
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 15 February 2022
Sec. Viral Immunology
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.841459
This article is part of the Research TopicHost-microbe Interaction in SARS-CoV-2 Infection: Mechanism and InterventionView all 14 articles
 Susana Santos1,2*
Susana Santos1,2* Pedro Barata3,4
Pedro Barata3,4 Adilia Charmier1,2Inês Lehmann1Suzilaine Rodrigues1Matteo M. Melosini1
Adilia Charmier1,2Inês Lehmann1Suzilaine Rodrigues1Matteo M. Melosini1 Patrick J. Pais3,4
Patrick J. Pais3,4 André P. Sousa3,4,5Catarina Teixeira3,4,5Inês Santos3,4,5
André P. Sousa3,4,5Catarina Teixeira3,4,5Inês Santos3,4,5 Ana Catarina Rocha3,4Pilar Baylina3,4,5
Ana Catarina Rocha3,4Pilar Baylina3,4,5 Ruben Fernandes3,4,5
Ruben Fernandes3,4,5In late 2019, COVID-19 emerged in Wuhan, China. Currently, it is an ongoing global health threat stressing the need for therapeutic compounds. Linking the virus life cycle and its interaction with cell receptors and internal cellular machinery is key to developing therapies based on the control of infectivity and inflammation. In this framework, we evaluate the combination of cannabidiol (CBD), as an anti-inflammatory molecule, and terpenes, by their anti-microbiological properties, in reducing SARS-CoV-2 infectivity. Our group settled six formulations combining CBD and terpenes purified from Cannabis sativa L, Origanum vulgare, and Thymus mastichina. The formulations were analyzed by HPLC and GC-MS and evaluated for virucide and antiviral potential by in vitro studies in alveolar basal epithelial, colon, kidney, and keratinocyte human cell lines.
Conclusions and Impact: We demonstrate the virucide effectiveness of CBD and terpene-based formulations. F2TC reduces the infectivity by 17%, 24%, and 99% for CaCo-2, HaCat, and A549, respectively, and F1TC by 43%, 37%, and 29% for Hek293T, HaCaT, and Caco-2, respectively. To the best of our knowledge, this is the first approach that tackles the combination of CBD with a specific group of terpenes against SARS-CoV-2 in different cell lines. The differential effectiveness of formulations according to the cell line can be relevant to understanding the pattern of virus infectivity and the host inflammation response, and lead to new therapeutic strategies.
Since the emergence of the SARS-CoV-2 outbreak, extensive efforts have been placed regarding antiviral research for compounds with effective antiviral or virucide activity. COVID-19 is a complex disease that afflicts respiratory and gastrointestinal tract and kidney function, being one of its main features the hyperstimulation of the immune system. The spectrum of medical therapies to treat COVID-19 is growing; however, there are no 100% effective therapeutic approaches for the prevention and treatment of COVID-19.
Although vaccination and preventive medications are recognized as the most effective means of combating a virus, the treatment of COVID-19 is a real challenge prompting the need for effective drugs (1). Natural compounds from medicinal plants, such as terpenes, have gained attention as potential inhibitors of coronaviruses being the possible mechanism in the inhibition of viral replication or targeting viral proteins relevant for virus adsorption and entry (1–7). Essential oils (EOs) exhibit antiviral (4, 8–12), immunomodulatory (8, 13), and anti-inflammatory (8, 14, 15) properties, namely, regarding virus infection as influenza or herpes simplex viruses 1 or 2 (16, 17). EOs from Origanum acutidens (18), Artemisia glabella (19), eucalyptus and tea tree (20), Thymus vulgaris, Melaleuca ericifolia, M. leucadendron, and M. armillaris (21), among many others, have been described against those viruses. Wen et al. (22) reported EO constituents inducing a cytopathogenic effect against SARS-CoV on Vero-E6 cells. From molecular modeling studies, several terpenoids were potent inhibitors of SARS-CoV-2 replication (23). Among medicinal plants, Origanum vulgare has been recognized for its potential therapeutic role mainly arising from terpenes and flavonoids (24, 25) and Thymus vulgaris EOs have been shown to be effective against several RNA viruses including coronaviruses (21, 26). Regarding Cannabis sativa, it is particularly rich in terpenes, typically monoterpenes and sesquiterpenes (27–30). This plant is mostly known for containing phytocannabinoids, mainly Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD), those widely accessed being medicinal compounds with known applications in several conditions, most of them related to inflammation processes (30–36). Phytocannabinoids are a group of terpenophenolic compounds with biological activities through interaction with the endocannabinoid system (ECS) in humans. CBD is a partial agonist for cannabinoid receptor 2 (CN2R), widely expressed in the immune system (37–39). In a mouse model for a respiratory syncytial virus (RSV) infection, CN2R activation reduced the signs of infection by modulating the immune system (10). One study demonstrates that a genetic polymorphism in CN2R, which reduces ECS-induced response (40), is associated with an increased risk of hospitalization in young children infected with RSV (n = 83), with up to 3-fold increased risk of developing severe acute respiratory tract infection. Rossi et al. (41) hypothesize that CN2R can be a therapeutic target for SARS-CoV-2 since its stimulation limits the release of pro-inflammatory cytokines, shifts the macrophage phenotype toward the anti-inflammatory M2 phenotype, and enhances the immune-modulating properties of mesenchymal stromal cells. The anti-inflammatory properties of CBD have been explored as antiviral agents for the treatment of HIV (42), influenza (43, 44), and most recently SARS-CoV-2 (45–48). CN2R increases as HIV infection progresses, and on infected macrophages, the exposure to CN2R to a selective agonist resulted in a dose-dependent decrease in reverse transcriptase activity/viral replication activity (17). Recently, researchers have tested CBD on 3D human models of oral, airways, and intestinal tissues and found that low THC/high CBD cultivars modulate ACE2 and TMPRSS2 levels, which might lower the virus load (49). By its turn, another study demonstrated that CBD reduced the secretion of pro-inflammatory cytokines IL-6, IL-8, CCL2, and CCL7 from the alveolar epithelial cell line A549 (50).
A similar study to the one proposed in this work was executed by Chatow et al. (51) who demonstrated an antiviral activity of a terpene formulation (30 terpenes) against HCoV-229E in human lung fibroblasts and the antiviral action during the viral multiplication cycle, in which the combination of the CBD with terpenes potentiated the antiviral effect. In another study, CBD exerted prolonged immunosuppression and hence might be used in chronic inflammation, and the terpenoids showed transient immunosuppression and might thus be used to relieve acute inflammation (52). Since terpenes are known to act as enhancing phytocannabinoid action (53–55), it is intended to query the action of a group of specific terpenes as virucide or antiviral agents acting in entourage effect with their selves and with the CBD as a therapeutic agent modulating inflammation. In detail, to exploit the biological action of the formulations on virus infectivity, it is a goal to understand their potential effect to act i) as a virucide agent blocking and inactivating the virus at early stages of infection, ii) as an antiviral agent blocking the virus cellular machinery, iii) as an agent against an overactive immune-inflammatory cascade at later stages of infection. This could be relevant in reversing the cytotoxic events induced by the virus and may contribute to the concept of ECS as a contributor for controlling the immune response from a virus infection.
CBD and terpenes identified from Cannabis sativa, Origanum vulgare subsp. virens, and Thymus mastichina provided the source for 6 proprietary formulations (F1T, F2T, F3T, F1TC, F2TC, F3TC). The CBD was purified from a Cannabis sativa distillate (FarmCeutica Wellness, Richmond, Canada). A purified CBD sample (>99.5%) was obtained by Centrifuge Partition Chromatography (CPC) technology using an rCPC device (RotaChrom, Purified Solutions, Budapest, Hungary). The method was internally developed through optimizing the best solvent combination and the solvent ratio (data to be published). Briefly, CPC is a liquid–liquid preparative chromatographic technique that makes use of two immiscible liquid phases, the solvent system, representing the stationary and mobile phases of a typical chromatographic apparatus. The target terpenes of EOs and CBD from the distillate were separated according to their partition coefficients. The main goal of this technology is to isolate specific compounds with a pharmaceutical purity grade (>99%) and high recovery mass yield (>95%). Relative to terpene purification, 3 groups of specific terpenes (19 in total) were purified from Origanum vulgare subsp. virens and Thymus mastichina (NBI-Natural Business Intelligence, Vila Real, Portugal) and from Cannabis sativa by combining a hydrodistillation clevenger with CPC (proprietary method). The formulations F1T, F2T, and F3T were prepared to contain a specific group of 7, 8, and 9 terpenes, respectively, comprising approximately 84% of terpene content and using MCT oil as carrier oil. The remaining terpenes were present at concentrations smaller than 2%. Moreover, CBD was added at a concentration of 1 µg/ml intending formulations F1TC, F2TC, and F3TC. Formulations and CBD samples were kept in dark amber glass flasks at room temperature. The EOs were obtained by the hydrodistillation clevenger, and the formulations comprising the terpenes were analyzed by gas chromatography-mass spectrometry (GC-MS, GC-MS-QP2020 NX Gas Chromatograph Mass Spectrometer, Shimadzu, Kyoto, Japan) (Figures S1A–C and Supplemental Information). The CBD isolate was analyzed by high-performance liquid chromatography (HPLC, Cannabis Analyzer™ for Potency, Shimadzu) (Figure S1D and Supplemental Information).
The Caco-2 cell line was cultured in MEM medium (VWR, Biowest, P0451-N10L, Riverside, MO, USA) supplemented with 20% of FBS (Gibco, Life Technologies, 10270, Grand Island, NY, USA) and 1% penicillin/streptomycin (Gibco, Life Technologies, 10270, USA). A549, HaCaT, and Hek293T cell lines were cultured in DMEM medium (VWR, Biowest, P0103-N10L, USA) supplemented with 10% of FBS and 1% penicillin/streptomycin. Cells were maintained at 37°C in a humidified chamber containing 5% CO2.
The B.1.1.7 strain of SARS-CoV-2, isolated in the laboratory, was clarified by centrifugation at 2,000 g for 15 min. The isolated virus was incubated in each cell line with 2% trypsin for 1 h, and then the cell culture was washed twice with PBS and incubated with complete cell culture medium.
Total RNA was extracted using Lab-Aid Virus RNA Extraction Kit (Zeesan, Xiamen, China). RNA purity was measured in a microdrop 16-well microplate spectrophotometer (Thermo Scientific™ Multiskan SkyHigh Microplate Spectrophotometer, Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA). Virus titer was determined by SARS-CoV-2 detection with Fosun COVID-19 RT-PCR Detection Kit (Fosun Pharma, Shanghai, China) and quantified using a calibration curve with Synthetic SARS-CoV-2 RNA SARS-CoV-2 positive control (SARS-CoV-2 Positive Control, Twist Synthetic, China).
The MTT assay (Life Technologies, Thermo Fisher Scientific, USA) was carried out following the manufacturer’s instruction. In brief, 1 × 105 cells/well were seeded and grown until 80% confluence. Each formulation and its isolated components were incubated with and without the predetermined virus titer for 24 h in cell culture. After the washing step with warm PBS, incomplete cell culture medium was added along with 0.5 mg/ml of MTT and incubated for 2 h at 37°C. The absorbance was measured using the microplate reader at 570 nm. Results were performed in triplicates and normalized to the control considered to be 100%.
Two different treatment approaches were employed: 1) treatment incubation of 24 h, followed by a rinsing step with warm PBS, and then the SARS-CoV-2 infection was executed for another 24 h; 2) incubation of SARS-CoV-2 for 24 h followed by a rinsing step with warm PBS and then the treatment incubation was employed during 24 h. Cell culture supernatant was harvested and submitted to a 3.2.3 process. The non-cytotoxic concentrations of the compounds and formulations were determined as the concentrations that did not lead to more than 50% cell death, as compared to untreated cells.
Four sets of primers and probes were designed based on the genome of SARS-CoV-2 (GenBank accession no. MN908947.3) using Primer Express Software (version 3.0.1 Applied Biosystems, Foster, CA, USA). The used primers and probes are identified in Table 1.

Table 1 Set of primer and probe sequences for the one-step multiplex RT-qPCR.
Gene expression was estimated measuring the mRNA from cell extraction by RT-qPCR with qTOWER (3) (Analytik Jena, Germany), using One-step NZYSpeedy RT-qPCR Probe Kit, ROX (NZYTech, Lisbon, Portugal). 10 ng/µl RNA was employed, and the threshold cycle (CT) values from each biological assay were plotted with two experimental replicates following the manufacturer’s procedure. Melting curve analysis was used to monitor the specificity of primers and probes. Results were normalized to the GAPDH housekeeping gene, and gene relative expression was employed by the ΔCT expression/ΔCT negative control ratio.
All experiments were performed in triplicate and normalized to a negative control. Statistical analysis was performed comparing the control group results with those of the different groups with two-way analysis of variance (ANOVA) multiple-comparison and Dunnett post hoc tests, using GraphPad Prism 8.0.1 software (GraphPad Software, San Diego, CA, USA). Normality of data distribution was assessed using the Shapiro–Wilk test and for the homogeneity of variance with Bartlett’s test. Results were considered statistically significant whenever p-value < 0.05.
F1T, F2T, and F3T were established using a mixture of specific terpenes purified from Origanum virens, Thymus mastichina, and Cannabis sativa as the genera of those plants are recognized to contain terpenes with reported antimicrobial properties (18, 21, 24–30). The concentration was based on published data and in data from our previous work (results from ecotoxicological and cytotoxic assays to be published, POCI-01-02B7-FEDER-053456-BIOBLOCKCOVID). Moreover, we intended to study the CBD-terpenes’ entourage action (53–55), and so we used a lower concentration of F1T, F2T, and F3T when in combination with CBD. Despite the respiratory tract being the dominant route in SARS-CoV-2 infection, the colon, kidney, and skin comprise COVID-associated symptoms (56–66). Moreover, the colon, kidney, and skin tissues present very considerable levels of ACE2 and TMPRSS2 expression (data obtained from Human Protein Atlas available from http://www.proteinatlas.org, Uhlén M et al., Tissue-based map of the human proteome. Science (2015) PubMed: 25613900 DOI: 10.1126/science.1260419) and hence potential targets for virus infection. We performed the viral reduction assays in several cell lines and quantified ACE2 and TMPRSS2 gene expressions. Both receptors can be considered as targets for SARS-CoV-2, expecting that the coding genes are upregulated. By its turn, it is expected that RdRp and Spike gene expression is upregulated as viral infectivity progress and that gene expression diminishes because of a virucide or antiviral action. As CN2R activation could limit the release of pro-inflammatory cytokines (50) associated with COVID-19, and in this context being a potential therapeutic target for SARS-CoV-2, CBD was included in the formulations (F1TC, F2TC, F3TC) as a partial agonist of CN2R (37–39).
F1T, F2T, and F3T were established using a mixture of specific terpenes, and the concentration was based on published data and in data from our previous work (results from ecotoxicological and cytotoxic assays to be published, POCI-01-02B7-FEDER-053456-BIOBLOCKCOVID). Moreover, we intended to study the CBD-terpene entourage effect, and so we used a lower concentration of F1T, F2T, and F3T when in combination with CBD. Despite the respiratory tract being the dominant route in SARS-CoV-2 infection, the kidney and colon tissues present very considerable levels of ACE2 and TMPRSS2 expression and hence potential targets for virus infection. We performed the viral reduction assays in several cell lines and quantified ACE2 and TMPRSS2 gene expression. Both receptors can be considered as targets for SARS-CoV-2, expecting that the coding genes are upregulated. By its turn, it is expected that RdRp and Spike gene expression is upregulated as viral infectivity progress and that gene expression diminishes because of a virucide or antiviral action. As CN2R activation could limit the release of pro-inflammatory cytokines associated with COVID-19, and in this context being a therapeutic target for SARS-CoV-2, CBD was included in the formulations (F1TC, F2TC, F3TC) as a partial agonist of CN2R. Figure 1 illustrates the adsorption and replication mechanisms as well as the potential action of CBD and terpenes and the ACE2, TMPRSS2, and CN2R expression in the lung, skin, colon, and kidney tissues. Table 2 shows the cytotoxicity evaluation of the formulations per cell line for both pretreatment and treatment conditions, and Table 3 shows the cytotoxicity of the components that comprises the formulations.
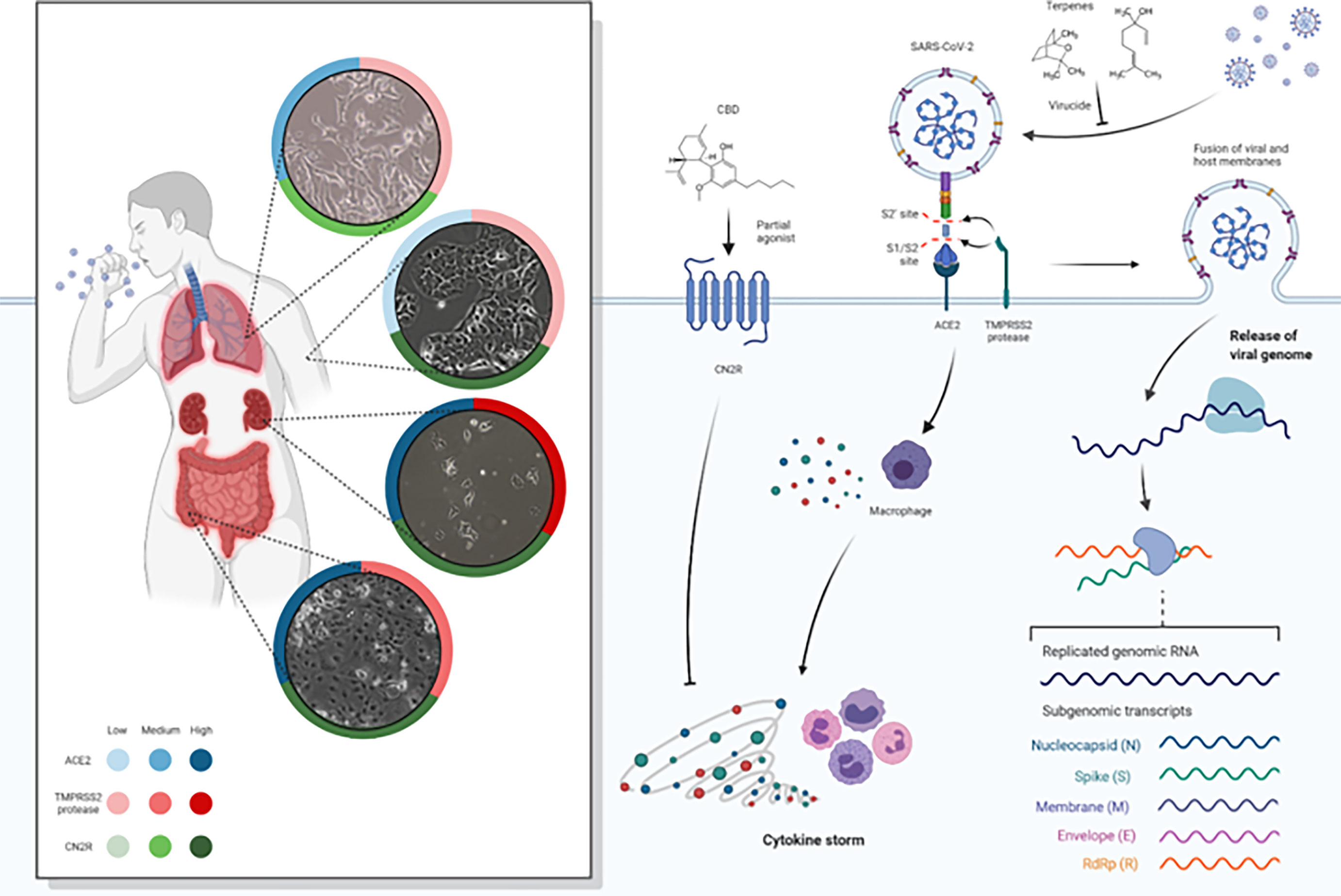
Figure 1 Spike protein of SARS-CoV-2 binds to ACE2 receptors, fusing to the cell membrane and releasing the viral RNA into the host cell. SARC-CoV-2 depends on cellular serine protease, TMPRSS2, for Spike priming. Viral replication in host cells is always associated with inflammation and immune activation being that virus–host–cell interaction produces a set of immune mediators, cytokines, against the virus. It represents the ACE2, TMPRSS2, and CN2R expression levels (low, medium, high) in lung, skin, colon, and kidney tissues (data obtained from Human Protein Atlas available from http://www.proteinatlas.org, Uhlén M et al. Tissue-based map of the human proteome. Science (2015) PubMed: 25613900 DOI: 10.1126/science.1260419). CBD and terpenes (linalool and 1.8-cineole as representative of components of the formulations) are represented as virucide agents blocking and inactivating the virus at an early stage of infection, as antiviral agents blocking the virus cellular machinery and as agents against an overactive immune-inflammatory cascade. As the efficiency of Spike–ACE2 interaction determines SARS-CoV-2 transmissibility, the expression of ACE2 and TMPRSS2 could represent a major risk factor for the susceptibility to SARS-CoV-2 infection. We evaluate the hypothesis of terpenes as virucide agents that could disrupt the interaction between the Spike/TMPRSS2 proteins and the host cell ACE2 receptor. Moreover, after virus entry, terpenes could potentially have an antiviral effect by inhibiting RdRp thus preventing viral replication. Also, it was intended to better understand if a specific group of terpenes and CBD have the potential to act synergistically as therapeutic agents for SARS-CoV-2 and if the action is at early stages or later stages of infection. We assess the hypothesis that CBD may have the potential for modulating the exacerbated inflammatory process typical of COVID-19. CBD is a partial agonist CN2R that is widely expressed in the immune system and, when stimulated, promotes the inhibition of proinflammatory cytokine production, the increase of anti-inflammatory cytokines, and the induction of regulatory T cells.
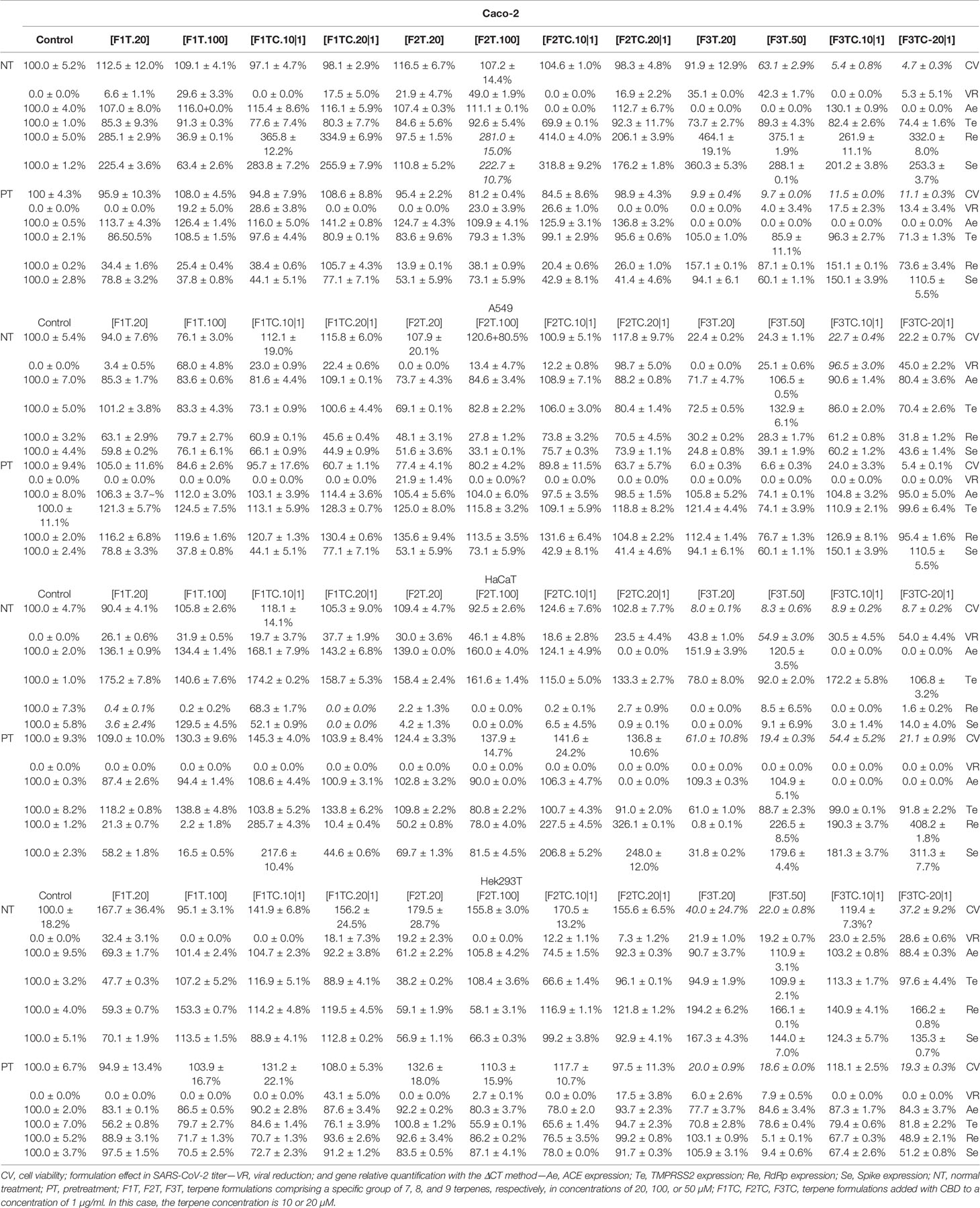
Table 2 Cytotoxicity evaluation of formulations.

Table 3 Formulation components evaluation regarding Caco-2, A549, HaCaT, and Hek293T cell line viability.
A two-way ANOVA analysis was performed to enquire the effect of treatment per cell line type on cytotoxicity, viral reduction, ACE2 expression, TMPRSS2 expression, RdRp expression, and Spike expression. The results of this analysis are presented in Table 4. A statistically significant interaction between the normal treatment and the A549 cell line (F (1, 12) = 3.773; p = 0.038) and between the pretreatment and Caco-2 (F (1, 12) = 8.181; p = 0.014), A549 (F (1, 12) = 7.406; p = 0.020), and HaCaT (F (1, 12) = 7,146; p = 0,020) is denoted.
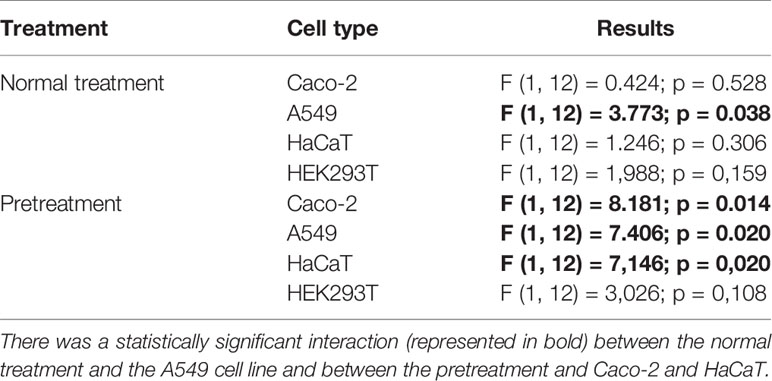
Table 4 A two-way ANOVA regarding the effect of treatment per cell line on cytotoxicity, viral reduction, and gene expression.
F3T is toxic at the concentration of 50 µM. However, F3T is a promising formulation as it reduces viral titer by 35% while cell viability is 91.9%. Interestingly by adding CBD, a much higher toxicity (94.5% of cell death) is promoted; indirectly, this could be the reason why the viral reduction is 0% when using F3T versus F3TC. The toxicity is higher in pretreatment. Comparing F1T.100 with F1TC.10|1, in treatment and pretreatment assays it is possible to conclude those formulations as promising as the viral reduction by 29.6% and 28.6%, respectively, and as Spike and RdRp gene expressions were reduced. Comparing F2T.100 with F2TC.10|1, in treatment and pretreatment assays it was possible to conclude those formulations as promising as the viral reduction by 49.0% and 26.6%, respectively. It is an advantage to use CBD, as the concentration of terpenes, for obtaining approximately the same viral reduction, is ten times lower, and thus it is possible to conclude about an additive effect, which is also denoted when comparing F1T.100 with F1TC.10|1. An interesting fact to be explored is to understand why, comparing F1TC.10|1 pretreatment with F1TC.10|1 or F1TC.20|1 treatment, the Spike and RdRp gene expression increases. This cell line was used as COVID-19 includes gastrointestinal symptoms, and it remains uncertain if they are caused by direct infection, as aerosol droplets can be swallowed and pass the gastrointestinal tract, or whether they are a consequence of immune system activation.
F3T is toxic at the concentration of 20 µM. Of relevance, regarding treatment assays, F3TC.10|1 reduces viral titer by 96.5% while cell viability is 22.7% and using F3TC.20|1 is less efficient in reducing viral titer. This fact is also denoted in Hek549 pointing out to a critical selection of the terpene concentration to be used when combined with CBD. By comparing F1T with F1TC and F2T with F2TC, in treatment assays, it is possible to conclude about the benefit of adding CBD as the viral titer increases, maintaining the cell viability. F2TC.20|1 is one of the most promising formulations as it reduces the viral titer by 98.7%, the best value obtained, and as expected Spike and RdRp gene expression is reduced. The additive effect of CBD is clear, as the concentration of terpenes, for obtaining approximately the same viral reduction, is ten times lower. For this cell line, the pretreatment assays have no impact in viral reduction. This cell line was used as SARS-CoV-2 propagates through aerosol droplets that can be inhaled and infect the upper airways. F2TC.20|1 could be exploited as a promising therapeutic for upper or lower airway infection.
F3T is toxic for the concentration of 20 µM. Pretreatment assays of this cell line are not adequate as viral reduction is 0%. The addition of CBD to F3T has no effect regarding viral reduction or cell viability in this cell line. A comparable effect is denoted regarding F1T.20 vs. F1TC.20|1, in that adding CBD has no (significant) additive effect in viral reduction either in Spike or in RdPp expression. F2T.20 is the most efficient formulation by reducing by 30% the viral titer and by promoting the downregulation of Spike and RdPp expression to less than 5%. The addition of CBD to F2T.20 has no additive effect. Importantly, and compared to the other cell lines, in HaCaT a higher expression of ACE2 and TMPRSS2 is verified. This cell line was used, as skin lesions and lesions of the vascular system in some SARS-CoV-2-positive patients have been reported. The high levels of ACE2 and TMPRSS2 expression could indicate that percutaneous transmission might be a potential risk route for SARS-CoV-2 infection, particularly in conditions of skin dysfunction. Also, the long-term wearing of protective clothing and having contact with disinfectants cause eczematoid dermatitis which might be a risk factor for percutaneous infection. F2T and F1T can be studied as treating skin lesions in SARS-CoV-2 patients. By its turn, F3T.50 could be used for SARS-CoV-2 control in surfaces as it reduces viral titer by 54.9%.
F3T is toxic for almost all the concentrations, except for F3TC.10|1, which is in fact a promising formulation. The addition of CBD to F1T, F2T, or F3T has no effect, regarding viral reduction or cell viability, and promotes upregulation of Spike and RdRp (F1T and F2T). F1T.20 and F2T.20 are the most promising formulations as the virus titer is 32% and 19%, respectively, and Spike and RdRp are downregulated. Similar to the effect in Caco-2, pretreatment and CBD addition is beneficial for this cell line as the virus titer is 43%. Comparing F1T.20 and F2T.20 with F1TC.20|1 and F2T.20|1, it is possible to conclude about a synergistic effect of CBD and terpenes, and that the terpenes from F1T have a higher effect in virus reduction compared to the terpenes from F2T. This cell line was used as COVID-19 includes kidney failure symptoms.
Table 5 intends to summarize per cell line the most promising formulations based on viral reduction and viral gene expression parameters. In Figure 2, it is intended to summarize the efficacy of each formulation per cell line.
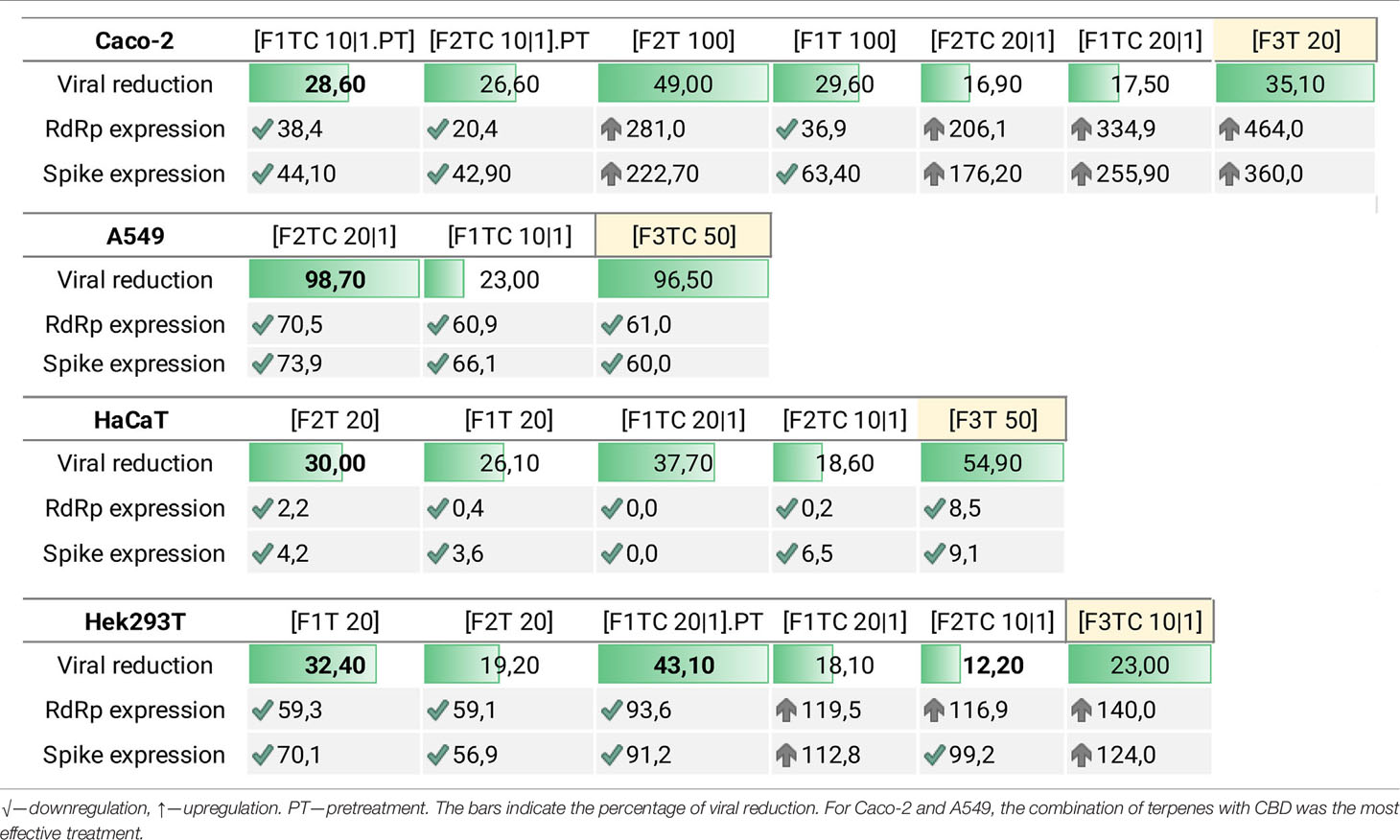
Table 5 Formulation efficacy considering viral reduction, RdRp expression, and Spike expression per cell line.
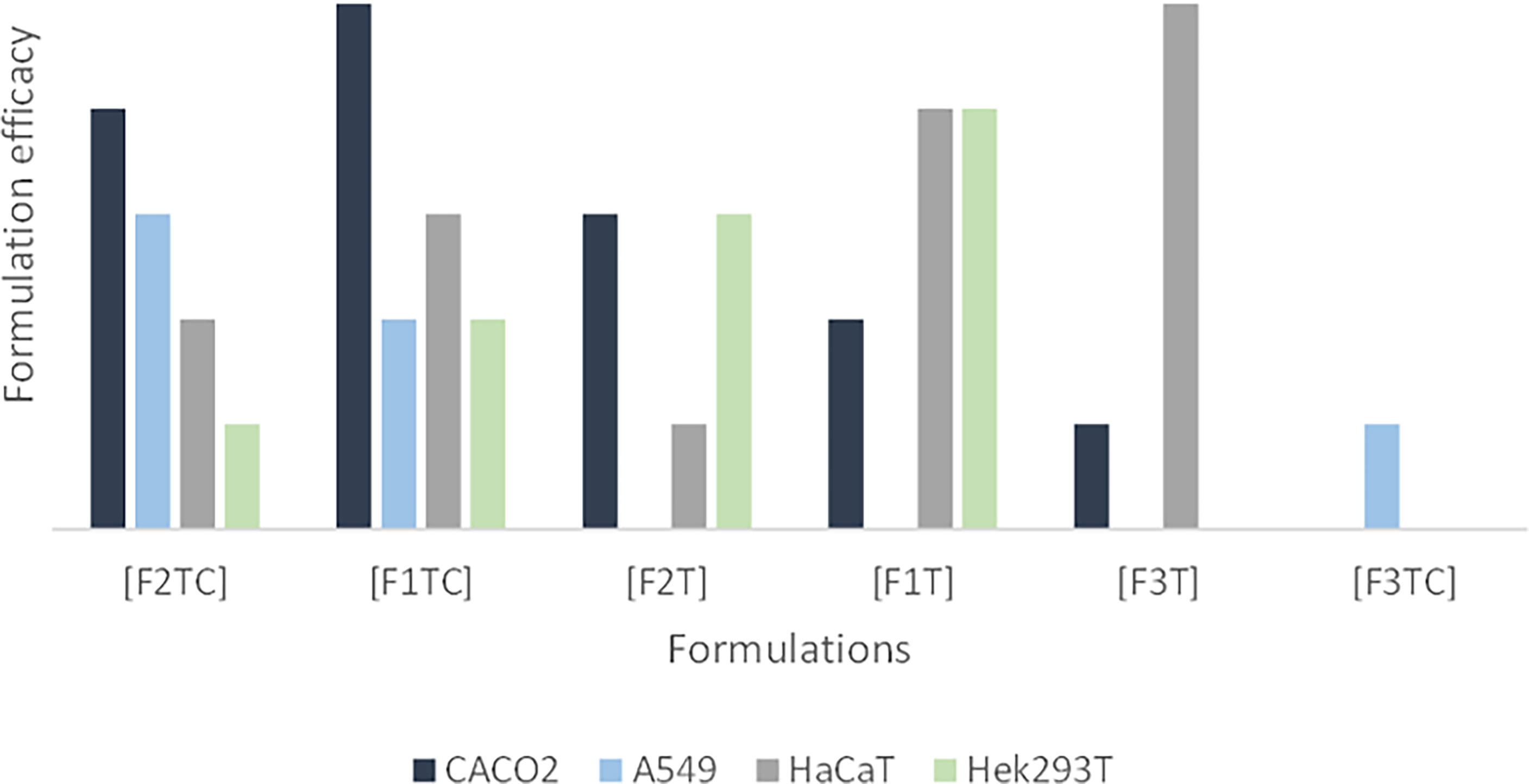
Figure 2 Formulation efficacy (in terms of viral reduction) for Caco-2, A549, HaCaT, and Hek293T cell lines. In this graphic, it is possible to observe i) an additive effect of CBD and terpenes in Caco-2 and A549, ii) that adding CBD to F1T is not advantageous regarding HaCaT, iii) that adding CBD to F2T is an advantage regarding HaCaT, iv) that the terpenes of F1T are more effective than the ones from F2T for HaCaT and Hek293T, vi) that the terpenes from F1T are more effective regarding Caco-2, vii) that F3T and F3TC include terpenes that are more toxic than the ones included in F1T and F2T, being that the toxicity is higher in combination with CBD.
Up to date, a fully effective treatment for COVID-19 is still a challenge. Although the interest in CBD and EOs as therapeutic strategies grows, no scientific studies were made to evaluate the role of CBD and specific terpenes in the progression of SARS-CoV-2 infection. Our group settled an approach focused on the inhibition of both virus entry and viral replication by using biobased formulations from cannabis, thyme, and oregano. The obtained data suggest these formulations to be exploited as new therapeutics targeting COVID-19, providing evidence that CBD and terpenes could be considered for further studies as effective anti-SARS-CoV-2 agents and potentially used for treatment or as adjuvants to conventional COVID-19 therapies. Also, it demonstrates that the selection of terpenes to be used combined with CBD is relevant and points out that treatment should be targeted for afflicted tissues. The proprietary formulations F2TC and F1TC could potentially be used for treating viral infections via modulation of the cytokine storm. Additional studies regarding the molecular mechanism explaining both the virucide or antiviral activity and the immunomodulatory effect will be exploited by our group. It will be interesting to explore the anti-inflammatory function of CBD concerning inflammatory events that happen during severe COVID-19 disease and how it might help to prevent the progression from mild to severe disease. In this context, the activation of the ECS could contribute to preventing the progress and the severity of COVID-19. The current study identifies CBD and a specific group of terpenes as a promising anti-COVID-19 therapeutic strategy that warrants further in vivo testing and preclinical trials.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
All authors certify that they have participated sufficiently in the work to take public responsibility for the content, including participation in the concept, design, analysis, interpretation, writing, or revision of the article. All authors contributed to the article and approved the submitted version.
This research was partially funded by the European Commission, European Regional Development, FEDER/02/SAICT/2020/072560 SI-B7-2020-15, POCI-01-02B7-FEDER-053456, BIOBLOCKCOVID. The funding allowed to perform the collection and harvesting of the medicinal plants, extract and purify the terpenes and formulations, execute the formulations, and execute preliminary assays related to toxicity. Considering in vitro virucide assays and gene expression assays, the work was partially funded by FCT – Fundação para a Ciência e Tecnologia (REF UID/BIM/04293/2019) and was partially supported by grants 104 and 112 of the 1st edition of RESEARCH4COVID (FCT) and by the grant 418 from the 2nd edition of RESEARCH4COVID-19 (FCT). This work was also partially supported by FEDER-European Regional Development Fund with the grant FEDER/02/SAICT/2020/072560.
Authors SS, AC, IL, SR and MM was employed by company EXMceuticals Portugal Lda.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to acknowledge the researchers that participated in the BioBlockCOVID project, especially Prof. João Ramalho Santos for providing the A549 and HaCaT cell lines and execution of preliminary cytotoxicity assays, Prof. José Paulo Sousa, and Dr. Tiago Luz for the ecotoxicity assays, and Prof. Rui Ribeiro and Dra. Matilde Moreira Santos for the aquatic toxicity assays. We acknowledge the assistance in hydrodistillation to Prof. Ana Cristina Figueiredo, Faculdade de Ciências da Universidade de Lisboa, CESAM Lisboa.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.841459/full#supplementary-material
Supplementary Figure 1 | – GC-MS quantitative chromatogram of formulations F1T (A), F2T (B) and F3T (C). Peak numbering represents terpenes ranked from the higher to the lower concentration. X axis - Retention Time (min), Y axis – Intensity. The terpenes are anonymized as F1T, F2T, F3T are proprietary formulations. HPLC-UV chromatogram analysis of a CBD isolate (D), X axis - Retention Time (min), Y axis - Intensity (mAU).
Supplementary Figure 2 | – Graphical representation of the cytotoxicity evaluation of formulations, formulation effect in SARS-CoV-2 titer and gene relative quantification with ΔCT method. Evaluation of the effect from formulations without (F1T=1, F2T=2, F3T=3) and with the addition of CBD (F1TC=1+, F2TC=2+, F3TC=3+). The terpenes were used in a concentration of 20 µM (L) and 100 µM (H) for F1T and F2T, 20 µM (L) and 50 µM (H) for F3T, 10 µM (L) and 20 µM (H) for F1TC and F2TC, 20 µM (H) and 10 µM (L) for F3TC. (A) Cytotoxicity effect; (B) Formulation effect on viral number copies reduction; (C and D) Gene expression of ACE2 and TMPRSS2 genes under the influence of the formulations; (E and F) Gene expression of RdRp and Spike genes under the influence of the formulations. Results are represented as mean with standard deviation and normalized to GAPDH expression. Symbols above bars represent statistical significance (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001). Figure 2.1 – Treatments administrated after SARS-CoV-2 infection in Caco-2 cell line. Figure 2.2 - Treatments administrated before SARS-CoV-2 infection in Caco-2 cell line. Figure 2.3 - Treatments administrated after SARS-CoV-2 infection in A549 cell line. Figure 2.4 –Treatments administrated before SARS-CoV-2 infection in A549 cell line. Figure 2.5 – Treatments administrated after SARS-CoV-2 infection in HaCaT cell line. Figure 2.6 - Treatments administrated before SARS-CoV-2 infection in HaCaT cell line. Figure 2.7 - Treatments administrated after SARS-CoV-2 infection in Hek293T cell line. Figure 2.8 –Treatments administrated before SARS-CoV-2 infection in Hek293T cell line.
ACE-2, angiotensin-converting enzyme 2; THC, tetrahydrocannabinol; ECS, endocannabinoid system; CNR2, cannabinoid receptor 2; RSV, respiratory syncytial virus; EOs, essential oils; HIV, human immunodeficiency virus; MCT, medium-chain triglyceride; CBD, cannabidiol; CPC, centrifuge partition chromatography; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; RdRp, RNA-dependent RNA polymerase; SARS-CoV, severe acute respiratory syndrome-CoV; SARS-CoV-2, severe acute respiratory syndrome-CoV-2.
1. Ford N, Vitoria M, Rangaraj A, Norris SL, Calmy A, Doherty M. Systematic Review of the Efficacy and Safety of Antiretroviral Drugs Against SARS, MERS or COVID-19: Initial Assessment. J Int AIDS Soc (2020) 23(4):e25489. doi: 10.1002/jia2.25489
2. Boukhatem MN, Setzer WN. Aromatic Herbs, Medicinal Plant-Derived Essential Oils, and Phytochemical Extracts as Potential Therapies for Coronaviruses: Future Perspectives. Plants (Basel) (2020) 9(6):800. doi: 10.3390/plants9060800
3. Elshabrawy HA. SARS-CoV-2: An Update on Potential Antivirals in Light of SARS-CoV Antiviral Drug Discoveries. Vaccines (Basel) (2020) 8(2):335. doi: 10.3390/vaccines8020335
4. Bergman ME, Davis B, Phillips MA. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules (2019) 24(21):3961. doi: 10.3390/molecules24213961
5. Jaeger R, Cuny E. Terpenoids With Special Pharmacological Significance: A Review. Nat Prod Commun (2016) 11(9):1373–90. doi: 10.1177/1934578X1601100946
6. Loizzo MR, Saab AM, Tundis R, Statti GA, Menichini F, Lampronti I, et al. Phytochemical Analysis and In Vitro Antiviral Activities of the Essential Oils of Seven Lebanon Species. Chem Biodivers (2008) 5(3):461–70. doi: 10.1002/cbdv.200890045
7. Schuhmacher A, Reichling J, Schnitzler P. Virucidal Effect of Peppermint Oil on the Enveloped Viruses Herpes Simplex Virus Type 1 and Type 2 In Vitro. Phytomedicine (2003) 10(6-7):504–10. doi: 10.1078/094471103322331467
8. Asif M, Saleem M, Saadullah M, Yaseen HS, Al Zarzour R. COVID-19 and Therapy With Essential Oils Having Antiviral, Anti-Inflammatory, and Immunomodulatory Properties. Inflammopharmacology (2020) 28(5):1153–61. doi: 10.1007/s10787-020-00744-0. Erratum in: Inflammopharmacology. 2021 Apr;29(2):577.
9. Reichling J. Plant-Microbe Interaction and Secondary Metabolites With Antiviral, Antibacterial and Antifungal Properties. In: Wink W, editor. Functions and Biotechnology of Plant Secondary Metabolites. West Sussex, United Kingdom: Wiley-Blackwell (2010). p. 214–347. doi: 10.1002/9781119312994.apr0420
10. Tahamtan A, Samieipoor Y, Nayeri FS, Rahbarimanesh AA, Izadi A, Rashidi-Nezhad A, et al. Effects of Cannabinoid Receptor Type 2 in Respiratory Syncytial Virus Infection in Human Subjects and Mice. Virulence (2018) 9(1):217–30. doi: 10.1080/21505594.2017.1389369
11. Sacerdote P, Martucci C, Vaccani A, Bariselli F, Panerai AE, Colombo A, et al. The Nonpsychoactive Component of Marijuana Cannabidiol Modulates Chemotaxis and IL-10 and IL-12 Production of Murine Macrophages Both In Vivo and In Vitro. J Neuroimmunol (2005) 159(1-2):97–105. doi: 10.1016/j.jneuroim.2004.10.003
12. Nadjib BM. Effective Antiviral Activity of Essential Oils and Their Characteristic Terpenes Against Coronaviruses: An Update. J Pharmacol Clin Toxicol (2020) 8(1):1138.
13. Sandner G, Heckmann M, Weghuber J. Immunomodulatory Activities of Selected Essential Oils. Biomolecules (2020) 10(8):1139. doi: 10.3390/biom10081139
14. Juergens UR, Dethlefsen U, Steinkamp G, Gillissen A, Repges R, Vetter H. Anti-Inflammatory Activity of 1.8-Cineol (Eucalyptol) in Bronchial Asthma: A Double-Blind Placebo-Controlled Trial. Respir Med (2003) 97(3):250–6. doi: 10.1053/rmed.2003.1432
15. Juergens LJ, Worth H, Juergens UR. New Perspectives for Mucolytic, Anti-Inflammatory and Adjunctive Therapy With 1,8-Cineole in COPD and Asthma: Review on the New Therapeutic Approach. Adv Ther (2020) 37(5):1737–53. doi: 10.1007/s12325-020-01279-0
16. Álvarez DM, Castillo E, Duarte LF, Arriagada J, Corrales N, Farías MA, et al. Current Antivirals and Novel Botanical Molecules Interfering With Herpes Simplex Virus Infection. Front Microbiol (2020) 11:139. doi: 10.3389/fmicb.2020.00139
17. Ramirez SH, Reichenbach NL, Fan S, Rom S, Merkel SF, Wang X, et al. Attenuation of HIV-1 Replication in Macrophages by Cannabinoid Receptor 2 Agonists. J Leukoc Biol (2013) 93(5):801–10. doi: 10.1189/jlb.1012523
18. Sökmen M, Serkedjieva J, Daferera D, Gulluce M, Polissiou M, Tepe B, et al. In Vitro Antioxidant, Antimicrobial, and Antiviral Activities of the Essential Oil and Various Extracts From Herbal Parts and Callus Cultures of Origanum Acutidens. J Agric Food Chem (2004) 52(11):3309–12. doi: 10.1021/jf049859g
19. Seidakhmetova RB, Beisenbaeva AA, Atazhanova GA, Suleimenov EM, Pak RN, Kulyyasov AT, et al. Chemical Composition and Biological Activity of the Essential Oil From Artemisia Glabella. Pharma Chem J (2002) 36:135–138.40. doi: 10.1023/A:1019630327576
20. Schnitzler P, Schön K, Reichling J. Antiviral Activity of Australian Tea Tree Oil and Eucalyptus Oil Against Herpes Simplex Virus in Cell Culture. Pharmazie (2001) 56(4):343–7.
21. Farag RS, Shalaby AS, El-Baroty GA, Ibrahim NA, Ali MA, Hassan EM. Chemical and Biological Evaluation of the Essential Oils of Different Melaleuca Species. Phytother Res (2004) 18(1):30–5. doi: 10.1002/ptr.1348
22. Wen CC, Kuo YH, Jan JT, Liang PH, Wang SY, Liu HG, et al. Specific Plant Terpenoids and Lignoids Possess Potent Antiviral Activities Against Severe Acute Respiratory Syndrome Coronavirus. J Med Chem (2007) 50(17):4087–95. doi: 10.1021/jm070295s
23. Diniz LRL, Perez-Castillo Y, Elshabrawy HA, Filho CDSMB, de Sousa DP. Bioactive Terpenes and Their Derivatives as Potential SARS-CoV-2 Proteases Inhibitors From Molecular Modeling Studies. Biomolecules (2021) 11(1):74. doi: 10.3390/biom11010074
24. Kintzios SE. The Biotechnology of Oregano (Origanum Sp. And Lippia Sp.). In: Kintzios SE, editor. Medicinal and Aromatic Plants-Industrial Profiles, Oregano. The Genera Origanum and Lippia, vol. 25. London: Taylor and Francis (2002). p. 237–42.
25. Pereira MMA, Morais LC, Zeneratto NJ, Reis WSM, Gómez OC, Luiz JHH, et al. Organic Management vs. Conventional Management Influence the Antimicrobial Activity of Essential Oils of Origanum Vulgare L. Research Soc Dev (2020) 9(11):e4239118504.
26. Catella C, Camero M, Lucente MS, Fracchiolla G, Sblano S, Tempesta M, et al. Virucidal and Antiviral Effects of Thymus Vulgaris Essential Oil on Feline Coronavirus. Res Vet Sci (2021) 137:44–7. doi: 10.1016/j.rvsc.2021.04.024
27. Russo EB. Taming THC: Potential Cannabis Synergy and Phytocannabinoid-Terpenoid Entourage Effects. Br J Pharmacol (2011) 163(7):1344–64. doi: 10.1111/j.1476-5381.2011.01238
28. Mediavilla V, Steinemann S. Essential Oil of Cannabis Sativa L. Strains. J Int Hemp Assoc (1997) 4:80–2.
29. Elsohly MA, Slade D. Chemical Constituents of Marijuana: The Complex Mixture of Natural Cannabinoids. Life Sci (2005) 78(5):539–48. doi: 10.1016/j.lfs.2005.09.011
30. Sommano SR, Chittasupho C, Ruksiriwanich W, Jantrawut P. The Cannabis Terpenes. Molecules (2020) 25(24):5792. doi: 10.3390/molecules25245792
31. Fasinu PS, Phillips S, ElSohly MA, Walker LA. Current Status and Prospects for Cannabidiol Preparations as New Therapeutic Agents. Pharmacotherapy (2016) 36(7):781–96. doi: 10.1002/phar.1780
32. Bridgeman MB, Abazia DT. Medicinal Cannabis: History, Pharmacology, And Implications for the Acute Care Setting. P T (2017) 42(3):180–8.
33. Burstein S. Cannabidiol (CBD) and Its Analogs: A Review of Their Effects on Inflammation. Bioorg Med Chem (2015) 23(7):1377–85. doi: 10.1016/j.bmc.2015.01.059
34. Amin MR, Ali DW. Pharmacology of Medical Cannabis. Adv Exp Med Biol (2019) 1162:151–65. doi: 10.1007/978-3-030-21737-2_8
35. De Petrocellis L, Ligresti A, Moriello AS, Allarà M, Bisogno T, Petrosino S. Effects of Cannabinoids and Cannabinoid-Enriched Cannabis Extracts on TRP Channels and Endocannabinoid Metabolic Enzymes. Br J Pharmacol (2011) 163:1479–94. doi: 10.1111/j.1476-5381.2010.01166.x
36. Robaina Cabrera CL, Keir-Rudman S, Horniman N, Clarkson N, Page C. The Anti-Inflammatory Effects of Cannabidiol and Cannabigerol Alone, and in Combination. Pulm Pharmacol Ther (2021) 69:102047. doi: 10.1016/j.pupt.2021.102047
37. An D, Peigneur S, Hendrickx LA, Tytgat J. Targeting Cannabinoid Receptors: Current Status and Prospects of Natural Products. Int J Mol Sci (2020) 21(14):5064. doi: 10.3390/ijms21145064
38. Zou S, Kumar U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int J Mol Sci (2018) 19(3):833. doi: 10.3390/ijms19030833
39. Jordan CJ, Xi ZX. Progress in Brain Cannabinoid CB2 Receptor Research: From Genes to Behavior. Neurosci Biobehav Rev (2019) 98:208–20. doi: 10.1016/j.neubiorev.2018.12.026
40. Gui H, Sun Y, Luo ZM, Su DF, Dai SM, Liu X. Cannabinoid Receptor 2 Protects Against Acute Experimental Sepsis in Mice. Mediators Inflamm (2013) 2013:741303. doi: 10.1155/2013/741303
41. Rossi F, Tortora C, Argenziano M, Di Paola A, Punzo F. Cannabinoid Receptor Type 2: A Possible Target in SARS-CoV-2 (CoV-19) Infection? Int J Mol Sci (2020) 21(11):3809. doi: 10.3390/ijms21113809
42. Costiniuk CT, Saneei Z, Routy JP, Margolese S, Mandarino E, Singer J, et al. Oral Cannabinoids in People Living With HIV on Effective Antiretroviral Therapy: CTN PT028-Study Protocol for a Pilot Randomised Trial to Assess Safety, Tolerability and Effect on Immune Activation. BMJ Open (2019) 9(1):e024793. doi: 10.1136/bmjopen-2018-024793
43. Karmaus PW, Chen W, Crawford R, Kaplan BL, Kaminski NE. Δ9-Tetrahydrocannabinol Impairs the Inflammatory Response to Influenza Infection: Role of Antigen-Presenting Cells and the Cannabinoid Receptors 1 and 2. Toxicol Sci (2013) 131(2):419–33. doi: 10.1093/toxsci/kfs315
44. Buchweitz JP, Karmaus PW, Williams KJ, Harkema JR, Kaminski NE. Targeted Deletion of Cannabinoid Receptors CB1 and CB2 Produced Enhanced Inflammatory Responses to Influenza A/PR/8/34 in the Absence and Presence of Delta9-Tetrahydrocannabinol. J Leukoc Biol (2008) 83(3):785–96. doi: 10.1189/jlb.0907618
45. Brown JD. Cannabidiol as Prophylaxis for SARS-CoV-2 and COVID-19? Unfounded Claims Versus Potential Risks of Medications During the Pandemic. Res Soc Adm Pharm (2021) 17(1):2053. doi: 10.1016/j.sapharm.2020.03.020
46. Onay A, Ertaş A, Süzerer V, Yener İ, Yilmaz MA, Ayaz-Tilkat E, et al. Cannabinoids for SARS-CoV-2 and Is There Evidence of Their Therapeutic Efficacy? Turk J Biol (2021) 45(4):570–87. doi: 10.3906/biy-2105-73
47. Paland N, Pechkovsky A, Aswad M, Hamza H, Popov T, Shahar E, et al. The Immunopathology of COVID-19 and the Cannabis Paradigm. Front Immunol (2021) 12:631233. doi: 10.3389/fimmu.2021.631233
48. El Biali M, Broers B, Besson M, Demeules J. Cannabinoids and COVID-19. Med Cannabis Cannabinoids (2020) 3(2):111–5. doi: 10.1159/000510799
49. Wang B, Kovalchuk A, Li D, Rodriguez-Juarez R, Ilnytskyy Y, Kovalchuk I, et al. In Search of Preventive Strategies: Novel High-CBD Cannabis Sativa Extracts Modulate ACE2 Expression in COVID-19 Gateway Tissues. Aging (Albany NY) (2020) 12(22):22425–44. doi: 10.18632/aging.202225
50. Anil SM, Shalev N, Vinayaka AC, Nadarajan S, Namdar D, Belausov E, et al. Cannabis Compounds Exhibit Anti-Inflammatory Activity In Vitro in COVID-19-Related Inflammation in Lung Epithelial Cells and Pro-Inflammatory Activity in Macrophages. Sci Rep (2021) 11(1):1462. doi: 10.1038/s41598-021-81049-2
51. Chatow L, Nudel A, Nesher I, Hayo Hemo D, Rozenberg P, Voropaev H, et al. In Vitro Evaluation of the Activity of Terpenes and Cannabidiol Against Human Coronavirus E229. Life (Basel) (2021) 11(4):290. doi: 10.3390/life11040290
52. Gallily R, Yekhtin Z, Hanuš LO. The Anti-Inflammatory Properties of Terpenoids From Cannabis. Cannabis Cannabinoid Res (2018) 3(1):282–90. doi: 10.1089/can.2018.0014
53. Baron EP. Medicinal Properties of Cannabinoids, Terpenes, and Flavonoids in Cannabis, and Benefits in Migraine, Headache, and Pain: An Update on Current Evidence and Cannabis Science. Headache (2018) 58(7):1139–86. doi: 10.1111/head.13345
54. Maayah ZH, Takahara S, Ferdaoussi M, Dyck JRB. The Molecular Mechanisms That Underpin the Biological Benefits of Full-Spectrum Cannabis Extract in the Treatment of Neuropathic Pain and Inflammation. Biochim Biophys Acta Mol Basis Dis (2020) 1866(7):165771. doi: 10.1016/j.bbadis.2020.165771
55. Namdar D, Voet H, Ajjampura V, Nadarajan S, Mayzlish-Gati E, Mazuz M, et al. Terpenoids and Phytocannabinoids Co-Produced in Cannabis Sativa Strains Show Specific Interaction for Cell Cytotoxic Activity. Molecules (2019) 24(17):3031. doi: 10.3390/molecules24173031
56. Bojkova D, Klann K, Koch B, Widera M, Krause D, Ciesek S, et al. Proteomics of SARS-CoV-2-Infected Host Cells Reveals Therapy Targets. Nature (2020) 583(7816):469–72. doi: 10.1038/s41586-020-2332-7
57. Rosa RB, Dantas WM, do Nascimento JCF, da Silva MV, de Oliveira RN, Pena LJ. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses (2021) 13(3):379. doi: 10.3390/v13030379
58. Xiao F, Tang M, Zheng X, Liu Y, Li X, Shan H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology (2020) 158(6):1831–1833.e3. doi: 10.1053/j.gastro.2020.02.055
59. Lee S, Yoon GY, Myoung J, Kim SJ, Ahn DG. Robust and Persistent SARS-CoV-2 Infection in the Human Intestinal Brush Border Expressing Cells. Emerg Microbes Infect (2020) 9(1):2169–79. doi: 10.1080/22221751.2020.1827985
60. Kellum JA, van Till JWO, Mulligan G. Targeting Acute Kidney Injury in COVID-19. Nephrol Dial Transplant (2020) 35(10):1652–62. doi: 10.1093/ndt/gfaa231
61. Liakopoulos V, Roumeliotis S, Papachristou S, Papanas N. COVID-19 and the Kidney: Time to Take a Closer Look. Int Urol Nephrol (2021), 1–5. doi: 10.1007/s11255-021-02976-7
62. Novak N, Peng W, Naegeli MC, Galvan C, Kolm-Djamei I, Brüggen C, et al. SARS-CoV-2, COVID-19, Skin and Immunology - What Do We Know So Far? Allergy (2021) 76(3):698–713. doi: 10.1111/all.14498
63. Recalcati S. Cutaneous Manifestations in COVID-19: A First Perspective. J Eur Acad Dermatol Venereol (2020) 34(5):e212–3. doi: 10.1111/jdv.16387
64. Wollina U, Karadağ AS, Rowland-Payne C, Chiriac A, Lotti T. Cutaneous Signs in COVID-19 Patients: A Review. Dermatol Ther (2020) 33(5):e13549. doi: 10.1111/dth.13549
65. Matar S, Oulès B, Sohier P, Chosidow O, Beylot-Barry M, Dupin N, et al. Cutaneous Manifestations in SARS-CoV-2 Infection (COVID19): A French Experience and a Systematic Review of the Literature. J Eur Acad Dermatol Venereol (2020) 34(11):e686–9. doi: 10.1111/jdv.16775
Keywords: CBD - cannabidiol, endocannabinoid system (ECS), SARS-CoV-2, therapeutics, terpenes, formulations, essential oil (EO)
Citation: Santos S, Barata P, Charmier A, Lehmann I, Rodrigues S, Melosini MM, Pais PJ, Sousa AP, Teixeira C, Santos I, Rocha AC, Baylina P and Fernandes R (2022) Cannabidiol and Terpene Formulation Reducing SARS-CoV-2 Infectivity Tackling a Therapeutic Strategy. Front. Immunol. 13:841459. doi: 10.3389/fimmu.2022.841459
Received: 22 December 2021; Accepted: 21 January 2022;
Published: 15 February 2022.
Edited by:
Tengchuan Jin, University of Science and Technology of China, ChinaReviewed by:
Khalid Bougrin, Mohammed V University, MoroccoCopyright © 2022 Santos, Barata, Charmier, Lehmann, Rodrigues, Melosini, Pais, Sousa, Teixeira, Santos, Rocha, Baylina and Fernandes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susana Santos, c2lyc2FudG9zc0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.