
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 20 April 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.804822
 Natalia Sherina1
Natalia Sherina1 Charlotte de Vries1
Charlotte de Vries1 Nastya Kharlamova1
Nastya Kharlamova1 Natalie Sippl1
Natalie Sippl1 Xia Jiang2Boel Brynedal3Elin Kindstedt1,4Monika Hansson1Linda Mathsson-Alm5,6Lena Israelsson1Ragnhild Stålesen1
Xia Jiang2Boel Brynedal3Elin Kindstedt1,4Monika Hansson1Linda Mathsson-Alm5,6Lena Israelsson1Ragnhild Stålesen1 Saedis Saevarsdottir7,8
Saedis Saevarsdottir7,8 Rikard Holmdahl9Aase Hensvold1,10
Rikard Holmdahl9Aase Hensvold1,10 Gunnar Johannsen11,12
Gunnar Johannsen11,12 Kaja Eriksson11,13
Kaja Eriksson11,13 Federica Sallusto14,15Anca I. Catrina1,10†
Federica Sallusto14,15Anca I. Catrina1,10† Johan Rönnelid6
Johan Rönnelid6 Caroline Grönwall1
Caroline Grönwall1 Tülay Yucel-Lindberg11,13
Tülay Yucel-Lindberg11,13 Lars Alfredsson3,16
Lars Alfredsson3,16 Lars Klareskog1
Lars Klareskog1 Luca Piccoli14
Luca Piccoli14 Vivianne Malmström1Khaled Amara1
Vivianne Malmström1Khaled Amara1 Karin Lundberg1*
Karin Lundberg1*Based on the epidemiological link between periodontitis and rheumatoid arthritis (RA), and the unique feature of the periodontal bacterium Porphyromonas gingivalis to citrullinate proteins, it has been suggested that production of anti-citrullinated protein antibodies (ACPA), which are present in a majority of RA patients, may be triggered in the gum mucosa. To address this hypothesis, we investigated the antibody response to a citrullinated P. gingivalis peptide in relation to the autoimmune ACPA response in early RA, and examined citrulline-reactivity in monoclonal antibodies derived from human gingival B cells. Antibodies to a citrullinated peptide derived from P. gingivalis (denoted CPP3) and human citrullinated peptides were analyzed by multiplex array in 2,807 RA patients and 372 controls; associations with RA risk factors and clinical features were examined. B cells from inflamed gingival tissue were single-cell sorted, and immunoglobulin (Ig) genes were amplified, sequenced, cloned and expressed (n=63) as recombinant monoclonal antibodies, and assayed for citrulline-reactivities by enzyme-linked immunosorbent assay. Additionally, affinity-purified polyclonal anti-cyclic-citrullinated peptide (CCP2) IgG, and monoclonal antibodies derived from RA blood and synovial fluid B cells (n=175), were screened for CPP3-reactivity. Elevated anti-CPP3 antibody levels were detected in RA (11%), mainly CCP2+ RA, compared to controls (2%), p<0.0001, with a significant association to HLA-DRB1 shared epitope alleles, smoking and baseline pain, but with low correlation to autoimmune ACPA fine-specificities. Monoclonal antibodies derived from gingival B cells showed cross-reactivity between P. gingivalis CPP3 and human citrullinated peptides, and a CPP3+/CCP2+ clone, derived from an RA blood memory B cell, was identified. Our data support the possibility that immunity to P. gingivalis derived citrullinated antigens, triggered in the inflamed gum mucosa, may contribute to the presence of ACPA in RA patients, through mechanisms of molecular mimicry.
Seropositive rheumatoid arthritis (RA) is characterized by presence of ACPA, autoantibodies targeting proteins citrullinated by peptidyl arginine deiminase (PAD) enzymes (1). ACPA associate with RA risk factors HLA-DRB1 shared epitope (SE) and smoking (2), and with a more severe disease course (3). Data from experimental models suggest ACPA may directly contribute to pain and bone loss (4).
Since ACPA can be detected years before clinical symptoms (5), it has been suggested that break of tolerance to citrullinated proteins may occur at extra-articular (mucosal) sites. While many studies have focused on the lungs (2, 4, 6), the gum mucosa has raised increasing interest in this context. There is an epidemiological link between periodontitis (PD), gingival inflammation causing tooth loss as a result of degraded jawbone, and RA (7). Jawbone loss has also been found increased in ACPA+ individuals before RA symptom onset (8), and higher prevalence of PD was reported for ACPA+ at-risk of RA individuals versus population controls (9, 10), implicating that PD may precede RA onset.
Notably, citrullinated proteins are present in PD gingival tissue (GT), and increased PAD activity has been detected in PD gingival crevicular fluid (11, 12). Moreover, Porphyromonas gingivalis (Pg), a key pathogen driving PD (13), is the only pathogen known to express a PAD enzyme (P.PAD) (14), capable of citrullinating bacterial as well as host proteins, including the RA candidate autoantigens fibrinogen, α-enolase, vimentin and histones (15–18). A number of reports also suggest P.PAD can autocitrullinate (14, 15, 17–19). With citrullinated proteins exposed in the bacteria-rich inflamed gingiva, we hypothesize that this milieu may facilitate break of tolerance and production of ACPA (20), possibly by mechanisms of molecular mimicry. Notably, we have previously shown that circulating ACPA targeting human citrullinated α-enolase cross-react with the Pg counterpart (21).
Increased antibody responses to Pg antigens, including citrullinated P.PAD, have been shown in RA, up to 10 years before diagnosis, and in individuals at-risk of RA, compared to population controls (15, 17, 22–24). However, it has not been reported how the antibody response to citrullinated P.PAD relates to the autoimmune ACPA response, RA risk factors and clinical features, or whether such antibodies are produced locally in the inflamed periodontium of PD patients. Hence, in the present study, we have investigated the antibody response to citrullinated Pg PAD in a large and well-characterized early RA cohort, by screening for reactivity against a citrullinated P.PAD peptide denoted CPP3, previously identified as an immunodominant epitope in RA (15, 23). Additionally, for the first time we have generated single-cell derived monoclonal antibodies (mAbs) from gingival tissue B cells of PD patients, and analyzed their reactivity to Pg and human citrullinated antigens.
Newly diagnosed, disease modifying anti-rheumatic drug (DMARD)-naïve, RA patients and population controls from the Epidemiological Investigation of RA (EIRA) study (25) were included. Data on HLA-DRB1 SE and PTPN22 (2,807 RA; 1,936 controls), smoking (2,807 RA; 4,864 controls), anti-CCP2 IgG (2,859 RA), and 5-year clinical data (1,600 RA), captured through linkage with the Swedish Rheumatology Quality Register, were generated previously and retrieved from the EIRA database (2, 26, 27). Periodontal status was not available for EIRA cases and controls.
Biopsies were obtained from inflamed gingival tissue (GT) during surgical treatment of periodontitis (n=3 ACPA+ RA/PD patients; n=4 non-RA/PD patients) at the Department of Dental Medicine, Karolinska Institutet, and specialist clinic Danakliniken Danderyd, Sweden.
All RA patients fulfilled the ACR 1987 and/or ACR/EULAR 2010 RA classification criteria. Samples were collected in compliance with the Declaration of Helsinki, with informed consent and ethical approval (Regional Ethics Review Board Stockholm).
Gingival tissue biopsies were either collected in ice-cold PBS, 2% human sera (HS), and processed directly (n=4: GT01, GT03, GT09 and GT10), or snap-frozen in liquid nitrogen and stored at -80°C (12-184 days) before processed (n=4: GT05-GT08). According to a modified protocol (28), the gingival tissue biopsies were rinsed in PBS, 2% HS, digested mechanically on ice (in 200µl PBS, 2% HS) and treated with 0.5mg Liberase™ Dispase-High (RocheDiagnostics GmbH, Mannheim, Germany) for 1h at 37°C, 200rpm. Dissociated GT cells were then labelled for flow cytometry analysis and single-cell sorting. See Supplementary Methods for detailed staining protocols.
Single viable CD3-/CD14-/CD19+ B cells were sorted into 96-well PCR plates containing 5μl 0.5×PBS, 10mM DTT, 8U RNAsin inhibitor/well (Promega, Madison, WI, USA); variable region immunoglobulin (Ig) cDNA was synthesized by reverse transcription and amplified using multiplex PCR, as previously described (29). Immunoglobulin heavy (IgH) and light (IgL) chains were sequenced (Eurofins Genomics) and annotated using IgBLAST, and IMGT/V-QUEST; variable region N-linked glycosylation sites were identified in translated sequences of V-(D)-J regions using the NetNGlyc1.0 server (www.cbs.dtu.dk/services/NetNGlyc/).
Recombinant human monoclonal antibodies (mAbs) were generated from one ACPA+RA/PD patient (GT01; fresh biopsy) and one non-RA/PD patient (GT06; frozen biopsy) as previously described (29–31). In brief, Ig gene-specific PCR was run to introduce restriction sites for expression vector cloning. Purified digested PCR products were ligated into expression vectors containing human Igγ1, Igλ or Igκ constant regions, and transformed into DH5α bacteria; purified IgH and IgL plasmids were sequenced to confirm identity with the original PCR products, and mAbs were produced by transient co-transfection in high-density suspension cultures of Expi293 cells using the polyethyleneimine (PEI)-precipitation method. Supernatants were collected after 5 days of culture, and mAbs purified on protein G-Sepharose® 4 Fast flow beads, eluted with 0.1M glycine buffer into 1M Tris-HCl, pH 8.0, following buffer exchange to PBS. IgG1 concentrations were determined by IgG ELISA, as previously described (29). Monoclonal antibodies derived from RA synovial fluid (n=139) and peripheral blood (n=36) plasma or memory B cells from 11 ACPA+ RA patients, including 13 CCP2+ clones, were produced previously, following the same protocol (29–33).
Sera from 2,859 RA patients and 372 controls were screened on a custom-made multiplex microarray (ThermoFisher Scientific, ImmunoDiagnostics Division, Uppsala, Sweden) for IgG reactivity against citrullinated peptides derived from Pg PAD (CPP3) and human proteins (Supplementary Table 1), as previously described (26, 34). Two different CPP3-cutoff values, corresponding to the 98th and 80th percentile reactivity among controls, were used.
Monoclonal antibodies (5µg/ml) were assayed for CCP2-reactivity using the Immunoscan CCPlus® assay (Euro-Diagnostica, Malmoü, Sweden), in accordance with the kit instructions, and for reactivity against CPP3 and human protein-derived citrullinated peptides (Supplemental Table 1) using in-house ELISAs as previously described (15, 23, 30, 35). See Supplementary Methods for a detailed protocol. Polyreactivity/unspecific binding was analyzed using uncoated wells blocked with 2% BSA, and in LPS-, insulin-, and dsDNA ELISAs, following established protocols (29). In addition, polyreactivity was evaluated in an ELISA based on the soluble membrane protein fraction of Hek293 cells (fractionated using ProteoExtract subcellular proteome extraction kit, Calbiochem, Darmstadt, Germany), as previously described (36). See Supplementary Methods for a detailed protocol.
Affinity-purified anti-CCP2 IgG, and corresponding anti-CCP2 IgG-depleted IgG pools (35), were also analyzed in the CPP3 ELISA.
Mann-Whitney U test was used to assess anti-CPP3 IgG levels as well as clinical variables, with adjustment for age, sex and smoking (in multivariant analyses). ACPA co-occurrence was calculated using pairwise Pearson correlation (using Rv.3.3.3), among RA patients expressing > one ACPA fine-specificity. Associations between RA risk factors and anti-CPP3 IgG was calculated in SAS (version 9.4) using unconditional logistic regression and presented as odds ratios (OR) with 95% confidence intervals (CI); adjusted for age, sex and residential area. Statistical differences for Ig gene characteristics were not evaluated due to the limited number of patients (n=2).
We first screened 2,859 early RA serum samples on a multiplex array containing Pg CPP3 and eight citrullinated peptides derived from human proteins previously identified as RA candidate autoantigens (Supplementary Table 1). With a cut-off set at the 98th percentile among controls, 11% of RA patients were anti-CPP3 IgG positive, with no significant reactivity to the corresponding non-modified counterpart RPP3 (Figure 1A).
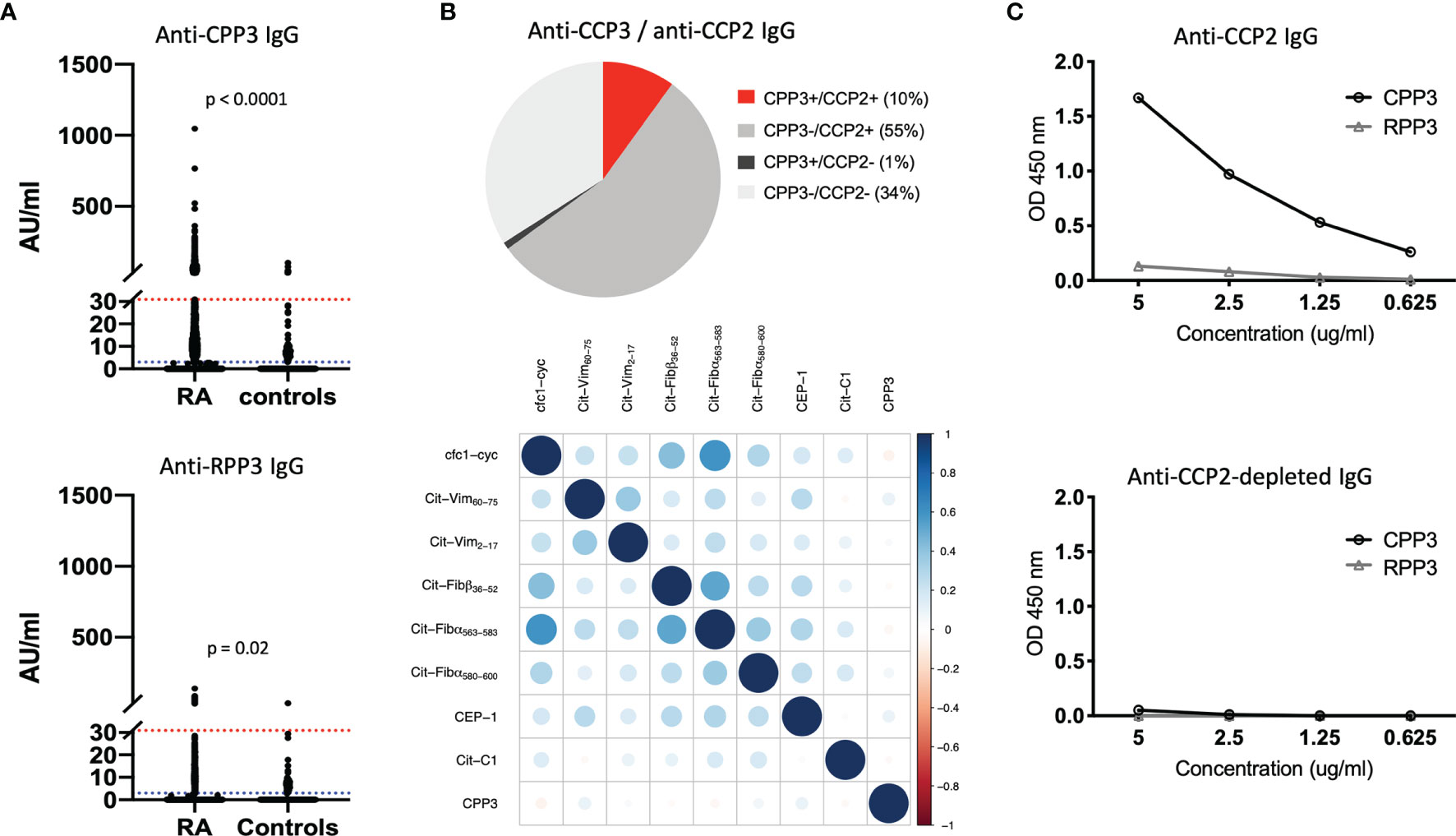
Figure 1 The anti-CPP3 antibody response in relation to the autoimmune ACPA response. (A) Anti-CPP3 (top panel) and anti-RPP3 (bottom panel) IgG levels in EIRA RA patients (n=2,859) and controls (n=372), measured by multiplex microarray; 98th percentile cut-off (red line), and 80th percentile cut-off (blue line) are shown; p-values indicate differences in CPP3/RPP3 IgG levels between groups. (B) Pie chart illustrating the frequency (%) of CPP3+ RA patients (98th percentile cut-off) in relation to CCP2 status (n=2,859) (top panel), and co-occurrence of anti-CPP3 IgG (98th percentile cut-off) in relation to eight autoimmune ACPA fine-specificities (measured by multiplex microarray) in EIRA RA patients (n=2,859) (bottom panel); correlation between antibody expression vectors (corrplot package v. 0.77) was used to illustrate co-occurrence, colours signify degree of correlation according to Pearson correlation coefficient (full scale shown on the right). (C) CPP3/RPP3 reactivity in a pool of affinity-purified anti-CCP2 IgG (top panel) and corresponding anti-CCP2 IgG-depleted IgG pool (bottom panel), at different IgG concentrations. AU, arbitrary units; CPP3, citrullinated P.PAD peptide; RPP3, arginine-containing version of CPP3; RA, rheumatoid arthritis; CCP2, cyclic citrullinated peptide(s); OD, optical density; cfc1-cyc, cyclic citrullinated filaggrin peptide 1; Cit-Vim, citrullinated vimentin; Cit-Fib, citrullinated fibrinogen; CEP-1, citrullinated α-enolase peptide 1; Cit-C1, citrullinated triple helical C1 epitope on collagen type II.
Since CPP3/RPP3 are oral bacteria-derived antigens, we hypothesize that some CPP3/RPP3 reactivity seen in controls may reflect a true bacterial immune response rather than background reactivity. Hence, we also analyzed the dataset using a lower cut-off (80th percentile), allowing 20% CPP3+ controls. With this lower cut-off, anti-CPP3 IgG was detected in 42% of RA patients, while anti-RPP3 IgG was detected in 11% of RA and 15% of controls. Both anti-CPP3 and anti-RPP3 IgG levels were significantly higher in RA patients than in controls.
Similar to the autoimmune ACPA fine-specificities (26), a majority of CPP3+ RA patients (91%) were confined to the CCP2+ subset, but showed weak correlation to other ACPA-reactivities (Pearson correlation coefficients between -0.074 and 0.098) (Figure 1B).
With access to affinity-purified polyclonal anti-CCP2 IgG (35), we could also show binding to CPP3 but not RPP3, demonstrating cross-reactivity between Pg CPP3 and CCP2, i.e. the antigenic peptide(s) used in the gold standard clinical ACPA test (Figure 1C).
To further characterize the CPP3+ RA subset, we analyzed RA risk factors. HLA-DRB1 SE showed a stronger association with CPP3+ (OR=6.07) compared to CPP3- RA (OR=2.55), p<0.0001 (Table 1). However, the significance for the association was lost when analyzing the CCP2+ subset only. PTPN22 polymorphism associated with both CPP3+ (OR=1.85) and CPP3- RA (OR=1.60), with no difference between subsets (p=0.5). Smoking on the other hand, showed a stronger association with CPP3+ compared to CPP3- RA, even within the CCP2+ subset (OR=2.88 for CPP3+ RA vs. OR=1.75 for CPP3- RA, p=0.0012).
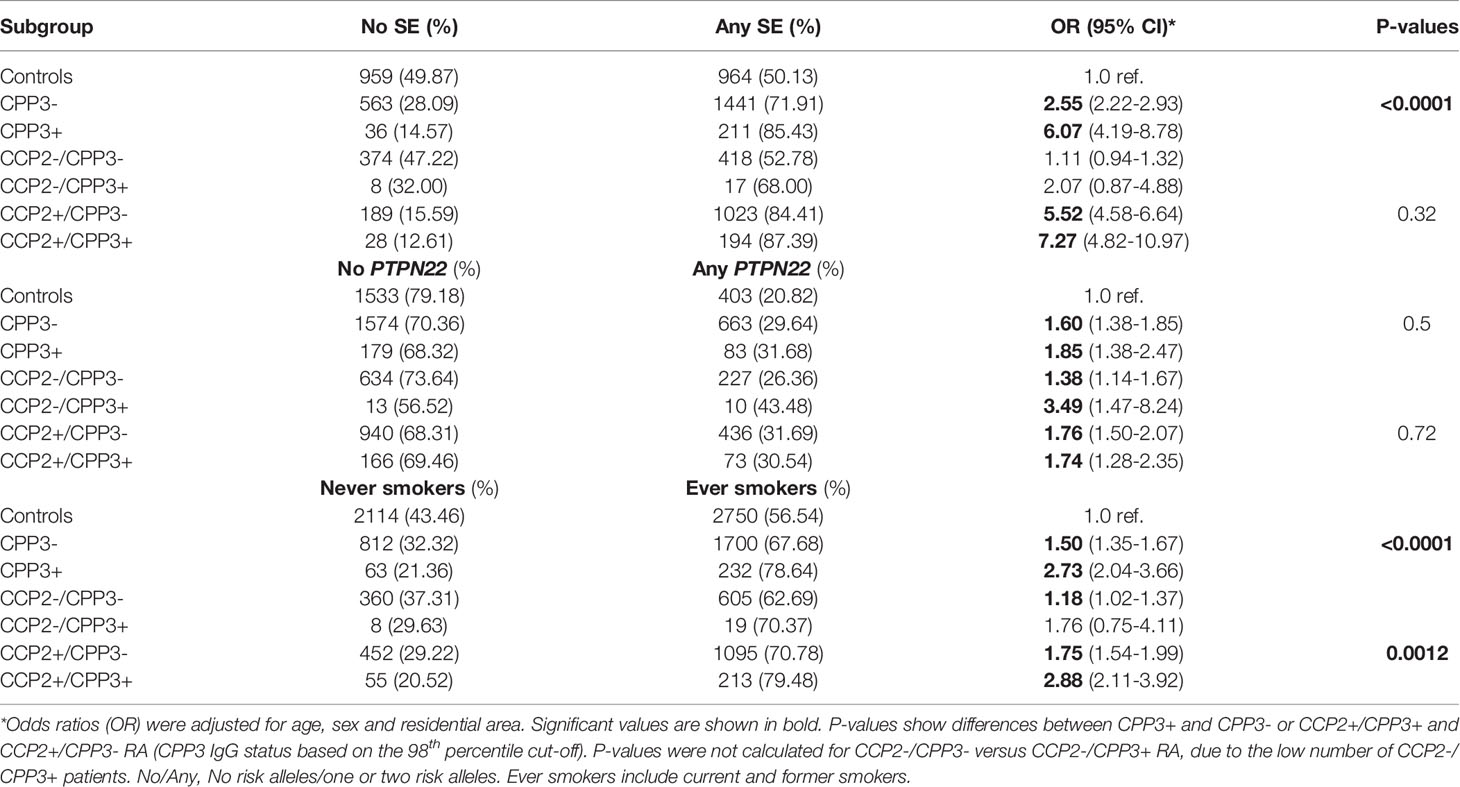
Table 1 Associations between anti-CPP3 IgG and RA risk factors in EIRA.
We also co-analyzed clinical features. Patient-reported baseline pain and global assessment were significantly higher in CPP3+ compared to CPP3- RA, but notably not in CCP2+ compared to CCP2- RA (Table 2). The difference for pain, but not patient global, remained significant after adjustment for age, sex and smoking. The same results were obtained when anti-CPP3 IgG was analyzed within the CCP2+ subset (Supplementary Table 2). Anti-CPP3 IgG also associated with significantly higher ESR at 12 and 48 months, but not with CRP, swollen-/tender joint counts, disease activity score in 28 joints (DAS28) or the health assessment questionnaire (HAQ). Most of these other clinical variables were significantly higher in CCP2+ versus CCP2- RA over time (Supplementary Tables 3–6).
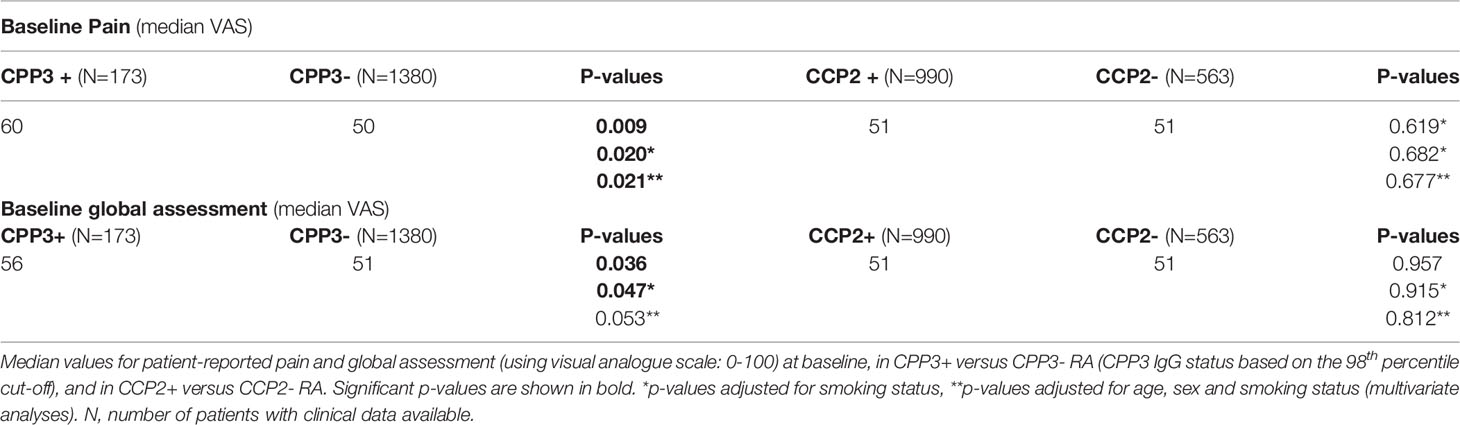
Table 2 Baseline pain and global assessment, stratified for CPP3 or CCP2 status in EIRA.
We next set out to investigate presence of Pg CPP3-reactive B cells in the gum mucosa. Thus, we sorted single B cells from four fresh (Figure 2) and four frozen (Supplementary Figure 1) gingival tissue biopsies, of PD patients with (n=3) and without (n=4) ACPA+ RA, and retrieved between 108 and 576 live CD3-/CD14-/CD19+ cells from each biopsy. Cell yields from frozen samples were variable and consistently lower than from fresh samples, and the lymphocyte population was generally less clear, hence B-cell subsets were analyzed in fresh samples only.

Figure 2 Flow cytometry sorting of CD19+ B cells from gingival tissue biopsies. (A) Representative general gating strategy for CD19+ B cells from fresh GT biopsies. (B) Flow cytometry plots showing the frequency of CD19+ B cells after exclusion of dead, CD3+ and CD14+ cells; GT01 and GT10 originate from PD patients with ACPA+ RA (PD/ACPA+ RA), GT03 and GT09 originate from PD patients without RA (PD). (C) Flow cytometry plots showing the frequency of memory B cells (CD19+/CD27+/CD138-) and plasma cells (CD19+/CD27+/CD138+) in GT01 and GT03, or memory B cells (CD19+/CD27+/IgD-), naïve B cells (CD19+/CD27-/IgD+) and unswitched memory B cells (CD19+/CD27+/IgD+) in GT10 and GT09. Red boxes show the number of CD19+ B cells sorted from each biopsy. FSC, forward scatter; SSC, side scatter; GT, gingival tissue; PD, periodontitis; ACPA, anti-citrullinated protein antibody; RA, rheumatoid arthritis.
Using two different B-cell panels, we compared memory and plasma cell frequencies in one ACPA+ RA/PD (GT01; fresh biopsy) versus one non-RA/PD (GT03; fresh biopsy) patient, and memory, naïve and unswitched memory B cells in another ACPA+ RA/PD (GT10; fresh biopsy) versus non-RA/PD (GT09; fresh biopsy) patient. With the first panel, memory B cells (CD19+/CD27+/CD138-) and plasma cells (CD19+/CD27+/CD138+) were detected in both the ACPA+ RA and non-RA sample. With the other panel, we detected memory B cells (CD19+/CD27+/IgD-), naïve (CD19+/CD27-/IgD+), and unswitched memory B cells (CD19+/CD27+/IgD+) in both the ACPA+ RA and non-RA sample.
We subsequently analyzed GT B-cell receptor variable gene region sequences, and distribution of Ig subclasses, in one ACPA+ RA/PD patient (GT01; fresh biopsy) and one non-RA/PD patient (GT06; frozen biopsy), selected based on the successful recovery of heavy and light chain PCR product from the sorted B cells. Ninety-four matched variable heavy (VH) and light (VL) sequences were generated from GT01, and 54 from GT06. VH3 gene family representation was higher in the ACPA+ RA/PD patient, and by individual variable region genes, IGHV4– 31, IGKV1-33 and IGLV1-47 were overrepresented, while IGHV4-4, IGHV4-38, IGHV1-69 and IGLV1-51 were underrepresented (Supplementary Figure 2).
The complementary determining region 3 (CDR3) length and number of negatively and positively charged amino acids were similar, and the GT immunoglobulins displayed similarly high numbers of somatic hypermutations (SHM), with a median of 27.0 (VH) and 20.0 (VL) for the ACPA+ RA/PD patient, and 27.5 (VH) and 16.0 (VL) for the non-RA/PD patient; mutations were distributed within the variable region with enrichment of replacement (R) over silent (S) mutations in the CDRs. The frequency of N-glycosylation sites was higher in the ACPA+ RA/PD patient (24.5%) than in the non-RA/PD patient (14.7%). The distribution of Ig subclasses was dominated by IgG1 followed by IgA in the ACPA+ RA/PD patient, and IgG1 and IgG2 in the non-RA/PD patient, with lower numbers of IgG3 and IgG4 (Supplementary Figure 3).
To assess presence of citrulline-reactive GT B cells, we randomly selected B-cell clones from the ACPA+ RA/PD patient (GT01; fresh biopsy) and recombinantly expressed 70 mAbs. We also expressed 16 GT mAbs from the non-RA/PD patient (GT06; frozen biopsy) after preselecting clones with a high level of SHM (>15) and/or N-glycosylation sites [i.e. previously described ACPA characteristics (30, 31, 33)]. Antibodies expressed at >100 μg/ml (52 from the ACPA+ RA/PD patient; 11 from the non-RA/PD patient) were analyzed.
Our mAb ELISA data show that 10 GT B cells were positive for the Pg CPP3 peptide derived from P.PAD (Figure 3A). Four CPP3+ mAbs also showed reactivity with the non-modified counterpart RPP3. Additionally, eight CPP3+ clones (and seven CPP3- clones) showed reactivity with citrullinated peptides derived from human proteins, mainly histone-4 and filaggrin, but for most of these peptide-reactivities, also the arginine counterparts were targeted. None of the mAbs were polyreactive/unspecific when analyzed by LPS-, insulin- and dsDNA ELISAs (data not shown). However, when tested in an ELISA based on soluble HEK293 cell membrane proteins, 6/53 clones from GT01 and 7/11 clones from GT06 were positive, suggesting an enrichment of unspecific autoreactivity in preselected clones from the non-RA/PD patient (GT06; frozen biopsy). Still, three CPP3+ clones from the ACPA+ RA/PD patient (GT01; fresh biopsy) had no polyreactivity or RPP3-reactivity. Two of these clones (GT01:04A03 and GT01:01E09) showed cross-reactivity with citrullinated filaggrin and/or histone-4 peptides. None of the clones were positive for CCP2 (data not shown). Phylogenetic trees of VH gene sequences showed that 9.6% of the ACPA+ RA/PD and 3.7% of the non-RA/PD GT B cells were clonally related (Figure 3B).
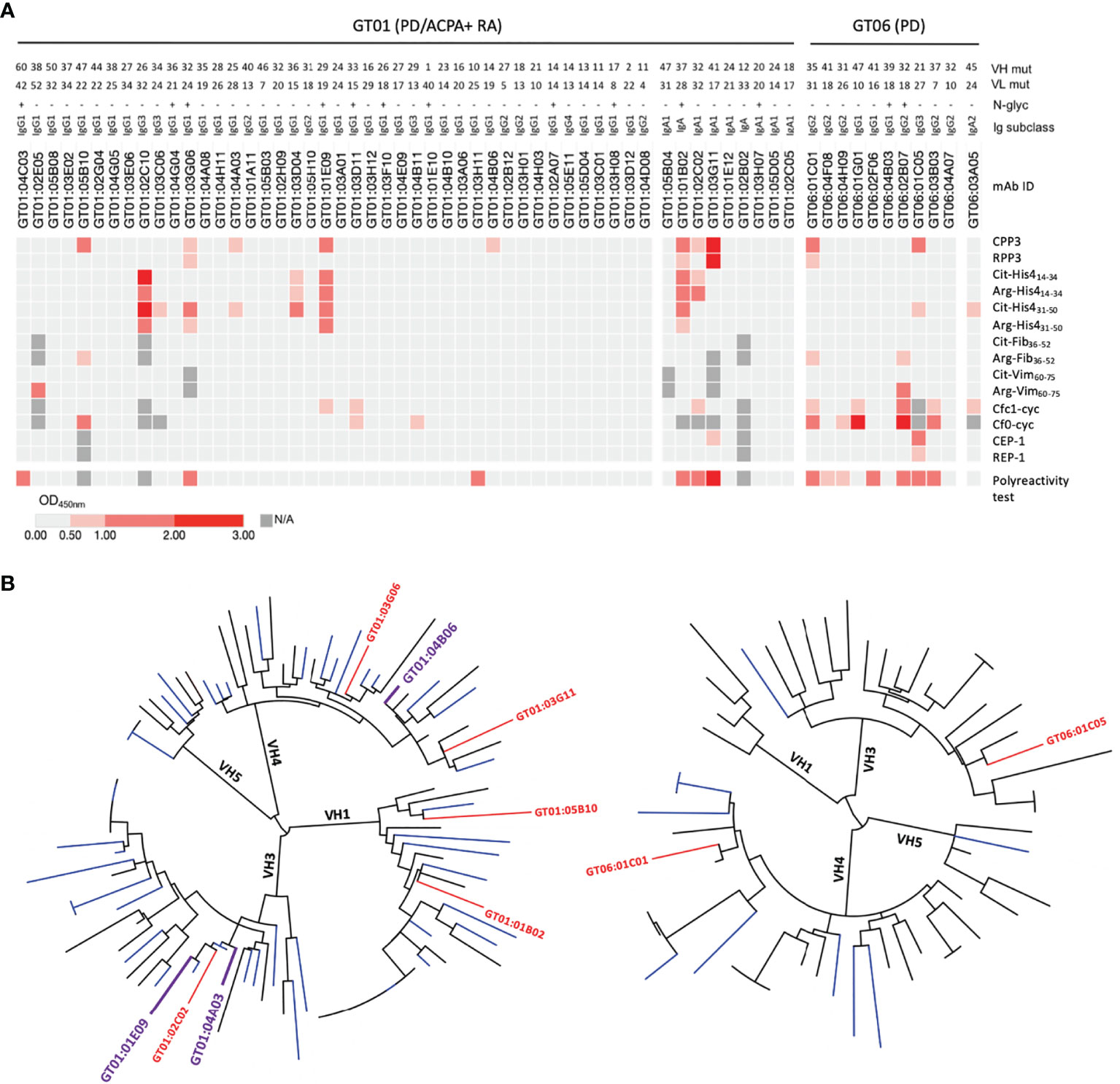
Figure 3 Recombinant gingival tissue mAbs exhibit citrulline-reactivity. (A) ELISA results shown as a heat map, for 52 mAbs generated from a fresh GT biopsy of a PD patient with ACPA+ RA (GT01) and 11 GT mAbs generated from a frozen GT biopsy of a PD patient without RA (GT06); colors indicate binding strength (based on OD values; scale shown at the bottom). Peptide antigens include: citrullinated peptides (and arginine-containing equivalents) from Pg (CPP3) and human proteins. The number of mutations (mut) in the variable heavy (VH) and variable light (VL) chains, presence/absence (+/-) of predicted N-linked glycosylation (N-glyc) motifs (N-X-S/T), and the original Ig subclasses are shown. (B) Phylogenetic trees of the VH chain sequences of GT01 (94 sequences) (left), and GT06 (54 sequences) (right), constructed using Phylogeny.fr with MUSCLE alignment and visualized by iTOL v4.4.2. CPP3+ clones are highlighted in plum (no polyreactivity detected) or in red (show polyreactivity). CPP3, citrullinated P.PAD peptide 3; RPP3, arginine-containing version of CPP3; Cit, citrulline; Arg, arginine; His4, histone-4; Fib, fibrinogen; Vim, vimentin; Cfc1-cyc, cyclic citrullinated filaggrin peptide 1; Cf0-cyc; arginine-containing version of Cfc1-cyc; CEP-1, citrullinated α-enolase peptide 1; REP, arginine-containing version of CEP-1; OD, optical density; N/A, not analysed. Polyreactivity test, ELISA based on the soluble membrane protein fraction of Hek293 cells.
To investigate presence of CPP3+ B cells in the circulation and joints of RA patients, we next analyzed in-house generated mAbs derived from single peripheral blood (PB) (n=36) and synovial fluid (n=139) plasma and memory B cells of 11 ACPA+ RA patients, including 13 CCP2+ clones (30, 31, 33), (Supplementary Table 7). We identified one CCP2+ PB mAb (denoted BVCA1) with strong CPP3-reactivity and no unspecific polyreactivity. BVCA1, originally identified as a strong binder to a citrullinated vimentin peptide (Cit-Vim60-75), also showed multireactivity to citrullinated histone-4 and filaggrin peptides (Figure 4A). Thus, BVCA1 demonstrates cross-reactivity on the monoclonal level, between a bacterial epitope, human citrullinated peptides, and the CCP2 peptide(s). Peptide alignment of CPP3 and Cit-Vim60-75 revealed two identical amino acids adjacent to citrulline (Figure 4B). Comparison of SHM show more mutations and higher R/S ratio in the CPP3+/CCP2+ BVCA1 clone, compared to the CPP3+/CCP2- GT clones (Figure 4C).
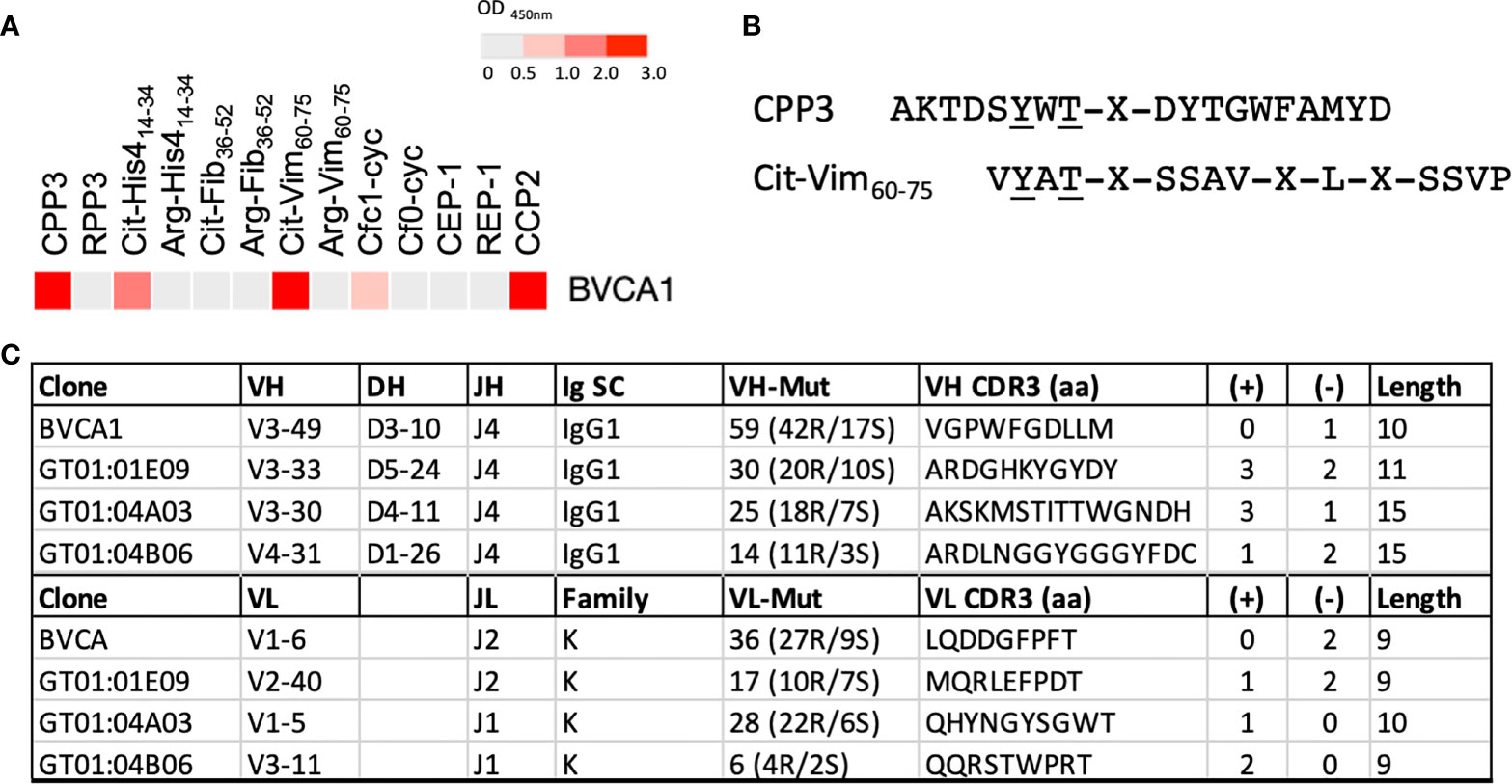
Figure 4 Binding pattern for BVCA1, CPP3/Cit-Vim60-75 peptide alignment, and Ig gene features of BVCA1 versus CPP3+ GT mAbs. (A) ELISA results for BVCA1 shown as a heat map; colors indicate binding strength (based on OD values; scale on top). (B) Peptide alignment of the citrullinated P.PAD peptide CPP3 and a citrullinated human vimentin peptide (Cit-Vim60-75); identical amino acids underlined, X=citrulline. (C) Immunoglobulin (Ig) heavy (H) and light (L) chain gene features of BVCA1 and three CPP3+ GT mAbs; Ig gene variable (V), diversity (D) and joining (J) segment usage, Ig subclass (SC), L chain family, complementary determining region (CDR3) features, and number of mutations (mut) are shown. CPP3, citrullinated P.PAD peptide 3; RPP3, arginine-containing version of CPP3; Cit, citrulline; Arg, arginine; His4, Histone-4; Fib, Fibrinogen; Vim, Vimentin; Cfc1-cyc, cyclic citrullinated filaggrin peptide 1; Cf0-cyc; arginine-containing version of Cfc1-cyc; CEP, citrullinated α-enolase peptide 1; REP, arginine-containing version of CEP-1; CCP2, cyclic citrullinated peptide(s); S, silent; R, replacement; aa, amino acids; (+)/(-), positively/negatively charged aa; Length, number of aa in the CDR3 region.
Based on an epidemiological link between PD and ACPA+ RA, and the unique feature of the periodontal pathogen Porphyromonas gingivalis to generate citrullinated proteins, it has been hypothesised that break of tolerance to citrullinated proteins and production of ACPA may occur in the gum mucosa (7, 14, 20). With the present study, we show an antibody response against the citrullinated Pg PAD enzyme in a subset of early RA, and presence of citrulline-reactive gingival B cells, which display cross-reactivity between Pg and human peptide antigens, supporting the concept of a bacterial/periodontal origin for the ACPA response.
We have focused our investigations on the antibody response to citrullinated P.PAD, first described in RA by Quirke et al. (15). It has been suggested that citrullinated P.PAD could be generated in vivo by autocitrullination, as studies of recombinant full-length P.PAD show autocitrullination (14, 15, 37), similar to human PAD enzymes (38). However, during infection, P.PAD is secreted in vesicles in a truncated form (14, 39, 40) and Konig et al. could show that the truncated form was not autocitrullinated (37), questioning whether autocitrullination would occur in vivo. Still, autocitrullination of the truncated form was reported by Rodriguez et al. (19) and more recent studies show autocitrullinated P.PAD in clinical Pg isolates (17, 18). Citrullination of P.PAD could possibly also occur in the context of NETosis, which is extensive during PD, when secreted P.PAD is likely in close proximity to human PAD enzymes (41). Based on these different scenarios, we hypothesise that an antibody response towards citrullinated P.PAD could be triggered in the inflamed periodontium during Pg infection.
In the large EIRA cohort (2,807 RA patients and 372 controls), we found high serum levels of antibodies to the citrullinated Pg CPP3 peptide derived from P.PAD only in RA patients, but low levels of anti-CPP3 IgG were detected also in a subset of controls. Additionally, low levels of antibodies to the corresponding arginine-containing peptide RPP3 were observed in both RA patients and controls. Periodontal status in EIRA is not known, but we hypothesize that the CPP3+/RPP3+ subsets have (or have had) a Pg infection. In support of this hypothesis, we have previously shown an anti-CPP3 IgG response in rats after Pg infection (42).
High anti-CPP3 IgG levels were mainly recorded in the CCP2+ subset, but did not show a strong correlation with the autoimmune ACPA fine-specificities. Moreover, anti-CPP3 IgG associated with HLA-DRB1 SE, but the association was likely dependent on the co-occurrence of autoimmune ACPA fine-specificities (here detected by the CCP2 test) with a well-known SE-association (26). Smoking on the other hand, a major risk factor for both RA (2, 25) and PD (43), associated significantly with anti-CPP3 antibodies even within the CCP2+ subset. In line with this observation, we have previously seen an increased prevalence of PD specifically in ACPA+ RA patients who smoke (44). Furthermore, anti-CPP3 IgG associated with higher baseline pain, which was not seen for anti-CCP2 IgG. We speculate that this association may reflect a pain-mediating effect of the anti-CPP3 antibodies, something that has previously been shown for ACPA in a mouse model (45).
Importantly, for the first time, we identified gingiva-residing B cells reactive to the citrullinated Pg CPP3 peptide. A number of CPP3+ and CPP3- gingival tissue B-cell clones also showed reactivity with citrullinated peptides derived from human proteins, in particular filaggrin and histone-4, known targets of the ACPA response (1, 46). Notably, the first ACPA test, the so-called anti-perinuclear factor, detected antibodies in RA sera that bound human buccal mucosa, later identified as citrullinated filaggrin (47). Moreover, NETosis with the release of citrullinated histones is a prominent feature of PD (41, 48), and presence of ACPA targeting citrullinated histones have been described in PD (49), in support of a link between periodontal infection and RA-related autoimmunity.
The GT mAbs frequently bound both citrullinated and corresponding arginine-containing peptides, in line with a previous report demonstrating elevated levels of antibodies recognizing both citrullinated and arginine-containing RA candidate autoantigens in PD patients versus non-PD controls (50). Regarding the high frequency of gingival tissue B cells with unspecific polyreactivity observed in our study, we speculate that this could reflect the bacteria-rich environment. Polyreactive germline-encoded IgM that bind both microbes and self-proteins are present in the natural antibody repertoire, and thought to have dual functions in immune defence and -homeostasis (51). Interestingly, the polyreactive gingival tissue clones were class-switched IgG or IgA with SHM, something that has previously also been described for polyreactive broadly neutralizing anti-HIV antibodies (52). Whether the polyreactive gingival tissue clones have functions in immune defence and homeostasis is not known and will be further investigated.
In addition to the CPP3+ gingival tissue B-cell clones, we identified an antibody derived from an RA blood memory B cell that showed cross-reactivity between Pg CPP3, human citrullinated peptides and the CCP2 peptide(s) used in the gold standard clinical test for RA. Peptide alignment pinpoints a shared “citrulline-motif”, previously described for this antibody (53), which could explain the cross-reactivity. In line with our data, Li et al. generated CCP2+ mAbs from circulating plasmablasts of ACPA+ RA patients, and could show cross-reactivity to Pg outer membrane proteins. Interestingly, the CCP2-reactivity was lost in the germline version of the antibody while reactivity to Pg remained (54), suggesting that autoimmunity may have evolved from an initial immune response against the bacteria. Future studies should address the clonal expansion of citrulline-reactive B cells in gingival tissue versus blood and joints.
Based on our serological and molecular data, we propose an etiological model where Pg triggers loss of citrulline-tolerance, via production of anti-Pg antibodies (i.e. RPP3/CPP3) with a potential to cross-react with human citrullinated proteins by mechanisms of molecular mimicry. In genetically susceptible individuals (i.e. HLA-DRB1 SE), autoreactive T cells may drive affinity maturation of this ACPA response towards citrullinated self-proteins (Figure 5).
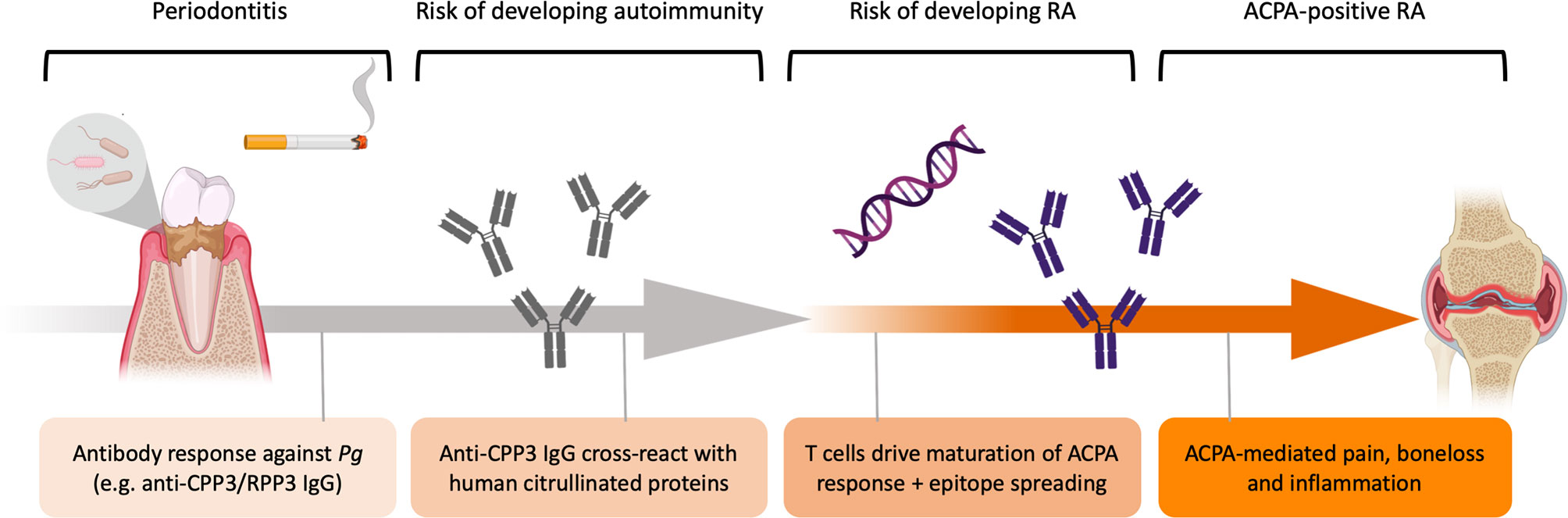
Figure 5 Schematic illustration of the etiological hypothesis linking Pg to ACPA+ RA. P. gingivalis drives gingival inflammation with increased protein citrullination, in the context of cigarette smoking. Immune responses against Pg includes anti-P.PAD antibody responses, such as anti-CPP3/RPP3 IgG, which could cross-react with human citrullinated proteins exposed in the inflamed periodontium by mechanisms of molecular mimicry. In genetically susceptible individuals (e.g. HLA-DRB1 SE), autoreactive T cells drive affinity maturation of the ACPA response targeting self-proteins. High-affinity autoreactive ACPA subsequently bind proteins expressed in the joints and trigger pain, bone loss and synovial inflammation.
An important technical achievement in our study, was the successful isolation of live B cells from frozen gingival biopsies. A similar protocol has been used for single-cell analyses of cells dissociated from cryopreserved synovial tissue (55). However, we observed lower leukocyte yields in our study, which may be explained by long-term storage at -80, and the enzymatic digestion may have degraded cell surface markers, hampering characterization of B-cell subsets from frozen biopsies. Still, B cells from one of the frozen gingival biopsies were successfully used to generate recombinant mAbs.
Analysis of B cells from fresh gingival tissue biopsies showed presence of both memory and plasma cells, in line with previous reports (56). Notably, our sorting strategy focused on CD19+ cells, yet recent reports have described long-lived CD19- plasma cells in bone marrow and intestine (57, 58). When analysing CD138 expression on CD19-/CD3-/CD14- cells in two of the GT biopsies, we were not able to detect a clear population (data not shown). However, patients were few and cell numbers low, thus further investigation is needed to examine the possible presence of long-lived CD19- plasma cells in inflamed gingiva. Our Ig repertoire analysis, demonstrating high numbers of SHM and replacement mutations in the CDRs, are supported by deep sequencing data of PD gingival tissue biopsies, revealing upregulation of genes involved in B-cell activation (59), and suggests antigen-dependent and T-cell driven B-cell responses in the inflamed periodontium. Although we observed some differences regarding B-cell subsets and BCR repertoire between ACPA+ RA/PD and non-RA/PD patients, we could not draw any conclusions due to the small cohort and inclusion of both frozen and fresh biopsies. In addition, we lacked detailed information on the patients donating gingival tissues, including pocked depth at the site of surgery, presence of Pg, age and smoking status, which would be relevant information if making such comparisons.
Another limitation of our study is the lack of periodontal data in the EIRA cohort, which prevented us from studying CPP3/RPP3 IgG in relation to PD status. Also, we did not detect any CCP2+ B cells in gingival tissue, but considering how rare CCP2+ B cells are (31, 60), and the limited number of GT mAbs analysed in our study (n=63 from two PD patients), the presence of CCP2+ B cells in the gum mucosa should be further explored.
Taken together, we show the presence of citrulline-reactive gingival tissue B cells, cross-reactivity between P. gingivalis and human citrullinated peptides on a monoclonal level, and an elevated antibody response to citrullinated P.PAD in CCP2+ RA patients, supporting a scenario where the ACPA response may be partly triggered by oral infection.
The raw data supporting the conclusions of this article will be made available by the authors, upon reasonable request.
The studies involving human participants were reviewed and approved by Regional Ethics Review Board Stockholm, Stockholm, Sweden. The patients/participants provided their informed consent to participate in this study. In line with Swedish law, patient consent in the EIRA cohort was documented in the medical records by respective treating physician. This was done after the patient had received information about the study and after approving participation (consent) in the study.
NS, CV, and NK performed a majority of the laboratory work, with practical guidance from NSi, KA, and KL, and additional scientific guidance from CG and VM. XJ and BB performed association and correlation statistics. EK and LI contributed to ELISA screening and validation. MH and LM-A generated the multiplex array data. RS contributed to the mAb production. SS retrieved EIRA clinical data from the Swedish Rheumatology Quality Register. RH provided the Cit-C1 peptide and related validation. FS provided the BVCA1 Ig sequence, and supervised the work of LP. TY-L had overall responsibility for the collection of GT biopsies and supervised the work of KE, and together with AH, GJ, and AC identified PD and RA/PD patients for the study. JR analysed EIRA clinical data. LA administrates the EIRA study and supervised the work of XJ, BB, and SS. LK administrates the EIRA study together with LA, and initiated development of the multiplex array. LP identified and cloned BVCA1. KA, VM, and CG contributed to the design, discussions and interpretation of data in relation to the B cell, Ig and mAb experiments. KL conceived the study, supervised the work of NS, CV, NK, and EK, and had overall responsibility for the study, including writing the manuscript together with NS and CV. All authors have read, critically reviewed, and approved the final manuscript.
This work was supported by grants from the Swedish Research Council (2017-01696), King Gustav V:s 80-year foundation (FAI-2016-0273), the Swedish Rheumatism foundation (R-931647), Professor Nanna Svartz’s foundation (2019-00292), and the EU/EFPIA Innovative Medicines Initiative Joint Undertaking RTCure (777357). FS and the Institute for Research in Biomedicine are supported by the Helmut Horten Foundation.
Karolinska Institutet has been a partner with Thermo Fisher Scientific within the Innovative Medicines Initiative BTCure, a public–private partnership between the EU and the European Federation of Pharmaceutical Industries (www.BTcure.eu). Thermo Fisher Scientific has contributed to this consortium with in-kind contributions for the development of the multiplex assay used in the study. LM-A is employed by Thermo Fischer Scientific. JR is member of the Thermo Fisher Scientific advisory board. KL is co-inventor of patent: US12/524,465, describing the diagnostic use of the CEP-1 epitope. RH is a co-inventor of patent: US Patent 7 148 020, protecting the use of the CitC1 peptide. GJ was employed by Praktikertjänst AB.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank EIRA study participants, the EIRA study group, and patients donating tissue biopsies to the project, for their contributions. We would also like to thank Gloria Rostvall, Susana Hernandez Machado and Julia Norkko at Karolinska Institutet for managing the EIRA blood samples and biobank, as well as scientists involved in the generation of EIRA data: Dr Leonid Padyukov, Dr Patrick Stolt, and Dr Camilla Bengtsson. Also, Per Matsson, Mats Nystrand and Thomas Schlederer at Thermo Fisher Scientific for scientific support regarding the multiplex platform, and Michael Kramer, Institute for Research in Biomedicine, for work on BVCA1.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.804822/full#supplementary-material
ACPA, anti-citrullinated protein antibodies; CCP, cyclic citrullinated peptide; CDR, complementary determining region; CPP3, citrullinated Porphyromonas gingivalis peptidylarginine deiminase peptide 3; EIRA, epidemiological investigation of rheumatoid arthritis; GT, gingival tissue; mAb, monoclonal antibody; P. PAD, Porphyromonas gingivalis peptidylarginine deiminase; PD, periodontitis; Pg, Porphyromonas gingivalis; RA, rheumatoid arthritis; RPP3, arginine (R)-containing Porphyromonas gingivalis peptide 3; SE, shared epitope (specific HLA-DRB1 alleles); SHM, somatic hypermutation; VH, variable heavy (chain); VL, variable light (chain).
1. Schellekens GA, de Jong BA, van den Hoogen FH, van de Putte LB, van Venrooij WJ. Citrulline is an Essential Constituent of Antigenic Determinants Recognized by Rheumatoid Arthritis-Specific Autoantibodies. 1998. J Immunol (2015) 195(1):8–16. doi: 10.1172/JCI1316
2. Klareskog L, Stolt P, Lundberg K, Källberg H, Bengtsson C, Grunewald J, et al. A New Model for an Etiology of Rheumatoid Arthritis: Smoking May Trigger HLA-DR (Shared Epitope)-Restricted Immune Reactions to Autoantigens Modified by Citrullination. Arthritis Rheumatol (2006) 54(1):38–46. doi: 10.1002/art.21575
3. Berglin E, Johansson T, Sundin U, Jidell E, Wadell G, Hallmans G, et al. Radiological Outcome in Rheumatoid Arthritis Is Predicted by Presence of Antibodies Against Cyclic Citrullinated Peptide Before and at Disease Onset, and by IgA-RF at Disease Onset. Ann Rheum Dis (2006) 65(4):453–8. doi: 10.1136/ard.2005.041376
4. Catrina AI, Svensson CI, Malmström V, Schett G, Klareskog L. Mechanisms Leading From Systemic Autoimmunity to Joint-Specific Disease in Rheumatoid Arthritis. Nat Rev Rheumatol (2017) 13(2):79–86. doi: 10.1038/nrrheum.2016.200
5. Rantapää-Dahlqvist S, de Jong BA, Berglin E, Hallmans G, Wadell G, Stenlund H, et al. Antibodies Against Cyclic Citrullinated Peptide and IgA Rheumatoid Factor Predict the Development of Rheumatoid Arthritis. Arthritis Rheumatol (2003) 48(10):2741–9. doi: 10.1002/art.11223
6. Makrygiannakis D, Hermansson M, Ulfgren AK, Nicholas AP, Zendman AJ, Eklund A, et al. Smoking Increases Peptidylarginine Deiminase 2 Enzyme Expression in Human Lungs and Increases Citrullination in BAL Cells. Ann Rheum Dis (2008) 67(10):1488–92. doi: 10.1136/ard.2007.075192
7. Fuggle NR, Smith TO, Kaul A, Sofat N. Hand to Mouth: A Systematic Review and Meta-Analysis of the Association Between Rheumatoid Arthritis and Periodontitis. Front Immunol (2016) 780:80. doi: 10.3389/fimmu.2016.00080
8. Kindstedt E, Johansson L, Palmqvist P, Koskinen Holm C, Kokkonen H, Johansson I, et al. Association Between Marginal Jawbone Loss and Onset of Rheumatoid Arthritis and Relationship to Plasma Levels of RANKL. Arthritis Rheumatol (2018) 70(4):508–15. doi: 10.1002/art.40394
9. Loutan L, Alpizar-Rodriguez D, Courvoisier DS, Finckh A, Mombelli A, Giannopoulou C. Periodontal Status Correlates With Anti-Citrullinated Protein Antibodies in First-Degree Relatives of Individuals With Rheumatoid Arthritis. J Clin Periodontol (2019) 46(7):690–8. doi: 10.1111/jcpe.13117
10. Mankia K, Cheng Z, Do T, Hunt L, Meade J, Kang J, et al. Prevalence of Periodontal Disease and Periodontopathic Bacteria in Anti-Cyclic Citrullinated Protein Antibody-Positive At-Risk Adults Without Arthritis. JAMA Netw Open (2019) 2(6):e195394. doi: 10.1001/jamanetworkopen.2019.5394
11. Laugisch O, Wong A, Sroka A, Kantyka T, Koziel J, Neuhaus K, et al. Citrullination in the Periodontium–a Possible Link Between Periodontitis and Rheumatoid Arthritis. Clin Oral Investig (2016) 20(4):675–83. doi: 10.1007/s00784-015-1556-7
12. Harvey GP, Fitzsimmons TR, Dhamarpatni AA, Marchant C, Haynes DR, Bartold PM. Expression of Peptidylarginine Deiminase-2 and -4, Citrullinated Proteins and Anti-Citrullinated Protein Antibodies in Human Gingiva. J Periodontal Res (2013) 48(2):252–61. doi: 10.1111/jre.12002
13. How KY, Song KP, Chan KG. Porphyromonas Gingivalis: An Overview of Periodontopathic Pathogen Below the Gum Line. Front Microbiol (2016) 9(7):53. doi: 10.3389/fmicb.2016.00053
14. McGraw WT, Potempa J, Farley D, Travis J. Purification, Characterization, and Sequence Analysis of a Potential Virulence Factor From Porphyromonas Gingivalis, Peptidylarginine Deiminase. Infect Immun (1999) 67(7):3248–56. doi: 10.1128/IAI.67.7.3248-3256.1999
15. Quirke AM, Lugli EB, Wegner N, Hamilton BC, Charles P, Chowdhury M, et al. Heightened Immune Response to Autocitrullinated Porphyromonas Gingivalis Peptidylarginine Deiminase: A Potential Mechanism for Breaching Immunologic Tolerance in Rheumatoid Arthritis. Ann Rheum Dis (2014) 73(1):263–9. doi: 10.1136/annrheumdis-2012-202726
16. Wegner N, Wait R, Sroka A, Eick S, Nguyen KA, Lundberg K, et al. Peptidylarginine Deiminase From Porphyromonas Gingivalis Citrullinates Human Fibrinogen and α-Enolase: Implications for Autoimmunity in Rheumatoid Arthritis. Arthritis Rheumatol (2010) 62(9):2662–72. doi: 10.1002/art.27552
17. Jenning M, Marklein B, Ytterberg J, Zubarev RA, Joshua V, van Schaardenburg D, et al. Bacterial Citrullinated Epitopes Generated by Porphyromonas Gingivalis Infection-a Missing Link for ACPA Production. Ann Rheum Dis (2020) 79(9):1194–202. doi: 10.1136/annrheumdis-2019-216919
18. Stobernack T, Glasner C, Junker S, Gabarrini G, de Smit M, de Jong A, et al. Extracellular Proteome and Citrullinome of the Oral Pathogen Porphyromonas Gingivalis. J Proteome Res (2016) 15(12):4532–43. doi: 10.1021/acs.jproteome.6b00634
19. Rodríguez SB, Stitt BL, Ash DE. Expression of Peptidylarginine Deiminase From Porphyromonas Gingivalis in Escherichia Coli: Enzyme Purification and Characterization. Arch Biochem Biophys (2009) 488(1):14–22. doi: 10.1016/j.abb.2009.06.010
20. Rosenstein ED, Greenwald RA, Kushner LJ, Weissmann G. Hypothesis: The Humoral Immune Response to Oral Bacteria Provides a Stimulus for the Development of Rheumatoid Arthritis. Inflamm (2004) 28(6):311–8. doi: 10.1007/s10753-004-6641-z
21. Lundberg K, Kinloch A, Fisher BA, Wegner N, Wait R, Charles P, et al. Antibodies to Citrullinated Alpha-Enolase Peptide 1 are Specific for Rheumatoid Arthritis and Cross-React With Bacterial Enolase. Arthritis Rheumatol (2008) 58(10):3009–19. doi: 10.1002/art.23936
22. Kharlamova N, Jiang X, Sherina N, Potempa B, Israelsson L, Quirke AM, et al. Antibodies to Porphyromonas Gingivalis Indicate Interaction Between Oral Infection, Smoking, and Risk Genes in Rheumatoid Arthritis Etiology. Arthritis Rheumatol (2016) 68(3):604–13. doi: 10.1002/art.39491
23. Johansson L, Sherina N, Kharlamova N, Potempa B, Larsson B, Israelsson L, et al. Concentration of Antibodies Against Porphyromonas Gingivalis Is Increased Before the Onset of Symptoms of Rheumatoid Arthritis. Arthritis Res Ther (2016) 18(1):201. doi: 10.1186/s13075-016-1100-4
24. Mikuls TR, Thiele GM, Deane KD, Payne JB, O'Dell JR, Yu F, et al. Porphyromonas Gingivalis and Disease-Related Autoantibodies in Individuals at Increased Risk of Rheumatoid Arthritis. Arthritis Rheumatol (2012) 64(11):3522–30. doi: 10.1002/art.34595
25. Stolt P, Bengtsson C, Nordmark B, Lindblad S, Lundberg I, Klareskog L, et al. Quantification of the Influence of Cigarette Smoking on Rheumatoid Arthritis: Results From a Population Based Case-Control Study, Using Incident Cases. Ann Rheum Dis (2003) 62(9):835–41. doi: 10.1136/ard.62.9.835
26. Reed E, Hedström AK, Hansson M, Mathsson-Alm L, Brynedal B, Saevarsdottir S, et al. Presence of Autoantibodies in “Seronegative” Rheumatoid Arthritis Associates With Classical Risk Factors and High Disease Activity. Arthritis Res Ther (2020) 22(1):170. doi: 10.1186/s13075-020-02191-2
27. Saevarsdottir S, Wedrén S, Seddighzadeh M, Bengtsson C, Wesley A, Lindblad S, et al. Patients With Early Rheumatoid Arthritis Who Smoke are Less Likely to Respond to Treatment With Methotrexate and Tumor Necrosis Factor Inhibitors: Observations From the Epidemiological Investigation of Rheumatoid Arthritis and the Swedish Rheumatology Register Cohorts. Arthritis Rheumatol (2011) 63(1):26–36. doi: 10.1002/art.27758
28. Mizraji G, Segev H, Wilensky A, Hovav AH. Isolation, Processing and Analysis of Murine Gingival Cells. J Vis Exp (2013) 77):e50388. doi: 10.3791/50388
29. amara k, Israelsson L, Stålesen R, Sahlström P, Steen J, Malmström V, et al. A Refined Protocol for Identifying Citrulline-Specific Monoclonal Antibodies From Single Human B Cells From Rheumatoid Arthritis Patient Material. Bio-protocol (2019) 9(17):e3347. doi: 10.21769/BioProtoc.3347
30. Steen J, Forsström B, Sahlström P, Odowd V, Israelsson L, Krishnamurthy A, et al. Recognition of Amino Acid Motifs, Rather Than Specific Proteins, by Human Plasma Cell–Derived Monoclonal Antibodies to Posttranslationally Modified Proteins in Rheumatoid Arthritis. Arthritis Rheumatol (2019) 71(2):196–209. doi: 10.1002/art.40699
31. Titcombe PJ, Wigerblad G, Sippl N, Zhang N, Shmagel AK, Sahlström P, et al. Pathogenic Citrulline-Multispecific B Cell Receptor Clades in Rheumatoid Arthritis. Arthritis Rheumatol (2018) 70(12):1933–45. doi: 10.1002/art.40590
32. Germar K, Fehres CM, Scherer HU, van Uden N, Pollastro S, Yeremenko N, et al. Generation and Characterization of Anti-Citrullinated Protein Antibody-Producing B Cell Clones From Rheumatoid Arthritis Patients. Arthritis Rheumatol (2019) 71(3):340–50. doi: 10.1002/art.40739
33. Lloyd KA, Steen J, Amara K, Titcombe PJ, Israelsson L, Lundström SL, et al. Variable Domain N-Linked Glycosylation and Negative Surface Charge Are Key Features of Monoclonal ACPA: Implications for B-Cell Selection. Eur J Immunol (2018) 48(6):1030–45. doi: 10.1002/eji.201747446
34. Hansson M, Mathsson L, Schlederer T, Israelsson L, Matsson P, Nogueira L, et al. Validation of a Multiplex Chip-Based Assay for the Detection of Autoantibodies Against Citrullinated Peptides. Arthritis Res Ther (2012) 14(5):R201. doi: 10.1186/ar4039
35. Ossipova E, Cerqueira CF, Reed E, Kharlamova N, Israelsson L, Holmdahl R, et al. Affinity Purified Anti-Citrullinated Protein/Peptide Antibodies Target Antigens Expressed in the Rheumatoid Joint. Arthritis Res Ther (2014) 16(4):R167. doi: 10.1186/ar4683
36. Jardine JG, Sok D, Julien JP, Briney B, Sarkar A, Liang CH, et al. Minimally Mutated HIV-1 Broadly Neutralizing Antibodies to Guide Reductionist Vaccine Design. PloS Pathog (2016) 12(8):e1005815. doi: 10.1371/journal.ppat.1005815
37. Konig MF, Paracha AS, Moni M, Bingham CO 3rd, Andrade F. Defining the Role of Porphyromonas Gingivalis Peptidylarginine Deiminase (PPAD) in Rheumatoid Arthritis Through the Study of PPAD Biology. Ann Rheum Dis (2015) 74(11):2054–61. doi: 10.1136/annrheumdis-2014-205385
38. Andrade F, Darrah E, Gucek M, Cole RN, Rosen A, Zhu X. Autocitrullination of Human Peptidyl Arginine Deiminase Type 4 Regulates Protein Citrullination During Cell Activation. Arthritis Rheumatol (2010) 62(6):1630–40. doi: 10.1002/art.27439
39. Veith PD, Chen YY, Gorasia DG, Chen D, Glew MD, O'Brien-Simpson NM, et al. Porphyromonas Gingivalis Outer Membrane Vesicles Exclusively Contain Outer Membrane and Periplasmic Proteins and Carry a Cargo Enriched With Virulence Factors. J Proteome Res (2014) 13(5):2420–32. doi: 10.1021/pr401227e
40. Sato K, Yukitake H, Narita Y, Shoji M, Naito M, Nakayama K. Identification of Porphyromonas Gingivalis Proteins Secreted by the Por Secretion System. FEMS Microbiol Letters (2013) 338(1):68–76. doi: 10.1111/1574-6968.12028
41. Magán-Fernández A, Rasheed Al-Bakri SM, O'Valle F, Benavides-Reyes C, Abadía-Molina F, Mesa F. Neutrophil Extracellular Traps in Periodontitis. Cells (2020) 9(6). doi: 10.3390/cells9061494
42. Eriksson K, Lönnblom E, Tour G, Kats A, Mydel P, Georgsson P, et al. Effects by Periodontitis on Pristane-Induced Arthritis in Rats. J Transl Med (2016) 14(1):311. doi: 10.1186/s12967-016-1067-6
43. Tomar SL, Asma S. Smoking-Attributable Periodontitis in the United States: Findings From NHANES III. National Health and Nutrition Examination Survey. J Periodontol (2000) 71(5):743–51. doi: 10.1902/jop.2000.71.5.743
44. Eriksson K, Nise L, Alfredsson L, Catrina AI, Askling J, Lundberg K, et al. Seropositivity Combined With Smoking is Associated With Increased Prevalence of Periodontitis in Patients With Rheumatoid Arthritis. Ann Rheum Dis (2018) 77(8):1236. doi: 10.1136/annrheumdis-2017-212091
45. Jurczak A, Delay L, Barbier J, Simon N, Krock E, Sandor K, et al. Antibody-Induced Pain-Like Behavior and Bone Erosion: Links to Subclinical Inflammation, Osteoclast Activity, and Acid-Sensing Ion Channel 3-Dependent Sensitization. Pain (2021). doi: 10.1097/j.pain.0000000000002543
46. Pratesi F, Dioni I, Tommasi C, Alcaro MC, Paolini I, Barbetti F, et al. Antibodies From Patients With Rheumatoid Arthritis Target Citrullinated Histone 4 Contained in Neutrophils Extracellular Traps. Ann Rheum Dis (2014) 73(7):1414–22. doi: 10.1136/annrheumdis-2012-202765
47. Sebbag M, Simon M, Vincent C, Masson-Bessière C, Girbal E, Durieux JJ, et al. The Antiperinuclear Factor and the So-Called Antikeratin Antibodies are the Same Rheumatoid Arthritis-Specific Autoantibodies. J Clin Invest (1995) 95(6):2672–9. doi: 10.1172/JCI117969
48. Vitkov L, Hartl D, Minnich B, Hannig M. Janus-Faced Neutrophil Extracellular Traps in Periodontitis. Front Immunol (2017) 8:1404. doi: 10.3389/fimmu.2017.01404
49. Janssen KMJ, de Smit MJ, Withaar C, Brouwer E, van Winkelhoff AJ, Vissink A, et al. Autoantibodies Against Citrullinated Histone H3 in Rheumatoid Arthritis and Periodontitis Patients. J Clin Periodontol (2017) 44(6):577–84. doi: 10.1111/jcpe.12727
50. de Pablo P, Dietrich T, Chapple IL, Milward M, Chowdhury M, Charles PJ, et al. The Autoantibody Repertoire in Periodontitis: A Role in the Induction of Autoimmunity to Citrullinated Proteins in Rheumatoid Arthritis? Ann Rheum Dis (2014) 73(3):580–6. doi: 10.1136/annrheumdis-2012-202701
51. Grönwall C, Vas J, Silverman G. Protective Roles of Natural IgM Antibodies. Front Immunol (2012) 3:66. doi: 10.3389/fimmu.2012.00066
52. Haynes BF, Fleming J, St Clair EW, Katinger H, Stiegler G, Kunert R, et al. Cardiolipin Polyspecific Autoreactivity in Two Broadly Neutralizing HIV-1 Antibodies. Science (2005) 308(5730):1906–8. doi: 10.1126/science.1111781
53. Sahlström P, Hansson M, Steen J, Amara K, Titcombe PJ, Forsström B, et al. Different Hierarchies of Anti-Modified Protein Autoantibody Reactivities in Rheumatoid Arthritis. Arthritis Rheumatol (2020) 72(10):1643–57. doi: 10.1002/art.41385
54. Li S, Yu Y, Yue Y, Liao H, Xie W, Thai J, et al. Autoantibodies From Single Circulating Plasmablasts React With Citrullinated Antigens and Porphyromonas Gingivalis in Rheumatoid Arthritis. Arthritis Rheumatol (2016) 68(3):614–26. doi: 10.1002/art.39455
55. Donlin LT, Rao DA, Wei K, Slowikowski K, McGeachy MJ, Turner JD, et al. Methods for High-Dimensional Analysis of Cells Dissociated From Cryopreserved Synovial Tissue. Arthritis Res Ther (2018) 20(1):139. doi: 10.1186/s13075-018-1631-y
56. Mahanonda R, Champaiboon C, Subbalekha K, Sa-Ard-Iam N, Rattanathammatada W, Thawanaphong S, et al. Human Memory B Cells in Healthy Gingiva, Gingivitis, and Periodontitis. J Immunol (2016) 197(3):715–25. doi: 10.4049/jimmunol.1600540
57. Landsverk OJ, Snir O, Casado RB, Richter L, Mold JE, Réu P, et al. Antibody-Secreting Plasma Cells Persist for Decades in Human Intestine. J Exp Med (2017) 214(2):309–17. doi: 10.1084/jem.20161590
58. Halliley JL, Tipton CM, Liesveld J, Rosenberg AF, Darce J, Gregoretti IV, et al. Long-Lived Plasma Cells Are Contained Within the CD19(-)CD38(hi)CD138(+) Subset in Human Bone Marrow. Immunity (2015) 43(1):132–45. doi: 10.1016/j.immuni.2015.06.016
59. Kim YG, Kim M, Kang JH, Kim HJ, Park JW, Lee JM, et al. Transcriptome Sequencing of Gingival Biopsies From Chronic Periodontitis Patients Reveals Novel Gene Expression and Splicing Patterns. Hum Genomics (2016) 10(1):28. doi: 10.1186/s40246-016-0084-0
Keywords: rheumatoid arthritis (RA), anti-citrullinated protein antibodies (ACPA), Porphyromonas gingivalis (Pg), periodontitis (PD), monoclonal antibodies (mAbs), B cells
Citation: Sherina N, de Vries C, Kharlamova N, Sippl N, Jiang X, Brynedal B, Kindstedt E, Hansson M, Mathsson-Alm L, Israelsson L, Stålesen R, Saevarsdottir S, Holmdahl R, Hensvold A, Johannsen G, Eriksson K, Sallusto F, Catrina AI, Rönnelid J, Grönwall C, Yucel-Lindberg T, Alfredsson L, Klareskog L, Piccoli L, Malmström V, Amara K and Lundberg K (2022) Antibodies to a Citrullinated Porphyromonas gingivalis Epitope Are Increased in Early Rheumatoid Arthritis, and Can Be Produced by Gingival Tissue B Cells: Implications for a Bacterial Origin in RA Etiology. Front. Immunol. 13:804822. doi: 10.3389/fimmu.2022.804822
Received: 29 October 2021; Accepted: 17 March 2022;
Published: 20 April 2022.
Edited by:
Carmelo Carmona-Rivera, National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIH), United StatesReviewed by:
Piotr Mydel, University of Bergen, NorwayCopyright © 2022 Sherina, de Vries, Kharlamova, Sippl, Jiang, Brynedal, Kindstedt, Hansson, Mathsson-Alm, Israelsson, Stålesen, Saevarsdottir, Holmdahl, Hensvold, Johannsen, Eriksson, Sallusto, Catrina, Rönnelid, Grönwall, Yucel-Lindberg, Alfredsson, Klareskog, Piccoli, Malmström, Amara and Lundberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karin Lundberg, S2FyaW4uTHVuZGJlcmdAa2kuc2U=
†Professor Anca I. Catrina passed away in February 2021 during the preparation of the manuscript
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.