
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 26 July 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.842651
This article is part of the Research Topic Towards Precision Medicine for Immune-Mediated Disorders: Advances in Using Big Data and Artificial Intelligence to Understand Heterogeneity in Inflammatory Responses View all 19 articles
 Yale Liu1,2,3
Yale Liu1,2,3 Hao Wang4
Hao Wang4 Christopher Cook2,3Mark A. Taylor3,5Jeffrey P. North3
Christopher Cook2,3Mark A. Taylor3,5Jeffrey P. North3 Ashley Hailer2,3
Ashley Hailer2,3 Yanhong Shou6
Yanhong Shou6 Arsil Sadik2Esther Kim7Elizabeth Purdom4
Arsil Sadik2Esther Kim7Elizabeth Purdom4 Jeffrey B. Cheng2,3*†
Jeffrey B. Cheng2,3*† Raymond J. Cho3*†
Raymond J. Cho3*†Identifying genetic variation underlying human diseases establishes targets for therapeutic development and helps tailor treatments to individual patients. Large-scale transcriptomic profiling has extended the study of such molecular heterogeneity between patients to somatic tissues. However, the lower resolution of bulk RNA profiling, especially in a complex, composite tissue such as the skin, has limited its success. Here we demonstrate approaches to interrogate patient-level molecular variance in a chronic skin inflammatory disease, psoriasis vulgaris, leveraging single-cell RNA-sequencing of CD45+ cells isolated from active lesions. Highly psoriasis-specific transcriptional abnormalities display greater than average inter-individual variance, nominating them as potential sources of clinical heterogeneity. We find that one of these chemokines, CXCL13, demonstrates significant correlation with severity of lesions within our patient series. Our analyses also establish that genes elevated in psoriatic skin-resident memory T cells are enriched for programs orchestrating chromatin and CDC42-dependent cytoskeleton remodeling, specific components of which are distinctly correlated with and against Th17 identity on a single-cell level. Collectively, these analyses describe systematic means to dissect cell type- and patient-level differences in cutaneous psoriasis using high-resolution transcriptional profiles of human inflammatory disease.
Individuals with psoriasis vulgaris broadly share cutaneous features such as erythema, micaceous scale, and induction at skin sites affected by friction. While the role of Th17 cell-produced cytokines such as IL17F and IL26 in generating these phenotypes is well-established (1, 2), the distinctive morphology of these lesions suggests that a broad array of yet uncharacterized downstream effector genes are also specific to and shared by psoriatic lesions. Conversely, individual cases of psoriasis can markedly differ in presentation. Each patient develops lesions in distinct anatomic patterns, for example whether the scalp or intertriginous skin is involved, and lesional itch is also highly variable. These patterns of difference must reflect underlying molecular heterogeneity, potentially related to other clinical features such as involvement of other organ systems (e.g. psoriatic arthritis) or response to the many pathway-targeting agents now available for treatment. One well-established example is the involvement of germline CARD14 variants in psoriasis patients with presentations overlapping with or including the disease state pityriasis rubra pilaris (3). However, many more yet undiscovered gene-based variances on the genetic and epigenetic level are likely to determine an individual patient’s clinical state.
In the past, bulk RNA-sequencing of tissue obtained from lesional skin has been used to detect and define such commonalities and differences, enabling rough estimates of genetic variance in both psoriasis vulgaris and atopic dermatitis (4–6). Such approaches, however, conflate gene expression values from many different immune and stromal cell types, providing relatively crude estimates of genetic similarity and variance. The recent emergence of single-cell profiling technologies, such as single cell RNA sequencing (scRNA-seq) and Cellular Indexing of Transcriptomes and Epitopes (CITE-seq) (7), offers the ability to compare instances of chronic skin inflammatory disease with far greater resolution. We can now ask, for example, what molecular abnormalities are shared by effector immune cells in most psoriasis patients, regardless of clinical presentation? Such recurrent derangements might suggest treatment of psoriasis with existing drugs affecting those targets. Alternatively, certain molecular abnormalities are likely to be found in only a subset of individual cases, nominating them as candidates for specific targeted therapies.
To formally deconstruct discrete levels of molecular heterogeneity underlying cutaneous psoriatic inflammation, we analyzed data from a recent study profiling 8 psoriasis samples and 7 normal controls using single-cell RNAseq (scRNA-seq) and CITE-seq based on the 10X Genomics Chromium platform (8). We intended to develop and test approaches to scRNA-seq datasets profiling chronic inflammatory disease that could be practically and widely applied as similar datasets become published.
Patient recruitment and methods are detailed in our companion publication (8). Briefly, written informed consent was procured from donors providing both normal and psoriatic lesional skin under protocols approved by the University of California, San Francisco Institutional Review Board. Full thickness punch biopsies (6 mm) were obtained from psoriasis lesions; discards from abdominoplasties and mammoplasties were used as normal controls. All patients had not used topical immunosuppressives for at least 2 weeks before biopsy. All patients were naïve to targeted biologic medications or disease-modifying non-steroidal agents except for Patient 5, who was under systemic immunosuppression following a liver transplant. Clinical details of psoriasis samples in our series are described in Supplementary Table 1.
Details of skin biopsy sample processing, CD45+ immune cell isolation, 10X Genomics 3’ scRNA-seq and CITE-seq library preparation, and data analysis are further described in a recent prior publication (8). Briefly, we initially performed high-resolution clustering and eliminated populations corresponding to non-immune and low quality cells (mitochondrial genes percentage <20%, 100 < nFeatures < 6000). With the remaining cells, we performed unsupervised clustering with the following in Seurat (15 harmonies to run UMAP() and 1.0 resolution for FindClusters()to obtain the final 20 clusters used in this analysis. Marker transcripts for each cluster were identified using the FindAllMarkers function in Seurat (results are in Supplementary Table 2). Cluster identities were then manually annotated based on canonical immune cell population markers.
We created pseudo-bulk counts for each patient for the cells that were mapped to CD45+ cell subpopulations using the package muscat (9) in Bioconductor. The muscat method aggregates the single-cell data at the cluster-sample level to create pseudo-bulk data and then applies the methods of edgeR (10) to pseudo-bulk calculations to identify DEGs between normal and psoriasis samples (volcano plot, Figure 2A). To calculate dispersion values of the psoriasis samples, we applied the function estimateTagwiseDisp from the edgeR package in Bioconductor to the pseudo-bulk counts from the psoriasis samples. To identify abnormally elevated, functionally related gene sets (e.g. Gene Ontology (GO), Reactome) in Trm2, we applied the Metascape package (11, 12) to significant DEGs identified by FindMarkers() in comparison to grouped healthy controls (p < 0.05).
Although all psoriasis biopsies were 0.6 cm in diameter, different proportions of isolated cells were scRNA-seq processed for each sample. To determine the number of T cells expressing each DEG in each biopsy, we took the assessed number of expressing T cells for a given DEG and adjusted by total number of CD45+ cells obtained from each biopsy/total cell number processed in Seurat.
Gene values were batch-corrected at the sample level using the CPCA method in the R package iCellR; missing gene values were independently imputed within inflamed and unflamed states of sample-aligned matrices using the PCA method in iCellR/run.impute. Resulting matrices were then used for the correlation matrix. Rstudio v.1.4.1717 and GraphPad Prism (version 8.0; GraphPad Software, La Jolla, California) were used for statistical analysis and heatmap generation. Pearson correlation coefficients were calculated for gene-gene comparisons using the R function cor(). Adjusted p < 0.05 was considered significant for Seurat-based analyses, while p < 0.05 was used for other analyses.
We focused on 7 normal and 8 psoriasis samples from the Liu et al. study (8) (Supplementary Table 1). Diagnoses were based on clinical evaluation by a board-certified dermatologist and confirmed by formal histopathological reading. Six of eight patients were judged to have moderate to severe disease based on Psoriasis Area and Severity Index (PASI) scores and two (Patients 2 and 5) were in the mild range (Supplementary Table 1). The only patient known to be taking systemic immunosuppressive treatments within 4 weeks of biopsy was Patient 5, who was maintained daily on 4 mg of tacrolimus and 1250 mg of mycophenolate mofetil following a liver transplant. Normal controls were taken from discarded tissue obtained from mammoplasties and abdominoplasties.
Briefly, skin biopsies were enzymatically digested and flow sorted for live CD45+ cells, which were then subjected to Chromium 3’ single cell RNA-seq and CITE-seq protein epitope sequencing. Single-cell transcriptomic data was obtained from an average of ~5,200 single cells per sample after eliminating doublets, poor-quality, as well as non-immune cells. To classify cells, a graph-based clustering approach using Louvain community detection-based modularity optimization, available in the Seurat package, was utilized.
We obtained 20 cell types based on previously described unsupervised clustering approaches (8). Robust representation of each sample was observed (Supplementary Data 1). As shown in Figure 1, the most upregulated transcripts in each cluster (so-called marker genes) define a central memory cell population (CD3D+/CCR7+/SELL+/KLF2+) we call Tcm, as well as a migratory memory class Tmm (CD3D+/CCR7+/SELL-). Based on expression of ITGAE (CD103), CXCR6, and CD69, we identified three resident memory populations (Trm1, Trm2, and Trm3). A CD4+ regulatory T cell (Treg) population was noted based on the expression of FOXP3, TIGIT, CTLA4, IL2RA (CD25), and IKZF2 (Helios).
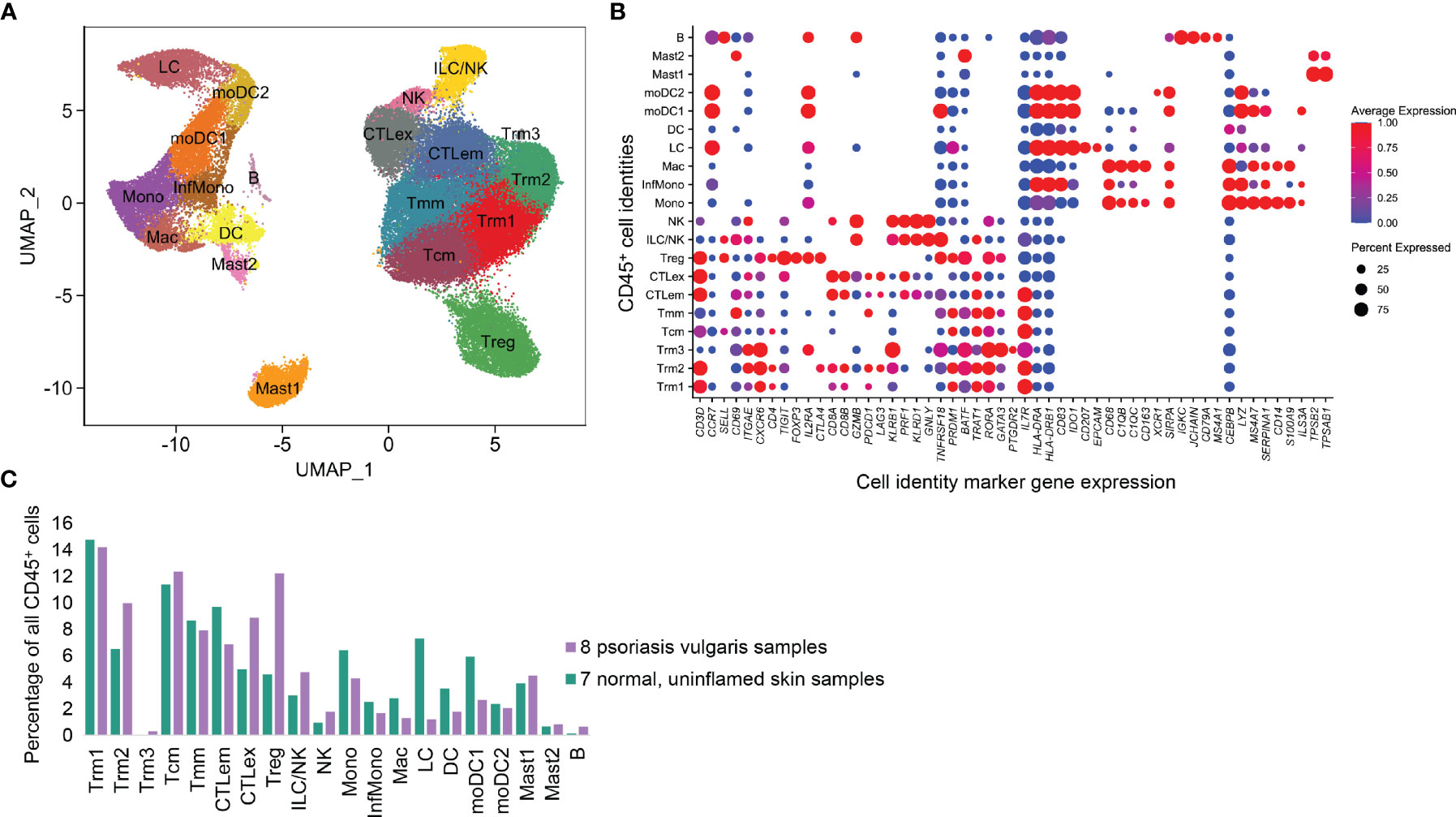
Figure 1 CD45+ immune cell types identified from 8 psoriasis vulgaris lesions and 7 normal skin samples. (A) UMAP representation of 11 T cell and 9 APC classes based on scRNA-seq transcriptional data, in which each point represents a single cell. (B) Expression of critical marker transcripts distinguishing immune cell classes. (C) Proportion of each immune class in total CD45+ cell populations.
Two cytotoxic (CD8A+CD8B+) T cell clusters expressing CCL5, GZMB, and NKG7 were identified. One we annotated as cytotoxic effector memory cells (CTLem) due to expression of effector molecules including TNFRSF18 and CD96, as well as resident markers CD69 and ITGAE. Interestingly, the second cytotoxic T cell population was quantitatively enriched in the psoriasis vs. normal samples and contained elevated canonical exhaustion markers such as PDCD1 and LAG3. Accordingly, this population was classified as exhausted T cells (CTLex). There were also two populations with high KLRD1+, GNLY+, PRF1+, and GZMB+ expression, one with high levels of the CD56 epitope by CITE-seq (NK cells) and the other defined as ILC/NK cells.
Antigen-presenting cell types (APCs) were also classified based on canonical markers. A macrophage population was enriched for CD68, CEBPB, and FCER1G, as well as complement transcripts C1QB and C1QC and the scavenger receptor CD163 (Mac). We also examined four monocyte or monocyte-derived cell populations with elevated MS4A7, LYZ, and SERPINA1. There was an inflammatory monocyte (InfMono) population characterized by increased IL1B and IL23A and another cluster of classical monocytes (Mono) which expressed higher S100A9 and CD14. Two of these clusters also expressed very high MHCII molecule levels (HLA-DRA, HLA-DRB1) and were identified as monocyte-derived DC (moDC1 and moDC2). A dendritic cell (DC) class (HLA-DRA+) was enriched in CLEC10A. A population with EPCAM, and CD207 was defined as Langerhans cells (LC). A small population comported with the B cell lineage, with high expression of IGHG, IGHA, IGKC, JCHAIN, CD19, and MA4A2). Two clusters of Mast cells (Mast) were distinguished by expression of TPSB2 and TPSAB1 (Mast1 and Mast2).
We next applied a pseudo-bulk method to identify differentially related genes (DEGs) that distinguished immune cell populations in our 8 psoriasis samples from 7 grouped healthy control biopsies. This approach aggregates scRNA-seq-derived gene counts for each cell subpopulation in each individual sample. Standard bulk mRNA-Seq computational approaches for differential expression were then applied, thereby allowing for patient-level variance to influence the significance of individual DEGs (9). One notable feature of our recent comparisons of psoriasis and other rash types such as atopic dermatitis is that the large majority of psoriasis-specific transcriptional changes are detected in Trm (8). For example, in the Tcm compartment, excluding mitochondrial and ribosomal transcripts, only KLRB1, IL17R, and JUN were expressed at greater than 0.5 logFC in psoriasis compared to normal samples. In Tregs, only CPM, TNFRSF, CD7, FTH1, IL7R, MAGEH1, MAL, TBC1D4, met these criteria. For APC classes, the far smaller number of cells captured in our CD45+ cell-centric approach led to detection of even fewer highly specific DEGs.
Consistent with these recent findings, our pseudo-bulk analysis primarily detected upregulation of Th17 cytokines such as IL17F and IL26, as well as established psoriasis inflammatory markers such as IFNG and CXCL13 (13), in a skin-resident memory T cell compartment (Trm2). Therefore, we mainly focused on this T cell class for further analysis. Overall, in Trm2 we identified 1,425 transcripts that distinguished psoriasis from healthy controls at a p value of < 0.05 (Supplementary Table 3).
Understanding patient-level variance, alongside fold-change magnitudes, is foundational to the conceptualization and use of disease biomarkers. Transcripts that distinguish psoriatic from normal tissue at higher log-fold change and relatively low patient-specific variance may perform well in broad screening efforts. Conversely, DEGs with high patient-level variance should be investigated as possible sources of phenotypic variance between affected individuals. To assess the contribution of patient-level variation to DEG identification in the skin-resident memory T cell population Trm2, we calculated dispersion values using edgeR (14) across our dataset. Lower dispersion scores correlate with lesser patient-level variation, which increases the significance of a pseudo-bulk-identified DEGs at a given logFC. Figure 2A plots logFC (x-axis) and dispersion score (y-axis), with transcripts with p < 0.05 adjusted value shown in red. In addition to the Th17 cytokines noted above, this representation shows significant elevation of immune activation markers such as CTLA4, CCR5, CD109, and ZEB2. We also saw clear suppression of other inflammatory pathways, including the interferon signaling genes IFITM1, IFITM3, and IFI6 and the chemokines CXCL3 and CXCL8. This representation also illustrates how greater patient-level variation for a given DEG (higher dispersion score along the y-axis) decreases its significance. For example, IL1R1, implicated in licensing Th17 cytokine production (15), ADA2, an adenosine deaminase central to T cell maturation (16), and the psoriasis-associated CD161 receptor gene KLRB1 (17) show psoriasis-specific elevation in the logFC 0.5 range, but Log10 dispersion scores of greater than -1, likely contributing to their failure to reach statistical significance in comparison to healthy control skin-residency T cells (annotated in blue in Figure 2A, data in Supplementary Table 3).
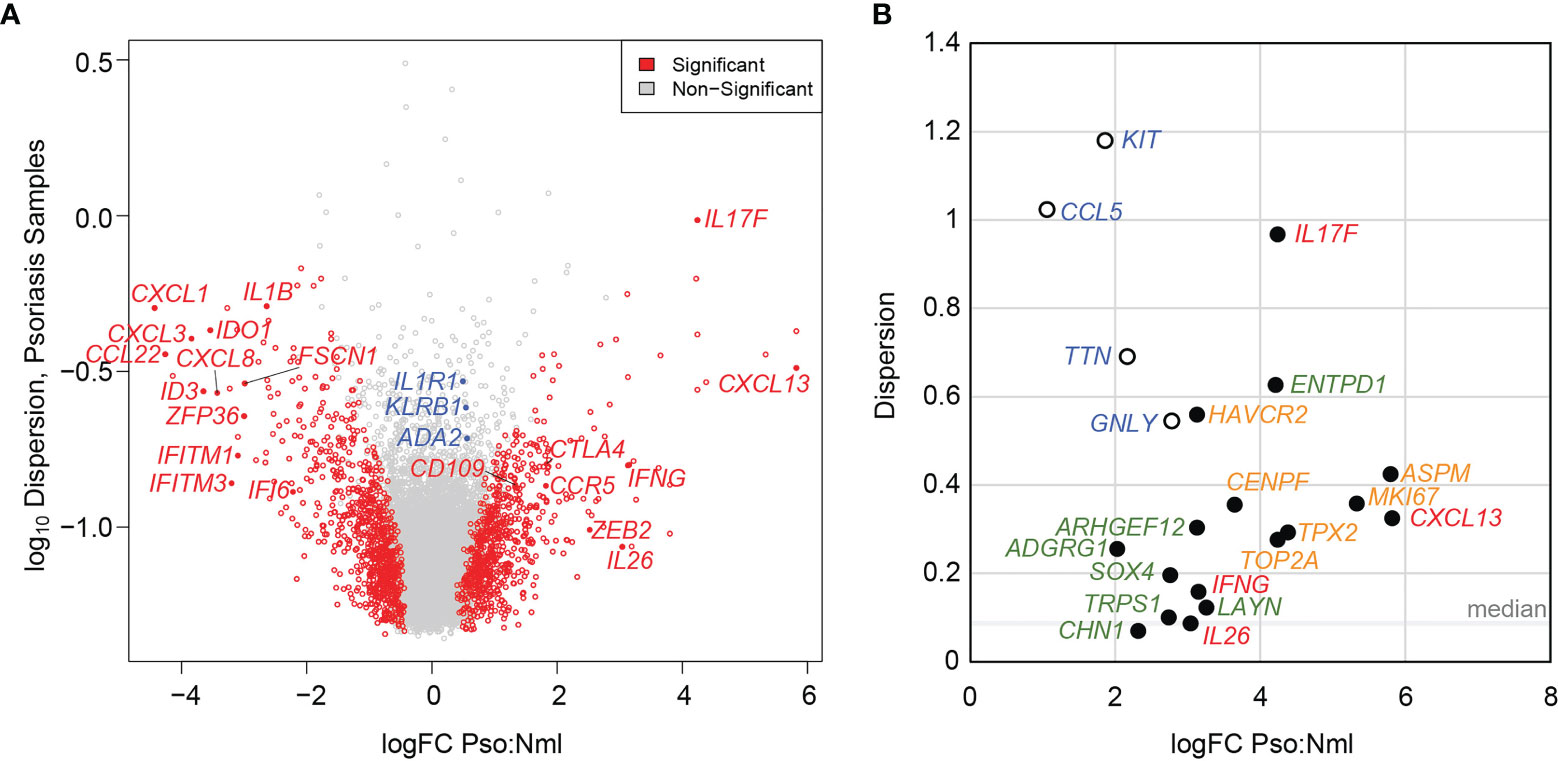
Figure 2 Elevated patient level variance in psoriasis-specific skin-resident memory T cell (Trm2) DEGs. (A) Volcano plot showing psoriasis DEGs identified using a pseudo-bulk approach charted as a function of logFC difference from normal, uninflamed cells (x-axis) and the log of the dispersion score (a proxy for patient-level variation, y-axis). Significant DEGs are shown in red, non-significant DEGs in grey. Labelled in blue are immune activation genes with relatively high dispersion scores, which may have prevented them from reaching statistical significance. (B) LogFC (x-axis) and dispersion score (y-axis) shown for established pathogenic psoriatic cytokines (red), mitotic cell division transcripts (green), psoriasis-specific abnormalities not elevated in atopic dermatitis (orange), and as in (A), immunologically activating DEGs with high end dispersion scores (blue).
We more closely examined inter-individual variance in psoriasis DEGs that were identified in the prior analysis as elevated not only relative to normal controls, but also to atopic dermatitis samples, indicating greater disease-specificity (8). Notably, many of these genes showed dispersion scores greater than the median of 0.086 (Figure 2B). In fact, for the six psoriasis DEGs with a logFC > 3 and significantly elevated compared to atopic dermatitis, the average dispersion score was 0.484 with a standard deviation of 0.273. In addition to the cytokines noted above such as IL17F (0.967), and CXCL13 (0.325), this set contained identified psoriasis-specific genes with less established functional roles, such as ARHGEF12 (0.303), ENTPD1 (0.628), LAYN (0.122) and HAVCR2 (0.560).
Cell cycle transcripts, which are elevated in both psoriasis and atopic dermatitis Trm2 compared to healthy controls, also show higher than median dispersion scores, including MKI67 (0.358), TOP2A (0.276), and CENPF (0.356). Similar to Figure 2A, Figure 2B displays examples of psoriasis-implicated genes whose expression is elevated in Trm2, but whose high inter-individual variance reduces their overall significance level (i.e. KIT, CCL5, TTN, and GNLY, blue, open circles).
We were next curious to understand if expression of psoriasis-specific immune DEGs correlated with clinical features such as PASI score. Such relationships might further narrow the search for genetic factors influencing clinical heterogeneity in psoriasis. We chose 16 established immune activation genes from our Trm2 DEGs including IL17F, CXCL13, IL26, CCR5, and CD82 (Supplementary Table 4) and quantified the T cells in each sample that detectably expressed each. We normalized these cell numbers between patients by bioinformatically deducing the total number of such cells existing in each sample, based on the total number of CD45+ cells obtained from each biopsy, as well as the total number of scRNA-seq profiled cells processed in Seurat (Materials and Methods, Supplementary Table 4). To generate accompanying measures of clinical severity, we reasoned that the phenotype of a biopsied and molecularly profiled lesion would be best represented by summing its individual Erythema, Induration, and Desquamation PASI descriptors, rather than the overall patient score, and derived such a lesion-specific severity score for each sample (Supplementary Table 1).
We then assessed Spearman correlation of T cell expression of all 16 immune cell DEGs with lesion-specific severity score. Three of these genes correlated strongly with lesion-specific scores: a single gene coefficient of 0.851 for CXCL13, and IKZF4 (p = 7.3 x 10-3) and 0.801 for CD84 (p = 1.7 x 10-2) (Bonferroni unadjusted, eight selected genes displayed in Figure 3; Supplementary Table 5). When the patient-level PASI score was used as an alternative comparator, none of the 16 immune genes showed significant correlations at unadjusted p values.
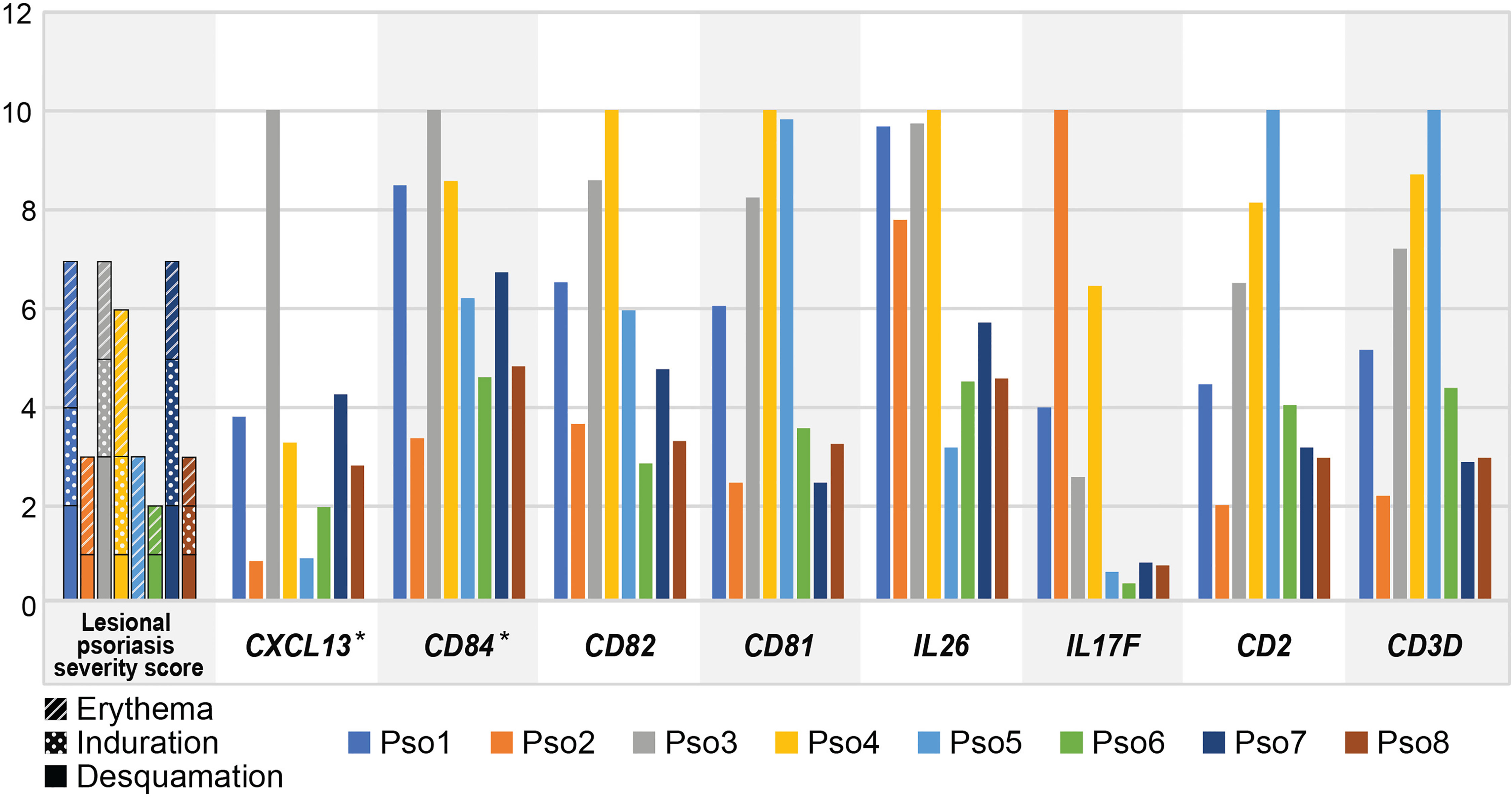
Figure 3 Inter-individual variation in lesional psoriasis severity score parallels that of CXCL13 and CD84. Leftmost graph shows severity scores for biopsied lesion for each patient. Subsequent graphs display deduced number of cells with positive expression for each gene, as a percentage of the maximum number of positive cells in any sample, multiplied by 1 x 103 (i.e. normalized to 10). Significant correlations for CXCL13 and CD84 are denoted by asterisks.
We next asked how functionally related groups of genes activated during psoriatic inflammation might vary in expression from patient to patient. We first applied the Metascape analysis package to detect overrepresentation of Gene Ontology and Reactome functional categories in the 662 genes significantly elevated (logFC > 0.4) in psoriatic skin-resident memory cells (Trm2), compared to healthy, controls, identifying 316 functional categories with a log (q value) < -2 (Supplementary Table 6; Supplementary Data 2). Statistically significant functional classes, included expected categories such as mitotic cell division and leukocyte activation (21 members, log (q value) < -9.97), but also highlighted the role of cytoskeletal reorganization (CDC42 signaling) and chromatin remodeling (Figure 4A). For example, ARHGEF12 selectively regulates RhoA subfamily GTPases to coordinate cell migration and invasion (19), while PAK2 influences actin cytoskeleton reorganization (20). DOCK8 deficiencies impair immune cell migration in both the innate and adaptive immune system (21). Changes in psoriatic Trm also include elevated transcripts levels of the linker histone H1FX (22), histone chaperone NAP1L4 (23), and the chromatin-modifying enzyme SMARCA5 (24). Figure 4B globally displays psoriasis Trm2 abnormalities in these four programs on a per-patient level.
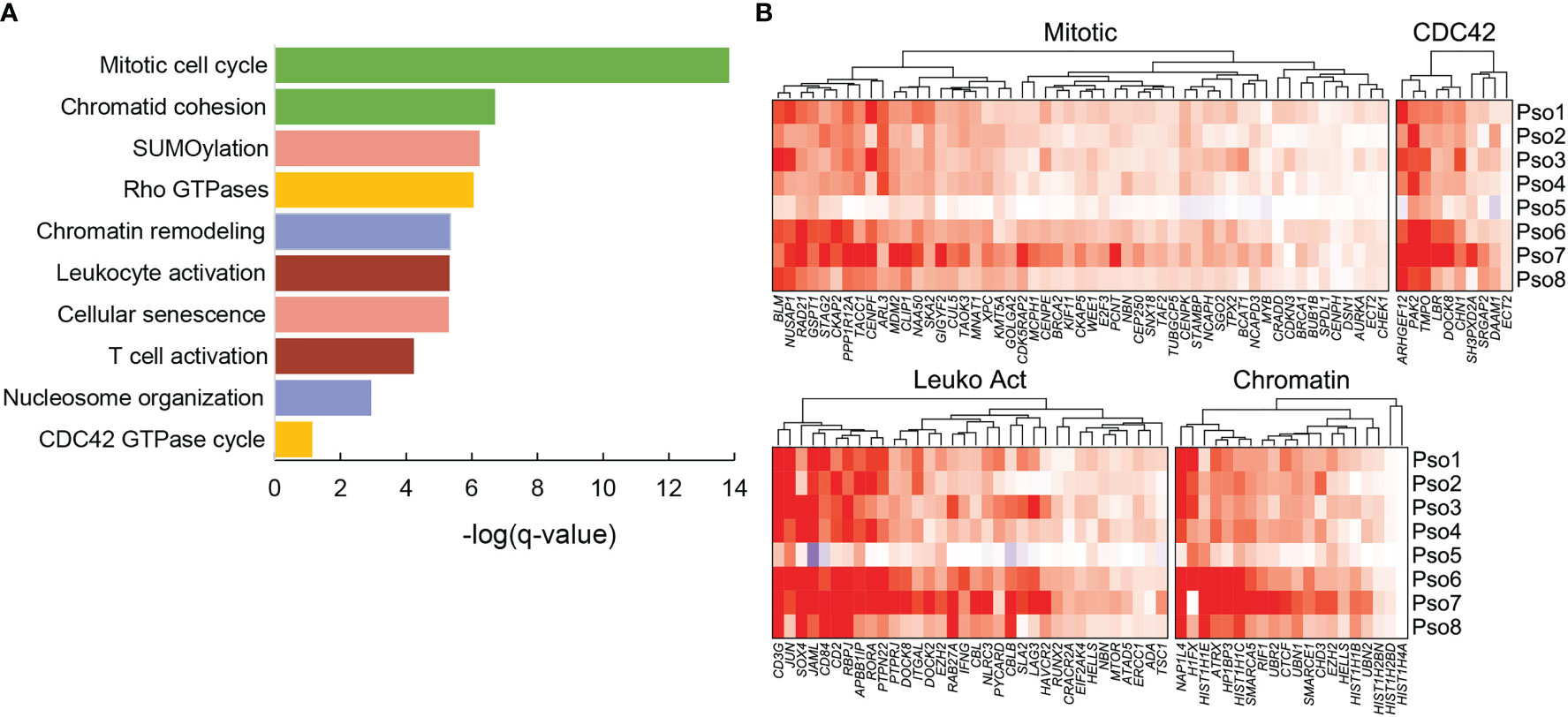
Figure 4 Significant functional associations for the 662 genes significantly elevated in psoriasis samples compared to grouped healthy controls in Trm2. (A) Ten example classifications are shown, with functions such as immune cell activation, mitotic cell division, and cytoskeletal reorganization. (B) Heatmaps visually represent average log2FC between individual psoriasis samples and normal controls using ComplexHeatmap (18). Heterogeneity is detected between patients, most prominently the dampened amplitude of transcript abnormalities in Patient 5, who was on systemic tacrolimus and mycophenolate at time of biopsy.
Considerable patient-specific fluctuations in these functionally related gene sets were easily appreciable. Most obviously, patient 5, the lone patient with psoriasis who was under systemic immunosuppression (mycophenolate and cyclosporine for a liver transplant) showed substantial attenuation of all these programs, corresponding to the lowest lesional psoriasis severity score (Supplementary Table 1). We sought to systematically assess these correlations between expression and phenotype, first averaging transcriptional log2FC for all genes in each of the four individual functional programs. Average scores for all four programs showed positive correlation with lesional severity score: CDC42 cytoskeletal reorganization at a Spearman rho value of 0.57, cell division at 0.55, chromatin reorganization at 0.48, and leukocyte activation at 0.36. None of these associations reached statistical significance, likely a factor of our limited sample size. However, in each pathway, activation parallels severity of individual lesions, revealing a potential source of some proportion of clinical heterogeneity.
While pervasive elevation of transcripts regulating mitosis or CDC42-centric functional reorganization coincided with induction of pathogenic cytokines in the Trm compartment, we were uncertain whether these programs were related on the single-cell level. In one model, members of these programs might simply be stochastically elevated in any given, pathogenically IL23-polarized single T cell. Alternatively, we hypothesized that some of these transcriptional programs could be shared on the single-cell level, a pattern that could impact approaches to therapeutic targeting. For example, if the single T cells most likely to express Th17 cytokines also showed robust reprogramming of cytoskeleton genes, strategies restraining actin reorganization might impede the mobility and infiltration of the most pathogenic skin-resident T cells.
We therefore calculated the Pearson correlation coefficients for expression of pathogenic cytokines in single Trm2 cells against those of genes in our cytoskeletal and secretory classes, finding striking instances of both positive and negative correlation (Figure 5). For example, Figure 5A shows positive correlation of the RORA transcription factor with IL17F expression, as would be expected given its role in Th17 programming (R = 0.3, p = 2.2 x 10-16) (25), as well as for the TCR component CD3G (R = 0.29, p = 2.2 x 10-16). Similarly, in Figure 5B, the cytoskeletal re-organization genes PAK2 and APBB1IP robustly positively correlate with IL17F expression in single skin-resident memory T cells, supporting a model in which more highly pathogenically activated cells are also more motile and capable of tissue infiltration. In sharp contrast, the single cells expressing maximum IL17F and those expressing elevated levels of a number of chromatin-modifying transcripts are negatively correlated, for example, a R of -0.30 for HIST1H1E (Figure 5C). Such instances of mutual exclusivity suggest the presence of a second, abnormal, non-Th17 population within psoriatic Trm, whose influence on disease state is yet undetermined. A comprehensive single-cell correlation table in Trm2 for IL17F, CXCL13, and IL26 is available in Supplementary Table 7.
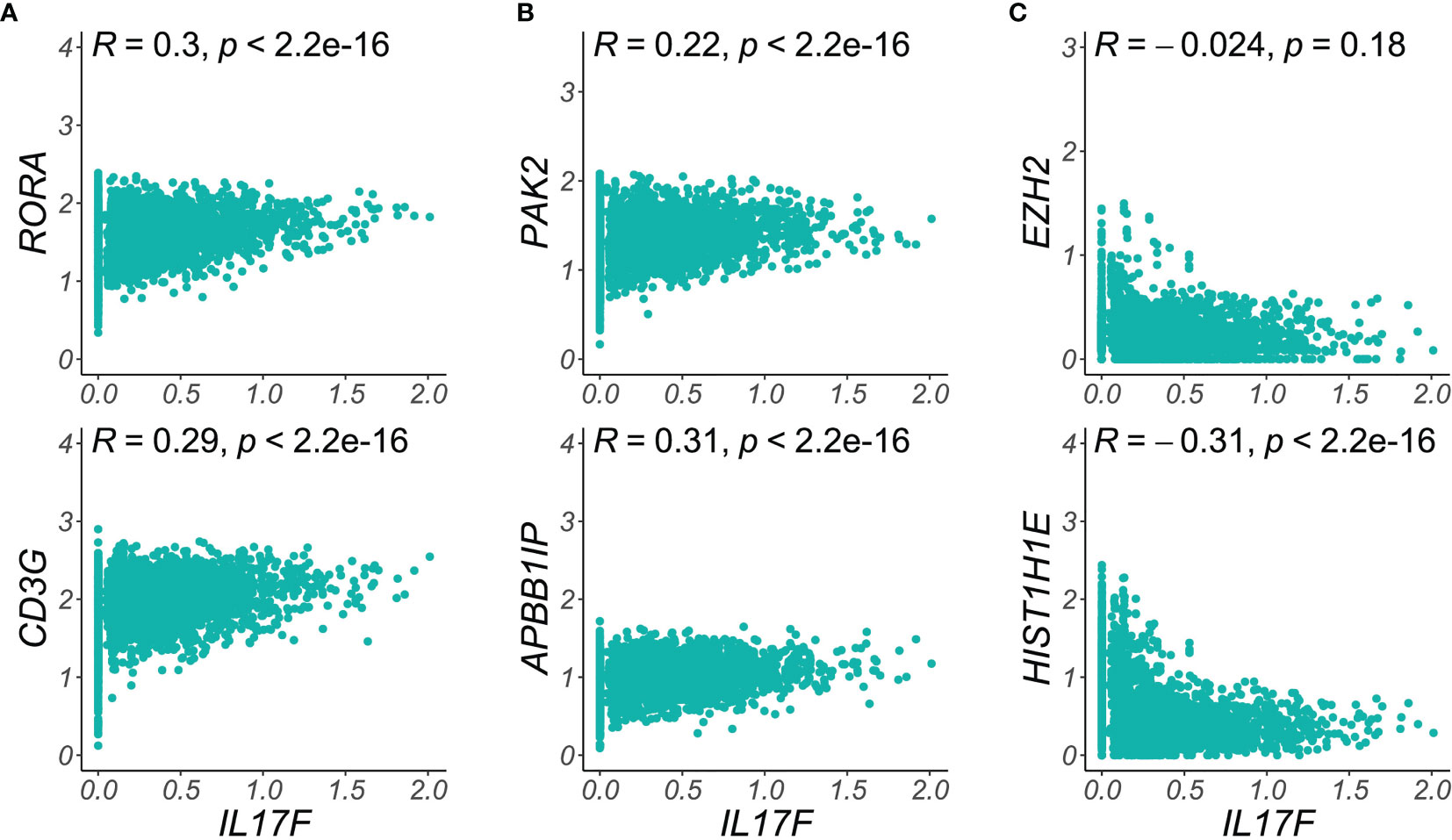
Figure 5 Single-cell correlations and anti-correlations between functional class transcripts and IL17F expression. (A) T cell activation markers like RORA and CD3G are elevated in the highest IL17F expressing cells, (B) Key cytoskeletal reorganization transcripts (PAK2, APBB1IB) are most elevated in the single skin-resident T cells expressing maximal psoriatic inflammatory mediators. (C) Chromatin remodeling transcripts (EZH2, HIST1H1E), are elevated in the lowest IL17F expression cells, suggesting a distinct, pathologic cell population in psoriatic Trm. Density plots show imputed single cell expression of T cell activation, chromatin remodeling, or cytoskeletal transcripts (y-axis) vs. IL17F (x-axis). Dots represent single Trm2 cells (psoriasis samples).
While a vast landscape of transcriptional abnormalities in immune and stromal cell types characterizes chronic inflammatory skin disease (26, 27), clinical improvement following inhibition of the IL12/23 pathway or blockade of IL17 isoforms validates the central role of psoriatic T cells. Our single-cell profiles of 8 psoriasis samples, along with normal controls, begin to illuminate patient-specific variation of transcriptional abnormalities in psoriatic Trm (Figure 2). Psoriasis DEGs with greater than average patient-level variance, reflected in higher dispersion scores, include the most recognizable Th17 cytokines such as IL17F and IL26, recently implicated inflammatory psoriatic mediators such as CXCL13 (13), and genes orchestrating cell division in mitotically active T cells. Such psoriasis DEGs with higher dispersion scores may represent sources of patient-specific phenotypic and clinical variability, such as lesion intensity or anatomic distribution.
Expression of CXCL13 and CD84 correlated significantly with lesional severity score in our study, a predicate for further investigation as sources or important associations of disease state. Our data adds to increasing evidence that CXCL13 represents a particularly Th17-specific abnormality (8) and positively associates with psoriasis severity (13, 28), nominating it as a clinically useful biomarker for cutaneous disease. CD84 is a known T cell activation marker, genetic variants of which have been associated with response in psoriasis to TNF blockade (29). Interestingly, IL17F expression in our series correlated poorly with lesional severity score but was highly elevated in scalp psoriasis, suggesting it might show anatomic specificity in more highly powered studies. This finding comports with an earlier scRNA-seq report that Th17 cytokine expression and overall inflammatory state is surprisingly prominent in healthy scalp cells (27). The key constraint of our study is patient number, limited by the current costs of scRNA-seq. It is very likely additional such correlations will reach significance as these approaches are extended to larger data sets.
Conversely, psoriasis-specific DEGs harboring lower dispersion scores may be more suitable for broader screening to identify psoriasis-like molecular profiles, a feature that may help direct biological treatment for the subset of rashes demonstrating both psoriasiform and spongiotic histopathology (30). Within the set of psoriasis-specific skin-resident DEGs that are overexpressed relative to analogous T cells in atopic dermatitis, examples of such lower variance Th17 biomarkers include the GTPase-activator CHN1 (0.069) and PTMS (0.077).
We also undertook a systematic search of coordinated functional derangements in skin-resident T cells, based on the increased resolution afforded by single-cell transcriptomics. Such groups of pathologic transcriptional alternations may function as quantitative traits, collectively modifying disease phenotype beyond the impact of dysregulated single genes. Applying this method, we detected not only expected elevations in inflammatory signalling and cell division, but also global increases in pathways coordinating CDC42-centric cytoskeletal reorganization and chromatin remodeling. In one sense, broad alterations in these programs are not surprising, given the profound changes in cell polarity and motility that accompany T cell activation. However, this is the first report describing recurrent upregulation of dozens of these transcripts in pathologically inflamed T cells. All 8 patients in our series show abnormalities in these programs (Figure 4), whose elevation trends with lesional psoriasis severity scores, supporting a role in the pathogenicity of skin inflammation.
We also show that single T cells expressing the highest levels of psoriatic inflammatory mediators such as IL17F are markedly enriched for cytoskeletal remodeling transcripts, suggesting such programs may facilitate tissue infiltration and cytokine secretion. Combination therapeutic approaches targeting both Th17 polarization and cytoskeletal activity may thus synergistically target a common population of particularly pathogenic skin T cells. We also find that certain chromatin remodeling DEGs peak in single T cells distinct from those maximally expressing IL17F, indicating these data can also identify additional, abnormally reprogrammed subpopulations within the Trm compartment.
In summary, the analyses presented here describe a suite of quantitative approaches to evaluate high-resolution transcriptional variation between psoriasis patients. The most distinguishing abnormalities are identified in skin-resident T cells, and even our limited test set identifies credible associations between specific genes and lesion phenotype. Greater numbers of scRNA-seq datasets are now becoming publicly accessible. Systematic identification of such instances of inter-individual molecular heterogeneity will make it possible to test clinically predictive associations for both single genes and aggregate molecular disease signatures.
Sequencing BAM files are deposited at the European Genome-Phenome Archive (EGA) under accession number EGAS00001005271. The processed Seurat object with cell identities is available at Zenodo (https://zenodo.org/record/6529821#.Ynh_aoxBwuW).
The studies involving human participants were reviewed and approved by University of California, San Francisco Institutional Review Board. The patients/participants provided their written informed consent to participate in this study.
YL, JC, EP, and RC designed the study. EK, JC, and RC supervised sample collection and processing. YL, CC, AH, and AS performed sample preparation and analysis. HW, MT, CC, YS, and EP performed computational analysis. JN performed histopathology analysis. YL, HW, EP, JC, and RC wrote the original manuscript with contributions from CC, JN, and AH. All authors contributed to the article and approved the submitted version.
This study received funding from Sun Pharmaceutical Industries. The funder was not involved in the study design, collection, analysis, and interpretation of data, the writing of this article or the decision to submit it for publication. JC and RC are funded in part by grants from the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health K08AR067243, the National Psoriasis Foundation, and a LEO foundation grant.
JC and RC receive support (research grants to their institution) from LEO Pharmaceuticals, Sanofi, and Pfizer.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge Rachel Sevey for essential assistance with graphic representation.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.842651/full#supplementary-material
Supplementary Table 1 | Clinical Characteristics of Psoriasis Patients.
Supplementary Table 2 | Cell Type Marker Transcripts (Seurat-derived).
Supplementary Table 3 | Trm2 Pseudo-bulk DEGs Normal Controls vs. Psoriasis.
Supplementary Table 4 | Deduced Cell Number Expression for each Gene.
Supplementary Table 5 | Correlation of Gene Expressing Cells with Lesional Psoriasis Severity Scores.
Supplementary Table 6 | Metascape Functional Analysis of Trm2 Psoriasis DEGs.
Supplementary Table 7 | Single-cell gene correlations (non-imputed) in Trm2 for IL17F, CXCL13, and IL26.
Supplementary Data Sheet 1 | uMAP representation of donors, showing high representation for each sample in key immune cell classes.
Supplementary Data Sheet 2 | Box plots of selected genes from Trm2 DEG enriched functional classes on a per-sample basis.
1. Bugaut H, Aractingi S. Major Role of the IL17/23 Axis in Psoriasis Supports the Development of New Targeted Therapies. Front Immunol (2021) 12:621956. doi: 10.3389/fimmu.2021.621956
2. Itoh T, Hatano R, Komiya E, Otsuka H, Narita Y, Aune TM, et al. Biological Effects of IL-26 on T Cell-Mediated Skin Inflammation, Including Psoriasis. J Invest Dermatol (2019) 139(4):878–89. doi: 10.1016/j.jid.2018.09.037
3. Jordan CT, Cao L, Roberson EDO, Duan S, Helms CA, Nair RP, et al. Rare and Common Variants in CARD14, Encoding an Epidermal Regulator of NF-kappaB, in Psoriasis. Am J Hum Genet (2012) 90(5):796–808. doi: 10.1016/j.ajhg.2012.03.013
4. Li B, Tsoi LC, Swindell WR, Gudjonsson JE, Tejasvi T, Johnston A, et al. Transcriptome Analysis of Psoriasis in a Large Case-Control Sample: RNA-seq Provides Insights Into Disease Mechanisms. J Invest Dermatol (2014) 134(7):1828–38. doi: 10.1038/jid.2014.28
5. Keermann M, Kõks S, Reimann E, Prans E, Abram K, Kingo K. Transcriptional Landscape of Psoriasis Identifies the Involvement of IL36 and IL36RN. BMC Genomics (2015) 16(1):322. doi: 10.1186/s12864-015-1508-2
6. Visvanathan S, Baum P, Vinisko R, Schmid R, Flack M, Lalovic B, et al. Psoriatic Skin Molecular and Histopathologic Profiles After Treatment With Risankizumab Versus Ustekinumab. J Allergy Clin Immunol (2019) 143(6):2158–69. doi: 10.1016/j.jaci.2018.11.042
7. Stoeckius M, Hafemeister C, Stephenson W, Houck-Loomis B, Chattopadhyay PK, Swerdlow H, et al. Simultaneous Epitope and Transcriptome Measurement in Single Cells. Nat Methods (2017), 14(9):865–8. doi: 10.1038/nmeth.4380.
8. Liu Y, Wang H, Taylor MA, Cook C, Martinez-Berdeja A, North JP, et al. Classification of Human Chronic Inflammatory Skin Disease Based on Immune Single-Cell Profiling. Sci Immunol.
9. Crowell HL, Soneson C, Germain P-L, Calini D, Collin L, Raposo C, et al. Muscat Detects Subpopulation-Specific State Transitions From Multi-Sample Multi-Condition Single-Cell Transcriptomics Data. Nat Commun (2020) 11(1):6077. doi: 10.1038/s41467-020-19894-4
10. edgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data (2022). Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2796818/.
11. Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, et al. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat Commun (2019) 10(1):1523. doi: 10.1038/s41467-019-09234-6
12. Metascape (2021). Available at: https://metascape.org/gp/index.html#/main/step1.
13. Liu J, Chang H-W, Huang Z-M, Nakamura M, Sekhon S, Ahn R, et al. Single-Cell RNA Sequencing of Psoriatic Skin Identifies Pathogenic Tc17 Cell Subsets and Reveals Distinctions Between CD8+ T Cells in Autoimmunity and Cancer. J Allergy Clin Immunol (2021) 147(6):2370–80. doi: 10.1016/j.jaci.2020.11.028
14. estimateTagwiseDisp Function - Rdocumentation (2022). Available at: https://www.rdocumentation.org/packages/edgeR/versions/3.14.0/topics/estimateTagwiseDisp.
15. Mahil SK, Catapano M, Di Meglio P, Dand N, Ahlfors H, Carr IM, et al. An Analysis of IL-36 Signature Genes and Individuals With IL1RL2 Knockout Mutations Validates IL-36 as a Psoriasis Therapeutic Target. Sci Transl Med (2017) 9(411):eaan2514. doi: 10.1126/scitranslmed.aan2514
16. Yap JY, Moens L, Lin M-W, Kane A, Kelleher A, Toong C, et al. Intrinsic Defects in B Cell Development and Differentiation, T Cell Exhaustion and Altered Unconventional T Cell Generation Characterize Human Adenosine Deaminase Type 2 Deficiency. J Clin Immunol (2021) 41(8):1915–35. doi: 10.1007/s10875-021-01141-0
17. Cosmi L, De Palma R, Santarlasci V, Maggi L, Capone M, Frosali F, et al. Human Interleukin 17-Producing Cells Originate From a CD161+CD4+ T Cell Precursor. J Exp Med (2008) 205(8):1903–16. doi: 10.1084/jem.20080397
18. Gu Z, Eils R, Schlesner M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinf (2016) 32(18):2847–9. doi: 10.1093/bioinformatics/btw313
19. Ghanem NZ, Matter ML, Ramos JW. Regulation of Leukaemia Associated Rho Gef (Larg/Arhgef12). Small GTPases (2021) 1–9. doi: 10.1080/21541248.2021.1951590
20. Wang Y, Zeng C, Li J, Zhou Z, Ju X, Xia S, et alPak2 Haploinsufficiency Results in Synaptic Cytoskeleton Impairment and Autism-Related Behavior. (2022).
21. Biggs CM, Keles S, Chatila TA. DOCK8 Deficiency: Insights Into Pathophysiology, Clinical Features and Management. Clin Immunol (2017) 181:75–82. doi: 10.1016/j.clim.2017.06.003
22. Ichihara-Tanaka K, Kadomatsu K, Kishida S. Temporally and Spatially Regulated Expression of the Linker Histone H1fx During Mouse Development. J Histochem Cytochem (2017) 65(9):513–30. doi: 10.1369/0022155417723914
23. Rodriguez P, Munroe D, Prawitt D, Chu LL, Bric E, Kim J, et al. Functional Characterization of Human Nucleosome Assembly Protein-2 (NAP1L4) Suggests a Role as a Histone Chaperone. Genomics (1997) 44(3):253–65. doi: 10.1006/geno.1997.4868
24. Ding Y, Li Y, Zhao Z, Cliff Zhang Q, Liu F. The Chromatin-Remodeling Enzyme Smarca5 Regulates Erythrocyte Aggregation via Keap1-Nrf2 Signaling. Elife (2021) 10:e72557. doi: 10.1101/2021.09.08.459391
25. Yang XO, Pappu B, Nurieva R, Akimzhanov A, Kang HS, Chung Y, et al. TH17 Lineage Differentiation Is Programmed by Orphan Nuclear Receptors Rorα and Rorγ. Immun (2008) 28(1):29–39. doi: 10.1016/j.immuni.2007.11.016
26. Reynolds G, Vegh P, Fletcher J, Poyner EFM, Stephenson E, Goh I, et al. Developmental Cell Programs Are Co-Opted in Inflammatory Skin Disease. Science (2021) 371(6527):eaba6500. doi: 10.1126/science.aba6500
27. Cheng JB, Sedgewick AJ, Finnegan AI, Harirchian P, Lee J, Kwon S, et al. Transcriptional Programming of Normal and Inflamed Human Epidermis at Single-Cell Resolution. Cell Rep (2018) 25(4):871–83. doi: 10.1016/j.celrep.2018.09.006
28. Liu W, Zhou X, Wang A, Ma J, Bai Y. Increased Peripheral Helper T Cells Type 17 Subset Correlates With the Severity of Psoriasis Vulgaris. Immunol Lett (2021) 229:48–54. doi: 10.1016/j.imlet.2020.11.005
29. van den Reek JMPA, Coenen MJH, van de L’Isle Arias M, Zweegers J, Rodijk-Olthuis D, Schalkwijk J, et al. Polymorphisms in CD84, IL12B and TNFAIP3 Are Associated With Response to Biologics in Patients With Psoriasis. Br J Dermatol (2017) 176(5):1288–96. doi: 10.1111/bjd.15005
30. Cohen JN, Bowman S, Laszik ZG, North JP. Clinicopathologic Overlap of Psoriasis, Eczema, and Psoriasiform Dermatoses: A Retrospective Study of T Helper Type 2 and 17 Subsets, Interleukin 36, and β-Defensin 2 in Spongiotic Psoriasiform Dermatitis, Sebopsoriasis, and Tumor Necrosis Factor α Inhibitor-Associated Dermatitis. J Am Acad Dermatol (2020) 82(2):430–9. doi: 10.1016/j.jaad.2019.08.023
Keywords: single-cell RNA-sequencing, psoriasis vulgaris, heterogeneity, cytoskeleton, chromatin
Citation: Liu Y, Wang H, Cook C, Taylor MA, North JP, Hailer A, Shou Y, Sadik A, Kim E, Purdom E, Cheng JB and Cho RJ (2022) Defining Patient-Level Molecular Heterogeneity in Psoriasis Vulgaris Based on Single-Cell Transcriptomics. Front. Immunol. 13:842651. doi: 10.3389/fimmu.2022.842651
Received: 24 December 2021; Accepted: 19 April 2022;
Published: 26 July 2022.
Edited by:
Xu-jie Zhou, Peking University, ChinaReviewed by:
Anna Cogen, University of Washington, United StatesCopyright © 2022 Liu, Wang, Cook, Taylor, North, Hailer, Shou, Sadik, Kim, Purdom, Cheng and Cho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raymond J. Cho, cmF5bW9uZC5jaG9AdWNzZi5lZHU=; Jeffrey B. Cheng, amVmZnJleS5jaGVuZ0B1Y3NmLmVkdQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.