
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 24 October 2022
Sec. Multiple Sclerosis and Neuroimmunology
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.1033774
This article is part of the Research Topic Understanding and Targeting Neuro-Immune Interactions within Disease and Inflammation View all 8 articles
 Pedro L. Katayama1*
Pedro L. Katayama1* Isabela P. Leirão1
Isabela P. Leirão1 Alexandre Kanashiro2
Alexandre Kanashiro2 José V. Menani1
José V. Menani1 Daniel B. Zoccal1
Daniel B. Zoccal1 Débora S. A. Colombari1
Débora S. A. Colombari1 Eduardo Colombari1
Eduardo Colombari1The idea that the nervous system communicates with the immune system to regulate physiological and pathological processes is not new. However, there is still much to learn about how these interactions occur under different conditions. The carotid body (CB) is a sensory organ located in the neck, classically known as the primary sensor of the oxygen (O2) levels in the organism of mammals. When the partial pressure of O2 in the arterial blood falls, the CB alerts the brain which coordinates cardiorespiratory responses to ensure adequate O2 supply to all tissues and organs in the body. A growing body of evidence, however, has demonstrated that the CB is much more than an O2 sensor. Actually, the CB is a multimodal sensor with the extraordinary ability to detect a wide diversity of circulating molecules in the arterial blood, including inflammatory mediators. In this review, we introduce the literature supporting the role of the CB as a critical component of neuroimmune interactions. Based on ours and other studies, we propose a novel neuroimmune pathway in which the CB acts as a sensor of circulating inflammatory mediators and, in conditions of systemic inflammation, recruits a sympathetic-mediated counteracting mechanism that appears to be a protective response.
The nervous and immune systems interact to maintain physiological functions and to regulate pathological processes. These interactions occur in a bidirectional fashion – the nervous system can modulate the immune system and the immune cells and molecules can affect nervous system functions (1–7). Our understanding of neuroimmune interactions has substantially increased in the last years due to advancing technologies and combined efforts from the neuroscience, immunology, and physiology fields. It is currently known that local (i.e., within organs) and systemic (i.e., inter-organs) neuroimmune interactions govern many physiological functions as well as contribute to the development and maintenance of several diseases (8–11). Very illustrative examples of local neuroimmune interactions are those occurring within the gut, where its intrinsic nervous system (i.e., enteric nervous system) continuously modulates the activity of gut-resident immune cells to provide host defense against pathogens (12, 13). Differently, systemic neuroimmune interactions involve nervous and immune cells across different organs and can depend on central nervous system processing. For example, during acute systemic inflammation, the increased levels of circulating inflammatory mediators activate brainstem areas that are able to generate a counteracting protective response through changes in autonomic (i.e., parasympathetic and sympathetic) outflows that, in turn, modulate the function of immune cells within different peripheral organs to resolve inflammation (3, 7, 14–16).
In the latter example, because many immune cells and inflammatory molecules cannot readily cross the blood-brain barrier (17), some mechanisms have been identified to explain how brainstem areas could be activated in response to acute peripheral inflammation. In this context, the vagus nerve (X cranial nerve), a major component of the parasympathetic nervous system, was found to play an important role. The vagus is a mixed nerve, constituted by both afferent (from the body to the brain) and efferent (from the brain to the body) fibers, in a proportion of approximately 80% and 20%, respectively (18). Vagal afferent fibers carry important visceral sensory information from most peripheral organs to the central nervous system and, therefore, are essential players in the communication between the body and the brain (19). Regarding peripheral inflammation, several lines of evidence have indicated that vagal afferents are excited by circulating inflammatory mediators and relay this information to the central nervous system (1, 14, 20, 21). Indeed, it has been extensively reported that endogenous or exogenous stimulation of vagal afferents suppresses inflammation, likely by activating parasympathetic-related brainstem areas that generate a vagal efferent outflow to modulate peripheral immune cells (2, 7, 22–25).
Nevertheless, although the roles of vagal afferents as peripheral sensors of inflammation and vagal efferents as modulators of immune cells are well documented, much less is known about alternative, vagal-independent mechanisms in the context of systemic inflammation. An increasing body of new evidence has shown that several neuroimmune mechanisms do not depend on the vagus nerve or could compensate for its absence (3, 5, 16, 26). For instance, the intravenous administration of lipopolysaccharide (LPS) promoted systemic inflammation and increased splanchnic sympathetic nerve activity (3). This heightened sympathetic activity appears to be an anti-inflammatory response since in animals subjected to splanchnic sympathetic denervation, the LPS-induced inflammation was exacerbated, as reflected by higher levels of tumor necrosis factor-alpha (TNF-α) in the plasma (3). It is important to note that, although this effect was attributed to the lack of the inhibitory role played by splanchnic efferents on the splenic production of TNF-α (3), the absence of splanchnic afferents could have also contributed to exacerbating inflammation. Afferent fibers within the splanchnic nerves convey a great variety of sensory (mechanical, chemical, and noxious) information from visceral organs to the brain and, importantly, can be sensitized by inflammatory mediators (27, 28). Therefore, in the study of Martelli et al. (3), a contribution of splanchnic afferents to body-brain communication during LPS-induced systemic inflammation cannot be excluded. Another remarkable finding of the study was that bilateral vagotomy did not affect the LPS-induced increase in splanchnic sympathetic nerve activity nor the LPS-induced increase in TNF-α levels (3). Collectively, these findings support the existence of a neuroimmune mechanism that detects systemic inflammation and increases sympathetic outflow to modulate inflammation independently of vagal afferents (as peripheral sensors of inflammatory mediators) and vagal efferents (as neural modulators of peripheral immune cells). These results (3) also support a role of the sympathetic nervous system as a brain output in controlling inflammation (5, 16, 29, 30). However, a question remains: what could be the sensing mechanisms responsible for detecting systemic inflammation and driving the activation of brain circuits that increase sympathetic outflow to modulate the function of peripheral immune cells when the vagus nerves are absent?
Here, we review the literature supporting that the carotid bodies, a pair of small sensory organs located in the neck, are crucial players in body-brain communications in the context of inflammation. The carotid body (CB) is classically known as the primary sensor of the oxygen (O2) levels in the arterial blood, by sensing hypoxemia and activating brainstem autonomic areas to promote adequate cardiorespiratory adjustments to maintain homeostasis (31–33). However, it is now clear that the CB is much more than a hypoxemia/hypoxia sensor. Instead, the CB can monitor the levels of several molecules in the arterial blood (34, 35). This multisensorial ability confers to the CB the extraordinary capacity for integrating different systems, including interactions between the nervous and the immune systems. For instance, we recently found that the CB cells detect increased levels of TNF-α in the circulation and drives a sympathetic-mediated response to suppress systemic inflammation (26). Based on this and other studies, we propose that the CB is a key player in neuroimmune interactions and might be involved in the pathophysiology of inflammation-mediated diseases, representing a potential novel target for the development of new therapies.
Before going into details about the role of the CB in neuroimmune interactions, a brief overview of CB anatomy and function is provided in this and the following section. The CB is a small paired organ located in the carotid bifurcations, the point where the common carotid artery divides into external and internal carotid arteries to supply blood to the brain (33, 34, 36, 37). This anatomical location is strategic: since the CB is the main sensor of the O2 levels in the body, one of its primordial tasks is monitoring the composition of the arterial blood flowing into the brain, which functioning is critically dependent on adequate O2 supply (35, 38). The CB is comprised by glomus cells (type I) and sustentacular cells (type II). The glomus cells are the most predominant and form clusters wrapped by sustentacular cells (33, 39). The glomus cells are responsible for detecting low levels of O2 in the arterial blood (hypoxemia), transducing this chemical information into neural signals that reach the central nervous system. In brief, when the partial pressure of O2 in the arterial blood falls, the CB glomus cells depolarize and release neurotransmitters that excite terminal afferents of the carotid sinus nerve (CSN) (34, 37, 38, 40). The CSN, which is a branch of the glossopharyngeal nerve (IX cranial nerve), conveys the signals from the CB to the nucleus tractus solitarius (NTS) in the brainstem (32, 37, 41). The NTS neurons, in turn, sends projections to brainstem areas related to the control of autonomic activity to promote cardiorespiratory adjustments to counteract hypoxemia and ensure the adequate delivery of O2 to the brain and body (32, 37, 42, 43).
The cardiorespiratory responses to hypoxemia include increases in arterial blood pressure and pulmonary ventilation and a decrease in heart rate (44, 45). The increase in arterial blood pressure results from a rapid elevation in sympathetic activity, driven mainly by a CB-mediated activation of NTS excitatory projections to the rostral ventrolateral medulla (RVLM), where most pre-sympathetic neurons are found (32, 46, 47). Notably, CB-mediated sympathetic effects during hypoxia are mostly excitatory, increasing lumbar, renal, and splanchnic sympathetic nerve activities (26, 48), only decreasing brown adipose tissue sympathetic nerve activity (49). Interestingly, CB stimulation by hypoxia also activates parasympathetic-controlling brainstem areas such as the nucleus ambiguus (NA) to increase parasympathetic activity resulting in bradycardia and, possibly, other effects (50–52). These observations demonstrate that the CB has a particular capacity for modulating both sympathetic and parasympathetic arms of autonomic outflows, impacting the function of different organs/systems. In the next section, we introduce the current concept that the CB contributes to regulating several body functions in a number of different conditions rather than only during hypoxia.
Over the last 30 years, it has been increasingly discovered that the sensory function of the CB is not limited to hypoxia – besides those, the CB can detect an impressive variety of molecules (or their absence) in the circulation. For example, the CB is able to sense the plasma levels of angiotensin II (53), endothelin (54), leptin (55), cytokines (26, 56–59), epinephrine (60, 61), sodium chloride (62), glucose (63–65), insulin (66), and glucagon like peptide-1 (67) in the arterial blood. This very unique ability supports the current view that the CB is a multimodal sensory organ that monitors blood composition and informs the central nervous system on whether a given molecule is at low or high levels in the circulation (Figure 1). The central nervous system, in turn, organize the reflex adjustments, mainly through changes in autonomic outflows to regulate various body functions (34, 69).
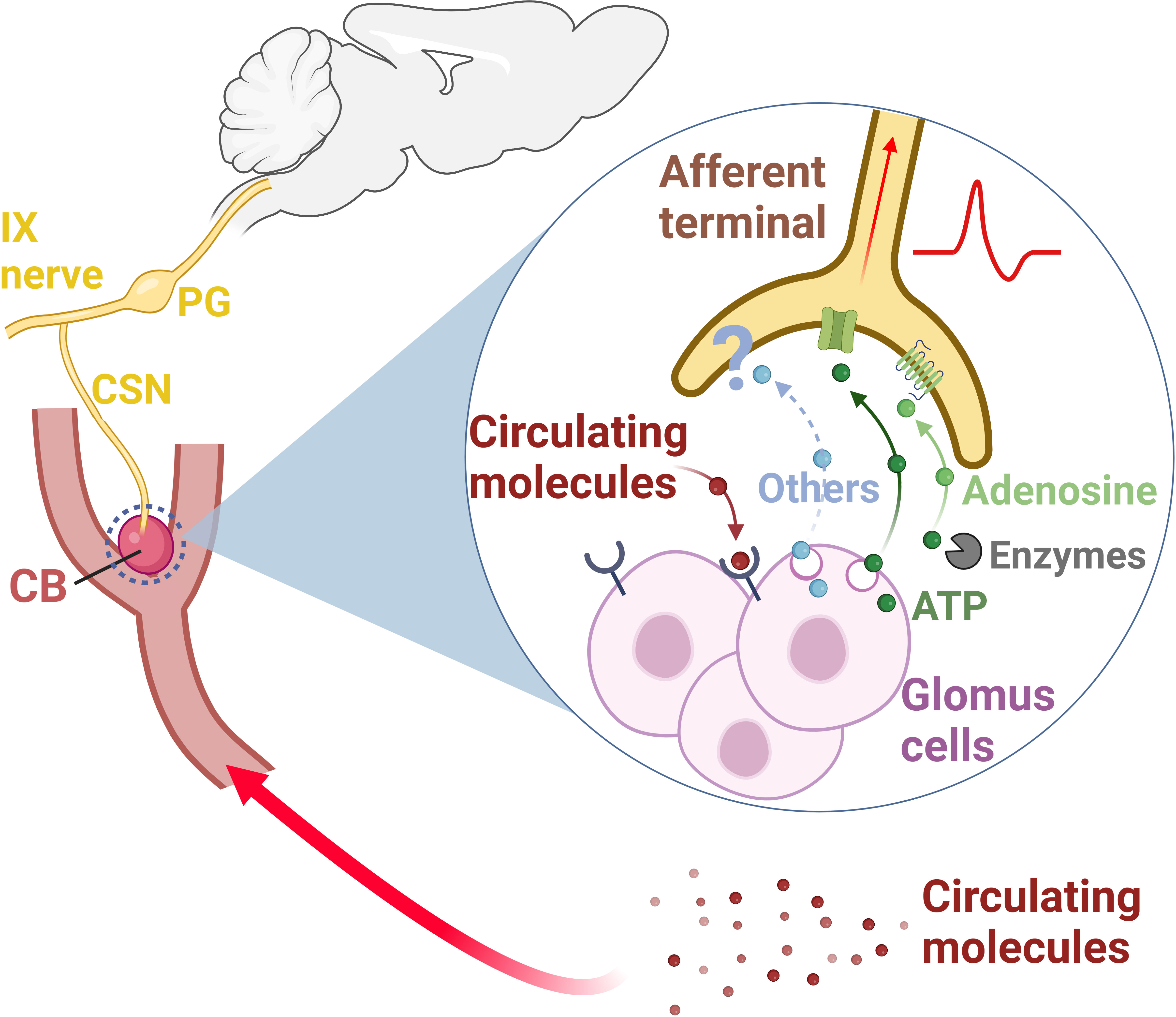
Figure 1 The carotid body (CB) is a sensory organ located at the carotid bifurcation (33, 34, 36). The carotid sinus nerve (CSN), a branch of the IX nerve (glossopharyngeal nerve), provides the sensory innervation to the CB (34, 36). Pseudo-unipolar neurons with cell bodies located within the petrosal ganglion (PG) convey the sensory information from the CB to the central nervous system (32, 34, 36, 37). This sensory information is multimodal, since the CB is responsive not only to changes in the partial pressure of oxygen in the arterial blood, but also to changes in the levels of several circulating molecules (epinephrine, angiotensin II, cytokines, endothelin, glucagon like peptide-1, glucose, insulin, leptin, and sodium chloride) (26, 53–55, 57–59, 61–64, 66, 67). Once depolarized, CB glomus cells release neurotransmitters, primarily ATP (39, 68). ATP binds to P2X2/P2X3 receptors in the afferent terminals of the CSN, generating action potentials which are propagated to the central nervous system (33, 38, 39, 68). Some of the ATP released by glomus cells is converted into adenosine by extracellular enzymes (NTPDase2,3 and ecto-5-nucleotidase) (38). Adenosine acts on A2a receptors on CSN afferent terminals, generating action potentials, and contributing to CB signaling to the central nervous system (38–40). Other neurotransmitters (Others) such as acetylcholine, dopamine, and serotonin, have been also found to modulate CSN activity (33, 37, 39). CB, carotid body; CSN, carotid sinus nerve; IX nerve, glossopharyngeal nerve; PG, petrosal ganglion. Created with BioRender.com.
One interesting example of a CB-mediated integrative homeostatic mechanism can be observed during hypoglycemia. Previous in vitro investigations have shown that CB glomus cells are sensitive to low levels of glucose (63, 64) encouraging later in vivo studies to explore the potential contribution of the CB in the homeostatic responses to hypoglycemia (61, 70). One of these studies, performed in humans, demonstrated that the hypoglycemia-induced acute release of counter-regulatory hormones that promote glycogenolysis and gluconeogenesis such as epinephrine, norepinephrine, cortisol, and glucagon is impaired by inhibition of the CB (70). The lower levels of epinephrine and norepinephrine in the plasma reflect a reduced sympathetic activation which, in turn, could have partially accounted for the lower levels of cortisol and glucagon when the CB is silenced (70). These findings illustrate how the CB plays an essential role in integrating different organs and systems. In this case, the CB acts as a sensor of blood glucose levels and coordinates a homeostatic response involving the sympathetic nervous and endocrine systems (70). In the following sections of this review, we focus on previous and new evidence indicating that the CB cells are also important for linking the nervous and the immune systems.
In this section, we examine the growing body of evidence supporting that the CB is a sensor of peripheral inflammation and plays a pivotal role in recruiting a sympathetic-mediated anti-inflammatory response. As a starting point, it should be noted that the presence of receptors that allow for the recognition of pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), and inflammatory mediators was reported in CB glomus cells and in PG neurons that provide CB sensory innervation. More precisely, it has been demonstrated that the mammalian CB and its innervating sensory neurons express toll-like receptors (TLR) 1 and 4, and receptors for inflammatory ligands such as TNF-α, interleukin 1-beta (IL-1β), interleukin 6 (IL-6), and lysophosphatidic acid (LPA) (26, 56, 71–75). Additional supportive evidence comes from studies showing that the CB is intrinsically responsive to several inflammatory mediators (56–58). For instance, whole-cell patch-clamp and calcium imaging experiments in cultured glomus cells demonstrated that IL-1β depolarizes the CB, inhibiting outward potassium currents and promoting calcium influx (58). In a recent study, it was shown that various pro-inflammatory cytokines (IL-4, IL-5, IL-13, IL-1β, IL-6, and TNF-α) and other inflammatory mediators (eotaxin and LPA) increase CSN activity in an isolated CB/CSN preparation (57). Notably, the same research group previously demonstrated that the LPA-induced CB activation stimulates phrenic and vagus nerve activities in an in situ decerebrated rat preparation (56), confirming that inflammatory mediators can directly activate the CB that, in turn, stimulates motor/autonomic outflows.
While these essential studies demonstrated that the CB has the machinery to intrinsically detect and respond to inflammatory mediators, other studies provided indirect but equally important evidence about the role of the CB in neuroimmune interactions, specifically in the context of systemic inflammation (71, 76, 77). In cats, the intravenous administration of LPS, resulting in systemic inflammation with high levels of various circulating pro-inflammatory cytokines, potently increased CSN discharge and caused tachypnea, tachycardia, and hypotension (71). Interestingly, the bilateral CSN section abolished the LPS-induced increase in respiratory frequency, indicating that the CB activates central respiratory pathways and stimulates breathing during systemic inflammation (71). A subsequent study provided further evidence that the CB is involved in brain-body communication during LPS-induced sepsis (76). The authors demonstrated that the intravenous administration of LPS in rats activates neurons in the NTS, the first relay site for CB afferents (76). Remarkably, LPS-induced NTS activation was profoundly suppressed in animals with bilateral CB denervation, strongly indicating that the CB is a crucial player in central nervous system activation during systemic inflammation (76). Furthermore, a later study from the same research group, suggested that the CB has a protective role during LPS-induced sepsis since bilateral CB denervation resulted in exacerbated plasma TNF-α, blunted plasma corticosterone, and faster progression to multiple organs dysfunction in comparison to rats with intact CBs (77).
Although the presented evidence seems sufficient to acknowledge the involvement of the CB in neuroimmune interactions, several questions still remain. Since intravenous administration of LPS increases the circulating levels of several inflammatory mediators such as TNF-α, IL1β, and IL-6 (78), what molecule (s) is (are) actually acting on the CB in vivo? What are the connections of the NTS neurons activated by CB afferents during inflammation? Is the CB-mediated activation of NTS neurons a counteracting response to systemic inflammation?
Therefore, in a recent study, we sought to further dissect the role of the CB in neuroimmune interactions (26). More specifically, we investigated if the CB could detect increased levels of TNF-α in the circulation and activate central autonomic pathways to potentially modulate inflammation. We decided to focus on TNF-α because its type I receptor (TNFR1) was already reported to be found in CB glomus cells of several mammalian species (including rats) (71, 73, 79) and, also, because TNF-α is a well-known critical mediator of inflammation (80). We first examined and confirmed that the TNF-α receptor type I is expressed in the CB glomus cells of rats at mRNA and protein levels (26). Next, we found that the intravenous administration of TNF-α increased CSN afferent discharge (26), suggesting that the CB can detect increased levels of TNF-α in the circulation. Since the first central synapse of CB afferents occurs in the NTS (32, 81), and because CB stimulation by hypoxia activates monosynaptic excitatory projections from NTS to RVLM, increasing sympathetic outflow (32, 46), we hypothesized that the observed TNF-α-induced increase in CSN afferent activity could stimulate this pathway (26). We found that intravenous TNF-α activated a high number of RVLM-projecting NTS glutamatergic neurons and that this activation was dependent on CB input since the number of activated neurons was drastically reduced in rats previously subjected to bilateral CB ablation (26). These results suggest that the CB detects circulating TNF-α and activates a central sympathoexcitatory pathway. Consistent with the observation that circulating TNF-α activates the CB to recruit a NTS-RVLM sympathoexcitatory pathway, we found that systemic TNF-α increased splanchnic sympathetic nerve activity, and importantly, CB ablation strongly attenuated this response (26). This observation further supports the role of the CB as a sensor of circulating TNF-α and a key player in body-brain communication in this model. Finally, because the splanchnic sympathetic nerve was shown to exert an anti-inflammatory and protective role during systemic inflammation (3, 15, 16), we hypothesized that the TNF-α-induced activation of the sympathetic circuits and splanchnic sympathetic nerve activity would counteract inflammation. Confirming this assumption, the levels of cytokines measured in the plasma and in the spleen 2 hours after the TNF-α administration were significantly increased in animals previously subjected to CB removal or splanchnic sympathetic denervation compared to sham animals (26). In summary, our results suggest a novel neuroimmune mechanism in which the CB acts as a sensor of peripheral inflammation, detecting increased levels of circulating TNF-α and communicating with the brain to activate a counteracting anti-inflammatory reflex mediated by the splanchnic sympathetic nerve. A schematic illustration of the novel mechanism is presented in Figure 2.
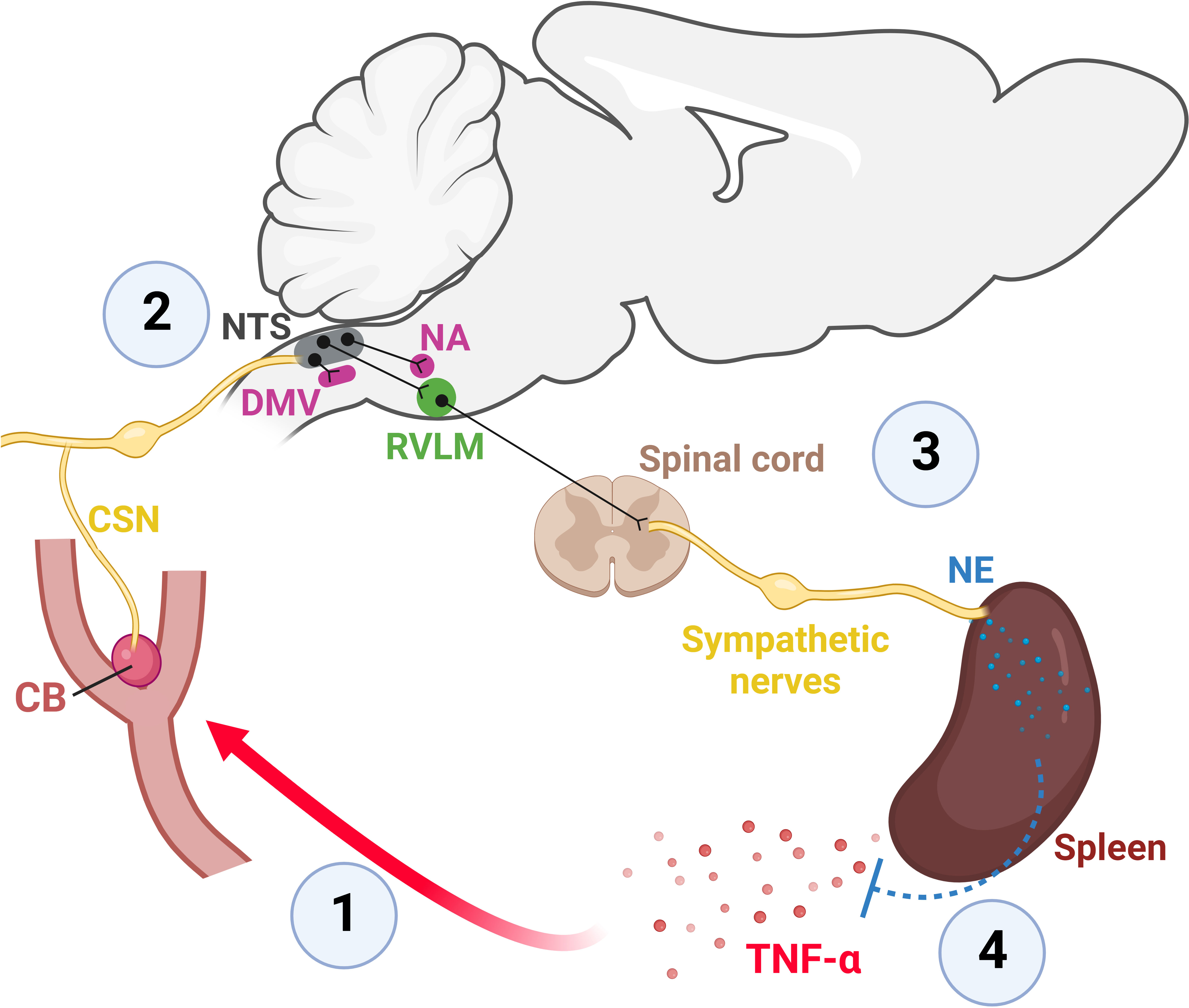
Figure 2 The CB is a sensor of peripheral inflammation and initiates a sympathetic-mediated anti-inflammatory response (26). 1: Elevated levels of TNF-α in the blood depolarizes CB glomus cells which widely express TNF-α receptors type I (26, 71–73). Depolarization of CB glomus cells by TNF-α generates action potentials that propagate along axons within the CSN towards the brainstem (26). 2: The first central synapse of CB-originated axons occurs in the NTS, a major integrative brainstem region that receives the sensory information from peripheral organs and projects to several brainstem autonomic areas that control parasympathetic (DMV and NA) and sympathetic (RVLM) functions (32, 41, 46, 50, 52). 3: The TNF-α-induced activation of the CB-NTS-RVLM circuit increases the activity of the splanchnic sympathetic nerve which innervates the celiac ganglia, from where the splenic nerve originates and projects to the spleen, releasing norepinephrine (NE) (26). 4: The release of NE into the spleen reduces both splenic and plasmatic levels of TNF-α (26). CB, carotid body; CSN, carotid sinus nerve; DMV, the dorsal motor nucleus of the vagus; NA, nucleus ambiguus; NTS, nucleus tractus solitarius; NE, norepinephrine; RVLM, rostral ventrolateral medulla; TNF-α, tumor necrosis factor-alpha. Created with BioRender.com.
The discovery of this novel neuroimmune mechanism raises numerous possibilities and questions. As well pointed out by a recent report (82): could other sympathetic nerves (besides the splanchnic) and/or premotor sympathetic areas (besides the RVLM) also contribute to regulating inflammation? In addition, we also add the following questions: What is the role of this mechanism in chronic inflammatory conditions? Are CB-mediated activation of NTS projections to brainstem parasympathetic nuclei (DMV and NA) activated during inflammation? Does the parasympathetic nervous system also play a role in this neuroimmune mechanism? Does chronic inflammation cause CB-mediated sympathetic overactivation in disease states? Although further investigations are needed to answer all the raised questions described above, in the next section of this review, we briefly discuss the potential role of inflammation in the pathological overactivation of the CB observed in several diseases.
Because of its multisensory ability and integrative role, it is somewhat expected that a CB dysfunction could impact the activity of different organs/systems and, thus, contribute to various diseases. Numerous pre-clinical and clinical studies have shown that CB dysfunction, usually characterized by its overactivity, plays a role in hypertension (83–85), heart failure (86, 87), apneas of prematurity (88, 89), and metabolic diseases (90, 91). In hypertension and heart failure, for example, a chronic CB overactivation has been associated with the sustained activation of the sympathetic nervous system, which contributes to the development and maintenance of these conditions (83, 87, 92–95).
Over the last decade, the surgical removal of the CB has been considered a therapeutic option to overcome this CB-mediated sympathetic overactivation in hypertension (83, 85, 92, 96) and heart failure (86, 95). Although the outcomes of these studies were very promising, concerns about the safety of removing the CBs were raised (45, 97). In spontaneously hypertensive rats, the bilateral CB removal abolished the pressor responses to hypoxia and exacerbated the pressor and respiratory responses to hypercapnia, indicating an impaired capacity for regulating cardiorespiratory functions in CB-ablated animals (45). In addition, based on the supposed protective role of the CB during systemic inflammation and sepsis (26, 77), CB ablation could result in catastrophic effects if the individual is exposed to these conditions.
In this context, pharmacological approaches have been described (93, 98) as promising options for reducing CB overactivity. However, to the best of our knowledge, there are no reports considering the targeting of inflammatory signaling within the CB in diseases such as hypertension and heart failure. Since a chronic state of inflammation is a common feature in these conditions (99, 100) and because inflammation-related molecules activate the CB (26, 57), we hypothesize that chronic inflammation contributes to the excessive CB activation observed in some diseases and thus, propose that targeting inflammation within the CB might represent a new therapeutic approach. However, studies are needed to test this hypothesis.
If, on one side, a great effort has been made to suppress CB activity in chronic conditions such as hypertension and heart failure, on the other side, much less is known about strategies to enhance CB activity that could be favorable in some situations such as in acute systemic inflammation (26, 77, 101, 102). Preclinical studies conducted in rodents have provided exciting evidence for considering the electrical stimulation of the CB to counteract acute systemic inflammation (101, 102). One of these studies employed a technique that allows the electrical stimulation of the carotid sinus/CSN in unanesthetized rats (103) and found that CB activation attenuates LPS-induced systemic inflammation likely by recruiting parasympathetic- and sympathetic-mediated mechanisms (101). Another study demonstrated that CSN stimulation in mice efficiently reduced LPS-induced inflammation by activating the hypothalamus-pituitary-adrenal axis, promoting corticosterone release, which, in turn, suppressed the activity of myeloid cells (102). Nevertheless, some questions remain. First, which type(s) of fiber(s) in the CSN (A-fibers, C-fibers, or both) mediate the observed anti-inflammatory effects following CB/CSN electrical stimulation in the abovementioned studies? Second, could different electrical stimulation parameters selectively activate CSN A-fibers, C-fibers, or both? Last, would the activation of specific CSN fiber types promote differential effects on the control of inflammation? For example, studies conducted on electrical stimulation of the vagus nerve have demonstrated that specific stimulation parameters can recruit different fiber types (104, 105). Moreover, it has been shown that specific parameters for electrical stimulation of the vagus nerve results in differential effects on circulating cytokines (24). Therefore, although the outcomes of CB activation in rodents to modulate inflammation seem encouraging, the translation of electrical stimulation of the CB/CSN to the clinical scenario still requires further investigation.
Besides the need for more robust evidence on the mechanisms underlying the electrical activation of the CB, as discussed above, safety concerns are probably the main challenge for implementing this approach in clinical settings. A surgical, invasive approach is necessary to access the CB and the CSN. Since the CB and surrounding areas are highly vascularized and innervated (34, 106), potential inaccuracies during the surgical procedure could result in serious consequences. In this context, ultrasound-based neural stimulation could be explored as a non-invasive alternative for activating the CB/CSN. Recent studies have shown successful applications of ultrasound stimulation for targeting neural pathways within specific organs, such as the spleen and the liver, modulating inflammation and glucose homeostasis, respectively (107, 108). A challenge for this, however, may rely on the fact that the carotid baroreceptor fibers and CB fibers are in close proximity and run together within the CSN (34, 109), making it very difficult to specifically target one and not the other.
In this review, we highlight the CB as a unique organ with extraordinary capacities for sensing a great variety of molecules and integrating different organs/systems. We focus on the role of the CB in mediating the integration between the nervous and the immune systems. In this context, we introduce a novel mechanism of neuroimmune interaction in which the CB acts as a sensor of inflammatory ligands in the circulation and recruits central sympathetic networks that counteract inflammation. Furthermore, based on a growing body of preclinical and clinical research, we propose new perspectives on managing inflammatory diseases by targeting the CB.
It should be noted that although most of the studies discussed in this review reported a sympathetic-mediated suppression of immune responses (3, 5, 16, 26), the sympathetic nervous system, in some conditions, can actually activate the immune system (110–112). Therefore, despite the potential therapeutic possibilities highlighted in the present study seem promising, there is still a lot to explore regarding the intricate interactions between the nervous and immune systems.
PK conceived and designed the manuscript. PK, IL, AK, JM, DZ, DC, and EC drafted and revised the manuscript. PK prepared the figures. All authors read and approved the final version of the manuscript.
This work was supported by the Sao Paulo Research Foundation (FAPESP; grants #2019/11196-0, #2015/23467-7, and #2019/24154-3) and PROPG-UNESP. Figures were created with BioRender.com.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Watkins LR, Goehler LE, Relton JK, Tartaglia N, Silbert L, Martin D, et al. Blockade of interleukin-1 induced hyperthermia by subdiaphragmatic vagotomy: evidence for vagal mediation of immune-brain communication. Neurosci Lett (1995) 183:27–31. doi: 10.1016/0304-3940(94)11105-R
2. Borovikova LV, Ivanova S, Zhang M, Yang H, Botchkina GI, Watkins LR, et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature (2000) 405:458–62. doi: 10.1038/35013070
3. Martelli D, Yao ST, Mckinley MJ, Mcallen RM. Reflex control of inflammation by sympathetic nerves, not the vagus. J Physiol (2014) 592:1677–86. doi: 10.1113/jphysiol.2013.268573
4. Talbot S, Abdulnour REE, Burkett PR, Lee S, Cronin SJF, Pascal MA, et al. Silencing nociceptor neurons reduces allergic airway inflammation. Neuron (2015) 87:341–54. doi: 10.1016/j.neuron.2015.06.007
5. Abe C, Inoue T, Inglis MA, Viar KE, Huang L, Ye H, et al. C1 neurons mediate a stress-induced anti-inflammatory reflex in mice. Nat Neurosci (2017) 20:700–7. doi: 10.1038/nn.4526
6. Hoover DB, Brown TC, Miller MK, Schweitzer JB, Williams DL. Loss of sympathetic nerves in spleens from patients with end stage sepsis. Front Immunol (2017) 8:1712. doi: 10.3389/fimmu.2017.01712
7. Kressel AM, Tsaava T, Levine YA, Chang EH, Addorisio ME, Chang Q, et al. Identification of a brainstem locus that inhibits tumor necrosis factor. Proc Natl Acad Sci United States America (2020) 117:29803–10. doi: 10.1073/pnas.2008213117
8. Chavan SS, Pavlov VA, Tracey KJ. Mechanisms and therapeutic relevance of neuro-immune communication. Immunity (2017) 46:927–42. doi: 10.1016/j.immuni.2017.06.008
9. Chu C, Artis D, Chiu IM. Neuro-immune interactions in the tissues. Immunity (2020) 52:464–74. doi: 10.1016/j.immuni.2020.02.017
10. Huh JR, Veiga-Fernandes H. Neuroimmune circuits in inter-organ communication. Nat Rev Immunol (2020) 20:217–28. doi: 10.1038/s41577-019-0247-z
11. Lowy DB, Makker PGS, Moalem-Taylor G. Cutaneous neuroimmune interactions in peripheral neuropathic pain states. Front Immunol (2021) 12:660203. doi: 10.3389/fimmu.2021.660203
12. Hagbom M, De Faria FM, Winberg ME, Westerberg S, Nordgren J, Sharma S, et al. Neurotrophic factors protect the intestinal barrier from rotavirus insult in mice. mBio (2020) 11 (1):e028345–19. doi: 10.1128/mBio.02834-19
13. Yu HB, Yang H, Allaire JM, Ma C, Graef FA, Mortha A, et al. Vasoactive intestinal peptide promotes host defense against enteric pathogens by modulating the recruitment of group 3 innate lymphoid cells. Proc Natl Acad Sci United States America (2021) 118(41):e21106634118. doi: 10.1073/pnas.2106634118
14. Tracey KJ. Anti-inflammatory responses normally inhibit inflammation. Nature (2002) 420:853–9. doi: 10.1038/nature01321
15. Martelli D, Farmer DGS, McKinley MJ, Yao ST, McAllen RM. Anti-inflammatory reflex action of splanchnic sympathetic nerves is distributed across abdominal organs. Am J Physiol - Regul Integr Comp Physiol (2019) 316:R235–42. doi: 10.1152/ajpregu.00298.2018
16. Lankadeva YR, May CN, McKinley MJ, Neeland MR, Ma S, Hocking DM, et al. Sympathetic nerves control bacterial clearance. Sci Rep (2020) 10:1–8. doi: 10.1038/s41598-020-72008-4
17. Muldoon LL, Alvarez JI, Begley DJ, Boado RJ, Del Zoppo GJ, Doolittle ND, et al. Immunologic privilege in the central nervous system and the blood-brain barrier. J Cereb Blood Flow Metab (2013) 33:13–21. doi: 10.1038/jcbfm.2012.153
18. Foley JO, DuBois FS. Quantitative studies of the vagus nerve in the cat. i. the ratio of sensory to motor fibers. J Comp Neurol (1937) 67:49–67. doi: 10.1002/cne.900670104
19. Karemaker JM. An introduction into autonomic nervous function. Physiol Meas (2017) 38:R89–R118. doi: 10.1088/1361-6579/aa6782
20. Ek M, Kurosawa M, Lundeberg T, Ericsson A. Activation of vagal afferents after intravenous injection of interleukin-1β: Role of endogenous prostaglandins. J Neurosci (1998) 18:9471–9. doi: 10.1523/jneurosci.18-22-09471.1998
21. Steinberg BE, Silverman HA, Robbiati S, Gunasekaran MK, Tsaava T, Battinelli E, et al. Cytokine-specific neurograms in the sensory vagus nerve. Bioelectron Med (2016) 3:7–17. doi: 10.15424/bioelectronmed.2016.00007
22. Bernik TR, Friedman SG, Ochani M, DiRaimo R, Ulloa L, Yang H, et al. Pharmacological stimulation of the cholinergic antiinflammatory pathway. J Exp Med (2002) 195:781–8. doi: 10.1084/jem.20011714
23. Rosas-Ballina M, Olofsson PS, Ochani M, Valdés-Ferrer SI, Levine YA, Reardon C, et al. Acetylcholine-synthesizing T cells relay neural signals in a vagus nerve circuit. Science (2011) 334:98–101. doi: 10.1126/science.1209985
24. Tsaava T, Datta-Chaudhuri T, Addorisio ME, Masi EB, Silverman HA, Newman JE, et al. Specific vagus nerve stimulation parameters alter serum cytokine levels in the absence of inflammation. Bioelectron Med (2020) 6:0–9. doi: 10.1186/s42234-020-00042-8
25. Murray K, Rude KM, Sladek J, Reardon C. Divergence of neuroimmune circuits activated by afferent and efferent vagal nerve stimulation in the regulation of inflammation. J Physiol (2021) 599:2075–84. doi: 10.1113/JP281189
26. Katayama PL, Leirão IP, Kanashiro A, Luiz JPM, Cunha FQ, Navegantes LCC, et al. The carotid body detects circulating tumor necrosis factor-alpha to activate a sympathetic anti-inflammatory reflex. Brain Behav Immun (2022) 102:370–86. doi: 10.1016/j.bbi.2022.03.014
27. Grundy D. Neuroanatomy of visceral nociception: Vagal and splanchnic afferent. Gut (2002) 51:2–5. doi: 10.1136/gut.51.suppl_1.i2
28. Ozaki N, Gebhart GF. Characterization of mechanosensitive splanchnic nerve afferent fibers innervating the rat stomach. Am J Physiol - Gastrointest Liver Physiol (2001) 281:1449–59. doi: 10.1152/ajpgi.2001.281.6.g1449
29. Kox M, Van Eijk LT, Zwaag J, Van Den Wildenberg J, Sweep FCGJ, van der Hoeven JG, et al. Voluntary activation of the sympathetic nervous system and attenuation of the innate immune response in humans. Proc Natl Acad Sci United States America (2014) 111:7379–84. doi: 10.1073/pnas.1322174111
30. van Westerloo DJ, Choi G, Löwenberg EC, Truijen J, de Vos AF, Endert E, et al. Acute stress elicited by bungee jumping suppresses human innate immunity. Mol Med (2011) 17:180–8. doi: 10.2119/molmed.2010.00204
31. Barros RCH, Bonagamba LGH, Okamoto-canesin R, De Oliveira M, Branco LGS, Machado BH. Cardiovascular responses to chemoref lex activation with potassium cyanide or hypoxic hypoxia in awake rats. Autonomic Neuroscience: Basic and Clinical (2002) 97:110–5. doi: 10.1016/s1566-0702(02)00050-4
32. Kline DD, King TL, Austgen JR, Heesch CM, Hasser EM. Sensory afferent and hypoxia-mediated activation of nucleus tractus solitarius neurons that project to the rostral ventrolateral medulla. Neuroscience (2010) 167:510–27. doi: 10.1016/j.neuroscience.2010.02.012
33. López-Barneo J, Ortega-Sáenz P, Pardal R, Pascual A, Piruat JI. Carotid body oxygen sensing. Eur Respir J (2008) 32:1386–98. doi: 10.1183/09031936.00056408
34. Kumar P, Prabhakar NR. Peripheral chemoreceptors: Function and plasticity of the carotid body. Compr Physiol (2012) 2:141–219. doi: 10.1002/cphy.c100069
35. Prabhakhar NR, Joyner MJ. Tasting arterial blood: What do the carotid chemoreceptors sense? Front Physiol (2015) 6:524. doi: 10.3389/fphys.2014.00524
36. de Castro F. Towards the sensory nature of the carotid body: Hering, de Castro and heymans. Front Neuroanat (2009) 3:23.2009. doi: 10.3389/neuro.05.023.2009
37. Zera T, Moraes DJA, da Silva MP, Fisher JP, Paton JFR. The logic of carotid body connectivity to the brain. Physiology (2019) 34:264–82. doi: 10.1152/physiol.00057.2018
38. Leonard EM, Salman S, Nurse CA. Sensory processing and integration at the carotid body tripartite synapse: Neurotransmitter functions and effects of chronic hypoxia. Front Physiol (2018) 9:225. doi: 10.3389/fphys.2018.00225
39. Nurse CA, Piskuric NA. Signal processing at mammalian carotid body chemoreceptors. Semin Cell Dev Biol (2013) 24:22–30. doi: 10.1016/j.semcdb.2012.09.006
40. Nurse CA. Synaptic and paracrine mechanisms at carotid body arterial chemoreceptors. J Physiol (2014) 592:3419–26. doi: 10.1113/jphysiol.2013.269829
41. Colombari E, Menani JV, Talman WT. Commissural NTS contributes to pressor responses to glutamate injected into the medial NTS of awake rats. Am J Physiol-Regul Integr Comp Physiol (1996) 270:R1220–5. doi: 10.1152/ajpregu.1996.270.6.R1220
42. Malheiros-Lima MR, Silva JN, Souza FC, Takakura AC, Moreira TS. C1 neurons are part of the circuitry that recruits active expiration in response to peripheral chemoreceptors activation. eLife (2020) 9:1–23. doi: 10.7554/eLife.52572
43. Yang CF, Kim EJ, Callaway EM, Feldman JL. Monosynaptic projections to excitatory and inhibitory preBötzinger complex neurons. Front Neuroanat (2020) 14:58. doi: 10.3389/fnana.2020.00058
44. Kara T, Narkiewicz K, Somers VK. Chemoreflexes - physiology and clinical implications. Acta Physiol Scandinavica (2003) 177:377–84. doi: 10.1046/j.1365-201X.2003.01083.x
45. Pijacka W, Katayama PL, Salgado HC, Lincevicius GS, Campos RR, McBryde FD, et al. Variable role of carotid bodies in cardiovascular responses to exercise, hypoxia and hypercapnia in spontaneously hypertensive rats. J Physiol (2018) 596:3201–16. doi: 10.1113/JP275487
46. Koshiya N, Guyenet PG. Tonic sympathetic chemoreflex after blockade of respiratory rhythmogenesis in the rat. J Physiol (1996) 491:859–69. doi: 10.1113/jphysiol.1996.sp021263
47. Aicher SA, Saravay RH, Cravo S, Jeske I, Morrison SF, Reis DJ, et al. Monosynaptic projections from the nucleus tractus solitarii to C1 adrenergic neurons in the rostral ventrolateral medulla: Comparison with input from the caudal ventrolateral medulla. J Comp Neurol (1996) 373:62–75. doi: 10.1002/(SICI)1096-9861(19960909)373:1<62::AID-CNE6>3.0.CO;2-B
48. Ferreira CB, Cravo SL, Stocker SD. Airway obstruction produces widespread sympathoexcitation: role of hypoxia, carotid chemoreceptors, and NTS neurotransmission. Physiol Rep (2018) 6(3):e13536. doi: 10.14814/phy2.13536
49. Madden CJ, Morrison SF. Hypoxic activation of arterial chemoreceptors inhibits sympathetic outflow to brown adipose tissue in rats. J Physiol (2005) 566:559–73. doi: 10.1113/jphysiol.2005.086322
50. Erickson JT, Millhorn DE. Hypoxia and electrical stimulation of the carotid sinus nerve induce fos-like immunoreactivity within catecholaminergic and serotoninergic neurons of the rat brainstem. J Comp Neurol (1994) 348:161–82. doi: 10.1002/cne.903480202
51. Song G, Xu H, Wang H, MacDonald SM, Poon CS. Hypoxia-excited neurons in NTS send axonal projections to kölliker-fuse/parabrachial complex in dorsolateral pons. Neuroscience (2011) 175:145–53. doi: 10.1016/j.neuroscience.2010.11.065
52. Zyuzin J, Jendzjowsky N. Neuroanatomical and neurophysiological evidence of pulmonary nociceptor and carotid chemoreceptor convergence in the nucleus tractus solitarius and nucleus ambiguus. J Neurophysiol (2022) 127:1511–8. doi: 10.1152/jn.00125.2022
53. Allen AM. Angiotensin AT1 receptor-mediated excitation of rat carotid body chemoreceptor afferent activity. J Physiol (1998) 510:773–81. doi: 10.1111/j.1469-7793.1998.773bj.x
54. Chen J, He L, Dinger B, Stensaas L, Fidone S. Role of endothelin and endothelin a-type receptor in adaptation of the carotid body to chronic hypoxia. Am J Physiol - Lung Cell Mol Physiol (2002) 282:1314–23. doi: 10.1152/ajplung.00454.2001
55. Shin MK, Eraso CC, Mu YP, Gu C, Yeung BHY, Kim LJ, et al. Leptin induces hypertension acting on transient receptor potential melastatin 7 channel in the carotid body. Circ Res (2019) 125:989–1002. doi: 10.1161/CIRCRESAHA.119.315338
56. Jendzjowsky NG, Roy A, Barioni NO, Kelly MM, Green FHY, Wyatt CN, et al. Preventing acute asthmatic symptoms by targeting a neuronal mechanism involving carotid body lysophosphatidic acid receptors. Nat Commun (2018) 9:1–15. doi: 10.1038/s41467-018-06189-y
57. Jendzjowsky NG, Roy A, Iftinca M, Barioni NO, Kelly MM, Herrington BA, et al. PKCϵ stimulation of TRPV1 orchestrates carotid body responses to asthmakines. J Physiol (2021) 599:1335–54. doi: 10.1113/JP280749
58. Shu HF, Wang BR, Wang SR, Yao W, Huang HP, Zhou Z, et al. IL-1β inhibits IK and increases [Ca2+] i in the carotid body glomus cells and increases carotid sinus nerve firings in the rat. Eur J Neurosci (2007) 25:3638–47. doi: 10.1111/j.1460-9568.2007.05586.x
59. Rio R, Moya EA, Madrid C, Iturriaga R. Carotid body inflammation and cardiorespiratory alterations in intermittent hypoxia. European Respiratory Journal (2012) 39:1492–500. doi: 10.1183/09031936.00141511
60. Katayama PL. Adrenaline and the carotid body during hypoglycaemia: an amplifying mechanism? J Physiol (2016) 594:7161–2. doi: 10.1113/JP273238
61. Thompson EL, Ray CJ, Holmes AP, Pye RL, Wyatt CN, Coney AM, et al. Adrenaline release evokes hyperpnoea and an increase in ventilatory CO2 sensitivity during hypoglycaemia: a role for the carotid body. J Physiol (2016) 594:4439–52. doi: 10.1113/JP272191
62. da Silva EF, Bassi M, Menani JV, Colombari DSA, Zoccal DB, Pedrino GR, et al. Carotid bodies contribute to sympathoexcitation induced by acute salt overload. Exp Physiol (2019) 104:15–27. doi: 10.1113/EP087110
63. Pardal R, López-Barneo J. Low glucose-sensing cells in the carotid body. Nat Neurosci (2002) 5:197–8. doi: 10.1038/nn812
64. Zhang M, Buttigieg J, Nurse CA. Neurotransmitter mechanisms mediating low-glucose signalling in cocultures and fresh tissue slices of rat carotid body. J Physiol (2007) 578:735–50. doi: 10.1113/jphysiol.2006.121871
65. Gao L, Ortega-Sáenz P, García-Fernández M, González-Rodríguez P, Caballero-Eraso C, López-Barneo J. Glucose sensing by carotid body glomus cells: Potential implications in disease. Front Physiol (2014) 5:398. doi: 10.3389/fphys.2014.00398
66. Ribeiro MJ, Sacramento JF, Gonzalez C, Guarino MP, Monteiro EC, Conde SV. Carotid body denervation prevents the development of insulin resistance and hypertension induced by hypercaloric diets. Diabetes (2013) 62:2905–16. doi: 10.2337/db12-1463
67. Pauza AG, Thakkar P, Tasic T, Felippe I, Bishop P, Greenwood MP, et al. GLP1R attenuates sympathetic response to high glucose via carotid body inhibition. Circ Res (2022) 130:694–707. doi: 10.1161/CIRCRESAHA.121.319874
68. Prasad M, Fearon IM, Zhang M, Laing M, Vollmer C, Nurse CA. Expression of P2X2 and P2X3 receptor subunits in rat carotid body afferent neurones: role in chemosensory signalling(2001) (Accessed December 10, 2017).
69. Joyner MJ, Limberg JK, Wehrwein EA, Johnson BD. Role of the carotid body chemoreceptors in glucose homeostasis and thermoregulation in humans. J Physiol (2018) 596:3079–85. doi: 10.1113/JP274354
70. Wehrwein EA, Basu R, Basu A, Curry TB, Rizza RA, Joyner MJ. Hyperoxia blunts counterregulation during hypoglycaemia in humans: Possible role for the carotid bodies? J Physiol (2010) 588:4593–601. doi: 10.1113/jphysiol.2010.197491
71. Fernández R, González S, Rey S, Cortés PP, Maisey KR, Reyes EP, et al. Lipopolysaccharide-induced carotid body inflammation in cats: Functional manifestations, histopathology and involvement of tumour necrosis factor-α. Exp Physiol (2008) 93:892–907. doi: 10.1113/expphysiol.2008.041152
72. Lam SY, Liu Y, Ng KM, Lau CF, Liong EC, Tipoe GL, et al. Chronic intermittent hypoxia induces local inflammation of the rat carotid body via functional upregulation of proinflammatory cytokine pathways. Histochem Cell Biol (2012) 137:303–17. doi: 10.1007/s00418-011-0900-5
73. Mkrtchian S, Kåhlin J, Ebberyd A, Gonzalez C, Sanchez D, Balbir A, et al. The human carotid body transcriptome with focus on oxygen sensing and inflammation - a comparative analysis. J Physiol (2012) 590:3807–19. doi: 10.1113/jphysiol.2012.231084
74. Wang X, Wang BR, Duan XL, Zhang P, Ding YQ, Jia Y, et al. Strong expression of interleukin-1 receptor type I in the rat carotid body. J Histochem Cytochem (2002) 50:1677–84. doi: 10.1177/002215540205001213
75. Wang X, Zhang XJ, Xu Z, Li X, Li GL, Ju G, et al. Morphological evidence for existence of IL-6 receptor alpha in the glomus cells of rat carotid body. Anat Rec - Part A Discov Mol Cell Evol Biol (2006) 288:292–6. doi: 10.1002/ar.a.20310
76. Reyes EP, Abarzúa S, Martin A, Rodríguez J, Cortés PP, Fernández R. LPS-induced c-fos activation in NTS neurons and plasmatic cortisol increases in septic rats are suppressed by bilateral carotid chemodenervation. Adv Exp Med Biol (2012) 758:185–90. doi: 10.1007/978-94-007-4584-1_26
77. Nardocci G, Martin A, Abarzúa S, Rodríguez J, Simon F, Reyes EP, et al. Sepsis progression to multiple organ dysfunction in carotid chemo/baro-denervated rats treated with lipopolysaccharide. J Neuroimmunol (2015) 278:44–52. doi: 10.1016/j.jneuroim.2014.12.002
78. Mota CMD, Borges GS, Amorim MR, Carolino ROG, Batalhão ME, Anselmo-Franci JA, et al. Central serotonin prevents hypotension and hypothermia and reduces plasma and spleen cytokine levels during systemic inflammation. Brain Behav Immun (2019) 80:255–65. doi: 10.1016/j.bbi.2019.03.017
79. Lam SY, Tipoe GL, Liong EC, Fung ML. Chronic hypoxia upregulates the expression and function of proinflammatory cytokines in the rat carotid body. Histochem Cell Biol (2008) 130:549–59. doi: 10.1007/s00418-008-0437-4
80. Webster JD, Vucic D. The balance of TNF mediated pathways regulates inflammatory cell death signaling in healthy and diseased tissues. Front Cell Dev Biol (2020) 8:365. doi: 10.3389/fcell.2020.00365
81. Cruz J de C, Bonagamba LGH, Stern JE, Machado BH. Fos expression in the NTS in response to peripheral chemoreflex activation in awake rats. Auton Neurosci: Basic Clin (2010) 152:27–34. doi: 10.1016/j.autneu.2009.08.016
82. Martelli D. The inflammatory reflex reloaded. Brain Behav Immun (2022) 104:137–8. doi: 10.1016/j.bbi.2022.06.001
83. McBryde FD, Abdala AP, Hendy EB, Pijacka W, Marvar P, Moraes DJA, et al. The carotid body as a putative therapeutic target for the treatment of neurogenic hypertension. Nat Commun (2013) 4:1–11. doi: 10.1038/ncomms3395
84. Siński M, Lewandowski J, Przybylski J, Bidiuk J, Abramczyk P, Ciarka A, et al. Tonic activity of carotid body chemoreceptors contributes to the increased sympathetic drive in essential hypertension. Hypertens Res (2012) 35:487–91. doi: 10.1038/hr.2011.209
85. Narkiewicz K, Ratcliffe LEK, Hart EC, Briant LJB, Chrostowska M, Wolf J, et al. Unilateral carotid body resection in resistant hypertension: A safety and feasibility trial. JACC: Basic to Trans Sci (2016) 1:313–24. doi: 10.1016/j.jacbts.2016.06.004
86. Marcus NJ, Del Rio R, Schultz EP, Xia XH, Schultz HD. Carotid body denervation improves autonomic and cardiac function and attenuates disordered breathing in congestive heart failure. J Physiol (2014) 592:391–408. doi: 10.1113/jphysiol.2013.266221
87. Niewinski P, Janczak D, Rucinski A, Tubek S, Engelman ZJ, Piesiak P, et al. Carotid body resection for sympathetic modulation in systolic heart failure: results from first-in-man study. Eur J Heart Fail (2017) 19:391–400. doi: 10.1002/ejhf.641
88. Nock ML, DiFiore JM, Arko MK, Martin RJ. Relationship of the ventilatory response to hypoxia with neonatal apnea in preterm infants. J Pediatr (2004) 144:291–5. doi: 10.1016/j.jpeds.2003.11.035
89. Katayama PL, Abdala AP, Charles I, Pijacka W, Salgado HC, Gever J, et al. P2X3 receptor antagonism reduces the occurrence of apnoeas in newborn rats. Respir Physiol Neurobiol (2020) 277:103438. doi: 10.1016/j.resp.2020.103438
90. Melo BF, Sacramento JF, Capucho AM, Sampaio-Pires D, Prego CS, Conde SV. Long-term hypercaloric diet consumption exacerbates age-induced dysmetabolism and carotid body dysfunction: Beneficial effects of CSN denervation. Front Physiol (2022) 13:889660. doi: 10.3389/fphys.2022.889660
91. Conde SV, Sacramento JF, Guarino MP, Gonzalez C, Obeso A, Diogo LN, et al. Carotid body, insulin and metabolic diseases: Unravelling the links. Front Physiol (2014) 5:418. doi: 10.3389/fphys.2014.00418
92. Abdala AP, McBryde FD, Marina N, Hendy EB, Engelman ZJ, Fudim M, et al. Hypertension is critically dependent on the carotid body input in the spontaneously hypertensive rat. J Physiol (2012) 590:4269–77. doi: 10.1113/jphysiol.2012.237800
93. Pijacka W, Moraes DJA, Ratcliffe LEK, Nightingale AK, Hart EC, Da Silva MP, et al. Purinergic receptors in the carotid body as a new drug target for controlling hypertension. Nat Med (2016) 22:1151–9. doi: 10.1038/nm.4173
94. Fitzgerald RS. Carotid body: A new target for rescuing neural control of cardiorespiratory balance in disease. Front Physiol (2014) 5:304. doi: 10.3389/fphys.2014.00304
95. Schultz HD, Li YL, Ding Y. Arterial chemoreceptors and sympathetic nerve activity: Implications for hypertension and heart failure. Hypertension (2007) 50:6–13. doi: 10.1161/HYPERTENSIONAHA.106.076083
96. Pijacka W, Mcbryde FD, Marvar PJ, Lincevicius GS, Abdala APL, Woodward L, et al. Carotid sinus denervation ameliorates renovascular hypertension in adult wistar rats key points. The Journal of Physiology (2016) 21:6255–66. doi: 10.1113/JP272708
97. Conde SV. Ablation of the carotid bodies in disease: meeting its adverse effects. J Physiol (2018) 596:2955. doi: 10.1113/JP275796
98. Langner-Hetmańczuk A, Tubek S, Niewiński P, Ponikowski P. The role of pharmacological treatment in the chemoreflex modulation. Front Physiol (2022) 13:912616. doi: 10.3389/fphys.2022.912616
99. Bautista LE, Vera LM, Arenas IA, Gamarra G. Independent association between inflammatory markers ( c-reactive protein , interleukin-6 , and TNF- a ) and essential hypertension. Journal of Human Hypertension (2005) 19, 149–54. doi: 10.1038/sj.jhh.1001785
100. Rauchhaus M, Doehner W, Francis DP, Davos C, Kemp M, Liebenthal C, et al. Plasma cytokine parameters and mortality in patients with chronic heart failure. Circulation (2000) 102:3060–7. doi: 10.1161/01.CIR.102.25.3060
101. Santos-Almeida FM, Domingos-Souza G, Meschiari CA, Fávaro LC, Becari C, Castania JA, et al. Carotid sinus nerve electrical stimulation in conscious rats attenuates systemic inflammation via chemoreceptor activation. Sci Rep (2017) 7:1–8. doi: 10.1038/s41598-017-06703-0
102. Falvey A, Duprat F, Simon T, Hugues-Ascery S, Conde SV, Glaichenhaus N, et al. Electrostimulation of the carotid sinus nerve in mice attenuates inflammation via glucocorticoid receptor on myeloid immune cells. J Neuroinflamm (2020) 17:1–12. doi: 10.1186/s12974-020-02016-8
103. Katayama PL, Castania JA, Dias DPM, Patel KP, Fazan R, Salgado HC. Role of chemoreceptor activation in hemodynamic responses to electrical stimulation of the carotid sinus in conscious rats. Hypertension (2015) 66:598–603. doi: 10.1161/HYPERTENSIONAHA.115.05316
104. Mughrabi IT, Hickman J, Jayaprakash N, Thompson D, Ahmed U, Papadoyannis ES, et al. Development and characterization of a chronic implant mouse model for vagus nerve stimulation. eLife (2021) 10:1–24. doi: 10.7554/ELIFE.61270
105. Ahmed U, Chang Y-C, Zafeiropoulos S, Nassrallah Z, Miller L, Zanos S. Strategies for precision vagus neuromodulation. Bioelectron Med (2022) 8:1–20. doi: 10.1186/s42234-022-00091-1
106. Kim GY, Lawrence PF, Moridzadeh RS, Zimmerman K, Munoz A, Luna-Ortiz K, et al. New predictors of complications in carotid body tumor resection. J Vasc Surg (2017) 65:1673–9. doi: 10.1016/j.jvs.2016.12.124
107. Ahmed U, Graf JF, Daytz A, Yaipen O, Mughrabi I, Jayaprakash N, et al. Ultrasound neuromodulation of the spleen has time-dependent anti-inflammatory effect in a pneumonia model. Front Immunol (2022) 13:892086. doi: 10.3389/fimmu.2022.892086
108. Cotero V, Fan Y, Tsaava T, Kressel AM, Hancu I, Fitzgerald P, et al. Noninvasive sub-organ ultrasound stimulation for targeted neuromodulation. Nat Commun (2019) 10:1–12. doi: 10.1038/s41467-019-08750-9
109. Porzionato A, Macchi V, Stecco C, Caro R. The carotid sinus nerve – structure, function and clinical implications. Anat Rec (2018) 302:587. doi: 10.1002/ar.23829
110. Case AJ, Zimmerman MC. Sympathetic-mediated activation versus suppression of the immune system: Consequences for hypertension. J Physiol (2016) 594:527–36. doi: 10.1113/JP271516
111. Carnevale D, Pallante F, Fardella V, Fardella S, Iacobucci R, Federici M, et al. The angiogenic factor PIGF mediates a neuroimmune interaction in the spleen to allow the onset of hypertension. Immunity (2014) 41:737–52. doi: 10.1016/j.immuni.2014.11.002
Keywords: Carotid body, inflammation, sympathetic nervous system, neuroimmunomodulation, neuroimmune interactions
Citation: Katayama PL, Leirão IP, Kanashiro A, Menani JV, Zoccal DB, Colombari DSA and Colombari E (2022) The carotid body: A novel key player in neuroimmune interactions. Front. Immunol. 13:1033774. doi: 10.3389/fimmu.2022.1033774
Received: 31 August 2022; Accepted: 11 October 2022;
Published: 24 October 2022.
Edited by:
Eric H. Chang, Feinstein Institute for Medical Research, United StatesReviewed by:
Umair Ahmed, Lenox Hill Hospital, United StatesCopyright © 2022 Katayama, Leirão, Kanashiro, Menani, Zoccal, Colombari and Colombari. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro L. Katayama, a2F0YXlhbWFwbEBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.