 David G. Hernández-Mejía
David G. Hernández-Mejía Iván Aurelio Páez-Gutiérrez
Iván Aurelio Páez-Gutiérrez Valerie Dorsant Ardón
Valerie Dorsant Ardón Nathalie Camacho RamírezMelissa Mosquera
Nathalie Camacho RamírezMelissa Mosquera Paola Andrea Cendales*
Paola Andrea Cendales* Bernardo Armando Camacho
Bernardo Armando Camacho- Instituto Distrital de Ciencia, Biotecnología e Innovación en Salud - IDCBIS, Bogotá, Colombia
The HLA compatibility continues to be the main limitation when finding compatible donors, especially if an identical match is not found within the patient’s family group. The creation of bone marrow registries allowed a therapeutic option by identifying 10/10 compatible unrelated donors (URD). However, the availability and frequency of haplotypes and HLA alleles are different among ethnic groups and geographical areas, increasing the difficulty of finding identical matches in international registries. In this study, the HLA-A, -B, -C, -DRB1, and -DQB1 loci of 1763 donors registered in the Colombian Bone Marrow Registry were typed by next-generation sequencing. A total of 52 HLA-A, 111 HLA-B, 41 HLA-C, 47 HLA-DRB1, and 20 HLA-DQB1 alleles were identified. The 3 most frequent alleles for each loci were A*24:02g (20,8%), A*02:01g (16,1%), A*01:01g (7.06%); B*35:43g (7.69%), B*40:02g (7.18%), B*44:03g (6.07%); C*04:01g (15.40%), C*01:02g (10.49%), C*07:02g (10.44%); DRB1*04:07g (11.03%), DRB1*07:01g (9.78%), DRB1*08:02g (6.72%); DQB1*03:02g (20.96%), DQB1*03:01g (17.78%) and DQB1*02:01g (16.05%). A total of 497 HLA-A-C-B-DRB1-DQB1 haplotypes were observed with a frequency greater than or equal to 0.05% (> 0.05%); the haplotypes with the highest frequency were A*24:02g~B*35:43g~C*01:02g~DQB1*03:02g~DRB1*04:07g (3.34%), A*29:02g~B*44:03g~C*16:01g~DQB1*02:01g~DRB1*07:01g (2.04%), and A*01:01g~B*08:01g~C*07:01g~DQB1*02:01g~DRB1*03:01g (1.83%). This data will allow the new Colombian Bone Marrow Donor Registry to assess the genetic heterogeneity of the Colombian population and serve as a tool of interest for future searches of unrelated donors in the country.
1 Introduction
Hematopoietic stem cell transplantation continues to be one of the most widely used treatments for both malignant and benign hematological diseases (1–4). In recent decades, unrelated donors (URD) have been the first option for transplantation when an HLA-compatible family donor is not available. In countries with a National Bone Marrow (BM) Donors Registry, finding an 8/8 or 10/10 compatible URD can be reduced by several months, having a positive effect on the treatment and quality of life of patients (5–13). In the absence of a National BM Donors Registry, searching for compatible URD can take a longer time, leading medical treatment toward alternative sources of hematopoietic stem cells such as haploidentical transplant or umbilical cord blood (UCB) units, which are associated with higher risks of graft-versus-host disease (GVHD), graft failure or transplant-related death (TRM) (14–18). Since 2018, the Instituto Distrital de Ciencia, Biotecnología e Innovación en Salud (IDCBIS) has been promoting the creation of a National BM Donor Registry in Colombia to provide hematopoietic stem cells from URD to the country’s transplant centers, ultimately shortening the waiting time of patients requiring HSC transplants. Currently, 4435 donors have registered in the Colombia BM Donor Registry, and, by the time of this study 1779 were HLA typed.
The major histocompatibility complex (MHC), known in humans as the human leukocyte antigen (HLA) system, includes proteins responsible for mediating antigen presentation to CD8+ and CD4+ lymphocytes and thus activating the cellular and humoral immune responses in individuals. These genes are contained in approximately 4 MB located on the short arm of chromosome 6 (6p21). The MHC is composed of 3 subregions: HLA classes I, II, and III. Within class I are classical (HLA-A, -B and -C) and nonclassical (HLA-E, -F, -G and HFE) HLA genes. Likewise, within class II, classic genes such as HLA-DRB1, -DRB3/4/5, -DQB1, and -DPB1, and nonclassical genes (HLA-DM and HLA-DO). In between the HLA class III region is found, where there are genes participating in the immune response, such as complement System proteins and cytokines; however, class III genes differ in their structure and function compared to the genes class I and II. The HLA system has the highest density of genes in the human genome which are highly polymorphic. HLA alleles and haplotype frequencies vary widely between different populations, and even in the same territory may be conditioned by ethnicities and geographical regions (19–24).
Access to next-generation sequencing (NGS) technologies allowed the identification of a vast number of new HLA alleles, thus expanding the records in the IPD-IMGT/HLA database (25, 26), 36016 HLA alleles are recorded in the latest version (V 3.50) of the database released in October 2022, 25019 HLA class I alleles, and 10201 HLA class II alleles. HLA compatibility between the recipient and the donor is currently the main criterion for donor selection since numerous reports documented that a disparity in any of these genes leads to a worse transplant outcome and an increase in GVHD (15, 16, 27–33).
Colombia is in the extreme northeast of the South American continent, bordered by Panamá to the northeast, Perú and Ecuador to the south, Venezuela to the west, and Brazil to the southwest, in addition to having access to the Pacific Ocean on the eastern side and the Caribbean Sea to the north. This exceptional geographical position made Colombia a necessary passage between the pre-Columbian cultures that migrated from the south to the north of the American continent and vice versa (34, 35). Due to the colonization process in the fifteenth century by the Spanish empire, the first European population settlement occurred, as well as the first inhabitants of African origin arrived in the territory as a result of the slave trade. These Afro-American populations ended up settling mainly on the Pacific and Caribbean coasts of the country. In the territory, processes of miscegenation occurred over several generations between the European Caucasian, Native American, and African American populations, giving way to mestizo (Caucasian with Native American), mulatto (Caucasian with African American), and Zambo (Native American with African American) populations (36, 37).
Since the independence of the country in the 19th century, there have been continuous displacements of the population from rural areas to the urban centers of the country as a result of different civil wars, political violence in the mid-20th century, and in recent decades, the conflict between different armed groups (guerrillas, paramilitaries, and drug traffickers) (38, 39). Noteworthy during the 20th century occurs the migration of Turkish-Lebanese people who settled in the commercial ports of the Atlantic coast (40). The current population of Colombia is 50,372,424, concentrated in urban centers of the Andean region and the Caribbean coast, with the most populated cities being Bogotá (7,743,955 people), Medellín (2,533,424 people), Cali (2,252,616 people) and Barranquilla (1,274,250). Regarding their ethnic composition, 88.9% of the population is mestizo or non-Native American, 6.7% are Afro-descendant, 4.31% are Native American, 0.06% are Raizal, and 0.01% are Roma-Gypsy (41, 42).
This complex sociocultural, geographical and historical context in Colombia has produced a gene pool that currently has not been fully characterized. The objective of this study is to describe the allele frequencies and haplotypes of the classic HLA genes, using high-resolution NGS techniques, from donors registered in the Colombian BM Donor Registry; the information obtained will allow a better understanding of the distribution of HLA genes in different geographical regions of the country. Furthermore, the data obtained will be a resource for the national BM donor registry in Colombia to establish donor recruitment strategies.
2 Materials and methods
2.1 Population sample
This study included 1779 samples from donors enrolled from 2018 to 2021 in the Colombian BM donor registry. All donors signed an informed consent form approved by the Ethics Committee of the Secretaría Distrital de Salud de Bogotá. Additionally, each donor completed a form containing demographic data such as place of birth, ethnicity, age, and gender. The registration process was carried out in 4 of the largest cities of the country: 1329 in Bogotá, 150 in Medellín, 150 in Cali, and 150 in Barranquilla. 1749 of the recruited donors, were Colombian nationals (Figure 1; Supplementary Table 1), while the other 30 were foreigners (Supplementary Figure 1). Donors self-identify as mestizo (97.4%), Caucasian (1.3%), African American (0.9%), and Native American (0.4%). For the genotyping of the HLA-A, HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 loci, samples of blood stored in Protein Saver® cards, or buccal swabs were taken from 1347 women and 416 men, for a total of 1763 registries analyzed. 16 results were pending, 3 of them for confirmation of possible new HLA alleles. These 16 results were not included in the statistical analysis.
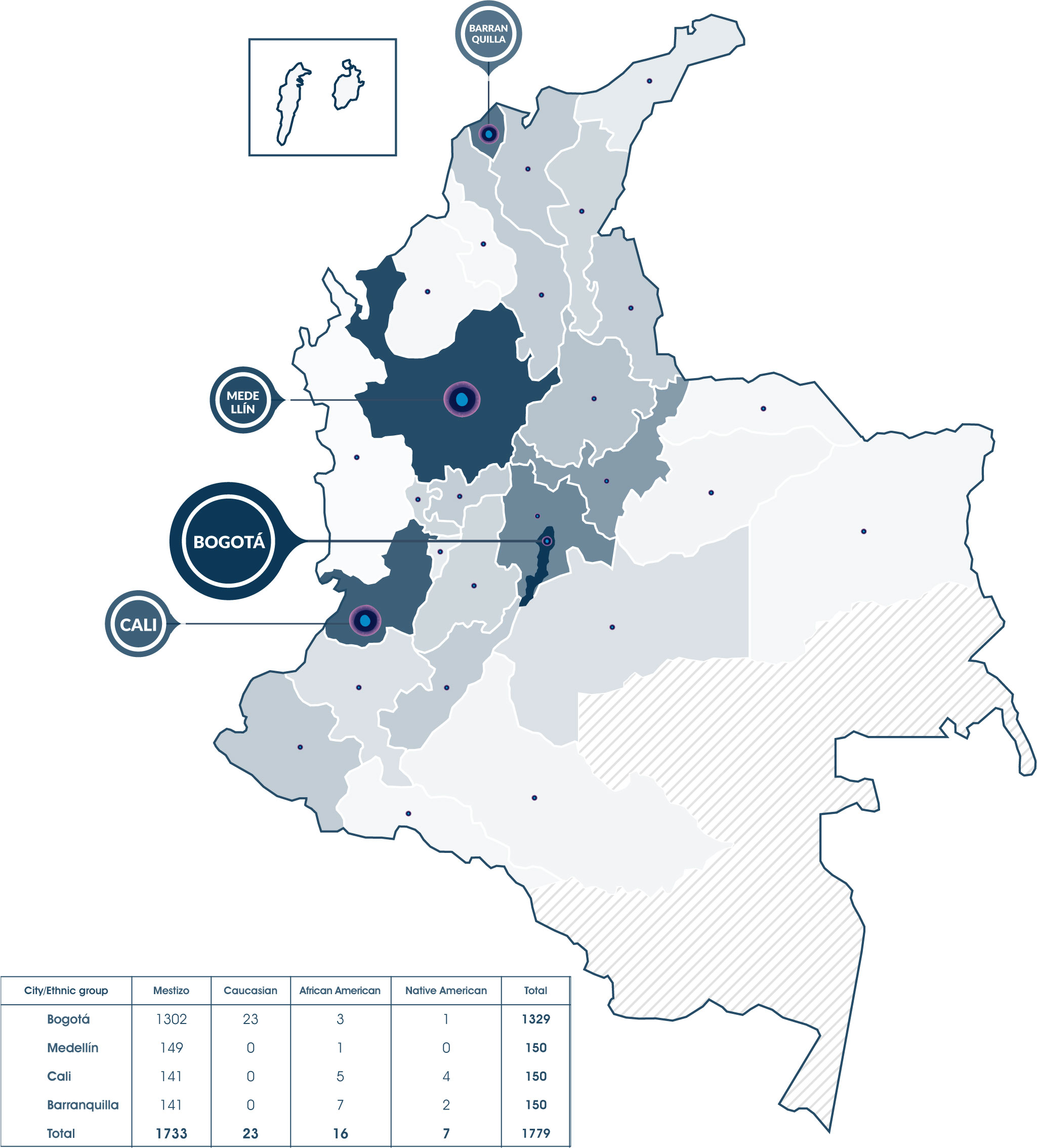
Figure 1 1779 donors were registered in the Colombian Bone Marrow Donor Registry, 1749 of them are Colombian and 30 are foreigners residing in the country, they all were recruited in 4 major cities: Bogotá, Cali, Medellín and Barranquilla. Currently, people from Bogotá and 28 of the 32 Colombian departments (except Amazonas, Vaupés, Guaviare, and Guainía) had registered.
2.2 HLA typing
Donors were typed at high resolution for the HLA-A, HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 loci. The blood samples were sent for sequencing to Histogenetics (Ossining, NY), and the buccal cell swabs were sent to DKMS Life Science Lab (Dresden, Germany). Both laboratories used Illumina platforms for sequencing exons 2 and 3 of HLA class I loci, and exon 2 of HLA class II loci, following the protocols standardized by each laboratory (43, 44). Typing results for each HLA allele were coded by G groups, with at least the first 2 fields being always resolved. New alleles identified at typing were confirmed and submitted by each laboratory to the IMGT/HLA database. Reports of these new alleles were subsequently published by the Colombian BM Donor Registry.
To corroborate the homozygous typing results, these labs use multiple sequencing technologies and platforms during the HLA testing. Using a combination of generic and group-specific primers complementary to sequences in the exon 2 and 3 and intron 2 for HLA class I genes and the exon 2 for HLA class II can generate amplicons to confirm the homozygosity. In case of doubt, the largest amplicons are repeated with PacBio sequencing technology. In addition, known associations between the different genes are checked before to reaffirm their homozygous associations.
2.3 Statistical analysis
HLA typing results were coded by g-groups for statistical analysis. The allele frequencies (AFs) of HLA-A, HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 loci were calculated by direct counting using Arlequin 3.5.2.2 (45), and haplotype frequencies (HFs) were calculated using the expectation-maximization (EM) algorithm from the open-source software Hapl-o-Mat (46). The haplotypes with a frequency lower than 10-6 were not reported. Hardy-Weinberg equilibrium (HWE) was determined for each locus using Fisher’s exact test and, linkage disequilibrium (LD) between allele pairs was also estimated using Arlequin 3.5.2.2 (45). A p-value of < 0.05 was considered significant.
3 Results
3.1 Allele frequency
In total 52 HLA-A alleles, 111 HLA-B alleles, 41 HLA-C alleles, 47 HLA-DRB1 alleles, and 20 HLA-DQB1 alleles were identified for the 1763 samples analyzed. Complete AFs results are provided in Supplementary Table 2.
The 20 most frequent alleles for HLA class I loci are shown in Table 1. For the HLA-A locus, 21 alleles had a frequency higher than 1%, and the most frequent alleles in this locus were A*24:02g (20.87%), A*02:01g (16.11%), A*01:01g (7.06%), A*03:01g (6.83%), and A*29:02g (5.22%). For the HLA-B locus, 27 alleles occurred at a frequency higher than 1%, and the most frequent alleles were B*35:43g (7.69%), B*40:02g (7.18%), B*44:03g (6.07%), B*51:01g (5.90%), and B*07:02g (5.36%). Finally, for HLA-C locus, 17 alleles had a frequency greater than 1%, being the most frequent alleles C*04:01g (15.40%), C*01:02g (10.49%), C*07:02g (10.44%), C*07:01g (8.93%), and C*03:04g (8.05%).
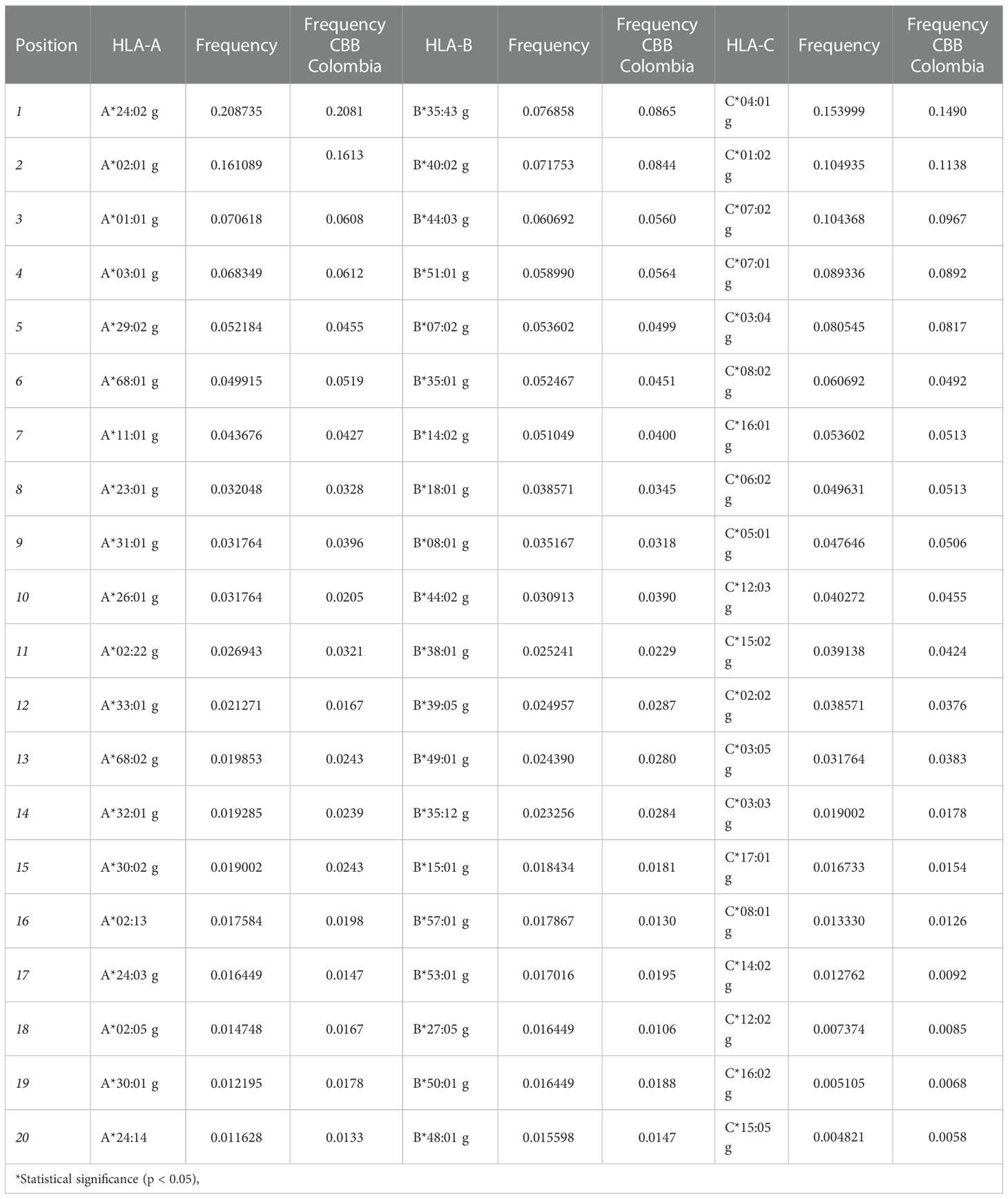
Table 1 List of the 20 most frequent alleles for HLA class I loci for the population in the study and a comparison with allele frequencies reported by the Colombian UCB Bank.
Table 2 provides the 20 most frequent alleles for HLA class II loci. The most common HLA-DRB1 alleles were DRB1*04:07g (11.03%), DRB1*07:01g (9.78%), DRB1*08:02g (6.72%), DRB1*03:01g (6.58%), and DRB1*15:01g (6.38%). While the 5 most frequent alleles for HLA-DQB1 were DQB1*03:02g (20.96%), DQB1*03:01g (17.78%), DQB1*02:01g (16.05%), DQB1*05:01g (11.91%), and DQB1*04:02g (11.11%). Furthermore, 25 HLA-DRB1, and 11 HLA-DQB1 alleles were counted with frequencies greater than 1%. Figure 2 shows the total number of alleles for each HLA loci and their individual frequency.
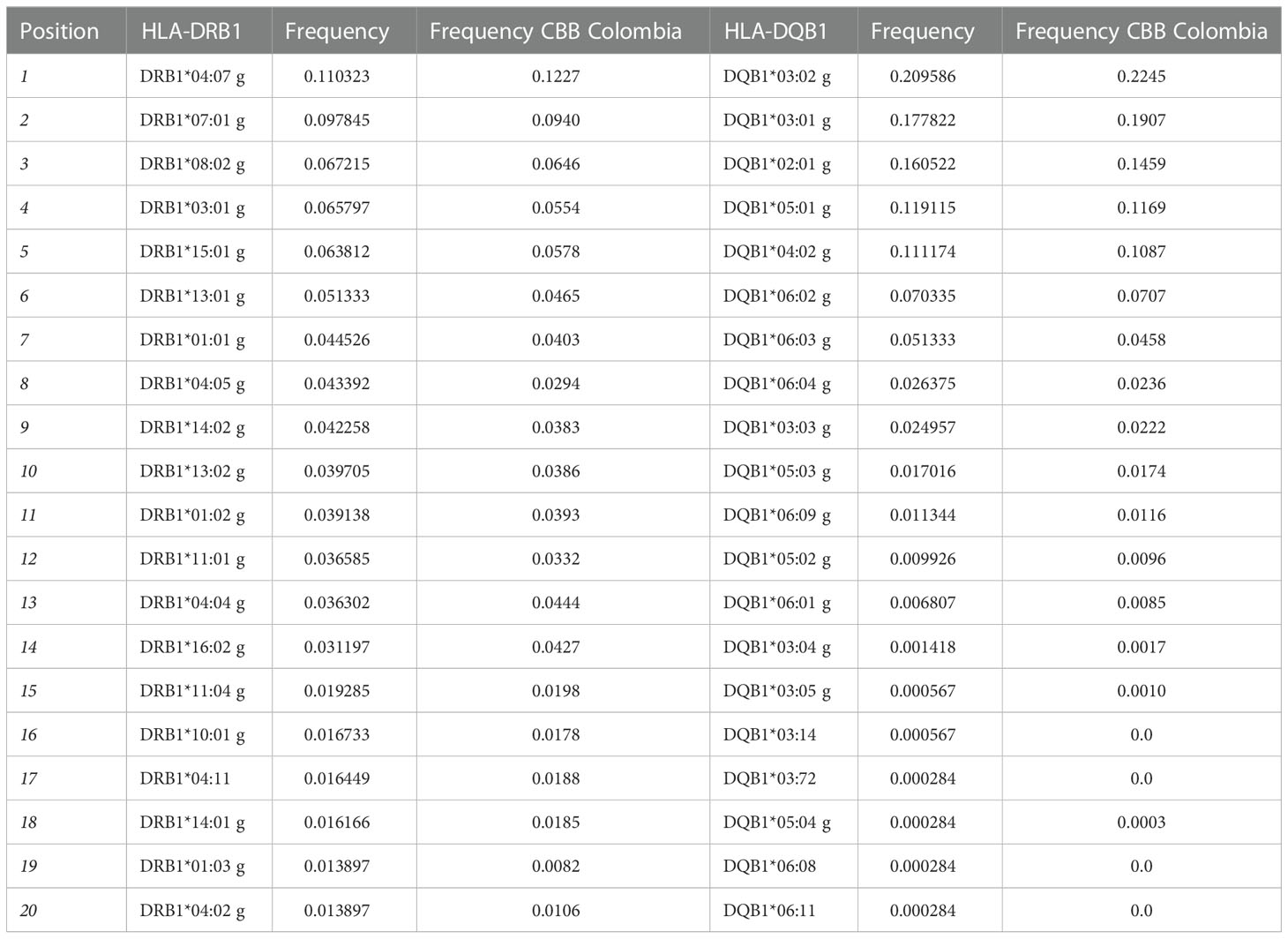
Table 2 List of the 20 most frequent alleles for HLA class II loci for the population in the study and a comparison with allele frequencies reported by the Colombian UCB Bank.
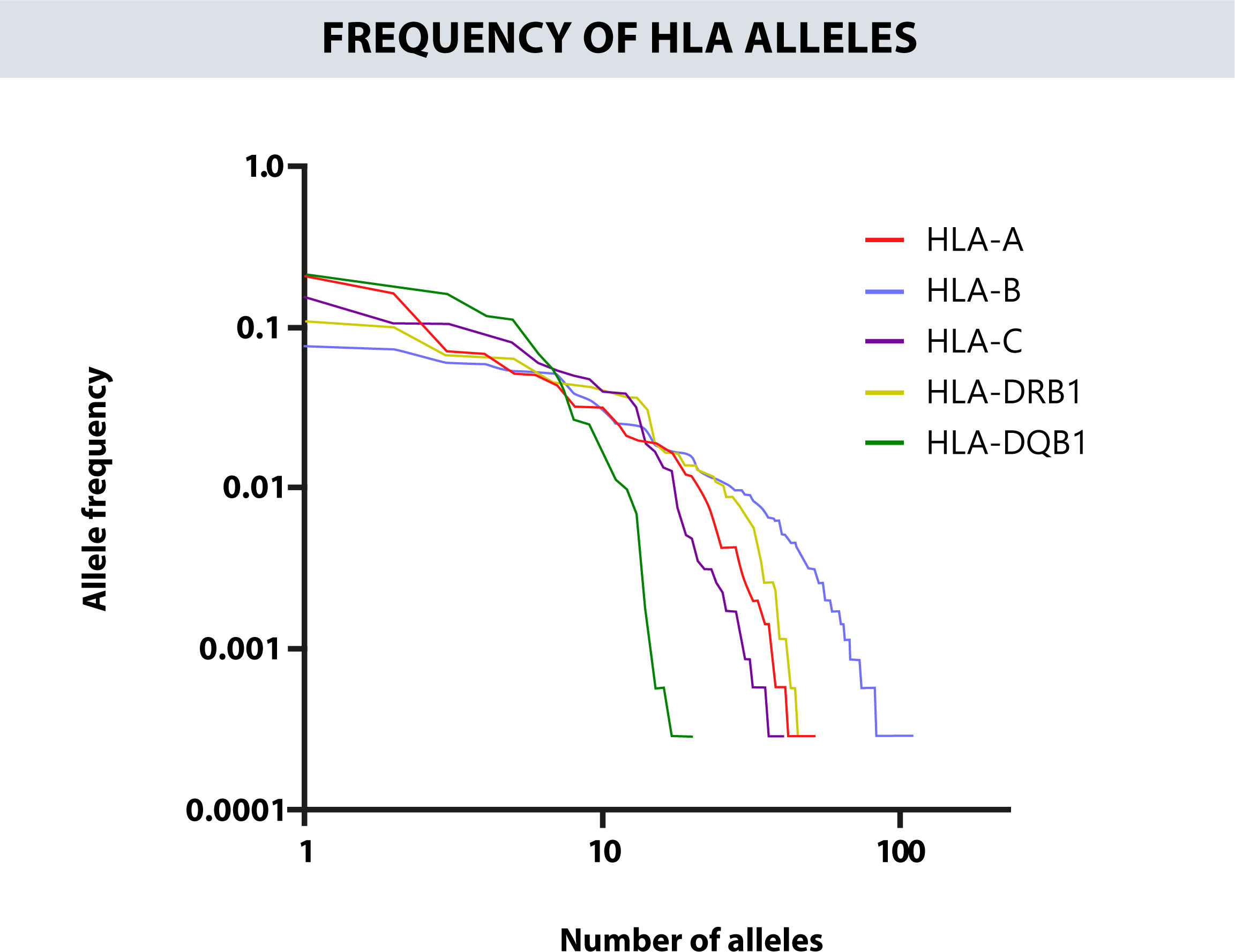
Figure 2 Represents the total number of alleles for each HLA loci versus its individual frequencies.
Both Tables 1, 2, include the AFs obtained previously by the UCB Bank from Bogotá, as we expected, by increasing the number of samples and the cities where were collected, we found a higher number of alleles, except for HLA-DRB1. In terms of the AFs, the most common alleles remain the same in both studies, however, some frequencies and their orders change slightly.
The cumulative frequency of each locus for the first 20 alleles was 92.91% for the HLA-A, 72.97% for the HLA-B, 97.36% for the HLA-C, 86.59% for the HLA-DRB1, and 100% for HLA-DQB1 (Figure 3). The zygosity is similar between the different HLA loci. Most are heterozygous, especially in HLA-B where the percent of heterozygous reach 96% of the results. On the other hand, the HLA locus that shows more homozygosity is HLA-DRB1 with a 14% (Supplementary Figure 2).
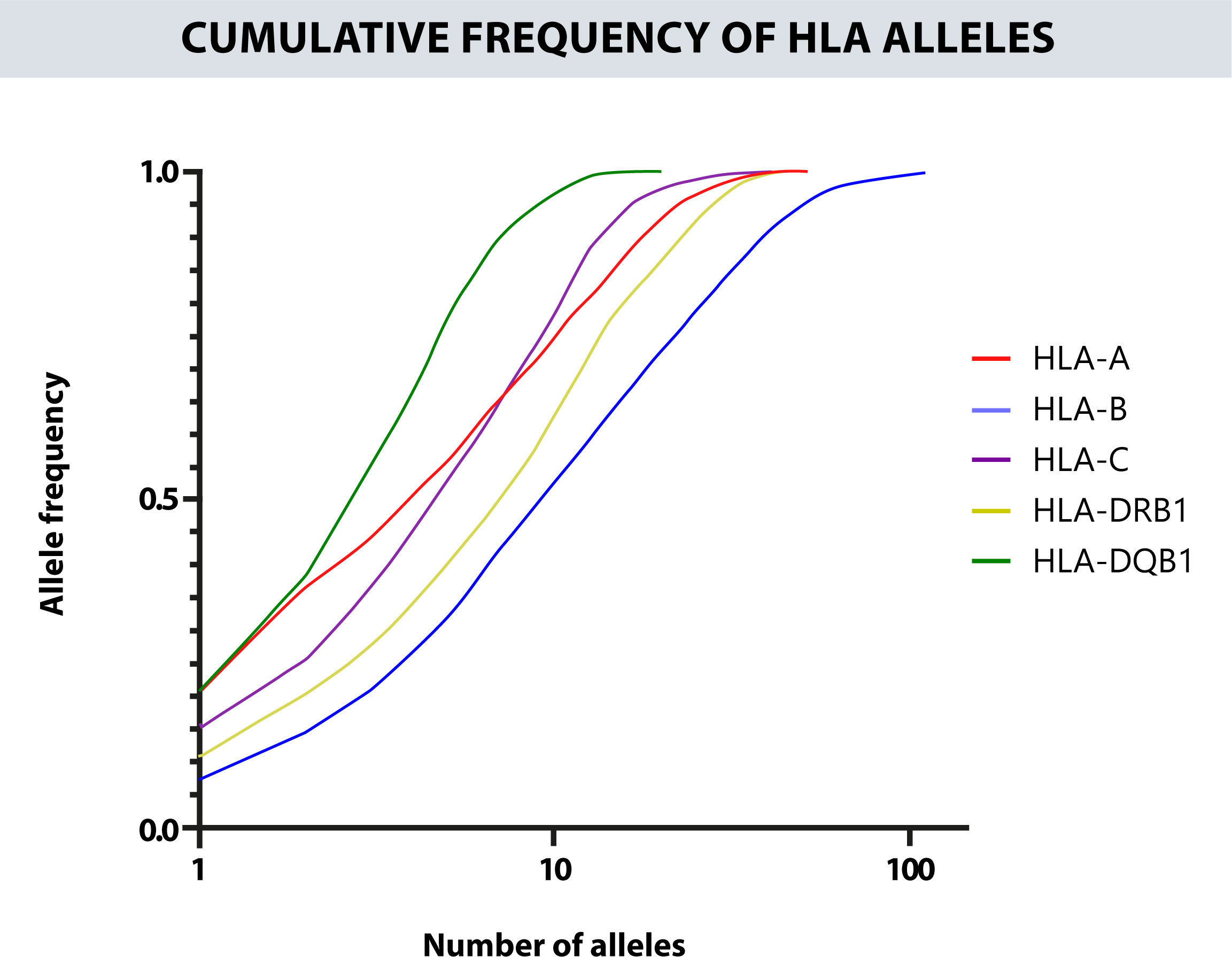
Figure 3 Cumulative frequency of the alleles for each HLA locus. The closer the curve is to the upper left end, the more homogeneous the locus will be. The x-axis is on a logarithmic scale. The most polymorphic locus is HLA-B, followed by HLA-DRB1 and HLA-A, and the least polymorphic is HLA-DQB1.
Finally, we compared the eleven most frequent HLA alleles between the 1347 women and 416 men to evaluate if gender might be influencing HLA segregation and how the AFs could be altered by it. The ratio of male/female is close to one in most of the cases for the 5 HLA loci, except for the alleles A*03:01g (ratio 0,703) and DQB1*05:03g (ratio 0,648) where the percent of males is meaningfully lower than females and, contrary, alleles DRB1*14:02g (ratio 1,357) and DRB1*08:02g (ratio 1,250) have a higher proportion of males than females (Supplementary Table 3).
3.2 Haplotype frequency
1626 different A~B~C~DQB1~DRB1 haplotypes were estimated with a frequency above 1e-6 among the 1763 typed samples. The haplotype A*24:02g~B*35:43g~C*01:02g~DQB1*03:02g~DRB1*04:07g occurred at the highest frequency (3.33%). There were 4 other haplotypes with a frequency higher than 1%:
A*29:02g~B*44:03g~C*16:01g~DQB1*02:01g~DRB1*07:01g (2.04%), A*01:01g~B*08:01g~C*07:01g~DQB1*02:01g~DRB1*03:01g (1.83%), A*33:01g~B*14:02g~C*08:02g~DQB1*05:01g~DRB1*01:02g (1.13%), and A*03:01g~B*07:02g~C*07:02g~DQB1*06:02g~DRB1*15:01g (1.05%). The 20 most frequent haplotypes are summarized in Table 3, and the haplotypes with a frequency higher than 0.1% are listed in Supplementary Table 4. In addition, in Table 3 we compared these 20 most frequent haplotypes with the haplotype data of the UCB Bank of Bogotá and the most frequent haplotypes registered in the United States Bone Marrow Registry (USA NMDP). Haplotypic frequencies of 4 different ethnic groups registered in the USA NMDP (Caucasian, African American, Middle Eastern and North African, and Hispanic from Central and South America), were accessed via the Internet portal, allelefrequiencies.net. Our 20 most frequent haplotypic frequencies are in the haplotypic frequencies reported in the UCB Bank of Bogota, although the frequencies are similar in both studies, the inclusion of records from other Colombian cities generates slight changes in haplotypic frequencies due to possible regional haplotypes not represented in Bogotá. On the other hand, it is remarkable, when we compare our most frequent haplotypes in the USA NMDP records for the 4 ethnic groups chosen, only 9 of the 20 haplotypes are found in these populations, even in the Hispanic population where we would expect similar results. The most frequent haplotype A*24:02g~B*35:43g~C*01:02g~DQB1*03:02g~DRB1*04:07g is much lower in all populations, even in the Hispanic population where the difference is almost 3 percentage points. The frequencies of the following 4 frequent haplotypes have similar proximity as expected to the Hispanic population.
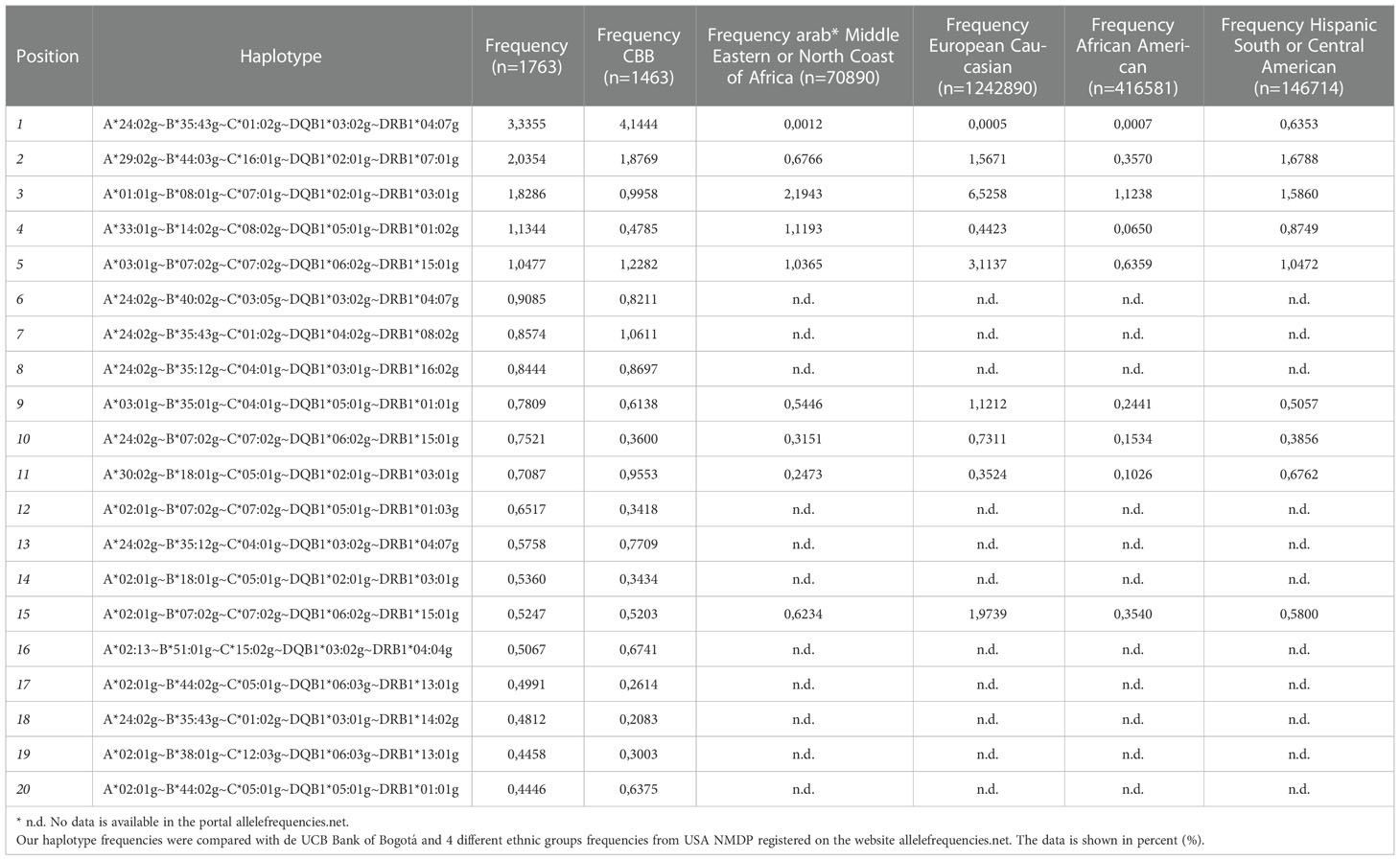
Table 3 The 20 most frequent A~B~C~DQB1~DRB1 haplotypes were identified for the donors from the Colombian BM Donor Registry.
Likewise, a frequency estimation of haplotypes A~B, A~B~C, A~B~DRB1, DRB1~DQB1, and A~B~C~DRB1 was performed. The Cumulative frequencies for each haplotype are shown in Figure 4. The most frequent haplotypes were A*24:02g~B*35:43g (5.23%), A*29:02g~B*44:03g (3.03%), A*24:02g~B*40:02g (2.89%); A*24:02g~B*35:43g~C*01:02g (5.09%), A*29:02g~B*44:03g~C*16:01g (2.94%), A*01:01g~B*08:01g~C*07:01g (2.31%); A*24:02g~B*35:43g~DRB1*04:07g (3.45%), A*29:02g~B*44:03g~DRB1*07:01g (2.07%), A*01:01g~B*08:01g~DRB1*03:01g (1.85%); A*24:02g~B*35:43g~C*01:02g~DRB1*04:07g (3.34%), A*29:02g~B*44:03g~C*16:01g~DRB1*07:01g (2.03%), A*01:01g~B*08:01g~C*07:01g~DRB1*03:01g (1.82%); DQB1*03:02g~DRB1*04:07g (10.86%), DQB1*02:01g~DRB1*07:01g (8.43%), and DQB1*04:02g~DRB1*08:02g (6.63%). Figure 5 shows the observed frequencies for each of the estimated haplotypes and the variation with the different combinations of loci.
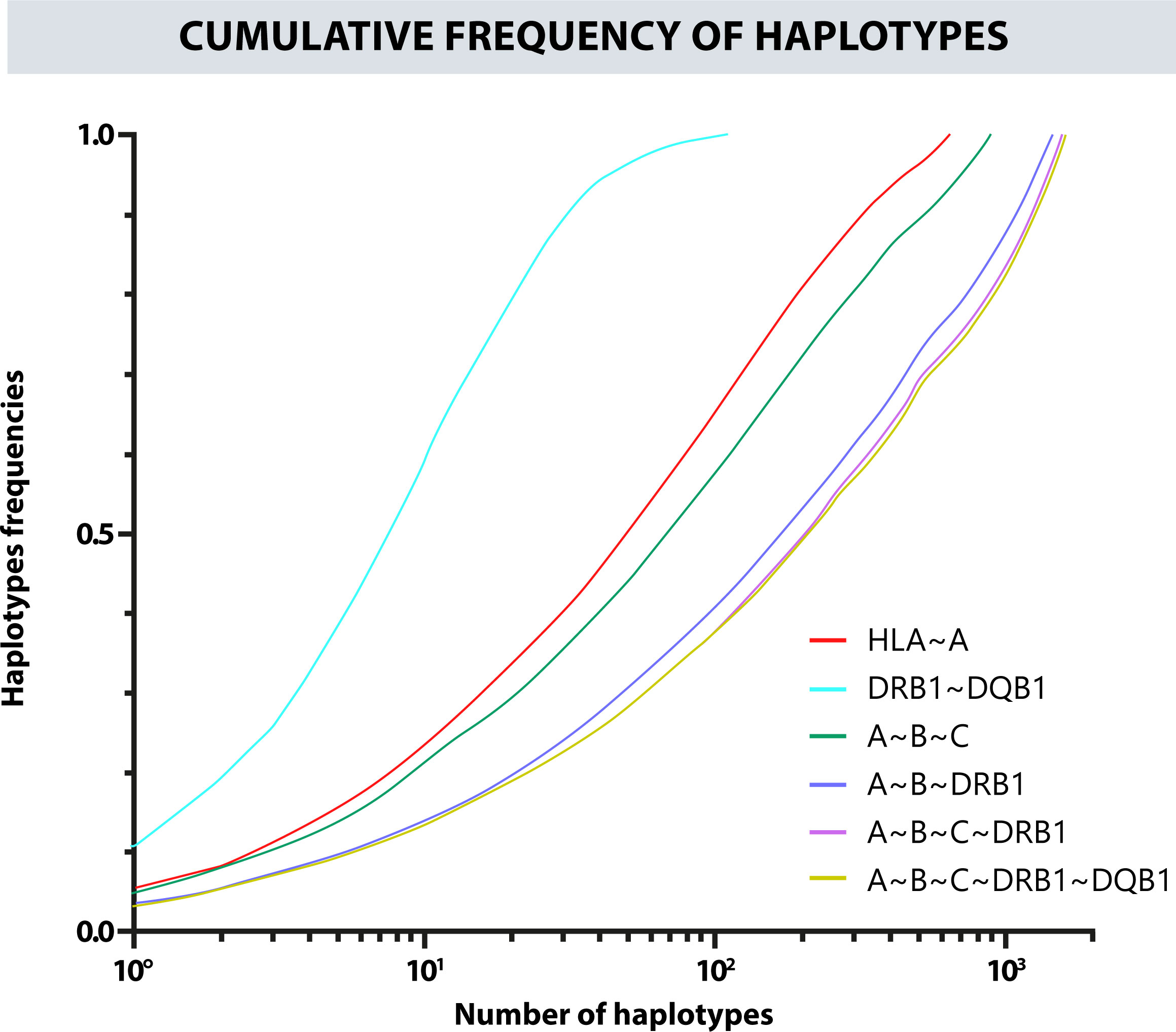
Figure 4 Cumulative frequency of the haplotypes for the different combinations of classic HLA loci.
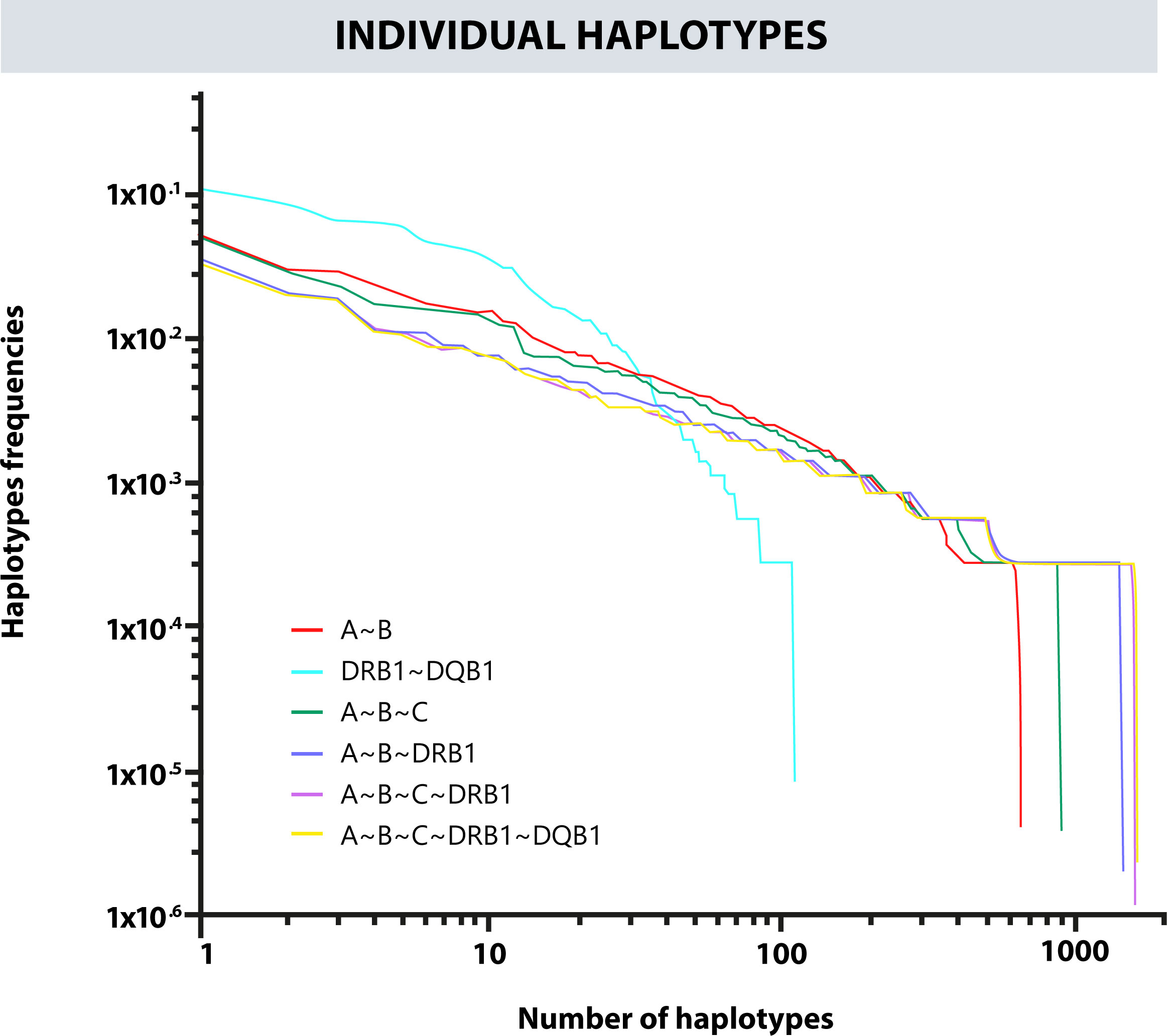
Figure 5 Frequency variation in each observed haplotype and their variation with different combinations of loci.
Table 4 provides the observed and expected heterozygosity, p-value, and standard deviation. Considering the allelic distributions of the HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 loci of the donors, the genotypic frequencies met HWE (p > 0.05); however, the distribution of the HLA-A locus showed significant deviation (p = 0.01440). In addition, LD was detected between HLA class I loci as well as class II loci (Supplementary Table 5).

Table 4 Hardy-Weinberg equilibrium (HWE) for the 1763 donors.
3.3 Detection of new alleles
Ten new HLA alleles were identified and submitted to the IPD-IMGT/HLA database. These alleles were named by the nomenclature committee of the World Health Organization (WHO). Seven new HLA alleles were described for class I; 6 of the 7 alleles, had a substitution of a nitrogenous base, generating an amino acid change, the remaining new allele being a synonymous substitution. Most of the substitutions occurred in exon 3 (4 changes), exon 4 (2 changes), and exon 2 (1 change). For HLA class II alleles, 3 changes occurred in exon 3, 2 of these substitutions generated an amino acid change; the remaining is a null allele generated by a 2-base deletion that causes a change in the reading frame, producing a premature stop codon (Table 5).
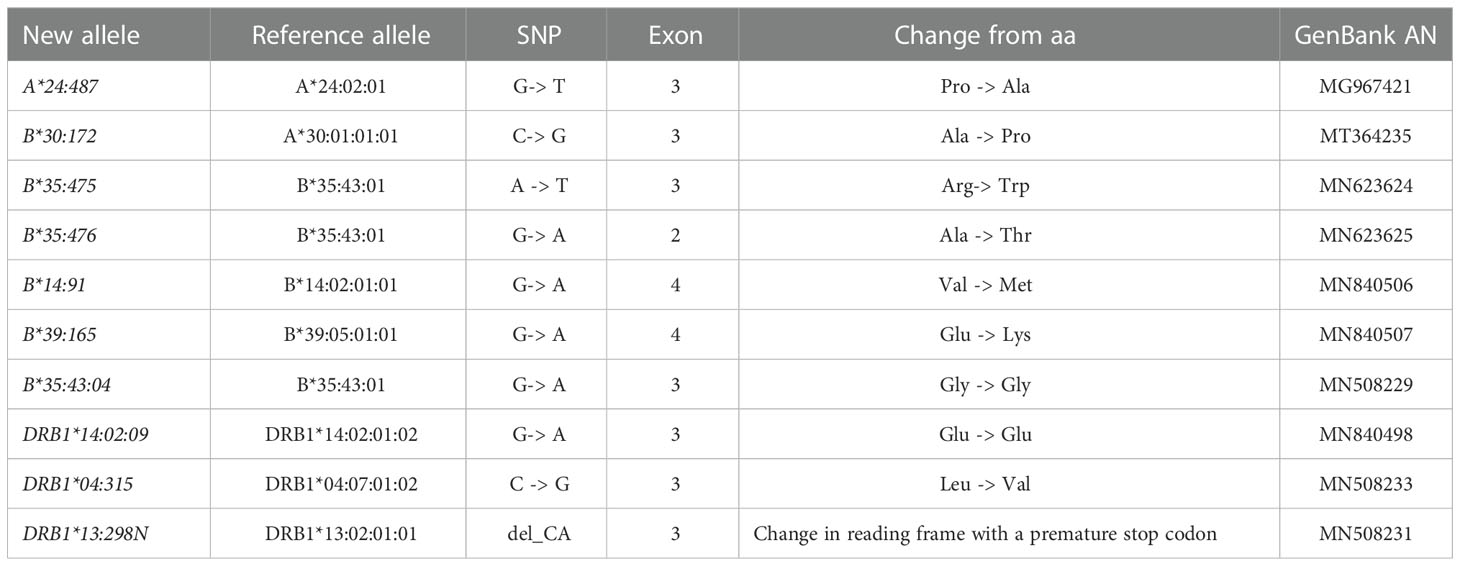
Table 5 New HLA alleles submitted currently.
3.4 AFs of class II loci and extended haplotypes
488 donor samples were typed at high resolution by DKMS Life Science Lab (Dresden, Germany) for classic HLA loci, including class II loci (HLA-DPB1, HLA-DPA1, and HLA-DQA1). The frequencies observed for each of these class II loci are summarized in Table 6. Among the donor samples, DRB3*02:02:01G (15.57%), DRB3*01:01:02G (14.14%), DRB3* 03:01:01G (6.04%), DRB4*01:01:01G (32.07%), and DRB5*01:01:01G (6.97%) occurred at a frequency higher than 5% (Supplementary Table 6). Subsequently, HFs were estimated, including the HLA-DPB1, HLA-DPA1, and HLA-DQA1 loci, generating an extended haplotype of 8 HLA loci for the donors. A total of 733 extended haplotypes were estimated, and the 20 most frequent haplotypes are summarized in Table 7, with the most frequent haplotype being A*24:02g~B*35:43g~C*01:02g~DPA1*02:01g~DPB1*14:01g~DQA1*03:01g~DQB1*03:02g~DRB1*04:07g, with a frequency of 1.437%.
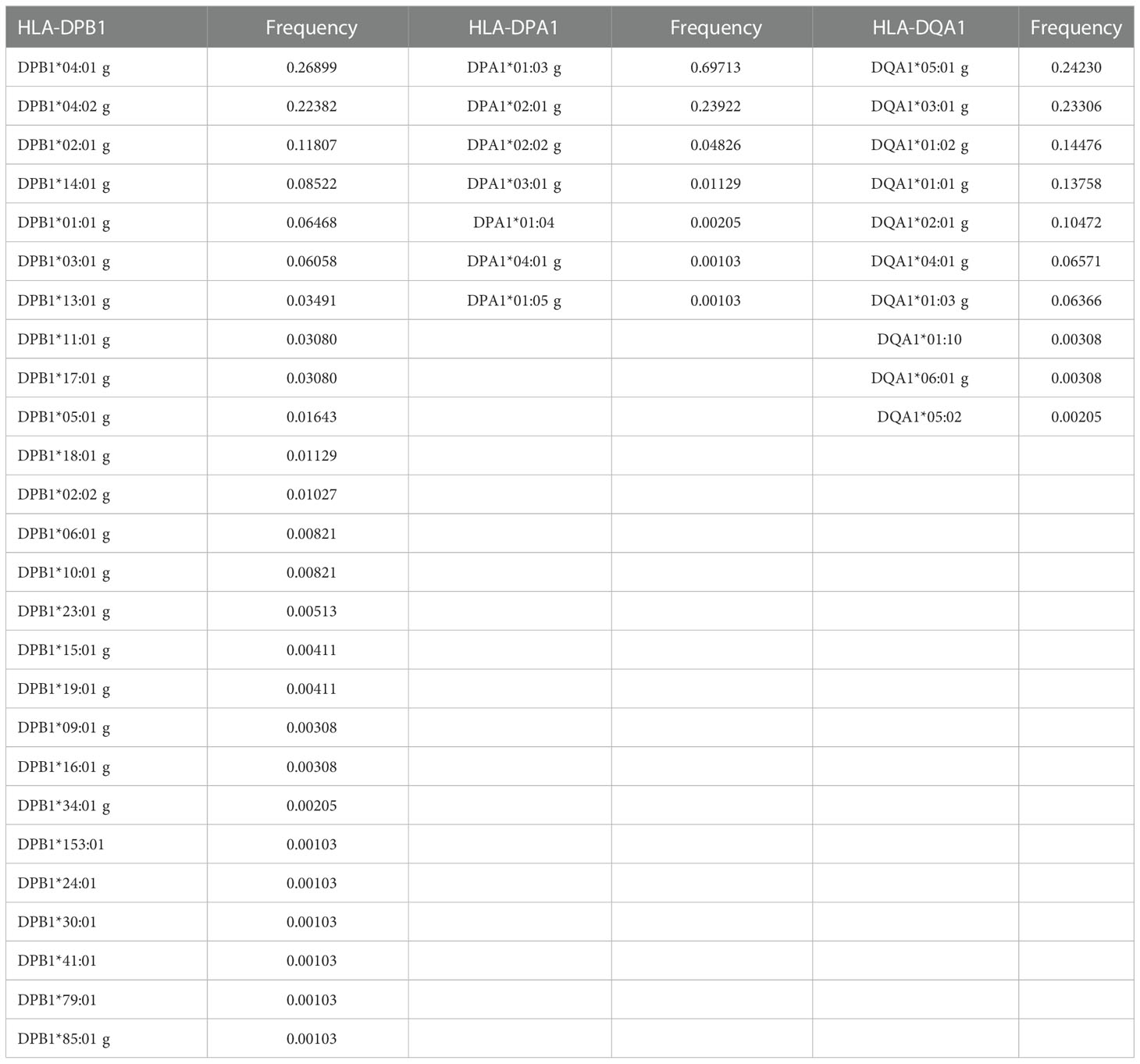
Table 6 Allele frequencies for class II, HLA-DPB1, HLA-DPA1, and HLA-DQA1 loci.

Table 7 20 extended haplotypes of the 8 most frequent classic HLA loci.
3.5 Distribution of HLA alleles and HF in Colombia
Within the project aiming to create a national BM Donor Registry in Colombia, a donor recruitment program was established in the 4 main cities of the country: Bogotá, Medellín, Cali, and Barranquilla. This program’s purpose was to increase the representation of the HLA alleles in the Registry and gain an overall view of the allelic distribution throughout the different regions in Colombia. For this analysis, the AFs of the HLA-A, -B, -C, -DRB1, and -DQB1 loci of the donors from the departments with the highest number of records were included, i.e., Antioquia, Valle del Cauca, Atlántico, Cundinamarca, Boyacá, Santander, Tolima, and Bogotá. Table 8 shows the 11 most frequent alleles for each HLA locus in Bogotá, which is the city with most of the records. These frequencies were compared by departments regarding their frequency position, for each population. Using the HLA AFs of the 5 loci for each department, principal component analysis (PCA) was performed to determine whether the distribution of these frequencies would allow differentiating the populations (Figure 6). Populations were categorized into 3 groups using PCA: Bogotá was alone in the first group; donors from Tolima, Boyacá, Santander, and Cundinamarca were in the second group; and the population from Atlántico, Antioquia, and Valle del Cauca, which were more dispersed, were in the third group.
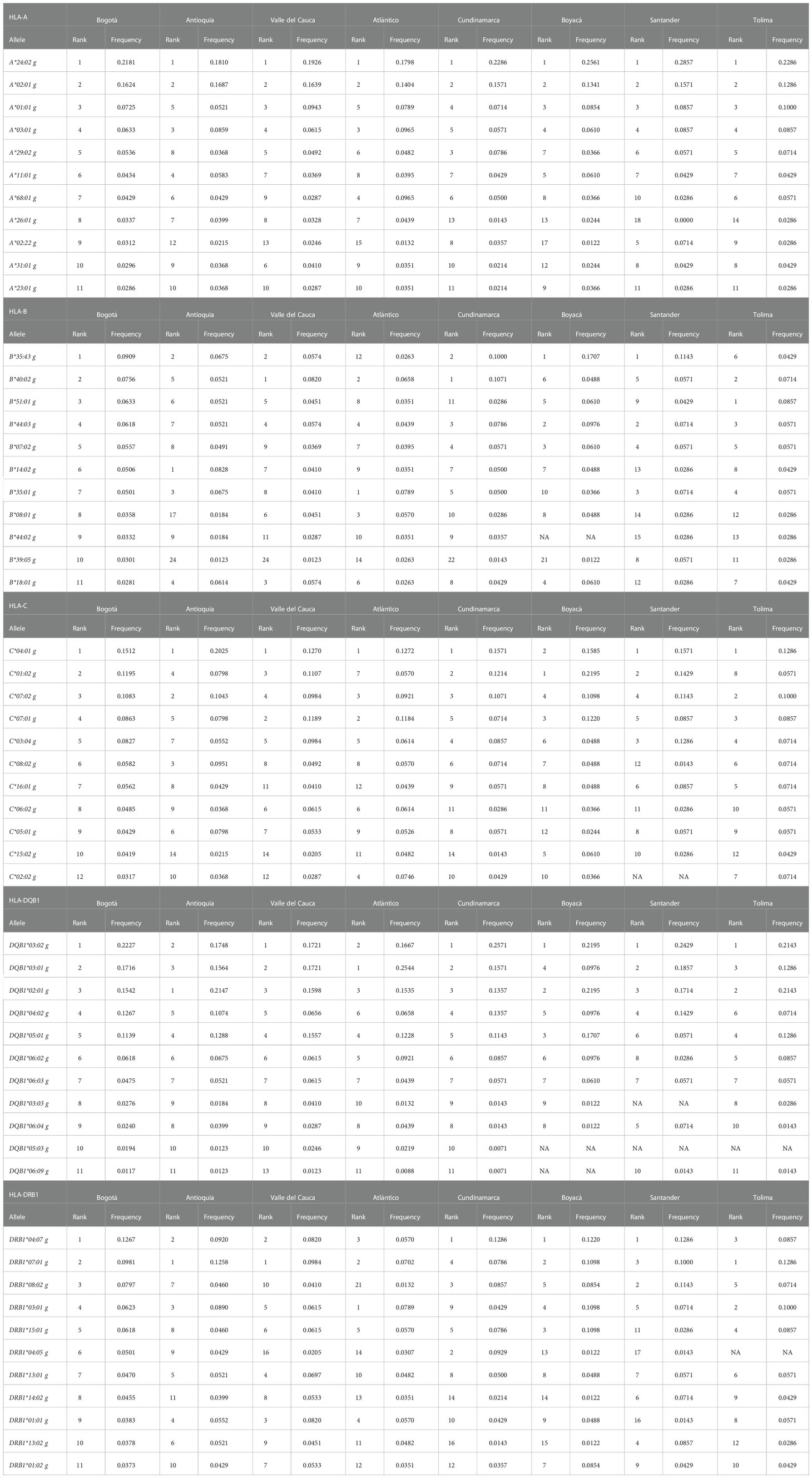
Table 8 The 11 most frequent HLA-A, HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 alleles in the city of Bogotá and the rank that these alleles occupied in the departments of Antioquia, Valle del Cauca, Atlántico, Cundinamarca, Boyacá, Santander and Tolima.
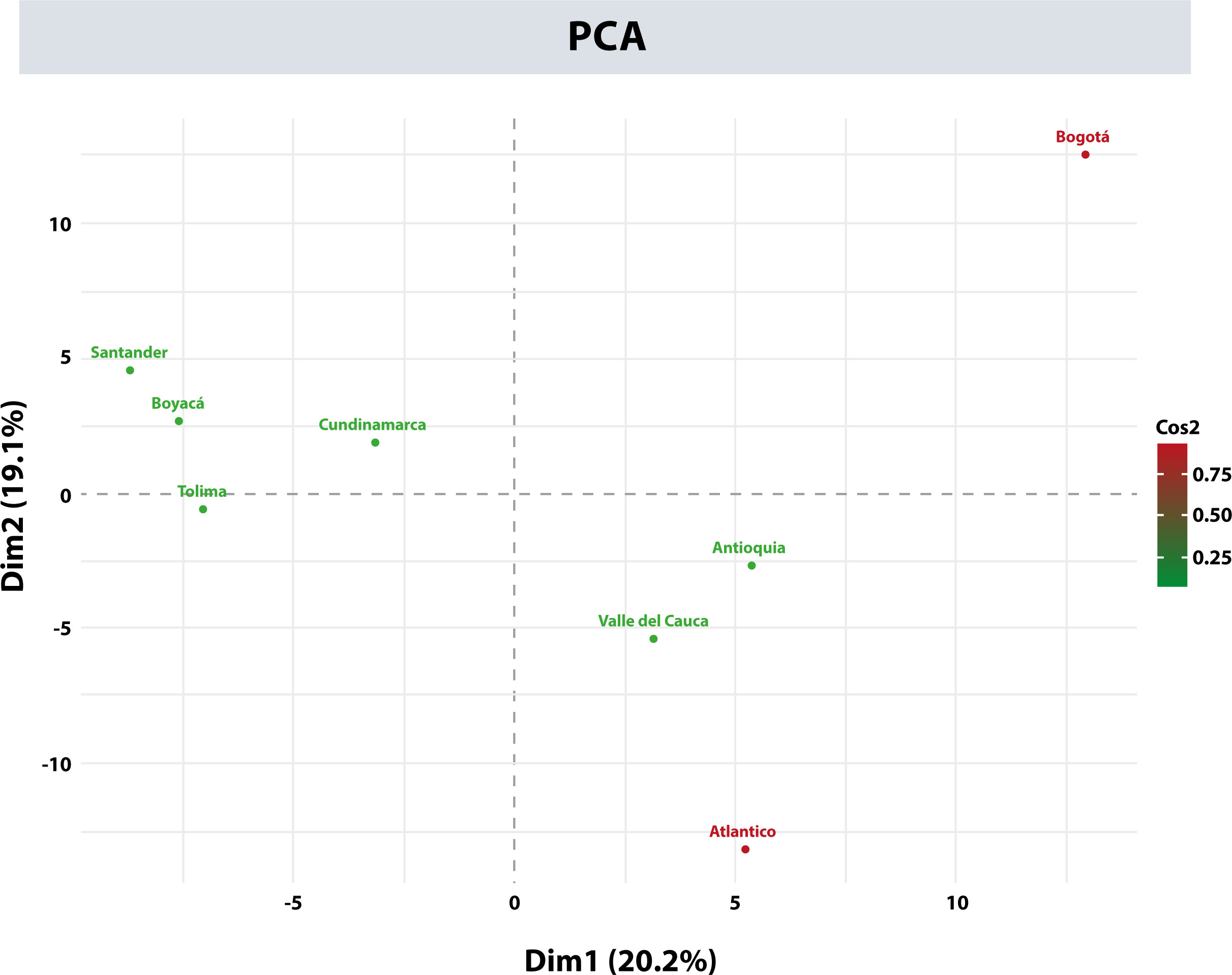
Figure 6 Principal component analysis (PCA) with allele frequencies for the departments of Antioquia, Valle del Cauca, Atlántico, Cundinamarca, Boyacá, Santander, Tolima, and Bogotá.
The A~B~C~DQB1~DRB1 haplotypes from each department were also estimated. The most frequent haplotypes are compiled in Supplementary Table 7. Notably, the A*24:02g~B*35:43g~C*01:02g~DQB1*03:02g~DRB1*04:07g haplotype was the most frequent in all departments except for Atlántico, Santander, and Tolima.
4 Discussion
This study is part of the initiative led in the last 5 years by the IDCBIS, which aims to create and consolidate a National BM Donor Registry in Colombia, willing to provide citizens with an affordable therapeutic option for allogeneic HSC transplant because this alternative cannot always be considered due to economic, medical or logistical reasons. Despite the limitations associated with the sample size, the results and analyses in this work are intended to have a better understanding of the genetic diversity in Colombia. This is the largest high-resolution study for classic HLA loci, including donors recruited in the 4 main cities of the country: Bogotá, Medellín, Cali, and Barranquilla carried out by the Colombian BM Donor Registry.
Páez-Gutiérrez et al. (47) reported a number of similar alleles in Bogotá to those in our sample. A slight increase in the number of alleles was observed for the HLA-A (+1), HLA-C (+5), and HLA-DQB1 (+2) loci, and a striking increase was observed for the most polymorphic locus HLA-B (+13). The class II locus, HLA-DRB1 (-4) was the only one with a reduction in the number of alleles observed. In both studies, alleles A*24:02g (20.87 - 0.06%), B*35:43g (7.69 + 0.96%), C*04:01g (15.40 – 0,50%), DRB1*04:07g (11.03 + 1.24%) and DQB1*03:02g (20.96 + 1.49%) were the most frequent. For the A~B~C~DQB1~DRB1 haplotypes, 187 more were detected in our study than in the study by Páez-Gutiérrez et al. (47); nevertheless, the A*24:02g~B*35:43g~C*01:02g~DQB1*03:02g~DRB1*04:07g haplotype was the most frequent in both studies, although the standard deviation was close to a percentage point (3.33 + 0.81%). The frequencies of the HLA-DRB1 and HLA-DQB1 loci and the DRB1*04:07g~DQB1*0302g haplotype in the study by Del Río-Ospina et al. (48) are consistent with our results. We also found similarities in the reported HLA alleles and their frequencies in other published studies involving the Colombian population; however, these studies were not performed in high-resolution typing (49–52).
The fact that the most frequent haplotype in our population is not represented in the compared populations from the USA NMDP, including the Hispanic population, and in addition, the absence of 11 of the 20 most frequent haplotypes compared, leads us to hypothesize that the migratory processes occurred in Colombia and the component of native groups have generated haplotypic combinations unusual or infrequent in other populations. As the size of the Colombian registry increases and its scope is extended to different regions and different populations, it will be possible to confirm whether these AFs and HFs are really particular to Colombians.
NGS used for typing the HLA loci led to the identification of 10 new HLA alleles among the 1763 samples analyzed. Most new alleles occurred in the HLA-B locus, and a null HLA-DRB1 allele was identified. The nucleotide sequences of the 10 new alleles were submitted to GenBank with the accession numbers MG967421, MT364235, MN23624, MN623625, MN840506, MN 840507, MN84508229, MN840498, MN508233, and MN508231 and were assigned the names A*24:487, B*30:172, B*:35:475, B*35:376, B*14:91, B*39:165, B*35:53:04, DRB1*14:02:09, DRB1*04:315, and DRB1*13:298N by the HLA system nomenclature committee of the WHO (53–58). The detection of these 10 alleles in a sample that represents approximately 0.01% of the Colombian population may indicate that the number of possible new alleles not reported circulating in the country is in the thousands if the trend found herein were maintained. In addition, the majority of the donors included in the registry corresponds to the mestizo population (97.41%), while the representation in the registry of other minority ethnic groups, such as Afro-American and Native American, is only 0.89% and 0.39%, respectively. Therefore, it would be expected that the identification of new HLA alleles in these minority groups will be higher as their participation in the Colombian registry increases.
The role of the HLA-DPB1 has gained greater relevance due to its association with various pathologies, including cancer (59), autoimmune diseases, such as rheumatoid arthritis (60), responses to viral infection (61), and vaccines (62, 63). Regarding stem cell transplants, there is still controversy about the inclusion of the HLA-DPB1 locus in the search algorithms for 12/12 histocompatibility (64) due to the increasing number of reports indicating that a mismatch in this locus increases the risk of developing GVHD (65). Data for HLA-DPB1 and other HLA class II loci is reported for the first time in high-resolution in Colombia. Notably, the most frequent alleles were DPB1*04:01g (26, 90%), DPB1*04:02g (22.38%), and DPB1*02:01g (11.81%); however, when analyzing the extended haplotype of the 8 most frequent loci, i.e., A*24:02g~B*35:43g~C*01:02g~DPA1*02:01g~DPB1*14:01g~DQA1*03:01g~DQB1*03:02g~DRB1*04:07g, the accompanying HLA-DPB1 allele was DPB1*14:01g, which had an AF of 8.52%. Likewise, from the analysis of the data of 488 donors, the first report of frequencies of the DRB3/4/5 alleles in Colombia was generated, with the most frequent alleles for each locus being DRB3*02:02:01G (15.57%), DRB4*01:01:01G (32.07%) and DRB5*01:01:01G (6.97%). Although these genes are not currently included in the selection of unrelated donors, there is increasing evidence of their importance in the incidence of acute GVHD (66).
Recruiting donors in several cities of Colombia allowed the registration of the HLA typing of donors from almost all departments in the national territory, except the Amazonian departments. In addition, it allowed an exploratory analysis of the behavior of AFs and HFs among the 8 departments with the most records: Antioquia, Valle del Cauca, Atlántico, Cundinamarca, Boyacá, Santander, Tolima, and Bogotá. Based on these AFs and HFs, the allele and haplotype distribution are quite heterogeneous in Colombia, especially in the department of Atlántico. In this department, the most frequent alleles and haplotypes vary significantly from those in the rest of the departments analyzed, especially the frequencies of the HLA-B, -DRB1, and -DQB1 loci.
Contrasting our data represents a challenge due to the lack of high-resolution studies of HLA loci frequencies in Colombia; nevertheless, the results published by the public UCB Bank of Bogotá (47) were similar to ours in number of alleles, and their frequencies for the city. We propose that the heterogeneous behavior observed among departments concerning AFs and HFs are mainly generated by differences in the population for each region. Increase the number of donors in the other departments might confirm this heterogeneous behavior and will be interesting to combine with other studies such as STRs or AIMs to confirm the differences in the region populations (67).
In Latin America AFs and HFs reported by countries bordering Colombia, such as Panamá (68), Venezuela (69), and Ecuador (70), showed frequencies similar to those described in our study, where alleles HLA-A*2, -A*24, -B*35, -DRB1*04 and -DQB1*0302 are the most frequent, and haplotype A*24~B*35~DRB1*04~DQB1*03:02 is the most frequent in Ecuador, Panamá, and Colombia. This coincidence may be explained by common migratory and historical processes over the centuries. In contrast, in the south of the continent the haplotype A*29~B*44~DRB1*07, the most frequent in Spain (71) and also found in other European populations, was reported. Other European haplotypes such as A*01~B*08~DRB1*03, and A*03~B*07~DRB1*15 occur frequently in Brazilian (REDOME) (72), Argentina (INCUCAI) BM Donor Registries (73), and the UCB banks from Buenos Aires (Garrahan Pediatric Hospital) (74), and Santiago de Chile (75). The presence of a greater European component in the south of the continent shows the difference in the processes of miscegenation that occurred in the north and the south.
This study is the first approach of the Colombian BM Donor Registry toward understanding the genetic diversity in Colombia, a result of migratory processes and centuries of miscegenation among different ethnic groups that have coexisted in the national territory. Although the number of registered and typed donors is still insufficient to determine this genetic variety, it is a starting point for better understanding the distribution of the alleles and haplotypes that compose the genes of the MHC in the country. This work will allow the recent Colombian BM Donor Registry to design donor recruitment strategies, both to increase the number of donors registered, and to target regions where non-common HLA alleles and haplotypes are found more frequently, to offer a therapeutic transplant option to all Colombians regardless their origin or ethnic group.
Likewise, we hope that these results will be a source of reference for subsequent studies of populations in Colombia and become a useful tool to determine the size of the national registry, moreover, allowing Colombia’s national BM Donor Registry to establish future collaborations with other centers, institutes and organizations worldwide.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by Comité de Ética de la Secretaría Distrital de Salud. The patients/participants provided their written informed consent to participate in this study.
Author contributions
DH-M, IP-G, and VD contributed to the initial conception of analyzing the HLA frequencies and haplotypes in Colombia and reviewing the available bibliography. NC, MM, and PC contributed with the BM Donor Registry perspective for distribution within the country. DH-M, IP-G, VD, NC, MM, and PC drafted, reviewed, and approved the submitted version of the manuscript. BC had financial support and supervised the project.
Funding
This research was conducted within the project entitled “Estudios técnicos para el establecimiento y organización de un Registro Nacional de Donantes de Células Progenitoras Hematopoyéticas en Colombia” [Technical studies for the establishment of a National Bone Marrow Donor Registry in Colombia] - BPIN2015000100054, funded by the Sistema General de Regalías and executed by the Instituto Distrital de Ciencia, Biotecnología and Innovación en Salud – IDCBIS.
Acknowledgments
We thank the donors of DarCélulas program from the Instituto Distrital de Ciencia, Biotecnología e Innovación en Salud (IDCBIS), for their valuable contributions by being part of the first bone marrow donor registry in Colombia.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.1057657/full#supplementary-material
References
1. Bazinet A, Popradi G. A general practitioner's guide to hematopoietic stem-cell transplantation. Curr Oncol (2019) 26:187–91. doi: 10.3747/co.26.5033
2. Duarte RF, Labopin M, Bader P, Basak GW, Bonini C, Chabannon C, et al. Indications for haematopoietic stem cell transplantation for haematological diseases, solid tumours and immune disorders: current practice in Europe, 2019. Bone Marrow Transplant (2019) 54:1525–52. doi: 10.1038/s41409-019-0516-2
3. Lee SJ, Klein J, Haagenson M, Baxter-Lowe LA, Confer DL, Eapen M, et al. High-resolution donor-recipient HLA matching contributes to the success of unrelated donor marrow transplantation. Blood (2007) 110:4576–83. doi: 10.1182/blood-2007-06-097386
4. Morishima Y, Kashiwase K, Matsuo K, Azuma F, Morishima S, Onizuka M, et al. Biological significance of HLA locus matching in unrelated donor bone marrow transplantation. Blood (2015) 125:1189–97. doi: 10.1182/blood-2014-10-604785
5. Kanakry CG, Fuchs EJ, Luznik L. Modern approaches to HLA-haploidentical blood or marrow transplantation. Nat Rev Clin Oncol (2016) 13:10–24. doi: 10.1038/nrclinonc.2015.128
6. Petersdorf EW. Which factors influence the development of GVHD in HLA-matched or mismatched transplants? Best Pract Res Clin Haematol (2017) 30:333–5. doi: 10.1016/j.beha.2017.09.003
7. Debals-Gonthier M, Siani C, Faucher C, Touzani R, Lemarié-Basset C, Chabannon C, et al. Cost-effectiveness analysis of haploidentical vs matched unrelated allogeneic hematopoietic stem cells transplantation in patients older than 55 years. Bone Marrow Transplant (2018) 53:1096–104. doi: 10.1038/s41409-018-0133-5
8. Dehn J, Buck K, Maiers M, Confer D, Hartzman R, Kollman C, et al. 8/8 and 10/10 high-resolution match rate for the be the match unrelated donor registry. Biol Blood Marrow Transplant (2015) 21:137–41. doi: 10.1016/j.bbmt.2014.10.002
9. Gragert L, Eapen M, Williams E, Freeman J, Spellman S, Baitty R, et al. HLA match likelihoods for hematopoietic stem-cell grafts in the U.S. registry. N Engl J Med (2014) 371:339–48. doi: 10.1056/NEJMsa1311707
10. Al Malki MM, Tsai N-C, Palmer J, Mokhtari S, Tsai W, Cao T, et al. Posttransplant cyclophosphamide as GVHD prophylaxis for peripheral blood stem cell HLA-mismatched unrelated donor transplant. Blood Adv (2021) 5:2650–9. doi: 10.1182/bloodadvances.2021004192
11. Petersdorf EW. Optimal HLA matching in hematopoietic cell transplantation. Curr Opin Immunol (2008) 20:588–93. doi: 10.1016/j.coi.2008.06.014
12. Piemontese S, Ciceri F, Labopin M, Arcese W, Kyrcz-Krzemien S, Santarone S, et al. A comparison between allogeneic stem cell transplantation from unmanipulated haploidentical and unrelated donors in acute leukemia. J Hematol Oncol (2017) 10:1–8. doi: 10.1186/s13045-017-0394-2
13. Wagner JE, Barker JN, DeFor TE, Baker KS, Blazar BR, Eide C, et al. Transplantation of unrelated donor umbilical cord blood in 102 patients with malignant and nonmalignant diseases: influence of CD34 cell dose and HLA disparity on treatment-related mortality and survival. Blood (2002) 100:1611–8. doi: 10.1182/blood-2002-01-0294
14. Ciurea SO, Cao K, Fernandez-Vina M, Kongtim P, Malki MA, Fuchs E, et al. The European society for blood and marrow transplantation (EBMT) consensus guidelines for the detection and treatment of donor-specific anti-HLA antibodies (DSA) in haploidentical hematopoietic cell transplantation. Bone Marrow Transplant (2018) 53:521–34. doi: 10.1038/s41409-017-0062-8
15. Dehn J, Spellman S, Hurley CK, Shaw BE, Barker JN, Burns LJ, et al. Selection of unrelated donors and cord blood units for hematopoietic cell transplantation: guidelines from the NMDP/CIBMTR. Blood (2019) 134:924–34. doi: 10.1182/blood.2019001212
16. Kanda J, Umeda K, Kato K, Murata M, Sugita J, Adachi S, et al. Effect of graft-versus-host disease on outcomes after pediatric single cord blood transplantation. Bone Marrow Transplant (2020) 55:1430–7. doi: 10.1038/s41409-020-0853-1
17. Kekre N, Antin JH. Hematopoietic stem cell transplantation donor sources in the 21st century: choosing the ideal donor when a perfect match does not exist. Blood (2014) 124:334–43. doi: 10.1182/blood-2014-02-514760
18. Rafiee M, Abbasi M, Rafieemehr H, Mirzaeian A, Barzegar M, Amiri V, et al. A concise review on factors influencing the hematopoietic stem cell transplantation main outcomes. Health Sci Rep (2021) 4:1–9. doi: 10.1002/hsr2.282
19. Antunes DA, Abella JR, Devaurs D, Rigo MM, Kavraki LE. Structure-based methods for binding mode and binding affinity prediction for peptide-MHC complexes. Curr Top Med Chem (2019) 18:2239–55. doi: 10.2174/1568026619666181224101744
20. Taylor BC, Balko JM. Mechanisms of MHC-I downregulation and role in immunotherapy response. Front Immunol (2022) 13:844866. doi: 10.3389/fimmu.2022.844866
21. Douillard V, Castelli EC, Mack SJ, Hollenbach JA, Gourraud P-A, Vince N, et al. Approaching genetics through the MHC lens: tools and methods for HLA research. Front Genet (2021) 12:774916. doi: 10.3389/fgene.2021.774916
22. Morishima S, Kashiwase K, Matsuo K, Azuma F, Yabe T, Sato-Otsubo A, et al. High-risk HLA alleles for severe acute graft-versus-host disease and mortality in unrelated donor bone marrow transplantation. Haematologica (2016) 101:491–8. doi: 10.3324/haematol.2015.136903
23. Naito T, Okada Y. HLA imputation and its application to genetic and molecular fine-mapping of the MHC region in autoimmune diseases. Semin Immunopathol (2022) 44:15–28. doi: 10.1007/s00281-021-00901-9
24. Scavuzzi BM, van Drongelen V, Holoshitz J. HLA-G and the MHC cusp theory. Front Immunol (2022) 13:814967. doi: 10.3389/fimmu.2022.814967
25. Rigden DJ, Fernández XM. The 27th annual nucleic acids research database issue and molecular biology database collection. Nucleic Acids Res (2020) 48:D1–8. doi: 10.1093/nar/gkz1161
26. Robinson J, Barker DJ, Georgiou X, Cooper MA, Flicek P, Marsh SGE. IPD-IMGT/HLA database. Nucleic Acids Res (2020) 48:D948–55. doi: 10.1093/nar/gkz950
27. Andreani M, Gregori S. The study of engraftment after hematopoietic stem cell transplantation: from the presence of mixed chimerism to the development of immunological tolerance. HLA (2018) 92:57–9. doi: 10.1111/tan.13402
28. Baumeister SHC, Rambaldi B, Shapiro RM, Romee R. Key aspects of the immunobiology of haploidentical hematopoietic cell transplantation. Front Immunol (2020) 11:191. doi: 10.3389/fimmu.2020.00191
29. Fürst D, Müller C, Vucinic V, Bunjes D, Herr W, Gramatzki M, et al. High-resolution HLA matching in hematopoietic stem cell transplantation: a retrospective collaborative analysis. Blood (2013) 122:3220–9. doi: 10.1182/blood-2013-02-482547
30. Ghimire S, Weber D, Mavin E, Wang XN, Dickinson AM, Holler E. Pathophysiology of GvHD and other HSCT-related major complications. Front Immunol (2017) 8:79. doi: 10.3389/fimmu.2017.00079
31. Kekre N, Mak KS, Stopsack KH, Binder M, Ishii K, Brånvall E, et al. Impact of HLA-mismatch in unrelated donor hematopoietic stem cell transplantation: a meta-analysis. Am J Hematol (2016) 91:551–5. doi: 10.1002/ajh.24342
32. Kunze-Schumacher H, Blasczyk R, Bade-Doeding C. Soluble HLA technology as a strategy to evaluate the impact of HLA mismatches. J Immunol Res (2014) 2014:246171. doi: 10.1155/2014/246171
33. Verneris MR, Lee SJ, Ahn KW, Wang H-L, Battiwalla M, Inamoto Y, et al. HLA mismatch is associated with worse outcomes after unrelated donor reduced-intensity conditioning hematopoietic cell transplantation: an analysis from the center for international blood and marrow transplant research. Biol Blood Marrow Transplant (2015) 21:1783–9. doi: 10.1016/j.bbmt.2015.05.028
34. Landaburu J. Las lenguas indígenas de Colombia: Presentación y estado del Arte Vol. N° 29/30. . París: Amerindia (2006) p. 1–22.
35. Noguera-Santamaría MC, Rivera-Sandoval J, Martín JG, Briceño-Balcázar I, Gómez-Gutiérrez A. Análisis genético de restos humanos precolombinos del bajo Magdalena sugiere una ruta migratoria y continuidad genética matrilineal en el norte de suramérica. Rev Acad Colomb Cienc Exactas Fís Nat (2020) 44:704–15. doi: 10.18257/raccefyn.973
36. Bolívar FJF. Celebrating and redefining the mestizaje: race and nation during the liberal republic, Colombia, 1930-1946. Mem Rev Digit Hist Arqueol Desde Caribe (2019) 15:93–116. doi: 10.14482/memor.37.986.104
37. Wade P. Blackness, indigeneity, multiculturalism and genomics in Brazil, Colombia and Mexico. J Lat Am Stud (2013) 45:205–33. doi: 10.1017/s0022216x13000011
38. Lecciones YMB. Desplazamientos forzados aprendidas en colombia forced displacements and migrations. Br Lessons Learn Colomb (2019) 25:1–18.
39. Centro Nacional de Memoria Histórica. Una nación desplazada: informe nacional del desplazamiento forzado en Colombia (2015). Available at: http://babel.banrepcultural.org/cdm/ref/collection/p17054coll2/id/63 (Accessed June 15, 2022).
40. Wabgou M, Vargas D, Carabalí J. International migrations in Colombia. Investig Desarro (2012) 20:142–67.
42. DANE. Proyecciones de población (2018). Available at: https://www.dane.gov.co/index.php/estadisticas-por-tema/demografia-y-poblacion/proyecciones-de-poblacion (Accessed June 15, 2022).
43. Cereb N, Kim HR, Ryu J, Yang SY. Advances in DNA sequencing technologies for high resolution HLA typing. Hum Immunol (2015) 76:923–7. doi: 10.1016/j.humimm.2015.09.015
44. Lange V, Böhme I, Hofmann J, Lang K, Sauter J, Schöne B, et al. Cost-efficient high-throughput HLA typing by MiSeq amplicon sequencing. BMC Genom (2014) 15:63. doi: 10.1186/1471-2164-15-63
45. Excoffier L, Lischer HEL. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and windows. Mol Ecol Resour (2010) 10:564–7. doi: 10.1111/j.1755-0998.2010.02847.x
46. Schäfer C, Schmidt AH, Sauter J. Hapl-o-Mat: open-source software for HLA haplotype frequency estimation from ambiguous and heterogeneous data. BMC Bioinform (2017) 18:1–10. doi: 10.1186/s12859-017-1692-y
47. Páez-Gutiérrez IA, Hernández-Mejía DG, Vanegas D, Camacho-Rodríguez B, Perdomo-Arciniegas AM. HLA-a, -b, -c, -DRB1 and -DQB1 allele and haplotype frequencies of 1463 umbilical cord blood units typed in high resolution from bogotá, Colombia. Hum Immunol (2019) 80:425–6. doi: 10.1016/j.humimm.2019.03.006
48. Del Río-Ospina L, Camargo M, Soto-De León SC, Robayo-Calderón KL, Garzón-Ospina D, Patarroyo ME, et al. Using next-generation sequencing for characterising HLA-DRB1 and DQB1 loci in a cohort of Colombian women. HLA (2019) 94:425–34. doi: 10.1111/tan.13672
49. Arias YR, Osorio-Arango K, Bayona B, Ercilla G, Beltrán-Durán M. Determination of HLA -a, -b, -DRB1 polymorphism in brain dead organ donors representative of the Colombian general population, 2007-2014. Biomédica (2017) 37:184–90. doi: 10.7705/biomedica.v37i2.3263
50. Arrunategui AM. Frecuencias alélicas, genotípicas y haplotípicas del sistema HLA clase I y II en donantes en una población del suroccidente colombiano. Acta Méd Colomb (2013) 38:16–21. doi: 10.36104/amc.2013.69
51. Flórez O, Cabrales CC, Pinto JA, Ramírez G, Flórez JA, Capella A, et al. HLA-matched donor-recipient combinations and kidney transplant probabilities in a specific colombian population. Rev Cienc Salud (2016) 14:147–60. doi: 10.12804/revsalud14.02.2016.01
52. Reyes HO, Manrique A, Quintanilla S, Peña A. Polimorfismos del sistema HLA (loci a*, b* y DRB1*) en población colombiana. Nova (2007) 5:25. doi: 10.22490/24629448.369
53. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. Identification of a novel HLA-DRB1 null allele, HLA-DRB1*13:298N, in two siblings from Colombia. HLA (2022) 99:659–60. doi: 10.1111/tan.14564
54. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. Identification of five novel HLA-b alleles in donors from the Colombian bone marrow donor registry. HLA (2022) 99:380–2. doi: 10.1111/tan.14516
55. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. Description of HLA-DRB1*14:02:09, a novel HLA allele identified in a Colombian donor. HLA (2022) 99:405–7. doi: 10.1111/tan.14519
56. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. The novel HLA-A*30:172 allele identified in a donor from the Colombian bone marrow donor registry. HLA (2022) 100:155–6. doi: 10.1111/tan.14627
57. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. The novel HLA-A*24 allele, HLA-A*24:487, identified in three unrelated bone marrow donors in Colombia. HLA (2021) 98:538–40. doi: 10.1111/tan.14432
58. Hernández DG, Ramírez NC, Martínez MM, Cendales PA, Camacho BA. Characterization of the novel HLA-DRB1*04:315 allele by next-generation sequencing. HLA (2022) 99:654–5. doi: 10.1111/tan.14534
59. Li H, Liu D, Li X. HLA-DPB1 and Epstein-Barr virus gp42 protein jointly contribute to the development of Hodgkin lymphoma. Transl Cancer Res (2020) 9:4424–32. doi: 10.21037/tcr-20-2070
60. Jiang L, Jiang D, Han Y, Shi X, Ren C. Association of HLA-DPB1 polymorphisms with rheumatoid arthritis: a systemic review and meta-analysis. Int J Surg (2018) 52:98–104. doi: 10.1016/j.ijsu.2018.01.046
61. Ou G, Liu X, Xu H, Ji X, Liu X, Wang J. Variation and expression of HLA-DPB1 gene in HBV infection. Immunogenetics (2021) 73:253–61. doi: 10.1007/s00251-021-01213-w
62. Ou G, Liu X, Jiang Y. HLA-DPB1 alleles in hepatitis b vaccine response: a meta-analysis. Medicine (2021) 100:e24904. doi: 10.1097/MD.0000000000024904
63. Yao Y, Yang H, Shi L, Liu S, Li C, Chen J, et al. HLA class II genes HLA-DRB1, HLA-DPB1, and HLA-DQB1 are associated with the antibody response to inactivated Japanese encephalitis vaccine. Front Immunol (2019) 10:428. doi: 10.3389/fimmu.2019.00428
64. Jaime-Pérez JC, Cancela-Murrieta CO, Salazar-Cavazos L, Aguilar- Calderón PE, Salazar-Riojas R, Méndez-Ramírez N, et al. Impact of HLA-DPB1 matching on clinical outcomes after haploidentical-related hematopoietic stem cell transplantation. Rev Invest Clin (2020) 72:69–79. doi: 10.24875/ric.19003215
65. Rutten CE, van Luxemburg-Heijs SAP, van der Meijden ED, Griffioen M, Oudshoorn M, Willemze R, et al. HLA-DPB1 mismatching results in the generation of a full repertoire of HLA-DPB1-specific CD4+ T cell responses showing immunogenicity of all HLA-DPB1 alleles. Biol Blood Marrow Transplant (2010) 16:1282–92. doi: 10.1016/j.bbmt.2010.03.018
66. Ducreux S, Dubois V, Amokrane K, Yakoub-Agha I, Labalette M, Michallet M, et al. HLA-DRB3/4/5 mismatches are associated with increased risk of acute GVHD in 10/10 matched unrelated donor hematopoietic cell transplantation. Am J Hematol (2018) 93:994–1001. doi: 10.1002/ajh.25133
67. Olivares FM, Madero JM, Casas-Vargas A, Montoya SZ, Medellín DS, Gusmão L, et al. Contrasting the ancestry patterns of three distinct population groups from the northernmost region of south America. Am J Phys Anthropol (2020) 173:437–47. doi: 10.1002/ajpa.24130
68. Llanes A, Ortiz L, Moscoso J, Gutiérrez G, Blake E, Restrepo CM, et al. HLA allele and haplotype frequencies in the Panamanian population. Hum Immunol (2021) 82:5–7. doi: 10.1016/j.humimm.2020.11.006
69. del Pilar Fortes M, Gill G, Paredes ME, Gamez LE, Palacios M, Blanca I, et al. Allele and haplotype frequencies at human leukocyte antigen class I and II genes in venezuela’s population. Ann Biol Clin (2012) 70:175–81. doi: 10.1684/abc.2012.0663
70. Galarza JM, Barquera R, Álvarez AMT, Zaragoza DIH, Sevilla GP, Tamayo A, et al. Genetic diversity of the HLA system in human populations from the Sierra (Andean), oriente (Amazonian) and Costa (Coastal) regions of Ecuador. Hum Immunol (2018) 79:639–50. doi: 10.1016/j.humimm.2018.06.004
71. Montero-Martín G, Mallempati KC, Gangavarapu S, Sánchez-Gordo F, Herrero-Mata MJ, Balas A, et al. High-resolution characterization of allelic and haplotypic HLA frequency distribution in a Spanish population using high-throughput next-generation sequencing. Hum Immunol (2019) 80:429–36. doi: 10.1016/j.humimm.2019.02.005
72. Torres L, da Silva Bouzas LF, Almada A, de Sobrino Porto LCM, Abdelhay E. Distribution of HLA-a, -b and -DRB1 antigenic groups and haplotypes from the Brazilian bone marrow donor registry (REDOME). Hum Immunol (2017) 78:602–9. doi: 10.1016/j.humimm.2017.08.002
73. Perusco A, Gamba C, Galarza P, Rojas F, Kalapodis A, Onofri A, et al. HLA antigenic and haplotype frequencies estimated in hematopoietic progenitor cell donors from Argentina. Transplant Proc (2014) 46:3064–7. doi: 10.1016/j.transproceed.2014.07.029
74. Souto DF, Rosello J, Lazo L, Veloso F, Gamba C, Kuperman S, et al. HLA diversity in the Argentinian umbilical cord blood bank: frequencies according to donor's reported ancestry and geographical distribution. Sci Rep (2021) 11:1–10. doi: 10.1038/s41598-021-83282-1
Keywords: allele frequency, Bone Marrow Registry, Colombia, HLA, next-generation sequencing
Citation: Hernández-Mejía DG, Páez-Gutiérrez IA, Dorsant Ardón V, Camacho Ramírez N, Mosquera M, Cendales PA and Camacho BA (2023) Distributions of the HLA-A, HLA-B, HLA-C, HLA-DRB1, and HLA-DQB1 alleles and haplotype frequencies of 1763 stem cell donors in the Colombian Bone Marrow Registry typed by next-generation sequencing. Front. Immunol. 13:1057657. doi: 10.3389/fimmu.2022.1057657
Received: 29 September 2022; Accepted: 19 December 2022;
Published: 09 January 2023.
Edited by:
Aziz Alami Chentoufi, University of Pretoria, South AfricaReviewed by:
Sara Lai, R.Binaghi Hospital, ItalyUma Kanga, All India Institute of Medical Sciences, India
Copyright © 2023 Hernández-Mejía, Páez-Gutiérrez, Dorsant Ardón, Camacho Ramírez, Mosquera, Cendales and Camacho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paola Andrea Cendales, cGFjZW5kYWxlc0BpZGNiaXMub3JnLmNv