
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 08 December 2022
Sec. Inflammation
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.1066036
This article is part of the Research TopicTargeting the microbiota to attenuate chronic inflammationView all 16 articles
 Carlos Blázquez-Bondia1
Carlos Blázquez-Bondia1 Mariona Parera1
Mariona Parera1 Francesc Català-Moll1
Francesc Català-Moll1 Maria Casadellà1
Maria Casadellà1 Aleix Elizalde-Torrent1
Aleix Elizalde-Torrent1 Meritxell Aguiló2
Meritxell Aguiló2 Jordi Espadaler-Mazo2José Ramon Santos3
Jordi Espadaler-Mazo2José Ramon Santos3 Roger Paredes1,3,4,5,6
Roger Paredes1,3,4,5,6 Marc Noguera-Julian1,4,6*
Marc Noguera-Julian1,4,6*Background: Some HIV-1 infected patients are unable to completely recover normal CD4+ T-cell (CD4+) counts after achieving HIV-1 suppression with combined Antiretroviral Therapy (cART), hence being classified as immuno-discordant. The human microbiome plays a crucial role in maintaining immune homeostasis and is a potential target towards immune reconstitution.
Setting: RECOVER (NCT03542786) was a double-blind placebo-controlled clinical trial designed to evaluate if the novel probiotic i3.1 (AB-Biotics, Sant Cugat del Vallès, Spain) was able to improve immune reconstitution in HIV-1 infected immuno-discordant patients with stable cART and CD4+ counts <500 cells/mm3. The mixture consisted of two strains of L. plantarum and one of P. acidilactici, given with or without a fiber-based prebiotic.
Methods: 71 patients were randomized 1:2:2 to Placebo, Probiotic or probiotic + prebiotic (Synbiotic), and were followed over 6 months + 3-month washout period, in which changes on systemic immune status and gut microbiome were evaluated. Primary endpoints were safety and tolerability of the investigational product. Secondary endpoints were changes on CD4+ and CD8+ T-cell (CD8+) counts, inflammation markers and faecal microbiome structure, defined by alpha diversity (Gene Richness), beta diversity (Bray-Curtis) and functional profile. Comparisons across/within groups were performed using standard/paired Wilcoxon test, respectively.
Results: Adverse event (AE) incidence was similar among groups (53%, 33%, and 55% in the Placebo, Probiotic and Synbiotic groups, respectively, the most common being grade 1 digestive AEs: flatulence, bloating and diarrhoea. Two grade 3 AEs were reported, all in the Synbiotic group: abdominal distension (possibly related) and malignant lung neoplasm (unrelated), and 1 grade 4 AE in the Placebo: hepatocarcinoma (unrelated). Synbiotic exposure was associated with a higher increase in CD4+/CD8+ T-cell (CD4/CD8) ratio at 6 months vs baseline (median=0.76(IQR=0.51) vs 0.72(0. 45), median change= 0.04(IQR=0.19), p = 0.03). At month 9, the Synbiotic group had a significant increase in CD4/CD8 ratio (0.827(0.55) vs 0.825(0.53), median change = 0.04(IQR=0.15), p= 0.02) relative to baseline, and higher CD4+ counts (447 (157) vs. 342(73) counts/ml, p = 0.03), and lower sCD14 values (2.16(0.67) vs 3.18(0.8), p = 0.008) than Placebo. No effect in immune parameters was observed in the Probiotic arm. None of the two interventions modified microbial gene richness (alpha diversity). However, intervention as categorical variable was associated with slight but significant effect on Bray-Curtis distance variance (Adonis R2 = 0.02, p = 0.005). Additionally, at month 6, Synbiotic intervention was associated with lower pathway abundances vs Placebo of Assimilatory Sulphate Reduction (8.79·10-6 (1.25·10-5) vs. 1.61·10-5 (2.77·10-5), p = 0.03) and biosynthesis of methionine (2.3·10-5 (3.17·10-5) vs. 4·10-5 (5.66·10-5), p = 0.03) and cysteine (1.83·10-5 (2.56·10-5) vs. 3.3·10-5 (4.62·10-5), p = 0.03). At month 6, probiotic detection in faeces was associated with significant decreases in C Reactive Protein (CRP) vs baseline (11.1(22) vs. 19.2(66), median change= -2.7 (13.2) ug/ml, p = 0.04) and lower IL-6 values (0.58(1.13) vs. 1.17(1.59) ug/ml, p = 0.02) when compared with samples with no detectable probiotic. No detection of the probiotic was associated with higher CD4/CD8 ratio at month 6 vs baseline (0.718(0.57) vs. 0.58(0.4), median change = 0.4(0.2), p = 0.02). After washout, probiotic non-detection was also associated with a significant increase in CD4+ counts (457(153) vs. 416(142), median change = 45(75), counts/ml, p = 0.005) and CD4/CD8 ratio (0.67(0.5) vs 0.59(0.49), median change = 0.04 (0.18), p = 0.02).
Conclusion: A synbiotic intervention with L. plantarum and P. acidilactici was safe and led to small increases in CD4/CD8 ratio and minor reductions in sCD14 of uncertain clinical significance. A probiotic with the same composition was also safe but did not achieve any impact on immune parameters or faecal microbiome composition.
One of the key aspects of HIV infection is a fast and widespread destruction of CD4+ T-lymphocytes (1), which becomes more exacerbated in the latest stages of infection (2, 3). Additionally, the virus presents the ability to establish reservoirs in which it can remain dormant mostly in high-CCR5 memory CD4+ T-cells (4, 5), where it remains integrated in the host genome (3–5). The ability to maintain a latency state has made it impossible to achieve a complete remission, although it can be life-long supressed in most patients with combined Antiretroviral Therapy (cART) (6). Nevertheless, even when the virus remains supressed, an important fraction of HIV-infected people will become immunodiscordant, as they fail to fully recover CD4+ counts and immune function (7–9) especially those who failed to receive cART on the early stages of infection.
This lack of recovery stems from the virus early replication site and its reservoir sanctuary: the Gut-Associated Lymphoid Tissue (GALT) (10), where the biggest population of high CCR5+ CD4+ T-cells resides. Depletion of such cells in the gut is coupled with a decrease in Treg numbers (11), but more slowly than its CD4+ counterpart, lowering the ratio between Treg and effector CD4+ cells, especially the Th17 subtypes (11, 12). A relative decrease of the Th17/Treg ratio has been correlated to a higher chance of disease progression (13). Th17 have also been shown to maintain gut barrier integrity, stimulating tight junction expression in epithelial cells (14), as well as modulating bacterial populations in the mucosa by secreting antimicrobial peptides (15). Hence, the skew in the Treg/Th17 ratio compromises the gut barrier integrity, and creates a feedback loop where dysbiosis and gut inflammation cause a leakage of bacterial compounds into the bloodstream (16), which increases residual systemic inflammation (17) and leads to further T-cell exhaustion and senescence (18), which further promotes dysbiosis and gut barrier. This vicious circle ends up causing immune exhaustion and may hinder any attempt of immune reconstitution.
While many approaches have been considered to recover immunity and gut integrity, modulation of the gut microbiome awakened great interest recently. It is now known that some bacterial species directly affect the immunologic makeup of the gut barrier (19) by modulating tryptophan to kynurenine catabolism (17). This pathway is thought to promote Treg differentiation (17) and has been shown to increase with presence of some Proteobacteria (20) and decrease with Lactobacillus species (20, 21). For this, probiotics have been widely tested with promising results for wide variety of ailments, both from the gut itself such as Inflammatory Bowel Disease (IBD) (22), diarrhoea, both HIV-induced (23) and by other pathogens (24) and even outside of the gut, such as allergies and upper respiratory infections (25).
Recently, the gut microbiome has awakened a great interest in the scope of HIV clinical management, as immune recovery has been shown to be affected by the state of the gut mucosa (26). Cross-sectional studies have found microbiome signatures correlated to immune reconstitution such as higher Prevotella/Bacteroides ratio and enrichment of Faecalibacterium prausnitzii and Coprococcus comes (27) or increased abundance of Fusobacterium negatively linked to immune recovery (28). Consequently, probiotic interventions have have become an interesting therapeutic target, as they promote tolerogenicity (29), displace pathogenic strains, and reduce inflammation (30), which could reduce T cell depletion and senescence, opening a way to improve immune reconstitution after viral suppression. However, many different combinations of probiotic strains and prebiotic substrates have been tested with mixed results (30, 31).
In this study, we performed a randomized double-blinded trial to test the safety, tolerability and effectiveness of a probiotic consisting of two strains of Lactobacillus plantarum and one of Pediococcus acidilactici, combined with prebiotic fibers over the course of 6 months, followed by a 3-month long washout period in immunodiscordant (<500 counts/ml) HIV patients with stable cART. The primary endpoints consisted of safety and tolerability. The secondary endpoints were changes in CD4+, CD8+ counts, CD4/CD8 ratio, inflammation, and gut permeability markers, as well as changes in the gut microbiome taxonomical and functional composition after 6 months of intervention + 3 of washout.
The study was reviewed and approved by the Institutional Review Board of the Hospital Universitari Germans Trias i Pujol (reference PI-13-046). All participants provided written informed consent in accordance with the World Medical Association Declaration of Helsinki, Fortaleza and Brazil, October 2013 and personal data was managed according to Spanish data protection law (LOPD 15/1999). The study concept, design, patient information and results were discussed with the FLSida Community Advisory Committee, in accordance with AB-Biotics internal QC auditing. All available information can be found in the protocol (Supplementary Methods), and the study is registered in clinicaltrials.gov, accession: NCT03542786.
This study took place in a two-year span between 2017 and 2019 and was designed as a masked randomized, placebo-controlled, double-blinded three-arm study in a cohort of 100 HIV+ patients with the following inclusion criteria: 18 years of age or older, chronic HIV infection with stable Anti-Retroviral Treatment (cART) ongoing for longer than a year prior to the start, peripheral CD4+ counts lower than 500 cells/ml in plasma, <50 HIV copies/ml in plasma for at least 6 months before the start, no antibiotic treatments at least 1 month before start, lack of severe AIDS-defining diseases and no pregnancy. An additional filter was later implemented, in which only those patients with at least 2 samples along the trial would be selected for further analysis, to preserve the longitudinal approach of the study.
After inclusion, patients were randomly assigned to one of the three following groups: Placebo, Probiotic or Synbiotic in a 1:2:2 ratio and matched by 3rd cART drug class: Integrase Strand Transfer Inhibitors (INSTI), Non-nucleoside reverse transcriptase inhibitors (NNRTI) or Protease Inhibitors (PI), and CD4+ nadir higher or lower than 200 cells/ml at the time of screening.
All recruited patients followed a 6-month treatment followed by a 3-month washout periods, with check-ups at months 0,1,3,6 and 9. Every follow-up visit consisted of a sample collection of both blood and faeces (except at the 1st month checkout, where only stool was collected), a physical examination and a questionnaire about quality of life, and self-reported treatment adherence since last check-up. During the treatment period, all participants received different formulation depending on whether they received a prebiotic + probiotic (Synbiotic), probiotic alone (Probiotic) or a placebo (Placebo), which was administered orally as dissolved powder sachets, daily.
The probiotic used in this study consisted of a mix of 3 Lactobacillales strains: L. plantarum (strains CECT7484 and CECT7485) and P. acidilactici (strain CECT7483). In the Synbiotic group, the probiotic was co-administered with two different mixes of vegetal fibers consisting of pectin, inulin, oat, acacia, maltodextrin polydextrose and Partially Hidrolyzed Guar Gum (PHGG) that were alternatively combined with the probiotic every other week (the exact formulation can be found in the protocol). The Probiotic and Placebo groups received excipient-containing envelopes that were identical to those of the Synbiotic group to preserve the double-blind. The exact composition and manufacturing process can be found in Supplementary Methods.
Blood samples were collected in fasting conditions at the same time as stool samples, when possible. A fraction of these samples was used as whole blood to perform peripheral CD4+ and CD8+ counts by flow cytometry at the Germans Trias i Pujol Hospital. Soluble markers of microbial translocation in plasma (sCD14 and LBP) and inflammation markers (IL-6, D-Dimer and CRP) were quantified using sCD14 and LBP DuoSet ELISA development system (pg/mL), R&D systems (Minneapolis, MN), Human IL6- High Sensitivity ELISA, Invitrogen (Waltham, Massachusetts, USA) and RayBio Human D-Dimer or CRP Elisa Kit, RayBiotech (Peachtree Corners, GA, USA), respectively.
Stool samples were collected by the patient or nursing staff according to the GUT (DNA Stabilized-frozen Inc., Ottawa, Ontario, Canada) extraction kit. Samples were stored at -80°C till processing. Faecal DNA was extracted using the PowerSoil DNA Extraction Kit (MO BIO Laboratories, Carlsbad, CA, USA), which was then fragmented into 300 bp clone-sized libraries using Nextera-XT Illumina kit (Illumina, Inc. San Diego, CA, USA) and sequenced in an Illumina HiSeq sequencer (Illumina, Inc. San Diego, CA, USA) with a sequencing depth target of 20 million reads.
Raw.fastq files were first processed for quality control. Read Quality filtering and trimming was performed with trimmomatic (32), with a 30-nt sliding window approach, trimming when the average phred score dropped below 20. Trimmed reads were then aligned against the human Hg19 genomic database using bowtie2 to remove any human DNA contamination.
Taxonomy assessment was performed with Metaphlan3 (33), performing a marker gene-based quantification, using the CHOCOPhlan 201901.1 database. The obtained data was packaged into an R phyloseq structure.
Parallel to taxonomic analysis, gene function and metabolic pathway quantification was performed from the raw, quality-filtered sequencing data using HUMAnN3.0 (33). The software was run with its standard configuration and built-in databases. Results from all samples were combined into a unique table using HUMAnN3 inner script merge_tables.sh and clustered into Metacyc pathway abundances.
In this study, ecological alpha diversity was studies as gene richness. To obtain it, the post-QC sequencing data was aligned against the Integrated Gene Catalog (IGC) database (34) using Bowtie2 (35). The output was sampled at different numbers of reads to obtain rarefaction curves, from which the minimum sampling threshold was defined as the 95th percentile of maximum per-sample coverages, which equalled 2·107 reads. Samples with max coverages below this threshold were discarded, and those above were subsampled to said value to remove coverage biases.
For beta diversity, the taxonomy tables were used to construct pairwise Bray-Curtis distance matrixes between samples. The obtain matrices were then projected into NMDS coordinates using the function MetaMDS from the R vegan (36) package using their default configuration.
All statistical testing was performed using R 4.0.2. Since most of the quantitative variables tested departed from normality, assessed by Shapiro-wilk test, comparisons across group were performed using Wilcoxon Rank-Sum test, while within-group, longitudinal comparisons were performed using Wilcoxon rank-sum matched-pairs test, with Benjamini-Hochberg correction. In the case of gene pathways, Kruskal-Wallis tests between groups were performed to filter out those pathways which changed in any timepoint other than basal, before proceeding to the pairwise group/timepoint comparisons using a p < 0.05 cut-off. Statistical results are reported as median (IQR), and longitudinal tests included median change (IQR). The number of patients per group for each statistical comparison performed in this study can be found in Supplementary Tables 3, 4, for categorical and longitudinal comparisons, respectively.
In addition, we studied the changes in time for numerical variables (diversity, inflammation, T-cells, and pathways) with Linear Mixed Models (LMM) using the Lme4 package. Models were built in two different approaches: one where the dataset was split by group, in which the LMM would be built for each using time as the only fixed effect, in order to study the magnitude of the change for each group and its significance, and another where an LMM was used with the entire dataset with both group and time as fixed effects, to assess the difference in change in slope between groups and its significance via two-way ANOVA. In all cases, the patients individual IDs were inputted as the random variable.
Bray Curtis distances were compared between groups using PERMANOVA. T-cell counts, inflammation and gut permeability markers were correlated with the NMDS coordinates using Spearman correlation.
From the original goal of 100 patients recruited of which 92 passed all the inclusion criteria, 89 patients were successfully randomised for the study. At study ending, 53 patients had complete follow-up and 36 had incomplete follow-up, of which 18 provided samples for at least two timepoints. Thus, 71 patients were finally selected for analysis, resulting in a proportion of 18 patients in the Placebo, 21 in the Probiotic and 32 in the Synbiotic groups (Figure 1). The self-reported mean adherences to the treatment showed no significant differences between groups (96% for the Placebo, 89.7% for Probiotic and 96.6% for the Synbiotic group). Demographic and clinical variables (Table 1, Supplementary Table 1) were tested with the corresponding statistical method depending on their normality according to a previous Shapiro test.
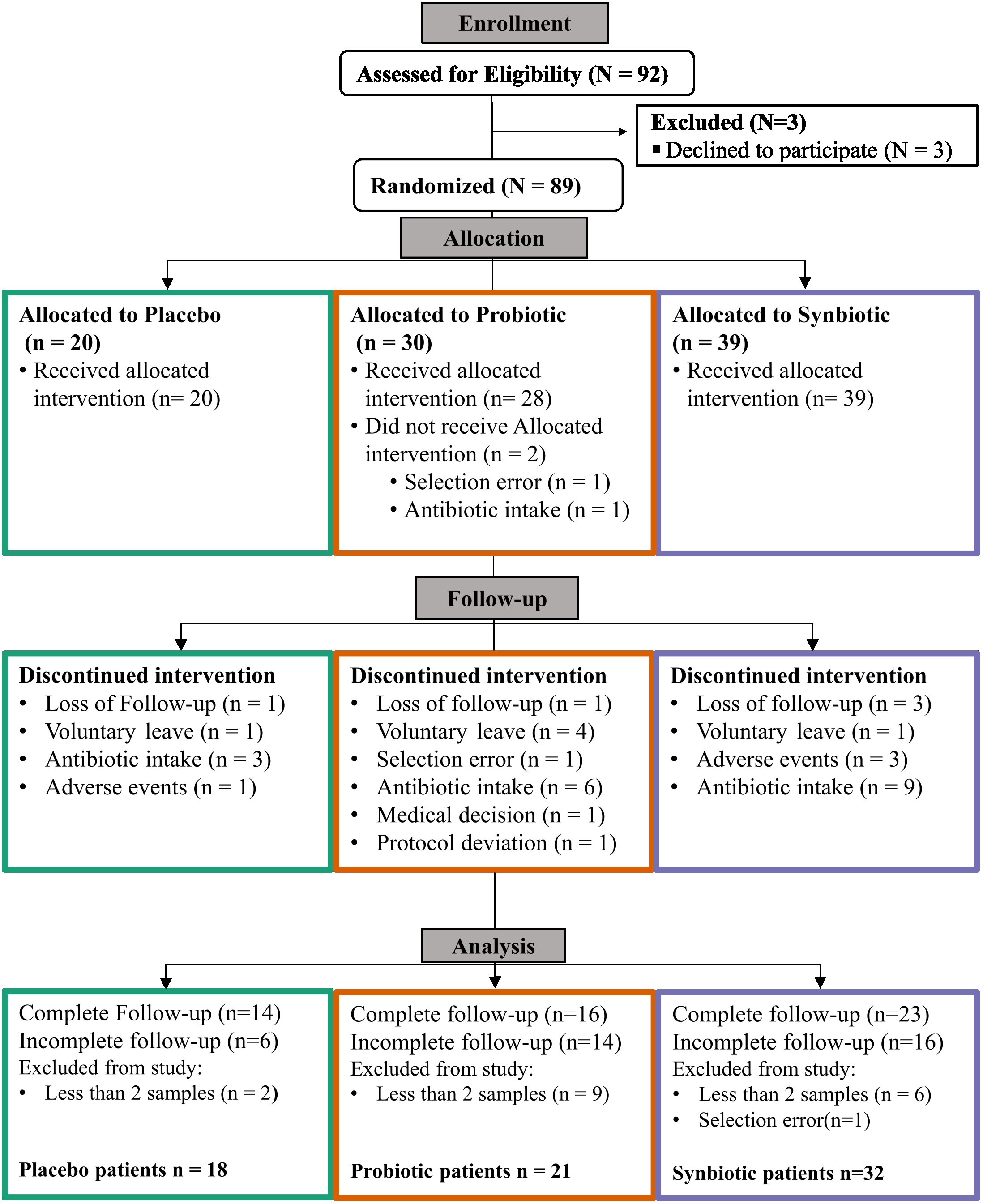
Figure 1 CONSORT (Consolidated Standards of Reporting Trials) Flow diagram.
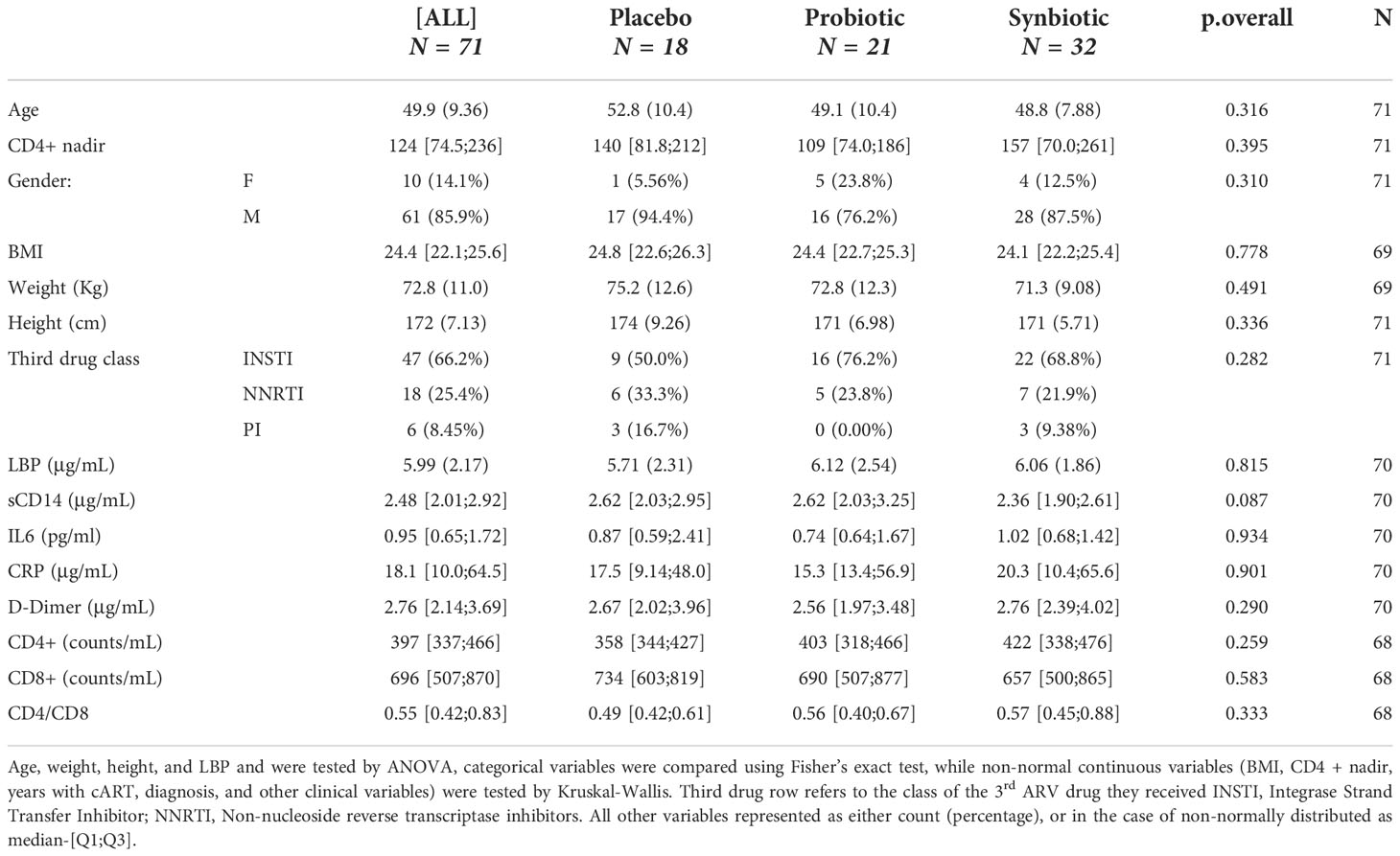
Table 1 Study population description of demographic variables and clinical markers at baseline: Age, height, LBP, and weight represented as mean-(sd), as they followed a continuous normal distribution (Shapiro test pval < 0.05).
The proportion of patients who suffered at least one Adverse Event (AE) during the study remained comparable between all arms, with no significant differences found: 9 patients with AE (50%) in the Placebo, 7 (33%) in the Probiotic and 17 (53%) in the Synbiotic groups (Supplementary Table 2). Most AEs consisted of severity grade 1 and 2, being the gastrointestinal conditions the most frequent, especially flatulence, dyspepsia, and diarrhoea, although other conditions were reported, but could not be associated with treatment group assignation. Two instances of severe grade 3 AEs were reported: abdominal distension (possibly related) and malignant lung neoplasm (unrelated), cancer (unrelated), while one grade 4 event (hepatocarcinoma, unrelated) was reported in the Placebo group.
CD4/CD8 ratio showed a slight albeit significant increase in the Synbiotic group at month 6 respect to baseline (median=0.76 (IQR=0.51) vs 0.72 (0.45), median change= 0.04 (0.19), p = 0.03). At month 9, the Synbiotic group still had increased CD4/CD8 vs baseline (0.827 (0.55) vs 0.825 (0.53), median change = 0.04 (0.15), p= 0.02), CD4+ was higher (447 (157) vs 342 (73) counts/ml, p = 0.03), and sCD14 was lower (2.16 (0.67) vs 3.18 (0.8) p = 0.008) than Placebo (Figure 2).
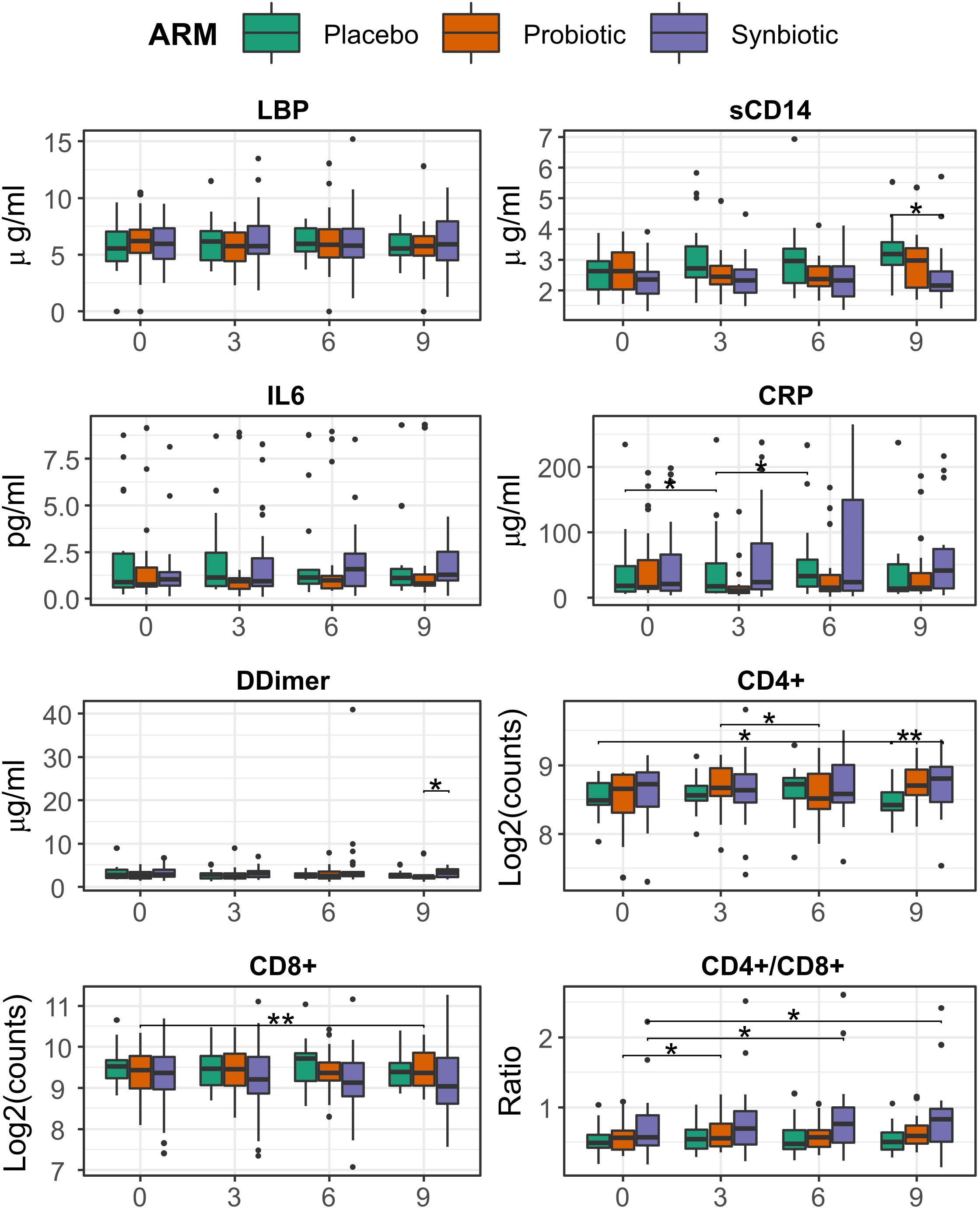
Figure 2 Evolution of T-cell counts, ratio, inflammation, and bacterial translocation markers between months 0 and 6, among treatment group. Comparisons within group and between time points were performed by Wilcoxon test, in its paired form for longitudinal differences and unpaired for cross-group comparisons. Significance was coded as follows: *(p < 0.05), **(p < 0.01).
Analysis with LMMs mirrored most of these previous findings, a significant increasing trend was found for both CRP (ANOVA p = 0.049, slope = 4.5) and CD4/CD8 ratio (p = 0.002, slope = 0.012) in the Synbiotic group (Supplementary Figure 2).
Finally, CD8+ counts appeared to be impacted by overall microbiome structure, as it was positively correlated with the 2nd coordinate of the NMDS (Supplementary Figure 1) and, as a result, CD4/CD8 ratio was negatively correlated (Supplementary Table 5)
No significant differences in gene richness were detected either between groups or longitudinally within groups (Figure 3A). Beta diversity did not increase either, as no changes in Bray Curtis distance vs each patient’s respective baseline could be observed (Figure 3B). NMDS showed no significant clustering based on Bray-Curtis distance and the intervention variable only explained 1.8% of the variance (Adonis R2 = 0.02, P=0.0051; Figure 3C).
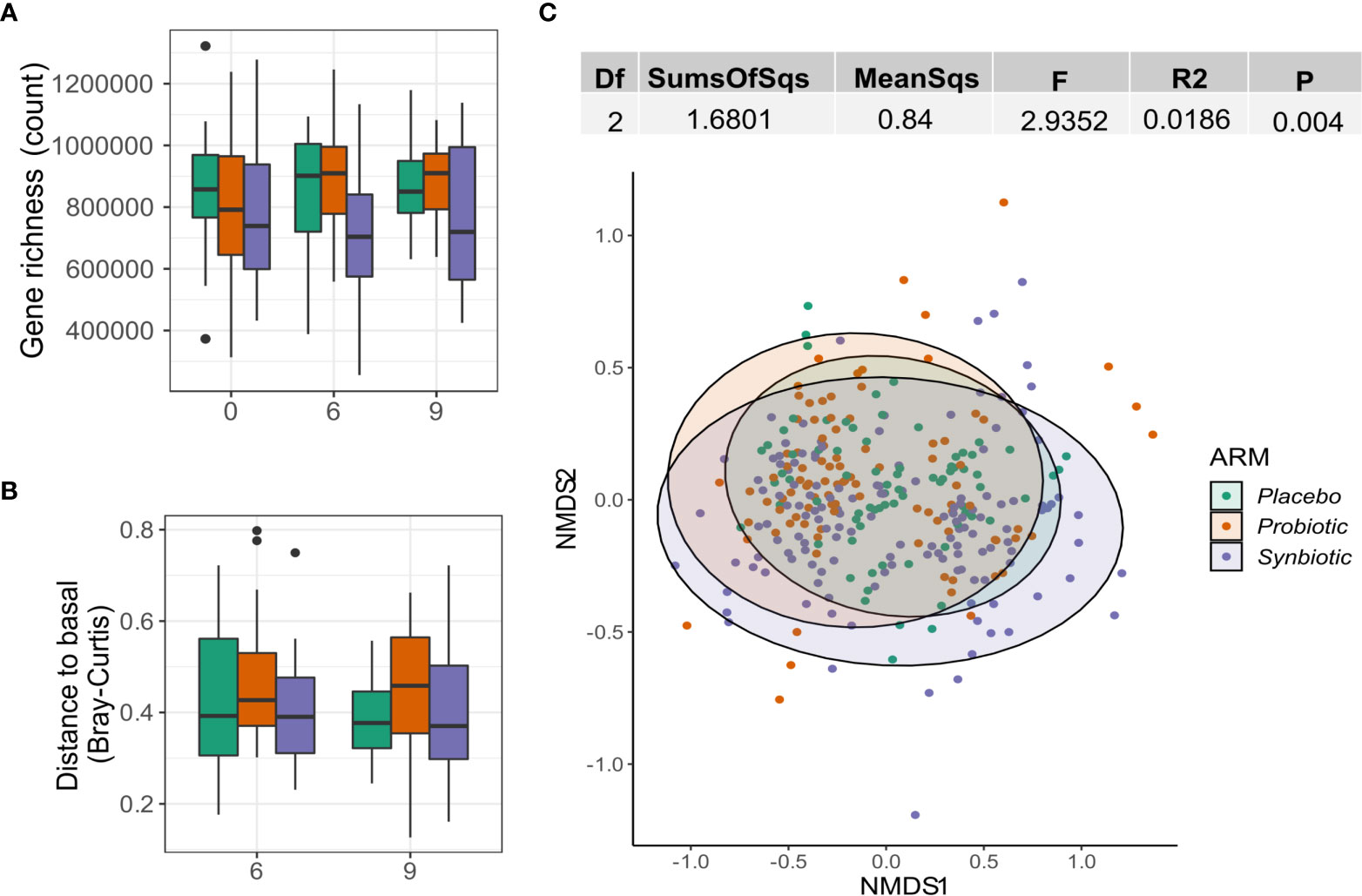
Figure 3 Effect of probiotics on the gut microbiome. (A) Gene richness counts for each intervention arm and month. Comparisons between group at each timepoint were performed using Wilcoxon test, while the paired version was used for within-group comparisons between months. (B) Distribution of Bray Curtis-Distances from baseline for each month and intervention arm. (C) NMDS showing sample ordination according to Bray-Curtis distance. Group distance dissimilarity was tested using ADONIS.
An exploratory analysis between groups identified 3 biochemical pathways with significant changes in relative abundance at month 6 (Kruskal-Wallis p < 0.01). Those pathways were consistently lower in the Synbiotic group respective to the Placebo: L-methionine biosynthesis (Met) (2.3·10-5 (3.17·10-5) vs 4·10-5 (5.66·10-5), p = 0.03), Assimilatory Sulphate Reduction (ASR) (median = 8.79·10-6 (IQR=1.25·10-5) vs 1.61·10-5 (2.77·10-5), p = 0.03), and cysteine biosynthesis (Cys) (1.83·10-5 (2.56·10-5) vs 3.3·10-5 (4.62·10-5), p = 0.03). At month 9, all three pathways increased in the Synbiotic group vs month 6 (Met: 3.8·10-5 (4.2·10-5) vs 2.3·10-5 (3.17·10-5), median change = 1.4·10-5 (3.12·10-5), p = 0.03; ASR: 1.5·10-5 (1.7·10-5) vs 8.79·10-6 (1.3·10-5) median change = 5.6·10-6 (1.3·10-5); p = 0.03; Cys: 3.1·10-5 (3.2·10-5) vs 1.8·10-5 (2.6·10-5), median change= 1.13·10-5 (2.42·10-5), p = 0.03), but no significant changes were found in the other groups (Figure 4).
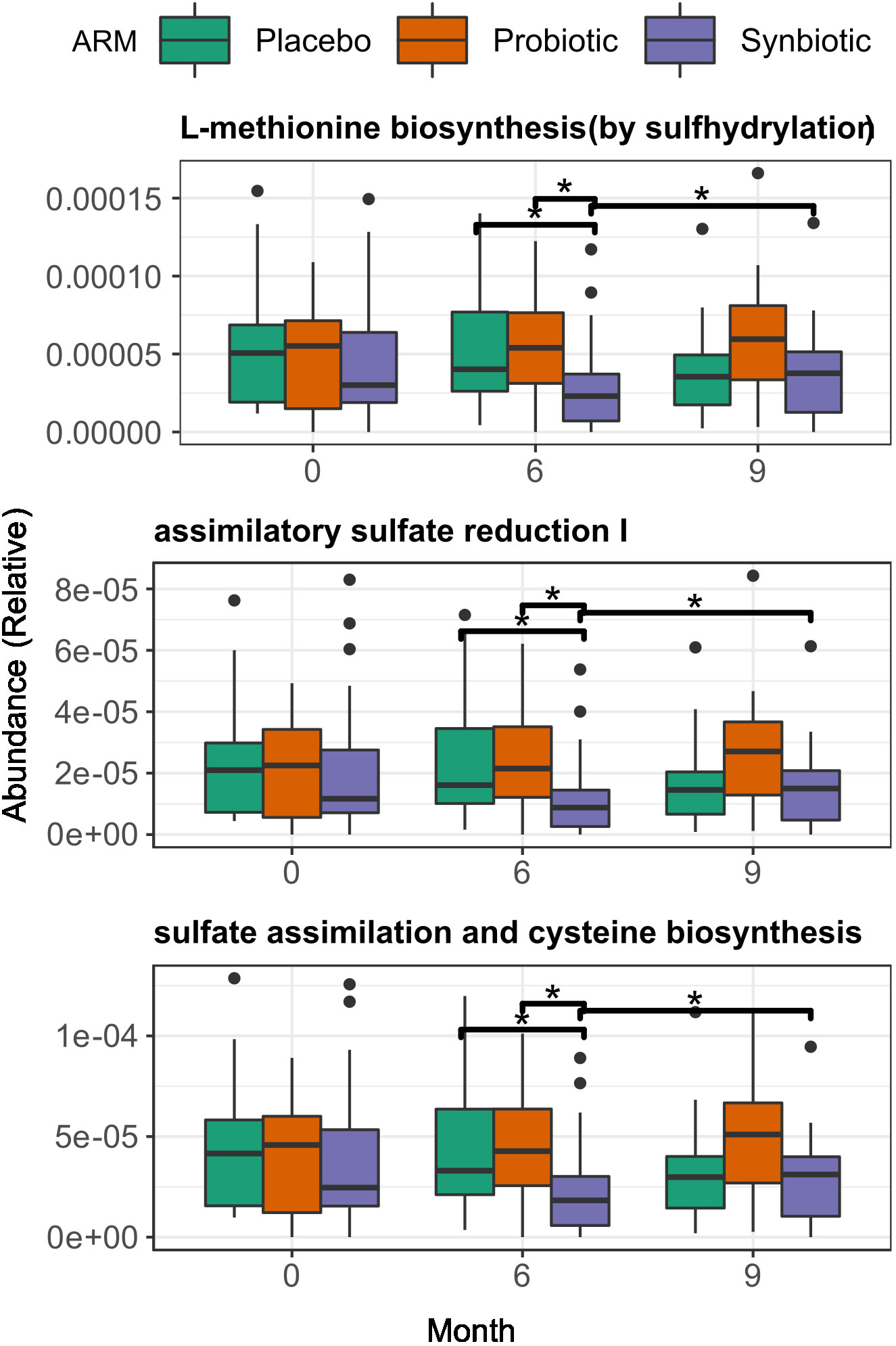
Figure 4 Changes in gene function at both start and end of the treatment period, among treatment group. Across group comparisons were performed with regular Wilcoxon test and longitudinal comparisons were performed using matched-samples Wilcoxon test. Significance was coded as follows: *(p < 0.05).
A more in-depth approach with LMMs suggested a significant decreasing trend in the relative abundances of all three pathways over time in the Synbiotic group (Met: p= 0.003, slope = -3.12·10-6; ASR: p = 0.003, slope = -1.59·10-6; Cys: p = 0.003, slope = -2.53·10-6) (Supplementary Figure 4). Additionally, a test of fixed effect interaction found a significant effect of intervention over the magnitude of change over time of pathway relative abundance (ANOVA = 0.008, Supplementary Figure 6A).
To better understand the lack of microbiome changes and the uncertain clinical changes, we tested how well the probiotic strains could maintain their presence in the gut. Only 6 (28.6%) patients in the Probiotic and 10 (31.3%) in the Synbiotic groups had detectable levels of either L. plantarum or P. acidilactici in at least two stool samples at months 1, 3 or 6, although in these cases, probiotic species median relative abundance started declining after the 3rd month of treatment, long before the wash-out phase. In the remaining patients, probiotic species were detected in only one of the timepoints in 5(23.8%) and 13(40%) of the Probiotic and Synbiotic, respectively. None of the probiotic strains was detected in any sample obtained at baseline, month 9 or in the Placebo group at any time point (Figure 5A).
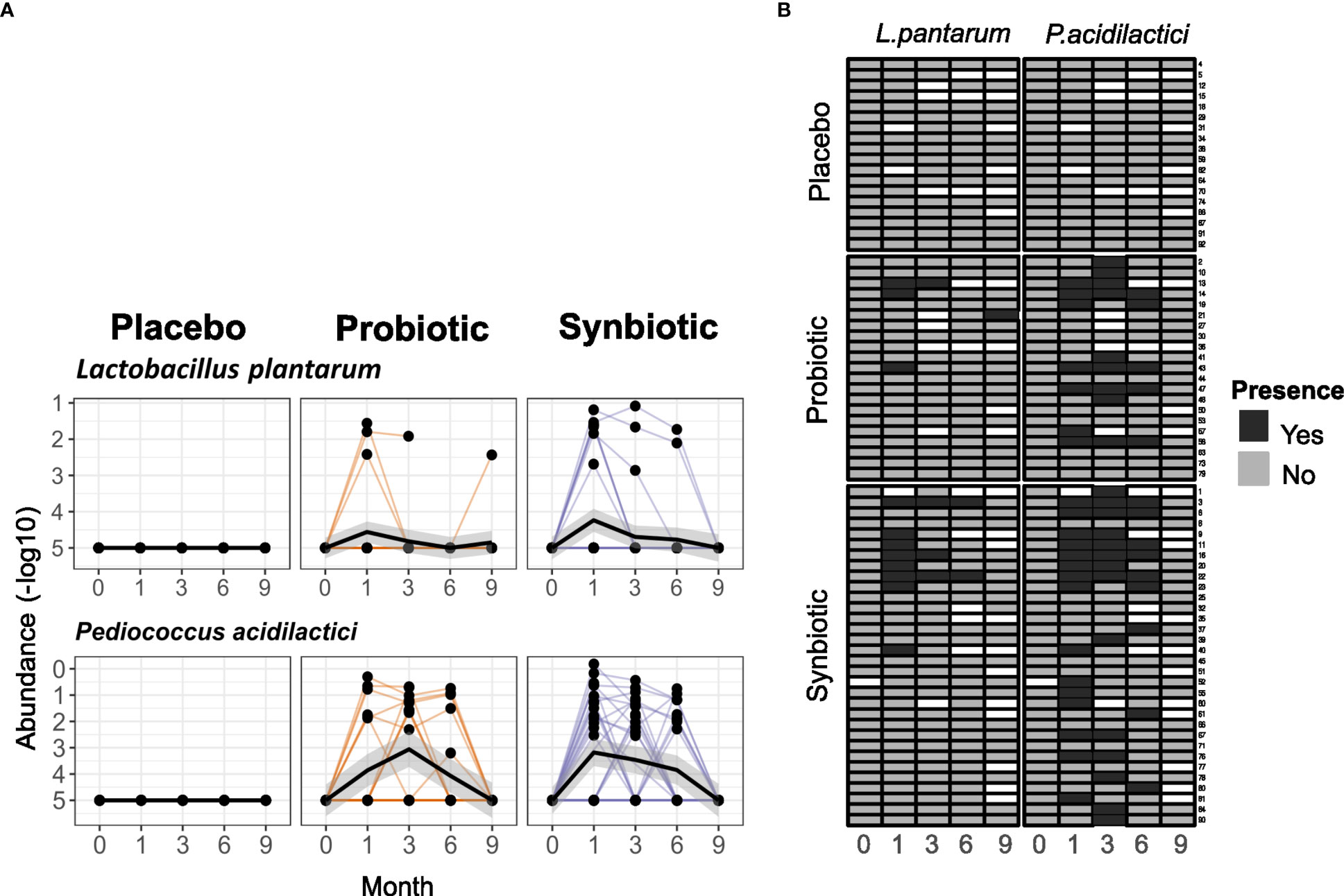
Figure 5 Detection of the probiotic strains in stool samples. (A) Relative abundances (in -log10) of P. acidilactici and L. plantarum in faeces for each of the intervention arms. (B) Heatmap representing presence/absence of each arm in each sample. Rows represent single patients and columns represent the month each sample belongs to. White cells represent lack of sample.
To better filter any potential probiotic strain-specific effect, as well as uncover potential factors that may affect probiotic engraftment, a new variable was defined. Two groups were created that separated patients with or without detectable levels of P. acidilactici in at least 2 stool samples belonging to different time points, excluding those patients in the Placebo group. P. acidilactici was chosen because it had presence in all samples where L. plantarum was detectable, but not the other way around (Figure 5B).
Microbiome analysis under this new stratification found no significant changes in alpha diversity as gene richness (Figure 6A) or beta diversity as Bray-Curtis distance from baseline (Figure 6B) over time between groups with (Present) and without (Absent) probiotic detection. This new variable only explained a 3.6% of the variance of Bray-Curtis distances between samples (Adonis R2 = 0.0362, p = 0.001). No clear group clustering could be observed under this new definition (Figure 6C)
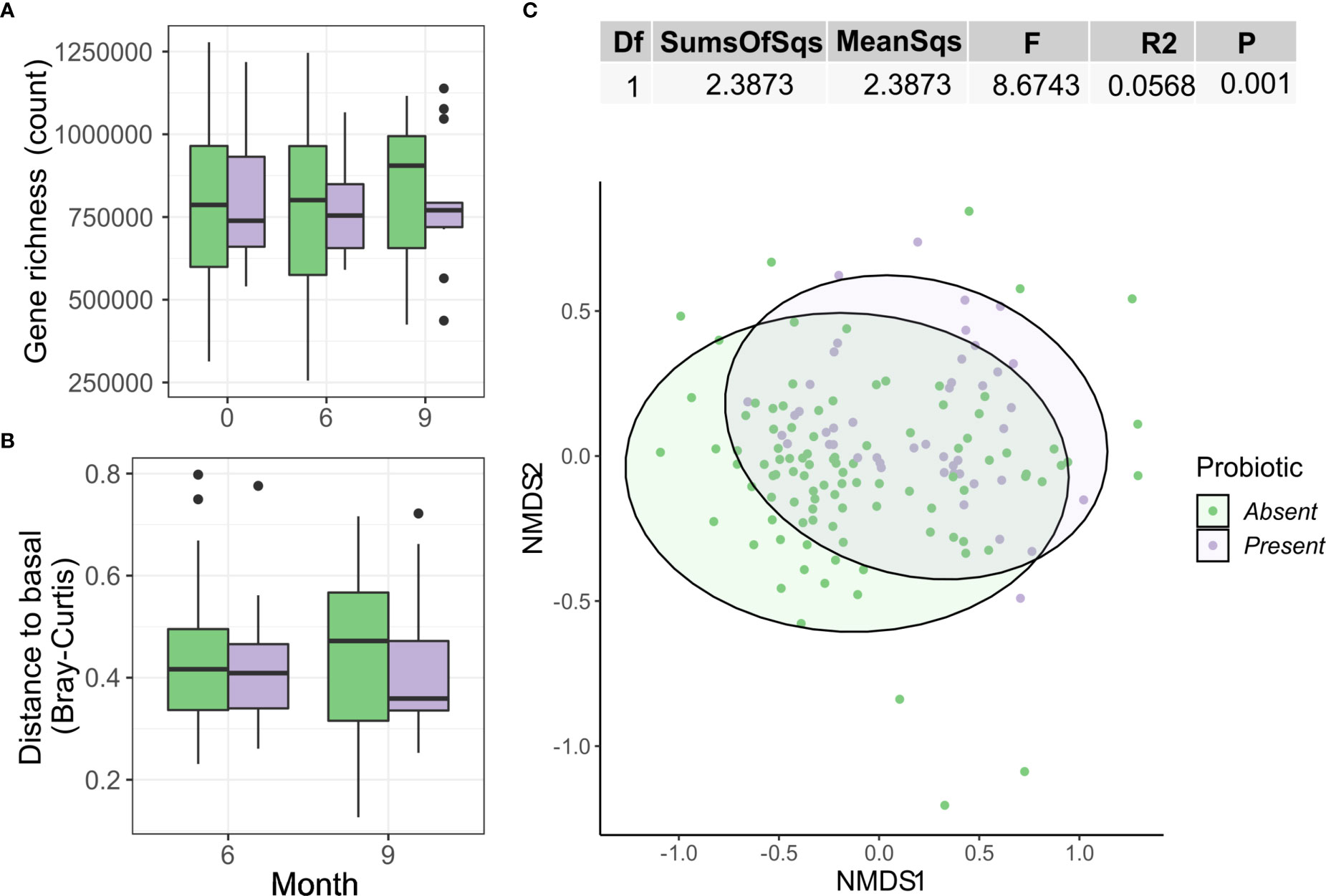
Figure 6 Differences by presence or absence of the probiotic strains on the gut microbiome. (A) Gene richness counts for each group arm and month. Comparisons between group at each timepoint were used simple Wilcoxon test, while the paired version was used for within-group comparisons between months. (B) Distribution of Bray Curtis-Distances from baseline for each month and group. (C) NMDS showing sample ordination according to Bray-Curtis distance. Group distance dissimilarity was tested using ADONIS.
At month 6, the Present group was associated with a significant decrease of CRP (median=11.1 (IQR=22) vs 19.2 (66), median change= -2.7 (13.2) ug/ml, p = 0.04), while the Absent group related to an increase in CD4/CD8 ratio (0.72 (0.57) vs 0.56 (0.4), median change = 0.4 (0.2) p=0.015) vs baseline. IL-6 differed between both groups at month 6, being lower in Present than Absent group (0.58 (1.13) vs 1.17 (1.59) ug/ml, p = 0.02). At month 9, an increase vs baseline was observed for CD4/CD8 ratio (0.67 (0.5) vs 0.59 ± (0.49), median change = 0.04 (0.18), p = 0.02), and CD4+ counts (457 (153) vs 416 (142), median change = 45 (75), counts/ml, p = 0.005) in Absent but not in Present group (Figure 7).
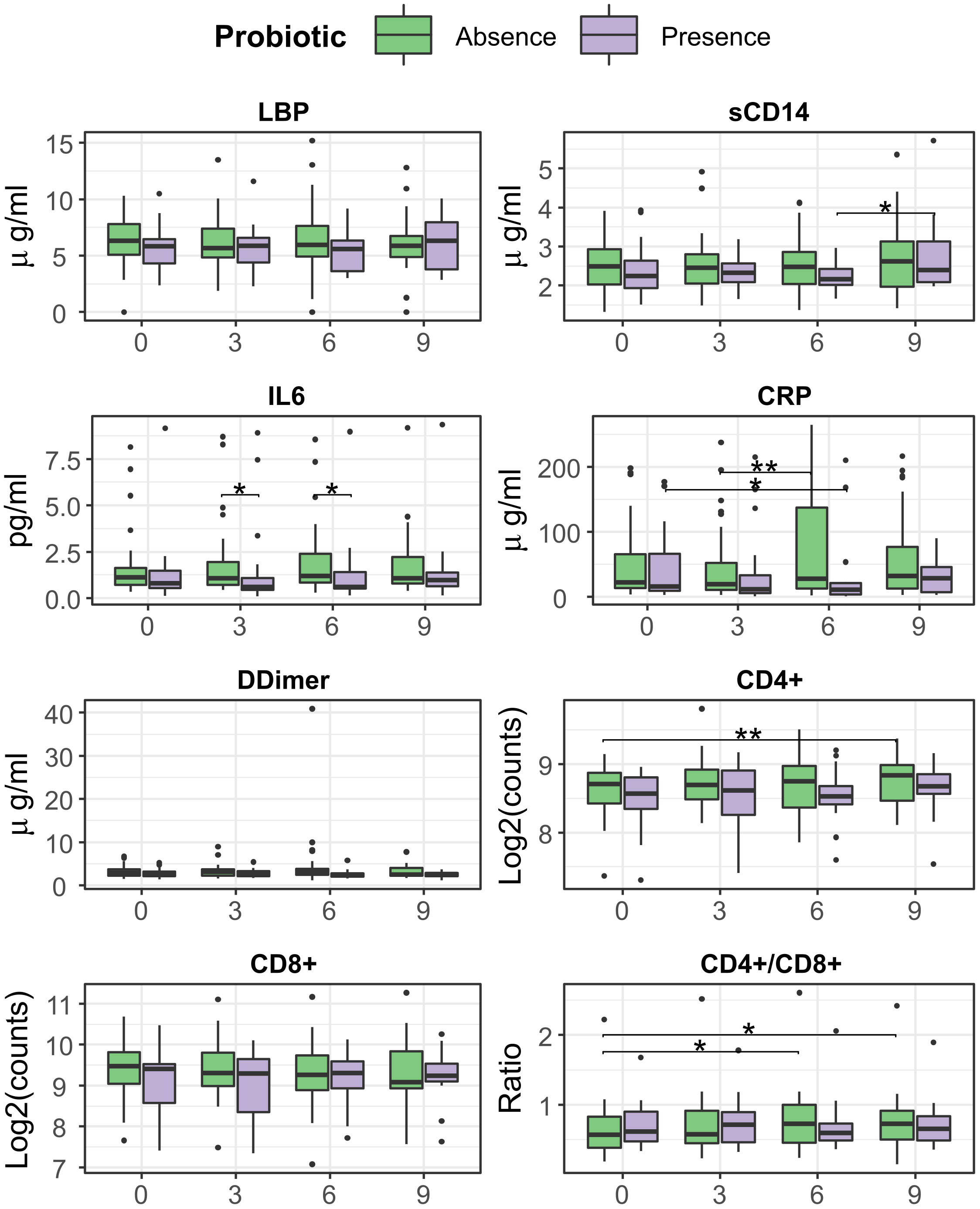
Figure 7 Evolution of T-cell counts, ratio, inflammation, and bacterial translocation markers between months 0 and 6, separated presence/absence of probiotic strains. Comparisons within group and between time points were performed by Wilcoxon test, in its paired form for longitudinal differences and unpaired for cross-group comparisons. Significance was coded as follows: *(p < 0.05), **(p < 0.01).
Analysis with LMM found little overall effect on immune status and inflammation, with a slight increase of CD4/CD8 ratio (p = 0.008, slope = 0.012) and D-Dimer (p = 0.047, slope = 0.236) in the Absent group (Supplementary Figure 3A). However, these trends changed when stratifying by treatment group, especially for CRP, which increased in the Synbiotic arm (p = 0.013, slope = 7.4) (Supplementary Figure 3C). Also, CD4/CD8 ratio increase was specific to the subjects in Absent group who were in the Synbiotic arm (p = 0.005, slope = 0.015).
Sulphate assimilation pathways seemed slightly reduced in the probiotic Present vs Absent group, but the differences were statistically non-significant (Figure 8). Analysis with LMMs found no significant trends for any of the three previously studied pathways neither in the Present nor in the Absent group. Interestingly, after stratifying by intervention, pathway relative abundances did not change over time in the Present patients within the Probiotic group but increased in the Absent group (Met: p = 0.026, slope = 2.13·10-6; ASR: p=0.024, slope = 1.11·10-6; Cys: p = 0.021, slope = 1.256·10-6), while the Synbiotic group showed a relative abundance decline in both Present and Absent groups (Supplementary Figure 5). Fixed effect interaction tests on unified models found no significant effect of presence/absence in any of the pathways, with and without stratification (Supplementary Figure 6).
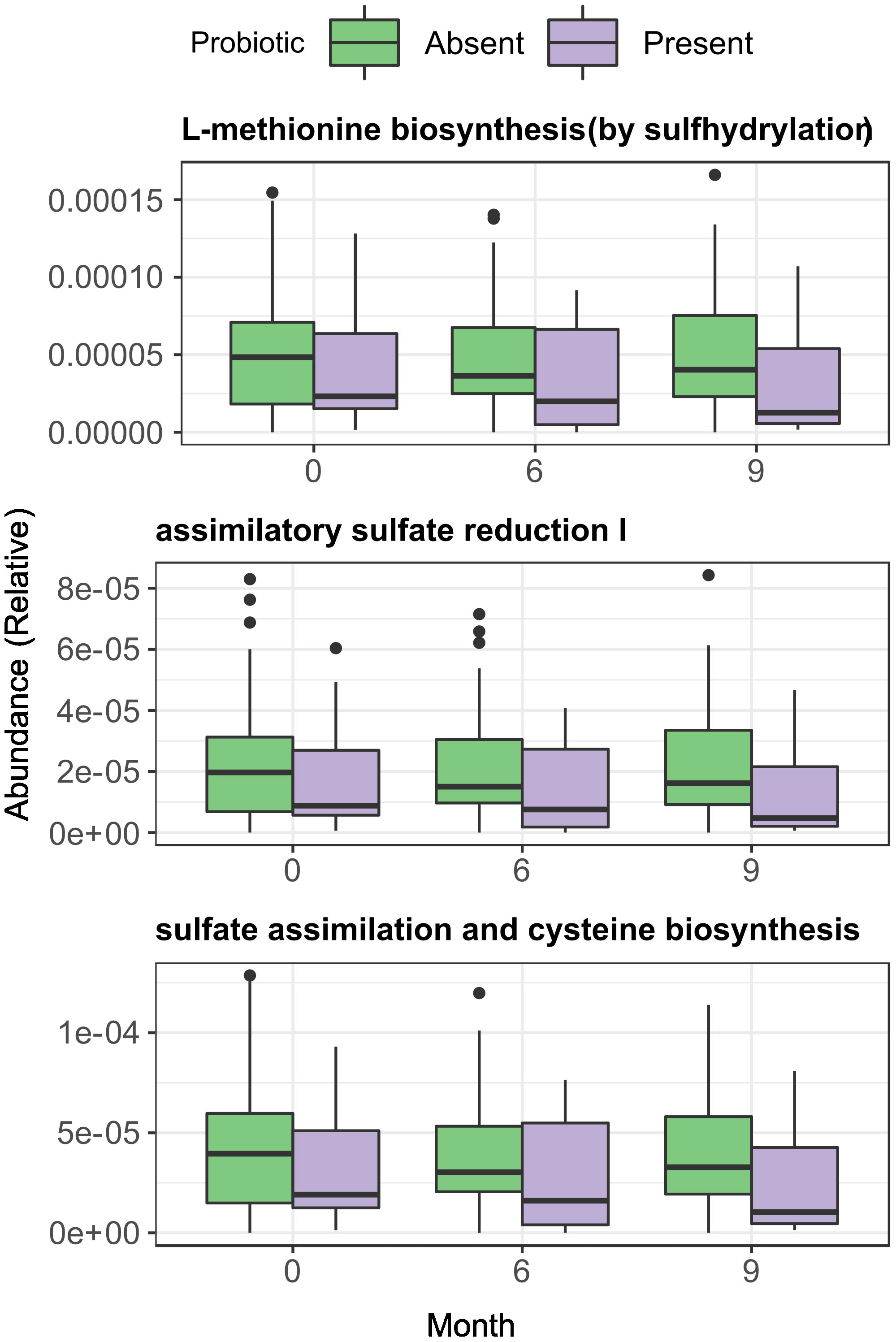
Figure 8 Longitudinal and across-group comparisons of Assimilatory Sulphate Reduction and sulphur amino acid pathways across intervention groups. Comparisons were performed across between groups by Wilcoxon test and longitudinally within-group with paired Wilcoxon test.
We assessed the efficacy and safety of long-term probiotic supplementation on immunodiscordant (CD4+ counts < 500) patients with supressed HIV after long, stable cART in a double-blind, randomized trial. We found that Synbiotic intervention with strains of L. plantarum and P. acidilactici along with prebiotic fibers is safe and associated with slight increases of CD4+ counts, CD4/CD8 ratio, and a decrease of the gut leakiness, as measured with sCD14 which is a proxy for bacterial endotoxin entering the bloodstream. However, the clinical significance of such improvements is uncertain, especially as they manifested 3 months after the intervention stopped. A more in-depth analysis suggests that the presence of the probiotic strains in stool was associated with lower levels of proinflammatory cytokines (IL-6 and CRP), while improvements in CD4/CD8 ratio appeared to be linked to the prebiotic fibers. The lowering in proinflammatory cytokines seems in agreement with previous reports using the same probiotic composition in animal models of IBD (37). This could point to a trade-off between immune activation and modulation by the prebiotic and probiotic, respectively. Of note, the reduction in inflammation markers happened during the intervention period, when no changes in gut permeability markers could be observed, suggesting that the immunomodulatory activity of the probiotics may be independent of the gut barrier status. Some gut barrier-unrelated anti-inflammatory mechanisms have been described, as Kawashima, Tadaomi et al. (38) found, lactic acid bacteria can induce IgA secretion to the gut lumen, which is a known immunomodulatory agent (39, 40).
The health-promoting properties of lactic acid bacteria have been widely reported in vivo and in vitro (41, 42). Nevertheless, the actual implantation and ecological viability of such strains is a multidimensional problem where host related factors and inherent ecological features of the host microbiome (such as normal ecological succession across the intestine) (43). Within the gastrointestinal (GI) tract, some lactobacilli show a nomadic behaviour (44) and tend to stay most commonly on the upper GI hence they tend to be underrepresented and transient in stool samples (45). In turn, implantation itself is not a strict requirement to generate change in the gut ecosystem or the host’s physiology (46). In this study, P. acidilactici was more consistently found than Lactobacillus plantarum in faeces but, unlike the later, the former has been reported as having good adhesion to the lower GI and higher presence in stool (47). Noteworthy, such differences may be strain-specific and not generalizable to other formulations (43). Hence, assessing the effectiveness of the probiotic strains in this study has been problematic, due to the low proportion of patients having a detectable presence of those, but also to the spatial and temporal complexity that characterizes the gut microbiome.
Additionally, we found that synbiotic intake was linked to decreases in the Assimilatory Sulphate Reduction (ASR) and sulphur-containing amino acids biosynthesis pathways. ASR, unlike the Dissimilatory Sulphate Reduction (DSR), takes sulphate without producing hydrogen sulphide as a final product, which has been shown to impair butyrate oxidation, the primary source of energy of enterocytes, and has been linked to gut inflammation and Ulcerative Colitis (48, 49). DSR is exclusive of anaerobic bacteria that undergo sulphate respiration, while ASR is more ubiquitous (50). Intake of Fructose and Glucose Oligosaccharides (FOS and GOS respectively) has been extensively used to modulate the microbiome, and has been linked to metabolic changes, especially of short chain fatty acids (51), but little is known about their relationship with sulphate metabolism. The fact that the Synbiotic group showed a consistent decline in such pathways could point to metabolic modulation and a population shift by the prebiotic fibers in the small intestine, although such changes may have been partially represented in the faecal samples.
Previous studies also found a lack of change in neither peripheral blood T-cells and/or gut permeability markers with probiotics only. Serrano-Villar et al. (52) found a significant decrease in inflammation markers (IL-6 and CRP) but no differential improvement in either circulatory T-cells nor sCD14 after 48 weeks, using a synbiotic formulation of Saccharomyces boulardii with various additives. However, this study was performed on late presenter, cART-naïve individuals, while our study was conducted on immunodiscordant patients with stable cART.
Previous studies using HIV infected immune discordant cohort reported diverse results. Presti et al. (53) tested a probiotic treatment consisting of different strains of streptococcus, bifidobacteria, and lactobacilli, without prebiotics, over 12 weeks, finding no differences in gut integrity (sCD14) and inflammation markers (D-Dimer, IP-10) but a significant decrease in Proteobacteria. Stiksrud et al. (54) found a decrease in inflammation (IL-6 and D-Dimer) but no differences in bacterial translocation markers and CD4+ counts after 8 weeks of intervention with formulation of skimmed milk, enriched with various species of Lactobacillus and Bifidobacterium vs a placebo of skimmed-milk only and a control groups. Geng et al. (55) reported an improvement in gut integrity (D-Dimer, DAO) and an enhanced CD4+ recovery in immune discordant patients using pre-digested protein supplementation. Despite the differences from the previously described studies (whether from study design or probiotic formulation), our study found improvement in inflammation and translocation markers but not an overall improvement in immune reconstitution. In addition, we could assess probiotic-related gut microbiome at species-level resolution of the microbiome, coupled with functional evidence and a washout period which adds robustness to any finding of potential signal of treatment effect.
Several limitations in this study should be considered. First, since patients were randomized by class of third antiretroviral drug, same-class ARV drgus might have different effects of the gut microbiome. Additionally, the cohort selected for this study was composed by immunodiscordant individuals, whose low CD4+ counts after viral suppression made them prone to infections that required antibiotic treatment, causing many dropouts. However, no significant biases were created by dropouts among the groups, although final sample size (n=71) was clearly smaller than original target (n=100) and may have been underpowered to detect some effects. Importantly, faecal samples hold an inherent bias and may not be representative of the actual gut microbiome composition, especially from the upper GI. Also, diet and concomitant treatments may affect faecal microbiome composition. While we found low rates of concomitant medications and these were balanced among groups, dietary information was not available, although extreme diets were excluded. This may affect our capability to detect any correlation between clinical outcomes and microbiome features. Finally, the fact that treatment intake was self-reported, could have led to many patients not taking the treatment but reporting otherwise during the 6 months follow-up, thus overestimating the actual intake of the probiotics. All these shortcomings add up to the limited effect of probiotics in immune reconstitution and affect the capability to translate research results from into clinical practice and warrant further research.
A synbiotic intervention with L. plantarum and P. acidilactici was safe and well tolerated. Synbiotic intervention led to small increases in CD4/CD8 ratio and minor reductions in sCD14 after 6 months and continued 3 months after discontinuing the intervention, but such changes are of uncertain clinical significance. A probiotic with the same composition but without prebiotics was also safe but did not achieve any impact on immune parameters or faecal microbiome composition.
The data presented in the study are deposited in the EBI-ENA repository, accession number PRJEB56022.
The study was reviewed and approved by the Institutional Review Board of the Hospital Universitari Germans Trias i Pujol (reference PI-13-046). All participants provided written informed consent in accordance with the World Medical Association Declaration of Helsinki, Fortaleza and Brazil, October 2013 and personal data was managed according to Spanish data protection law (LOPD 15/1999). The study concept, design, patient information and results were discussed with the FLSida Community Advisory Committee, in accordance with AB-Biotics internal QC auditing. All available information can be found in the protocol (Supplementary Methods), and the study is registered in clinicaltrials.gov, accession: NCT03542786. The patients/participants provided their written informed consent to participate in this study.
RP, JE-M, JRS, and MN-J conceived and designed the study. RP and JRS recruited the study participants and performed their clinical evaluations. MA performed study monitoring. MP, AE-T, and MC performed sample processing for sequencing and inflammation markers experiments, under the supervision of MN-J and RP. CB and FC-M performed the bioinformatic and statistical analyses with the supervision of RP, MN-J, CB, RP, and MN-J wrote the paper, which was reviewed, edited, and approved by all authors.
We thank all the enrolled participants who contributed to this study, the staff from Fundació Lluita contra la Sida (FLS), IrsiCaixa and Hospital Germans Trias I Pujol. We also thank AB-Biotics for their non-financial support and feedback, and the staff from MISTRAL.
This project has received funding from the European Union’s Horizon 2020 Research and Innovation Programme under Grant Agreement N° 847943, and the Spanish Ministry of Economy and Competitiveness (MINECO), reference RTC-2016-5223-1. This research was supported by CIBER -Consorcio Centro de Investigación Biomédica en Red- (CB 2021), Instituto de Salud Carlos III, Ministerio de Ciencia e Innovación and Unión Europea – NextGenerationEU.
JE-M and MA are full-time employees of AB-BIOTICS SA, the company owning IP rights for the probiotic strains used in this study. MN-J is co-founder and shareholder of Nano1Health SL, outside the scope of the submitted work.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.1066036/full#supplementary-material
1. Doitsh G, Galloway NLK, Geng X, Yang Z, Monroe KM, Zepeda O, et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature (2014) 505:509–14. doi: 10.1038/nature12940
2. Pantazis N, Papastamopoulos V, Paparizos V, Metallidis S, Adamis G, Antoniadou A, et al. Long-term evolution of CD4+ cell count in patients under combined antiretroviral therapy. AIDS (2019) 33:1645–55. doi: 10.1097/QAD.0000000000002248
3. Deeks SG, Overbaugh J, Phillips A, Buchbinder S. HIV Infection. Nat Rev Dis Prim (2015) 1:1–22. doi: 10.1038/nrdp.2015.35
4. Chomont N, El-Far M, Ancuta P, Trautmann L, Procopio FA, Yassine-Diab B, et al. HIV Reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med (2009) 15:893–900. doi: 10.1038/nm.1972
5. Kuo HH, Lichterfeld M. Recent progress in understanding HIV reservoirs. Curr Opin HIV AIDS (2018) 13:137–42. doi: 10.1097/COH.0000000000000441
6. Collaboration TATC. Life expectancy of individuals on combination antiretroviral therapy in high-income countries: A collaborative analysis of 14 cohort studies. Lancet (2008) 372:293–9. doi: 10.1016/S0140-6736(08)61113-7
7. Gazzola L, Tincati C, Bellistrì GM, Monforte ADA, Marchetti G. The absence of CD4+ T cell count recovery despite receipt of virologically suppressive highly active antiretroviral therapy: Clinical risk, immunological gaps, and therapeutic options. Clin Infect Dis (2009) 48:328–37. doi: 10.1086/595851
8. Benveniste O, Flahault A, Rollot F, Elbim C, Estaquier J, Pédron B, et al. Mechanisms involved in the low-level regeneration of CD4+ cells in HIV-1-infected patients receiving highly active antiretroviral therapy who have prolonged undetectable plasma viral loads. J Infect Dis (2005) 191:1670–9. doi: 10.1086/429670
9. Negredo E, Massanella M, Puig J, Pérez-Álvarez N, Gallego-Escuredo JM, Villarroya J, et al. Nadir CD4 T cell count as predictor and high CD4 T cell intrinsic apoptosis as final mechanism of poor CD4 T cell recovery in virologically suppressed HIV-infected patients: Clinical implications. Clin Infect Dis (2010) 50:1300–8. doi: 10.1086/651689
10. D’Angelo C, Reale M, Costantini E. Microbiota and probiotics in health and HIV infection. Nutrients (2017) 9(6):615. doi: 10.3390/nu9060615
11. Kanwar B, Favre D, McCune JM. Th17 and regulatory T cells: Implications for AIDS pathogenesis. Curr Opin HIV AIDS (2010) 5:151–7. doi: 10.1097/COH.0b013e328335c0c1
12. Hartigan-O’Connor DJ, Hirao LA, McCune JM, Dandekar S. Th17 cells and regulatory T cells in elite control over HIV and SIV. Curr Opin HIV AIDS (2011) 6:221–7. doi: 10.1097/COH.0b013e32834577b3
13. Nilsson J, Boasso A, Velilla PA, Zhang R, Vaccari M, Franchini G, et al. HIV-1-driven regulatory T-cell accumulation in lymphoid tissues is associated with disease progression in HIV/AIDS. Blood (2006) 108:3808–17. doi: 10.1182/blood-2006-05-021576
14. Kinugasa T, Sakaguchi T, Gu X, Reinecker HC. Claudins regulate the intestinal barrier in response to immune mediators. Gastroenterology (2000) 118:1001–11. doi: 10.1016/S0016-5085(00)70351-9
15. Salzman NH, Hung K, Haribhai D, Chu H, Karlsson-Sjöberg J, Amir E, et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nat Immunol (2010) 11:76–83. doi: 10.1038/ni.1825
16. Marchetti G, Bellistrì GM, Borghi E, Tincati C, Ferramosca S, La Francesca M, et al. Microbial translocation is associated with sustained failure in CD4+ T-cell reconstitution in HIV-infected patients on long-term highly active antiretroviral therapy. AIDS (2008) 22:2035–8. doi: 10.1097/QAD.0b013e3283112d29
17. Vujkovic-Cvijin I, Swainson LA, Chu SN, Ortiz AM, Santee CA, Petriello A, et al. Gut-resident lactobacillus abundance associates with IDO1 inhibition and Th17 dynamics in SIV-infected macaques. Cell Rep (2015) 13:1589–97. doi: 10.1016/j.celrep.2015.10.026
18. Vallejo AN, Weyand CM, Goronzy JJ. T-Cell senescence: A culprit of immune abnormalities in chronic inflammation and persistent infection. Trends Mol Med (2004) 10:119–24. doi: 10.1016/j.molmed.2004.01.002
19. Blaschitz C, Raffatellu M. Th17 cytokines and the gut mucosal barrier. J Clin Immunol (2010) 30:196–203. doi: 10.1007/s10875-010-9368-7
20. Vujkovic-Cvijin I, Dunham RM, Iwai S, Maher MC, Albright RG, Broadhurst MJ, et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci Transl Med (2013) 5:193ra91–193ra91. doi: 10.1126/scitranslmed.3006438
21. Jenabian M-A, El-Far M, Vyboh K, Kema I, Costiniuk CT, Thomas R, et al. Immunosuppressive tryptophan catabolism and gut mucosal dysfunction following early HIV infection. J Infect Dis (2015) 212:355–66. doi: 10.1093/infdis/jiv037
22. Sheil B, Shanahan F, O’Mahony L. Probiotic effects on inflammatory bowel disease. J Nutr (2007) 137(3 Suppl 2):819S–24S. doi: 10.1093/jn/137.3.819S
23. Zhang XL, Chen MH, Geng ST, Yu J, Kuang YQ, Luo HY, et al. Effects of probiotics on diarrhea and CD4 cell count in people living with HIV: A systematic review and meta-analysis. Front Pharmacol (2021) 12:570520/BIBTEX. doi: 10.3389/FPHAR.2021.570520/BIBTEX
24. Monachese M, Cunningham-Rundles S, Diaz MA, Guerrant R, Hummelen R, Kemperman R, et al. Probiotics and prebiotics to combat enteric infections and HIV in the developing world: A consensus report. Gut Microbes (2011) 2(3):198–207. doi: 10.4161/GMIC.2.3.16106
25. Wang Y, Li X, Ge T, Xiao Y, Liao Y, Cui Y, et al. Probiotics for prevention and treatment of respiratory tract infections in children: A systematic review and meta-analysis of randomized controlled trials. Med (United States) (2016) 95(31):e4509. doi: 10.1097/MD.0000000000004509
26. Meyer-Myklestad MH, Medhus AW, Lorvik KB, Seljeflot I, Hansen SH, Holm K, et al. Human immunodeficiency virus–infected immunological nonresponders have colon-restricted gut mucosal immune dysfunction. J Infect Dis (2022) 225:661. doi: 10.1093/INFDIS/JIAA714
27. Lu W, Feng Y, Jing F, Han Y, Lyu N, Liu F, et al. Association between gut microbiota and CD4 recovery in HIV-1 infected patients. Front Microbiol (2018) 9:1451/FULL. doi: 10.3389/FMICB.2018.01451/FULL
28. Lee SC, Chua LL, Yap SH, Khang TF, Leng CY, Raja Azwa RI, et al. Enrichment of gut-derived fusobacterium is associated with suboptimal immune recovery in HIV-infected individuals. Sci Rep (2018) 8(1):14277. doi: 10.1038/s41598-018-32585-x
29. Zeuthen LH, Fink LN, Frokiaer H. Epithelial cells prime the immune response to an array of gut-derived commensals towards a tolerogenic phenotype through distinct actions of thymic stromal lymphopoietin and transforming growth factor-β. Immunology (2008) 123:197–208. doi: 10.1111/j.1365-2567.2007.02687.x
30. Hummelen R, Changalucha J, Butamanya NL, Koyama TE, Habbema JDF, Reid G. Efect of 25 weeks probiotic supplementation on immune function of HIV patients. Gut Microbes (2011) 2:80–5. doi: 10.4161/gmic.2.2.15787
31. Rawi MH, Zaman SA, Pa’ee KF, Leong SS, Sarbini SR. Prebiotics metabolism by gut-isolated probiotics. J Food Sci Technol (2020) 57:2786. doi: 10.1007/S13197-020-04244-5
32. Bolger AM, Lohse M, Usadel B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics (2014) 30:2114–20. doi: 10.1093/bioinformatics/btu170
33. Beghini F, McIver LJ, Blanco-Míguez A, Dubois L, Asnicar F, Maharjan S, et al. Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery 3. Elife (2021) 10:e65088. doi: 10.7554/eLife.65088
34. Li J, Jia H, Cai X, Zhong H, Feng Q, Sunagawa S, et al. An integrated catalog of reference genes in the human gut microbiome. Nat Biotechnol (2014) 32:834–41. doi: 10.1038/NBT.2942
35. Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol (2009) 10:R25. doi: 10.1186/gb-2009-10-3-r25
36. Oksanen J, Simpson GL, Blanchet FG, Kindt R, Legendre P, Minchin PR, et al. Vegan: Community ecology package. R package version 2.6-4. In: Vegan community ecol (2012). Available at: https://cran.r-project.org/package=vegan.
37. Lorén V, Manyé J, Fuentes MC, Cabré E, Ojanguren I, Espadaler J. Comparative effect of the I3.1 probiotic formula in two animal models of colitis. Probiotics Antimicrob Proteins (2017) 9:71–80. doi: 10.1007/S12602-016-9239-5
38. Kawashima T, Ikari N, Kouchi T, Kowatari Y, Kubota Y, Shimojo N, et al. The molecular mechanism for activating IgA production by pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci Rep (2018) 8(1):5065. doi: 10.1038/S41598-018-23404-4
39. Guo Y, Wang B, Wang T, Gao L, Yang ZJ, Wang FF, et al. Biological characteristics of IL-6 and related intestinal diseases. Int J Biol Sci (2021) 17:204. doi: 10.7150/IJBS.51362
40. Wang Y, Liu L, Moore DJ, Shen X, Peek RM, Acra SA, et al. An LGG-derived protein promotes IgA production through upregulation of APRIL expression in intestinal epithelial cells. Mucosal Immunol (2017) 10:373–84. doi: 10.1038/MI.2016.57
41. Smits HH, Engering A, van der Kleij D, De Jong EC, Schipper K, Van Capel TMM, et al. Selective probiotic bacteria induce IL-10-producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin. J Allergy Clin Immunol (2005) 115:1260–7. doi: 10.1016/J.JACI.2005.03.036
42. Kim HG, Lee SY, Kim NR, Lee HY, Ko MY, Jung BJ, et al. Lactobacillus plantarum lipoteichoic acid down-regulated shigella flexneri peptidoglycan-induced inflammation. Mol Immunol (2011) 48:382–91. doi: 10.1016/J.MOLIMM.2010.07.011
43. Walter J. Ecological role of lactobacilli in the gastrointestinal tract: Implications for fundamental and biomedical research. Appl Environ Microbiol (2008) 74:4985. doi: 10.1128/AEM.00753-08
44. Martino ME, Bayjanov JR, Caffrey BE, Wels M, Joncour P, Hughes S, et al. Nomadic lifestyle of lactobacillus plantarum revealed by comparative genomics of 54 strains isolated from different habitats. Environ Microbiol (2016) 18:4974–89. doi: 10.1111/1462-2920.13455
45. Zmora N, Zilberman-Schapira G, Suez J, Mor U, Dori-Bachash M, Bashiardes S, et al. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell (2018) 174:1388–1405.e21. doi: 10.1016/J.CELL.2018.08.041/ATTACHMENT/39F12DBE-F56A-4CD7-9089-962332F15476/MMC7.PDF
46. Wieërs G, Belkhir L, Enaud R, Leclercq S, Philippart de Foy JM, Dequenne I, et al. How probiotics affect the microbiota. Front Cell Infect Microbiol (2020) 9:454. doi: 10.3389/FCIMB.2019.00454
47. Balgir PP, Kaur B, Kaur T, Daroch N, Kaur G. In vitro and In vivo survival and colonic adhesion of pediococcus acidilactici MTCC5101 in human gut. BioMed Res Int (2013) 2013:583850. doi: 10.1155/2013/583850
48. Jørgensen J, Mortensen PB. Hydrogen sulfide and colonic epithelial metabolism: implications for ulcerative colitis. Dig Dis Sci (2001) 46:1722–32. doi: 10.1023/A:1010661706385
49. Figliuolo VR, dos Santos LM, Abalo A, Nanini H, Santos A, Brittes NM, et al. Sulfate-reducing bacteria stimulate gut immune responses and contribute to inflammation in experimental colitis. Life Sci (2017) 189:29–38. doi: 10.1016/J.LFS.2017.09.014
50. Kushkevych I, Cejnar J, Treml J, Dordević D, Kollar P, Vítězová M. Recent advances in metabolic pathways of sulfate reduction in intestinal bacteria. Cells (2020) 9(3):698. doi: 10.3390/CELLS9030698
51. Serrano-Villar S, Vázquez-Castellanos JF, Vallejo A, Latorre A, Sainz T, Ferrando-Martínez S, et al. The effects of prebiotics on microbial dysbiosis, butyrate production and immunity in HIV-infected subjects. Mucosal Immunol (2016) 10(5):1279–93. doi: 10.1038/mi.2016.122
52. Serrano-Villar S, De Lagarde M, Vázquez-Castellanos J, Vallejo A, Bernadino JI, Madrid N, et al. Effects of immunonutrition in advanced human immunodeficiency virus disease: A randomized placebo-controlled clinical trial (Promaltia study). Clin Infect Dis (2019) 68(1):120–30. doi: 10.1093/cid/ciy414
53. Presti RM, Yeh E, Williams B, Landay A, Jacobson JM, Wilson C, et al. A randomized, placebo-controlled trial assessing the effect of VISBIOME ES probiotic in people with HIV on antiretroviral therapy. Open Forum Infect Dis (2021) 8(12):ofab550. doi: 10.1093/OFID/OFAB550
54. Stiksrud B, Nowak P, Nwosu FC, Kvale D, Thalme A, Sonnerborg A, et al. Reduced levels of d-dimer and changes in gut microbiota composition after probiotic intervention in HIV-infected individuals on stable ART. J Acquir Immune Defic Syndr (2015) 70:329–37. doi: 10.1097/QAI.0000000000000784
55. Geng ST, Zhang JB, Wang YX, Xu Y, Lu D, Zhang Z, et al. Pre-digested protein enteral nutritional supplementation enhances recovery of CD4+ T cells and repair of intestinal barrier in HIV-infected immunological non-responders. Front Immunol (2021) 12:757935/FULL. doi: 10.3389/FIMMU.2021.757935/FULL
Keywords: probiotics, prebiotics, synbiotics, HIV, immune reconstitution
Citation: Blázquez-Bondia C, Parera M, Català-Moll F, Casadellà M, Elizalde-Torrent A, Aguiló M, Espadaler-Mazo J, Santos JR, Paredes R and Noguera-Julian M (2022) Probiotic effects on immunity and microbiome in HIV-1 discordant patients. Front. Immunol. 13:1066036. doi: 10.3389/fimmu.2022.1066036
Received: 10 October 2022; Accepted: 17 November 2022;
Published: 08 December 2022.
Edited by:
Marius Trøseid, Oslo University Hospital, NorwayReviewed by:
Piotr Nowak, Karolinska Institutet (KI), SwedenCopyright © 2022 Blázquez-Bondia, Parera, Català-Moll, Casadellà, Elizalde-Torrent, Aguiló, Espadaler-Mazo, Santos, Paredes and Noguera-Julian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marc Noguera-Julian, bW5vZ3VlcmFAaXJzaWNhaXhhLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.