu J, Ruan J, Weir MD, Ren K, Schneider A, Wang P, et al. Periodontal Bone-Ligament-Cementum Regeneration via Scaffolds and Stem Cells. Cells (2019) 8(6):537.
112. Ding T, Kang W, Li J, Yu L, Ge S. An in Situ Tissue Engineering Scaffold With Growth Factors Combining Angiogenesis and Osteoimmunomodulatory Functions for Advanced Periodontal Bone Regeneration. J Nanobiotechnol (2021) 19(1):247.
113. Qin Y, Sun R, Wu C, Wang L, Zhang C. Exosome: A Novel Approach to Stimulate Bone Regeneration Through Regulation of Osteogenesis and Angiogenesis. Int J Mol Sci (2016) 17(5):712.
114. Rahimi Ghiasi M, Rahimi E, Amirkhani Z, Salehi R. Leucine-Rich Repeat-Containing G-Protein Coupled Receptor 5 Gene Overexpression of the Rat Small Intestinal Progenitor Cells in Response to Orally Administered Grape Exosome-Like Nanovesicles. Adv BioMed Res (2018) 7:125.
115. Watanabe Y, Tsuchiya A, Terai S. The Development of Mesenchymal Stem Cell Therapy in the Present, and the Perspective of Cell-Free Therapy in the Future. Clin Mol Hepatol (2021) 27(1):70–80.
116. Fernandes M, Lopes I, Teixeira J, Botelho C, Gomes AC. Exosome-Like Nanoparticles: A New Type of Nanocarrier. Curr Med Chem (2020) 27(23):3888–905.
117. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering Precision Nanoparticles for Drug Delivery. Nat Rev Drug Discov (2021) 20(2):101–24.
118. Arrighetti N, Corbo C, Evangelopoulos M, Pasto A, Zuco V, Tasciotti E. Exosome-Like Nanovectors for Drug Delivery in Cancer. Curr Med Chem (2019) 26(33):6132–48.
119. Umezu T, Takanashi M, Murakami Y, Ohno SI, Kanekura K, Sudo K, et al. Acerola Exosome-Like Nanovesicles to Systemically Deliver Nucleic Acid Medicine via Oral Administration. Mol Ther Methods Clin Dev (2021) 21:199–208.
120. Shi X, Cheng Q, Zhang Y. Reprogramming Extracellular Vesicles With Engineered Proteins. Methods (2020) 177:95–102.
121. Mukherjee A, Waters AK, Kalyan P, Achrol AS, Kesari S, Yenugonda VM. Lipid-Polymer Hybrid Nanoparticles as a Next-Generation Drug Delivery Platform: State of the Art, Emerging Technologies, and Perspectives. Int J Nanomedicine (2019) 14:1937–52.
Keywords: plant-derived exosome-like nanoparticles, periodontitis, exosomes, inflammation, oral tissue regeneration, drug delivery systems, extracellular vesicles
Citation: Zhang Z, Yu Y, Zhu G, Zeng L, Xu S, Cheng H, Ouyang Z, Chen J, Pathak JL, Wu L and Yu L (2022) The Emerging Role of Plant-Derived Exosomes-Like Nanoparticles in Immune Regulation and Periodontitis Treatment. Front. Immunol. 13:896745. doi: 10.3389/fimmu.2022.896745
Received: 15 March 2022; Accepted: 16 May 2022;
Published: 10 June 2022.
Edited by:
Elham Hosseini-Beheshti, The University of Sydney, AustraliaReviewed by:
Subhash Chand, University of Nebraska Medical Center, United StatesMichael Merchant, University of Louisville, United States
Copyright © 2022 Zhang, Yu, Zhu, Zeng, Xu, Cheng, Ouyang, Chen, Pathak, Wu and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Janak L. Pathak, ai5wYXRoYWtAZ3pobXUuZWR1LmNu; Lihong Wu, d2NhbmhvbmdAMTYzLmNvbQ==; Lina Yu, ODQwODE2ODFAcXEuY29t
†These authors have contributed equally to this work and share first authorship