 Güliz Tuba Barut
Güliz Tuba Barut Marco Kreuzer
Marco Kreuzer Rémy Bruggmann
Rémy Bruggmann Artur Summerfield1,2
Artur Summerfield1,2 Stephanie C. Talker
Stephanie C. Talker- 1Institute of Virology and Immunology, Bern, Switzerland
- 2Department of Infectious Diseases and Pathobiology, Vetsuisse Faculty, University of Bern, Bern, Switzerland
- 3Interfaculty Bioinformatics Unit and Swiss Institute of Bioinformatics, University of Bern, Bern, Switzerland
Dendritic and monocytic cells co-operate to initiate and shape adaptive immune responses in secondary lymphoid tissue. The complexity of this system is poorly understood, also because of the high phenotypic and functional plasticity of monocytic cells. We have sequenced mononuclear phagocytes in mesenteric lymph nodes (LN) of three adult cows at the single-cell level, revealing ten dendritic-cell (DC) clusters and seven monocyte/macrophage clusters with clearly distinct transcriptomic profiles. Among DC, we defined LN-resident subsets and their progenitors, as well as subsets of highly activated migratory DC differing in transcript levels for T-cell attracting chemokines. Our analyses also revealed a potential differentiation path for cDC2, resulting in a cluster of inflammatory cDC2 with close transcriptional similarity to putative DC3 and monocyte-derived DC. Monocytes and macrophages displayed sub-clustering mainly driven by pro- or anti-inflammatory expression signatures, including a small cluster of cycling, presumably self-renewing, macrophages. With this transcriptomic snapshot of LN-derived mononuclear phagocytes, we reveal functional properties and differentiation trajectories in a “command center of immunity”, and identify elements that are conserved across species.
1 Introduction
Dendritic cells (DC) are known as central instructors of adaptive immunity (1), which is initiated in specialized areas of secondary lymphoid tissues (2). It is widely accepted that also monocytic cells contribute to shape adaptive immune responses (3, 4). Being about ten times more frequent than DC in peripheral blood, monocytes fulfill multiple functions in the induction and resolution of inflammation (5), and have also been implicated in antigen presentation in lymph nodes (6).
Bona fide DCs can be delineated from other mononuclear phagocytes by their surface expression of Flt3 receptor tyrosine kinase, a hematopoietic cytokine receptor essential for DC differentiation (7–11). Distinction of three DC subsets (cDC1, cDC2, pDC) is well established and based on differences in ontogeny, expression of key genes, phenotype, and function (12–14). However, discussions are ongoing if pDC should be re-classified as innate lymphocytes (15). Murine studies have suggested that cDC1 and cDC2 are specialized to induce CD8-T cell/Th1 responses and Th2/Th17 responses, respectively (16, 17). As such, murine cDC1 were described to be especially efficient at cross-presentation (18). Recent findings have strengthened the idea that also pDC engage in T-cell stimulation, notably following initial IFN-I production (19). To identify DC subsets in non-mouse/non-human species (namely pig, horse, and cow), we have previously analyzed expression of key genes that were found to be evolutionarily conserved in DC subsets, including essential transcription factors and FLT3 (9–11).
Across species, monocyte subsets are less well defined and most likely represent a differentiation continuum rather than developmentally distinct populations (20). Nevertheless, in humans and cattle, nonclassical and intermediate monocytes (ncM and intM) can be distinguished from classical monocytes (cM) based on expression of CD14/CD16 and certain key genes such as CCR2 (cM) and NR4A1/CX3CR1 (ncM and intM) (11, 21–23). Notably, we recently demonstrated that bovine monocyte subsets appear to be very similar to their human counterparts (23, 24), with transcriptomic analyses suggesting a role of ncM in antiviral immunity and a specialization of intM in antigen presentation. In tissues, however, the subset identity of monocytes gets blurred, as they are differentiating towards monocyte-derived DC (moDC) and monocyte-derived macrophages, the latter of which can be transcriptionally very similar to long-lived and self-renewing tissue-resident macrophages of embryonic origin (3, 25, 26).
Single-cell RNA sequencing (scRNA-seq) has enabled a relatively unbiased view on immune-cell composition in complex tissues (27, 28), revealing for example considerable heterogeneity within human cDC2, including an inflammatory DC subset with close similarity to moDC (29–32), and contributing to the recent discovery of the separate DC lineage DC3 – cells described as phenotypic and functional intermediates between cDC2 and monocytes (33–35).
Moreover, scRNA-seq enables high-level comparative immunology through in-depth analysis of non-model species, promising exciting insights into basic and evolutionarily conserved functions of immune cells. Having previously performed detailed transcriptomic analyses of dendritic and monocytic cells isolated from blood of cattle, both ex vivo and following in-vitro stimulation (11, 23, 36), we provide here the first scRNA-seq analysis of the mononuclear phagocyte compartment in bovine lymph node. With the mesenteric lymph node we have chosen a secondary lymphoid tissue that is expected to contain a considerable fraction of activated immune cells under steady-state conditions – giving us the opportunity to study the mononuclear phagocyte system at its best – where antigens are presented, T cells get instructed, and apoptotic cells are disposed of.
2 Materials and methods
2.1 Isolation of bovine mesenteric-lymph-node cells
Mesenteric lymph nodes (mesLN) draining the small intestine were collected from three cows that were slaughtered at a nearby butchery in three consecutive weeks (MLN2306: RedHolstein, 3.5 years; MLN3006: Simmental, 8.5 years; MLN0707: Holstein, 4 years), and immediately placed into ice-cold PBS containing 1 mM EDTA (Invitrogen, ThermoFisher, Basel, Switzerland) (PBS-EDTA). After removal of fat and connective tissue, lymph nodes were cut into small pieces and minced using a gentleMACS Dissociator (Miltenyi Biotec Swiss AG, Solothurn, Switzerland). After a washing step with cold PBS-EDTA (300 x g, 10 min, 4°C), the cells were incubated in PBS containing 0.1 mg/mL DNase I (Worthington-Biochem, BioConcept, Basel, Switzerland), for 15 min at room temperature. Thereafter, cells were washed (300 x g, 10 min, 4°C), with DMEM supplemented with 5% FBS (Gibco, Life Technologies, Basel, Switzerland), and the suspension was passed through a cell strainer (70 µm). This washing step was repeated, and cells were resuspended in PBS-EDTA (room temperature) and layered onto Ficoll Paque (1.077 g/mL; GE Healthcare Europe GmbH). After centrifugation (800 x g, 25 min, 20°C), cells at the interface were collected and washed twice with cold PBS-EDTA (400 x g, 10 min, 4°C). Subsequently, the cells were counted and stained for fluorescence-activated cell sorting (FACS) as described below.
2.2 Fluorescence-activated cell sorting (FACS) of Flt3+ and CD172ahigh cells
Staining of isolated lymph-node cells was performed with 2 x 108 cells in 50 mL Falcon tubes and encompassed three incubation steps, each for 20 min at 4°C (incubation volume 2 mL). In-between incubation steps, cells were washed (300 x g, 10 min, 4°C) with 40 mL of BD Cell Wash (BD Biosciences, Allschwil, Switzerland). In order to block Fc receptors, cells were incubated with purified 50 µg/mL bovine IgG (Bethyl laboratories, Montgomery, USA), before adding CD172a (CC149, mIgG2b; Bio Rad, Cressier, Switzerland) and His-tagged Flt3L in a subsequent incubation step. Bovine His-tagged Flt3L (NCBI NM_181030.2) was produced as previously described (8, 10).
In the third and final step, cells were incubated with anti-IgG2b-AF647 (Molecular Probes, Thermo Fischer, Basel, Switzerland), anti-His-PE (mIgG1) (Miltenyi Biotec Swiss AG, Solothurn, Switzerland), and LIVE/DEAD™ Fixable Near-IR stain (Thermo Fisher Scientific, Basel, Switzerland). After a final washing step, cells were resuspended in BD Cell Wash and delivered to the Flow Cytometry and Cell Sorting Facility (FCCS) of the University of Bern, where 1x105 viable cells (Flt3+ and/or CD172ahigh) were sorted using a MoFlo Astrios EQ cell sorter equipped with five lasers (Beckman Coulter Eurocenter SA, Nyon, Switzerland). After sorting, cells were spun down (300 x g, 10 min, 4°C), re-suspended in 200 µl of PBS containing 0.4% BSA and viability was assessed microscopically using Trypan Blue staining. All three samples were confirmed to be free of visible debris and doublets and to have a viability > 95%. Immediately after counting, cells were submitted to the Next Generation Sequencing Platform of the University of Bern for generation of 10x Genomics sequencing libraries and subsequent sequencing.
2.3 Single-cell RNA sequencing (10x Genomics)
Library preparation was done in three consecutive weeks at the Next Generation Sequencing (NGS) Platform at the University of Bern. The three libraries were stored at minus 80°C, before being pooled and sequenced in one run. GEM generation & barcoding, reverse transcription, cDNA amplification and 3’ gene expression library generation steps were all performed according to the Chromium Next GEM Single Cell 3’ Reagent Kits v3.1 User Guide (10x Genomics CG000204 Rev D) with all stipulated 10x Genomics reagents. Specifically, 33.0 µL of each cell suspension (500 cells/µL) and 10.2 µL of nuclease-free water were used for a targeted cell recovery of 10’000 cells. GEM generation was followed by a GEM-reverse transcription incubation, a clean-up step and 11 cycles of cDNA amplification. The resulting cDNA was evaluated for quantity and quality using a Thermo Fisher Scientific Qubit 4.0 fluorometer with the Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, Q32854) and an Advanced Analytical Fragment Analyzer System using a Fragment Analyzer NGS Fragment Kit (Agilent, DNF-473), respectively. Thereafter, 3′ gene expression libraries were constructed using a sample index PCR step of 13-15 cycles. At the end of the protocol, an additional 0.8x bead-based cleanup of the libraries was performed. The generated cDNA libraries were tested for quantity and quality using fluorometry and capillary electrophoresis as described above. The cDNA libraries were pooled and sequenced with a loading concentration of 300 pM, paired-end and single-indexed, on an illumina NovaSeq 6000 sequencer with a shared NovaSeq 6000 S2 Reagent Kit (100 cycles; illumina 20012862). The read set-up was as follows: read 1: 28 cycles, i7 index: 8 cycles, i5: 0 cycles and read 2: 91 cycles. The quality of the sequencing runs was assessed using illumina Sequencing Analysis Viewer (illumina version 2.4.7) and all base-call files were demultiplexed and converted into FASTQ files using illumina bcl2fastq conversion software v2.20. An average of 787,553,242 reads/library were obtained.
2.4 Analysis of scRNA-seq data
Mapping and counting of the UMIs were performed using Cell Ranger (version 3.0.2, 10x Genomics) with the reference genome ARS-UCD1.2 from Ensembl to build the necessary index files. Subsequent analysis was performed in R (version 4.0.2) (37). The Scater package (version 1.14) (38) was used to assess the proportion of ribosomal and mitochondrial genes as well as the number of detected genes. Cells were considered as outliers and filtered out if the value of the proportion of expressed mitochondrial genes or the number of detected genes deviated more than three median absolute deviations from the median across all cells. After quality control, the sample from MLN2306 retained 6604 cells, the sample from MLN3006 retained 3956 cells, and the sample from MLN0707 retained 3649 cells. Normalization between samples was done with the deconvolution method of Lun et al. (39) using the package Scran (version 1.14) (40). Samples were integrated with the FindIntegrationAnchors function of the package Seurat (version 3.1) based on the first 30 principal components (PCs; Seurat default options) (41). Graph-based clustering was done with the FindNeighbors and FindClusters functions of the Seurat package using the first 40 PCs from the dimensionality reduction step, as determined by the JackStraw procedure. The Clustree package (version 0.4) (42) was used to determine the resolution resulting in clustering concurring with the presumed cell types, which was 1.2. In order to identify up- or down-regulated genes between clusters, FindAllMarkers was applied to the dataset. This function returns all differentially expressed genes per cluster. Clusters were then manually annotated on the basis of these marker genes. Trajectory analysis was performed using the R package Monocle 3 (43–45). Following dimensionality reduction (UMAP) and Louvain clustering (k = 10 nearest neighbors) using the function “cluster_cells”, the trajectory was calculated using the function “learn_graph”. In order to find genes differentially expressed along selected trajectories, the function “choose_cells” was applied followed by the function “graph_test”. Default parameters were used unless stated otherwise.
2.5 Preparation of figures
Figures were prepared using FlowJo version 10 (FlowJo LLC, Ashland, OR), R version 4.1.1, and Inkscape (https://www.inkscape.org). Single-cell RNA-seq data was visualized in feature plots, dot plots, scatter plots, violin plots, and single-cell heatmaps using the R packages “Seurat” (default parameters, unless stated otherwise) and “ggplot2”. Prior to visualization in heatmaps, data of subsetted clusters was scaled and centered using the ScaleData function of Seurat. For improved contrast in feature plots, feature-specific contrast levels were calculated based on quantiles (q10, q90) of non-zero expression. Code is available from the visualization vignette of the Seurat package (https://www.satijalab.org/seurat/).
3 Results
3.1 Distinct clustering of dendritic and monocytic cells
Single-cell transcriptomes were obtained from sorted Flt3+ and/or CD172ahigh mesenteric-lymph-node cells of three healthy cows (Figure 1A and Supplementary File 1) and bioinformatically processed as outlined in Materials and Methods. The integrated dataset was used for further analyses. A resolution of 1.2 was chosen, resulting in 24 different clusters (Figure 1B and Supplementary File 1). The integrated dataset split by sample, as well as the non-integrated datasets are shown in Supplementary File 1.
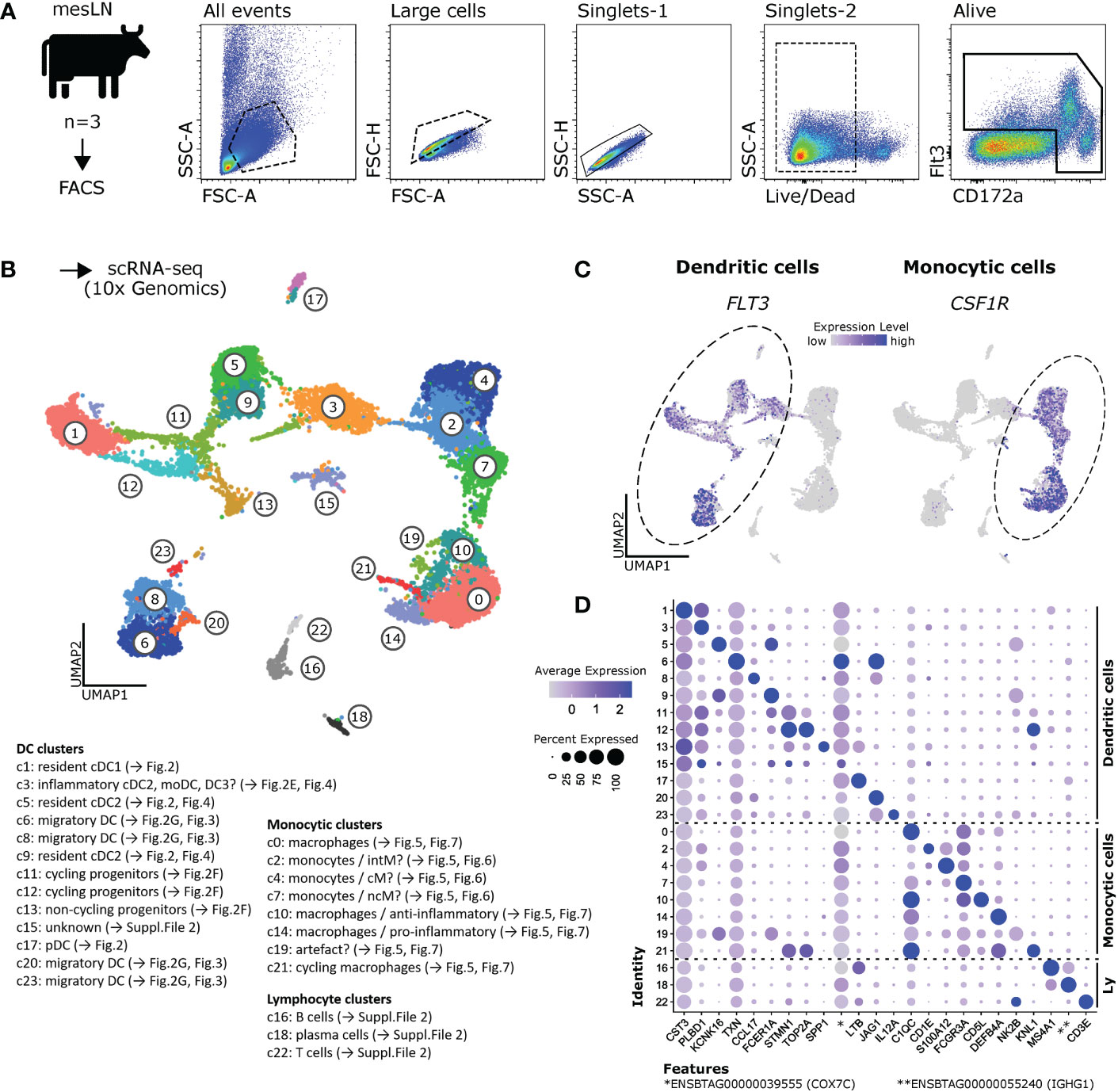
Figure 1 Single-cell RNA-seq of bovine mononuclear phagocytes. (A) Mononuclear phagocytes were sorted by FACS as CD172ahigh and/or Flt3+ cells from mesenteric lymph nodes of three cows and subjected to 10x Genomics scRNA-seq. (B) Data of three animals were integrated and analyzed at a resolution of 1.2, resulting in 24 different clusters. (C) Visualization of FLT3 and CSF1R in feature plots, revealing clusters of dendritic and monocytic cells, respectively. (D) Dot plot shows expression of top marker gene (lowest p_val_adj, followed by highest avg_log2FC), as determined by Seurat’s FindAllMarkers, across all clusters. Complete gene lists are given in Supplementary File 2.
When visualizing the expression of the DC marker FLT3 and the monocytic marker CSF1R in UMAP plots (Figure 1C), we found that most of these 24 clusters could be grouped into major clusters of putative DC (c1, c3, c5, c6, c8, c9, c11, c12, c13, c15, c17, c20, c23) and putative monocytic cells (clusters c0, c2, c4, c7, c10, c14, c19, c21). Notably, clusters 3 and 15, both assigned to DC, contained a subset of cells expressing CSF1R and lacking FLT3 expression.
Clusters lacking both FLT3 and CSF1R expression, were found to express CD79B (c16 and c18) or CD3E (c22), alongside other B- and T-cell markers, respectively. Notably, c18 likely contained plasma cells, as indicated by the expression of IRF4, PRDM1 (Blimp-1), JCHAIN and immunoglobulin genes. These plasma cells expressed CD27 and TNFRSF17, two surface molecules that could be targeted for flow cytometric detection of plasma cells in cattle.
Cluster-defining marker genes, as determined by Seurat’s FindAllMarkers function, are listed in Supplementary File 2. Expression of the top marker gene of each cluster (lowest p_val_adj, followed by highest avg_log2FC) is visualized in a dotplot for all clusters in Figure 1D.
3.2 Subset-specific gene transcription defines resident cDC1, cDC2 and pDC
In accordance with the phenotype and transcriptome of bona fide DC subsets recently identified in blood of cattle (11), distinct clusters of lymph-node-derived DC could be defined by expression of ANPEP (CD13), FCER1A and CD4 (Figure 2A).
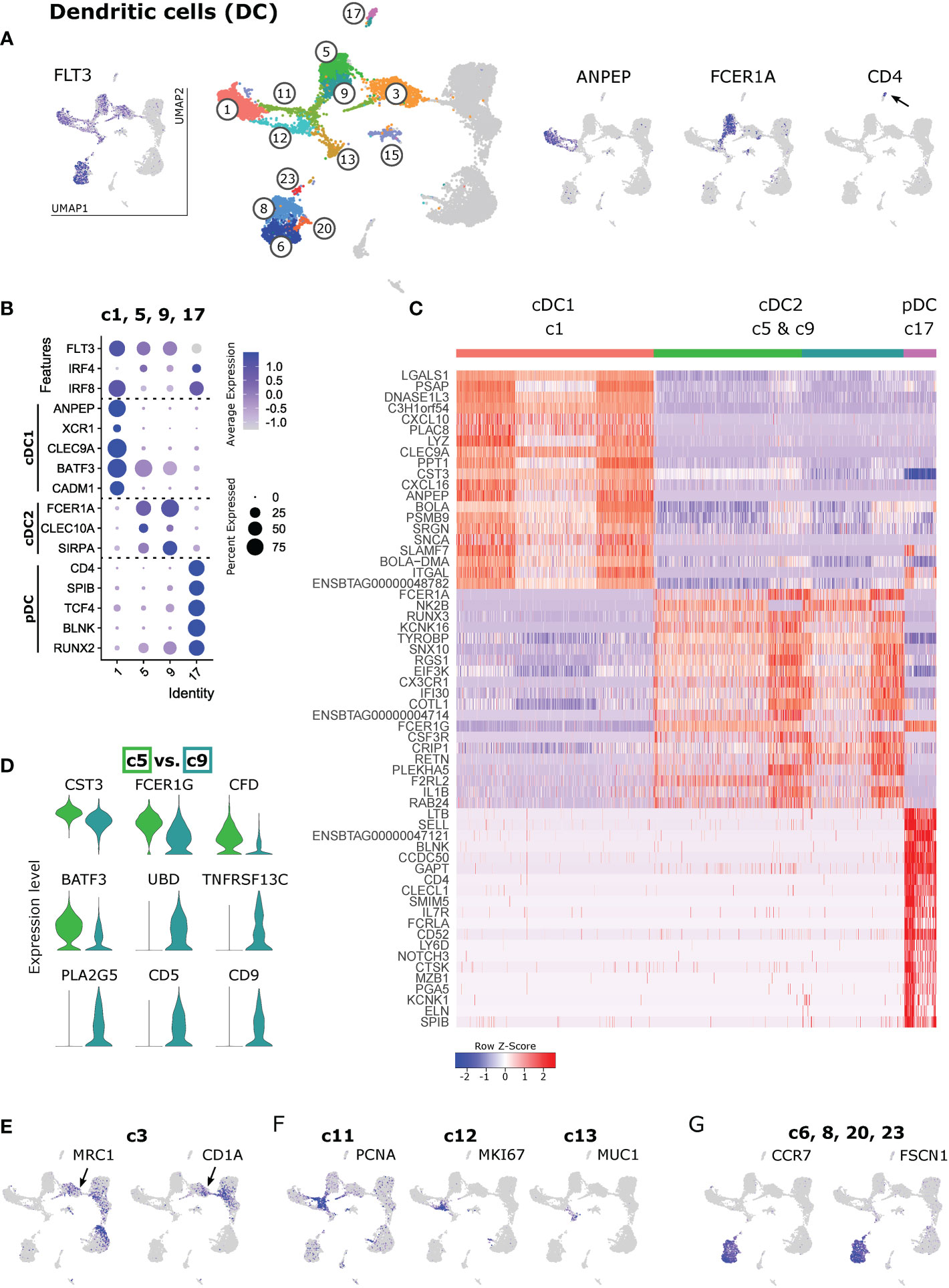
Figure 2 Dendritic cells. (A) Visualization of key genes in feature plots to identify clusters containing total DCs (FLT3), cDC1 (ANPEP, cluster 1), cDC2 (FCER1A, clusters 5 and 9) and pDC (CD4, cluster 17). (B) Dot plot of key subset-defining genes in selected DC clusters. (C) Heatmap of the top 20 (p_adj) differentially expressed genes in clusters identified as cDC1 (c1), cDC2 (c5 & c9) and pDC (c17). (D) Top genes differentially expressed between cDC2 clusters c5 and c9, visualized as violin plots (linear y-axis). (E–G) Expression of selected signature genes for DC clusters that could not be clearly assigned to a DC subset based on key gene expression.
As shown in Figure 2B, the co-expression of subset-specific key genes clearly confirmed cluster 1 as cDC1 (ANPEP, XCR1, CLEC9A, BATF3, CADM1), clusters 5 and 9 as cDC2 (FCER1A, CLEC10A, SIRPA), and cluster 17 as pDC (CD4, SPIB, TCF4, BLNK, RUNX2). A heatmap of the top 20 (adjusted p-value) differentially expressed genes between these clusters (c1, c5 & c9, c17) is shown in Figure 2C. The complete gene list is given in Supplementary File 3. Apart from the genes mentioned above, the top subset-specific genes included LGALS1, PSAP, DNASE1L3, CXCL10, LYZ for cDC1, NK2B, RUNX3, CX3CR1 for cDC2, and LTB, SELL, CLECL1 and IL7R for pDC.
The two cDC2 clusters (c5 and c9) differed in the expression of various immune-relevant genes (Figure 2D and Supplementary File 4), such as CST3, FCER1G, CFD, and BATF3 (higher expression in c5), and UBD, TNFRSF13C, PLA2G5, CD5 and CD9 (clearly expressed in c9, but almost absent from c5). Notably, this difference in CD5 and CD9 transcription should be evaluated at the protein level for suitability to distinguish bovine cDC2 subsets with flow cytometry.
Selected subset-specific key genes shown in Figure 2B were also visualized for the complete dataset (Supplementary File 5). Clusters 3, 6, 8, 13, 15, 20 and 23 lacked transcripts for most of these subset-defining marker genes. Among DC, c3 exclusively contained cells expressing MRC1 and CD1A (Figure 2E). Clusters 11 and 12 appeared to contain both cDC1 and cDC2 and stood out by their high expression of cell cycle genes such as PCNA (c11), and MKI67 (c12) (Figure 2F). In fact, c11 and c12 predominantly expressed genes associated with the G1-S phase and the G2-M phase of the cell cycle, respectively (Supplementary File 2). Furthermore, as presented in the UMAP projection (Figure 2A), c13 conveyed the impression of giving rise to c11 and c12 that later diverge to meet cDC1 (c1) and cDC2 (c9). In cluster 13, however, we did not detect transcripts related to an active cell cycle. Instead, c13 stood out by high expression of AATF (cell-cycle control) as well as of CD9, CD164L2, ANXA1, ANXA2, and CLEC10A, and exclusive expression of MUC1, RORC and SLC14A1 among other genes (Figure 2F and Supplementary File 2).
Finally, despite their lack of DC-subset defining transcripts (Supplementary File 5), clusters 6 and 8 showed by far the highest levels of FLT3 expression, alongside exclusive expression of CCR7 and FSCN1 (Figure 2G). In fact, their high levels of CCR7 and FSCN1 expression suggest that these cells comprise migratory DC that have recently migrated from intestinal tissue to the mesenteric lymph node.
3.3 Subsets of CCR7high migratory DC defined by chemokine expression
Cells in clusters 6, 8, 20 and 23, showing an isolated projection in the UMAP plot, stood out by expressing high levels of various activation- and maturation-related genes (Figure 3A and Supplementary File 2). Over 1500 genes were exclusively expressed or significantly upregulated (p_adj < 0.05) in these migratory DC (c6 & c8 & c20 & c23) when compared against resident DC (c1 & c5 & c9 & c17) (Supplementary File 6). This includes genes involved in cytoskeleton regulation (FSCN1, SAMSN1, MARCKSL1), T-cell or B-cell co-stimulation (CD83, TNFSF13B, CD40) and T-cell regulation (IDO1, CD274, IL4I1).
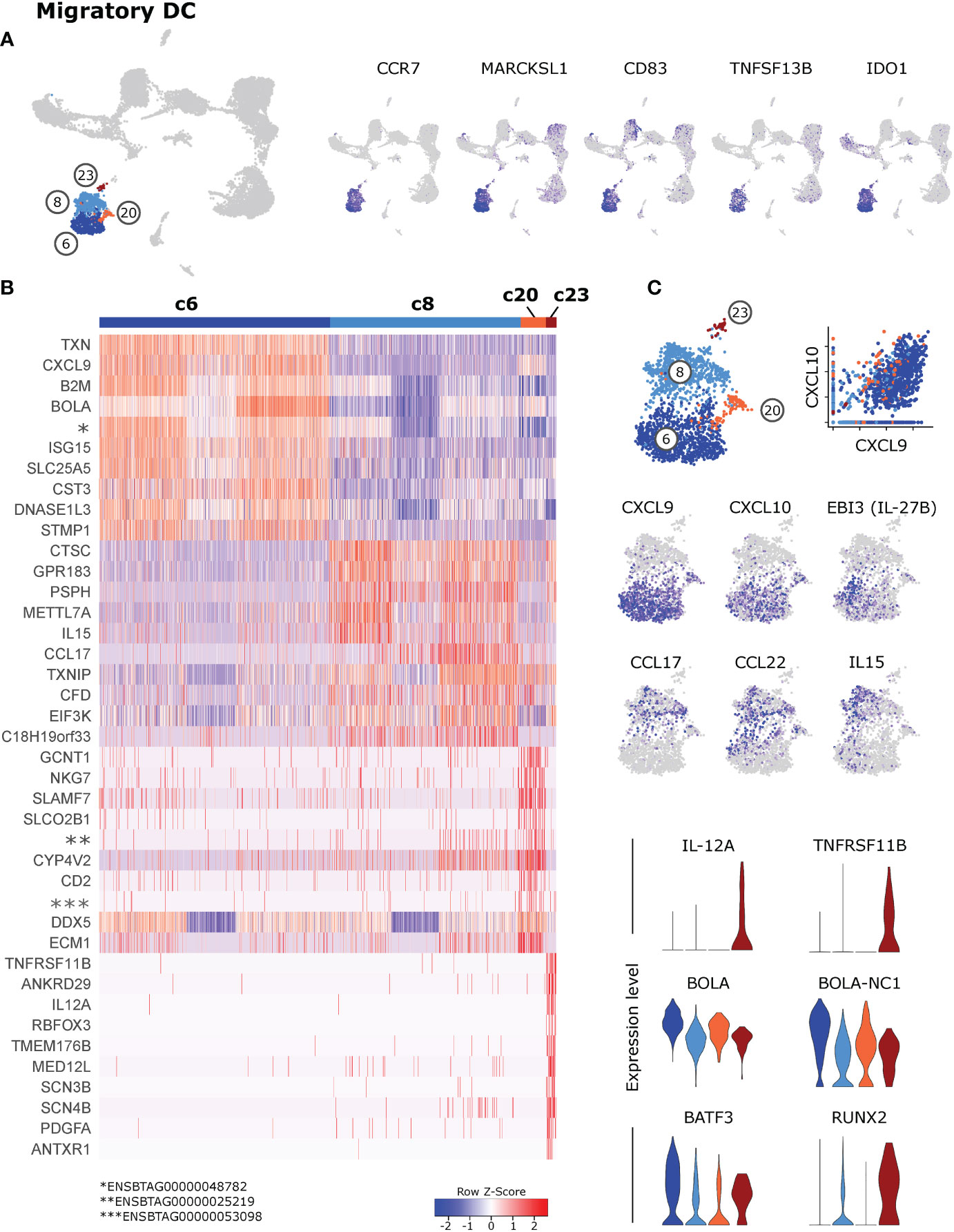
Figure 3 Migratory dendritic cells. (A) Feature plots illustrating high expression of selected activation-related genes in CCR7-expressing migratory DC (c6, c8, c20, c23). (B) Heatmap of the top 10 (p_adj) differentially expressed genes in clusters of CCR7-expressing DC. (C) Visualization of selected genes specifically expressed in clusters of CCR7-expressing DC.
Subclusters within CCR7high migratory DC differed in the expression of T-cell attracting chemokines and T-cell activating cytokines (Figures 3B, C), suggesting a division of labor between subsets of migratory DC (for complete gene lists see Supplementary File 7).
Cluster 6 stood out by high expression of T-cell attracting CXCL9 and CXCL10 (46, 47), alongside ISG15 and MHC-I-associated genes. Notably, also TXN was specifically expressed in c6, encoding for thioredoxin, which has been suggested to regulate the Th1/Th2 balance, among other immuno-relevant effects (48). Moreover, IL27 and EBI3 (IL-27B) were specifically expressed in a subcluster of c6. Cells in cluster 8 were clearly enriched in transcripts for CCL17 and CCL22, presumably attracting Th2 and Th17 cells as well as Tregs (49), and in IL15 transcripts, promoting survival and proliferation of T cells and NK cells. Notably, the very small but distinct cluster 23, exclusively displayed expression of IL12A and TNFRSF11B. Although transcripts for most DC-lineage defining key genes were only weakly detected in these migratory DC clusters, overall gene expression would suggest that cDC2 are enriched in cluster 8 and that cDC1 are enriched in c6. Also the relatively high level of BATF3, BOLA and BOLA-NC1 expression in cluster 6 would point towards cDC1, whereas the high level of RUNX2 in cluster 23 may indicate the presence of pDC (Figure 3C and Supplementary File 7).
3.4 Co-clustering of inflammatory cDC2, monocyte-derived DC and putative DC3
Positioned in-between cDC2 (c5, c9) and monocytes (c2) in the UMAP projection, FLT3+ dendritic cells in cluster 3 expressed genes exclusively shared with either cDC2 (e.g. F2RL2) or monocytic cells (e.g. MRC1). Notably, a considerable fraction of cells in cluster 3 appeared to co-express these genes, including FLT3 and CSF1R (Figure 4A).
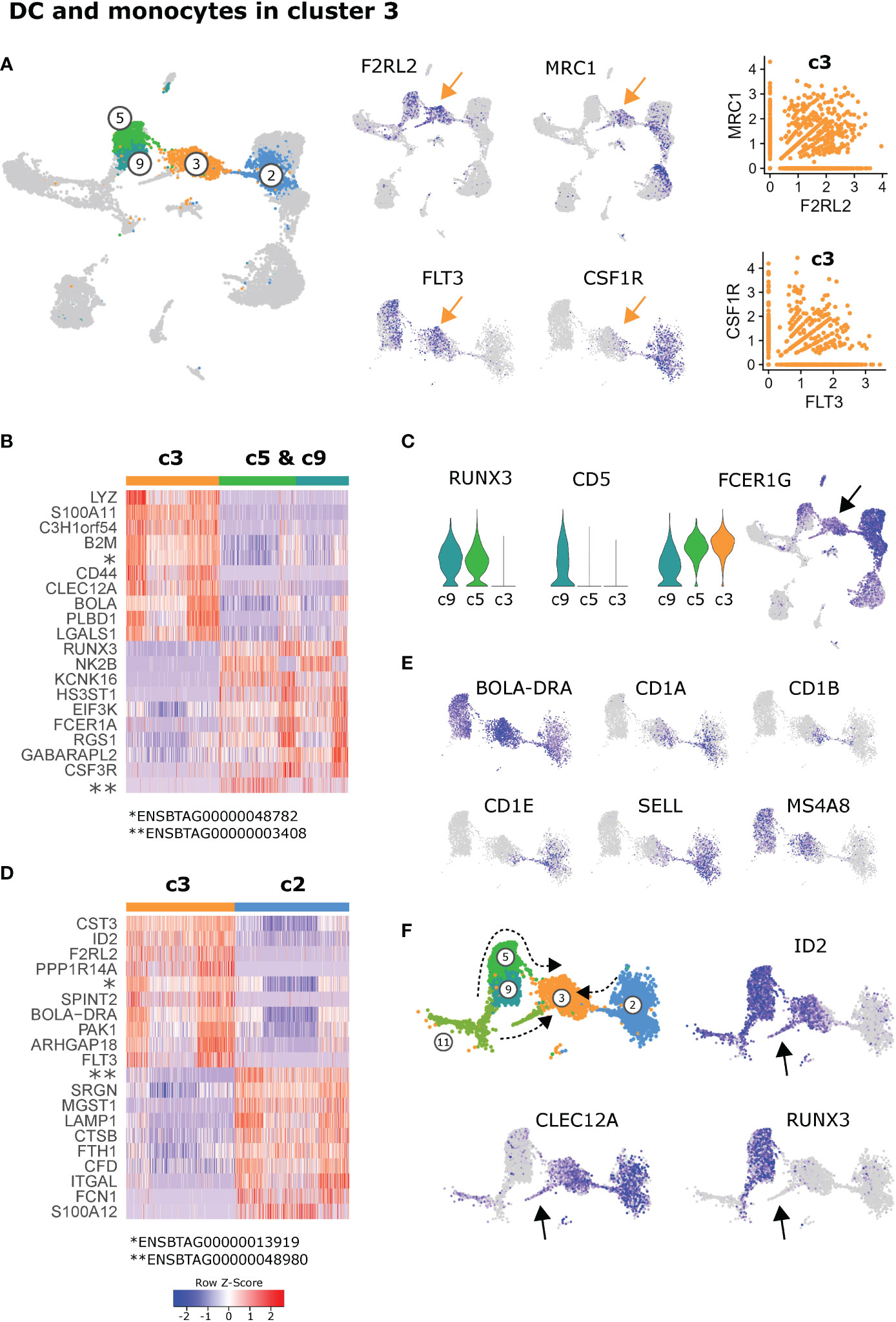
Figure 4 Dendritic cells and monocytes in cluster 3. (A) Expression of DC-associated and monocyte-associated genes in cluster 3. (B) Heatmap of the top 10 (p_adj) differentially expressed genes between cluster 3 and the cDC2 clusters c5 & c9. (C) Visualization of selected genes differentially expressed between c3, c5 and c9. Arrow indicates cluster 3. (D) Heatmap of the top 10 (p_adj) differentially expressed genes between cluster 3 and the monocytic cluster 2. (E) Visualization of selected genes in feature plots showing c2, c3, c5, and c9. (F) Proposed differentiation pathways of inflammatory cDC2 (c11→c9→c5→c3), moDC (c2→c3) and DC3 (c11→c3) indicated by dashed arrows. Arrows in feature plots indicate location of putative DC3 progenitors.
When compared against all other clusters, the most significant signature genes of cluster 3 included PLBD1 (shared with cDC1), CKB and VIM (Supplementary File 2). While PLBD1 might be involved in the generation of lipid-based inflammatory mediators (50), CKB is implicated in immunometabolism of cells with high and fluctuating energy demands (51, 52), and vimentin (VIM) is described to be required for activation of the NLRP3 inflammasome (53).
When cluster 3 was compared with cDC2 (c5 & c9), the antibacterial and pro-inflammatory gene transcription of c3 became even more apparent, with high expression of LYZ (lysozyme) and S100A11 (an alarmin), alongside XDH, described to promote NLRP3 inflammasome activation (54). (Figure 4B and Supplementary File 8). Among the most significant genes with lower expression in c3 compared to cDC2 (c5 & c9) were RUNX3, NK2B, FCER1A and CSF3R.
It is likely that cluster 3 represents inflammatory cDC2, which have recently been described in mice (31). While we could hardly detect any expression of FCER1A, we found high expression of FCER1G in putative inflammatory cDC2 (c3). Notably, FCER1G was also expressed in cluster 5 of resident cDC2, with expression increasing from c9 via c5 towards c3. With RUNX3 and CD5 expression absent from c3, this may indicate a differentiation pathway from c9 (RUNX3+CD5+FCER1Gdim) via c5 (RUNX3+CD5-FCER1G+) towards c3 (RUNX3-CD5-FCER1Ghigh) (Figures 4C, F).
Compared to the monocyte cluster 2, c3 had higher expression of e.g. CST3 and ID2 (Figure 4D), both predominantely expressed by DC clusters, and higher expression of a number of genes coding for MHC-II molecules (Supplementary File 8). Genes with lower expression in c3 compared to monocytes (c2) coded for C-C motif chemokine 23 (ENSBTAG00000048980), a granule-associated proteoglycan (SRGN) and CD107a (LAMP1), as well as cathepsin B (CTSB), numerous inflammation-related proteins, and surface molecules CD11a (ITGAL) and CD172a (SIRPA) (Figure 4D and Supplementary File 8).
Moreover, while all cells in cluster 3 expressed high levels of BOLA-DRA, mainly cells bridging between c3 and c2 were found to express CD1A, CD1B and CD1E (Figure 4E), suggesting the presence of monocyte-derived DC (moDC) specialized in lipid antigen presentation in cluster 3. In line with their blood-borne monocyte-origin, these putative moDC expressed SELL (CD62L) which mediates LN entry via high endothelial venules.
Notably, while MS4A8 was found to be highly expressed selectively in cDC2 (c5, c9) and putative inflammatory cDC2 (c3, left part), it appeared to be absent from putative moDC (c3, right part). The function of MSA48 is poorly characterized, but it was recently found to be highly upregulated in bovine pregnant endometrium (55) and in blood of Salmonella-infected pigs (56).
Alongside inflammatory DC originating from resident cDC2, and moDC originating from blood-borne monocytes, cells of the recently described distinct DC lineage DC3 (33, 34) may be present in cluster 3. Indeed, the observation that cycling (PCNA expressing) pre-DC (c11) branch towards cluster 3 (Figures 2A, F), would support the presence of DC3 as a developmentally distinct linage within cluster 3 (Figure 4F). In line with DC3 progenitors described in humans (34), this separate branch of DC progenitors expressed CLEC12A alongside ID2. Moreover it lacked detectable RUNX3 expression (indicated by black arrows in Figure 4F). Certainly, more detailed analyses are warranted to untangle what seems to be transcriptomic co-clustering of inflammatory cDC2, monocyte-derived DC and DC3 in bovine mesenteric lymph node.
Cluster 15, another cluster positioned in-between DC and monocytes in the UMAP plot, did not show any specific expression of marker genes (Supplementary File 2). Instead, these cells stood out by a low number of detected UMIs and a low number of detected genes, while the proportion of reads mapping to ribosomal and mitochondrial genes was comparable to other clusters (data not shown).
3.5 Separate clustering of monocytes and macrophages
Like in humans, monocytes in blood of cattle are divided into classical and nonclassical monocytes, and an intermediate subset, based on CD14 and CD16 expression (11, 22, 23). In bovine lymph nodes, CSF1R and SIRPA expressing monocytic cells appeared to predominantly express either CD14 (c4, c14) or FCGR3A (CD16; c7, c10) (Figure 5A).
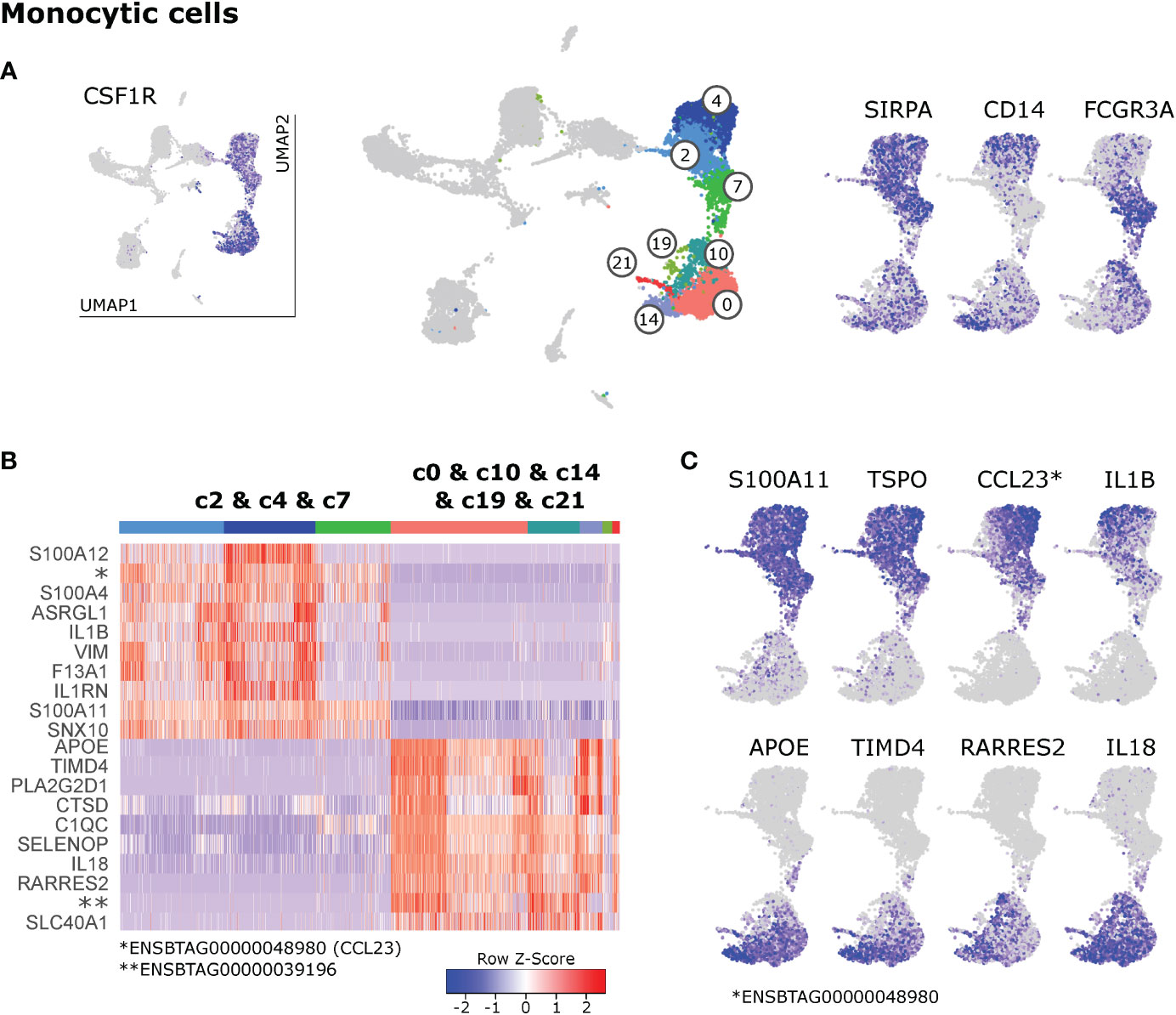
Figure 5 Monocytic cells. (A) Visualization of key genes that define monocytes and their subsets in blood of cattle. (B) Heatmap of the top 10 (p_adj) differentially expressed genes between monocyte clusters (c2 & c4 & c7) and macrophage clusters (c0 & c10 & c14 & c19 & c21). (C) Visualization of selected genes enriched in monocytes (top row) and macrophages (bottom row).
However, the pattern of CD14/FCGR3A expression did not reflect the observed division of monocytic clusters into two main islands in the UMAP projection: clusters 2 & 4 & 7 and clusters 0 & 10 & 14 & 19 & 21. Differential gene expression between these two groups of clusters, revealed that one group (2&4&7) was clearly enriched in genes associated with classical monocytes found in blood of cattle (23) such as S100A11, S100A12 and VIM, whereas the other group (0&10&14&19&21) was strongly enriched in transcripts for genes commonly associated with macrophages (APOE, TIMD4, CD68) (Figure 5B and Supplementary File 9). Notably, putative monocytes exclusively expressed high levels of a gene recently annotated as CCL23 (ENSBTAG00000048980) and expressed high levels of TSPO and pro-inflammatory IL1B, whereas putative macrophages stood out by expressing RARRES2 (Chemerin), IL18, and anti-inflammatory PLA2G2D1 (Figures 5B, C).
3.6 Pro-and anti-inflammatory monocyte clusters
Monocyte clusters (c2, c4, c7) differed in the expression of genes related to inflammation and antigen presentation (Figure 6 and Supplementary File 10). Top gene transcripts enriched in cluster 4 were reminiscent of pro-inflammatory classical monocytes in blood of cattle (Figure 6A). When visualizing these genes in feature plots, expression gradients became apparent with highest expression levels for S100A8, S100A12, VCAN, and DEFB7 in the top right corner of cluster 4 and tendentially higher expression of IL1B and IL1RN in the top left corner of cluster 4. Yet another expression pattern could be observed for HIF1A and SLC2A3, with decreasing expression from top to bottom of cluster 4 (Figure 6B and Supplementary File 10). Cluster 2 was significantly enriched in transcripts related to antigen presentation such as CD1E, BOLA-DRA, CD1A, CD74, and BOLA-DQA1 (Figure 6A). Notably, expression of CD1E, CD1A and CD1B was mainly detected in cells of cluster 2 that seemed to bridge towards dendritic cells (i.e. cluster 3, see also Figure 4E). Moreover, several C-type lectin receptors, most prominently CLEC6A, showed increased expression in cells of cluster 2 (Figure 6C and Supplementary File 10). Cluster 7 was significantly enriched in gene transcripts associated with nonclassical monocytes in blood of cattle, such as C1QA, C1QB, C1QC, FCGR3A, and CX3CR1. Notably, expression of MS4A7 was shared between clusters 2 and 7, and was almost absent from pro-inflammatory cluster 4 (Figure 6D and Supplementary File 10).
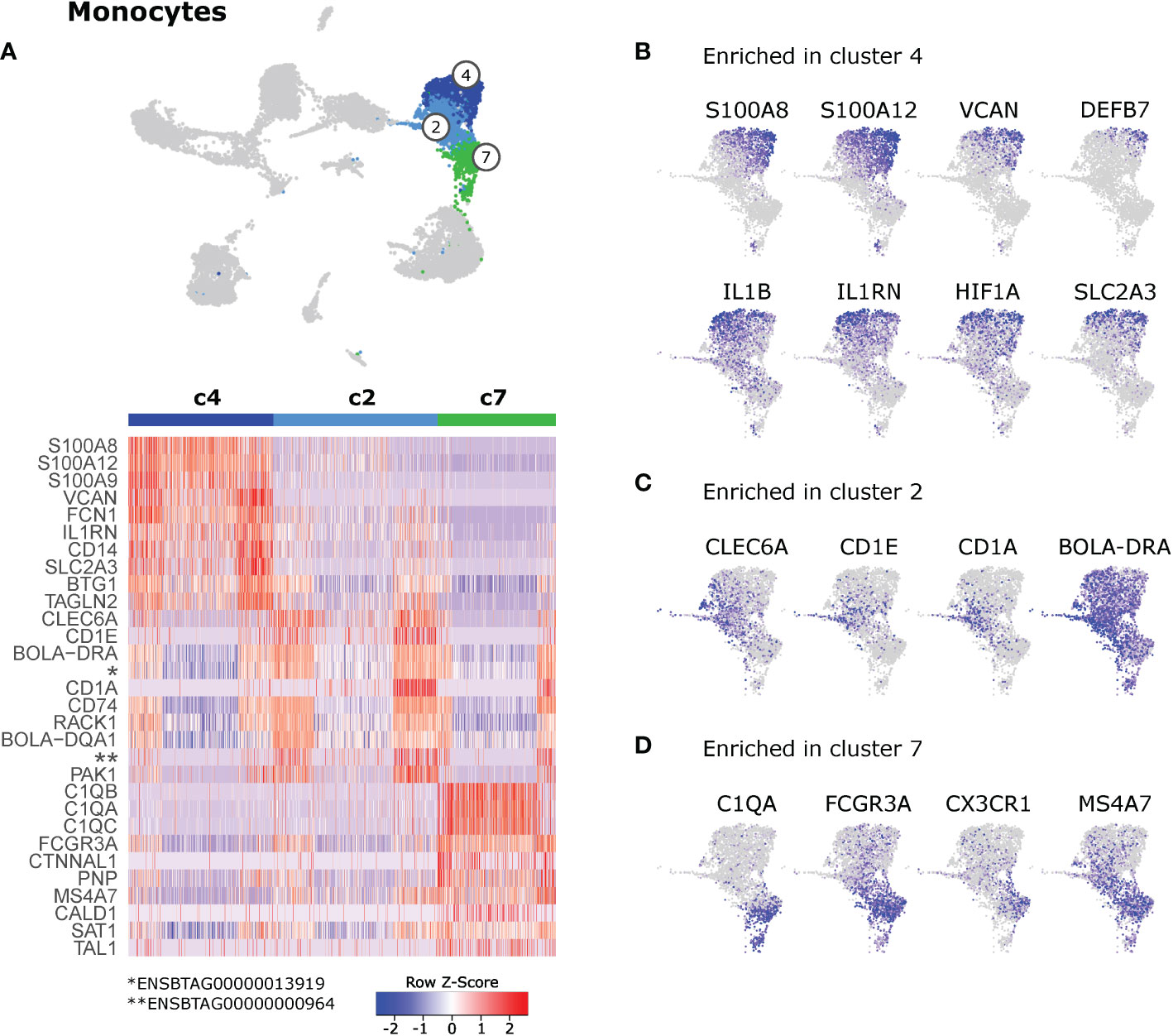
Figure 6 Monocytes. (A) Heatmap of the top 10 (p_adj) differentially expressed genes between monocyte clusters (c2, c4, c7). (B–D) Visualization of selected genes enriched in cluster 4 (B), cluster 2 (C), and cluster 7 (D).
3.7 Macrophage clusters
The majority of macrophages clustered together as cluster 0. While no genes were found to be exclusively expressed in c0, the clusters surrounding c0 showed more pronounced differential gene expression signatures (Figure 7A). Notably, cells in cluster 10 appeared to uniquely express CD5L, SIGLEC1 (CD169) and CD163 (Figures 7A, B). Also genes encoding the phagocytic receptors MRC1 (CD206) and CLEC4F (57) were predominantly expressed in c10, as was HMOX1, encoding a heme-degrading enzyme with anti-inflammatory effects (58). Moreover, cells in c10 expressed significantly increased levels of regakine-1 (ENSBTAG00000010155), a CC-chemokine found to be constitutively present at high concentrations in bovine plasma and to attract neutrophils and lymphocytes in vitro (59).
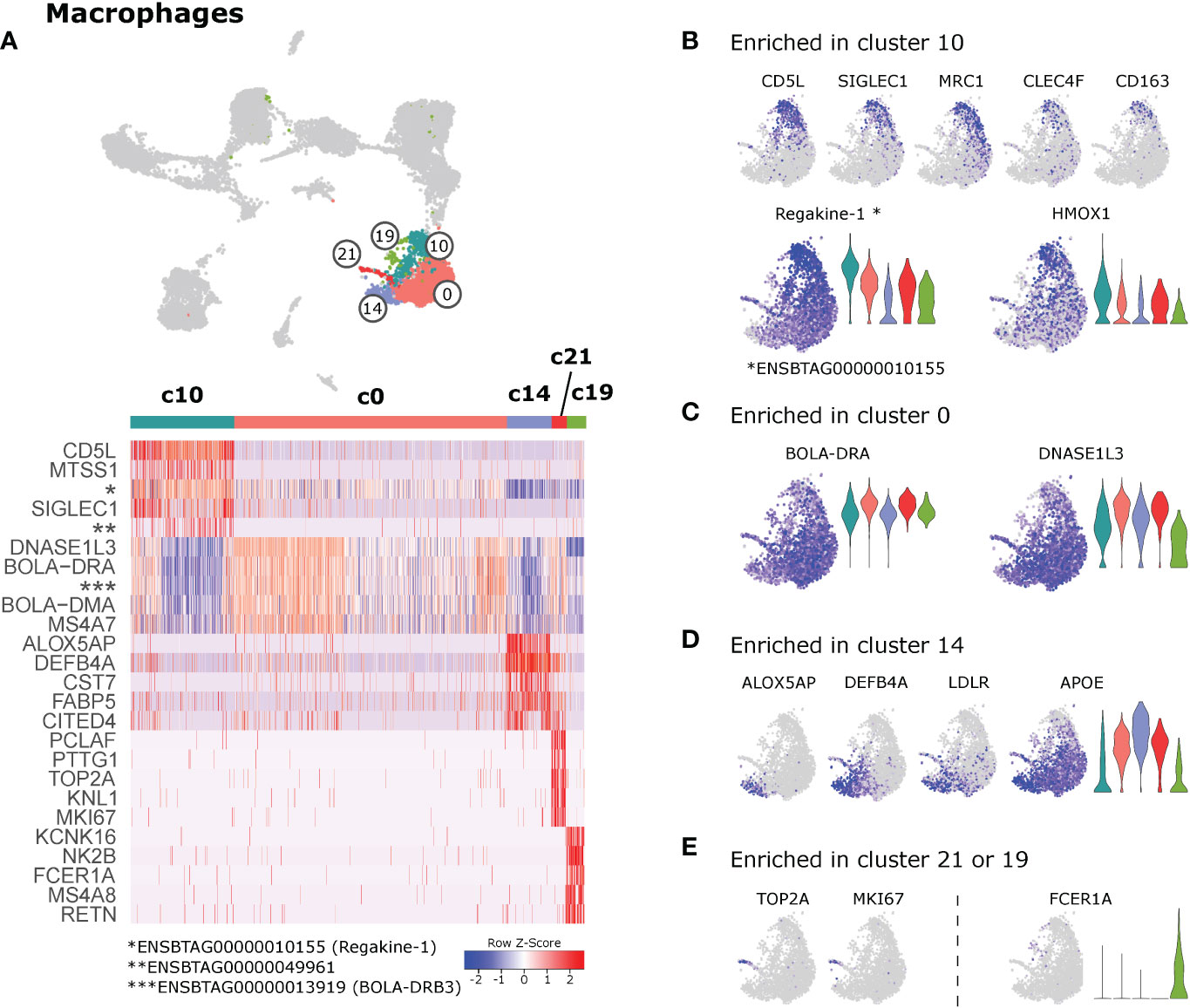
Figure 7 Macrophages. (A) Heatmap of the top 10 (p_adj) differentially expressed genes between macrophage clusters. (B–E) Visualization of selected genes enriched in cluster 10 (B), cluster 0 (C), cluster 14 (D) , and in clusters 21 or 19 (E).
Similar to monocytes in cluster 2, macrophages in cluster 0 expressed higher levels of genes related to antigen presentation (BOLA-DRA, BOLA-DMA, BOLA-DQA1, BLA-DQB, CD74) (Figure 7C and Supplementary File 11). High expression levels of DNASE1L3, MERTK and AXL in cluster 0, point towards a prominent role of these macrophages in efferocytosis.
Together with pro-inflammatory monocytes, cells in cluster 14 uniquely expressed ALOX5AP and DEFB4A (Figure 7D). While ALOX5AP is crucial for leukotriene biosynthesis, DEFB4A is described to have both antimicrobial and chemotactic functions (60). Other genes predominantly detected in cluster 14 encoded cystatin F (CST7), reported to regulate proteolytic activity during monocyte-to-macrophage differentiation (61), the receptor for low-density lipoprotein (LDLR), and the fatty-acid binding protein 5 (FABP5). Expression of FABP5 is in line with the idea that c14 represents pro-inflammatory macrophages, as this fatty-acid binding protein was shown to limit the anti-inflammatory response of murine macrophages (62).
Cells in cluster 21 stood out by high expression of proliferation-associated genes such as TOP2A and MKI67 (Figure 7E). When c21 was compared against the proliferating DC clusters 11 & 12, the clear macrophage identity (e.g. CD68, CSF1R) of c21 became apparent (data not shown). The presence of proliferating macrophages in the current dataset may suggest that some macrophages in lymph node of cattle are replaced by self-renewal, as reported for bona fide tissue-resident macrophages (26).
Cells in cluster 19 shared a number of genes with cDC2. Cells of this cluster were also located together with cDC2 and monocytes in the UMAP projection. Due to the small size of cluster 19 and the widespread positioning of its cells, we concluded that c19 might represent an artefact and should be interpreted with caution.
3.8 Genes of interest
While differential expression testing is restricted to and limited by cluster definitions, visualization of genes in feature plots provides unbiased and cluster-independent information, which is highly valuable to reveal sub-clustering and expression patterns that are hidden in the cluster analysis. So in addition to the differential expression testing described above, we examined genes associated with important immunological functions, such as pattern recognition, adhesion, migration, and antigen presentation, as well as genes coding for cytokines and their receptors and other immuno-relevant molecule classes, such as solute carriers, tetraspanins, semaphorins, metalloproteinases and purinergic receptors.
Very interesting patterns of gene expression became apparent which cannot all be described in detail here. Selected feature plots are shown in Figure 8. The complete collection of feature plots is given in Supplementary File 12. When interpreting these feature plots, the reader should keep in mind that dropout in single-cell data might contribute to low detection and apparent lack of expression.

Figure 8 Genes of interest. The complete collection of feature plots is given in Supplementary File 12 and includes the following categories: 1) Pattern recognition receptors, 2) Fc receptors, 3) Purinergic receptors, 4) Chemokines, 5) Chemokine receptors, 6) Integrins, 7) Galectins, 8) Antigen presentation, 9) T-cell modulation, 10) Interleukins, 11) Interleukin receptors, 12) TNF superfamily, 13) TNF receptor superfamily, 14) Tetraspanins, 15) Metalloproteinases, 16) Metabolism (misc.), 17) Glycolysis, 18) Solute carriers, 19) Complement system, 20) Interferon-associated, 21) Retinoic-acid production and signaling, 22) Semaphorins and receptors.
Transcripts for pattern-recognition-receptor genes were predominantly detected in monocytic cells. Apart from for example TLR3 transcripts, which were mainly detected in dendritic cells, and CLEC9A transcripts, which were exclusively detected in resident cDC1 (c1 and part of c11/c12), who also expressed high levels of CLEC12A alongside cluster 3 (inflammatory cDC2) and monocytes. Interestingly, NLRP3 expression appeared to be limited to monocytic cells and cDC2 (including c3), while NLRP1 was also highly expressed in cDC1 (c1).
Looking at Fc receptors, we found FCER2 to be expressed predominantly in monocytes and in cluster 3 (inflammatory cDC2), as well as in putative migratory cDC2 (c8). Notably, while FCER1G expression was detected in CD5- cDC2 (c5), but not in CD5+ cDC2 (c9), expression of FCRL3 and FCRL4 followed the opposite pattern, with clear transcript enrichment in CD5+ cDC2 (c9).
Among transcripts encoding purinergic receptors, involved in immune regulation by sensing extracellular nucleotides (63), P2RY6 transcripts were clearly enriched in resident cDC1 (c1). In line with our previous results obtained with blood-derived cells of cattle (11), P2RY1 and P2RY10 transcripts were primarily detected in monocytic and dendritic cells, respectively.
Also chemokines and chemokine receptors showed clear cluster-specific expression patterns. Most strikingly, CCR7-expressing DC could be divided according to CCL17/CCL22 expression (c8, putative migratory cDC2) and CXCL9/CXCL10 expression (c6, putative migratory cDC1). Apart from c6, CXCL9 and CXCL10 were also highly expressed in what appears to be a subcluster of c1 (resident cDC1), presumably containing activated resident cDC1. Notably, CX3CR1 was mainly expressed by DC progenitors, cDC2 (including c3) and monocytes. Apart from CX3CR1 and CCR5, we could hardly detect any transcripts coding for chemokine receptors in progenitor cells (c11, c12, c13), cDC2 (c5, c9) and cluster 3. Remarkable is also the high expression of CCR1 and CXCR4 in pro-inflammatory monocytes (c4). However, while CXCR4 expression decreased towards anti-inflammatory monocytes and macrophages, CCR1 expression was again detected at high levels throughout macrophage clusters. Moreover, expression of the follicle-homing receptor CXCR5 was exclusively detected in putative migratory cDC2, suggesting that these cells locate close to follicles to interact with B- and T-cells, as described for the murine system (64).
Among the investigated integrins, ITGAL (CD11a) stood out by exclusive high expression in resident cDC1 (c1) and monocytes (c4, c2, c7). And transcripts for ITGAV (CD51) were almost exclusively detected in pro-inflammatory macrophages (c14).
In addition, galectins showed specific expression patterns. Most notably Galectin-1 (LGALS1), which was highly expressed in both resident and migratory cDC1 (c1, c6) and in inflammatory and migratory cDC2 (c3, c8). Galectin-1 expression in DC is reported to induce IL-27 expressing regulatory DC (65). Transcripts for LGALS3 and LGALS9 were enriched in macrophages and monocytes, respectively.
Moreover, genes related to antigen presentation and T-cell modulation showed interesting cluster-specific patterns. Notably, cluster 6 (putative cDC1 within migratory DC), clearly showed the highest levels of BOLA, BOLA-NC1 and IDO1, suggesting that these cells interact with and regulate CD8 T cells. Resident cDC1 (c1) were clearly enriched for WDFY4 transcripts, presumably indicating their potential for cross presentation. Notably, while CD1A, CD1B and CD1E were exclusively expressed by putative moDC (bridging between c2 and c3), CD1D was also highly expressed by resident cDC1 (c1) and to a lesser extent by cDC2 (c3, c5, c9).
Looking at interleukins, we found that IL1B was expressed mainly by monocytic cells and cDC2 (including c3), while the gene for the IL1 receptor antagonist (IL1RN) was mainly detected in pro-inflammatory monocytes (c4). Transcripts for IL-18 (IL18) showed a prominent and specific expression in macrophages. Notably, subclusters of migratory DC clearly differed in their expression of IL15 (c8, putative migratory cDC2) and EBI3 (IL-27B) (c6, putative migratory cDC1). Among interleukin receptor genes, IL21R and IL13RA1 stood out by their exclusive expression in migratory DC and monocytes, respectively.
Genes encoding members of the TNF- and TNF-receptor superfamily were also differentially expressed, with TNFSF13 (APRIL) and TNFSF10 (TRAIL) being detected primarily in pro- and anti-inflammatory monocytes (c4 vs. c7), respectively. Notably, transcripts for TNF-α (TNF) were only poorly detected, but expression of the TNF receptor genes TNFRSF1A (TNFR1) and TNFRSF1B (TNFR2) was detected at high levels in monocytic cells (TNFRSF1A/B) and migratory DC (TNFRSF1B). Moreover, while TNFSF13B (BAFF) was highly and almost exclusively expressed in all migratory DC, transcripts of the TNF receptor genes TNFRSF9 and TNFRSF13C were enriched in putative migratory cDC1 (c6) and CD5 expressing resident cDC2 (c9)/putative migratory cDC2 (c8), respectively.
Also tetraspanins showed pronounced cell-type specific expression, for example the macrophage-restricted expression of TSPAN4, recently shown to interact with Histamin H4 receptor (66), or the high transcription of TSPAN3 in migratory DC, among which migratory cDC1 (c6) were also enriched in CD151 (TSPAN24) and CD81 (TSPAN28) transcripts. Other tetraspanin genes highly expressed in migratory DC included TSPAN13, TSPAN17, and TSPAN33, the two latter of which are known to interact with ADAM10, a metalloproteinase mediating ectodomain shedding (67). While ADAM10 transcripts were detected in all clusters with higher levels in monocytic cells, other metalloproteinases showed more specific patterns of expression. Transcripts for ADAM8 and ADAM9 were enriched in migratory DC (c6, c8), and transcripts for MMP9 and MMP14 were almost exclusively detected in resident cDC2 (c5, c9) and macrophages (c0, c10, c14, c21), respectively.
Among genes involved in metabolism and previously reported to be overexpressed in cM compared to ncM in bovine blood (23), KHK and SORD (fructose metabolism) stood out by selectively higher expression in monocytic clusters and in resident cDC2 (c5, c9), respectively. Transcripts involved in glycolysis showed surprisingly heterogeneous expression patterns, but were mostly detected in monocytes and cDC2 (including c3), consistent with the highest transcript levels of HIF1A in these clusters. In line with the pro-inflammatory signature of monocytes in c4, these cells transcribed the highest levels of SLC2A3, a high-affinity glucose transporter.
Visualization of SLC (solute carrier) gene expression revealed interesting patterns that are also reported for humans, like high SLCO2B1 expression in macrophages (68) or high SLCO5A1 expression in mature dendritic cells (69). Notably, transcripts for the iron transporter SLC40A1 were only detected in certain cluster-independent regions within macrophages, and the urea transporter SLC14A1 was exclusively detected in putative early DC progenitors (c13). Moreover we found that migratory DC exclusively expressed high levels of SLC7A1, which was recently described as a cellular receptor for bovine leukemia virus (70). Infection of migratory DC by bovine leukemia virus may have important implications for pathogenesis of the disease. Notably, bovine leukemia virus has recently also gained attention as a potential causative agent in human breast cancer (71).
Interesting expression patterns were also apparent for genes related to the complement system. Transcripts for C1Q, the sensory component initiating the formation of the C1 complex (C1Q+C1R+C1S), were exclusively expressed in macrophages and anti-inflammatory monocytes. The latter (c7) also expressed the highest levels of C2, while transcripts for C3 were exclusively detected in rather pro-inflammatory monocytes (c4, c2). Conversely, C3AR1 transcription appeared to be selectively absent from pro-inflammatory monocytes (c4). Notably, detection of CD55 transcripts was limited to monocytes. Complement factor D (CFD) and CFP, involved in the alternative pathway of complement activation, were also detected in dendritic cells, CFD most prominently in non-proliferating progenitors (c13), putative migratory cDC2 (c8), as well as in CD5- resident cDC2 (c5) and cluster 3 (inflammatory cDC2).
Among interferon-associated genes, ISG15 and IFI6 stood out by their almost exclusive and high expression in putative migratory cDC1 (c6) and macrophages, respectively.
Retinoic acid (RA) is reported to imprint gut homing in T cells (72, 73) and – as recently reviewed – to control IgA switch in B cells (74). As reported in these studies, mucosal subsets of myeloid cells appear to be specialized to produce RA, reflected by their unique expression of required enzymes. Notably, in our dataset we found high expression of ALDH1A1 exclusively in macrophages, and some ALDH1A2 expression in cells assigned to resident cDC1 and migratory DC clusters. We also detected CD103 (ITGAE) expression in DC, which is regarded as a marker for “RA-DC” in mice, alongside expression of retinoic acid receptors (RARA, RARB, RARG, RXRA, RXRB). Notably, the RA-responsive genes RARRES1 and RARRES2 (Chemerin) were predominantly expressed in macrophages, the latter also in migratory cDC1.
Finally, genes for semaphorins (SEMA4A, SEMA4D, SEMA4F, SEMA7A) and their receptors (plexins such as PLXNB2 and PLXNC1 and neuropilins such as NRP1) showed unique expression patterns across dendritic and monocytic clusters. Notably, semaphorin signaling is regarded as highly conserved across species (75) and its regulatory roles in innate immunity are only beginning to be elucidated (76).
3.9 Trajectories and sources of MPS heterogeneity in bovine mesenteric lymph node
Our scRNA-seq analysis of mononuclear phagocytes in bovine mesenteric lymph nodes revealed clusters of resident DC (progenitors, cDC1, cDC2, pDC), as well as migratory DC (putative cDC1 and cDC2), and a cluster of inflammatory cDC2 containing moDC and potentially DC3 (Figure 9A). Monocytic cells could be clearly separated into monocytes and macrophages, both clustering according to pro-and anti-inflammatory gene expression, and included a cluster of proliferating macrophages, presumably giving rise to bona fide lymph-node resident macrophages. As illustrated in Figure 9B, heterogeneity of mononuclear phagocytes in bovine lymph nodes may originate from blood-borne progenitors (S1) that differentiate into resident cDC1 (T1a and T1b), resident cDC2 (T2) and resident DC3 (T3). Highly activated migratory DC (S2) may enter via afferent lymph. Furthermore, cDC2 may differentiate into inflammatory cDC2 (T4). Monocytic cells may originate from cM that enter the lymph node (S3) and differentiate into antigen-presenting moDC (T5) or via antigen-presenting monoctyes (resembling intM in blood) towards increasingly anti-inflammatory monocytes (resembling ncM in blood) and further into macrophages (T6). Macrophages may aquire anti-inflammatory or pro-inflammatory gene expression depening on the niche they occupy. Alternatively, they may aquire an increasingly pro-inflammatory gene expression over time (T7). Lastly, self-renewing tissue macrophages (S4) may contribute to the pool of macrophages (T8), displaying a transcriptome indistinguishable from terminally differentiated monocyte-derived macrophages.
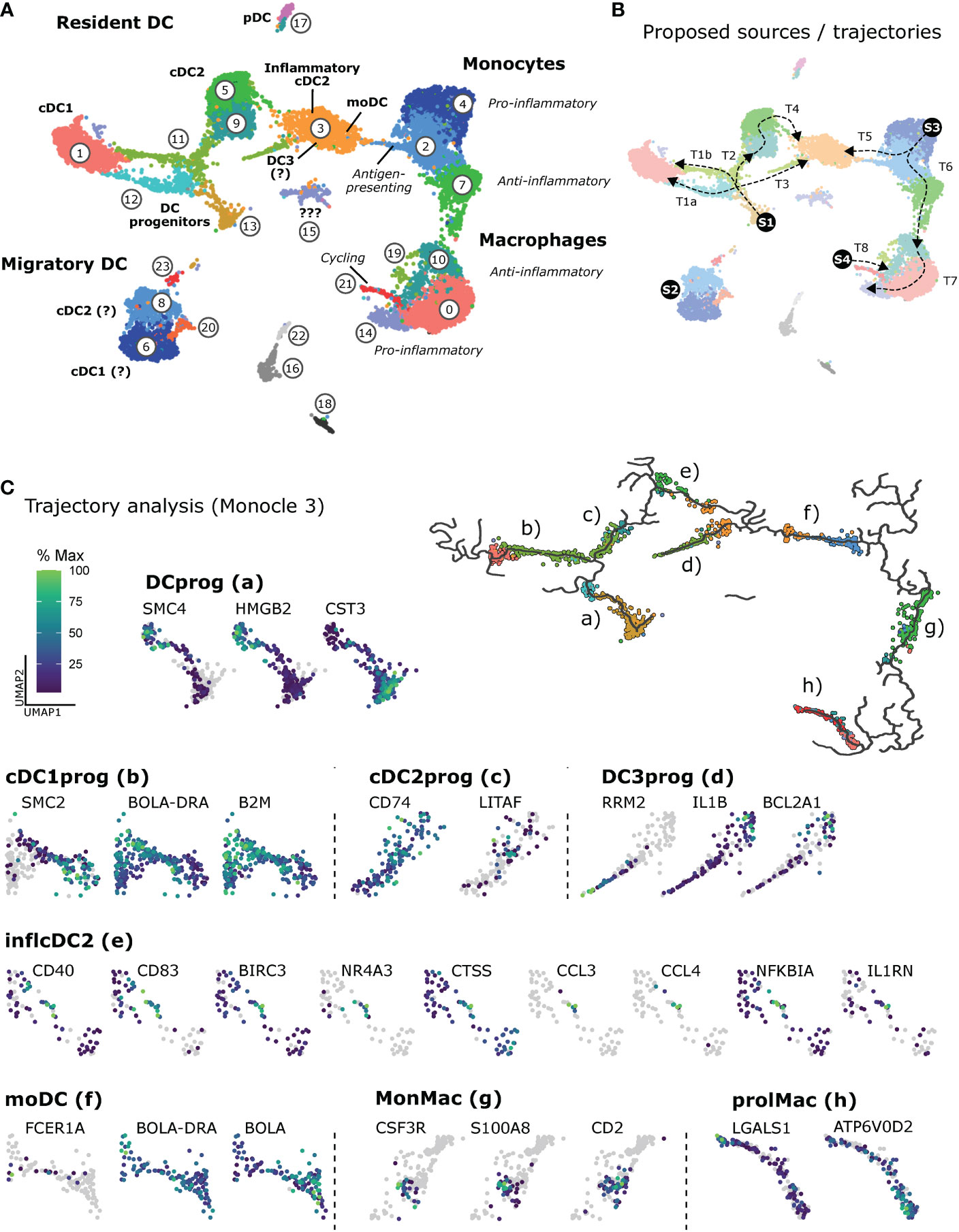
Figure 9 Heterogeneity of mononuclear phagocytes in bovine mesenteric lymph node. (A) Overview of cluster assignment. (B) Proposed source populations (S1-S4) and proposed differentiation trajectories (T1-T8). (C) Trajectories calculated by Monocle 3 and visualization of exemplary genes differentially expressed along selected trajectories (a-h). The complete list of differentially expressed genes (q < 0.05) is given in Supplementary File 13.
The R package Monocle 3 was used to analyze expression patterns along selected trajectories of interest (Figure 9C and Supplementary File 13). Calculated graphs do not allow for conclusions on type or direction of differentiation, biological processes, or cell ontogeny.
The number of differentially expressed genes (q < 0.05) differed markedly between trajectories (Supplementary File 13), spanning from 101 genes (f, moDC) to 2370 genes (a, DCprog). Selected genes are visualized in Figure 9C, showing for example dominant expression of CST3 in putative early progenitors of dendritic cells (Figure 9C, a), and a gradual increase of BOLA-DRA and B2M expression in progenitors developing towards cDC1 (b). Progenitors developing towards cDC2 (c) gradually upregulated CD74 and LITAF, the latter encoding an LPS-induced transcription factor (77) reported to (co-) regulate inflammatory cytokine production (78). Notably, IL1B and anti-apoptotic BCL2A1 were clearly upregulated towards putative DC3 (d), and the trajectory leading to inflammatory cDC2 (e) contained a subcluster of cells expressing high levels of genes commonly associated with an activated state, including expression of CCL3, CCL4 and regulatory NFKBI and IL1RN. Monocytes differentiating towards moDC (f) appeared to at least transiently express FCER1A and to upregulate BOLA-DRA while downregulating BOLA. On the trajectory bridging betweeen monocytes and macrophages (g), clusters of cells became apparent expressing high levels of CSF3R, S100A8 and/or CD2. In fact, for trajectories originating at progenitor cells, the majority of differentially expressed genes were cell-cycle genes beeing downregulated. The same was observed for proliferating macrophages (h). Moreover, LGALS1 was highly expressed at the beginning of this trajectory (prolMac, h). Notably, ATP6V0D2, encoding a lysosomal ATPase suggested to have crucial roles for endosomal TLR signaling (79, 80) and antiviral responses (81), was increasingly upregulated along this trajectory leading to putative bona fide macrophages.
4 Discussion
In the present study, we have applied 10x Genomics and Illumina sequencing to decipher the single-cell transcriptome of mononuclear phagocytes in bovine mesenteric lymph nodes.
One particularly interesting aspect of the present dataset is the collection of single-cell transcriptomes from migratory DC subsets, enabling an unbiased view on in-vivo activated DC and their profound transcriptional reprogramming. We identified two subclusters of migratory DC that clearly differ in their expression of chemokine genes and likely contain cDC1 (c6; CXCL9, CXCL10) and cDC2 (c8; CCL17, CCL22). High expression of interferon-inducible genes CXCL9 and CXCL10 might indicate that migratory cDC1 occupy a niche where they interact with CXCR3-expressing Th1 and CD8 T cells. Indeed, high expression of BOLA (MHC-I) and B2M would support a specialization towards CD8-T-cell interaction. Notably, CXCR3 transcripts were found in resident cDC1 alongside CXCL9 and CXCL10 transcripts, suggesting a common niche of resident and migratory cDC1 for promoting antiviral responses. A recent study performed in mice suggested that CXCL9 and CXCL10 are produced by spatially distinct DC subsets, creating distinct microenvironments and favoring distinct effector and memory T-cell fates (82). In our dataset, most migratory DC contained transcripts for both CXCL9 and CXCL10, making compartmentalization purely based on differential expression of these two chemokines rather unlikely, at least in the bovine system.
Dominant expression of CCL17 and CCL22 detected in putative migratory cDC2 would suggest that these cells are specialized in attracting CCR4-expressing Th2/Th17 cells as well as Tregs (49), presumably creating a niche supporting survival and proliferation of these cells by IL-15 expression. Moreover, some of these putative migratory cDC2 were enriched in CXCR5 transcripts, suggesting that they locate close to B-cell follicles to support follicular T-helper cell (Thf) differentiation (2). In this regard, production of soluble CD25 by DC has been proposed as a means to support Thf differentiation (83). Transcripts for CD25 (IL2RA) were poorly detected in the current dataset, but were primarily found in clusters associated with cDC2, including cluster 8. We have previously reported pronounced upregulation of surface CD25 expression after 4-hour in-vitro stimulation of bovine DC subsets (36), making expression of membrane-bound and/or soluble CD25 by activated DC a likely mechanism to control Thf differentiation also in bovine lymph nodes.
Apart from activation in the periphery, some CCR7+ DC may also have differentiated from resident DC that were activated in the lymph node and have upregulated CCR7 expression in order to migrate towards T-cell zones, as recently described for mice (4). In fact, a few cells adjacent to resident cDC1 also expressed CCR7, and displayed a more activated transcriptome. Moreover, both tolerogenic and immunogenic DC may be present in the migratory DC clusters, as molecular changes have been shown to be highly similar for both activation states (84). Along this line, recently described mregDC (85) may be included as well, given the prominent expression of regulatory genes such as CD274 (PDL-1), PDCD1LG2 (PDL-2), FAS, and SOCS2 in CCR7-expressing DC.
The recent discovery of DC3 as a separate DC lineage (33, 34) prompted us to watch out for these cells in the current dataset. Looking at the branching pattern of DC progenitors, it is conceivable that some cells in cluster 3 constitute DC3, however we could not find a clear separation from putative inflammatory cDC2 and monocyte-derived DC. Certainly, the seemingly convergent differentiation pathways of several cell types might indicate crucial roles of cells with this transcriptomic makeup – it remains to be determined if these inflammatory cells act in different niches of the lymph node with nuanced differences in their specialization, or if there is pronounced redundancy in the system because the tasks they fulfill are crucial for proper functioning of the immune system and thus survival.
The high proportion of monocytic cells in bovine mesenteric lymph nodes, and reportedly also in murine and human lymph nodes (6, 86–88), raises several questions regarding their functional roles, their origins, and their fates. The idea that dendritic and monocytic cells cooperate in the lymph node to optimize adaptive immune responses, is supported by recent findings in mice, where inflammatory monocytes were shown to enter the lymph node from blood via high endothelial venules (HEV) and accumulate in the T-cell zone, where they provide polarizing cytokines to optimize effector T-cell differentiation (4). In line with this, we found monocytic cells in bovine mesLN to express high levels of SELL (CD62L) alongside CXCL16, attracting activated T cells (89), and transcripts for T-cell modulating cytokines, such as IL-16, IL-12B/IL-23A and IL-27/IL-27B (EBI3). Classical monocytes were also described to travel to lymph nodes via tissue-draining lymph, both under inflammatory and steady-state conditions (90). Accordingly, murine moDC were reported to be capable of CCR7 upregulation in vivo (91). It is unclear if bovine monocytes can upregulate CCR7 in vivo. At least in-vitro activation with TLR-ligands did not lead to an increase of CCR7 transcripts and protein expression in bovine cM (36). Moreover, in the present study we did not detect any CCR7 transcripts in lymph-node derived monocytic cells.
Our recent scRNA-seq analyses of bovine blood monocyte subsets support the idea of continuous differentiation from cM via intM to ncM (23). It remains to be determined if classical pro-inflammatory monocytes that enter the lymph node can differentiate to cells that resemble intM and ncM found in peripheral blood of cattle, or if intM and ncM differentiated in blood can enter lymph nodes themselves. The almost complete absence of CD62L expression (mRNA and protein) from bovine intM and ncM in blood (23), would argue against them having access via HEV, or at least indicates differential regulation of LN entry.
The biology of tissue-resident macrophages is described to be shaped by four different factors: i) origin (embryonic vs. monocyte-derived), ii) tissue-specific environment (e.g. lung vs. liver), iii) inflammatory environment, and iv) time spent in the tissue (92). Consistent with described functions of macrophages, our data suggests a division of labor between rather anti-inflammatory macrophages engaging in efferocytosis, and macrophages with antibacterial activity and a rather pro-inflammatory profile. Furthermore, the detection of cycling macrophages in the present dataset clearly supports the hypothesis that self-renewing macrophages of embryonic origin are present in bovine lymph nodes. The fact that these bona fide macrophages don’t segregate as a separate cluster in our dataset, is in line with the idea that monocyte-derived macrophages, educated by niche-specific signals, acquire a very similar and thus undistinguishable transcriptome over time. Expression of TIMD4 has been associated with long-term residence under steady-state conditions – thus being upregulated on monocyte-derived macrophages over time (26). In the current dataset, the continuous increase of TIMD4 expression across macrophage clusters may therefore indicate a differentiation path for monocyte-derived macrophages. The expression of APOE followed the same pattern, suggesting that also APOE might serve as a time-dependent marker for macrophage differentiation.
It has been suggested that differentiation of monocytes to tissue macrophages can be split into two phases (92): a rapid differentiation that would instruct cells to stay in the tissue niche (“stay-here” signals), and a second phase, where monocyte-derived macrophages would adapt to their environment integrating information on tissue type and inflammatory status (“learn-this” signals). It is intriguing to speculate that this first rapid phase of differentiation is visible in the UMAP projection of our dataset, where monocytes and macrophages are connected with a narrow bridge of cells. The low frequency of transitional cells would be in line with rapid differentiation. Similarly, differentiation towards moDC appears to be a rapid process, leading to a narrow cellular bridge between monocytes and cluster 3 in our dataset.
Further macrophage heterogeneity is introduced by niches within the same tissue and stromal and immune cells present therein. For the lymph node, several niche-specific subsets of macrophages have been described (93), such as subcapsular and medullary sinus macrophages, capturing lymph-borne antigens (94, 95), macrophages in the lymph node parenchyma such as in germinal centers and medullary cords, and recently described efferocytotic T-cell zone macrophages (96). Our data reveals cluster-specific heterogeneity in the expression of chemokine receptor genes such as CCR1, CCR5, CXCR4 and CX3CR1, presumably guiding monocytic cells to their “niche of residence”, where further differentiation may also depend on available space in that niche (97).
One limitation of the current study is that pDC were only detected in a very small cluster (c17). In fact, flow cytometric analyses suggest that the frequency of pDC in bovine mesLN is considerably higher than suggested by our scRNA-seq dataset (unpublished data). With the gating strategy employed to sort mononuclear phagocytes in the present study we aimed to reduce contamination with lymphocytes, but at the same time we excluded most pDC that express comparatively low levels of Flt3. A future study will have to address pDC in the lymph node and investigate for example if pDC subsets are present in a special cDC2-like activation state, as described for human and murine transitional DC (19, 29, 98–101). An indication for these transitional DC in the current dataset may be that a small subset of cells spatially clustering with pDC (c17) got assigned to a cDC2 cluster (c9). In general, the authors cannot rule out that enrichment of cells by FACS may have specifically excluded certain populations of DC or monocytes/macrophages. Moreover, having a large enough sample size and/or integration of multiple samples is crucial for making conclusions on complex cell composition and trajectories involving rare transitional cell states, as evident from comparing individual samples to our integrated dataset.
With the present study we provide the first in-depth single-cell analysis of the mononuclear phagocyte compartment in bovine lymph nodes. Trajectories of differentiation became apparent that may well reflect general principles of MPS dynamics in lymph nodes across species. Some of them previously reported, such as the differentiation of resident DC from blood-borne progenitors in the lymph node (102, 103), others less well understood, such as the origin and differentiation pathways of monocytic cells and bona fide macrophages in secondary lymphoid tissue, or the seemingly convergent functions of inflammatory DC subsets (including DC3) and monocyte-derived DC.
The hypotheses generated in this manuscript are an important contribution towards a better understanding of the mononuclear phagocyte compartment in lymph nodes, especially when acknowledging that basic DC and monocyte biology appears to be largely conserved across mammalian species. Spatial analyses performed in future studies should help to define niche-specific transcriptomes of macrophages and to characterize microenvironments where dendritic cells, monocytes, and T-cells interact to shape adaptive immune responses – fundamental insights into MPS biology that will benefit human and veterinary medicine alike.
Data availability statement
The data presented in the study are deposited in the ENA repository, accession number PRJEB57581 (https://www.ebi.ac.uk/ena/browser/view/PRJEB57581).
Ethics statement
Ethical review and approval was not required for the animal study because lymph nodes were collected at a slaughter house from the by-products of slaughter.
Author contributions
GB performed laboratory work, analyzed data, and wrote a first draft of the manuscript. ST performed data analysis, prepared the figures, and wrote the final manuscript. MK and RB performed bioinformatic analyses. ST and AS designed and supervised the overall project. All authors contributed to the article and approved the submitted version.
Acknowledgments
We thank Corinne Hug (IVI) for producing recombinant bovine Flt3L, Stefan Müller (FCCS, University of Bern) for sorting, and Pamela Nicholson, Catia Coito and Tosso Leeb from the NGS Platform of the University of Bern for single-cell RNA sequencing. A previous version of the manuscript is available on the preprint server bioRxiv (104).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.1099357/full#supplementary-material
References
1. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature (1998) 392:245–52. doi: 10.1038/32588
2. Eisenbarth SC. Dendritic cell subsets in T cell programming: location dictates function. Nat Rev Immunol (2019) 19:89–103. doi: 10.1038/s41577-018-0088-1
3. Guilliams M, Mildner A, Yona S. Developmental and functional heterogeneity of monocytes. Immunity (2018) 49:595–613. doi: 10.1016/j.immuni.2018.10.005
4. Leal JM, Huang JY, Kohli K, Stoltzfus C, Lyons-Cohen MR, Olin BE, et al. Innate cell microenvironments in lymph nodes shape the generation of T cell responses during type I inflammation. Sci Immunol (2021) 6(56):eabb9435. doi: 10.1126/sciimmunol.abb9435
5. Shi C, Pamer EG. Monocyte recruitment during infection and inflammation. Nat Rev Immunol (2011) 11:762–74. doi: 10.1038/nri3070
6. Jakubzick CV, Randolph GJ, Henson PM. Monocyte differentiation and antigen-presenting functions. Nat Rev Immunol (2017) 17:349–62. doi: 10.1038/nri.2017.28
7. Karsunky H, Merad M, Cozzio A, Weissman IL, Manz MG. Flt3 ligand regulates dendritic cell development from Flt3+ lymphoid and myeloid-committed progenitors to Flt3+ dendritic cells in vivo. J Exp Med (2003) 198:305–13. doi: 10.1084/jem.20030323
8. Guzylack-Piriou L, Alves MP, McCullough KC, Summerfield A. Porcine Flt3 ligand and its receptor: generation of dendritic cells and identification of a new marker for porcine dendritic cells. Dev Comp Immunol (2010) 34:455–64. doi: 10.1016/j.dci.2009.12.006
9. Auray G, Keller I, Python S, Gerber M, Bruggmann R, Ruggli N, et al. Characterization and transcriptomic analysis of porcine blood conventional and plasmacytoid dendritic cells reveals striking species-specific differences. J Immunol (2016) 197:4791–806. doi: 10.4049/jimmunol.1600672
10. Ziegler A, Marti E, Summerfield A, Baumann A. Identification and characterization of equine blood plasmacytoid dendritic cells. Dev Comp Immunol (2016) 65:352–7. doi: 10.1016/j.dci.2016.08.005
11. Talker SC, Baumann A, Barut GT, Keller I, Bruggmann R, Summerfield A. Precise delineation and transcriptional characterization of bovine blood dendritic-cell and monocyte subsets. Front Immunol (2018) 9:2505. doi: 10.3389/fimmu.2018.02505
12. Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai N, Schraml BU, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol (2014) 14:571–8. doi: 10.1038/nri3712
13. Guilliams M, Dutertre CA, Scott CL, McGovern N, Sichien D, Chakarov S, et al. Unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity (2016) 45:669–84. doi: 10.1016/j.immuni.2016.08.015
14. Collin M, Bigley V. Human dendritic cell subsets: an update. Immunology (2018) 154:3–20. doi: 10.1111/imm.12888
15. Ziegler-Heitbrock L, Ohteki T, Ginhoux F, Shortman K, Spits H. Reclassifying plasmacytoid dendritic cells as innate lymphocytes. Nat Rev Immunol (2022). doi: 10.1038/s41577-022-00806-0
16. Durai V, Murphy KM. Functions of murine dendritic cells. Immunity (2016) 45:719–36. doi: 10.1016/j.immuni.2016.10.010
17. Yin X, Chen S, Eisenbarth SC. Dendritic cell regulation of T helper cells. Annu Rev Immunol (2021) 39:759–90. doi: 10.1146/annurev-immunol-101819-025146
18. Theisen D, Murphy K. The role of cDC1s in vivo: CD8 T cell priming through cross-presentation. F1000Res (2017) 6:98. doi: 10.12688/f1000research.9997.1
19. Abbas A, Vu Manh TP, Valente M, Collinet N, Attaf N, Dong C, et al. The activation trajectory of plasmacytoid dendritic cells in vivo during a viral infection. Nat Immunol (2020) 21:983–97. doi: 10.1038/s41590-020-0731-4
20. Yona S, Kim KW, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity (2013) 38:79–91. doi: 10.1016/j.immuni.2012.12.001
21. Ziegler-Heitbrock L. Monocyte subsets in man and other species. Cell Immunol (2014) 289:135–9. doi: 10.1016/j.cellimm.2014.03.019
22. Hussen J, Schuberth HJ. Heterogeneity of bovine peripheral blood monocytes. Front Immunol (2017) 8:1875. doi: 10.3389/fimmu.2017.01875
23. Talker SC, Barut GT, Lischer HEL, Rufener R, von Munchow L, Bruggmann R, et al. Monocyte biology conserved across species: Functional insights from cattle. Front Immunol (2022) 13:889175. doi: 10.3389/fimmu.2022.889175
24. Kapellos TS, Bonaguro L, Gemund I, Reusch N, Saglam A, Hinkley ER, et al. Human monocyte subsets and phenotypes in major chronic inflammatory diseases. Front Immunol (2019) 10:2035. doi: 10.3389/fimmu.2019.02035
25. Hume DA, Irvine KM, Pridans C. The mononuclear phagocyte system: The relationship between monocytes and macrophages. Trends Immunol (2019) 40:98–112. doi: 10.1016/j.it.2018.11.007
26. Jenkins SJ, Allen JE. The expanding world of tissue-resident macrophages. Eur J Immunol (2021) 51:1882–96. doi: 10.1002/eji.202048881
27. Papalexi E, Satija R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat Rev Immunol (2018) 18:35–45. doi: 10.1038/nri.2017.76
28. Ginhoux F, Yalin A, Dutertre CA, Amit I. Single-cell immunology: Past, present, and future. Immunity (2022) 55:393–404. doi: 10.1016/j.immuni.2022.02.006
29. Villani AC, Satija R, Reynolds G, Sarkizova S, Shekhar K, Fletcher J, et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science (2017) 356(6335):eaah4573. doi: 10.1126/science.aah4573
30. Dutertre CA, Becht E, Irac SE, Khalilnezhad A, Narang V, Khalilnezhad S, et al. Single-cell analysis of human mononuclear phagocytes reveals subset-defining markers and identifies circulating inflammatory dendritic cells. Immunity (2019) 51:573–589 e8. doi: 10.1016/j.immuni.2019.08.008
31. Bosteels C, Neyt K, Vanheerswynghels M, van Helden MJ, Sichien D, Debeuf N, et al. Inflammatory type 2 cDCs acquire features of cDC1s and macrophages to orchestrate immunity to respiratory virus infection. Immunity (2020) 52:1039–1056 e9. doi: 10.1016/j.immuni.2020.04.005
32. Tussiwand R, Rodrigues PF. Where's Waldo: Identifying DCs within mononuclear phagocytes during inflammation. Immunity (2020) 52:892–4. doi: 10.1016/j.immuni.2020.05.006
33. Cytlak U, Resteu A, Pagan S, Green K, Milne P, Maisuria S, et al. Differential IRF8 transcription factor requirement defines two pathways of dendritic cell development in humans. Immunity (2020) 53:353–370 e8. doi: 10.1016/j.immuni.2020.07.003
34. Bourdely P, Anselmi G, Vaivode K, Ramos RN, Missolo-Koussou Y, Hidalgo S, et al. Transcriptional and functional analysis of CD1c(+) human dendritic cells identifies a CD163(+) subset priming CD8(+)CD103(+) T cells. Immunity (2020) 53:335–352 e8. doi: 10.1016/j.immuni.2020.06.002
35. Ginhoux F, Guilliams M, Merad M. Expanding dendritic cell nomenclature in the single-cell era. Nat Rev Immunol (2022) 22:67–8. doi: 10.1038/s41577-022-00675-7
36. Barut GT, Lischer HEL, Bruggmann R, Summerfield A, Talker SC. Transcriptomic profiling of bovine blood dendritic cells and monocytes following TLR stimulation. Eur J Immunol (2020) 50(11):1691–711. doi: 10.1002/eji.202048643
37. Team RC. R: A language and environment for statistical computing (2016). Available at: https://www.R-project.org/.
38. McCarthy DJ, Campbell KR, Lun AT, Wills QF. Scater: pre-processing, quality control, normalization and visualization of single-cell RNA-seq data in r. Bioinformatics (2017) 33:1179–86. doi: 10.1093/bioinformatics/btw777
39. Lun AT, Bach K, Marioni JC. Pooling across cells to normalize single-cell RNA sequencing data with many zero counts. Genome Biol (2016) 17:75. doi: 10.1186/s13059-016-0947-7
40. Lun AT, McCarthy DJ, Marioni JC. A step-by-step workflow for low-level analysis of single-cell RNA-seq data with bioconductor. F1000Res (2016) 5:2122. doi: 10.12688/f1000research.9501.2
41. Stuart T, Butler A, Hoffman P, Hafemeister C, Papalexi E, Mauck WM, et al. Comprehensive integration of single-cell data. Cell (2019) 177:1888–1902 e21. doi: 10.1016/j.cell.2019.05.031
42. Zappia L, Oshlack A. Clustering trees: a visualization for evaluating clusterings at multiple resolutions. Gigascience (2018) 7(7):giy083. doi: 10.1093/gigascience/giy083
43. Trapnell C, Cacchiarelli D, Grimsby J, Pokharel P, Li S, Morse M, et al. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat Biotechnol (2014) 32:381–6. doi: 10.1038/nbt.2859
44. Qiu X, Mao Q, Tang Y, Wang L, Chawla R, Pliner HA, et al. Reversed graph embedding resolves complex single-cell trajectories. Nat Methods (2017) 14:979–82. doi: 10.1038/nmeth.4402
45. Cao J, Spielmann M, Qiu X, Huang X, Ibrahim DM, Hill AJ, et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature (2019) 566:496–502. doi: 10.1038/s41586-019-0969-x
46. Yoneyama H, Narumi S, Zhang Y, Murai M, Baggiolini M, Lanzavecchia A, et al. Pivotal role of dendritic cell-derived CXCL10 in the retention of T helper cell 1 lymphocytes in secondary lymph nodes. J Exp Med (2002) 195:1257–66. doi: 10.1084/jem.20011983
47. Groom JR, Richmond J, Murooka TT, Sorensen EW, Sung JH, Bankert K, et al. CXCR3 chemokine receptor-ligand interactions in the lymph node optimize CD4+ T helper 1 cell differentiation. Immunity (2012) 37:1091–103. doi: 10.1016/j.immuni.2012.08.016
48. Wang J, Zhou J, Wang C, Fukunaga A, Li S, Yodoi J, et al. Thioredoxin-1: A promising target for the treatment of allergic diseases. Front Immunol (2022) 13:883116. doi: 10.3389/fimmu.2022.883116
49. Rapp M, Wintergerst MWM, Kunz WG, Vetter VK, Knott MML, Lisowski D, et al. CCL22 controls immunity by promoting regulatory T cell communication with dendritic cells in lymph nodes. J Exp Med (2019) 216:1170–81. doi: 10.1084/jem.20170277
50. Xu S, Zhao L, Larsson A, Venge P. The identification of a phospholipase b precursor in human neutrophils. FEBS J (2009) 276:175–86. doi: 10.1111/j.1742-4658.2008.06771.x
51. Kazak L, Cohen P. Creatine metabolism: energy homeostasis, immunity and cancer biology. Nat Rev Endocrinol (2020) 16:421–36. doi: 10.1038/s41574-020-0365-5
52. Samborska B, Roy DG, Rahbani JF, Hussain MF, Ma EH, Jones RG, et al. Creatine transport and creatine kinase activity is required for CD8(+) T cell immunity. Cell Rep (2022) 38:110446. doi: 10.1016/j.celrep.2022.110446
53. dos Santos G, Rogel MR, Baker MA, Troken JR, Urich D, Morales-Nebreda L, et al. Vimentin regulates activation of the NLRP3 inflammasome. Nat Commun (2015) 6:6574. doi: 10.1038/ncomms7574
54. Ives A, Nomura J, Martinon F, Roger T, LeRoy D, Miner JN, et al. Xanthine oxidoreductase regulates macrophage IL1beta secretion upon NLRP3 inflammasome activation. Nat Commun (2015) 6:6555. doi: 10.1038/ncomms7555
55. Adhikari B, Lee CN, Khadka VS, Deng Y, Fukumoto G, Thorne M, et al. RNA-Sequencing based analysis of bovine endometrium during the maternal recognition of pregnancy. BMC Genomics (2022) 23:494. doi: 10.1186/s12864-022-08720-4
56. Huang T, Huang X, Shi B, Liang X, Luo J, Yao M, et al. Relationship among MS4A8 expression, its variants, and the immune response in a porcine model of salmonella. Can J Anim Sci (2018) 98:778–86. doi: 10.1139/cjas-2017-0037
57. Jiang Y, Tang Y, Hoover C, Kondo Y, Huang D, Restagno D, et al. Kupffer cell receptor CLEC4F is important for the destruction of desialylated platelets in mice. Cell Death Differ (2021) 28:3009–21. doi: 10.1038/s41418-021-00797-w
58. Wu B, Wu Y, Tang W. Heme catabolic pathway in inflammation and immune disorders. Front Pharmacol (2019) 10:825. doi: 10.3389/fphar.2019.00825
59. Struyf S, Proost P, Lenaerts JP, Stoops G, Wuyts A, Van Damme J. Identification of a blood-derived chemoattractant for neutrophils and lymphocytes as a novel CC chemokine, regakine-1. Blood (2001) 97:2197–204. doi: 10.1182/blood.V97.8.2197
60. Rohrl J, Yang D, Oppenheim JJ, Hehlgans T. Specific binding and chemotactic activity of mBD4 and its functional orthologue hBD2 to CCR6-expressing cells. J Biol Chem (2010) 285:7028–34. doi: 10.1074/jbc.M109.091090
61. Dautovic E, Perisic Nanut M, Softic A, Kos J. The transcription factor C/EBP alpha controls the role of cystatin f during the differentiation of monocytes to macrophages. Eur J Cell Biol (2018) 97:463–73. doi: 10.1016/j.ejcb.2018.07.002
62. Moore SM, Holt VV, Malpass LR, Hines IN, Wheeler MD. Fatty acid-binding protein 5 limits the anti-inflammatory response in murine macrophages. Mol Immunol (2015) 67:265–75. doi: 10.1016/j.molimm.2015.06.001
63. Cekic C, Linden J. Purinergic regulation of the immune system. Nat Rev Immunol (2016) 16:177–92. doi: 10.1038/nri.2016.4
64. Leon B, Ballesteros-Tato A, Browning JL, Dunn R, Randall TD, Lund FE. Regulation of T(H)2 development by CXCR5+ dendritic cells and lymphotoxin-expressing b cells. Nat Immunol (2012) 13:681–90. doi: 10.1038/ni.2309
65. Ilarregui JM, Croci DO, Bianco GA, Toscano MA, Salatino M, Vermeulen ME, et al. Tolerogenic signals delivered by dendritic cells to T cells through a galectin-1-driven immunoregulatory circuit involving interleukin 27 and interleukin 10. Nat Immunol (2009) 10:981–91. doi: 10.1038/ni.1772
66. Ma X, Verweij EWE, Siderius M, Leurs R, Vischer HF. Identification of TSPAN4 as novel histamine H4 receptor interactor. Biomolecules (2021) 11(8):1127. doi: 10.3390/biom11081127
67. Harrison N, Koo CZ, Tomlinson MG. Regulation of ADAM10 by the TspanC8 family of tetraspanins and their therapeutic potential. Int J Mol Sci (2021) 22(13):6707. doi: 10.3390/ijms22136707
68. Skazik C, Heise R, Bostanci Ö, Paul N, Denecke B, Joussen S, et al. Differential expression of influx and efflux transport proteins in human antigen presenting cells. Exp Dermatol (2008) 17:739–47. doi: 10.1111/j.1600-0625.2008.00745.x
69. Sebastian K, Detro-Dassen S, Rinis N, Fahrenkamp D, Muller-Newen G, Merk HF, et al. Characterization of SLCO5A1/OATP5A1, a solute carrier transport protein with non-classical function. PloS One (2013) 8:e83257. doi: 10.1371/journal.pone.0083257
70. Bai L, Sato H, Kubo Y, Wada S, Aida Y. CAT1/SLC7A1 acts as a cellular receptor for bovine leukemia virus infection. FASEB J (2019) 33:14516–27. doi: 10.1096/fj.201901528R
71. Lawson JS, Salmons B, Glenn WK. Oncogenic viruses and breast cancer: Mouse mammary tumor virus (MMTV), bovine leukemia virus (BLV), human papilloma virus (HPV), and Epstein-Barr virus (EBV). Front Oncol (2018) 8:1. doi: 10.3389/fonc.2018.00001
72. Iwata M, Hirakiyama A, Eshima Y, Kagechika H, Kato C, Song SY. Retinoic acid imprints gut-homing specificity on T cells. Immunity (2004) 21:527–38. doi: 10.1016/j.immuni.2004.08.011
73. Bakdash G, Vogelpoel LT, van Capel TM, Kapsenberg ML, de Jong EC. Retinoic acid primes human dendritic cells to induce gut-homing, IL-10-producing regulatory T cells. Mucosal Immunol (2015) 8:265–78. doi: 10.1038/mi.2014.64
74. Bos A, van Egmond M, Mebius R. The role of retinoic acid in the production of immunoglobulin a. Mucosal Immunol (2022) 15:562–72. doi: 10.1038/s41385-022-00509-8
75. Junqueira Alves C, Yotoko K, Zou H, Friedel RH. Origin and evolution of plexins, semaphorins, and met receptor tyrosine kinases. Sci Rep (2019) 9:1970. doi: 10.1038/s41598-019-38512-y
76. Kanth SM, Gairhe S, Torabi-Parizi P. The role of semaphorins and their receptors in innate immune responses and clinical diseases of acute inflammation. Front Immunol (2021) 12:672441. doi: 10.3389/fimmu.2021.672441
77. Myokai F, Takashiba S, Lebo R, Amar S. A novel lipopolysaccharide-induced transcription factor regulating tumor necrosis factor alpha gene expression: molecular cloning, sequencing, characterization, and chromosomal assignment. Proc Natl Acad Sci U.S.A. (1999) 96:4518–23. doi: 10.1073/pnas.96.8.4518
78. Tang X, Marciano DL, Leeman SE, Amar S. LPS induces the interaction of a transcription factor, LPS-induced TNF-alpha factor, and STAT6(B) with effects on multiple cytokines. Proc Natl Acad Sci U.S.A. (2005) 102:5132–7. doi: 10.1073/pnas.0501159102
79. Murase M, Kawasaki T, Hakozaki R, Sueyoshi T, Putri DDP, Kitai Y, et al. Intravesicular acidification regulates lipopolysaccharide inflammation and tolerance through TLR4 trafficking. J Immunol (2018) 200:2798–808. doi: 10.4049/jimmunol.1701390
80. Zainol MIB, Kawasaki T, Monwan W, Murase M, Sueyoshi T, Kawai T. Innate immune responses through toll-like receptor 3 require human-antigen-R-mediated Atp6v0d2 mRNA stabilization. Sci Rep (2019) 9:20406. doi: 10.1038/s41598-019-56914-w
81. Shen L, Hu P, Zhang Y, Ji Z, Shan X, Ni L, et al. Serine metabolism antagonizes antiviral innate immunity by preventing ATP6V0d2-mediated YAP lysosomal degradation. Cell Metab (2021) 33:971–987 e6. doi: 10.1016/j.cmet.2021.03.006
82. Duckworth BC, Lafouresse F, Wimmer VC, Broomfield BJ, Dalit L, Alexandre YO, et al. Effector and stem-like memory cell fates are imprinted in distinct lymph node niches directed by CXCR3 ligands. Nat Immunol (2021) 22:434–48. doi: 10.1038/s41590-021-00878-5
83. Li J, Lu E, Yi T, Cyster JG. EBI2 augments tfh cell fate by promoting interaction with IL-2-quenching dendritic cells. Nature (2016) 533:110–4. doi: 10.1038/nature17947
84. Ardouin L, Luche H, Chelbi R, Carpentier S, Shawket A, Montanana Sanchis F, et al. Broad and largely concordant molecular changes characterize tolerogenic and immunogenic dendritic cell maturation in thymus and periphery. Immunity (2016) 45:305–18. doi: 10.1016/j.immuni.2016.07.019
85. Maier B, Leader AM, Chen ST, Tung N, Chang C, LeBerichel J, et al. A conserved dendritic-cell regulatory program limits antitumour immunity. Nature (2020) 580:257–62. doi: 10.1038/s41586-020-2134-y
86. Blecher-Gonen R, Bost P, Hilligan KL, David E, Salame TM, Roussel E, et al. Single-cell analysis of diverse pathogen responses defines a molecular roadmap for generating antigen-specific immunity. Cell Syst (2019) 8:109–121.e6. doi: 10.1016/j.cels.2019.01.001
87. Lee A, Scott MKD, Wimmers F, Arunachalam PS, Luo W, Fox CB, et al. A molecular atlas of innate immunity to adjuvanted and live attenuated vaccines, in mice. Nat Commun (2022) 13:549. doi: 10.1038/s41467-022-28197-9
88. James KR, Gomes T, Elmentaite R, Kumar N, Gulliver EL, King HW, et al. Distinct microbial and immune niches of the human colon. Nat Immunol (2020) 21:343–53. doi: 10.1038/s41590-020-0602-z
89. Di Pilato M, Kfuri-Rubens R, Pruessmann JN, Ozga AJ, Messemaker M, Cadilha BL, et al. CXCR6 positions cytotoxic T cells to receive critical survival signals in the tumor microenvironment. Cell (2021) 184:4512–4530 e22. doi: 10.1016/j.cell.2021.07.015
90. Jakubzick C, Gautier EL, Gibbings SL, Sojka DK, Schlitzer A, Johnson TE, et al. Minimal differentiation of classical monocytes as they survey steady-state tissues and transport antigen to lymph nodes. Immunity (2013) 39:599–610. doi: 10.1016/j.immuni.2013.08.007
91. Tamoutounour S, Guilliams M, Montanana Sanchis F, Liu H, Terhorst D, Malosse C, et al. Origins and functional specialization of macrophages and of conventional and monocyte-derived dendritic cells in mouse skin. Immunity (2013) 39:925–38. doi: 10.1016/j.immuni.2013.10.004
92. Bleriot C, Chakarov S, Ginhoux F. Determinants of resident tissue macrophage identity and function. Immunity (2020) 52:957–70. doi: 10.1016/j.immuni.2020.05.014
93. Bellomo A, Gentek R, Bajenoff M, Baratin M. Lymph node macrophages: Scavengers, immune sentinels and trophic effectors. Cell Immunol (2018) 330:168–74. doi: 10.1016/j.cellimm.2018.01.010
94. Moran I, Grootveld AK, Nguyen A, Phan TG. Subcapsular sinus macrophages: The seat of innate and adaptive memory in murine lymph nodes. Trends Immunol (2019) 40:35–48. doi: 10.1016/j.it.2018.11.004
95. Louie DAP, Liao S. Lymph node subcapsular sinus macrophages as the frontline of lymphatic immune defense. Front Immunol (2019) 10:347. doi: 10.3389/fimmu.2019.00347
96. Baratin M, Simon L, Jorquera A, Ghigo C, Dembele D, Nowak J, et al. T Cell zone resident macrophages silently dispose of apoptotic cells in the lymph node. Immunity (2017) 47:349–362 e5. doi: 10.1016/j.immuni.2017.07.019
97. Guilliams M, Scott CL. Does niche competition determine the origin of tissue-resident macrophages? Nat Rev Immunol (2017) 17:451–60. doi: 10.1038/nri.2017.42
98. Leylek R, Alcantara-Hernandez M, Granja JM, Chavez M, Perez K, Diaz OR, et al. Chromatin landscape underpinning human dendritic cell heterogeneity. Cell Rep (2020) 32:108180. doi: 10.1016/j.celrep.2020.108180
99. Leylek R, Alcantara-Hernandez M, Lanzar Z, Ludtke A, Perez OA, Reizis B, et al. Integrated cross-species analysis identifies a conserved transitional dendritic cell population. Cell Rep (2019) 29:3736–3750 e8. doi: 10.1016/j.celrep.2019.11.042
100. Alcantara-Hernandez M, Leylek R, Wagar LE, Engleman EG, Keler T, Marinkovich MP, et al. High-dimensional phenotypic mapping of human dendritic cells reveals interindividual variation and tissue specialization. Immunity (2017) 47:1037–1050 e6. doi: 10.1016/j.immuni.2017.11.001
101. See P, Dutertre CA, Chen J, Günther P, McGovern N, Irac SE, et al. Mapping the human DC lineage through the integration of high-dimensional techniques. Science (2017) 356(6342):eaag3009. doi: 10.1126/science.aag3009
102. Diao J, Winter E, Cantin C, Chen W, Xu L, Kelvin D, et al. In situ replication of immediate dendritic cell (DC) precursors contributes to conventional DC homeostasis in lymphoid tissue. J Immunol (2006) 176:7196–206. doi: 10.4049/jimmunol.176.12.7196
103. Liu K, Victora GD, Schwickert TA, Guermonprez P, Meredith MM, Yao K, et al. In vivo analysis of dendritic cell development and homeostasis. Science (2009) 324:392–7. doi: 10.1126/science.1170540
Keywords: single-cell RNA-seq (scRNA-seq), dendritic cells, monocytes, macrophages, mesenteric lymph node, cattle
Citation: Barut GT, Kreuzer M, Bruggmann R, Summerfield A and Talker SC (2023) Single-cell transcriptomics reveals striking heterogeneity and functional organization of dendritic and monocytic cells in the bovine mesenteric lymph node. Front. Immunol. 13:1099357. doi: 10.3389/fimmu.2022.1099357
Received: 15 November 2022; Accepted: 13 December 2022;
Published: 06 January 2023.
Edited by:
Elodie Segura, Institut Curie, FranceReviewed by:
Julie Helft, Institut National de la Santé et de la Recherche Médicale (INSERM), FrancePaul Gueguen, Université de Lausanne, Switzerland
Copyright © 2023 Barut, Kreuzer, Bruggmann, Summerfield and Talker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephanie C. Talker, c3RlcGhhbmllLnRhbGtlckB1bmliZS5jaA==