
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 05 April 2022
Sec. Viral Immunology
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.857322
This article is part of the Research TopicDeleterious and Beneficial Humoral Immune Response in Viral Diseases: Two Sides of the Same CoinView all 9 articles
 Edmilson F. de Oliveira-Filho1
Edmilson F. de Oliveira-Filho1 Otávio V. de Carvalho2
Otávio V. de Carvalho2 Ianei O. Carneiro3
Ianei O. Carneiro3 Fagner D’ambroso Fernandes4
Fagner D’ambroso Fernandes4 Sara Nunes Vaz5Célia Pedroso5Lilian Gonzalez-Auza1Victor Carvalho Urbieta1Arne Kühne1
Sara Nunes Vaz5Célia Pedroso5Lilian Gonzalez-Auza1Victor Carvalho Urbieta1Arne Kühne1 Rafaela Mayoral3
Rafaela Mayoral3 Wendy K. Jo1
Wendy K. Jo1 Andrés Moreira-Soto1Chantal B. E. M. Reusken6Christian Drosten1
Andrés Moreira-Soto1Chantal B. E. M. Reusken6Christian Drosten1 Carlos Brites5Klaus Osterrieder7Eduardo Martins Netto5Luiz Eduardo Ristow2Rita de Cassia Maia8Fernanda S. Flores Vogel4
Carlos Brites5Klaus Osterrieder7Eduardo Martins Netto5Luiz Eduardo Ristow2Rita de Cassia Maia8Fernanda S. Flores Vogel4 Nadia Rossi de Almeida3Carlos Roberto Franke3
Nadia Rossi de Almeida3Carlos Roberto Franke3 Jan Felix Drexler1,9*
Jan Felix Drexler1,9*Carnivores such as cats and minks are highly susceptible to SARS-CoV-2. Brazil is a global COVID-19 hot spot and several cases of human-to-cat transmission have been documented. We investigated the spread of SARS-CoV-2 by testing 547 domestic cats sampled between July-November 2020 from seven states in southern, southeastern, and northeastern Brazil. Moreover, we investigated whether immune responses elicited by enzootic coronaviruses affect SARS-CoV-2 infection in cats. We found infection with significantly higher neutralizing antibody titers against the Gamma variant of concern, endemic in Brazil during 2020, than against an early SARS-CoV-2 B.1 isolate (p<0.0001), validating the use of Gamma for further testing. The overall SARS-CoV-2 seroprevalence in Brazilian cats during late 2020 validated by plaque reduction neutralization test (PRNT90) was 7.3% (95% CI, 5.3-9.8). There was no significant difference in SARS-CoV-2 seroprevalence in cats between Brazilian states, suggesting homogeneous infection levels ranging from 4.6% (95% CI, 2.2-8.4) to 11.4% (95% CI, 6.7-17.4; p=0.4438). Seroprevalence of the prototypic cat coronavirus Feline coronavirus (FCoV) in a PRNT90 was high at 33.3% (95% CI, 24.9-42.5) and seroprevalence of Bovine coronavirus (BCoV) was low at 1.7% (95% CI, 0.2-5.9) in a PRNT90. Neutralizing antibody titers were significantly lower for FCoV than for SARS-CoV-2 (p=0.0001), consistent with relatively more recent infection of cats with SARS-CoV-2. Neither the magnitude of SARS-CoV-2 antibody titers (p=0.6390), nor SARS-CoV-2 infection status were affected by FCoV serostatus (p=0.8863). Our data suggest that pre-existing immunity against enzootic coronaviruses neither prevents, nor enhances SARS-CoV-2 infection in cats. High SARS-CoV-2 seroprevalence already during the first year of the pandemic substantiates frequent infection of domestic cats and raises concerns on potential SARS-CoV-2 mutations escaping human immunity upon spillback.
SARS-CoV-2 has evolutionary origins in bats and emerged in humans during late 2019. Beyond humans, carnivores are particularly susceptible to SARS-CoV-2 (1). Among carnivores, natural SARS-CoV-2 infection has been shown in dogs, cats, lions, tigers, ferrets, and minks by late 2021 (2–7). Clinical presentation of SARS-CoV-2-infected cats includes respiratory and gastroenteric symptoms such as sneezing, coughing, nasal and ocular discharge, anorexia, vomiting, diarrhea and appetite loss, but COVID-19 in cats remains poorly described (4, 8). Molecular and serological evidence from several countries showed that cats can easily be infected with SARS-CoV-2, predominantly from infected humans, but also from minks and other cats (3, 5, 9, 10). During the 2003-2004 outbreak of SARS-CoV, it was speculated that mutations enabling efficient infection of humans may have arisen in carnivore intermediate hosts (1). In minks infected with SARS-CoV-2, mutations have arisen in a variant strain capable of infecting humans and showing reduced antibody-mediated neutralization by human serum samples (11–13).
Different from minks that are kept as farmed animals in some countries, over 370 million cats globally live in close contact with humans (see: https://www.statista.com/statistics/1044386/dog-and-cat-pet-population-worldwide/). Brazil, with roughly 24 million cats and 210 million inhabitants, is one of the countries most affected by the COVID-19 pandemic with high incidence, different variants of concern (VOC), and more than 600,000 deaths by the end of 2021 (14, 15). Household transmission from humans to cats has been reported in Brazil (16), but the epidemiology of SARS-CoV-2 infection in cats and its distribution over the country remain unknown.
Pre-existing coronavirus immunity may be protective against subsequent infection with SARS-CoV-2 (17, 18). In contrast, pre-existing immunity can also have deleterious effects, such as antibody-dependent enhancement (ADE) known to contribute to severe Dengue (19, 20). Among coronaviruses, ADE has been described during experimental Feline Coronavirus (FCoV) infection (21) and in consequence of vaccination with a recombinant vaccinia virus-based vaccine (22, 23). It is not known whether pre-existing immunity against enzootic carnivore coronaviruses would provide protection against or immune enhancement of SARS-CoV-2 infection. Additionally, previous infection by endemic human coronaviruses and other infection can cause false-positive results in SARS-CoV-2 antibody tests (18, 24–27), and careful assessments of coronavirus immunity are thus required irrespective of the host species. Here, we investigated SARS-CoV-2 seroprevalence in domestic cats from different Brazilian states and analyzed whether pre-existing coronavirus immunity affects SARS-CoV-2 diagnostics and infection.
A total of 547 pandemic and 52 pre-pandemic serum samples from cats presenting miscellaneous clinical signs, including, among others, fever, weakness, lack of appetite, gastroenteritis, were sampled for routine laboratory diagnostics. Pandemic sera were sampled from July-November 2020 from seven states in Brazil (Table 1) and pre-pandemic sera were sampled in the states of Bahia and Rio Grande do Sul between May 2018 and November 2019. Based on the information available for 172 animals, the median age was 4 years, and the sex ratio (F/M) was 0.68 (70/102), suggesting our samples comprised mainly adult female animals. Sampling was approved by the Federal University of Bahia’s animal ethics committee under authorization no. 74/2019.
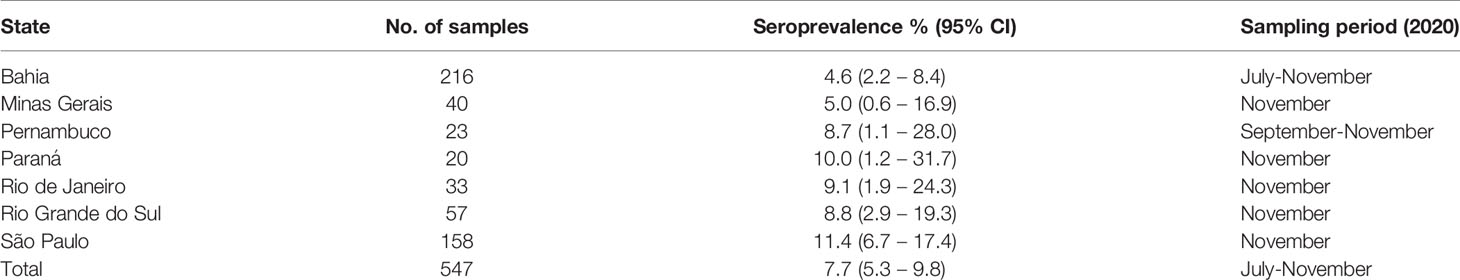
Table 1 SARS-CoV-2 seroprevalence in cats from different states in Brazil.
For antibody screening we used a highly specific receptor-binding domain (RBD)-based SARS-CoV-2 surrogate virus neutralization test (sVNT) (cPass, GenScript, https://www.genscript.com), which does not require species-specific detection antibodies and should thus be usable in cats.
We performed plaque reduction neutralization tests (PRNT) and compared the reciprocal endpoint titers against SARS-CoV-2 with those against enzootic coronaviruses belonging to the viral species Alphacoronavirus 1 and Betacoronavirus 1 (Figure 1A). For SARS-CoV-2 we used the Gamma variant of concern (VOC) (strain hCoV-19/Netherlands/NoordHolland_10915/2021, purchased from the European Virus Archive Global) and an early B.1 isolate (Munich/ChVir929/2020 strain, sampled in January 2020; Gisaid accession: EPI_ISL_406862). For enzootic coronaviruses, we used FCoV (ATCC VR-989, WSU 79-1683 strain) and Bovine Coronavirus (BCoV) (strain Kakegawa) (Figure 1A). PRNT90 (a serum dilution reducing viral plaques by ≥90% is considered positive) was conducted in cell monolayers of 1.6 × 105 Vero E6 for SARS-CoV-2, 3.5 × 105 Crandell-Rees Feline Kidney cells (CRFK) for FCoV and 3.0 × 105 sheep epithelioid cells (PT) (CCLV-RIE 0011) for BCoV. Cells were seeded in 12-well plates one day before the infection. Sixty plaque-forming units were incubated with serum dilutions of 1:20, 1:80, 1:320, and 1:1280 for one hour, added onto the cell monolayer, incubated again for one hour before adding the overlayer containing DMEM with 1% FCS and 2% Avicell for SARS-CoV-2 and 1.25% carboxymethyl cellulose (CMC) for FCoV and BCoV. After two days for SARS-CoV-2, four days for BCoV and five days for FCoV, the overlayer was removed, cells fixated with 6% paraformaldehyde and stained with crystal violet.
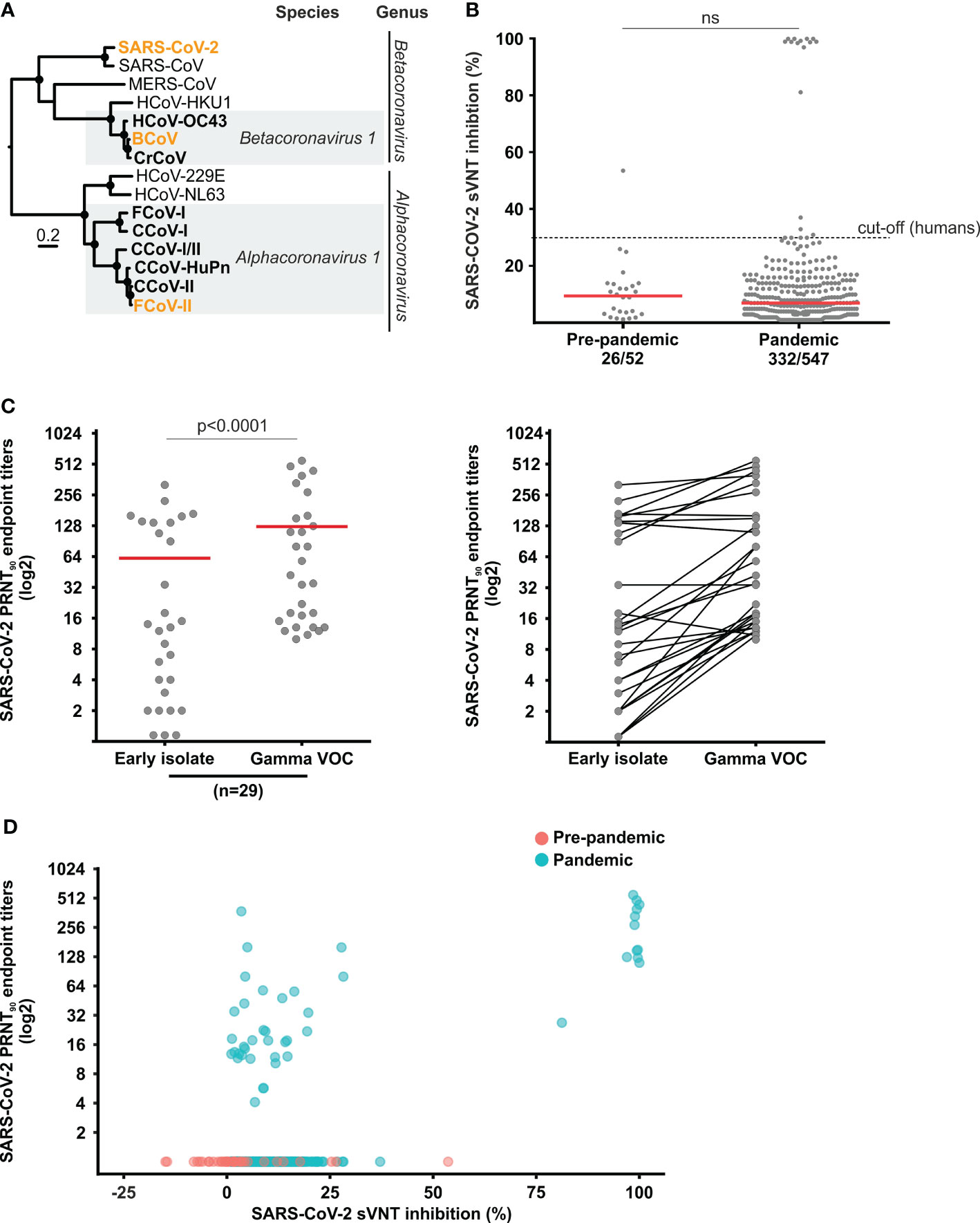
Figure 1 Coronavirus phylogeny and set-up of SARS-CoV-2 serology. (A) Maximum-likelihood tree based on translated spike gene sequences of human, dog and cat coronaviruses. The strains used for testing in this study are highlighted in orange. WAG+G+I was used as a substitution model and a complete deletion option was chosen. Scale bar indicates amino acid substitutions per site. Circles at nodes indicate support of grouping in ≥75% from 1,000 bootstrap replicates. (B) sVNT inhibition rate in sera from cats sampled before and during the pandemic. The 30% inhibition cut-off above which a human serum sample is considered positive is given for comparison. (C) Comparison of SARS-CoV-2 PRNT90 endpoint titers between an early isolate and a Gamma VOC strain. Statistical significance was determined using the Wilcoxon matched-pairs signed rank test. (D) Comparison of sVNT inhibition rates and reciprocal PRNT endpoint titers. Red bars in panels (B, C) indicate medians; ns, not statistically significant.
Endpoint reciprocal titers were calculated using a logistic regression function and statistical tests as indicated in the text and figure legends were done in GraphPad prism 6 (GraphPad Software, www.graphpad.com).
To investigate whether pre-existing immune responses against enzootic coronaviruses or other pathogens affect SARS-CoV-2 antibody testing, we tested 52 pre-pandemic sera in a the receptor-binding domain (RBD)-based SARS-CoV-2 surrogate virus neutralization test (sVNT) considered highly specific for human antibody responses (28). Surprisingly, one cat sample (1.9%, 95% CI, 0.1-10.6) was clearly positive (signal inhibition ≥30.0%) and several other samples showed reactivity close to the sVNT threshold above which test results are considered positive in human-derived samples (Figure 1B). This suggested that sVNT results needed to be confirmed by PRNT. Because the emergence of the Gamma VOC in Brazil between June-November 2020 (14) overlaps with our sampling, we compared the PRNT90 endpoint titers against the Gamma VOC and an early SARS-CoV-2 isolate in a subset of PRNT-positive serum samples with sufficient volume. Our data confirmed significantly higher PRNT90 endpoint titers for the Gamma VOC in 24 out of 29 (82.8%) positive samples (Wilcoxon matched-pairs signed rank test, p<0.0001) (Figure 1C), which led us to conduct subsequent confirmatory testing only with the Gamma VOC strain. Because of relatively lower sVNT sensitivity due to usage of a spike protein RBD from an early isolate that may afford decreased reactivity with antibodies raised by contemporary strains in Brazil in the sVNT, all reactive sera (signal inhibition ratio ≥1.0) were confirmed by plaque reduction neutralization test (PRNT90) using the Gamma VOC, instead of using the 30% threshold used for testing of humans. Lastly, none of the pre-pandemic sera showing reactivity in the sVNT could be confirmed in the SARS-CoV-2 PRNT90, highlighting the risk of false-positive test results even when using a test validated for confirmatory human testing in cats (Figure 1D).
To assess the SARS-CoV-2 seroprevalence in cats in Brazil, we tested a total of 547 sera sampled during the second half of 2020 in seven Brazilian states spanning 3000 km longitude. The overall SARS-CoV-2 seroprevalence confirmed by PRNT90 was 7.3% (95% CI, 5.3-9.8), ranging from 4.6 (95% CI, 2.2-8.4) to 11.4 (95% CI, 6.7-17.4) in different states of Brazil (Table 1 and Figure 2A). The regional differences in seroprevalence in cats were not statistically significant (chi-square, p=0.4438), despite differences between the COVID-19 incidence in humans in those Brazilian states (Figure 2B). Cumulative incidence in humans and cat seroprevalence were not significantly correlated (rho=-0.0982, p=0.8341; Figure 2C), although the rates were similar in some Brazilian states (Supplementary Figure 1). Whether cats can thus be used as a proxy for human infection remains to be determined using representative samples.
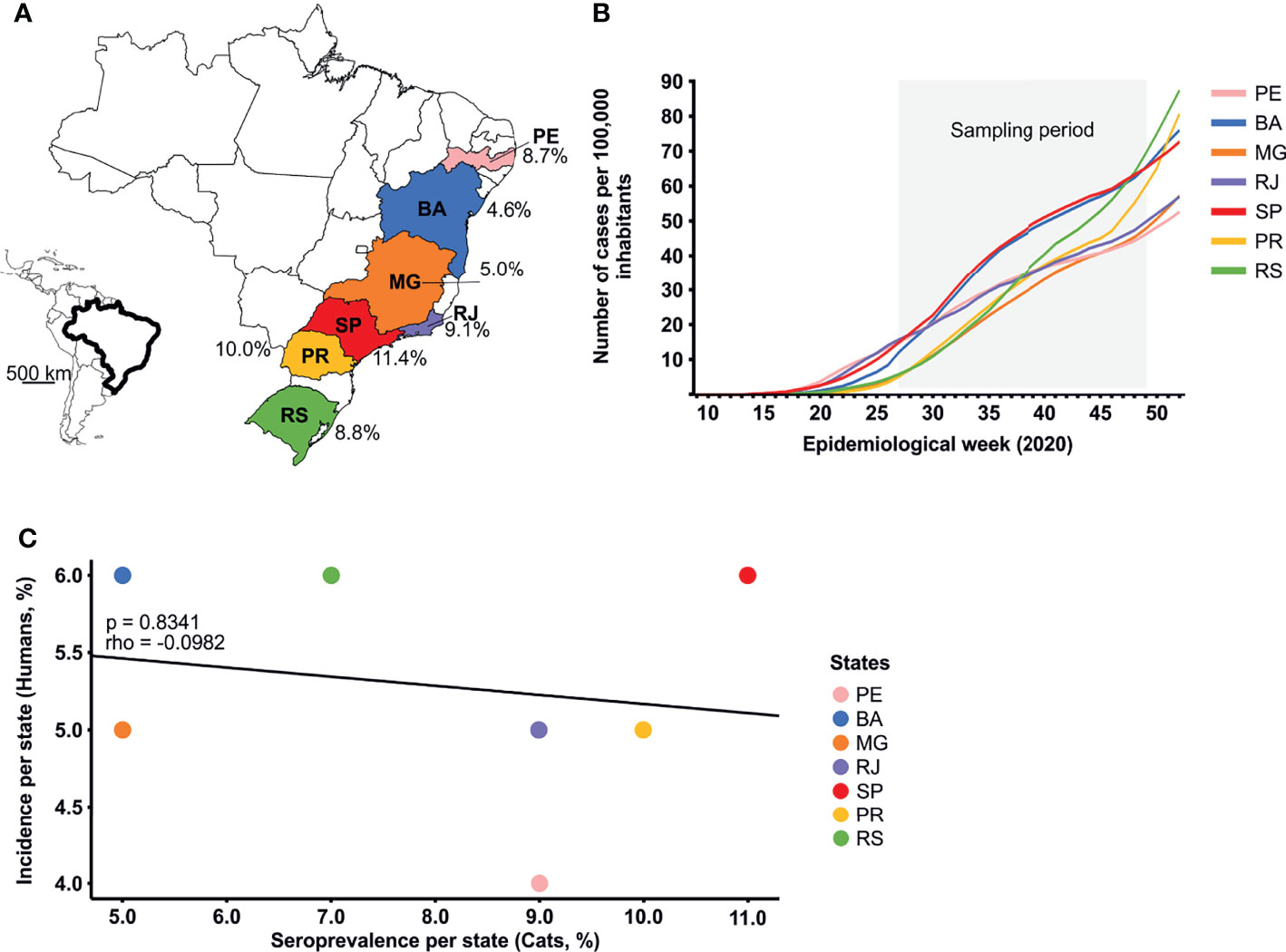
Figure 2 Sampling sites and COVID-19 incidence. (A) SARS-CoV-2 seroprevalence in cats in different states in Brazil. (B) Weekly SARS-CoV-2 incidence per 100,000 habitants among different states in Brazil. (C) Spearman’s rank correlation comparing SARS-CoV-2 seroprevalence in cats and cumulative cases in humans per state by November 2020 in Brazil. BA, Bahia; MG, Minas Gerais; PE, Pernambuco; PR, Paraná; RJ, Rio de Janeiro; RS, Rio Grande do Sul; SP, São Paulo.
We tested all 52 pre-pandemic sera and a subset of 68 pandemic sera showing sufficient volumes against FCoV and BCoV, including all 42 samples that were seropositive for SARS-CoV-2.
Only two of those 120 sera contained BCoV-specific neutralizing antibodies (1.7%; 95% CI, 0.2-5.9). To our knowledge, infection of cats with viral strains belonging to the species Betacoronavirus-1 suggested by our data has not been reported. The only coronavirus comprised in the viral species Betacoronavirus-1 reported in carnivores so far is Canine Respiratory coronavirus (CrCoV) in dogs (1). Whether and which viruses comprised in the Betacoronavirus-1 species may infect cats thus requires further investigation. Interestingly, the two cat sera that tested positive for BCoV presented inhibition of 16.7% and 53.6% in the sVNT, including the pre-pandemic serum that would have been considered positive upon using criteria established for testing humans (Figure 1B). Our data thus suggest that the sporadic presence of neutralizing antibodies against members of the species Betacoronavirus-1 may elicit cross-reactive antibodies interfering with serological detection of antibodies against the antigenically related betacoronavirus SARS-CoV-2. Our interpretation is supported by a previous study reporting cross-reactivity of immune responses against the endemic HCoV-OC43 and SARS-CoV-2 in humans interfering with antibody detection, particularly when using spike-based serological assays (18).
In contrast, FCoV-specific neutralizing antibodies were detected in 33.3% of cats (40/120, 95% CI, 24.9-42.5) (Figure 3A). The FCoV seroprevalence found here is comparable with data reported previously from Brazil, whereas seroprevalence rates in cats ranged from 13.2 to 56.2% in different age groups (29, 30). The disparities of seroprevalence by age are because over 70% of FCoV infections occur in animals with less than 1 year of age and FCoV antibody titers decrease with time (31, 32). Cats remain seropositive for prolonged time spans following SARS-CoV-2 and FCoV infection (33, 34), however around 12% of FCoV-infected animals are lifelong carriers and antibody responses among those animals can differ from animals who have cleared FCoV infection (32, 34). Therefore, comparisons between FCoV and SARS-CoV-2 titers over time should be interpreted with caution. Notably, our results are unlikely to be related with vaccination of cats, since the single existing FCoV vaccine is not available in Brazil (32). Moreover, the median PRNT90 endpoint titer among seropositive sera was about 3.5x higher for SARS-CoV-2 than for FCoV (Figure 3A, p=0.0001), supporting a relatively more recent infection of cats with SARS-CoV-2 in contrast to prior infection with the enzootic FCoV (Figure 3A). Neither the SARS-CoV-2 seroprevalence (chi-square, p=0.8863), nor the magnitude of SARS-CoV-2 endpoint titers (Mann-Whitney test, p=0.6492) differed between FCoV-positive and FCoV-negative sera (Figures 3B, C), suggesting that pre-existing immunity against enzootic carnivore coronaviruses did not affect subsequent SARS-CoV-2 infection.
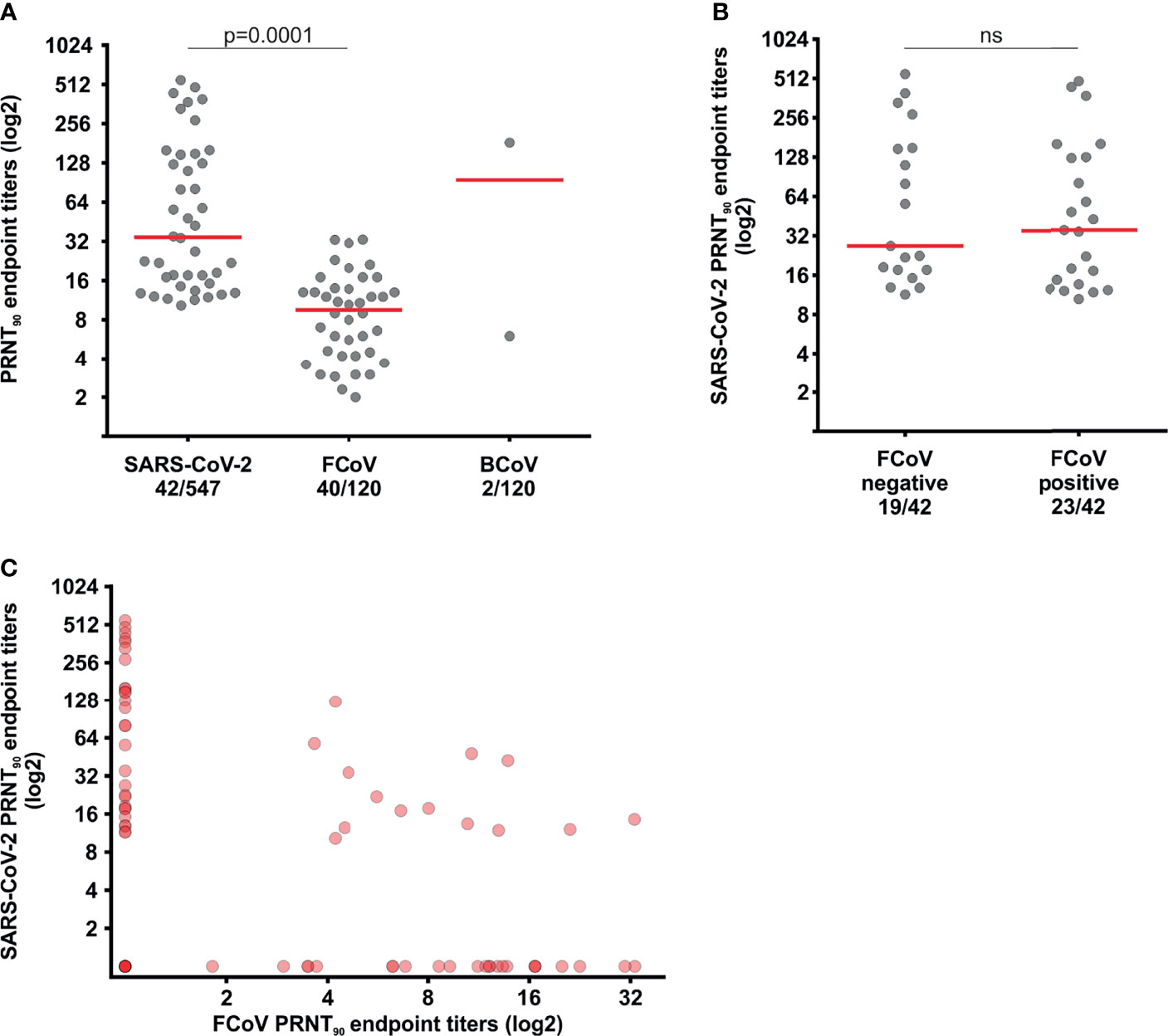
Figure 3 Neutralizing antibodies against SARS-CoV-2 and enzootic coronaviruses. (A) Comparison of the reciprocal FCoV, BCoV and SARS-CoV-2 PRNT90 endpoint titers in cat sera. (B) Comparison of SARS-CoV-2 and FCoV PRNT90 endpoint titers. (C) Reciprocal SARS-CoV-2 PRNT90 endpoint titers among FCoV-positive and -negative sera. Statistical significance was inferred using the Mann-Whitney test in (A) and (B). The red bars indicate medians; ns, not statistically significant.
We revealed geographically widespread and frequent infection of cats from Brazil with SARS-CoV-2. The few available studies on SARS-CoV-2 epidemiology in cats globally show considerable discrepancy in seroprevalence rates (9, 35–39). Those discrepancies are likely related to different antibody detection techniques and sampling strategies. The highest seroprevalence rates were described upon testing cats from COVID-19 positive households, e.g., 14.8% of cats in Wuhan, China (9) and, 58.8% of cats in France (39). Regardless, the seroprevalence in Brazil was higher than those reported from domestic cats in Europe during early 2020, namely 0.7 to 1.4% in Germany, 0.8% in Croatia and 3.3% in the UK (5, 35–38) and 0.8% found in cat shelters in the Netherlands (40), likely consistent with less intense SARS-CoV-2 circulation in humans during those studies. On the other hand, our data is consistent with seroprevalence rates of 4.2 to 5.8% in Italy (5, 38), 4.2% Germany and 6.4% in Spain (38), likely due to relatively higher SARS-CoV-2 incidence during late 2020 and in those countries. Altogether, those comparisons substantiate the robustness of our data showing a relatively high seroprevalence in Brazilian cats likely facilitated by intense SARS-CoV-2 transmission in Brazil during the study period.
The immune interplay between different coronaviruses in different hosts is poorly understood. In humans, immunity against common cold coronaviruses has been reported to afford cross-protection against subsequent SARS-CoV-2 infection (41). In contrast, some studies have reported a tentative association between prior immunity against common cold coronaviruses and COVID-19 severity, hypothetically either via enhancement or via insufficient mounting of SARS-CoV-2 specific immune responses (1, 18, 25, 42). We found no evidence that pre-existing immunity against enzootic carnivore coronaviruses protects against or enhances SARS-CoV-2 infection. Our interpretation is supported by previous findings in humans showing the absence of neutralizing activity against SARS-CoV-2 among individuals recently infected with endemic human coronaviruses (43). It is feasible to hypothesize that cross-protection between FCoV and SARS-CoV-2 is unlikely, however, since ADE may be related to antibody concentrations (20, 44) and very few clinically symptomatic SARS-CoV-2 infections have been so far reported in cats (4, 8), it remains unclear whether ADE pottentially impacts the clinical outcome of SARS-CoV-2 infection in cats. Extrapolating from other viruses known for potential ADE such as flaviviruses, it seems likely that the chronology and sequence of prior infections, in addition to inter-individual variation of immune responses may affect the coronaviral immune interplay (45).
Our study was limited by our sampling of animals presenting clinical disease. It is possible that common cat pathogens that were not tested for here are more frequent in our samples than in clinically healthy cats. For instance, those pathogens could have elicited unspecific antibody responses due to acute infections potentially causing polyclonal B-cell stimulation such as observed in humans (46).
In summary, our results substantiate high SARS-CoV-2 seroprevalence in cats from Brazil already in late 2020. It seems plausible that seroprevalence has increased since the time of our study. Genetic characterizations of SARS-CoV-2 strains from cats and humans in a One Health framework to monitor the emergence of potential mutations of concern may allow early detection of cat-associated SARS-CoV-2 variants, and potential preventive control measures such as vaccination of animals (47).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by the Federal University of Bahia’s animal ethics committee under authorization no. 74/2019. Written informed consent for participation was not obtained from the owners because it was not required by the Ethical Committee because the sera used in this study were leftovers of samples sent for medical diagnostics.
EO-F, OC, IC, FF, SN, CP, LG-A, VC, AK, RM, WJ, AM-S, CR, CD, KO, CB, EN, LR, RM, FF, NR, CF, and JD conceived, planned and performed the experiments. EO-F and AK carried out the BSL-3 experiments. EO-F, OC, FF, VU, RM, IC, AM-S, and JD planned and carried out the analyses and contributed to the interpretation of the results. EO-F and JD took the lead in writing the manuscript. All authors provided critical feedback, helped shape the research, analysis, and manuscript. All authors contributed to the article and approved the submitted version.
Authors OC and LR were employed by company Tecsa Laboratorios.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Dr. Matthias König from the Institute of Virology at the Justus-Liebig University Giessen for providing the CRFK cells and Friedrich-Loeffler-Institute, Bundesforschungsinstitut für Tiergesundheit, Insel Riems for providing the PT (CCLV-RIE 0011) cells.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.857322/full#supplementary-material
1. Jo WK, de Oliveira-Filho EF, Rasche A, Greenwood AD, Osterrieder K, Drexler JF. Potential Zoonotic Sources of SARS-CoV-2 Infections. Transbound Emerg Dis (2020). doi: 10.1111/tbed.13872
2. McAloose D, Laverack M, Wang L, Killian ML, Caserta LC, Yuan F, et al. From People to Panthera: Natural SARS-CoV-2 Infection in Tigers and Lions at the Bronx Zoo. mBio (2020) 11. doi: 10.1128/mBio.02220-20
3. Decaro N, Balboni A, Bertolotti L, Martino PA, Mazzei M, Mira F, et al. SARS-CoV-2 Infection in Dogs and Cats: Facts and Speculations. Front Vet Sci (2021) 8:619207. doi: 10.3389/fvets.2021.619207
4. Garigliany M, Van Laere AS, Clercx C, Giet D, Escriou N, Huon C, et al. SARS-CoV-2 Natural Transmission From Human to Cat, Belgium, March 2020. Emerg Infect Dis (2020) 26:3069–71. doi: 10.3201/eid2612.202223
5. Patterson EI, Elia G, Grassi A, Giordano A, Desario C, Medardo M, et al. Evidence of Exposure to SARS-CoV-2 in Cats and Dogs From Households in Italy. Nat Commun (2020) 11:6231. doi: 10.1038/s41467-020-20097-0
6. Decaro N, Vaccari G, Lorusso A, Lorusso E, De Sabato L, Patterson EI, et al. Possible Human-To-Dog Transmission of SARS-CoV-2, Italy, 2020. Emerg Infect Dis (2021) 27:1981–4. doi: 10.3201/eid2707.204959
7. Romito G, Bertaglia T, Bertaglia L, Decaro N, Uva A, Rugna G, et al. Myocardial Injury Complicated by Systolic Dysfunction in a COVID-19-Positive Dog. Anim (Basel) (2021) 11. doi: 10.3390/ani11123506
8. Newman A, Smith D, Ghai RR, Wallace RM, Torchetti MK, Loiacono C, et al. First Reported Cases of SARS-CoV-2 Infection in Companion Animals - New York, March-April 2020. MMWR Morb Mortal Wkly Rep (2020) 69:710–3. doi: 10.15585/mmwr.mm6923e3
9. Zhang Q, Zhang H, Gao J, Huang K, Yang Y, Hui X, et al. A Serological Survey of SARS-CoV-2 in Cat in Wuhan. Emerg Microbes Infect (2020) 9:2013–9. doi: 10.1080/22221751.2020.1817796
10. van Aart AE, Velkers FC, Fischer EAJ, Broens EM, Egberink H, Zhao S, et al. SARS-CoV-2 Infection in Cats and Dogs in Infected Mink Farms. Transbound Emerg Dis (2021). doi: 10.22541/au.161821264.49927405/v1
11. Hoffmann M, Zhang L, Kruger N, Graichen L, Kleine-Weber H, Hofmann-Winkler H, et al. SARS-CoV-2 Mutations Acquired in Mink Reduce Antibody-Mediated Neutralization. Cell Rep (2021) 35:109017. doi: 10.1016/j.celrep.2021.109017
12. Larsen HD, Fonager J, Lomholt FK, Dalby T, Benedetti G, Kristensen B, et al. Preliminary Report of an Outbreak of SARS-CoV-2 in Mink and Mink Farmers Associated With Community Spread, Denmark, June to November 2020. Euro Surveill (2021) 26. doi: 10.2807/1560-7917.ES.2021.26.5.210009
13. Oude Munnink BB, Sikkema RS, Nieuwenhuijse DF, Molenaar RJ, Munger E, Molenkamp R, et al. Transmission of SARS-CoV-2 on Mink Farms Between Humans and Mink and Back to Humans. Science (2021) 371:172–7. doi: 10.1126/science.abe5901
14. Faria NR, Mellan TA, Whittaker C, Claro IM, Candido DDS, Mishra S, et al. Genomics and Epidemiology of the P.1 SARS-CoV-2 Lineage in Manaus, Brazil. Science (2021) 372:815–21. doi: 10.1126/science.abh2644
15. Worldometer. Coronavirus Updates. (2021). Available online at: https://www.worldometers.info/coronavirus/ (accessed December 30, 2021).
16. Calvet GA, Pereira SA, Ogrzewalska M, Pauvolid-Correa A, Resende PC, Tassinari WS, et al. Investigation of SARS-CoV-2 Infection in Dogs and Cats of Humans Diagnosed With COVID-19 in Rio De Janeiro, Brazil. PloS One (2021) 16:e0250853. doi: 10.1371/journal.pone.0250853
17. Monto AS, DeJonge PM, Callear AP, Bazzi LA, Capriola SB, Malosh RE, et al. Coronavirus Occurrence and Transmission Over 8 Years in the HIVE Cohort of Households in Michigan. J Infect Dis (2020) 222:9–16. doi: 10.1093/infdis/jiaa161
18. Guo L, Wang Y, Kang L, Hu Y, Wang L, Zhong J, et al. Cross-Reactive Antibody Against Human Coronavirus OC43 Spike Protein Correlates With Disease Severity in COVID-19 Patients: A Retrospective Study. Emerg Microbes Infect (2021) 10:664–76. doi: 10.1080/22221751.2021.1905488
19. Fischer C, de Oliveira-Filho EF, Drexler JF. Viral Emergence and Immune Interplay in Flavivirus Vaccines. Lancet Infect Dis (2020) 20:15–7. doi: 10.1016/S1473-3099(19)30697-8
20. Marques ETA, Drexler JF. Complex Scenario of Homotypic and Heterotypic Zika Virus Immune Enhancement. mBio (2019) 10:e01849-19. doi: 10.1128/mBio.01849-19
21. Takano T, Kawakami C, Yamada S, Satoh R, Hohdatsu T. Antibody-Dependent Enhancement Occurs Upon Re-Infection With the Identical Serotype Virus in Feline Infectious Peritonitis Virus Infection. J Vet Med Sci (2008) 70:1315–21. doi: 10.1292/jvms.70.1315
22. Huisman W, Martina BE, Rimmelzwaan GF, Gruters RA, Osterhaus AD. Vaccine-Induced Enhancement of Viral Infections. Vaccine (2009) 27:505–12. doi: 10.1016/j.vaccine.2008.10.087
23. Vennema H, de Groot RJ, Harbour DA, Dalderup M, Gruffydd-Jones T, Horzinek MC, et al. Early Death After Feline Infectious Peritonitis Virus Challenge Due to Recombinant Vaccinia Virus Immunization. J Virol (1990) 64:1407–9. doi: 10.1128/jvi.64.3.1407-1409.1990
24. Crowley AR, Natarajan H, Hederman AP, Bobak CA, Weiner JA, Wieland-Alter W, et al. Boosting of Cross-Reactive Antibodies to Endemic Coronaviruses by SARS-CoV-2 Infection But Not Vaccination With Stabilized Spike. medRxiv (2021). doi: 10.1101/2021.10.27.21265574
25. Sagar M, Reifler K, Rossi M, Miller NS, Sinha P, White LF, et al. Recent Endemic Coronavirus Infection Is Associated With Less-Severe COVID-19. J Clin Invest (2021) 131. doi: 10.1172/JCI143380
26. Yadouleton A, Sander AL, Moreira-Soto A, Tchibozo C, Hounkanrin G, Badou Y, et al. Limited Specificity of Serologic Tests for SARS-CoV-2 Antibody Detection, Benin. Emerg Infect Dis (2021) 27. doi: 10.3201/eid2701.203281
27. Nath H, Mallick A, Roy S, Sukla S, Basu K, De A, et al. Archived Dengue Serum Samples Produced False-Positive Results in SARS-CoV-2 Lateral Flow-Based Rapid Antibody Tests. J Med Microbiol (2021) 70. doi: 10.1099/jmm.0.001369
28. Tan CW, Chia WN, Qin X, Liu P, Chen MI, Tiu C, et al. A SARS-CoV-2 Surrogate Virus Neutralization Test Based on Antibody-Mediated Blockage of ACE2-Spike Protein-Protein Interaction. Nat Biotechnol (2020) 38:1073–8. doi: 10.1038/s41587-020-0631-z
29. Johann J, Caetano C, Hass R, Guim T, Fischer G, Vargas G, et al. Serum Survey for Antibodies to Coronavirus, Herpesvirus, Calicivirus, and Parvovirus in Domestics Cats From Rio Grande do Sul, Brazil. Arquivo Brasileiro Medicina Veterinária e Zootecnia (2009) 61:752. doi: 10.1590/S0102-09352009000300033
30. Almeida A, Galdino M, JP A. Seroepidemiological Study of Feline Coronavirus (FCoV) Infection in Domiciled Cats From Botucatu, São Paulo, Brazil. Pesquisa Veterinária Bras (2019) 39:5. doi: 10.1590/1678-5150-PVB-5706
31. Taharaguchi S, Soma T, Hara M. Prevalence of Feline Coronavirus Antibodies in Japanese Domestic Cats During the Past Decade. J Vet Med Sci (2012) 74:1355–8. doi: 10.1292/jvms.11-0577
32. Addie D, Belak S, Boucraut-Baralon C, Egberink H, Frymus T, Gruffydd-Jones T, et al. Feline Infectious Peritonitis. ABCD Guidelines on Prevention and Management. J Feline Med Surg (2009) 11:594–604. doi: 10.1016/j.jfms.2009.05.008
33. Decaro N, Grassi A, Lorusso E, Patterson EI, Lorusso A, Desario C, et al. Long-Term Persistence of Neutralizing SARS-CoV-2 Antibodies in Pets. Transbound Emerg Dis (2021). doi: 10.1111/tbed.14308
34. Addie DD, Jarrett O. Use of a Reverse-Transcriptase Polymerase Chain Reaction for Monitoring the Shedding of Feline Coronavirus by Healthy Cats. Vet Rec (2001) 148:649–53. doi: 10.1136/vr.148.21.649
35. Stevanovic V, Vilibic-Cavlek T, Tabain I, Benvin I, Kovac S, Hruskar Z, et al. Seroprevalence of SARS-CoV-2 Infection Among Pet Animals in Croatia and Potential Public Health Impact. Transbound Emerg Dis (2021) 68:1767–73. doi: 10.1111/tbed.13924
36. Michelitsch A, Hoffmann D, Wernike K, Beer M. Occurrence of Antibodies Against SARS-CoV-2 in the Domestic Cat Population of Germany. Vaccines (Basel) (2020) 8:772. doi: 10.3390/vaccines8040772
37. Michelitsch A, Schon J, Hoffmann D, Beer M, Wernike K. The Second Wave of SARS-CoV-2 Circulation-Antibody Detection in the Domestic Cat Population in Germany. Viruses (2021) 13:1009. doi: 10.3390/v13061009
38. Schulz C, Martina B, Mirolo M, Muller E, Klein R, Volk H, et al. SARS-CoV-2-Specific Antibodies in Domestic Cats During First COVID-19 Wave, Europe. Emerg Infect Dis (2021) 27:3115–8. doi: 10.3201/eid2712.211252
39. Fritz M, Rosolen B, Krafft E, Becquart P, Elguero E, Vratskikh O, et al. High Prevalence of SARS-CoV-2 Antibodies in Pets From COVID-19+ Households. One Health (2021) 11:100192. doi: 10.1016/j.onehlt.2020.100192
40. van der Leij WJR, Broens EM, Hesselink JW, Schuurman N, Vernooij JCM, Egberink HF. Serological Screening for Antibodies Against SARS-CoV-2 in Dutch Shelter Cats. Viruses (2021) 13:1634. doi: 10.3390/v13081634
41. Song G, He WT, Callaghan S, Anzanello F, Huang D, Ricketts J, et al. Cross-Reactive Serum and Memory B-Cell Responses to Spike Protein in SARS-CoV-2 and Endemic Coronavirus Infection. Nat Commun (2021) 12:2938. doi: 10.1038/s41467-021-23074-3
42. Ng KW, Faulkner N, Cornish GH, Rosa A, Harvey R, Hussain S, et al. Preexisting and De Novo Humoral Immunity to SARS-CoV-2 in Humans. Science (2020) 370:1339–43. doi: 10.1126/science.abe1107
43. Poston D, Weisblum Y, Wise H, Templeton K, Jenks S, Hatziioannou T, et al. Absence of Severe Acute Respiratory Syndrome Coronavirus 2 Neutralizing Activity in Prepandemic Sera From Individuals With Recent Seasonal Coronavirus Infection. Clin Infect Dis (2021) 73:e1208–11. doi: 10.1093/cid/ciaa1803
44. Katzelnick LC, Gresh L, Halloran ME, Mercado JC, Kuan G, Gordon A, et al. Antibody-Dependent Enhancement of Severe Dengue Disease in Humans. Science (2017) 358:929–32. doi: 10.1126/science.aan6836
45. Pedroso C, Fischer C, Feldmann M, Sarno M, Luz E, Moreira-Soto A, et al. Cross-Protection of Dengue Virus Infection Against Congenital Zika Syndrome, Northeastern Brazil. Emerg Infect Dis (2019) 25:1485–93. doi: 10.3201/eid2508.190113
46. Fischer C, Jo WK, Haage V, Moreira-Soto A, de Oliveira Filho EF, Drexler JF. Challenges Towards Serologic Diagnostics of Emerging Arboviruses. Clin Microbiol Infect (2021) 27:1221–9. doi: 10.1016/j.cmi.2021.05.047
Keywords: SARS-CoV-2, cats (felis catus), serology, zoonosis, cross-reactivity, coronavirus
Citation: Oliveira-Filho EF, de Carvalho OV, Carneiro IO, Fernandes FD’a, Vaz SN, Pedroso C, Gonzalez-Auza L, Urbieta VC, Kühne A, Mayoral R, Jo WK, Moreira-Soto A, Reusken CBEM, Drosten C, Brites C, Osterrieder K, Netto EM, Ristow LE, Maia RdC, Vogel FSF, Almeida NRd, Franke CR and Drexler JF (2022) Frequent Infection of Cats With SARS-CoV-2 Irrespective of Pre-Existing Enzootic Coronavirus Immunity, Brazil 2020. Front. Immunol. 13:857322. doi: 10.3389/fimmu.2022.857322
Received: 18 January 2022; Accepted: 21 February 2022;
Published: 05 April 2022.
Edited by:
Yuejin Liang, University of Texas Medical Branch at Galveston, United StatesReviewed by:
Jianping Ma, National Institutes of Health (NIH), United StatesCopyright © 2022 Oliveira-Filho, de Carvalho, Carneiro, Fernandes, Vaz, Pedroso, Gonzalez-Auza, Urbieta, Kühne, Mayoral, Jo, Moreira-Soto, Reusken, Drosten, Brites, Osterrieder, Netto, Ristow, Maia, Vogel, Almeida, Franke and Drexler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Felix Drexler, ZmVsaXguZHJleGxlckBjaGFyaXRlLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.