 Gatien A. G. Lokossou
Gatien A. G. Lokossou Léonce Kouakanou
Léonce Kouakanou Anne Schumacher
Anne Schumacher Ana C. Zenclussen
Ana C. Zenclussen
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 05 April 2022
Sec. Nutritional Immunology
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.849012
This article is part of the Research TopicBreast Milk: From Nutrition to Immunological ActionView all 9 articles
Breastfeeding is associated with long-term wellbeing including low risks of infectious diseases and non-communicable diseases such as asthma, cancer, autoimmune diseases and obesity during childhood. In recent years, important advances have been made in understanding the human breast milk (HBM) composition. Breast milk components such as, non-immune and immune cells and bioactive molecules, namely, cytokines/chemokines, lipids, hormones, and enzymes reportedly play many roles in breastfed newborns and in mothers, by diseases protection and shaping the immune system of the newborn. Bioactive components in HBM are also involved in tolerance and appropriate inflammatory response of breastfed infants if necessary. This review summarizes the current literature on the relationship between mother and her infant through breast milk with regard to disease protection. We will shed some light on the mechanisms underlying the roles of breast milk components in the maintenance of health of both child and mother.
Breastfeeding is considered as an optimal way to provide the developmental nutrients needs to newborns and shaping their immune system. Indeed, the immune system of an infant matures by learning to fight infections, establishing adequate tolerance to the environment and the developing microbiota. Breastfeeding is characterized by important psychological consequences on neonates and mothers (1–4).
Breastfeeding is the most optimal food for infants, and in this regard, the World Health Organization (WHO) and the United Nation International Children’s Emergency Fund (UNICEF) recommend exclusive breastfeeding of babies at least during the first six months after birth and add complementary foods in addition to breastfeeding for up to 2 years or beyond (2). In addition to provide nutrients, breast milk is rich in microbiota and non-immune and also immune components to ensure the infant protection against numerous diseases and support maturation of the developing immune system of an infant (5, 6). The roles of the different breast milk components are far from being completely understood. Of note, immune cells such as B and T lymphocytes, regulatory cells, monocytes/macrophages, neutrophils, natural killer cells and IgA, IgG and IgM antibodies are found in the breast milk (7). Breast milk-derived antibodies are produced as a result of pathogenic or non-pathogenic stimulation of mucosa-associated lymphoid tissues in the gut and airways (8). Therefore, breast milk-derived immune components are related to a wide range of intestinal and respiratory pathogens which the infant will encounter (8–10).
Studies in human have shown that many factors such as geography, infant sex or number of pregnancies influence the heterogeneity and composition of human breast milk (HBM) (11). These parameters shape the composition of HBM and therefore impact the maturation and the susceptibility of the human immune system. Indeed, Holmlund et al. have shown that the maternal country of birth influences the pro- and anti-inflammatory contents of HBM, resulting in susceptibility or not to immune-mediated diseases such as allergy or necrotizing enterocolitis (11).
However, breast milk may also transfer virus such as Epstein–Barr virus (EBV) and cytomegalovirus (CMV) from infected mothers to the infants (12), without infection in infants (13–15). Furthermore, as hepatitis B or C virus are lowly presented in breast milk, breastfeeding is not a contraindication by infected mothers (14). Influenza, rotavirus and HIV virus transmitted by breast milk from infected mothers could lead to infant increased disease susceptibility (12, 14, 16). Moreover, environmental chemicals are sometime also found in HBM leading to toxicity (17, 18). All of these environmental factors need to be assessed for a positive impact of HBM on dyad mother/infant.
Furthermore, maternal microbiota, especially breast-derived microbiota (BDM), is one of the relevant features in the development of the immune system of the infant and his microbiota colonization, affecting mucosal and systemic immunity (10). Bacteria strains are shared in the dyad mother/infant (19, 20). BDM is involved in shaping of the microbiota of offspring, resulting in important differences between breast-fed infant microbiota and those of formula-fed infant (21, 22). BDM include mostly Staphylococcus and Streptococcus bacteria (23), but other bacteria such as Bifidobacterium, Lactobacillus, and Acinetobacter are commonly detected (24, 25). Many factors, namely, geographic locations, maternal lifestyle, the delivery mode and contact to microorganisms influence BDM composition, with variable consequences on the infant immune system (26, 27).
In this review, we summarize the current literature on the breast milk with focus on both non-immune and immune compositions of the breast milk. We also shed some light on their roles in the maintenance of the health of both child and mother.
Breast milk can be subdivided into colostrum, transitional milk and mature milk that reflect the time of breastfeeding from the delivery and the needs for the newborn growth (28). Indeed, colostrum is produced from delivery until seven days while transitional milk is produced between days seven and fifteen after delivery. The mature milk production begins four weeks after postpartum (29). Colostrum is the most important type of HBM based on it high concentration of immune components (7). Overall, the different types of HBM is composed of factors, namely, breast-derived cells, blood-derived cells (30), antibodies, vitamins, and many other cells-derived factors (extracellular vesicles, nucleic acids, enzymes, polysaccharides, lipids, and hormones). All this is reinforced with the presence of probiotic bacterium (10, 31–35). Immune component levels decrease over time and become stable when breast milk is mature (29). Breast-derived cells included lactocytes and myoepithelial cells and also progenitor and stem cells (32). A sustainable proportion of breast milk stem cells is capable of crossing the intestine of nursed infants, entering their circulatory system, and populating distant organs (32, 36, 37). Molès and colleagues have proposed a specific mechanism termed maternal microchimerism (MMC) allowing the transfer and persistent nesting of maternal cells to infant intestinal mucosal tissue, and from there, to other infant immune tissues (38). Milk cells crossing through intestinal mucosal tissue is allowed by weakening interepithelial junctions (39). An important number of breast milk-derived cells are transferred to infant before gut closure and lead to more efficient and effective maturation of mucosal immunity and/or systemic immunity (38).
These natural human milk bioactive components enhanced immunity in the infant during the first years of life (40). High proportion of breast-derived immune cells is activated showing the transfer of active immunity to the infant. These active cells supports the immunological maturation and immune defense of newborns against pathogens (41–43). While it is expected that other components will be identified in the future, the overall picture indicates that HBM may have considerable impact on the dyad mother/infant health.
Multicolor flow cytometry has been used to identify in breast milk two major groups of cells, the breast-derived cells, and the blood-derived cells. Both of these groups contain heterogeneous progenitor stem cells, namely, early-stage stem cells, hematopoietic stem cells, mammary stem cells, mesenchymal stem cells, neuro-progenitor cells, and myoepithelial progenitor cells (44–52). Healthy infants consume between 0.5 and 1.5 L of breast milk per day. Immune cells constitute less than 2% of mature human milk-derived cells in healthy mothers. Therefore stem cells ingested daily will represent an important number of breast and blood-derived cells and may reach billions of cells (46, 48, 53). Indeed, in a recent publication, the higher number of hematopoietic stem cells in mature breast milk compared to colostrum was described (53). Moreover, Kakulas et al., formerly Hassiotou, using luminous mice have shown that viable multifunctional stem cells are in breast milk and can cross gastrointestinal (GI) tract mucosa and enter the systemic circulation of the breastfed neonates (48). After crossing the GI tract through diapedesis and entering the bloodstream, these cells immigrate into the different organs and differentiate into functional cells (Table 1) (32). Therefore, specific breast-derived stem cells are found in the thymus, liver, pancreas, brain, bone marrow, spleen, intestinal tissues, and lungs and remain unharmed after (32, 48, 54–57). For instance, the incorporation of breast-derived stem cells in the brain is followed by differentiation into the neuron and glial cells (57, 58).
Using gene expression in stem cells (47, 59), Li et al. have recently characterized stem cells in preterm and term mother’s milk (53). They have shown that gestational age at delivery but also body mass index (BMI), the mode of delivery and parity influenced the percentages of hematopoietic stem cells in colostrum and transitional milk. These observations confirmed the Briere et al. data in which breast milk samples obtained from feeding mothers of preterm and term offspring are obviously different in the proportion of stem cells (60). Moreover, stem cell numbers changed dynamically at different breastfeeding stages (53, 59). Mesenchymal stem cells (MSC) (CD44+, CD90+, and CD105+ cells) represent 10–15% of stem cells (50) and MSC were shown to be higher in mature milk than in colostrum (59). These changes in colostrum vs mature milk were not confirmed by Shujuan et al. (53), showing the need of more data to better describe the breast milk composition.
It is still a challenge to identify the origin of human breast milk stem cells (hBmSc). Hassiotou and Hartmann have suggested that at least some subpopulations of hBmSCs could have the same origin as hematopoietic stem cells (48). This suggestion is reinforced by the presence of other blood-derived cells in breast milk (51, 61). Unfortunately, the precise mechanisms by which they enter into breast milk and the physiological roles of hematopoietic stem/progenitor cells in early neonate development remain unclear.
The last decades allowed good documentation of the transmission of cells from mother to fetus and vice versa through placenta but also from mother to breastfed infants via breast milk (36, 37, 62–64). Breast milk-derived cells seem to cause microchimerism in breastfed infants and these cells may survive in the infants for several years as those cells exchanged between mother and fetus in utero (37, 65–68). This microchimerism is firmly involved in tissue repair and in the efficient maturation of infant immune system (38, 69), for instance, by inducing tolerance to mother noninherited maternal antigen (NIMA), especially in case of graft from mother to breastfed infants (37). This not fully understood physiological role of hBmSC becomes a great challenge to learn more about all the implications of breastfeeding and harness stem cells to provide significant therapies.
At birth, the newborn immunity is characterized by its remarkable difference compared to the adult immune system (70–72) placing an infant at an increased risk of infection (70, 73, 74). However, breastfeeding is perfectly adapted to provide needs to learn how to establish balance between fighting infections and tolerance to harmless environmental compounds (40). Furthermore, in some cases the immune system of the newborn is associated with developmental immune deficiencies which might render the newborn more susceptible to infections, eventually decreases vaccine efficacy and increases the susceptibility of the respiratory and gastrointestinal (GI) tract to infections (75). Breastfeeding will reinforce and educate neonatal immune trajectories allowing the best protection for the newborn (76, 77). In addition to stem cells (53, 54, 59), HBM is rich in immune cells such as monocytes/macrophages, neutrophils, cytotoxic, helper and regulatory T cells, natural killer (NK) cells and B cells. These cells provide active immunity to neonates by their abilities to produce bioactive molecules such as lactoferrin, lysozyme, oligosaccharides, cytokines and others. B cells complete these bioactive molecules by producing immunoglobulins (secretory IgA, IgG and IgM) (29, 30, 78). Breast milk shapes the development of the newborn immune system and is involved in the development of the microbiota of the infant (10). The proportion of leucocytes is significantly higher in colostrum than in transitional milk and in mature milk. More interesting, although the number of leucocytes/ml decreased significantly from colostrum to mature HBM, the variety of leucocytes and their phenotypic characteristics is associated with minor difference with the breastfeeding time, the length of pregnancy or delivery mode (79–82).
Macrophages are part of the innate immune system that detect pathogenic microbes or infected cells and trigger immune responses by producing inflammatory mediators and activating adaptive immune responses. These cells are professional antigen presenting cells and therefore important players during adaptive immune activation. Breast milk macrophages (BrMM) represent up to 80% of colostrum and transitional HBM, showing their involvement in the immune protection of the infant (83). Recent data have confirmed the abundance of monocytes/macrophages in HBM and are grouped into inflammatory (CD16−) and non-inflammatory (CD16+) macrophages (84–87). Although a consistent number of monocytes/macrophages found in breast tissue are originated from the peripheral blood, the gut, or the nasopharynx-associated lymphoid tissues, a fraction is locally constituted in the mammary tissue (8, 78). The strong presence of macrophages in HBM is due to their high phagocytic role (88) and BrMM were able to produce spontaneously granulocyte-macrophage colony-stimulating factor (GM-CSF) upon exposure to HBM components. In the presence of exogenous interleukin-4 (IL-4) alone, BrMM differentiated into CD1+ DCs showing their abilities to stimulate T cells (89, 90).
Zheng et al. reported that macrophages homing to breast, and are found in milk, in response to ongoing respiratory infections in the nursing infant, displayed an anti-inflammatory profile that correlated with protection of the infant against mucosal inflammation (84). Indeed, these breast macrophages of a mother are trafficking to the mucosal surfaces of the offspring in mice and other animal models (36, 39, 58, 62, 91). Therefore, it is clear that these cells play a crucial role in the gastrointestinal mucosa and are involved in the maturation of the immune system of the nursing infant without causing inflammation (73, 84, 92, 93). These breast-derived macrophages may therefore exert an important role in innate immunity but also triggering high antigen-specific T cells (89). These findings showed the uniqueness functions of BrMM in both normal and pathological conditions, which is not the case with other differentiated cells that are transferred into breast milk. Moreover, BrMM immunoprotective function is improved by opsonin-independent mechanisms than blood monocytes, by differentiating into lectinophagocytic properties (94). However, the mechanism underlying this function needs to be investigated.
Neutrophils are important sensor cells that help to prevent infections by blocking, disabling or digesting invading particles and microorganisms. These cells are key players during acute inflammation and also contribute to chronic inflammatory conditions and adaptive immune responses (95).
Neonatal neutrophils are in lower numbers and they are not able to effectively kill bacteria as neonatal neutrophils do not form neutrophil extracellular traps (NETs). Expression of adhesion molecules (e.g., L-Selectin: CD62L, Integrin: MAC-1) is also decreased on neonatal neutrophils, affecting their binding ability to the endothelium. They are also characterized by a lower expression of TLR2 and TLR4 as well as decreased level of phagocytosis, neutrophilic burst, and diminished capacity of degrading intracellular infective agents (96). HBM neutrophils will make up these deficiencies by being more active and motile, and presenting interactive features showing that HBM neutrophils are involved in active immune response in infants (44, 93, 97, 98). During breastfeeding, the relative median frequencies of neutrophils within total leukocytes significantly increased from colostrum to mature milk (79) showing their involvement in infant immune protection. Furthermore, milk from asthmatic mothers has demonstrated elevated proportions and activation of neutrophils. These changes of HBM neutrophils due to maternal asthma may result in alteration in the immune response of the infant (99). Besides, breastfeeding mothers with high levels of neutrophils in their milk (>20%) are more likely to have children who are allergic to cow’s milk than breastfeeding mothers with low neutrophil levels (100).
On the other hand, this does not question the fact that data from the systematic reviews have shown the breastfeeding protection of infants against the development of atopic diseases such as eczema and food and respiratory allergy, especially when there is a family history (101, 102).
Natural killer cells are important players in the innate immune response. These cells act directly against infection but also produce cytokines that activate others immune cells. Therefore, NK cells contribute to neonate protection (42, 45). Recent data have described NK cells in HBM (30, 44). NK cells represent on average 0.5% of colostrum-derived leucocytes, 1.3% of transitional breast milk cells, and 2% of mature breast milk cells (53). The proportion of NK cells was significantly lower in mature milk of preterm mother (79) and formula-fed newborns (103). Interestingly, Fernandez et al. have shown that human milk-derived Lactobacillus may enhance the release of Th1 cytokines, and activate NK cells, and others immune cells (104).
NK cells migrate to the breast from lymphatic vessels and systemic circulation (105), and arrive in mammary glands (106). These cells do not play a protective role only in the infant gastrointestinal tract but also protect the mammary gland from infection (42).
The involvement of breast milk-derived NK and NKT cells in antibody-dependent cellular cytotoxicity (ADCC) and antibody-dependent cellular phagocytosis (ADCP) should be more investigated to better understand how NK cells contribute to pathogen clearance in infant. For instance activated NK cells are found in HBM during CMV reactivation, supporting the immune response against CMV (15).
One of the most recent discoveries in the composition of breast milk is the presence of innate lymphoid cells (ILCs) that often were found near the lining of the aero-digestive tract (96, 107–109). These cells seem to be more activated and/or differentiated than their blood homologous (108) and constituted guard against breast infection by pathogens. They share functions with T cells (110) and may shape neonatal innate immunity (108) as they are important players in intestinal microbiota and the adaptive immunity of the newborn (111–113). Based on cytokines secretion and transcription factor profiles, ILCs were regrouped within 3 subtypes of ILCs with ILC1 counted 3 to 10 times of ILC3 which was 3 times the count of ILC2. More interesting, ILC1 produce higher levels of IFN-γ than IL-5 and IL-22 produced by ILC2 and ILC3 respectively (108). ILC2 and ILC3 are therefore important player in the GI tract health as IL-22 is highly associated with the maintenance of gut barrier integrity (113). Furthermore, ILCs play an important role during inflammation and tissue homeostasis (108, 112). As breast milk-derived ILCs modulate the immunity of the neonate, maternal ILCs may affect the infant ILCs. But it is not clear how this occurs. The next step will be to investigate the role of maternal ILCs in shaping infant ILCs and understand how; 1—they protect the infant from enteric infections and intestine inflammation, 2—they are harnessed to regulate the establishment of the gut microbiome of the infant.
As key actors of adaptive immunity, lymphocytes represent 5 to 10% of all HBM leucocytes (105). Among these lymphocytic populations, T cells represent more than 80% whereas B cells are only 4 to 6% (114). In HBM, CD4+ T cells are present in activated state and express activation markers such as CD40L, sCD30, IL-2 receptor, HLA-DR+, human mucosa lymphocyte antigen-1 (hMLA-1), or late activation protein-1 (LAP-1). These adaptive immune cells also express CD45RO which is associated with immunological memory functions (115–118). Low number of naïve CD4+ T cells were also described in HBM (117). In HBM, 26–76% of CD4+ T cells co-express the C chemokine receptor 5 (CCR5) and CXC chemokine receptor 4 (CXCR4), the major co-receptors used by HIV to attach and entry cells (119–121). Although the expression of CD103, the mucosal homing marker indicating their mucosal origin, these HIV target cells were preserved in HIV-infected mothers breast milk unlike in other mucosal immune areas in which these cells were quickly eliminated by HIV (122). Moreover, the HIV-infected women were clinically stable on highly-active antiretroviral therapy (HAART) allowing low or undetectable viral loads but do not achieve CD4+ T cell reconstitution in the periphery (122). As HBM contains memory CD45RO+ T cells with helper-inducer function, they may be resistant to killing by HIV (123) and shelter the mammary gland from HIV replication (124–127).
HBM-derived CD8+ T cells are homed from the mother mucosal immune system to the breast and are presented at a higher proportion than CD4+T cells. Indeed, Wirt et al. have shown that the HBM mean CD4/CD8 ratio of T cell was 0.88, ranging from 0.40 to 1.25 whereas it decreases in peripheral blood (116). CD8+ T cells express L-selectin, α4β7 integrin, mucosal addressin cell adhesion molecule-1, intestinal homing receptor CD103, mucosal homing receptor CCR9, high level of CD45RO, and HLA-DR markers showing their effector memory functions (117, 128, 129). Comparable to peripheral blood, HBM CD8+ T cells express IFN-γ alone or IFN-γ and granzyme together and increase during human cytomegalovirus, HIV, influenza, or EBV infection (80, 130). The increased number of CD8+ T cells were positively correlated with breast milk viral load showing that local virus replication induced homing of antigen-specific CD8+ T cells into the breast for local viral control and relative lack of viral transmission via mammary glands (131). As showed in murine pups, maternal breast milk cytotoxic T lymphocytes home to the Peyer’s Patches of the breastfed infant and this was due to the expression of gut-homing molecules α4β7 and CCR9. Moreover, breast milk cytotoxic T cells have a high capacity to produce potent cytolytic and inflammatory mediators when compared to those generated by the breastfed infant (130). These data suggest the compensation of the learning adaptive immune system of the infant in order to limit oral infectious during the postnatal phase (36).
Since the first description of presence of leucocytes in HBM, especially in colostrum (132, 133), subsequent findings have elucidated the HBM composition. It is established that breast milk is involved in initiating immunologic tolerance in newborns to antigens of both mother and environment to achieve lifelong homeostasis and prevent immune-related disorders (128, 134). It was reported that the thymus size of breast-fed infants is double to that of non-breast-fed infants (135–137) and supports the role of human milk on regulatory T cells expansion during the first three weeks after birth in breastfed human babies. These cells are nearly twice as abundant as in formula-fed babies. These cells also control the immune response of the baby against maternal cells transferred with breast milk and help to reduce inflammation (65, 66, 138, 139).
As the increased number and functionality of T regulatory cells (Treg) are crucial for establishing and maintaining the semi-allograft fetus (140–142), these cells are the main players for providing tolerance in the newborn (138, 139). Cérbulo-Vázquez et al. have demonstrated a higher frequency of CD127− CD25++ cells (Treg) in colostrum than peripheral blood, showing a passive transfer of tolerance mediated by memory cells and it is positively associated with vaginal delivery (143).
These tolerogenic cells also exhibited higher expression of CD25, CD152, CD279, and TGF-β than blood-derived Treg cells, suggesting the increased ability of colostrum-derived Treg cells to mediate immunomodulation and maternal microchimerism (37, 143).
Treg cells will play a beneficial role in the newborn by lowing inflammation in the gut mucosa (144) and reducing the risk of developing allergy, asthma, obesity, and diabetes type 1 during the short-and long-term life of the infants (145).
Moreover, it has been shown that maternal allotransplants from the human kidney are better tolerated if the graft recipient has been breastfed during early life (146, 147). This immunosuppressed anti-maternal immunity may be related to the ability of newborn naive CD4+ T cells to differentiate toward tolerogenic features and drive the suppression of normal immune responses leading to T cell anergy (148–150). The breakdown of tolerance to maternal antigens mechanism throughout the life can drive autoimmune diseases such as type 1 diabetes (T1D), multiple sclerosis (MS), and systemic lupus erythematous (SLE), suggesting the necessity of maintaining this maternal microchimerism induced by HBM (151–155). More recently, CD161+/TRAV1‐2+ mucosal‐associated invariant T cells (MAIT) and γδ T cells were found in breast milk. The study by Bedin et al., reported an increased frequency of CD161+/TRAV1‐2+ MAIT cells in HIV-infected mother breast milk compared to healthy mother (107). Furthermore, γδ T cells have selective compartmentalization and represent almost 9% of CD45+ breast milk leucocytes whereas they represent only 2.5% of blood-derived lymphocyte cells. γδT cells number decrease slowly in HIV-infected women breast milk and represent 7.26% of breast milk-derived leucocytes. In blood from HIV-infected cells, 3.31% of CD45+ cells were TCR γδ cells (107). This observation shows the detection of unconventional T cells that may also be involved in both the protection against infection of the lactating mammary gland and the maturation of infants’ gut and microbiota (107). One of the future challenges will be to focus on exploring their precise role in gut and microbiota integrity.
The most important thing we knew about breast milk is the presence of a variety of immunoglobulins. These antibodies are secreted by B lymphocytes or plasma cells (79).
After memory B cells or Ig-secreting cells were generated following B cells activation, they recirculate through peripheral lymph nodes, spleen, tonsils, and mucosal effector sites such as exocrine glands, including the lactating mammary gland (9, 156–159).
Phenotypically and functionally, two main subsets of memory B cells have been described in HBM (156, 160). The most abundant subset, class-switched B memory cells (CD27+IgD−) express surface IgG or IgA molecules, while non-class switched B cells, less abundant (CD27+IgD+) express IgM and are involved in the response to T cell-independent antigens (161, 162).
HBM-derived B cells are activated cells with high levels of CD38 expression, low expression of complement receptors, a feature of plasma cells (163–165). Mammary gland-homed memory B cells and plasma cells arise from other mucosal areas where they have encountered antigens (139–141). These B cell homing-molecule profiles are different from those of homed B cells in nasal-associated lymphoid tissue (NALT) or gut-associated lymphoid tissue (GALT) (166–169). However, HBM-derived memory B cells are α4β7+/−, α4β1+, CD44+,CD62L−, sharing their profile with GALT B cells which also bear α4 and β7 integrin chains and are CD62L− (168, 170–172). Based on homing profiles, these observations showed that the breast is more closely associated with intestinal mucosa than with upper respiratory mucosal sites (28). B cell migration to the mammary glands is mediated by chemokine CCL28 which is the mucosa-associated epithelial chemokine (111, 173, 174). Moreover, during lactation CCL28 is upregulated and is linked to the CCR10 receptor expression on IgA-secretory cells (IgA-SCs) in the mammary gland, allowing IgA-SCs accumulation in the mammary gland, unlike to IgG-SCs or IgM-SCs (173, 174). Since IgA-SCs are accumulated in the mammary gland, >90% of Igs secreted in breast milk are IgA (111, 175). The entero-mammary pathway appears to be another process by which the mother could also provide the protection to offspring (176). Indeed, during suckling, the mouth-derived pathogen of the infant may be retro-transferred via the nipple to breast inducing a mammary immune response (41, 177, 178). After the recognition of the pathogen by the mother immune system, specific antibodies are then produced in milk and fed to the infected infant (42, 111, 179). Cleary, maternal, and infant infections stimulate a rapid leukocyte response in breast milk (41). Moreover, in the milk of HIV-infected mothers and stools of their breastfed children, the levels of total IgA and IgG were increased indicating the activation and homing into the breast of gut mucosa-derived B cells (175, 180). Notably, breast milk samples and stool samples from HIV-negative and HIV-positive babies breastfed by their HIV-infected mothers, contained high levels of HIV-specific IgG. However, IgA antibodies were less frequent, and IgM are scarce (181). These antibodies, that specifically target gp120, purified from HIV-exposed breastfed infants, were able to inhibit HIV binding to HT29 cells and monocyte-derived macrophages (181). Moreover, the intestinal mucosa of infant exposed to HIV by breastfeeding produces HIV-specific antibodies with inhibitory properties to HIV, showing the effect of prophylactic breastfeeding on immune responses of an infant against natural immunization with HIV (181).
It is therefore clear that class-switched B memory cells may highly be involved to compensate the low antigen-presenting capacity of newborn macrophages.
Acquired immunity of a newborn is characterized by its excellent capacities to learn fighting pathogenic microbes and during this learning phase, it is supplemented by maternal antibodies (70–72, 182). These antibodies are provided first in utero and after birth by breast milk. These in utero maternal IgGs are actively transported through the placenta to the fetus via the Brambell receptor commonly called FcγRn (183). However, IgG with certain specificities and with high avidity are more efficiently transferred to the fetus than others (184). These IgG antibodies are involved in newborn protection and mediate antimicrobial and antibody-related cytotoxicity (185). These antibodies recognize microbes on intestinal mucosa but also in the circulation and tissues (43, 186).
Soluble milk IgAs also have antimicrobial properties and are higher in milk from very preterm delivered mothers compare to term milk (187–190) and in preterm milk vs term milk (185). sIgA could inhibit pathogen binding (79, 191) by complexing antigens and taking them up by intestinal DCs which will then process and present these antigens (192). IgM, antimicrobial antibodies, are also found in breast milk and mediate cytotoxicity (185).
Recently, extracellular vesicles (EV) have been identified in breast milk and seem to shape the breastfed infant immune system, especially his intestinal immune response (10, 124, 193, 194). Moreover, Hock et al. have shown that rat milk-derived exosomes promote intestinal epithelial cell viability, enhance proliferation, and stimulate intestinal stem cell activity. These findings support the fact that milk-derived exosomes play important roles in gut health and prevent necrotizing enterocolitis in infants (195).
Breast milk-derived EVs contain a rich cargo, namely, cytosolic and membrane proteins, mRNA, miRNA, and are oriented towards cell-to-cell communication and result in the immune shaping of newborns (196–198). EVs include two subsets such as microvesicles (100–1.000 nm) (199, 200) and exosomes (30–100 nm) (201, 202). Exosomes differ from the other cellular microvesicles according to 1—the endocytic pathway by which they are generated, 2—their homogenous cup-shaped structure, when observed by electron microscopy, and 3—their buoyant density ranges between 1.13 and 1.19 g/ml on a sucrose gradient (203). Although the protein composition of exosomes varies from their originating cell, proteome analyses shown that the tetraspanins, CD9, CD63, CD81, and the endosomal sorting complexes required for transport (ESCRT)-related proteins, Alix and TSG101, are constituents of nearly all exosomes and are markers used for the detection of exosomes (196, 203). The acetylcholine esterase (AChE) activity is also a useful tool to control for exosomes isolation and purification (204). Exosomes, are formed through the endosomal pathway and are released from cells following the fusion of multivesicular bodies with the plasma membrane (205). Exosomes are released by a large range of cells, namely, immune cells, neural cells, stem cells, placenta cells, and many cancer cells (reviewed here (205)), and can be isolated from different body fluids, such as serum, urine, cerebrospinal fluid, amniotic fluid and breast milk (10, 195, 203, 205–207). There are enriched by immunologically relevant components that direct immune responses (208). High concentration of exosomes is found in colostrum compared with mature milk. HLA-DR is highly incorporated in early milk-derived exosomes which displayed significantly lower levels of HLA-A, -B, and -C in relation to mature milk. Indeed, breast milk-derived exosomes phenotype depends on maternal sensitization and lifestyle, which might increase allergy sensibility in the offspring (209). Comparatively to breast milk volume, a high level of micro RNAs (miRNAs) is found in breast milk (196). These large various non-coding molecules (210) are stable in an acidic conditions of GI tract (211) and regulate post-transcriptional expression of genes and have biological activities in humans (210, 212, 213). According to the HBM immune cells miRNAs and mammary gland cells miRNAs, HBM-derived miRNAs seem to originate from immune-related and mammary gland cells (193). HBM-derive exosomes contain mostly miR-148a-3p, miR-22-3p, miR-200a-3p, miR-146b-5p, miR-30d-5p, let-7a-5p, miR-30a-5p, let-7f-5p, let-7b-5p, and miR-21-5p (193, 214–221). Upon in the GI tract, these microvesicles-derived microRNAs are taken up by intestinal cells (222). Moreover, some of these miRNAs seem to shape the neonate immune system (210, 214) by limiting cytokine production (223) and inhibiting T-cell proliferation induced by DCs (223) or B cell activation, TLR4 signaling, and macrophage activation (193, 224).
Since gestational age induces changes in the miRNA profile which influence metabolism and lipid biosynthesis (225), it becomes opportune to understand the impact of these changes on breast milk composition and offspring health (193). In the same way, other HBM exosome-derived miRNAs induced B- and T-cell activation, proliferation, and differentiation (193). They are able to protect newborn from infection by reducing inflammation and boosting the immune response by many ways (193, 222, 226).
Carr et al. have recently proposed that breast milk miRNAs may act as TLR7/TLR8 agonists and may boost the immune system (193). This opens the way for a better understanding of the specific role of HBM exosomes-derived miRNAs on the immune system and the neonatal microbiota (10, 193, 227).
HBM contains an array of cytokines and chemokines reviewed by Nolan et al. (228). These cytokines are primarily originated from the mammary glands, and include IL-1 β (229, 230), IL-2 (229), IL-4 (230), IL-5 (230), IL-6 (84, 229–231), IL-8 (232), IL-10 (189, 230), IL-12 (233), IL-13 (230, 233), TNFα (229, 231), TGF (transforming growth factor)-β (232), IFN-γ (235), granulocyte-colony stimulating factor (G-CSF) (234), monocytes chemotactic protein 1 (MCP-1) (115) and Regulated upon Activation, Normal T Cell Expressed and Presumably Secreted (RANTES) (93, 230). Purified immune cells from HBM have been shown to be able of producing several cytokines (231). For instance, ongoing infection in breastfeeding infant change milk-derived macrophage profiles into a more anti-inflammatory profile compared to the healthy breastfeeding infant (84). It will be very worthwhile to generate more data to understand how these cytokines stay stable and passage through the infant digestive system. These molecules may be embedded and protected until they reach the intestine (234, 235). The HBM-derived cytokines shape the maturation and development of immune cells in infants. For instance, maternal milk-derived TGF-β, IL-6, and IL-10 induce the development and differentiation of IgA-producing cells (230) and maturation of the naive intestinal immune system (236). Moreover, infant infection is associated to IL-6 and IL-8 increase in HBM that also contains CCL2 and CX3CL1 (84).
Many factors influence the cytokine/chemokine profile of breast milk. For example, geographic location, migration of women to a place different from the place of birth, the number of pregnancies, etc., determine the cytokine/chemokine profile of HBM. Thus, studies have shown that maternal country of birth may influence breast milk composition of cytokines. For instance, immigrant mothers from low income country with high exposure to pathogens, to low exposure developed country, continue to produce higher levels of pro-inflammatory cytokines (IL-6 and IL-8) and TGF-β in their breast milk when they gave birth in developed country (237). In countries with high pathogens exposure, it is relevant to produce higher levels of TGF-β1 and IL-6 in breast milk because of their involvement in stimulation of IgA synthesis (238, 239). Therefore, their offspring will be primed for more inflammatory cells to fight pathogens. However, when these mothers immigrate to developed countries with low exposure to pathogen, their infants continue to be primed with higher levels of IL-8, IL-6 and TGF-β1, resulting to higher incidence of inflammatory disease (237).
The number of pregnancies also influences the levels of cytokines in breast milk. Indeed, a larger number of previous pregnancies are associated to lower levels of IL-6, IL-8 TGF-β and soluble CD14 in breast milk (237), resulting in a global damping of the immune system. A lot of information is still needed in order to better appreciate the role of these cytokines/chemokines.
Immune development during infancy is in part related to bioactive factors provided by HBM. Among these factors, probiotic bacteria are important players and their variation due to geographic locations, maternal lifestyle, the delivery mode and contact to microorganisms can impact positively or negatively on the infant immune system (26, 27). Breast milk microbiota contains a large number of specific bacterial species with antimicrobial properties and health benefits (240–242). These bacteria include beneficial, commensal and probiotic bacteria (243, 244).
In regard to the number of bacteria consumed per day, about 8.105 bacteria/day, breast milk appear to be the second source of bacteria for the infant after normal delivery (21). Human breast milk-derived bacteria spectrum in healthy mothers is highly diverse. The major bacterial species belong to Lactobacillus (L. gasseri, L. fermentum, L. plantarum, L. rhamnosus, and L. salivarius) (25, 241, 245, 246) and Bifidobacterium (B. breve, B. bifidum, B. lungum, B. dentium, and B. adolescentis) (247, 248). Among Staphylococcus spp. presented in HBM, S. epidermidis was predominant while S. aureus was also found (249, 250). Streptococcus spp. included mostly S. salivarius, S. hominis, and S. mitis (249, 250). Recent analysis of HBM using sequencing technologies has provided more detail about the diversity of microbiota of milk with more than hundred bacterial species (10). Interestingly, these bacteria are not present at the same lactation time and their proportion among all bacterial is not identical in all women. It is therefore believed that variations in HBM microbiota are based on maternal diet, genetic, the mode of delivery, demographic or environmental differences (27, 251–253). Indeed, during the first month of lactation, full-term and preterm milk samples shared bacteria such as Bifidobacterium, Lactobacillus, Staphylococcus, Streptococcus, and Enterococcus with significantly lower number of Bifidobacterium spp. in preterm group (254). However, Urbaniak et al. did not found significant changes in the bacterial composition between term and preterm patients (255), showing that more research are needed to highlight the composition of breast milk with regard to the gestational age. According to the mode of delivery, cesarean sections were associated with higher overall bacterial concentration in colostrum and transitional milk. Moreover, higher concentration of Streptococcus spp. and lower concentration of Bifidobacterium spp. were found in cesarean sections than vaginal deliveries (254).
Anti-biotherapy during pregnancy could also influence microbiota diversity, leading to a significant decrease of Bifidobacterium and Lactobacterium spp (256).. Other factors that might impact breast milk microbiota are mother BMI and geographical locations. Indeed, high BMI is associated with lower bacterial diversity with reduction of Bifidobacterium spp. levels and this lower bacterial diversity is correlated to celiac disease in mothers (257). Mothers living in Africa, Asia, and Europe have different milk profiles (253, 258), even if Staphylococcus and Streptococcus were constant compounds of the milk microbiota regardless the geographical locations (253, 258).
Too much debate surrounded the origin of breast milk bacterial population. Similarities were found between the adult skin microbiome and milk microbiota, suggesting bacterial transfer from the skin of the mother during infant suckling (21, 242, 259). In addition, infant oral cavity also influences the breast milk bacterial composition based on retrograde back flow mechanism (179, 260, 261). This mechanism referred to a “retrograde transfer” of external bacteria, and the gut-mammary pathway for translocation of internal bacteria (261). Many data are expected from BDM analysis and the harnessing of BDM will be useful in disease prevention and therapy.
Mortality and risk of infection in breastfed infants vs. nonbreastfed infants is one of the markers of immunological benefits of breastfeeding (262). Indeed, Victora et al. meta-analysis shows that breastfeeding during the first six months of life is strongly protective against infectious diseases (101). Their analysis and others have demonstrated at least 88% reduction of mortality in breastfed compared to nonbreastfed infants (101, 263). Regarding the many advantageous anti-infection and immunological properties described in this review, HBM appear as the optimal food with strong potential to reduce infectious diseases and to optimize the well-being of offsprings, especially in premature neonates. Although, mishandling or misadministration of HBM may result in transmission of infection to neonate, the many immune compounds prevails the potential risk of infection. A recent study showed that mothers are able to significantly increase their breast milk-derived leucocytes in response to acute neonatal infection (41). Some chronic diseases and respiratory infections and also otitis and gastroenteritis rates are shown to be decreased in breastfed infants (264, 265). HBM is also associated with decreased risk of necrotizing enterocolitis and late-onset sepsis in premature neonates (266, 267). When mothers are infected by airbones diseases such as measles, varicella and tuberculosis, infants are recommended to be temporarily separated from their mother. Because these airbone diseases are not transmitted by their milk as long as there are no lesions in the breast area, infants can receive pressed milk from their mothers (268). Chronically infected mothers by hepatitis B virus can produce in their breast milk some virus even if the risk of transmissiom is low (269). Therefore, the benefits of breastfeeding prevails the risk of transmission and it is recommended for mothers to continue breastfeeding (270, 271). In case of Herpes simplex virus (HSV) which is responsible of perinatal infections and, even if less frequently in neonate, HSV infections result from infected maternal breast lesions. As there is no evidence of HSV transmission in HBM, breastfeeding or pumping milk in the absence of breast lesions is recommended when women are infected by HSV (268).
Some pieces of evidence suggest that HBM can provide immune protection against Severe Acute Respiratory Syndrome novel Coronavirus 2 (272, 273). Indeed, significant specific IgA to the full SARS-CoV-2 spike protein and significant IgA, IgG, and/or IgM directed to SARS-CoV-2 spike protein receptor-binding domain are found in HBM (272, 273). As SARS-CoV-2 RNA is expressed in HBM (274) without it being possible to demonstrate the viability and transmissibility of detected virus (275–277), the feeding of HBM should be encouraged, either via direct breastfeeding or via pumped HBM.
Acute gastroenteritis may be cause by norovirus (NV) resulting in diarrhea in infant. It has been shown that high positivity rates and titers of IgA directed to NV in HBM is associated with reduced diarrheal symptoms in NV infected infants (278). These data confirm the antiviral properties of HBM and taking more attention to breask milk composition will help fighting infectious disease in low income countries.
HBM is an antibacterial and anti-infectious food which reduces the possibility of pathogens transmission to neonate (270). These anti-infection properties are confirmed in in vitro studies that showed that HBM-derived Lactobacillus rhamnosus and Lactobacillus crispatus have anti-infection activities against Staphylococcus aureus (S. aureus). As S. aureus is involved in mastitis in lacting women (279), antibiotic-resistant nosocomial infections, and neonatal infections, breastfeeding is and important weapon to reduce the pathogen load owing to S. aureus (270, 280). Moreover, in vitro and in vivo studies in mice have shown that HBM-derived lactobacilli strains, especially, Lactobacillus salivarius, have antibacterial activity and that inhibit Salmonella enterica adhesion to mucins. This results in high survival proportion in infected mice (240, 280). In addition, lactic acid bacteria in HBM allow acid pH, resulting in inhibition of the growth of anaerobic bacteria and protection of the gut (240, 281).
Escherichia coli (E. coli) are responsible of neonatal systemic infection and urinary tract infection. However, HBM is not a source of E. coli due to the anti-infection properties of breask milk (282). Therefore, if a mother is experiencing E. coli infection, breastfeeding migh not be interrupted (270). HBM is also known to prevent oropharyngeal infant Haemophilus influenza type b (Hib) colonization. Therefore, breastfeeding migh be continued with careful masking and hand hygiene when mother is experiencing Hib disease (270, 283).
During delivery, the risk of transmission of Group B Streptococcus (GBS) is significant, resulting in neonatal late-onset sepsis. Although GBS can be found in HBM (284), the anti-infection properties of HBM justify the fact that breastfeeding is encouraged during GBS disease. However, in case of recurrent late-onset GBS disease in preterm neonate, it suggests avoiding breastfeeding until the disease is resolved (285).
Sexually transmitted diseases caused by Chlamydia or Neisseria gonorrhea can result in perinatal infection. Chlamydial acquired infection during delivery results in neonatal conjunctivitis and pneumonitis. While HBM is not a source of chlamydial transmission, specific secretory IgA has been found in colostrum and mature milk of infected women allowing the continuation of breastfeeding (286). N. gonorrhea also is not transmitted by HBM and breastfeeding is recommended if women experiencing infection are treated with appropriate medication (286).
While tuberculosis (TB) remains a burden in developing countries, there has been no demonstrated case of TB transmission through HBM, excepted during TB mastitis (287). Therefore, during separation between TB experienced woman and infant, infants are allowed to receive pumped milk with adequate precautions and are breastfed in case of asymptomatic TB (279).
The benefit of breastfeeding in both mother and infant health is demonstrated by recent data showing that HBM bacteriome are associated with anti-tumor properties (280). Indeed, live, heat-killed and cytoplasmic fractions of HBM-derived Enterococcus faecalis and Staphylococcus hominis have shown anti-tumor activity against a breast cancer cell line in vitro (288).
In vitro study has also demonstrated that Lactococccus lactis has therapeutic effects against colon cancer (289). HBM bacteriome has also relevant implication in mastitis that occurs in about 30% of breastfeeding women (290, 291). HBM-derived Lactobacillus salivarius and Lactobacillus gasseri are shown to be effective substitutes for the treatment and control of mastitis (292).
A collaborative study analyzing 47 epidemiological studies, including women with breast cancer and without the disease, has demonstrated an inverse relationship between breastfeeding and breast cancer. A reduction of 4.9% in the incidence of breast cancer is associated with the duration of breastfeeding (293). This observation was confirmed in a meta-analysis performed by Victora et al. who found that longer breastfeeding is associated with 7% reduction of breast cancer (101). Ovarian cancer is also reduced by 30% when infant was breastfed for a long duration (294).
Potential mechanisms underlining the relationships between breastfeeding and cancers may include inflammatory disorders and epigenetic reprogramming in the tissues, resulting in disorders in progenitor cell pools. For instance, in breast cancer, high FOXA1 methylation is found among women who did not breastfeed (295). Using technological advances, it will be possible to shed light on the mechanism(s) by which breastfeeding contributes to reduce specific cancers.
Compelling data have demonstrated the maternal health outcomes of breastfeeding. Victora et al. have well demonstrated how breastfeeding acts as natural contraceptive by spacing birth, especially when mother exclusive breastfed (101, 296).
Type 2 diabetes (T2D) risk is also reduced by breastfeeding (297, 298), while every 6 months an increase in breastfeeding duration is associated to 1% lower mean body maternal index (299). The mechanism underlining the benefit effect of breastfeeding includes the increase in the number of β cells into the pancreas. Indeed, during breastfeeding prolactin induces serotonin in β cells improving insulin secretory function and glucose tolerance (300). The impact of prolactin on the immune system has been described during the last 2 decades. This polypeptide can act as hormone when synthetized by hypophysis and as cytokine when it is provides by neurons, mammary epithelium, skin, prostate, immune organs, and cells (301). In addition to controlling lactation and maternal behaviors, polymorphisms in prolactin gene appear to play an important role in autoimmune disease including SLES (301, 302). Indeed, hyperprolactinemia impairs B-cell tolerance (303), and results in SLE (303).
T1D characterized by the destruction of pancreatic B cells (304), is strongly based on genetic susceptibility but also involves non-genetic factors (305–307) including breastfeeding which could protect against T1D in infants (308). T1D risk is reduced in newborn breastfed during more than 3 months or during minimum of 2 weeks exclusive breastfeeding after postpartum (309) while T1D risk is twofold increased in nonbreastfed infants (307). However, this study did not found positive association between the time of introduction of specific solid foods and T1D (307). In contract two important studies, the Diabetes and Autoimmunity Study in the Young (DAISY) (310) and the Type 1 Diabetes Prediction and Prevention Project (DIPP) (311) suggested that introduction of solid foods before 4 months and after 6 months of age increase the risk of T1D (303). The risk of T2D in childhood is also reduced by proper breastfeeding including the duration of breastfeeding and exclusive breastfeeding (312).
The protective role of breastfeeding in immune disease, including juvenile idiopathic arthritis (JIA) and rheumatoid arthritis (RA) is controversial (303). For instance, breastfeeding has been associated to protection against JIA (313) whereas other authors did not support this observation (314). It also has been shown that the gut microbiome is altered among JIA patients suggesting a role of breastfeeding (315). Futures studies are needed to better understand the impact of HBM on the risk of JIA.
The influence of HBM on the risk of developing RA is not clear (303) and a recent meta-analysis has concluded that the risk of RA is not affected by the duration of breastfeeding (316).
Most studies on the relationship between breast milk and allergy development in offspring often show contradictory results (317–320). It has been proposed that breastfeeding is associated with anti-allergic properties due to it bacteriome. For instance, HBM probiotic lactobacilli, especially Lactobacillus gasseri together with Lactobacillus coryniformis, decrease the frequency and severity of allergic responses to cow milk proteins in animal studies (321). Moreover, some studies have demonstrated the reduction of infant eczema between one and two years when mother are supplemented with Lactobacillus spp. and/or Bifidobacterium spp (322, 323). However, the supplementation with probiotic in the first six months after birth did not show a reduction of the risk to develop atopic eczema (324).
According to Victoria et al., there is no clear consistent evidence indicating a protective effect of breastfeeding towards either eczema or food allergy (101). More data are therefore needed to clearly understand the role of breastfeeding in allergy prevention.
Other immune disease such as Celiac disease (CD), inflammatory bowel diseases (IBD) and MS seem to be reduced by breastfeeding (303). Indeed, breastfed infants who have developed CD have better long term health (325) owing to the later introduction of gluten (326). Likewise, IBD (327) including ulcerative colitis and Crohn disease (328) is found to be reduced by breastfeeding (327). However, in other studies, no association was found between breastfeeding and IBD (329, 330). This controversy is based on difference in the definition of exclusive or nonexclusive and duration of breastfeeding (303). Breastfeeding during at least 4 months has been associated to protection against MS (152) whereas the impact of breastfeeding duration on the risk of developing MS continues to be questioned by others (331, 332).
The current understanding of HBM supports that it is biologically and temporally adapted to the breastfeed child. Breast milk composition varies greatly from one pregnancy to another, from one mother to another, and despite these differences, breast milk remains the best food for the infant. Immune benefits associated to breastfeeding are well documented, but the role of some immune cells is still unclear and need to be investigated. Moreover, how these immune cells especially effector memory cells and some innate immune cells such as macrophages are involved in infant protection against infection and immune-related diseases need to be more investigated. As stem cells are also present in breast milk and are able to differentiate into various organs, it become ambitious to consider breast milk-derived stem cell use in transplantation.
Overall, understanding mechanisms under HBM components in newborn protection and wellbeing may increase the promotion and support of breastfeeding to achieve the third sustainable Development Goal of The United nations, namely, maternal and child health. This third sustainable goal also includes non-communicable disease such as breast cancer, overweight, obesity, and diabetes. It is noteworthy that breastfeeding is not only relevant for nutrition, a part of the second Development Goal, but also for education which is a part of the fourth Goal. Therefore, breastfeeding can effectively contribute to the reduction of poverty.
LAG conceived and wrote the manuscript. LK, AS, and ACZ edited the manuscript and figures. All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors thank Adrian Luty and Justin Doritchamou for the critical review and editing of the manuscript.
HBM, human breast milk; BDM, Breast-derived microbiota; EBV, Epstein–Barr virus; CMV, cytomegalovirus; HIV, human immunodeficiency virus; MMC, maternal microchimerism; BMI, body mass index; GI, gastrointestinal; MSC, mesenchymal stem cells; hBmScs, human breast milk stem cells; NIMA, noninherited maternal antigen; EV, extracellular vesicles; ESCRT, endosomal sorting complex required for transport; AchE, acetylcholine esterase; HLA, human leucocyte antigen; DCs, dendritic cells; BrMM, breast milk macrophage; GM-CSF, granulocyte-macrophage colony-stimulating factor; IL, interleukin; CD, cluster of differentiation; NET, neutrophile extracellular traps; MAC-1, macrophage-1 antigen; ADCC, antibody dependent cellular cytotoxicity; ADCP, antibody dependent cellular phagocytosis, ILCs, innate lymphoid cells; HAART, highly-active antiretroviral therapy; Treg, T regulatory cells; TGF, tumor growth factor; MAIT, mucosal-associated invariant T cells; NALT, nasal-associated lymphoid tissue; GALT, gut-associated lymphoid tissue; SCs, secretory cells; IFN, interferon; TNF, tumor necrosis factor; G-CSF, granulocyte-colony stimulating factor, MCP-1, monocytes chemotactic protein 1, RANTES, Regulated upon Activation, Normal T Cell Expressed and Presumably Secreted; SLE, Systemic lupus erythematosus; T1D, Type 1 Diabetes; T2D, Type 2 Diabetes; DAISY, Diabetes and Autoimmunity Study in the Young; DIPP, Prediction and Prevention Project; JIA, Juvenile idiopathic arthritis; RA, Rheumatoid arthritis; CD, Celiac disease; IBD, Inflammatory bowel diseases; MS, Multiple sclerosis.
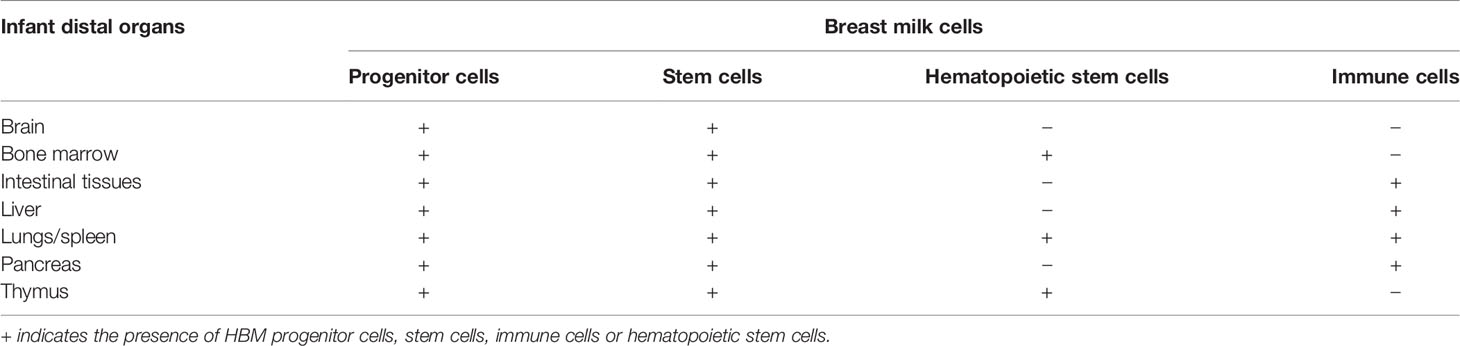
Table 1 Infant’s organs in which specific cells of HBM are found.
1. Krol KM, Grossmann T. Psychological Effects of Breastfeeding on Children and Mothers. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz (2018) 61:977–85. doi: 10.1007/s00103-018-2769-0
2. World Health Organization. Guideline, Counselling of Women to Improve Breastfeeding Practices (2018). Available at: http://www.ncbi.nlm.nih.gov/books/NBK539314/ (Accessed March 11, 2021).
3. Kennell J, McGrath S. Starting the Process of Mother-Infant Bonding. Acta Paediatr (2005) 94:775–7. doi: 10.1111/j.1651-2227.2005.tb01982.x
4. Raju TNK. Breastfeeding Is a Dynamic Biological Process–Not Simply a Meal at the Breast. Breastfeed Med (2011) 6:257–9. doi: 10.1089/bfm.2011.0081
5. Hinde K, German JB. Food in an Evolutionary Context: Insights From Mother’s Milk. J Sci Food Agric (2012) 92:2219–23. doi: 10.1002/jsfa.5720
6. Walker A. Breast Milk as the Gold Standard for Protective Nutrients. J Pediatr (2010) 156:S3–7. doi: 10.1016/j.jpeds.2009.11.021
7. Godhia M, Patel N. Colostrum - Its Composition, Benefits As A Nutraceutical : A Review. Curr Res Nutr Food Sci (2013) 1:37–47. doi: 10.12944/CRNFSJ.1.1.04
8. Brandtzaeg P. The Mucosal Immune System and Its Integration With the Mammary Glands. J Pediatr (2010) 156:S8–15. doi: 10.1016/j.jpeds.2009.11.014
9. Brandtzaeg P. Mucosal Immunity: Integration Between Mother and the Breast-Fed Infant. Vaccine (2003) 21:3382–8. doi: 10.1016/s0264-410x(03)00338-4
10. Le Doare K, Holder B, Bassett A, Pannaraj PS. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front Immunol (2018) 9:361. doi: 10.3389/fimmu.2018.00361
11. Holmlund U, Amoudruz P, Johansson MA, Haileselassie Y, Ongoiba A, Kayentao K, et al. Maternal Country of Origin, Breast Milk Characteristics and Potential Influences on Immunity in Offspring. Clin Exp Immunol (2010) 162:500–9. doi: 10.1111/j.1365-2249.2010.04275.x
12. Paquette SG, Banner D, Huang SSH, Almansa R, Leon A, Xu L, et al. Influenza Transmission in the Mother-Infant Dyad Leads to Severe Disease, Mammary Gland Infection, and Pathogenesis by Regulating Host Responses. PloS Pathog (2015) 11:e1005173. doi: 10.1371/journal.ppat.1005173
13. McCall SA, Lichy JH, Bijwaard KE, Aguilera NS, Chu W-S, Taubenberger JK. Epstein-Barr Virus Detection in Ductal Carcinoma of the Breast. JNCI J Natl Cancer Inst (2001) 93:148–50. doi: 10.1093/jnci/93.2.148
14. Turck D. Allaitement Maternel : Les Bénéfices Pour La Santé De L’enfant Et De Sa Mère. Arch Pédiatrie (2005) 12:S145–65. doi: 10.1016/j.arcped.2005.10.006
15. Lazar K, Kussmann T, Pawelec G, Pöschel S, Goelz R, Hamprecht K, et al. Immunomonitoring of Human Breast Milk Cells During HCMV-Reactivation. Front Immunol (2021) 12:723010. doi: 10.3389/fimmu.2021.723010
16. Parashar UD, Nelson EAS, Kang G. Diagnosis, Management, and Prevention of Rotavirus Gastroenteritis in Children. BMJ (2013) 347:f7204–4. doi: 10.1136/bmj.f7204
17. Lehmann GM, LaKind JS, Davis MH, Hines EP, Marchitti SA, Alcala C, et al. Environmental Chemicals in Breast Milk and Formula: Exposure and Risk Assessment Implications. Environ Health Perspect (2018) 126:096001. doi: 10.1289/EHP1953
18. Arnold C. Baby Steps Forward: Recommendations for Better Understanding Environmental Chemicals in Breast Milk and Infant Formula. Environ Health Perspect (2019) 127:064001. doi: 10.1289/EHP4804
19. Milani C, Duranti S, Bottacini F, Casey E, Turroni F, Mahony J, et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev (2017) 81(4):e00036–17. doi: 10.1128/MMBR.00036-17
20. Duranti S, Lugli GA, Mancabelli L, Armanini F, Turroni F, James K, et al. Maternal Inheritance of Bifidobacterial Communities and Bifidophages in Infants Through Vertical Transmission. Microbiome (2017) 5:66. doi: 10.1186/s40168-017-0282-6
21. Pannaraj PS, Li F, Cerini C, Bender JM, Yang S, Rollie A, et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA Pediatr (2017) 171:647. doi: 10.1001/jamapediatrics.2017.0378
22. Bode L. The Functional Biology of Human Milk Oligosaccharides. Early Hum Dev (2015) 91:619–22. doi: 10.1016/j.earlhumdev.2015.09.001
23. Fitzstevens JL, Smith KC, Hagadorn JI, Caimano MJ, Matson AP, Brownell EA. Systematic Review of the Human Milk Microbiota. Nutr Clin Pract (2017) 32:354–64. doi: 10.1177/0884533616670150
24. Gomez-Gallego C, Garcia-Mantrana I, Salminen S, Collado MC. The Human Milk Microbiome and Factors Influencing Its Composition and Activity. Semin Fetal Neonatal Med (2016) 21:400–5. doi: 10.1016/j.siny.2016.05.003
25. Hunt KM, Foster JA, Forney LJ, Schütte UME, Beck DL, Abdo Z, et al. Characterization of the Diversity and Temporal Stability of Bacterial Communities in Human Milk. PloS One (2011) 6:e21313. doi: 10.1371/journal.pone.0021313
26. Peroni DG, Pescollderungg L, Piacentini GL, Rigotti E, Maselli M, Watschinger K, et al. Immune Regulatory Cytokines in the Milk of Lactating Women From Farming and Urban Environments: Milk TGF-β1 and IL-10 and Environment. Pediatr Allergy Immunol (2010) 21:977–82. doi: 10.1111/j.1399-3038.2010.00995.x
27. Hermansson H, Kumar H, Collado MC, Salminen S, Isolauri E, Rautava S. Breast Milk Microbiota Is Shaped by Mode of Delivery and Intrapartum Antibiotic Exposure. Front Nutr (2019) 6:4. doi: 10.3389/fnut.2019.00004
28. Andreas NJ, Hyde MJ, Gomez-Romero M, Lopez-Gonzalvez MA, Villaseñor A, Wijeyesekera A, et al. Multiplatform Characterization of Dynamic Changes in Breast Milk During Lactation. Electrophoresis (2015) 36:2269–85. doi: 10.1002/elps.201500011
29. Ballard O, Morrow AL. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr Clin North Am (2013) 60:49–74. doi: 10.1016/j.pcl.2012.10.002
30. Witkowska-Zimny M, Kaminska-El-Hassan E. Cells of Human Breast Milk. Cell Mol Biol Lett (2017) 22:11. doi: 10.1186/s11658-017-0042-4
31. Wu F, Zhi X, Xu R, Liang Z, Wang F, Li X, et al. Exploration of microRNA Profiles in Human Colostrum. Ann Transl Med (2020) 8:1170. doi: 10.21037/atm-20-5709
32. Ninkina N, Kukharsky MS, Hewitt MV, Lysikova EA, Skuratovska LN, Deykin AV, et al. Stem Cells in Human Breast Milk. Hum Cell (2019) 32:223–30. doi: 10.1007/s13577-019-00251-7
33. Casavale KO, Ahuja JKC, Wu X, Li Y, Quam J, Olson R, et al. NIH Workshop on Human Milk Composition: Summary and Visions. Am J Clin Nutr (2019) 110:769–79. doi: 10.1093/ajcn/nqz123
34. Andreas NJ, Kampmann B, Mehring Le-Doare K. Human Breast Milk: A Review on Its Composition and Bioactivity. Early Hum Dev (2015) 91:629–35. doi: 10.1016/j.earlhumdev.2015.08.013
35. Lemay DG, Ballard OA, Hughes MA, Morrow AL, Horseman ND, Nommsen-Rivers LA. RNA Sequencing of the Human Milk Fat Layer Transcriptome Reveals Distinct Gene Expression Profiles at Three Stages of Lactation. PloS One (2013) 8:e67531. doi: 10.1371/journal.pone.0067531
36. Cabinian A, Sinsimer D, Tang M, Zumba O, Mehta H, Toma A, et al. Transfer of Maternal Immune Cells by Breastfeeding: Maternal Cytotoxic T Lymphocytes Present in Breast Milk Localize in the Peyer’s Patches of the Nursed Infant. PloS One (2016) 11:e0156762. doi: 10.1371/journal.pone.0156762
37. Dutta P, Burlingham WJ. Microchimerism: Tolerance vs. Sensitization. Curr Opin Organ Transplant (2011) 16:359–65. doi: 10.1097/MOT.0b013e3283484b57
38. Molès J-P, Tuaillon E, Kankasa C, Bedin A-S, Nagot N, Marchant A, et al. Breastmilk Cell Trafficking Induces Microchimerism-Mediated Immune System Maturation in the Infant. Pediatr Allergy Immunol (2018) 29:133–43. doi: 10.1111/pai.12841
39. Tuboly S, Bernáth S. Intestinal Absorption of Colostral Lymphoid Cells in Newborn Animals. Adv Exp Med Biol (2002) 503:107–14. doi: 10.1007/978-1-4615-0559-4_12
40. Camacho-Morales A, Caba M, García-Juárez M, Caba-Flores MD, Viveros-Contreras R, Martínez-Valenzuela C. Breastfeeding Contributes to Physiological Immune Programming in the Newborn. Front Pediatr (2021) 9:744104. doi: 10.3389/fped.2021.744104
41. Hassiotou F, Hepworth AR, Metzger P, Tat Lai C, Trengove N, Hartmann PE, et al. Maternal and Infant Infections Stimulate a Rapid Leukocyte Response in Breastmilk. Clin Transl Immunol (2013) 2:e3. doi: 10.1038/cti.2013.1
42. Hassiotou F, Geddes DT. Immune Cell-Mediated Protection of the Mammary Gland and the Infant During Breastfeeding. Adv Nutr (2015) 6:267–75. doi: 10.3945/an.114.007377
43. Hanson LA, Korotkova M, Lundin S, Håversen L, Silfverdal S-A, Mattsby-Baltzer I, et al. The Transfer of Immunity From Mother to Child. Ann N Y Acad Sci (2003) 987:199–206. doi: 10.1111/j.1749-6632.2003.tb06049.x
44. Hassiotou F, Geddes DT, Hartmann PE. Cells in Human Milk: State of the Science. J Hum Lact (2013) 29:171–82. doi: 10.1177/0890334413477242
45. Bode L, McGuire M, Rodriguez JM, Geddes DT, Hassiotou F, Hartmann PE, et al. It’s Alive: Microbes and Cells in Human Milk and Their Potential Benefits to Mother and Infant. Adv Nutr (2014) 5:571–3. doi: 10.3945/an.114.006643
46. Ghosh A. Breast Milk Stem Cell Survival in Neonate’s Gut, Entery Into Neonate Circulation and Adaption by the Body. Curr Stem Cell Res Ther (2020) 15:98–101. doi: 10.2174/1574888X14666191107095728
47. Twigger A-J, Hepworth AR, Lai CT, Chetwynd E, Stuebe AM, Blancafort P, et al. Gene Expression in Breastmilk Cells Is Associated With Maternal and Infant Characteristics. Sci Rep (2015) 5:12933. doi: 10.1038/srep12933
48. Hassiotou F, Hartmann PE. At the Dawn of a New Discovery: The Potential of Breast Milk Stem Cells. Adv Nutr (2014) 5:770–8. doi: 10.3945/an.114.006924
49. Thomas E, Zeps N, Cregan M, Hartmann P, Martin T. 14-3-3ς (Sigma) Regulates Proliferation and Differentiation of Multipotent P63-Positive Cells Isolated From Human Breastmilk. Cell Cycle (2011) 10:278–84. doi: 10.4161/cc.10.2.14470
50. Patki S, Kadam S, Chandra V, Bhonde R. Human Breast Milk Is a Rich Source of Multipotent Mesenchymal Stem Cells. Hum Cell (2010) 23:35–40. doi: 10.1111/j.1749-0774.2010.00083.x
51. Fan Y, Chong YS, Choolani MA, Cregan MD, Chan JKY. Unravelling the Mystery of Stem/Progenitor Cells in Human Breast Milk. PloS One (2010) 5:e14421. doi: 10.1371/journal.pone.0014421
52. Cregan MD, Fan Y, Appelbee A, Brown ML, Klopcic B, Koppen J, et al. Identification of Nestin-Positive Putative Mammary Stem Cells in Human Breastmilk. Cell Tissue Res (2007) 329:129–36. doi: 10.1007/s00441-007-0390-x
53. Li S, Zhang L, Zhou Q, Jiang S, Yang Y, Cao Y. Characterization of Stem Cells and Immune Cells in Preterm and Term Mother’s Milk. J Hum Lact (2019) 35:528–34. doi: 10.1177/0890334419838986
54. Abd Allah SH, Shalaby SM, El-Shal AS, El Nabtety SM, Khamis T, Abd El Rhman SA, et al. Breast Milk MSCs: An Explanation of Tissue Growth and Maturation of Offspring. IUBMB Life (2016) 68:935–42. doi: 10.1002/iub.1573
55. Jain N, Mathur NB, Sharma VK, Dwarkadas AM. Cellular Composition Including Lymphocyte Subsets in Preterm and Full Term Human Colostrum and Milk. Acta Paediatr Scand (1991) 80:395–9. doi: 10.1111/j.1651-2227.1991.tb11872.x
56. Aydın MŞ, Yiğit EN, Vatandaşlar E, Erdoğan E, Öztürk G. Transfer and Integration of Breast Milk Stem Cells to the Brain of Suckling Pups. Sci Rep (2018) 8:14289. doi: 10.1038/s41598-018-32715-5
57. Hosseini SM, Talaei-Khozani T, Sani M, Owrangi B. Differentiation of Human Breast-Milk Stem Cells to Neural Stem Cells and Neurons. Neurol Res Int (2014) 2014:807896. doi: 10.1155/2014/807896
58. Jain L, Vidyasagar D, Xanthou M, Ghai V, Shimada S, Blend M. In Vivo Distribution of Human Milk Leucocytes After Ingestion by Newborn Baboons. Arch Dis Child (1989) 64:930–3. doi: 10.1136/adc.64.7_spec_no.930
59. Kaingade P, Somasundaram I, Sharma A, Patel D, Marappagounder D. Cellular Components, Including Stem-Like Cells, of Preterm Mother’s Mature Milk as Compared With Those in Her Colostrum: A Pilot Study. Breastfeed Med (2017) 12:446–9. doi: 10.1089/bfm.2017.0063
60. Briere C-E, Jensen T, McGrath JM, Young EE, Finck C. Stem-Like Cell Characteristics From Breast Milk of Mothers With Preterm Infants as Compared to Mothers With Term Infants. Breastfeed Med (2017) 12:174–9. doi: 10.1089/bfm.2017.0002
61. Indumathi S, Dhanasekaran M, Rajkumar JS, Sudarsanam D. Exploring the Stem Cell and Non-Stem Cell Constituents of Human Breast Milk. Cytotechnology (2013) 65:385–93. doi: 10.1007/s10616-012-9492-8
62. Arvola M, Gustafsson E, Svensson L, Jansson L, Holmdahl R, Heyman B, et al. Immunoglobulin-Secreting Cells of Maternal Origin can be Detected in B Cell-Deficient Mice. Biol Reprod (2000) 63:1817–24. doi: 10.1095/biolreprod63.6.1817
63. Marleau AM, Greenwood JD, Wei Q, Singh B, Croy BA. Chimerism of Murine Fetal Bone Marrow by Maternal Cells Occurs in Late Gestation and Persists Into Adulthood. Lab Invest (2003) 83:673–81. doi: 10.1097/01.lab.0000067500.85003.32
64. Piotrowski P, Croy BA. Maternal Cells Are Widely Distributed in Murine Fetuses In Itero. Biol Reprod (1996) 54:1103–10. doi: 10.1095/biolreprod54.5.1103
65. Kinder JM, Stelzer IA, Arck PC, Way SS. Immunological Implications of Pregnancy-Induced Microchimerism. Nat Rev Immunol (2017) 17:483–94. doi: 10.1038/nri.2017.38
66. Kinder JM, Jiang TT, Ertelt JM, Xin L, Strong BS, Shaaban AF, et al. Tolerance to Noninherited Maternal Antigens, Reproductive Microchimerism and Regulatory T Cell Memory: 60 Years After “Evidence for Actively Acquired Tolerance to Rh Antigens.”. Chimerism (2015) 6:8–20. doi: 10.1080/19381956.2015.1107253
67. Kinder JM, Jiang TT, Ertelt JM, Xin L, Strong BS, Shaaban AF, et al. Cross-Generational Reproductive Fitness Enforced by Microchimeric Maternal Cells. Cell (2015) 162:505–15. doi: 10.1016/j.cell.2015.07.006
68. Ichinohe T. Long-Term Feto-Maternal Microchimerism Revisited: Microchimerism and Tolerance in Hematopoietic Stem Cell Transplantation. Chimerism (2010) 1:39–43. doi: 10.4161/chim.1.1.12743
69. Molès J-P, Tuaillon E, Kankasa C, Bedin A-S, Nagot N, Marchant A, et al. Breastfeeding-Related Maternal Microchimerism. Nat Rev Immunol (2017) 17:729–1. doi: 10.1038/nri.2017.115
70. Newburg DS, Walker WA. Protection of the Neonate by the Innate Immune System of Developing Gut and of Human Milk. Pediatr Res (2007) 61:2–8. doi: 10.1203/01.pdr.0000250274.68571.18
71. Adkins B. T-Cell Function in Newborn Mice and Humans. Immunol Today (1999) 20:330–5. doi: 10.1016/s0167-5699(99)01473-5
72. Schelonka RL, Infante AJ. Neonatal Immunology. Semin Perinatol (1998) 22:2–14. doi: 10.1016/S0146-0005(98)80003-7
73. Palmeira P, Carneiro-Sampaio M. Immunology of Breast Milk. Rev Assoc Med Bras (1992) (2016) 62:584–93. doi: 10.1590/1806-9282.62.06.584
74. Schanler RJ, Fraley JK, Lau C, Hurst NM, Horvath L, Rossmann SN. Breastmilk Cultures and Infection in Extremely Premature Infants. J Perinatol (2011) 31:335–8. doi: 10.1038/jp.2011.13
75. Cacho NT, Lawrence RM. Innate Immunity and Breast Milk. Front Immunol (2017) 8:584. doi: 10.3389/fimmu.2017.00584
76. Welsh JK, May JT. Anti-Infective Properties of Breast Milk. J Pediatr (1979) 94:1–9. doi: 10.1016/S0022-3476(79)80340-6
77. Goldman AS, Smith CW. Host Resistance Factors in Human Milk. J Pediatr (1973) 82:1082–90. doi: 10.1016/s0022-3476(73)80453-6
79. Trend S, de Jong E, Lloyd ML, Kok CH, Richmond P, Doherty DA, et al. Leukocyte Populations in Human Preterm and Term Breast Milk Identified by Multicolour Flow Cytometry. PloS One (2015) 10:e0135580. doi: 10.1371/journal.pone.0135580
80. Twigger A-J, Küffer GK, Geddes DT, Filgueria L. Expression of Granulisyn, Perforin and Granzymes in Human Milk Over Lactation and in the Case of Maternal Infection. Nutrients (2018) 10(9):1230. doi: 10.3390/nu10091230
81. Jin Y-Y, Zhao W, Cao R-M, Xi W, Wu S-M, Chen T-X. Characterization of Immunocompetent Cells in Human Milk of Han Chinese. J Hum Lact (2011) 27:155–62. doi: 10.1177/0890334410392041
82. Mandyla H, Xanthou M, Maravelias C, Baum D, Matsaniotis N. Antibody Dependent Cytotoxicity of Human Colostrum Phagocytes. Pediatr Res (1982) 16:995–9. doi: 10.1203/00006450-198212000-00004
83. Hughes A, Brock JH, Parrott DM, Cockburn F. The Interaction of Infant Formula With Macrophages: Effect on Phagocytic Activity, Relationship to Expression of Class II MHC Antigen and Survival of Orally Administered Macrophages in the Neonatal Gut. Immunology (1988) 64:213–8.
84. Zheng Y, Corrêa-Silva S, de Souza EC, Maria Rodrigues R, da Fonseca FAM, Gilio AE, et al. Macrophage Profile and Homing Into Breast Milk in Response to Ongoing Respiratory Infections in the Nursing Infant. Cytokine (2020) 129:155045. doi: 10.1016/j.cyto.2020.155045
85. Perdiguero EG, Geissmann F. The Development and Maintenance of Resident Macrophages. Nat Immunol (2016) 17:2–8. doi: 10.1038/ni.3341
86. Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K. Development of Monocytes, Macrophages, and Dendritic Cells. Science (2010) 327:656–61. doi: 10.1126/science.1178331
87. Weber C, Belge KU, von Hundelshausen P, Draude G, Steppich B, Mack M, et al. Differential Chemokine Receptor Expression and Function in Human Monocyte Subpopulations. J Leukoc Biol (2000) 67:699–704. doi: 10.1002/jlb.67.5.699
88. Hume DA. The Mononuclear Phagocyte System. Curr Opin Immunol (2006) 18:49–53. doi: 10.1016/j.coi.2005.11.008
89. Ichikawa M, Sugita M, Takahashi M, Satomi M, Takeshita T, Araki T, et al. Breast Milk Macrophages Spontaneously Produce Granulocyte-Macrophage Colony-Stimulating Factor and Differentiate Into Dendritic Cells in the Presence of Exogenous Interleukin-4 Alone. Immunology (2003) 108:189–95. doi: 10.1046/j.1365-2567.2003.01572.x
90. Randolph GJ, Inaba K, Robbiani DF, Steinman RM, Muller WA. Differentiation of Phagocytic Monocytes Into Lymph Node Dendritic Cells In Vivo. Immunity (1999) 11:753–61. doi: 10.1016/s1074-7613(00)80149-1
91. Ma LJ, Walter B, Deguzman A, Muller HK, Walker AM. Trans-Epithelial Immune Cell Transfer During Suckling Modulates Delayed-Type Hypersensitivity in Recipients as a Function of Gender. PloS One (2008) 3:e3562. doi: 10.1371/journal.pone.0003562
92. Medzhitov R, Schneider DS, Soares MP. Disease Tolerance as a Defense Strategy. Science (2012) 335:936–41. doi: 10.1126/science.1214935
93. Michie CA. The Long Term Effects of Breastfeeding: A Role for the Cells in Breast Milk? J Trop Pediatr (1998) 44:2–3. doi: 10.1093/tropej/44.1.2
94. Schroten H, Kuczera F, Köhler H, Adam R. Opsonophagocytosis Versus Lectinophagocytosis in Human Milk Macrophages. Adv Exp Med Biol (2000) 478:95–107. doi: 10.1007/0-306-46830-1_8
95. Kolaczkowska E, Kubes P. Neutrophil Recruitment and Function in Health and Inflammation. Nat Rev Immunol (2013) 13:159–75. doi: 10.1038/nri3399
96. Yu JC, Khodadadi H, Malik A, Davidson B, Salles É da SL, Bhatia J, et al. Innate Immunity of Neonates and Infants. Front Immunol (2018) 9:1759. doi: 10.3389/fimmu.2018.01759
97. Smith CW, Goldman AS. The Cells of Human ColostrumI. In Vitro Studies of Morphology and Functions. Pediatr Res (1968) 2:103–9. doi: 10.1203/00006450-196803000-00005
98. Weiler IJ, Hickler W, Sprenger R. Demonstration That Milk Cells Invade the Suckling Neonatal Mouse. Am J Reprod Immunol (1983) 4:95–8. doi: 10.1111/j.1600-0897.1983.tb00261.x
99. Dixon D-L, Forsyth KD. Leukocytes in Expressed Breast Milk of Asthmatic Mothers. Allergol Immunopathol (Madr) (2017) 45:325–32. doi: 10.1016/j.aller.2016.08.015
100. Järvinen K-M, Suomalainen H. Leucocytes in Human Milk and Lymphocyte Subsets in Cow’s Milk-Allergic Infants. Pediatr Allergy Immunol (2002) 13:243–54. doi: 10.1034/j.1399-3038.2002.00087.x
101. Victora CG, Bahl R, Barros AJD, França GVA, Horton S, Krasevec J, et al. Breastfeeding in the 21st Century: Epidemiology, Mechanisms, and Lifelong Effect. Lancet (2016) 387:475–90. doi: 10.1016/S0140-6736(15)01024-7
102. van Odijk J, Kull I, Borres MP, Brandtzaeg P, Edberg U, Hanson LA, et al. Breastfeeding and Allergic Disease: A Multidisciplinary Review of the Literature (1966-2001) on the Mode of Early Feeding in Infancy and Its Impact on Later Atopic Manifestations. Allergy (2003) 58:833–43. doi: 10.1034/j.1398-9995.2003.00264.x
103. Andersson M, Aeberli I, Wüst N, Piacenza AM, Bucher T, Henschen I, et al. The Swiss Iodized Salt Program Provides Adequate Iodine for School Children and Pregnant Women, But Weaning Infants Not Receiving Iodine-Containing Complementary Foods as Well as Their Mothers Are Iodine Deficient. J Clin Endocrinol Metab (2010) 95:5217–24. doi: 10.1210/jc.2010-0975
104. Fernández L, Langa S, Martín V, Jiménez E, Martín R, Rodríguez JM. The Microbiota of Human Milk in Healthy Women. Cell Mol Biol (Noisy-le-grand) (2013) 59:31–42.
105. Goldman AS, Goldblum RM. Transfer of Maternal Leukocytes to the Infant by Human Milk. Curr Top Microbiol Immunol (1997) 222:205–13. doi: 10.1007/978-3-642-60614-4_10
106. Hassiotou F, Geddes D. Anatomy of the Human Mammary Gland: Current Status of Knowledge. Clin Anat (2013) 26:29–48. doi: 10.1002/ca.22165
107. Bedin A-S, Molès J-P, Rutagwera D, Nagot N, Kankasa C, Tylleskär T, et al. MAIT Cells, TCR γδ+ Cells and ILCs Cells in Human Breast Milk and Blood From HIV Infected and Uninfected Women. Pediatr Allergy Immunol (2019) 30:479–87. doi: 10.1111/pai.13037
108. Baban B, Malik A, Bhatia J, Yu JC. Presence and Profile of Innate Lymphoid Cells in Human Breast Milk. JAMA Pediatr (2018) 172:594–6. doi: 10.1001/jamapediatrics.2018.0148
109. Fuchs A, Colonna M. Innate Lymphoid Cells in Homeostasis, Infection, Chronic Inflammation and Tumors of the Gastrointestinal Tract. Curr Opin Gastroenterol (2013) 29:581–7. doi: 10.1097/MOG.0b013e328365d339
110. Bostick JW, Zhou L. Innate Lymphoid Cells in Intestinal Immunity and Inflammation. Cell Mol Life Sci (2016) 73:237–52. doi: 10.1007/s00018-015-2055-3
111. Laouar A. Maternal Leukocytes and Infant Immune Programming During Breastfeeding. Trends Immunol (2020) 41:225–39. doi: 10.1016/j.it.2020.01.005
112. Artis D, Spits H. The Biology of Innate Lymphoid Cells. Nature (2015) 517:293–301. doi: 10.1038/nature14189
113. Sonnenberg GF, Artis D. Innate Lymphoid Cell Interactions With Microbiota: Implications for Intestinal Health and Disease. Immunity (2012) 37:601–10. doi: 10.1016/j.immuni.2012.10.003
114. Richie ER, Bass R, Meistrich ML, Dennison DK. Distribution of T Lymphocyte Subsets in Human Colostrum. J Immunol (1982) 129:1116–9.
115. Eglinton BA, Roberton DM, Cummins AG. Phenotype of T Cells, Their Soluble Receptor Levels, and Cytokine Profile of Human Breast Milk. Immunol Cell Biol (1994) 72:306–13. doi: 10.1038/icb.1994.46
116. Wirt DP, Adkins LT, Palkowetz KH, Schmalstieg FC, Goldman AS. Activated and Memory T Lymphocytes in Human Milk. Cytometry (1992) 13:282–90. doi: 10.1002/cyto.990130310
117. Bertotto A, Castellucci G, Scalise F, Vaccaro R. [Recent Findings on the Phenotype and Function of T-Lymphocytes in the Human Colostrum]. Pediatr Med Chir (1991) 13:325–9. doi: 10.1002/eji.1830200838
118. Bertotto A, Castellucci G, Scalise F, Tognellini R, Vaccaro R. “Memory” T Cells in Human Breast Milk. Acta Paediatr Scand (1991) 80:98–9. doi: 10.1111/j.1651-2227.1991.tb11738.x
119. Quayle AJ, Coston WM, Trocha AK, Kalams SA, Mayer KH, Anderson DJ. Detection of HIV-1-Specific CTLs in the Semen of HIV-Infected Individuals. J Immunol (1998) 161:4406–10.
120. Veazey RS, Marx PA, Lackner AA. Vaginal CD4+ T Cells Express High Levels of CCR5 and Are Rapidly Depleted in Simian Immunodeficiency Virus Infection. J Infect Dis (2003) 187:769–76. doi: 10.1086/368386
121. Poles MA, Elliott J, Taing P, Anton PA, Chen IS. A Preponderance of CCR5(+) CXCR4(+) Mononuclear Cells Enhances Gastrointestinal Mucosal Susceptibility to Human Immunodeficiency Virus Type 1 Infection. J Virol (2001) 75:8390–9. doi: 10.1128/jvi.75.18.8390-8399.2001
122. Kourtis AP, Jamieson DJ, de Vincenzi I, Taylor A, Thigpen MC, Dao H, et al. Prevention of Human Immunodeficiency Virus-1 Transmission to the Infant Through Breastfeeding: New Developments. Am J Obstet Gynecol (2007) 197:S113–122. doi: 10.1016/j.ajog.2007.03.003
123. Brenchley JM, Hill BJ, Ambrozak DR, Price DA, Guenaga FJ, Casazza JP, et al. T-Cell Subsets That Harbor Human Immunodeficiency Virus (HIV) In Vivo: Implications for HIV Pathogenesis. J Virol (2004) 78:1160–8. doi: 10.1128/jvi.78.3.1160-1168.2004
124. Henrick BM, Yao X-D, Nasser L, Roozrogousheh A, Rosenthal KL. Breastfeeding Behaviors and the Innate Immune System of Human Milk: Working Together to Protect Infants Against Inflammation, HIV-1, and Other Infections. Front Immunol (2017) 8:1631. doi: 10.3389/fimmu.2017.01631
125. Pedersen SH, Wilkinson AL, Andreasen A, Kinung’hi SM, Urassa M, Michael D, et al. Longitudinal Analysis of Mature Breastmilk and Serum Immune Composition Among Mixed HIV-Status Mothers and Their Infants. Clin Nutr (2016) 35:871–9. doi: 10.1016/j.clnu.2015.05.016
126. Teasdale CA, Marais BJ, Abrams EJ. HIV: Prevention of Mother-to-Child Transmission. BMJ Clin Evid (2011) 2011:0909.
127. Lewis P, Nduati R, Kreiss JK, John GC, Richardson BA, Mbori-Ngacha D, et al. Cell-Free Human Immunodeficiency Virus Type 1 in Breast Milk. J Infect Dis (1998) 177:34–9. doi: 10.1086/513816
128. Field CJ. The Immunological Components of Human Milk and Their Effect on Immune Development in Infants. J Nutr (2005) 135:1–4. doi: 10.1093/jn/135.1.1
129. Sabbaj S, Ghosh MK, Edwards BH, Leeth R, Decker WD, Goepfert PA, et al. Breast Milk-Derived Antigen-Specific CD8+ T Cells: An Extralymphoid Effector Memory Cell Population in Humans. J Immunol (2005) 174:2951–6. doi: 10.4049/jimmunol.174.5.2951
130. Ogra PL, Welliver RC. Effects of Early Environment on Mucosal Immunologic Homeostasis, Subsequent Immune Responses and Disease Outcome. Nestle Nutr Workshop Ser Pediatr Program (2008) 61:145–81. doi: 10.1159/000113492
131. Moylan DC, Pati SK, Ross SA, Fowler KB, Boppana SB, Sabbaj S. Breast Milk Human Cytomegalovirus (CMV) Viral Load and the Establishment of Breast Milk CMV-Pp65-Specific CD8 T Cells in Human CMV Infected Mothers. J Infect Dis (2017) 216:1176–9. doi: 10.1093/infdis/jix457
132. Kampen KR. The Discovery and Early Understanding of Leukemia. Leuk Res (2012) 36:6–13. doi: 10.1016/j.leukres.2011.09.028
133. Donné A. Cours De Microscopie Complémentaire Des Etudes Médicales [Electronic Resource] : Anatomie Microscopique Et Physiologie Des Fluides De L’économie. (1844).
134. MacGillivray DM, Kollmann TR. The Role of Environmental Factors in Modulating Immune Responses in Early Life. Front Immunol (2014) 5:434. doi: 10.3389/fimmu.2014.00434
135. Wood H, Acharjee A, Pearce H, Quraishi MN, Powell R, Rossiter A, et al. Breastfeeding Promotes Early Neonatal Regulatory T-Cell Expansion and Immune Tolerance of Non-Inherited Maternal Antigens. Allergy (2021) 76(8):2447–60. doi: 10.1111/all.14736
136. Jeppesen DL, Hasselbalch H, Lisse IM, Ersbøll AK, Engelmann MDM. T-Lymphocyte Subsets, Thymic Size and Breastfeeding in Infancy. Pediatr Allergy Immunol (2004) 15:127–32. doi: 10.1111/j.1399-3038.2004.00032.x
137. Hasselbalch H, Engelmann MD, Ersboll AK, Jeppesen DL, Fleischer-Michaelsen K. Breast-Feeding Influences Thymic Size in Late Infancy. Eur J Pediatr (1999) 158:964–7. doi: 10.1007/s004310051258
138. Kinder JM, Jiang TT, Clark DR, Chaturvedi V, Xin L, Ertelt JM, et al. Pregnancy-Induced Maternal Regulatory T Cells, Bona Fide Memory or Maintenance by Antigenic Reminder From Fetal Cell Microchimerism? Chimerism (2014) 5:16–9. doi: 10.4161/chim.28241
139. Safinia N, Leech J, Hernandez-Fuentes M, Lechler R, Lombardi G. Promoting Transplantation Tolerance; Adoptive Regulatory T Cell Therapy. Clin Exp Immunol (2013) 172:158–68. doi: 10.1111/cei.12052
140. Tian M, Zhang Y, Liu Z, Sun G, Mor G, Liao A. The PD-1/PD-L1 Inhibitory Pathway Is Altered in Pre-Eclampsia and Regulates T Cell Responses in Pre-Eclamptic Rats. Sci Rep (2016) 6:27683. doi: 10.1038/srep27683
141. Craenmehr MHC, Heidt S, Eikmans M, Claas FHJ. What Is Wrong With the Regulatory T Cells and Foetomaternal Tolerance in Women With Recurrent Miscarriages? HLA (2016) 87:69–78. doi: 10.1111/tan.12737
142. Somerset DA, Zheng Y, Kilby MD, Sansom DM, Drayson MT. Normal Human Pregnancy Is Associated With an Elevation in the Immune Suppressive CD25+ CD4+ Regulatory T-Cell Subset. Immunology (2004) 112:38–43. doi: 10.1111/j.1365-2567.2004.01869.x
143. Cérbulo-Vázquez A, Hernández-Peláez G, Arriaga-Pizano LA, Bautista-Pérez P, Romero-Venado J, Flores-González JC, et al. Characterization of CD127- CD25++ Treg From Human Colostrum. Am J Reprod Immunol (2018) 79(2). doi: 10.1111/aji.12806
144. Parigi SM, Eldh M, Larssen P, Gabrielsson S, Villablanca EJ. Breast Milk and Solid Food Shaping Intestinal Immunity. Front Immunol (2015) 6:415. doi: 10.3389/fimmu.2015.00415
145. Steer PJ, Modi N. Elective Caesarean Sections–Risks to the Infant. Lancet (2009) 374:675–6. doi: 10.1016/S0140-6736(09)61544-0
146. Zhang L, Miller RG. The Correlation of Prolonged Survival of Maternal Skin Grafts With the Presence of Naturally Transferred Maternal T Cells. Transplantation (1993) 56:918–21. doi: 10.1097/00007890-199310000-00027
147. Campbell DA, Lorber MI, Sweeton JC, Turcotte JG, Niederhuber JE, Beer AE. Breast Feeding and Maternal-Donor Renal Allografts. Possibly the Original Donor-Specific Transfusion. Transplantation (1984) 37:340–4. doi: 10.1097/00007890-198404000-00004
148. Chappert P, Leboeuf M, Rameau P, Lalfer M, Desbois S, Liblau RS, et al. Antigen-Specific Treg Impair CD8(+) T-Cell Priming by Blocking Early T-Cell Expansion. Eur J Immunol (2010) 40:339–50. doi: 10.1002/eji.200839107
149. Chappert P, Schwartz RH. Induction of T Cell Anergy: Integration of Environmental Cues and Infectious Tolerance. Curr Opin Immunol (2010) 22:552–9. doi: 10.1016/j.coi.2010.08.005
150. Xu Q, Lee J, Jankowska-Gan E, Schultz J, Roenneburg DA, Roennburg DA, et al. Human CD4+CD25low Adaptive T Regulatory Cells Suppress Delayed-Type Hypersensitivity During Transplant Tolerance. J Immunol (2007) 178:3983–95. doi: 10.4049/jimmunol.178.6.3983
151. Stevens AM. Maternal Microchimerism in Health and Disease. Best Pract Res Clin Obstet Gynaecol (2016) 31:121–30. doi: 10.1016/j.bpobgyn.2015.08.005
152. Conradi S, Malzahn U, Paul F, Quill S, Harms L, Then Bergh F, et al. Breastfeeding Is Associated With Lower Risk for Multiple Sclerosis. Mult Scler (2013) 19:553–8. doi: 10.1177/1352458512459683
153. Yamamoto T, Tsubota Y, Kodama T, Kageyama-Yahara N, Kadowaki M. Oral Tolerance Induced by Transfer of Food Antigens via Breast Milk of Allergic Mothers Prevents Offspring From Developing Allergic Symptoms in a Mouse Food Allergy Model. Clin Dev Immunol (2012) 2012:721085. doi: 10.1155/2012/721085
154. Radlovic NP, Mladenovic MM, Lekovic ZM, Stojsic ZM, Radlovic VN. Influence of Early Feeding Practices on Celiac Disease in Infants. Croat Med J (2010) 51:417–22. doi: 10.3325/cmj.2010.51.417
155. Verhasselt V, Milcent V, Cazareth J, Kanda A, Fleury S, Dombrowicz D, et al. Breast Milk-Mediated Transfer of an Antigen Induces Tolerance and Protection From Allergic Asthma. Nat Med (2008) 14:170–5. doi: 10.1038/nm1718
156. Tangye SG, Good KL. Human IgM+CD27+ B Cells: Memory B Cells or “Memory” B Cells? J Immunol (2007) 179:13–9. doi: 10.4049/jimmunol.179.1.13
157. Liu YJ, Barthélémy C, de Bouteiller O, Arpin C, Durand I, Banchereau J. Memory B Cells From Human Tonsils Colonize Mucosal Epithelium and Directly Present Antigen to T Cells by Rapid Up-Regulation of B7-1 and B7-2. Immunity (1995) 2:239–48. doi: 10.1016/1074-7613(95)90048-9
158. Salmi M, Jalkanen S. How do Lymphocytes Know Where to Go: Current Concepts and Enigmas of Lymphocyte Homing. Adv Immunol (1997) 64:139–218. doi: 10.1016/s0065-2776(08)60889-5
159. Roux ME, McWilliams M, Phillips-Quagliata JM, Weisz-Carrington P, Lamm ME. Origin of IgA-Secreting Plasma Cells in the Mammary Gland. J Exp Med (1977) 146:1311–22. doi: 10.1084/jem.146.5.1311
160. Shi Y, Agematsu K, Ochs HD, Sugane K. Functional Analysis of Human Memory B-Cell Subpopulations: IgD+CD27+ B Cells Are Crucial in Secondary Immune Response by Producing High Affinity IgM. Clin Immunol (2003) 108:128–37. doi: 10.1016/s1521-6616(03)00092-5
161. Klein U, Rajewsky K, Küppers R. Human Immunoglobulin (Ig)M+IgD+ Peripheral Blood B Cells Expressing the CD27 Cell Surface Antigen Carry Somatically Mutated Variable Region Genes: CD27 as a General Marker for Somatically Mutated (Memory) B Cells. J Exp Med (1998) 188:1679–89. doi: 10.1084/jem.188.9.1679
162. Weller S, Braun MC, Tan BK, Rosenwald A, Cordier C, Conley ME, et al. Human Blood IgM “Memory” B Cells Are Circulating Splenic Marginal Zone B Cells Harboring a Prediversified Immunoglobulin Repertoire. Blood (2004) 104:3647–54. doi: 10.1182/blood-2004-01-0346
163. Bush JF, Beer AE. Analysis of Complement Receptors on B-Lymphocytes in Human Milk. Am J Obstet Gynecol (1979) 133:708–12. doi: 10.1016/0002-9378(79)90023-1
164. Weisz-Carrington P, Roux ME, Lamm ME. Plasma Cells and Epithelial Immunoglobulins in the Mouse Mammary Gland During Pregnancy and Lactation. J Immunol (1977) 119:1306–7.
165. Tanneau GM, Hibrand-Saint Oyant L, Chevaleyre CC, Salmon HP. Differential Recruitment of T- and IgA B-Lymphocytes in the Developing Mammary Gland in Relation to Homing Receptors and Vascular Addressins. J Histochem Cytochem (1999) 47:1581–92. doi: 10.1177/002215549904701210
166. Fishaut M, Murphy D, Neifert M, McIntosh K, Ogra PL. Bronchomammary Axis in the Immune Response to Respiratory Syncytial Virus. J Pediatr (1981) 99:186–91. doi: 10.1016/s0022-3476(81)80447-7
167. Johansen F-E, Baekkevold ES, Carlsen HS, Farstad IN, Soler D, Brandtzaeg P. Regional Induction of Adhesion Molecules and Chemokine Receptors Explains Disparate Homing of Human B Cells to Systemic and Mucosal Effector Sites: Dispersion From Tonsils. Blood (2005) 106:593–600. doi: 10.1182/blood-2004-12-4630
168. Csencsits KL, Walters N, Pascual DW. Cutting Edge: Dichotomy of Homing Receptor Dependence by Mucosal Effector B Cells: Alpha(E) Versus L-Selectin. J Immunol (2001) 167:2441–5. doi: 10.4049/jimmunol.167.5.2441
169. Csencsits KL, Pascual DW. Absence of L-Selectin Delays Mucosal B Cell Responses in Nonintestinal Effector Tissues. J Immunol (2002) 169:5649–59. doi: 10.4049/jimmunol.169.10.5649
170. Kunkel EJ, Butcher EC. Plasma-Cell Homing. Nat Rev Immunol (2003) 3:822–9. doi: 10.1038/nri1203
171. Williams MB, Rosé JR, Rott LS, Franco MA, Greenberg HB, Butcher EC. The Memory B Cell Subset Responsible for the Secretory IgA Response and Protective Humoral Immunity to Rotavirus Expresses the Intestinal Homing Receptor, Alpha4beta7. J Immunol (1998) 161:4227–35.
172. Hamann A, Andrew DP, Jablonski-Westrich D, Holzmann B, Butcher EC. Role of Alpha 4-Integrins in Lymphocyte Homing to Mucosal Tissues In Vivo. J Immunol (1994) 152:3282–93.
173. Lazarus NH, Kunkel EJ, Johnston B, Wilson E, Youngman KR, Butcher EC. A Common Mucosal Chemokine (Mucosae-Associated Epithelial Chemokine/CCL28) Selectively Attracts IgA Plasmablasts. J Immunol (2003) 170:3799–805. doi: 10.4049/jimmunol.170.7.3799
174. Wilson E, Butcher EC. CCL28 Controls Immunoglobulin (Ig)A Plasma Cell Accumulation in the Lactating Mammary Gland and IgA Antibody Transfer to the Neonate. J Exp Med (2004) 200:805–9. doi: 10.1084/jem.20041069
175. Tuaillon E, Valea D, Becquart P, Al Tabaa Y, Meda N, Bollore K, et al. Human Milk-Derived B Cells: A Highly Activated Switched Memory Cell Population Primed to Secrete Antibodies. J Immunol (2009) 182:7155–62. doi: 10.4049/jimmunol.0803107
176. Kourtis AP, Ibegbu CC, Theiler R, Xu Y-X, Bansil P, Jamieson DJ, et al. Breast Milk CD4+ T Cells Express High Levels of C Chemokine Receptor 5 and CXC Chemokine Receptor 4 and Are Preserved in HIV-Infected Mothers Receiving Highly Active Antiretroviral Therapy. J Infect Dis (2007) 195:965–72. doi: 10.1086/512082
177. Petitjean G, Becquart P, Tuaillon E, Al Tabaa Y, Valea D, Huguet M-F, et al. Isolation and Characterization of HIV-1-Infected Resting CD4+ T Lymphocytes in Breast Milk. J Clin Virol (2007) 39:1–8. doi: 10.1016/j.jcv.2007.02.004
178. Ramsay DT, Kent JC, Owens RA, Hartmann PE. Ultrasound Imaging of Milk Ejection in the Breast of Lactating Women. Pediatrics (2004) 113:361–7. doi: 10.1542/peds.113.2.361
179. Moossavi S, Azad MB. Origins of Human Milk Microbiota: New Evidence and Arising Questions. Gut Microbes (2020) 12:1667722. doi: 10.1080/19490976.2019.1667722
180. Bourges D, Meurens F, Berri M, Chevaleyre C, Zanello G, Levast B, et al. New Insights Into the Dual Recruitment of IgA+ B Cells in the Developing Mammary Gland. Mol Immunol (2008) 45:3354–62. doi: 10.1016/j.molimm.2008.04.017
181. Moussa S, Jenabian M-A, Gody JC, Léal J, Grésenguet G, Le Faou A, et al. Adaptive HIV-Specific B Cell-Derived Humoral Immune Defenses of the Intestinal Mucosa in Children Exposedvia Breast-Feeding. PloS One (2013) 8:e63408. doi: 10.1371/journal.pone.0063408
182. Brandtzaeg P. Function of Mucosa-Associated Lymphoid Tissue in Antibody Formation. Immunol Invest (2010) 39:303–55. doi: 10.3109/08820131003680369
183. Junghans RP. Finally! The Brambell Receptor (FcRB). Mediator of Transmission of Immunity and Protection From Catabolism for IgG. Immunol Res (1997) 16:29–57. doi: 10.1007/BF02786322
184. Avanzini MA, Pignatti P, Chirico G, Gasparoni A, Jalil F, Hanson LA. Placental Transfer Favours High Avidity IgG Antibodies. Acta Paediatr (1998) 87:180–5. doi: 10.1080/08035259850157633
185. Koenig A, de Albuquerque Diniz EM, Barbosa SFC, Vaz FAC. Immunologic Factors in Human Milk: The Effects of Gestational Age and Pasteurization. J Hum Lact (2005) 21:439–43. doi: 10.1177/0890334405280652
186. Hanson LA. Breastfeeding Provides Passive and Likely Long-Lasting Active Immunity. Ann Allergy Asthma Immunol (1998) 81:523–33. doi: 10.1016/S1081-1206(10)62704-4
187. Nolan LS, Parks OB, Good M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients (2019) 12(1):14. doi: 10.3390/nu12010014
188. Castellote C, Casillas R, Ramírez-Santana C, Pérez-Cano FJ, Castell M, Moretones MG, et al. Premature Delivery Influences the Immunological Composition of Colostrum and Transitional and Mature Human Milk. J Nutr (2011) 141:1181–7. doi: 10.3945/jn.110.133652
189. Mehta R, Petrova A. Biologically Active Breast Milk Proteins in Association With Very Preterm Delivery and Stage of Lactation. J Perinatol (2011) 31:58–62. doi: 10.1038/jp.2010.68
190. Mehta R, Petrova A. Very Preterm Gestation and Breastmilk Cytokine Content During the First Month of Lactation. Breastfeed Med (2011) 6:21–4. doi: 10.1089/bfm.2010.0024
191. Hsu P, Nanan RKH. Innate and Adaptive Immune Interactions at the Fetal and Maternal Interface in Healthy Human Pregnancy and Pre-Eclampsia. Front Immunol (2014) 5:125. doi: 10.3389/fimmu.2014.00125
192. Kadaoui KA, Corthésy B. Secretory IgA Mediates Bacterial Translocation to Dendritic Cells in Mouse Peyer’s Patches With Restriction to Mucosal Compartment. J Immunol (2007) 179:7751–7. doi: 10.4049/jimmunol.179.11.7751
193. Carr LE, Virmani MD, Rosa F, Munblit D, Matazel KS, Elolimy AA, et al. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front Immunol (2021) 12:604080. doi: 10.3389/fimmu.2021.604080
194. Murphy CA, O’Reilly DP, Neary E, El-Khuffash A, NíAinle F, McCallion N, et al. A Review of the Role of Extracellular Vesicles in Neonatal Physiology and Pathology. Pediatr Res (2020) 90:289–99. doi: 10.1038/s41390-020-01240-5
195. Hock A, Miyake H, Li B, Lee C, Ermini L, Koike Y, et al. Breast Milk-Derived Exosomes Promote Intestinal Epithelial Cell Growth. J Pediatr Surg (2017) 52:755–9. doi: 10.1016/j.jpedsurg.2017.01.032
196. Spencer N, Yeruva L. Role of Bacterial Infections in Extracellular Vesicles Release and Impact on Immune Response. BioMed J (2020) 44(2):157–64. doi: 10.1016/j.bj.2020.05.006
197. Admyre C, Johansson SM, Qazi KR, Filén J-J, Lahesmaa R, Norman M, et al. Exosomes With Immune Modulatory Features Are Present in Human Breast Milk. J Immunol (2007) 179:1969–78. doi: 10.4049/jimmunol.179.3.1969
198. Admyre C, Bohle B, Johansson SM, Focke-Tejkl M, Valenta R, Scheynius A, et al. B Cell-Derived Exosomes can Present Allergen Peptides and Activate Allergen-Specific T Cells to Proliferate and Produce TH2-Like Cytokines. J Allergy Clin Immunol (2007) 120:1418–24. doi: 10.1016/j.jaci.2007.06.040
199. Cocucci E, Meldolesi J. Ectosomes and Exosomes: Shedding the Confusion Between Extracellular Vesicles. Trends Cell Biol (2015) 25:364–72. doi: 10.1016/j.tcb.2015.01.004
200. Taylor DD, Taylor CG, Jiang CG, Black PH. Characterization of Plasma Membrane Shedding From Murine Melanoma Cells. Int J Cancer (1988) 41:629–35. doi: 10.1002/ijc.2910410425
201. Zhang X, Yuan X, Shi H, Wu L, Qian H, Xu W. Exosomes in Cancer: Small Particle, Big Player. J Hematol Oncol (2015) 8:83. doi: 10.1186/s13045-015-0181-x
202. Bobrie A, Colombo M, Krumeich S, Raposo G, Théry C. Diverse Subpopulations of Vesicles Secreted by Different Intracellular Mechanisms Are Present in Exosome Preparations Obtained by Differential Ultracentrifugation. J Extracell Vesicles (2012) 1. doi: 10.3402/jev.v1i0.18397
203. Théry C, Amigorena S, Raposo G, Clayton A. Isolation and Characterization of Exosomes From Cell Culture Supernatants and Biological Fluids. Curr Protoc Cell Biol (2006) Chapter 3:Unit 3.22. doi: 10.1002/0471143030.cb0322s30
204. Vargas A, Zhou S, Éthier-Chiasson M, Flipo D, Lafond J, Gilbert C, et al. Syncytin Proteins Incorporated in Placenta Exosomes Are Important for Cell Uptake and Show Variation in Abundance in Serum Exosomes From Patients With Preeclampsia. FASEB J (2014) 28:3703–19. doi: 10.1096/fj.13-239053
205. Lokossou AG, Toudic C, Barbeau B. Implication of Human Endogenous Retrovirus Envelope Proteins in Placental Functions. Viruses (2014) 6:4609–27. doi: 10.3390/v6114609
206. Torregrosa Paredes P, Gutzeit C, Johansson S, Admyre C, Stenius F, Alm J, et al. Differences in Exosome Populations in Human Breast Milk in Relation to Allergic Sensitization and Lifestyle. Allergy (2014) 69:463–71. doi: 10.1111/all.12357
207. Simpson RJ, Jensen SS, Lim JWE. Proteomic Profiling of Exosomes: Current Perspectives. Proteomics (2008) 8:4083–99. doi: 10.1002/pmic.200800109
208. Lokossou AG, Toudic C, Nguyen PT, Elisseeff X, Vargas A, Rassart É, et al. Endogenous Retrovirus-Encoded Syncytin-2 Contributes to Exosome-Mediated Immunosuppression of T Cells†. Biol Reprod (2020) 102:185–98. doi: 10.1093/biolre/ioz124
209. Torregrosa Paredes P, Esser J, Admyre C, Nord M, Rahman QK, Lukic A, et al. Bronchoalveolar Lavage Fluid Exosomes Contribute to Cytokine and Leukotriene Production in Allergic Asthma. Allergy (2012) 67:911–9. doi: 10.1111/j.1398-9995.2012.02835.x
210. Golan-Gerstl R, Elbaum Shiff Y, Moshayoff V, Schecter D, Leshkowitz D, Reif S. Characterization and Biological Function of Milk-Derived miRNAs. Mol Nutr Food Res (2017) 61(10). doi: 10.1002/mnfr.201700009
211. Kosaka N, Iguchi H, Yoshioka Y, Takeshita F, Matsuki Y, Ochiya T. Secretory Mechanisms and Intercellular Transfer of microRNAs in Living Cells. J Biol Chem (2010) 285:17442–52. doi: 10.1074/jbc.M110.107821
212. Cui J, Zhou B, Ross SA, Zempleni J. Nutrition, microRNAs, and Human Health. Adv Nutr (2017) 8:105–12. doi: 10.3945/an.116.013839
213. Zempleni J, Aguilar-Lozano A, Sadri M, Sukreet S, Manca S, Wu D, et al. Biological Activities of Extracellular Vesicles and Their Cargos From Bovine and Human Milk in Humans and Implications for Infants. J Nutr (2017) 147:3–10. doi: 10.3945/jn.116.238949
214. van Herwijnen MJC, Driedonks TAP, Snoek BL, Kroon AMT, Kleinjan M, Jorritsma R, et al. Abundantly Present miRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Front Nutr (2018) 5:81. doi: 10.3389/fnut.2018.00081
215. Vaswani K, Koh YQ, Almughlliq FB, Peiris HN, Mitchell MD. A Method for the Isolation and Enrichment of Purified Bovine Milk Exosomes. Reprod Biol (2017) 17:341–8. doi: 10.1016/j.repbio.2017.09.007
216. Alsaweed M, Hepworth AR, Lefèvre C, Hartmann PE, Geddes DT, Hassiotou F. Human Milk MicroRNA and Total RNA Differ Depending on Milk Fractionation. J Cell Biochem (2015) 116:2397–407. doi: 10.1002/jcb.25207
217. Alsaweed M, Hartmann PE, Geddes DT, Kakulas F. MicroRNAs in Breastmilk and the Lactating Breast: Potential Immunoprotectors and Developmental Regulators for the Infant and the Mother. Int J Environ Res Public Health (2015) 12:13981–4020. doi: 10.3390/ijerph121113981
218. Lotvall J, Valadi H. Cell to Cell Signalling via Exosomes Through esRN. Available at: https://www.ncbi.nlm.nih.gov/pubmed/19262134.
219. Cheng L, Sun X, Scicluna BJ, Coleman BM, Hill AF. Characterization and Deep Sequencing Analysis of Exosomal and Non-Exosomal miRNA in Human Urine. Kidney Int (2014) 86:433–44. doi: 10.1038/ki.2013.502
220. Cheng L, Quek CYJ, Sun X, Bellingham SA, Hill AF. The Detection of microRNA Associated With Alzheimer’s Disease in Biological Fluids Using Next-Generation Sequencing Technologies. Front Genet (2013) 4:150. doi: 10.3389/fgene.2013.00150
221. Zhou Q, Li M, Wang X, Li Q, Wang T, Zhu Q, et al. Immune-Related microRNAs Are Abundant in Breast Milk Exosomes. Int J Biol Sci (2012) 8:118–23. doi: 10.7150/ijbs.8.118
222. Liao Y, Du X, Li J, Lönnerdal B. Human Milk Exosomes and Their microRNAs Survive Digestion In Vitro and Are Taken Up by Human Intestinal Cells. Mol Nutr Food Res (2017) 61(11). doi: 10.1002/mnfr.201700082
223. Liu X, Zhan Z, Xu L, Ma F, Li D, Guo Z, et al. MicroRNA-148/152 Impair Innate Response and Antigen Presentation of TLR-Triggered Dendritic Cells by Targeting Camkiiα. J Immunol (2010) 185:7244–51. doi: 10.4049/jimmunol.1001573
224. Wells AC, Daniels KA, Angelou CC, Fagerberg E, Burnside AS, Markstein M, et al. Modulation of Let-7 miRNAs Controls the Differentiation of Effector CD8 T Cells. Elife (2017) 6:e26398. doi: 10.7554/eLife.26398
225. Carney MC, Tarasiuk A, DiAngelo SL, Silveyra P, Podany A, Birch LL, et al. Metabolism-Related microRNAs in Maternal Breast Milk Are Influenced by Premature Delivery. Pediatr Res (2017) 82:226–36. doi: 10.1038/pr.2017.54
226. Corsetti PP, de Almeida LA, Gonçalves ANA, Gomes MTR, Guimarães ES, Marques JT, et al. miR-181a-5p Regulates TNF-α and miR-21a-5p Influences Gualynate-Binding Protein 5 and IL-10 Expression in Macrophages Affecting Host Control of Brucella Abortus Infection. Front Immunol (2018) 9:1331. doi: 10.3389/fimmu.2018.01331
227. Zhou F, Paz HA, Sadri M, Cui J, Kachman SD, Fernando SC, et al. Dietary Bovine Milk Exosomes Elicit Changes in Bacterial Communities in C57BL/6 Mice. Am J Physiol Gastrointest Liver Physiol (2019) 317:G618–24. doi: 10.1152/ajpgi.00160.2019
228. Nolan LS, Rimer JM, Good M. The Role of Human Milk Oligosaccharides and Probiotics on the Neonatal Microbiome and Risk of Necrotizing Enterocolitis: A Narrative Review. Nutrients (2020) 12(10):3052. doi: 10.3390/nu12103052
229. Ustundag B, Yilmaz E, Dogan Y, Akarsu S, Canatan H, Halifeoglu I, et al. Levels of Cytokines (IL-1beta, IL-2, IL-6, IL-8, TNF-Alpha) and Trace Elements (Zn, Cu) in Breast Milk From Mothers of Preterm and Term Infants. Mediators Inflammation (2005) 2005:331–6. doi: 10.1155/MI.2005.331
230. Böttcher MF, Jenmalm MC, Garofalo RP, Björkstén B. Cytokines in Breast Milk From Allergic and Nonallergic Mothers. Pediatr Res (2000) 47:157–62. doi: 10.1203/00006450-200001000-00026
231. Hawkes JS, Bryan D-L, Gibson RA. Cytokine Production by Human Milk Cells and Peripheral Blood Mononuclear Cells From the Same Mothers. J Clin Immunol (2002) 22:338–44. doi: 10.1023/a:1020652215048
232. Polat A, Tunc T, Erdem G, Yerebasmaz N, Tas A, Beken S, et al. Interleukin-8 and Its Receptors in Human Milk From Mothers of Full-Term and Premature Infants. Breastfeed Med (2016) 11:247–51. doi: 10.1089/bfm.2015.0186
233. Bryan DL, Hawkes JS, Gibson RA. Interleukin-12 in Human Milk. Pediatr Res (1999) 45:858–9. doi: 10.1203/00006450-199906000-00013
234. Calhoun DA, Lunøe M, Du Y, Staba SL, Christensen RD. Concentrations of Granulocyte Colony-Stimulating Factor in Human Milk After In Vitro Simulations of Digestion. Pediatr Res (1999) 46:767–71. doi: 10.1203/00006450-199912000-00021
235. Jung HH, Kim J-Y, Lim JE, Im Y-H. Cytokine Profiling in Serum-Derived Exosomes Isolated by Different Methods. Sci Rep (2020) 10:14069. doi: 10.1038/s41598-020-70584-z
236. Donnet-Hughes A, Duc N, Serrant P, Vidal K, Schiffrin EJ. Bioactive Molecules in Milk and Their Role in Health and Disease: The Role of Transforming Growth Factor-Beta. Immunol Cell Biol (2000) 78:74–9. doi: 10.1046/j.1440-1711.2000.00882.x
237. Amoudruz P, Holmlund U, Schollin J, Sverremark-Ekström E, Montgomery SM. Maternal Country of Birth and Previous Pregnancies Are Associated With Breast Milk Characteristics. Pediatr Allergy Immunol (2009) 20:19–29. doi: 10.1111/j.1399-3038.2008.00754.x
238. Saito S, Maruyama M, Kato Y, Moriyama I, Ichijo M. Detection of IL-6 in Human Milk and Its Involvement in IgA Production. J Reprod Immunol (1991) 20:267–76. doi: 10.1016/0165-0378(91)90051-Q
239. van Vlasselaer P, Punnonen J, de Vries JE. Transforming Growth Factor-Beta Directs IgA Switching in Human B Cells. J Immunol (1992) 148:2062–7.
240. Olivares M, Diaz-Ropero MP, Martin R, Rodriguez JM, Xaus J. Antimicrobial Potential of Four Lactobacillus Strains Isolated From Breast Milk. J Appl Microbiol (2006) 101:72–9. doi: 10.1111/j.1365-2672.2006.02981.x
241. Riaz Rajoka MS, Mehwish HM, Siddiq M, Haobin Z, Zhu J, Yan L, et al. Identification, Characterization, and Probiotic Potential of Lactobacillus Rhamnosus Isolated From Human Milk. LWT (2017) 84:271–80. doi: 10.1016/j.lwt.2017.05.055
242. Lyons KE, Ryan CA, Dempsey EM, Ross RP, Stanton C. Breast Milk, a Source of Beneficial Microbes and Associated Benefits for Infant Health. Nutrients (2020) 12:1039. doi: 10.3390/nu12041039
243. Lackey KA, Williams JE, Meehan CL, Zachek JA, Benda ED, Price WJ, et al. Corrigendum: What’s Normal? Microbiomes in Human Milk and Infant Feces Are Related to Each Other But Vary Geographically: The INSPIRE Study. Front Nutr (2020) 7:12. doi: 10.3389/fnut.2020.00012
244. Murphy K, Curley D, O’Callaghan TF, O’Shea C-A, Dempsey EM, O’Toole PW, et al. The Composition of Human Milk and Infant Faecal Microbiota Over the First Three Months of Life: A Pilot Study. Sci Rep (2017) 7:40597. doi: 10.1038/srep40597
245. Jiang M, Zhang F, Wan C, Xiong Y, Shah NP, Wei H, et al. Evaluation of Probiotic Properties of Lactobacillus Plantarum WLPL04 Isolated From Human Breast Milk. J Dairy Sci (2016) 99:1736–46. doi: 10.3168/jds.2015-10434
246. Li N, Pang B, Liu G, Zhao X, Xu X, Jiang C, et al. Lactobacillus Rhamnosus From Human Breast Milk Shows Therapeutic Function Against Foodborne Infection by Multi-Drug Resistant Escherichia Coli in Mice. Food Funct (2020) 11:435–47. doi: 10.1039/C9FO01698H
247. Kansandee W, Moonmangmee D, Moonmangmee S, Itsaranuwat P. Characterization and Bifidobacterium Sp. Growth Stimulation of Exopolysaccharide Produced by Enterococcus Faecalis EJRM152 Isolated From Human Breast Milk. Carbohydr Polymers (2019) 206:102–9. doi: 10.1016/j.carbpol.2018.10.117
248. Martïn R, Jimnez E, Heilig H, Fernïndez L, Marïn ML, Zoetendal EG, et al. Isolation of Bifidobacteria From Breast Milk and Assessment of the Bifidobacterial Population by PCR-Denaturing Gradient Gel Electrophoresis and Quantitative Real-Time PCR. Appl Environ Microbiol (2009) 75:965–9. doi: 10.1128/AEM.02063-08
249. Gavin A, Ostovar K. Microbiological Characterization of Human Milk1. J Food Prot (1977) 40:614–6. doi: 10.4315/0362-028X-40.9.614
250. Heikkila MP, Saris PEJ. Inhibition of Staphylococcus Aureus by the Commensal Bacteria of Human Milk. J Appl Microbiol (2003) 95:471–8. doi: 10.1046/j.1365-2672.2003.02002.x
251. Cabrera-Rubio R, Collado MC, Laitinen K, Salminen S, Isolauri E, Mira A. The Human Milk Microbiome Changes Over Lactation and Is Shaped by Maternal Weight and Mode of Delivery. Am J Clin Nutr (2012) 96:544–51. doi: 10.3945/ajcn.112.037382
252. Browne PD, Aparicio M, Alba C, Hechler C, Beijers R, Rodríguez JM, et al. Human Milk Microbiome and Maternal Postnatal Psychosocial Distress. Front Microbiol (2019) 10:2333. doi: 10.3389/fmicb.2019.02333
253. Li S-W, Watanabe K, Hsu C-C, Chao S-H, Yang Z-H, Lin Y-J, et al. Bacterial Composition and Diversity in Breast Milk Samples From Mothers Living in Taiwan and Mainland China. Front Microbiol (2017) 8:965. doi: 10.3389/fmicb.2017.00965
254. Khodayar-Pardo P, Mira-Pascual L, Collado MC, Martínez-Costa C. Impact of Lactation Stage, Gestational Age and Mode of Delivery on Breast Milk Microbiota. J Perinatol (2014) 34:599–605. doi: 10.1038/jp.2014.47
255. Urbaniak C, Angelini M, Gloor GB, Reid G. Human Milk Microbiota Profiles in Relation to Birthing Method, Gestation and Infant Gender. Microbiome (2016) 4:1. doi: 10.1186/s40168-015-0145-y
256. Soto A, Martín V, Jiménez E, Mader I, Rodríguez JM, Fernández L. Lactobacilli and Bifidobacteria in Human Breast Milk: Influence of Antibiotherapy and Other Host and Clinical Factors. J Pediatr Gastroenterol Nutr (2014) 59:78–88. doi: 10.1097/MPG.0000000000000347
257. Olivares M, Albrecht S, De Palma G, Ferrer MD, Castillejo G, Schols HA, et al. Human Milk Composition Differs in Healthy Mothers and Mothers With Celiac Disease. Eur J Nutr (2015) 54:119–28. doi: 10.1007/s00394-014-0692-1
258. Kumar H, du Toit E, Kulkarni A, Aakko J, Linderborg KM, Zhang Y, et al. Distinct Patterns in Human Milk Microbiota and Fatty Acid Profiles Across Specific Geographic Locations. Front Microbiol (2016) 7:1619. doi: 10.3389/fmicb.2016.01619
259. Dimitriu PA, Iker B, Malik K, Leung H, Mohn WW, Hillebrand GG. New Insights Into the Intrinsic and Extrinsic Factors That Shape the Human Skin Microbiome. mBio (2019) 10:e00839–19. doi: 10.1128/mBio.00839-19
260. Moossavi S, Sepehri S, Robertson B, Bode L, Goruk S, Field CJ, et al. Composition and Variation of the Human Milk Microbiota Are Influenced by Maternal and Early-Life Factors. Cell Host Microbe (2019) 25:324–35.e4. doi: 10.1016/j.chom.2019.01.011
261. Fernández L, Langa S, Martín V, Maldonado A, Jiménez E, Martín R, et al. The Human Milk Microbiota: Origin and Potential Roles in Health and Disease. Pharmacol Res (2013) 69:1–10. doi: 10.1016/j.phrs.2012.09.001
262. Lawrence RM, Pane CA. Human Breast Milk: Current Concepts of Immunology and Infectious Diseases. Curr Problems Pediatr Adolesc Health Care (2007) 37:7–36. doi: 10.1016/j.cppeds.2006.10.002
263. Sankar MJ, Sinha B, Chowdhury R, Bhandari N, Taneja S, Martines J, et al. Optimal Breastfeeding Practices and Infant and Child Mortality: A Systematic Review and Meta-Analysis. Acta Paediatr (2015) 104:3–13. doi: 10.1111/apa.13147
264. Lodge C, Tan D, Lau M, Dai X, Tham R, Lowe A, et al. Breastfeeding and Asthma and Allergies: A Systematic Review and Meta-Analysis. Acta Paediatr (2015) 104:38–53. doi: 10.1111/apa.13132
265. Binns C, Lee M, Low WY. The Long-Term Public Health Benefits of Breastfeeding. Asia Pac J Public Health (2016) 28:7–14. doi: 10.1177/1010539515624964
266. Sands BE. Inflammatory Bowel Disease: Past, Present, and Future. J Gastroenterol (2007) 42:16–25. doi: 10.1007/s00535-006-1995-7
267. von Hertzen LC, Haahtela T. Asthma and Atopy - the Price of Affluence? Allergy (2004) 59:124–37. doi: 10.1046/j.1398-9995.2003.00433.x
268. Aarnisalo J, Ilonen J, Vainionpää R, Volanen I, Kaitosaari T, Simell O. Development of Antibodies Against Cytomegalovirus, Varicella-Zoster Virus and Herpes Simplex Virus in Finland During the First Eight Years Of Life: A Prospective Study. Scand J Infect Dis (2003) 35:750–3. doi: 10.1080/00365540310015881
269. Petrova M. Breastfeeding and Chronic HBV Infection: Clinical and Social Implications. WJG (2010) 16:5042. doi: 10.3748/wjg.v16.i40.5042
270. Madore LS, Fisher DJ. The Role of Breast Milk in Infectious Disease. Clinics Perinatol (2021) 48:359–78. doi: 10.1016/j.clp.2021.03.008
271. Wegienka G, Ownby DR, Havstad S, Williams LK, Johnson CC. Breastfeeding History and Childhood Allergic Status in a Prospective Birth Cohort. Ann Allergy Asthma Immunol (2006) 97:78–83. doi: 10.1016/S1081-1206(10)61374-9
272. Dong Y, Chi X, Hai H, Sun L, Zhang M, Xie W-F, et al. Antibodies in the Breast Milk of a Maternal Woman With COVID-19. Emerging Microbes Infect (2020) 9:1467–9. doi: 10.1080/22221751.2020.1780952
273. Fox A, Marino J, Amanat F, Krammer F, Hahn-Holbrook J, Zolla-Pazner S, et al. Robust and Specific Secretory IgA Against SARS-CoV-2 Detected in Human Milk. iScience (2020) 23:101735. doi: 10.1016/j.isci.2020.101735
274. Zhang Z-J, Yu X-J, Fu T, Liu Y, Jiang Y, Yang BX, et al. Novel Coronavirus Infection in Newborn Babies Aged <28 Days in China. Eur Respir J (2020) 55:2000697. doi: 10.1183/13993003.00697-2020
275. Kimberlin DW, Puopolo KM. reast Milk and COVID-19: What Do We Know? Clin Infect Dis (2020) 72(1):131–2. doi: 10.1093/cid/ciaa800
276. Groß R, Conzelmann C, Müller JA, Stenger S, Steinhart K, Kirchhoff F, et al. Detection of SARS-CoV-2 in Human Breastmilk. Lancet (2020) 395:1757–8. doi: 10.1016/S0140-6736(20)31181-8
277. Tam PCK, Ly KM, Kernich ML, Spurrier N, Lawrence D, Gordon DL, et al. Detectable Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in Human Breast Milk of a Mildly Symptomatic Patient With Coronavirus Disease 2019 (COVID-19). Clin Infect Dis (2020) 72(1):128–30. doi: 10.1093/cid/ciaa673
278. Labayo HKM, Pajuelo MJ, Tohma K, Ford-Siltz LA, Gilman RH, Cabrera L, et al. Norovirus-Specific Immunoglobulin A in Breast Milk for Protection Against Norovirus-Associated Diarrhea Among Infants. EClinicalMedicine (2020) 27:100561. doi: 10.1016/j.eclinm.2020.100561
279. American Academy of Pediatrics. RED BOOK 2018: Report of the Committee on Infectious Diseases. Elk Grove Village, Illinois: Collaborative Group on Hormonal Factors in Breast CancerAmerican Academy of Pediatrics (2018).
280. Ojo-Okunola A, Nicol M, du Toit E. Human Breast Milk Bacteriome in Health and Disease. Nutrients (2018) 10:1643. doi: 10.3390/nu10111643
281. Fons AG, Tuomo Karjalainen M. Mechanisms of Colonisation and Colonisation Resistance of the Digestive Tract Part 2: Bacteria/Bacteria Interactions. Microb Ecol Health Dis (2000) 12:240–6. doi: 10.1080/089106000750060495
282. Lawrence RM, Lawrence RA. Breast Milk and Infection. Clinics Perinatol (2004) 31:501–28. doi: 10.1016/j.clp.2004.03.019
283. Hokama T, Sakamoto R, Yara A, Asato Y, Takamine F, Itokazu K. Incidence of Haemophilus Influenzae in the Throats of Healthy Infants With Different Feeding Methods. Pediatr Int (1999) 41:277–80. doi: 10.1046/j.1442-200x.1999.01068.x
284. Puopolo KM, Lynfield R, Cummings JJ. American Academy of Pediatrics, Committee on Fetus and Newborn, Committee on Infectious Diseases. Management of Infants at Risk for Group B Streptococcal Disease. Pediatrics (2019) 144(2):e20191881. doi: 10.1542/peds.2019-2350
285. Zimmermann P, Gwee A, Curtis N. The Controversial Role of Breast Milk in GBS Late-Onset Disease. J Infect (2017) 74:S34–40. doi: 10.1016/S0163-4453(17)30189-5
286. Breastfeeding and Human Lactation. 4th ed. Riordan J, Wambach K, editors. Sudbury, Mass: Jones and Bartlett Publishers (2010). 912 p.
287. Lawrence RM. Transmission of Infectious Diseases Through Breast Milk and Breastfeeding. In: Breastfeeding. Philadelphia, PA: Elsevier (2011). p. 406–73. doi: 10.1016/B978-1-4377-0788-5.10013-6
288. Hassan Z, Mustafa S, Rahim RA, Isa NM. Anti-Breast Cancer Effects of Live, Heat-Killed and Cytoplasmic Fractions of Enterococcus Faecalis and Staphylococcus Hominis Isolated From Human Breast Milk. In Vitro Cell Dev Biol-Animal (2016) 52:337–48. doi: 10.1007/s11626-015-9978-8
289. Kim JY, Woo HJ, Kim Y-S, Kim KH, Lee HJ. Cell Cycle Dysregulation Induced by Cytoplasm of Lactococcus Lactis Ssp. Lactis in SNUC2A, a Colon Cancer Cell Line. Nutr Cancer (2003) 46:197–201. doi: 10.1207/S15327914NC4602_13
290. Delgado S, Arroyo R, Martín R, Rodríguez JM. PCR-DGGE Assessment of the Bacterial Diversity of Breast Milk in Women With Lactational Infectious Mastitis. BMC Infect Dis (2008) 8:51. doi: 10.1186/1471-2334-8-51
291. Foxman B. Lactation Mastitis: Occurrence and Medical Management Among 946 Breastfeeding Women in the United States. Am J Epidemiol (2002) 155:103–14. doi: 10.1093/aje/155.2.103
292. Arroyo R, Martín V, Maldonado A, Jiménez E, Fernández L, Rodríguez JM. Treatment of Infectious Mastitis During Lactation: Antibiotics Versus Oral Administration of Lactobacilli Isolated From Breast Milk. Clin Infect Dis (2010) 50:1551–8. doi: 10.1086/652763
293. Collaborative Group on Hormonal Factors in Breast Cancer. Breast Cancer and Breastfeeding: Collaborative Reanalysis of Individual Data From 47 Epidemiological Studies in 30 Countries, Including 50 302 Women With Breast Cancer and 96 973 Women Without the Disease. Lancet (2002) 360:187–95. doi: 10.1016/S0140-6736(02)09454-0
294. Chowdhury R, Sinha B, Sankar MJ, Taneja S, Bhandari N, Rollins N, et al. Breastfeeding and Maternal Health Outcomes: A Systematic Review and Meta-Analysis. Acta Paediatr (2015) 104:96–113. doi: 10.1111/apa.13102
295. Ambrosone CB, Higgins MJ. Relationships Between Breast Feeding and Breast Cancer Subtypes: Lessons Learned From Studies in Humans and in Mice. Cancer Res (2020) 80:4871–7. doi: 10.1158/0008-5472.CAN-20-0077
296. Mosca F, Giannì ML. Human Milk: Composition and Health Benefits. Pediatr Med Chir (2017) 39(2):155. doi: 10.4081/pmc.2017.155
297. Aune D, Norat T, Romundstad P, Vatten LJ. Breastfeeding and the Maternal Risk of Type 2 Diabetes: A Systematic Review and Dose–Response Meta-Analysis of Cohort Studies. Nutrition Metab Cardiovasc Dis (2014) 24:107–15. doi: 10.1016/j.numecd.2013.10.028
298. Horta BL, de Lima NP. Breastfeeding and Type 2 Diabetes: Systematic Review and Meta-Analysis. Curr Diabetes Rep (2019) 19:1. doi: 10.1007/s11892-019-1121-x
299. for the Million Women Study Collaborators, Bobrow KL, Quigley MA, Green J, Reeves GK, Beral V. Persistent Effects of Women’s Parity and Breastfeeding Patterns on Their Body Mass Index: Results From the Million Women Study. Int J Obes (2013) 37:712–7. doi: 10.1038/ijo.2012.76
300. Moon JH, Kim H, Kim H, Park J, Choi W, Choi W, et al. Lactation Improves Pancreatic β Cell Mass and Function Through Serotonin Production. Sci Transl Med (2020) 12:eaay0455. doi: 10.1126/scitranslmed.aay0455
301. Buskila D, Shoenfeld Y. Prolactin, Bromocriptine and Autoimmune Diseases. Isr J Med Sci (1996) 32:23–7.
302. Anaya J-M, Shoenfeld Y. Multiple Autoimmune Disease in a Patient With Hyperprolactinemia. Isr Med Assoc J (2005) 7:740–1.
303. Vieira Borba V, Sharif K, Shoenfeld Y. Breastfeeding and Autoimmunity: Programing Health From the Beginning. Am J Reprod Immunol (2018) 79:e12778. doi: 10.1111/aji.12778
304. Atkinson MA, Eisenbarth GS, Michels AW. Type 1 Diabetes. Lancet (2014) 383:69–82. doi: 10.1016/S0140-6736(13)60591-7
305. Patterson CC, Gyürüs E, Rosenbauer J, Cinek O, Neu A, Schober E, et al. Trends in Childhood Type 1 Diabetes Incidence in Europe During 1989–2008: Evidence of Non-Uniformity Over Time in Rates of Increase. Diabetologia (2012) 55:2142–7. doi: 10.1007/s00125-012-2571-8
306. Norwegian Childhood Diabetes Study Group, Skrivarhaug T, Stene LC, Drivvoll AK, Strøm H, Joner G. Incidence of Type 1 Diabetes in Norway Among Children Aged 0–14 Years Between 1989 and 2012: Has the Incidence Stopped Rising? Results From the Norwegian Childhood Diabetes Registry. Diabetologia (2014) 57:57–62. doi: 10.1007/s00125-013-3090-y
307. Lund-Blix NA, Stene LC, Rasmussen T, Torjesen PA, Andersen LF, Rønningen KS. Infant Feeding in Relation to Islet Autoimmunity and Type 1 Diabetes in Genetically Susceptible Children: The MIDIA Study. Diabetes Care (2015) 38:257–63. doi: 10.2337/dc14-1130
308. Borch-Johnsen K, Mandrup-Poulsen T, Zachau-Christiansen B, Joner G, Christy M, Kastrup K, et al. Relation Between Breast-Feeding and Incidence Rates of Insulin-Dependent Diabetes Mellitus. Lancet (1984) 324:1083–6. doi: 10.1016/S0140-6736(84)91517-4
309. Cardwell CR, Stene LC, Ludvigsson J, Rosenbauer J, Cinek O, Svensson J, et al. Breast-Feeding and Childhood-Onset Type 1 Diabetes. Diabetes Care (2012) 35:2215–25. doi: 10.2337/dc12-0438
310. Smithers LG, Kramer MS, Lynch JW. Effects of Breastfeeding on Obesity and Intelligence: Causal Insights From Different Study Designs. JAMA Pediatr (2015) 169:707. doi: 10.1001/jamapediatrics.2015.0175
311. Nucci AM, Virtanen SM, Becker DJ. Infant Feeding and Timing of Complementary Foods in the Development of Type 1 Diabetes. Curr Diabetes Rep (2015) 15:62. doi: 10.1007/s11892-015-0628-z
312. Halipchuk J, Temple B, Dart A, Martin D, Sellers EAC. Prenatal, Obstetric and Perinatal Factors Associated With the Development of Childhood-Onset Type 2 Diabetes. Can J Diabetes (2018) 42:71–7. doi: 10.1016/j.jcjd.2017.04.003
313. Kindgren E, Fredrikson M, Ludvigsson J. Early Feeding and Risk of Juvenile Idiopathic Arthritis: A Case Control Study in a Prospective Birth Cohort. Pediatr Rheumatol (2017) 15:46. doi: 10.1186/s12969-017-0175-z
314. Shenoi S, Shaffer ML, Wallace CA. Environmental Risk Factors and Early-Life Exposures in Juvenile Idiopathic Arthritis: A Case-Control Study: JIA Environmental Risk Factors. Arthritis Care Res (2016) 68:1186–94. doi: 10.1002/acr.22806
315. Stoll ML, Kumar R, Morrow CD, Lefkowitz EJ, Cui X, Genin A, et al. Altered Microbiota Associated With Abnormal Humoral Immune Responses to Commensal Organisms in Enthesitis-Related Arthritis. Arthritis Res Ther (2014) 16:486. doi: 10.1186/s13075-014-0486-0
316. Chen H, Wang J, Zhou W, Yin H, Wang M. Breastfeeding and Risk of Rheumatoid Arthritis: A Systematic Review and Metaanalysis. J Rheumatol (2015) 42:1563–9. doi: 10.3899/jrheum.150195
317. Oddy W. TGF-β in Human Milk Is Associated With Wheeze in Infancy. J Allergy Clin Immunol (2003) 112:723–8. doi: 10.1016/S0091-6749(03)01941-9
318. Kalliomäki M, Ouwehand A, Arvilommi H, Kero P, Isolauri E. Transforming Growth Factor-β in Breast Milk: A Potential Regulator of Atopic Disease at an Early Age. J Allergy Clin Immunol (1999) 104:1251–7. doi: 10.1016/S0091-6749(99)70021-7
319. Böttcher MF, Abrahamsson TR, Fredriksson M, Jakobsson T, Björkstén B. Low Breast Milk TGF-β2 Is Induced by Lactobacillus Reuteri Supplementation and Associates With Reduced Risk of Sensitization During Infancy. Pediatr Allergy Immunol (2008) 19:497–504. doi: 10.1111/j.1399-3038.2007.00687.x
320. Snijders BEP, Damoiseaux JGMC, Penders J, Kummeling I, Stelma FF, van Ree R, et al. Cytokines and Soluble CD14 in Breast Milk in Relation With Atopic Manifestations in Mother and Infant (KOALA Study). Clin Exp Allergy (2006) 36:1609–15. doi: 10.1111/j.1365-2222.2006.02613.x
321. Lara-Villoslada F, Olivares M, Sierra S, Miguel Rodríguez J, Boza J, Xaus J. Beneficial Effects of Probiotic Bacteria Isolated From Breast Milk. Br J Nutr (2007) 98:S96–S100. doi: 10.1017/S0007114507832910
322. Abrahamsson TR, Jakobsson T, Böttcher MF, Fredrikson M, Jenmalm MC, Björkstén B, et al. Probiotics in Prevention of IgE-Associated Eczema: A Double-Blind, Randomized, Placebo-Controlled Trial. J Allergy Clin Immunol (2007) 119:1174–80. doi: 10.1016/j.jaci.2007.01.007
323. Kim JY, Kwon JH, Ahn SH, Lee SI, Han YS, Choi YO, et al. Effect of Probiotic Mix (Bifidobacterium Bifidum, Bifidobacterium Lactis, Lactobacillus Acidophilus) in the Primary Prevention of Eczema: A Double-Blind, Randomized, Placebo-Controlled Trial. Pediatr Allergy Immunol (2010) 21:e386–93. doi: 10.1111/j.1399-3038.2009.00958.x
324. Taylor AL, Dunstan JA, Prescott SL. Probiotic Supplementation for the First 6 Months of Life Fails to Reduce the Risk of Atopic Dermatitis and Increases the Risk of Allergen Sensitization in High-Risk Children: A Randomized Controlled Trial. J Allergy Clin Immunol (2007) 119:184–91. doi: 10.1016/j.jaci.2006.08.036
325. Cunningham AS, Jelliffe DB, Patrice Jelliffe EF. Breast-Feeding and Health in the 1980s: A Global Epidemiologic Review. J Pediatr (1991) 118:659–66. doi: 10.1016/S0022-3476(05)80023-X
326. Guandalini S. The Influence of Gluten: Weaning Recommendations for Healthy Children and Children at Risk for Celiac Disease. In: Agostoni C, Brunser O, editors. Nestlé Nutrition Workshop Series: Pediatric Program. Basel: Karger (2007). p. 139–55. doi: 10.1159/000106366
327. van der Sloot KWJ, Amini M, Peters V, Dijkstra G, Alizadeh BZ. Inflammatory Bowel Diseases: Review of Known Environmental Protective and Risk Factors Involved. Inflamm Bowel Dis (2017) 23:1499–509. doi: 10.1097/MIB.0000000000001217
328. Ek WE, D’Amato M, Halfvarson J. The History of Genetics in Inflammatory Bowel Disease. Ann Gastroenterol (2014) 27:294–303.
329. Khalili H, Ananthakrishnan AN, Higuchi LM, Richter JM, Fuchs CS, Chan AT. Early Life Factors and Risk of Inflammatory Bowel Disease in Adulthood. Inflamm Bowel Dis (2013) 19:542–7. doi: 10.1097/MIB.0b013e31828132f8
330. Roberts SE. Perinatal and Early Life Risk Factors for Inflammatory Bowel Disease. WJG (2011) 17:743. doi: 10.3748/wjg.v17.i6.743
331. Dick G, Piscane A, Impagliazzo N, Russo M, Valiani R, Mandarini A, et al. Breast Feeding and Multiple Sclerosis: Research Could Have Been Reviewed. BMJ (1994) 309:610–0. doi: 10.1136/bmj.309.6954.610a
Keywords: human breast milk, breastfeeding, mother and infant health, immunity, microbiota
Citation: Lokossou GAG, Kouakanou L, Schumacher A and Zenclussen AC (2022) Human Breast Milk: From Food to Active Immune Response With Disease Protection in Infants and Mothers. Front. Immunol. 13:849012. doi: 10.3389/fimmu.2022.849012
Received: 05 January 2022; Accepted: 07 March 2022;
Published: 05 April 2022.
Edited by:
Eduardo Luzía França, Federal University of Mato Grosso, BrazilReviewed by:
Gergely Toldi, University of Auckland, New ZealandCopyright © 2022 Lokossou, Kouakanou, Schumacher and Zenclussen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anne Schumacher, YW5uZS5zY2h1bWFjaGVyQHVmei5kZQ==; Gatien A. G. Lokossou, Z2F0aWVuLmxva29zc291QGVwYWMudWFjLmJq
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.