 Liping Luo1†
Liping Luo1† Qiaoyun Zheng1†
Qiaoyun Zheng1† Zhenyu Chen1,2
Zhenyu Chen1,2 Meijuan Huang1
Meijuan Huang1 Lin Fu3
Lin Fu3 Jianda Hu1
Jianda Hu1 Qizhen Shi4,5,6,7*‡
Qizhen Shi4,5,6,7*‡ Yingyu Chen1*‡
Yingyu Chen1*‡- 1Department of Hematology, Fujian Provincial Key Laboratory of Hematology, Fujian Institute of Hematology, Fujian Medical University Union Hospital, Fuzhou, Fujian, China
- 2Medical Technology and Engineering College of Fujian Medical University, Fuzhou, Fujian, China
- 3Department of Hematology, The Second Affiliated Hospital of Guangzhou Medical University, Guangzhou, Guangdong, China
- 4Department of Pediatrics, Medical College of Wisconsin, Milwaukee, WI, United States
- 5Blood Research Institute, Versiti, Milwaukee, WI, United States
- 6Children’s Research Institute, Children’s Wisconsin, Milwaukee, WI, United States
- 7Midwest Athletes Against Childhood Cancer (MACC) Fund Research Center, Milwaukee, WI, United States
The development of coagulation factor VIII (FVIII) inhibitory antibodies is a serious complication in hemophilia A (HA) patients after FVIII replacement therapy. Inhibitors render regular prophylaxis ineffective and increase the risk of morbidity and mortality. Immune tolerance induction (ITI) regimens have become the only clinically proven therapy for eradicating these inhibitors. However, this is a lengthy and costly strategy. For HA patients with high titer inhibitors, bypassing or new hemostatic agents must be used in clinical prophylaxis due to the ineffective ITI regimens. Since multiple genetic and environmental factors are involved in the pathogenesis of inhibitor generation, understanding the mechanisms by which inhibitors develop could help identify critical targets that can be exploited to prevent or eradicate inhibitors. In this review, we provide a comprehensive overview of the recent advances related to mechanistic insights into anti-FVIII antibody development and discuss novel therapeutic approaches for HA patients with inhibitors.
Introduction
Hemophilia A (HA) is an X-linked recessive genetic bleeding disorder caused by a deficiency of coagulation protein factor VIII (FVIII). If patients with severe HA do not undergo appropriate prophylaxis, they can suffer from spontaneous bleeding episodes and chronic damage to soft tissues, joints, and muscles. The latest global report from the World Federation of Hemophilia (WFH; compiled in 2019) indicates that the current prevalence of HA is 23.2 in 100000 live male births (1). The clinical phenotype and severity of bleeding in HA patients are generally correlated with residual FVIII levels: mild (5-40% FVIII), moderate (1-5%), or severe (< 1%) (2).
Current treatment approaches for HA involve intravenous infusion of clotting FVIII, which is the mainstay for hemophilia therapy. However, up to 30% of patients with severe HA develop inhibitors, rendering treatment with regular replacement factor concentrates ineffective and increasing morbidity and mortality rates (3). Several genetic and non-genetic factors could be involved in forming inhibitors, and there is currently no clear biomarker to predict whether a patient will produce FVIII inhibitors (4). In 1977, immune tolerance induction (ITI) regimens were first proposed for the treatment of HA patients with inhibitors. However, this regimen requires the frequent administration of relatively large doses of FVIII to downregulate the established immune response and induce immune tolerance to FVIII (5). Since then, this strategy has been modified, including variations in the FVIII dosage and product type, and combinations of additional immunosuppressive agents have been developed. The three most famous protocols are the Bonn (high-dose), Van Creveld (low-dose), and Malmo (refractory patients) (6). Of note, the ITI protocol is the only widely accepted method for establishing FVIII immunologic tolerance. Current consensus guidelines recommend that ITI should be first attempted in patients with FVIII inhibitors (7–9). However, ITI administration is generally provided via frequent central venous catheter infusion, which could increase the risks of thrombosis and infection (10). Moreover, approximately 30% of HA patients cannot achieve sustained immune tolerance to FVIII (11, 12). It remains unclear why some patients never trigger an anti-FVIII immune response, while others do. As such, the prevention and/or eradication of inhibitors to FVIII is important for managing HA.
The purpose of this review is to highlight the current concepts regarding the underlying pathogenesis of FVIII inhibitor development and the therapeutic advances in HA patients with FVIII immunity.
Pathophysiology of FVIII inhibitor development
Characteristics of FVIII inhibitors
FVIII inhibitors are polyclonal IgG antibodies with high affinity, among which IgG4 is typically a major component (13). Most inhibitors neutralize the procoagulant activity of FVIII through steric hindrance, and the most common target epitopes are the A2, A3 and C2 domains of FVIII, which prevent interaction with factor IIa, factor IXa, factor X, von Willebrand factor (VWF), and phospholipids (14). Lacroix-Desmazes et al. (15) found that an additional mechanism of anti-FVIII antibodies is hydrolysis. In 50% of inhibitor-positive patients, anti-FVIII antibodies have enzymatic activity and affect the procoagulant function of FVIII through catalytic hydrolysis of multiple FVIII cleavage sites. This type of anti-FVIII antibody can be clinically detected by the Bethesda or modified Nijmegen method. The titer is reported in the Bethesda units/ml (BU/ml). A high titer inhibitor is defined as an inhibitor level ≥ 5 BU/ml, and a low titer inhibitor is defined as a level < 5 BU/ml (3). Low-titer inhibitors comprise 25% to 50% of observed inhibitors (16), and standard alternative therapies are available for these patients (3). However, by-passing agents are required to treat bleeding episodes in patients with high-titer inhibitors (3, 16).
On the other hand, a few FVIII isoantibodies do not directly inhibit the coagulation activity of FVIII but bind to FVIII antigen epitopes to form circulating immune complexes that are cleared by the reticuloendothelial system, accelerating the metabolic processing of FVIII, shortening the half-life of the internal and external sources of FVIII, and affecting the efficacy of clinical treatment. In this regard, a competition-based enzyme-linked immunosorbent assay (ELISA) could help to differentiate neutralizing FVIII inhibitors from non-neutralizing FVIII-specific antibodies (NNAs) (17). However, as the class of NNAs has been detected in both healthy individuals and PUPs, the true clinical significance of NNAs remains unknown (18, 19). Cannavo et al. reported that the presence of NNAs substantially increased the risk of inhibitor development and that detection of NNAs could be an early predictor of inhibitor development (20). These reports provide further rationale for monitoring NNAs in HA patients. Of note, IgG4 has been identified as the most abundant IgG subclass in HA patients with FVIII inhibitors, though FVIII-specific IgG4 is absent in patients without FVIII inhibitors and in healthy subjects (13). The results from the Hemophilia Inhibitor Previously Untreated Patients (PUPs) Study (HIPS) demonstrated that the development of FVIII inhibitors within the first 50 exposure days (EDs) is associated with distinct antibody signatures (21). Patients with persistent FVIII inhibitors develop unique signatures of FVIII-binding IgG1, followed by IgG3 and IgG4. Low-affinity IgG1 maturing into high-affinity IgG1, IgG3, and IgG4 is correlated with the development of neutralizing FVIII-specific antibodies. High-affinity IgG4 can only be detected after or at the same time as the first FVIII inhibitor detection (21). The distinct antibody signatures could serve as candidates for early biomarkers of FVIII inhibitor development.
Risk factors for inhibitor development in severe HA
Inhibitor development is a complex and multifaceted process involving interactions between genetic (unchangeable) and nongenetic (changeable) factors. The former category includes F8 gene mutations, single nucleotide polymorphisms at the HLA locus, mutations in the immunomodulatory gene, race and family history; the latter category includes the age of first exposure, treatment intensity, type of FVIII preparation, presence of infection and history of surgery (22). Genetic factors are the basis and a prerequisite for the production of inhibitors, while nongenetic factors are typically the triggering factors. Both genetic and nongenetic factors mediate the development of inhibitors and determine their severity and duration.
Severe HA patients with large deletions, nonsense mutations, or intron 22 inversions in the F8 gene have 7-10 times as many inhibitors as those with a small deletion/insertion, missense mutation, or splicing-induced mutation of this gene (23, 24). In a cohort of 203 children with severe HA with inhibitors, 196 cases (96.6%) were identified to have F8 mutations. Of those patients, major mutation types include intron 22 inversions, nonsense mutations, and large deletions or insertions focusing on high- and medium-risk F8 gene mutation types. The authors found that large deletions or insertions encompassing multiple exons and nonsense mutations residing in the light chain contributed to the progression to a high-titer inhibitor and higher peak inhibitor titer in people with severe HA (25). As such, it is highly recommended that gene mutation detection should be conducted for all newly diagnosed HA patients, particularly since it is important for guiding subsequent clinical treatments.
A recent study further demonstrated a significant association of variants in the human leukocyte antigen (HLA) region (26). In particular, low-frequency variants in GRID2IP are closely related to high-titer inhibitors. Another report highlighted that HLA class II molecules play an essential role in inhibitor formation. This report demonstrated that the high-TNF-α/high-IL-10 genotype is associated with an increased risk of immune response to FVIII in severe HA (27). Although certain cytokine genetic polymorphisms have been implicated in inhibitor development, these results are inconsistent between populations with various genetic backgrounds. To further understand the roles of innate immune cells and mechanisms of inhibitor development versus immune tolerance, achieved with or without ITI therapy, researchers performed temporal transcriptomics profiling for HA subjects with and without a current or historic inhibitor using RNA-Seq. HA subjects with a current inhibitor showed differential expression of 56 genes and a clustering analysis identified three major temporal profiles. Interestingly, Gene Ontology (GO) enrichments featured innate immune modulators, including NLRP3, TLR8, IL32, CLEC10A, and COLEC12. NLRP3 and TLR8 are associated with enhanced secretion of the pro-inflammatory cytokines IL-1β and TNFα, while IL32 is associated with both inflammatory and regulatory immune processes. The inflammatory status of HA patients suffering from an ongoing inhibitor includes up-regulated innate immune modulators, which could negatively affect the responses and outcomes of ITI therapy (28).
Nongenetic risk factors for the occurrence of inhibitory antibodies are largely related to FVIII treatment. Lorenzo and colleagues reported that receiving initial treatments at a young age increase the incidence of FVIII inhibitor development in patients with severe HA (29). The other report confirmed that the risk of high inhibitor formation significantly increases in HA children by early exposure to recombinant FVIII (rFVIII) concentrates (30). In this report, the immunogenicity of plasma-derived FVIII (pdFVIII) is speculated to be weaker than that of rFVIII due to VWF protection (30). Multiple studies support that VWF protects FVIII from protease degradation and attenuates FVIII memory immune responses (31, 32). Indeed, the latest evidence from a global multicenter randomized controlled clinical trial (33, 34) showed that for PUPs, the incidence of inhibitors among those treated with rFVIII preparations was markedly higher than in those treated with pdFVIII preparations. This study also stressed that rFVIII should be avoided in the early stages of treatment. To reduce the incidence of inhibitors, the differences in the immunogenicity of the two FVIII preparations require further investigation.
The retrospective CANAL study investigated 366 PUPs with severe HA from 14 European and Canadian hemophilia treatment centers (35). The results demonstrated that the risk of inhibitor development could be explained by early, intensive treatment with FVIII. Intensive FVIII treatment caused by major bleeds or surgeries could become an independent risk factor for inhibitor development. Additionally, early, regular prophylaxis could protect patients with hemophilia against the development of inhibitors (35). Moreover, a large, international cohort study (RODIN study) further demonstrated that high-dose intensive FVIII treatments increase the risk of inhibitor development in PUPs with severe HA (36). The risk of inhibitor development is highest during the first 10 to 20 EDs and decreases to less than 1% after 50 EDs. Prophylactic FVIII treatment reduces inhibitor risk, especially in patients with low-risk F8 mutations (36). However, identifying those patients who could benefit from prophylaxis is challenging and should be addressed in additional studies.
Risk factors associated with inhibitor development in nonsevere HA
In contrast to severe HA, patients with moderate/mild HA have a life-long risk of inhibitor development. Both genetic and environmental factors influence the risk of inhibitor development in patients with nonsevere HA (37–39). The type of F8 mutations is a key risk factor for the occurrence of inhibitors, in patients with severe HA or nonsevere HA (40). In the INSIGHT study, the authors observed that nonsevere HA patients with splice-site mutations had a similar risk as patients with intron 22 inversions. This could be due to the effect of conserved nucleotide positions, which had a higher risk of inhibitor development than those in nonconserved nucleotide positions. The mutations associated with inhibitors in the INSIGHT cohort were all located within regions encoded for the light chain and the A2 domain of FVIII (41). This study further showed that the risk of inhibitor development was 6.7% at 50 ED and increased to 13.3% at 100 ED (41), which differs from the cases of severe HA mentioned above (36). As such, inhibitors should be routinely screened during the time of greatest risk for inhibitor development in patients after FVIII replacement therapy. In pediatric patients, inhibitor screening should be performed every 5 EDs for the first 20 EDs after the first treatment with clotting factor products (36, 40, 42, 43).
Inhibitors usually occur at an earlier age in nonsevere HA. Recently, a total of 6624 persons with nonsevere HA were investigated for an average of 8.5 years in the United States (38). The results demonstrated that the prevalence of inhibitors was 2.6% (n = 171), occurring at a median age of 13 years. However, the occurrence of inhibitors at an early age was not associated with increased mortality. Previous studies revealed that older age is another risk factor for inhibitor development (40, 44). Age-related immune dysregulation and late loss of tolerance could be particularly relevant for the nonsevere hemophilia cohort, as exposure to therapeutic FVIII concentrates is distributed throughout a lifetime and often skewed toward later decades for elective operative interventions. Moreover, both high-dose FVIII treatment and surgical interventions could increase the risk of inhibitor development in nonsevere HA patients (39, 45). In the INSIGHT study, the all-cause mortality rate in nonsevere HA patients with inhibitors was more than five times higher compared to those without inhibitors (46). Inhibitor-related mortality primarily increased in older patients with high inhibitor titers. These findings indicate that nonsevere hemophilia is not mild and highlight the importance of closely following-up with these patients.
Inhibitor occurrence connected with the surrounding microenvironment has recently raised concern. Researchers have found that inhibitors are often produced along with the surrounding danger signals. The “danger theory”, proposed by Polly Matzinger in 1994 (47–49), postulated that the human immune response is triggered by “danger signals” or “alarm signals”. Patients are often exposed to various endogenous or exogenous danger signals, such as those resulting from surgery, trauma, and infection, during treatment with FVIII. These danger signals, termed danger-associated molecular patterns (DAMPs), could become the major drivers to elicit an anti-FVIII immune response (48, 50–54). A clinical trial reported by Kurnik et al. (55) demonstrated that the production of FVIII inhibitors decreases during the administration of the first dose of FVIII by avoiding proinflammatory stimuli (bleeding, infection, surgery, vaccination, etc.). However, it remains difficult to predict the potential risks associated with individual patients’ inhibitor development. The available evidence is scant and inconclusive, and distinguishing the contribution of peak treatment from danger signals remains challenging. However, by clearly identifying danger signals, we could further explore the precise mechanisms underlying the risk effect, identify different biomarkers, and provide new targets to effectively prevent inhibitor development. As such, in the future clinical management of HA patients, the impacts of danger signals should be monitored in various risk scenarios, and therapeutic strategies can be improved by avoiding unnecessary risks.
Immunological pathophysiology of inhibitor development
Previous studies confirmed that FVIII inhibitor development is CD4+ T cell-dependent in both murine models and patients (56–60). T follicular helper cells (TFHs), a novel subset of CD4+ T cells, work with cognate follicular B cells to trigger a germinal center (GC) reaction that is ultimately responsible for the production of anti-FVIII antibodies in HA mice (58). In a multicenter study, significant reductions in levels or even complete elimination of FVIII inhibitors were observed in HA patients with high titers of inhibitors and concomitant HIV infection (61). Mechanistically, it is the antigen-presenting cells (APCs) that present FVIII peptides on major histocompatibility complex class II (MHC-II) to T cells, triggering FVIII immune responses. Indeed, several cell types, such as dendritic cells (DCs), macrophages, B lymphocytes and endothelial cells, could act as APCs that mediate FVIII-specific immune response (59, 62). Low-density lipoprotein receptor-related protein (LRP, CD91), the mannose receptor (MR), and membrane-bound heparan sulfate proteoglycans (HSPGs) play important roles in FVIII endocytosis by binding to the A2, A3 and C1 domains of FVIII (63–65), anti-FVIII C1 domain antibodies can prevent the endocytosis of FVIII by APCs (66). These in vitro and in vivo mouse model studies facilitate the discovery of the key factors involved in inhibitor development.
An in vitro study demonstrated that VWF reduced the endocytosis of FVIII by human DCs through steric hindrance effects (67). In murine HA models, researchers found that phosphatidylserine (PS) induced immune tolerance and reduced the production of inhibitors by affecting the maturation of DCs, inducing the generation of Tregs, and inhibiting the generation of memory B cells (68–70). More recently, studies have demonstrated that the binding of FVIII to lysophosphatidylserine (Lyso-PS) to form the Lyso-PS-rFVIII Fc complex significantly reduced inhibitor development in HA mice after either intravenous or oral administration of Lyso-PS (71, 72). In addition, researchers found that site N2118, containing high-mannose glycans, have a significant impact on FVIII immunogenicity in the murine HA model (73). Results from these studies can help identify valuable targets to reduce FVIII immunogenicity.
The spleen is the predominant organ involved in initiating the immune response. In a murine model study, researchers confirmed that APCs in the marginal zone (MZ) of the spleen play critical roles in the immune response to FVIII (74). Zerra and coworkers found that transfused FVIII was located in the marginal sinus of the spleen and colocalized with the B cells in the MZ of the HA mouse spleen, and the development of FVIII inhibitors could be completely blocked by specific removal of the MZ B cells (75). Moreover, the activation of APC costimulatory signals (CD40, CD40L, CD80/CD86, and CD28) is necessary for the complete activation of T cells. Indeed, blocking the CD40/CD40L interaction has been found to induce long-term immune tolerance in HA mice. Unfortunately, treatment with anti-CD40 can also activate platelets and increase the risk of thrombosis (76).
Activated T cells can differentiate into effector T cells (Teffs) and regulatory T cells (Tregs). Teffs are responsible for initiating and maintaining an effective anti-FVIII immune response by activating B cells to differentiate into mature plasma cells through a GC process. In contrast, Tregs could act as potent suppressors of the Teffs to inhibit antibody development. There is growing evidence that the generation of Tregs via either human samples or animal models could lead to FVIII tolerance, suggesting that the balance between Teffs and Tregs is another key determinant involved in the pathophysiology of FVIII inhibitor development (57, 77–80). A recent study showed that immune tolerance against FVIII under non-hemophilic conditions was maintained by programmed death (PD) ligand 1 (PD-L1)-expressing Treg (81). In addition, B cell-activating factor (BAFF) cytokine family is a key regulator of B cell differentiation in normal homeostasis and immune disorders. A recent study demonstrated that BAFF levels are elevated in pediatric HA inhibitor patients and in those who failed to achieve immune tolerance with anti-CD20-mediated B cell depletion. Using a mouse model, the authors found that BAFF modulated tolerance induction and inhibitor eradication by downregulating plasma cells, demonstrating the important role of BAFF in the modulation of anti-FVIII response (82).
Moreover, memory B and/or long-lived plasma cells (LLPCs) play a key role in maintaining established anti-FVIII immune responses. Importantly, FVIII-specific memory B cells are present in HA patients with inhibitors, while these cells are absent in healthy controls or patients without inhibitors (83). Following re-exposure to FVIII, the unique memory B cells can rapidly differentiate into plasma cells that produce high levels of anti-FVIII antibodies (84). Additionally, there is evidence that memory B cells contribute to replenishing the pool of LLPCs in the bone marrow (BM) (85). GC reaction memory B cells can develop into plasma blasts that migrate to the BM, and mature into LLPCs that maintain serum levels of antibodies for prolonged periods. LLPCs persistently producing anti-FVIII antibodies in BM are unlikely to be affected by conventional high dosages of FVIII infusions (84, 86). Therefore, for long-term induction FVIII tolerance, it is highly necessary to develop novel strategies targeting FVIII-specific memory B cells and LLPCs.
Novel strategies for treating HA with inhibitors
Conventional hemostatic bypassing agents
The development of FVIII inhibitors in HA patients remains challenging in clinical management. Currently, the mainstay of treating HA inhibitor patients requires controlling bleeding symptoms and eliminating alloantibodies against FVIII. Unfortunately, ITI protocols are not always effective in patients with high levels of inhibitors (87–89), making alternative strategies clinically necessary. In recent years, promising therapeutic strategies have been identified, including treatment with immunomodulatory drugs or molecules, oral or transplacental delivery of FVIII, and cell or gene therapy (90–92).
For patients who appear to be ineffective for standard replacement therapies and have confirmed inhibitors, a “bypass approach” should be adopted immediately to stop bleeding regardless if the patients undergo ITI or not (9); this approach includes treatment with recombinant activated factor VII (rFVIIa) and activated prothrombin complex concentrate (aPCC). Either rFVIIa or aPCC could be used to treat bleeding episodes (BEs) in both HA and hemophilia B (HB) patients with inhibitors (9). However, a major concern remains about the risks of thrombotic microangiopathy in aPCC-treated patients receiving non-factor prophylactic agents such as emicizumab (93). In addition, bypassing agents are expensive and provide incomplete hemostatic correction in some patients (94). As such, new strategies for the control and prevention of BEs are highly anticipated. Various new approaches based on non-factor replacement therapies have been developed and entered clinical trials, which include facilitating the coagulation pathway (e.g. emicizumab) and blocking the anticoagulant pathway (e.g. concizumab, fitusiran).
Emicizumab: Bispecific monoclonal antibody
The new hemostatic agent emicizumab is a humanized bispecific monoclonal antibody that binds to and bridges activated factor IX (FIXa) and factor X to simulate the physiological function of FVIII (95). It has been licensed for bleeding prophylaxis in HA patients with or without inhibitors, and is the first non-factor replacement therapy for HA (93). Subjects enrolled in early clinical trials tolerated once-weekly subcutaneous injections of emicizumab at various doses (0.3, 1.0, and 3.0 mg/kg); no bleeding occurred in 73% of subjects, and no neutralizing antibodies specific for emicizumab were observed during 12 weeks of treatment in the early phase of clinical trial (96, 97).
Data from the HAVEN 1 phase 3 trial showed that among the 109 HA patients aged 12 years and older with high titers of inhibitors, the bleeding rate in the trial group treated with emicizumab was 87% lower than in the untreated group (95, 98). Emicizumab significantly improved the quality of life of patients and had an obvious effect in preventing bleeding. The HAVEN 2 study enrolled 88 children with inhibitors with a median age of 7 years (range, 1-15 years) (99). In this trial, participants who received an emicizumab injection with 4 once-weekly loading doses of 3 mg/kg followed by a maintenance regimen of once weekly (1.5 mg/kg, QW) showed a low annualized rate of treated bleeding event (ABR), with 77% of participants experiencing no treated bleeding events. Notably, efficacy was maintained among those who received emicizumab every 2 weeks (3 mg/kg, Q2W) or every 4 weeks (6 mg/kg, Q4W). This large prospective study highlighted the special role of emicizumab as a highly effective novel medication for pediatric HA patients with anti-FVIII inhibitors.
The efficacy, safety, and pharmaceutics of these treatments have been further studied in the phase 3 HAVEN 3-5 studies, which included adults/adolescents ≥ 12 years of age with and without inhibitors (100–102). These trials demonstrated that emicizumab prophylaxis achieved remarkable efficacy for bleeding control and was well tolerated in HA patients, regardless of FVIII inhibitor status. Of note, the long-term efficacy and safety of emicizumab for up to 5.8 years were reported in patients with severe HA (103). Nevertheless, emicizumab provides more convenient and feasible administration way (maintenance dose of once every 1, 2 or 4 weeks) than conventional FVIII replacement therapy. No anti-FVIII antibodies developed among those participants during administered emicizumab, and neither serious nor thrombotic adverse events were reported in either of these clinical trials (104). However, as thrombosis and thrombotic microangiopathy occurred in some participants who had received aPCC in HAVEN 1 (95), WFH recommends the use of rFVIIa instead of aPCC for treating breakthrough bleeding during emicizumab prophylaxis (9). Moreover, 0.33% (25/7500) of subjects produced an immune response to emicizumab (93). In the HAVEN trials, 2 out of 88 pediatric participants in HAVEN 2 (99, 105) and 1 out of 64 evaluable participants in HAVEN 5 (102) developed anti-drug antibodies (ADAs) with neutralizing potential. Although this occurs infrequently, it is necessary to monitor the neutralizing ADAs to implement immunological surveillance in HA patients with emicizumab regimens.
The final analysis of the STASEY phase III clinical trial enrolled a total of 193 HA patients age ≥ 12 years with inhibitors who received emicizumab once weekly for up two years (106). The results revealed that emicizumab was well-tolerated in the cohort of participants. The most common AEs were arthralgria (17.1%), naspharyngitis (15.5%), and headache (15%). Emicizumab-related AEs were observed in 18.1% of participants, with injection site reactions being the most frequent (9.8%). No emicizumab-related thrombotic microangiopathies (TMAs) or thromboembolic events (TEs) were reported in five participants who also received aPCC. ADAs were found in 5.2% of participants, 2.6% of whom were classified as having ADAs that were neutralizing in vitro. Most importantly, emicizumab continued to present effective bleeding control, with 82.6% of participants achieving no bleeding episodes that required treatment. The final analysis of the phase IIIb STASEY study further confirmed the favorable safety profile of emicizumab, which is consistent with the phase III HAVEN clinical program.
The first case of an emicizumab ADA was reported in clinical trials (107). In this report, a 6-year-old boy with severe HA with inhibitors received standard loading doses of emicizumab and was then administered a 1.5 mg/kg/week regimen. However, 3 months into treatment the patient presented with a traumatic bleed due to the loss of emicizumab effect with a proven ADA. Emicizumab was discontinued and the patient resumed daily prophylaxis with rFVIIa. Therefore, it is important for clinicians to be aware that meaningful ADAs can occur, and to refer the patient to a special coagulation laboratory for ADAs testing if clinical suspicion arises. Moreover, real-world data from 52 pediatric patients with severe HA revealed that emicizumab was safe and well tolerated and that minor AEs, including headaches, abdominal pain and nausea, and injection site reactions, occurred in about 7.7% of patients. Four patients experienced major AEs, including severe headaches, major bleeding events, development of ADAs, and recurrence of inhibitors. Emicizumab prophylaxis was discontinued in three patients (5.7% of the cohort) due to AEs. No AEs were reported in four PUPs of the cohort (108).
Although emicizumab demonstrated significant reductions in ABR compared with standard prophylaxis, however, bypassing agents, such as aPCC, should be used with caution. As mentioned above, 3 patients developed TMA in HAVEN 1 and 2 additional TEs were reported among 401 participants in HAVEN 1-4 (109). For the 3 patients who developed TMA, one proposed explanation is that these patients concomitantly received aPCC, which contains the targets of emicizumab, FIX/IXa and FX/Xa. The excess substrate availability could have resulted in uncontrolled thrombin generation and subsequent thrombotic complications.
Currently, there are other FVIII mimetics under development. Mim8 is a next-generation bispecific to FIXa and FX, which shows enhanced thrombin generation in vivo compared with a sequence-identical analog of emicizumab and an extended half-life of 14 days in cynomolgus monkeys (110). Subcutaneous administration of Mim8 up to 3 mg/kg/week for 26 weeks resulted in relevant pharmacodynamic effects, with no signs of thrombi or excessive coagulation activation (111). The FRONTIER 1 study is an ongoing phase 1/2 study of Mim8 in the HA participants with or without inhibitors ≥12 years of age. Another FVIII mimetic bispecific antibody, BS-027125, is currently in preclinical evaluation with no human data yet available.
Targeting natural anticoagulants
In recent years, novel non-factor replacement therapies based on targeting of natural anticoagulants have been acknowledged; and the efficacy and safety in HA patients with inhibitors are currently ongoing in clinical trials. These new strategies consist of targeting negative clotting regulators such as tissue factor pathway inhibitor (TFPI), antithrombin (AT), or activated protein C (aPC).
TFPI presents two splicing forms TFPIα and TFPIβ. TFPIα is a soluble form of TFPI that is responsible for inhibiting prothrombin activity in the extrinsic coagulation system. It inhibits FVIIa via its K1 domain, FXa via its K2 domain, and protein S via its K3 domain (112). Current research focuses on monoclonal antibodies (mAbs) specific to the K2 domain. Studies on the evaluation of these novel molecules, including befovacimab, concizumab, and marstacimab, have been ongoing in participants with HA or HB, regardless of the presence of inhibitors. Of these, befovacimab also targets the K1 domain of the TFPI (113). While favorable results were obtained from the preclinical in vivo studies and phase 1 clinical trials, the phase 2 study was terminated early due to three befovacimab-related thrombotic serious adverse events (SAEs) (114). Therefore, the therapeutic window of anti-TFPI treatment must be further investigated.
Concizumab is an IgG4 humanized mAb selectively targeting the K2 domain of the TFPI. Results from phase 1-3 clinical trials in healthy volunteers and HA patients showed a dose-dependent increase in D-dimer and thrombin fragments after concizumab treatment, accompanied by significant hemostatic efficacy and tolerability. These results indicate the adequate safety and half-life characteristics of concizumab, whether administered subcutaneously or intravenously (115–117). However, in phase 3 of the clinical trial, three patients experienced five nonfatal thrombotic SAEs (118, 119). These AEs were acute myocardial infarction in one patient with HA, a renal infarction in one HB patient with inhibitors, and three TEs (deep venous thrombosis, pulmonary embolism, and superficial thrombosis of the vein) in one patient with HA. This trial had to be temporarily suspended and resumed after modifying the protocols for treating breakthrough bleeding. Shapiro and colleagues presented the results of concizumab prophylaxis from the extension parts of phase 2 explorer 4 (HA or HB with inhibitors) and explorer 5 (severe HA without inhibitors) trials (119). The long-term efficacy of concizumab was maintained during the trial, and estimated ABRs at the last concizumab dose after more than 76 weeks of treatment were comparable to those observed in the main parts (≥ 24 weeks). Although the authors found that around 25% of patients developed ADAs, the ADAs levels were low and transient, with no observed SAEs in most cases. Phase 3 trials are currently ongoing, and will provide further insight into the efficacy and safety of concizumab to treat hemophilia in the future. Moreover, other humanized mAbs, such as marstacimab, which specifically binds to the K1 domain of the TFPI, have shown the feasibility and efficacy of targeting TFPI therapies in hemophilia. A phase 1b/2 clinical study demonstrated clinically meaningful reductions in ABRs and treatment-related changes for all pharmacodynamics biomarkers across all marstacimab dose levels in participants with hemophilia (120). Together, therapeutic advances in hemophilia with targeting TFPI treatments are promising. Currently, two of the three novel anti-TFPI mAbs have progressed to phase 3 clinical trials. However, the potential thrombotic issue remains a key consideration and should be thoroughly evaluated in ongoing clinical trials.
AT is a member of a small protein family produced in liver cells and is an effective thrombin inhibitor. Fitusiran is an antithrombin RNA interference molecule that can silence AT mRNA expression in liver cells, increase the production of thrombin, and reduce bleeding tendency (121, 122). Fitusiran-induced reduction of AT levels can rebalance hemostasis in patients with HA or HB, with or without inhibitors. The phase 1 dose-escalation study enrolled 25 patients with moderate to severe HA or HB (122). The results demonstrated a fitusiran dose‐dependent mean maximum AT reduction of 70%–89% from baseline, with a reduction of > 75% enabling a similar degree of thrombin generation compared with healthy volunteers. One participant reported severe chest pain, and although thrombosis was ruled out, this event led to treatment discontinuation. No ADAs developed in the enrolled four healthy volunteers or the participants with hemophilia during the study course (122, 123). The results from the phase 2 and phase 3 clinical trials showed that the thrombin peak height in the enrolled participants was related to the degree of AT reduction when fistusiran was subcutaneously administered each month. AT reduction by ≥ 75% from baseline led to a significant improvement in thrombin generation and decreased bleeding frequencies (124, 125). The SAE was a fatal cerebral sinus thrombosis in one patient during the phase 2 trial, which occurred after repeated infusions of high FVIII concentrate doses over more than 24 hours. Three additional nonfatal vascular thrombotic events occurred in late phase 3 trials despite adherence to breakthrough bleed management guidelines. These were attributed to AT levels decreasing below 10% (126, 127). As a result, studies targeting higher AT levels have been resumed, seeking to maintain a favorable benefit-risk balance for patients. Overall, fitusiran could offer either HA or HB patients with inhibitor effective prophylaxis delivered through monthly, low volume, fixed-dose and subcutaneous administration.
In general, the aim of all therapeutic strategies in hemophilia is to increase the amount of thrombin generation at sites of vascular damage, which can be generally achieved by replacing missed clotting factors or by increasing the concentration of either the enzymes or substrates that result in the production of thrombin. Aymonnier and colleagues reported a novel approach consisting of targeting a natural and potent thrombin inhibitor, named protease nexin-1 (PN-1) (128). The authors showed that a PN-1-neutralizing antibody could significantly shorten the thrombin burst in response to tissue factor in platelet-rich plasma (PRP) from mild or moderate hemophilia patients. In contrast, neutralized PN-1 did not improve thrombin generation in PRP from severe hemophilia patients. However, with collagen-induced platelet activation, PN-1 deficiency in HA mice or PN-1 neutralization in patients with severe HA resulted in significantly improved thrombin generation in PRP. Recent experimental and clinical studies revealed that serine protease inhibitors (SERPINs), e.g. α1-antitrypsin and SerpinPC, could decrease bleeding in hemophilia via selective inhibition of aPC (129–132). Nevertheless, specific inhibition of the aPC anticoagulant function by the novel inhibitory mAb has been designed and developed in recent years (133–135). Indeed, the administration of mAb selectively inhibits aPC’s anticoagulant activity, but does not compromise its cytoprotective function, and offers a better therapeutic potential alternative for HA.
Novel strategies for eradicating inhibitors or immune tolerance induction
AAV-liver gene therapy
In recent years, hepatic in vivo gene therapy via adeno-associated virus (AAV) vectors has demonstrated promising results in clinical trials for hemophilia patients in the first year after treatment, but FVIII levels steadily decline afterward. Compared to those in HB, clinical trials in HA have been slower due to the large size of F8 cDNA (7 kb). Encouragingly, after removing the sequence encoding the nonfunctional domain (B-domain deletion, BDD), the truncation of F8 can be incorporated into AAV vectors (136). The first successful application of this approach was found in 6 of 7 severe HA participants who received a single high-dose cohort of an AAV5 vector encoding a BDD F8 (AAV5-hFVIII-SQ) (137). The FVIII activity (FVIII:C) level of these participants remained stable at or above the physiologic range 1 year after receiving AAV5-hFVIII-SQ liver-directed gene therapy. The annualized rates of FVIII concentrate use and treated bleeding significantly decreased in the participants (138). However, multiyear follow-up data from the participants receiving AAV5-hF8-SQ gene therapy showed that FVIII expression steadily declined (139). This raises concerns about the durability and safety of AAV-based liver-specific gene therapy. Currently, several AAV-based gene transfer strategies have been modified and clinical trials have been initiated, which could offer hope to improve the durability and safety of this therapy for patients with hemophilia (140–143).
To date, only adult men with endogenous factor levels ≤ 2% and without the advanced liver disease have received gene therapy. It remains unknown whether HA patients with inhibitors respond to AAV liver-directed gene therapy approach. Evidence from murine and canine models has demonstrated that liver-directed F8 gene therapy could induce tolerance towards a primed immune system with pre-existing inhibitors (144–147). In HA dogs with pre-existing inhibitors after canine(c) FVIII AAV-based liver-directed gene therapy, there is an early increase in CD4+ CD25+ FOXP3+ Tregs within the first few days (145, 146). This is associated with a decline in anti-cFVIII antibodies but was not observed in HA dogs without inhibitors receiving similar therapy. Furthermore, immune tolerance was established in treated dogs even after repeated challenges with cFVIII proteins. In mouse model studies, AAV-F8 gene therapy can not only correct FVIII levels, but also results in low- to undetectable inhibitor titers following subsequent challenges with rFVIII in HA mice on a BALB/c background (147, 148). The ability to induce FVIII tolerance in these mice occurred in both the presence and absence of transient immunosuppression mediated by B cell depletion with anti-CD20. In contrast, HA mice on a mixed BL/6-129/sv background following AAV-F8 gene transfer only resulted in modest correction of the clotting time below that of untreated mice regardless of anti-CD20 treatment. These mice lost bleeding phenotype correction along with the development of inhibitors when challenged with supplementary rFVIII. However, transient B cell depletion with anti-CD20 at the time of AAV-F8 gene transfer resulted in subsequent hyporesponsiveness to FVIII with significantly lower inhibitors (147). Thus, these results highlight the potential role of genetics in determining whether a patient will respond to AAV-F8 gene therapy. Importantly, targeting B cell method could further improve tolerance induction using this novel approach.
As mentioned above, a combination regimen targeting FVIII-specific memory B cells and LLPCs facilitate ITI to FVIII has recently attracted research interest. A study by Liu et al. found that anti-CD20 combined with IL-2/IL-2 mAb complexes plus rapamycin significantly reduced both FVIII-specific antibody-secreting cells and memory B-cells in HA inhibitor mice (148). The combination had a synergistic effect on the inhibition and neutralization of antibody production. Moreover, as LLPCs play a key role in maintaining established antibodies, the same group developed a novel strategy that could target LLPCs (149). In the regimen, AMD3100, an antagonist of CXCR4, was used to block the CXCL12/CXCR4 interaction and inhibit the homing and retention of LLPCs (150). The results demonstrated that the combined treatment using AMD3100, G-CSF, anti-CD20 and IL-2/IL-2 mAb complexes effectively eradicated LLPCs and inhibitor titers in either FVIII protein- or FVIII plasmid-primed HA inhibitor mice. Encouragingly, a recent study found that B cell depletion with anti-CD20 eliminates memory B cells and enhances FVIII tolerance when combined with rapamycin, preventing neutralization of the newly expressed clotting FVIII in HA mice following AAV8-coF8 gene therapy. In addition, it has been reported that combining rapamycin and anti-CD20 therapy with conventional FVIII ITI treatment could induce FVIII tolerance in patients with previously refractory FVIII inhibitors (151). Herein, a combination of anti-CD20/mTOR inhibition strategies could increase the success of ITI and allow for the inclusion of HA inhibitor patients for AAV-mediated F8 gene therapy or FVIII ITI in clinical trials.
LV-platelet gene therapy
Lentiviral vector (LV)-mediated gene therapy has demonstrated significant potential for treating FVIII deficiency. In particular, LV-mediated platelet-targeted gene transfer into hematopoietic stem cells (HSCs) results in stable integration of the F8 gene into the host genome, leading to persistent therapeutic effects in hemophilia models with and without inhibitors (152–157). The ectopic expression of FVIII under the platelet-specific promoter control enables FVIII storage together with VWF in α-granules in platelets. This could shield FVIII from the circulating neutralizing anti-FVIII antibodies until it is delivered to the site of injury upon platelet activation. Even relatively small numbers of activated platelets that locally excrete FVIII could be sufficient to promote efficient clot formation in the recipient mice. Notably, none of the transduced mice developed an anti-FVIII immune response even when challenged with rFVIII. The inhibitor titers decreased with time in the pre-existing anti-FVIII immunity models after LV-F8 gene therapy (57, 152, 155, 157, 158). Furthermore, infusion of platelets containing FVIII triggers neither primary nor memory anti-FVIII immune responses in HA mice (92), indicating that it could become a useful alternative treatment approach for HA inhibitor patients if the availability of AAV-based gene therapy is limited by certain circumstances. Importantly, novel gene therapy strategies could also induce Tregs expansion and CD4+ T cell-mediated immune tolerance (155).
Since the LV-F8 gene therapy approach targets HSCs, this strategy could result in lifelong FVIII production. Encouragingly, the LV-mediated therapeutic strategy could also provide an alternative gene therapy for patients with pre-existing immunity to the AAV. Currently, a phase 1 clinical trial using an LV-mediated targeting platelet F8 gene delivery system has been initiated at the Medical College of Wisconsin (ClinicalTrials.gov: NCT03818763) (141). However, either total body irradiation or chemotherapy plus immune suppression is required to facilitate engraftment and efficacy, which could limit widespread applications of this strategy in clinical settings.
Nonviral delivery system
Oral tolerance induction via transplastomic lettuce expressing FVIII or FIX fused to a transmucosal carrier is at the forefront of hemophilia treatment research. Such bioencapsulated factors can be ingested, cross the intestinal epithelium, and induce Treg cells within the gut-associated lymphoid tissue (141). Sherman et al. demonstrated that the FVIII heavy chain and C2 domain can be expressed as cholera toxin B subunit fusion proteins in tobacco chloroplasts (159). Oral delivery of a mixture of these bioencapsulated antigens could suppress and reverse inhibitor formation in HA mice. Intestinal microbiota plays a vital role in maintaining immune homeostasis through the interaction between the microbiome and the intestinal immune system (160). A recent study revealed that tolerance induction by oral delivery of antigens bioencapsulated in plant cells occurs via the unique immune system of the small intestine, and the inhibition of antibody formation primarily performed by CD4+ CD25− FoxP3− LAP+ Tregs (but not of CD4+ CD25+ FoxP3+ Tregs) (161). Antigen release from the plant cells occurs via bacteria with required enzymatic degradation activities in the small intestine and delivery to the associated immune system. The composition of this microbiome is distinct from that of the large intestine, and their augmentation could further promote plant cell-based oral tolerance induction in the treatment of hemophilia. The ongoing advances of the novel nonviral delivery system in animal HA models make it a likely candidate for clinical trials.
Since the goal of the development of tolerogenic therapies is to suppress antibody formation, Tregs have been proposed as a potential clinical therapy for various adverse immune disorders. Adoptive transfer of ex vivo-expanded Tregs improved immune cell engraftment and graft-versus-host disease after HSC transplantation in the clinical trial (162). Nevertheless, chimeric antigen receptor (CAR) Tregs expansion has been identified as a novel biomarker of response and toxicity after CAR T cell therapy, which raises the prospect that this subset could regulate CAR T cell response in humans (163). Interestingly, Yoon et al. demonstrated that FVIII-specific CAR (ANS8 CAR) Tregs can be engineered, and demonstrates their ability to inhibit T- and B-cell effector responses to FVIII (164). Importantly, ANS8 CAR-transduced Tregs can suppress the recall antibody response of murine splenocytes from FVIII knockout mice to FVIII in vitro and in vivo. Notably, CAR-modified Tregs engineered in a non-MHC restricted manner have widespread applications and have gained increasing attention in recent years (165, 166). This novel cellular therapy remains a promising approach for the future tolerogenic treatment of HA patients with existing inhibitors.
Closing remarks
Anti-FVIII immunity represents a major limitation of HA therapy. Multiple factors impact inhibitor development in patients with HA, although novel alternative therapeutic options can be used to prevent bleeding disorders in HA patients with and without harboring inhibitors. However, the benefits are only available for a small number of HA patients, and the lack of infrastructure facilitating the worldwide availability of novel strategies remains a barrier. There is also an unmet need to provide further insight into the longer-term efficacy and safety of these novel approaches. As such, mechanistic insights into humoral immunity to FVIII and identifying new biomarkers connecting inhibitor generation remain top priorities for clinical or preclinical investigations.
Author contributions
YC wrote, revised, edited, critically reviewed and submitted the manuscript. LL drafted part of the manuscript. QZ, ZC and MH provided comments and discussions for the manuscript. LF and JH critically reviewed the manuscript. QS critically reviewed, edited, and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was sponsored by Joint Funds for the Innovation of Science and Technology in Fujian Province (2020Y9056), Fujian Provincial Natural Science Foundation (2020J01996), National Key Clinical Specialty Discipline Construction Program (2021–76), Fujian Provincial Clinical Research Center for Hematological Malignancies (2020Y2006), National Institutes of Health grant HL-102035 (QS) and Midwest Athletes Against Childhood Cancer and Bleeding Disorders (MACC) Fund (QS).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
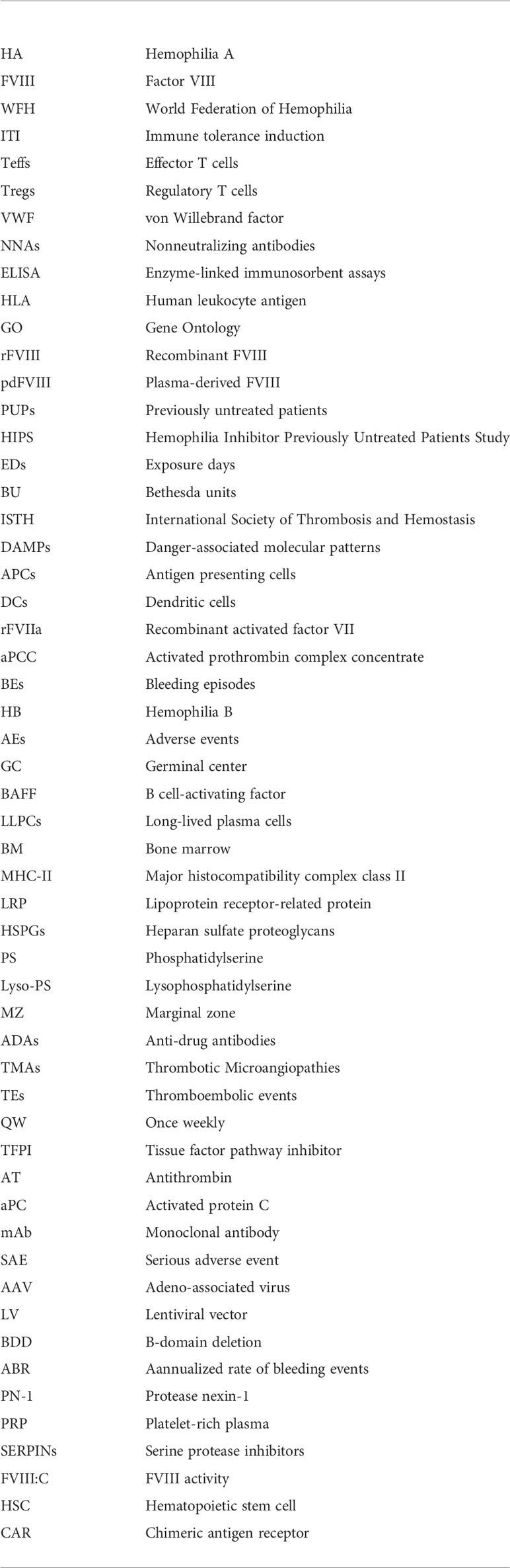
Glossary
References
1. Iorio A, Stonebraker JS, Chambost H, Makris M, Coffin D, Herr C, et al. Establishing the prevalence and prevalence at birth of hemophilia in males: A meta-analytic approach using national registries. Ann Intern Med (2019) 171:540–6. doi: 10.7326/M19-1208
2. White GC, Rosendaal F, Aledort LM, Lusher JM, Rothschild C, Ingerslev J, et al. Definitions in hemophilia. recommendation of the scientific subcommittee on factor VIII and factor IX of the scientific and standardization committee of the international society on thrombosis and haemostasis. Thromb Haemost (2001) 85:560. doi: 10.1097/00024382-200115030-00013
3. Srivastava A, Brewer AK, Mauser-Bunschoten EP, Key NS, Kitchen S, Llinas A, et al. Guidelines for the management of hemophilia. Haemophilia (2013) 19:e1–47. doi: 10.1111/j.1365-2516.2012.02909.x
4. Astermark J. FVIII inhibitors: Pathogenesis and avoidance. Blood (2015) 125:2045–51. doi: 10.1182/blood-2014-08-535328
5. Brackmann HH, Gormsen J. Massive factor-VIII infusion in haemophiliac with factor-VIII inhibitor, high responder. 1977. Haemophilia (2010) 16:2–3. doi: 10.1111/j.1365-2516.2010.02266.x
6. Hart DP, Alamelu J, Bhatnagar N, Biss T, Collins PW, Hall G, et al. Immune tolerance induction in severe haemophilia a: A UKHCDO inhibitor and paediatric working party consensus update. Haemophilia (2021) 27:932–7. doi: 10.1111/hae.14381
7. Valentino LA, Kempton CL, Kruse-Jarres R, Mathew P, Meeks SL, Reiss UM, et al. US Guidelines for immune tolerance induction in patients with haemophilia a and inhibitors. Haemophilia (2015) 21:559–67. doi: 10.1111/hae.12730
8. Srivastava A, Santagostino E, Dougall A, Kitchen S, Sutherland M, Pipe SW, et al. WFH guidelines for the management of hemophilia, 3rd edition. Haemophilia (2020) 26 Suppl 6:1–158. doi: 10.1111/hae.14046
9. Giangrande PLF, Hermans C, O'Mahony B, de Kleijn P, Bedford M, Batorova A, et al. European Principles of inhibitor management in patients with haemophilia. Orphanet J Rare Dis (2018) 13:66. doi: 10.1186/s13023-018-0800-z
10. Van Dijk K, van der Bom JG, Bax KN, van der Zee DC, Van Den Berg MH. Use of implantable venous access devices in children with severe hemophilia: Benefits and burden. Haematologica (2004) 89:189–94. doi: 10.3324/haematolo.2004.%25x
11. Liu W, Xue F, Poon MC, Chen L, Jin Z, Zhang L, et al. Current status of haemophilia inhibitor management in mainland China: a haemophilia treatment centres survey on treatment preferences and real-world clinical practices. Br J Haematol (2021) 194:750–8. doi: 10.1111/bjh.17677
12. Antun A, Monahan PE, Manco-Johnson MJ, Callaghan MU, Kanin M, Knoll C, et al. Inhibitor recurrence after immune tolerance induction: A multicenter retrospective cohort study. J Thromb Haemost (2015) 13:1980–8. doi: 10.1111/jth.13143
13. Whelan SF, Hofbauer CJ, Horling FM, Allacher P, Wolfsegger MJ, Oldenburg J, et al. Distinct characteristics of antibody responses against factor VIII in healthy individuals and in different cohorts of hemophilia a patients. Blood (2013) 121:1039–48. doi: 10.1182/blood-2012-07-444877
14. Reipert BM, van Helden PM, Schwarz HP, Hausl C. Mechanisms of action of immune tolerance induction against factor VIII in patients with congenital haemophilia a and factor VIII inhibitors. Br J Haematol (2007) 136:12–25. doi: 10.1111/j.1365-2141.2006.06359.x
15. Lacroix-Desmazes S, Moreau A, Sooryanarayana, Bonnemain C, Stieltjes N, Pashov A, et al. Catalytic activity of antibodies against factor VIII in patients with hemophilia a. Nat Med (1999) 5:1044–7. doi: 10.1038/12483
16. Kempton CL, White GC. How we treat a hemophilia a patient with a factor VIII inhibitor. Blood (2009) 113:11–7. doi: 10.1182/blood-2008-06-160432
17. Hofbauer CJ, Whelan SF, Hirschler M, Allacher P, Horling FM, Lawo JP, et al. Affinity of FVIII-specific antibodies reveals major differences between neutralizing and nonneutralizing antibodies in humans. Blood (2015) 125:1180–8. doi: 10.1182/blood-2014-09-598268
18. Lavigne-Lissalde G, Rothschild C, Pouplard C, Lapalud P, Gruel Y, Schved JF, et al. Characteristics, mechanisms of action, and epitope mapping of anti-factor VIII antibodies. Clin Rev Allergy Immunol (2009) 37:67–79. doi: 10.1007/s12016-009-8119-0
19. Lindgren A, Wadenvik H, Tengborn L. Characterization of inhibitors to FVIII with an ELISA in congenital and acquired haemophilia a. Haemophilia (2002) 8:644–8. doi: 10.1046/j.1365-2516.2002.00674.x
20. Cannavo A, Valsecchi C, Garagiola I, Palla R, Mannucci PM, Rosendaal FR, et al. Nonneutralizing antibodies against factor VIII and risk of inhibitor development in severe hemophilia a. Blood (2017) 129:1245–50. doi: 10.1182/blood-2016-06-720086
21. Reipert BM, Gangadharan B, Hofbauer CJ, Berg V, Schweiger H, Bowen J, et al. The prospective hemophilia inhibitor PUP study reveals distinct antibody signatures prior to FVIII inhibitor development. Blood Adv (2020) 4:5785–96. doi: 10.1182/bloodadvances.2020002731
22. Peyvandi F, Garagiola I, Young G. The past and future of haemophilia: Diagnosis, treatments, and its complications. Lancet (2016) 388:187–97. doi: 10.1016/s0140-6736(15)01123-x
23. Garagiola I, Palla R, Peyvandi F. Risk factors for inhibitor development in severe hemophilia a. Thromb Res (2018) 168:20–7. doi: 10.1016/j.thromres.2018.05.027
24. Gouw SC, van der Bom JG, Van Den Berg HM, Zewald RA, Ploos Van Amstel JK, Mauser-Bunschoten EP. Influence of the type of F8 gene mutation on inhibitor development in a single centre cohort of severe haemophilia a patients. Haemophilia (2011) 17:275–81. doi: 10.1111/j.1365-2516.2010.02420.x
25. Sun J, Li Z, Huang K, Ai D, Li G, Xie X, et al. F8 gene mutation spectrum in severe hemophilia a with inhibitors: A large cohort data analysis from a single center in China. Res Pract Thromb Haemost (2022) 6:e12723. doi: 10.1002/rth2.12723
26. Lessard S, He C, Rajpal DK, Klinger K, Loh C, Harris T, et al. Genome-wide association study and gene-based analysis of participants with hemophilia a and inhibitors in the my life, our future research repository. Front Med (Lausanne) (2022) 9:903838. doi: 10.3389/fmed.2022.903838
27. Pavlova A, Delev D, Lacroix-Desmazes S, Schwaab R, Mende M, Fimmers R, et al. Impact of polymorphisms of the major histocompatibility complex class II, interleukin-10, tumor necrosis factor-alpha and cytotoxic T-lymphocyte antigen-4 genes on inhibitor development in severe hemophilia a. J Thromb Haemost (2009) 7:2006–15. doi: 10.1111/j.1538-7836.2009.03636.x
28. Karim AF, Soltis AR, Sukumar G, Konigs C, Ewing NP, Dalgard CL, et al. Hemophilia a inhibitor subjects show unique PBMC gene expression profiles that include up-regulated innate immune modulators. Front Immunol (2020) 11:1219. doi: 10.3389/fimmu.2020.01219
29. Lorenzo JI, López A, Altisent C, Aznar JA. Incidence of factor VIII inhibitors in severe haemophilia: The importance of patient age. Br J Haematol (2001) 113:600–3. doi: 10.1046/j.1365-2141.2001.02828.x
30. Strauss T, Lubetsky A, Ravid B, Bashari D, Luboshitz J, Lalezari S, et al. Recombinant factor concentrates may increase inhibitor development: A single centre cohort study. Haemophilia (2011) 17:625–9. doi: 10.1111/j.1365-2516.2010.02464.x
31. Chen J, Schroeder JA, Luo XF, Shi QZ. The impact of von willebrand factor on factor VIII memory immune responses. Blood Adv (2017) 1:1565–74. doi: 10.1182/bloodadvances.2017009209
32. Delignat S, Repessé Y, Navarrete AM, Meslier Y, Gupta N, Christophe OD, et al. Immunoprotective effect of von willebrand factor towards therapeutic factor VIII in experimental haemophilia a. Haemophilia (2012) 18:248–54. doi: 10.1111/j.1365-2516.2011.02679.x
33. Peyvandi F, Mannucci PM, Garagiola I, El-Beshlawy A, Elalfy M, Ramanan V, et al. A randomized trial of factor VIII and neutralizing antibodies in hemophilia a. N Engl J Med (2016) 374:2054–64. doi: 10.1056/NEJMoa1516437
34. Peyvandi F, Mannucci PM, Palla R, Rosendaal FR. SIPPET: methodology, analysis and generalizability. Haemophilia (2017) 23:353–61. doi: 10.1111/hae.13203
35. Gouw SC, van der Bom JG, Marijke van den Berg H. Treatment-related risk factors of inhibitor development in previously untreated patients with hemophilia a: the CANAL cohort study. Blood (2007) 109:4648–54. doi: 10.1182/blood-2006-11-056291
36. Gouw SC, van den Berg HM, Fischer K, Auerswald G, Carcao M, Chalmers E, et al. Intensity of factor VIII treatment and inhibitor development in children with severe hemophilia a: The RODIN study. Blood (2013) 121:4046–55. doi: 10.1182/blood-2012-09-457036
37. Wight J, Paisley S. The epidemiology of inhibitors in haemophilia a: a systematic review. Haemophilia (2003) 9:418–35. doi: 10.1046/j.1365-2516.2003.00780.x
38. Lim MY, Cheng D, Recht M, Kempton CL, Key NS. Inhibitors and mortality in persons with nonsevere hemophilia a in the united states. Blood Adv (2020) 4:4739–47. doi: 10.1182/bloodadvances.2020002626
39. van Velzen AS, Eckhardt CL, Peters M, Leebeek FWG, Escuriola-Ettingshausen C, Hermans C, et al. Intensity of factor VIII treatment and the development of inhibitors in non-severe hemophilia a patients: Results of the INSIGHT case-control study. J Thromb Haemost (2017) 15:1422–9. doi: 10.1111/jth.13711
40. Abdi A, Linari S, Pieri L, Voorberg J, Fijnvandraat K, Castaman G. Inhibitors in nonsevere hemophilia a: What is known and searching for the unknown. Semin Thromb Hemost (2018) 44:568–77. doi: 10.1055/s-0037-1621717
41. Eckhardt CL, van Velzen AS, Peters M, Astermark J, Brons PP, Castaman G, et al. Factor VIII gene (F8) mutation and risk of inhibitor development in nonsevere hemophilia a. Blood (2013) 122:1954–62. doi: 10.1182/blood-2013-02-483263
42. Gouw SC, van den Berg HM, le Cessie S, van der Bom JG. Treatment characteristics and the risk of inhibitor development: A multicenter cohort study among previously untreated patients with severe hemophilia a. J Thromb Haemost (2007) 5:1383–90. doi: 10.1111/j.1538-7836.2007.02595.x
43. van den Berg HM, Fischer K, Carcao M, Chambost H, Kenet G, Kurnik K, et al. Timing of inhibitor development in more than 1000 previously untreated patients with severe hemophilia a. Blood (2019) 134:317–20. doi: 10.1182/blood.2019000658
44. Mauser-Bunschoten EP, Den Uijl IE, Schutgens RE, Roosendaal G, Fischer K. Risk of inhibitor development in mild haemophilia a increases with age. Haemophilia (2012) 18:263–7. doi: 10.1111/j.1365-2516.2011.02629.x
45. Abdi A, Eckhardt CL, van Velzen AS, Vuong C, Coppens M, Castaman G, et al. Treatment-related risk factors for inhibitor development in non-severe hemophilia a after 50 cumulative exposure days: A case-control study. J Thromb Haemost (2021) 19:2171–81. doi: 10.1111/jth.15419
46. Eckhardt CL, Loomans JI, van Velzen AS, Peters M, Mauser-Bunschoten EP, Schwaab R, et al. Inhibitor development and mortality in non-severe hemophilia a. J Thromb Haemost (2015) 13:1217–25. doi: 10.1111/jth.12990
47. Matzinger P. Tolerance, danger, and the extended family. Annu Rev Immunol (1994) 12:991–1045. doi: 10.1146/annurev.iy.12.040194.005015
48. Pradeu T, Cooper EL. The danger theory: 20 years later. Front Immunol (2012) 3:287. doi: 10.3389/fimmu.2012.00287
49. Schep SJ, Boes M, Schutgens REG, van Vulpen LFD. An update on the 'danger theory' in inhibitor development in hemophilia a. Expert Rev Hematol (2019) 12:335–44. doi: 10.1080/17474086.2019.1604213
50. Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know about danger. J Leukoc Biol (2007) 81:1–5. doi: 10.1189/jlb.0306164
51. Kaczorowski DJ, Mollen KP, Edmonds R, Billiar TR. Early events in the recognition of danger signals after tissue injury. J Leukoc Biol (2008) 83:546–52. doi: 10.1189/jlb.0607374
52. Pulendran B. Immune activation: death, danger and dendritic cells. Curr Biol (2004) 14:R30–32. doi: 10.1016/j.cub.2003.12.020
53. Lövgren KM, Christensen KR, Majewski W, Østrup O, Skov S, Wiinberg B. Acute haemarthrosis in the haemophilia a rat generates a local and systemic proinflammatory response. Thromb Haemost (2017) 117:2092–104. doi: 10.1160/th17-03-0149
54. Ragni MV. FVIII. CD4, and liaisons dangereuses. Blood (2011) 117:6060–1. doi: 10.1182/blood-2011-04-348458
55. Kurnik K, Bidlingmaier C, Engl W, Chehadeh H, Reipert B, Auerswald G. New early prophylaxis regimen that avoids immunological danger signals can reduce FVIII inhibitor development. Haemophilia (2010) 16:256–62. doi: 10.1111/j.1365-2516.2009.02122.x
56. Waters B, Qadura M, Burnett E, Chegeni R, Labelle A, Thompson P, et al. Anti-CD3 prevents factor VIII inhibitor development in hemophilia a mice by a regulatory CD4+CD25+-dependent mechanism and by shifting cytokine production to favor a Th1 response. Blood (2009) 113:193–203. doi: 10.1182/blood-2008-04-151597
57. Chen Y, Luo X, Schroeder JA, Chen J, Baumgartner CK, Hu J, et al. Immune tolerance induced by platelet-targeted factor VIII gene therapy in hemophilia a mice is CD4 T cell mediated. J Thromb Haemost (2017) 15:1994–2004. doi: 10.1111/jth.13800
58. Jing W, Chen J, Cai Y, Chen Y, Schroeder JA, Johnson BD, et al. Induction of activated T follicular helper cells is critical for anti-FVIII inhibitor development in hemophilia a mice. Blood Adv (2019) 3:3099–110. doi: 10.1182/bloodadvances.2019000650
59. Tieu P, Chan A, Matino D. Molecular mechanisms of inhibitor development in hemophilia. Mediterr J Hematol Infect Dis (2020) 12:e2020001. doi: 10.4084/MJHID.2020.001
60. Ragni MV, Bontempo FA, Lewis JH. Disappearance of inhibitor to factor VIII in HIV-infected hemophiliacs with progression to AIDS or severe ARC. Transfusion (1989) 29:447–9. doi: 10.1046/j.1537-2995.1989.29589284147.x
61. Bray GL, Kroner BL, Arkin S, Aledort LW, Hilgartner MW, Eyster ME, et al. Loss of high-responder inhibitors in patients with severe hemophilia a and human immunodeficiency virus type 1 infection: A report from the multi-center hemophilia cohort study. Am J Hematol (1993) 42:375–9. doi: 10.1002/ajh.2830420408
62. Shi Q, Carman CV, Chen YY, Sage PT, Xue F, Liang XM, et al. Unexpected enhancement of FVIII immunogenicity by endothelial expression in lentivirus-transduced and transgenic mice. Blood Adv (2020) 4:2272–85. doi: 10.1182/bloodadvances.2020001468
63. Bovenschen N, Mertens K, Hu L, Havekes LM, van Vlijmen BJ. LDL receptor cooperates with LDL receptor-related protein in regulating plasma levels of coagulation factor VIII. Blood (2005) 106:906–912. doi: 10.1182/blood-2004-11-4230
64. Sarafanov AG, Ananyeva NM, Shima M, Saenko EL. Cell surface heparan sulfate proteoglycans participate in factor VIII catabolism mediated by low density lipoprotein receptor-related protein. J Biol Chem (2001) 276:11970–9. doi: 10.1074/jbc.M008046200
65. van Haren SD, Wroblewska A, Fischer K, Voorberg J, Herczenik E. Requirements for immune recognition and processing of factor VIII by antigen-presenting cells. Blood Rev (2012) 26:43–9. doi: 10.1016/j.blre.2011.10.001
66. Herczenik E, van Haren SD, Wroblewska A, Kaijen P, van den Biggelaar M, Meijer AB, et al. Uptake of blood coagulation factor VIII by dendritic cells is mediated via its C1 domain. J Allergy Clin Immunol (2012) 129:501–9. doi: 10.1016/j.jaci.2011.08.029
67. Dasgupta S, Repessé Y, Bayry J, Navarrete AM, Wootla B, Delignat S, et al. VWF protects FVIII from endocytosis by dendritic cells and subsequent presentation to immune effectors. Blood (2007) 109:610–2. doi: 10.1182/blood-2006-05-022756
68. Ramakrishnan R, Davidowitz A, Balu-Iyer SV. Exposure of FVIII in the presence of phosphatidyl serine reduces generation of memory b-cells and induces regulatory T-Cell-Mediated hyporesponsiveness in hemophilia a mice. J Pharm Sci (2015) 104:2451–6. doi: 10.1002/jps.24513
69. Gaitonde P, Peng A, Straubinger RM, Bankert RB, Balu-Lyer ST. Phosphatidylserine reduces immune response against human recombinant factor VIII in hemophilia a mice by regulation of dendritic cell function. Clin Immunol (2011) 138:135–45. doi: 10.1016/j.clim.2010.10.006
70. Gaitonde P, Ramakrishnan R, Chin J, Kelleher RJ, Bankert RB, Balu-Iyer SV. Exposure to factor VIII protein in the presence of phosphatidylserine induces hypo-responsiveness toward factor VIII challenge in hemophilia a mice. J Biol Chem (2013) 288:17051–6. doi: 10.1074/jbc.C112.396325
71. Nguyen NH, Dingman RK, Balu-Iyer SV. Tolerogenic form of factor VIII to prevent inhibitor development in the treatment of hemophilia a. J Thromb Haemost (2021) 19:2744–50. doi: 10.1111/jth.15497
72. Glassman FY, Balu-Iyer SV. Subcutaneous administration of lyso-phosphatidylserine nanoparticles induces immunological tolerance towards factor VIII in a hemophilia a mouse model. Int J Pharm (2018) 548:642–8. doi: 10.1016/j.ijpharm.2018.07.018
73. Vander Kooi A, Wang S, Fan MN, Chen A, Zhang J, Chen CY, et al. Influence of n-glycosylation in the a and c domains on the immunogenicity of factor VIII. Blood Adv (2022) 6:4271–82. doi: 10.1182/bloodadvances.2021005758
74. Navarrete A, Dasgupta S, Delignat S, Caligiuri G, Christophe OD, Bayry J, et al. Splenic marginal zone antigen-presenting cells are critical for the primary allo-immune response to therapeutic factor VIII in hemophilia a. J Thromb Haemost (2009) 7:1816–23. doi: 10.1111/j.1538-7836.2009.03571.x
75. Zerra PE, Cox C, Baldwin WH, Patel SR, Arthur CM, Lollar P, et al. Marginal zone b cells are critical to factor VIII inhibitor formation in mice with hemophilia a. Blood (2017) 130:2559–68. doi: 10.1182/blood-2017-05-782912
76. Rossi G, Sarkar J, Scandella D. Long-term induction of immune tolerance after blockade of CD40-CD40L interaction in a mouse model of hemophilia a. Blood (2001) 97:2750–7. doi: 10.1182/blood.v97.9.2750
77. Wang X, Terhorst C, Herzog RW. In vivo induction of regulatory T cells for immune tolerance in hemophilia. Cell Immunol (2016) 301:18–29. doi: 10.1016/j.cellimm.2015.10.001
78. Kim YC, Zhang AH, Su Y, Rieder SA, Rossi RJ, Ettinger RA, et al. Engineered antigen-specific human regulatory T cells: Immunosuppression of FVIII-specific T- and b-cell responses. Blood (2015) 125:1107–15. doi: 10.1182/blood-2014-04-566786
79. Miao CH. Immunomodulation for inhibitors in hemophilia a: the important role of treg cells. Expert Rev Hematol (2010) 3:469–83. doi: 10.1586/ehm.10.33
80. Cao O, Loduca PA, Herzog RW. Role of regulatory T cells in tolerance to coagulation factors. J Thromb Haemost (2009) 7 Suppl 1:88–91. doi: 10.1111/j.1538-7836.2009.03417.x
81. Becker-Gotot J, Meissner M, Kotov V, Jurado-Mestre BJ, Maione A, Pannek A, et al. Immune tolerance against infused FVIII in hemophilia a is mediated by PD-L1 + regulatory T cells. J Clin Invest (2022) 132:e159925. doi: 10.1172/JCI159925
82. Doshi BS, Rana J, Castaman G, Shaheen MA, Kaczmarek R, Butterfield JS, et al. B cell-activating factor modulates the factor VIII immune response in hemophilia a. J Clin Invest (2021) 131:e142906. doi: 10.1172/JCI142906
83. van Helden PM, Kaijen PH, Fijnvandraat K, van den Berg HM, Voorberg J. Factor VIII-specific memory b cells in patients with hemophilia a. J Thromb Haemost (2007) 5:2306–8. doi: 10.1111/j.1538-7836.2007.02736.x
84. van Helden PM, Van Haren SD, Fijnvandraat K, van den Berg HM, Voorberg J. Factor VIII-specific b cell responses in haemophilia a patients with inhibitors. Haemophilia (2010) 16:35–43. doi: 10.1111/j.1365-2516.2010.02215.x
85. Bernasconi NL, Traggiai E, Lanzavecchia A. Maintenance of serological memory by polyclonal activation of human memory b cells. Science (2002) 298:2199–202. doi: 10.1126/science.1076071
86. Tarlinton D, Radbruch A, Hiepe F, Dorner T. Plasma cell differentiation and survival. Curr Opin Immunol (2008) 20:162–9. doi: 10.1016/j.coi.2008.03.016
87. Brackmann HH, White GC, Berntorp E, Andersen T, Escuriola-Ettingshausen C. Immune tolerance induction: What have we learned over time? Haemophilia (2018) 24:3–14. doi: 10.1111/hae.13445
88. Scott DW, Pratt KP. Factor VIII: Perspectives on immunogenicity and tolerogenic strategies. Front Immunol (2019) 10:3078. doi: 10.3389/fimmu.2019.03078
89. Hay CR, DiMichele DM. International immune tolerance s. the principal results of the international immune tolerance study: A randomized dose comparison. Blood (2012) 119:1335–44. doi: 10.1182/blood-2011-08-369132
90. Merlin S, Follenzi A. Escape or fight: Inhibitors in hemophilia a. Front Immunol (2020) 11:476. doi: 10.3389/fimmu.2020.00476
91. Lu Y, Chen Z, Dai J, Wu X, Gu H, Li Z, et al. Maternal microchimerism protects hemophilia a patients from inhibitor development. Blood Adv (2020) 4:1867–9. doi: 10.1182/bloodadvances.2020001832
92. Chen Y, Schroeder JA, Chen J, Luo X, Baumgartner CK, Montgomery RR, et al. The immunogenicity of platelet-derived FVIII in hemophilia a mice with or without preexisting anti-FVIII immunity. Blood (2016) 127:1346–54. doi: 10.1182/blood-2015-08-662916
93. Wieland I. Emicizumab for all pediatric patients with severe hemophilia a. Hamostaseologie (2022) 42:104–15. doi: 10.1055/a-1727-1384
94. Escuriola Ettingshausen C, Sidonio RF Jr. Design of an international investigator-initiated study on MOdern treatment of inhibitor-positiVe pATiEnts with haemophilia a (MOTIVATE). Ther Adv Hematol (2021) 12:20406207211032452. doi: 10.1177/20406207211032452
95. Oldenburg J, Mahlangu JN, Kim B, Schmitt C, Callaghan MU, Young G, et al. Emicizumab prophylaxis in hemophilia a with inhibitors. N Engl J Med (2017) 377:809–18. doi: 10.1056/NEJMoa1703068
96. Uchida N, Sambe T, Yoneyama K, Fukazawa N, Kawanishi T, Kobayashi S, et al. A first-in-human phase 1 study of ACE910, a novel factor VIII-mimetic bispecific antibody, in healthy subjects. Blood (2016) 127:1633–41. doi: 10.1182/blood-2015-06-650226
97. Shima M, Hanabusa H, Taki M, Matsushita T, Sato T, Fukutake K, et al. Factor VIII–mimetic function of humanized bispecific antibody in hemophilia a. N Engl J Med (2016) 374:2044–53. doi: 10.1056/NEJMoa1511769
98. Oldenburg J, Mahlangu JN, Bujan W, Trask P, Callaghan MU, Young G, et al. The effect of emicizumab prophylaxis on health-related outcomes in persons with haemophilia a with inhibitors: HAVEN 1 study. Haemophilia (2019) 25:33–44. doi: 10.1111/hae.13618
99. Young G, Liesner R, Chang T, Sidonio R, Oldenburg J, Jiménez-Yuste V, et al. A multicenter, open-label phase 3 study of emicizumab prophylaxis in children with hemophilia a with inhibitors. Blood (2019) 134:2127–38. doi: 10.1182/blood.2019001869
100. Mahlangu J, Oldenburg J, Paz-Priel I, Negrier C, Niggli M, Mancuso ME, et al. Emicizumab prophylaxis in patients who have hemophilia a without inhibitors. N Engl J Med (2018) 379:811–22. doi: 10.1056/NEJMoa1803550
101. Pipe SW, Shima M, Lehle M, Shapiro A, Chebon S, Fukutake K, et al. Efficacy, safety, and pharmacokinetics of emicizumab prophylaxis given every 4 weeks in people with haemophilia a (HAVEN 4): A multicentre, open-label, non-randomised phase 3 study. Lancet Haematol (2019) 6:e295–305. doi: 10.1016/s2352-3026(19)30054-7
102. Yang R, Wang S, Wang X, Sun J, Chuansumrit A, Zhou J, et al. Prophylactic emicizumab for hemophilia a in the Asia-pacific region: A randomized study (HAVEN 5). Res Pract Thromb Haemost (2022) 6:e12670. doi: 10.1002/rth2.12670
103. Shima M, Nagao A, Taki M, Matsushita T, Oshida K, Amano K, et al. Long-term safety and efficacy of emicizumab for up to 5.8 years and patients' perceptions of symptoms and daily life: A phase 1/2 study in patients with severe haemophilia a. Haemophilia (2021) 27:81–9. doi: 10.1111/hae.14205
104. Blair HA. Emicizumab: A review in haemophilia a. Drugs (2019) 79:1697–707. doi: 10.1007/s40265-019-01200-2
105. Valsecchi C, Gobbi M, Beeg M, Adams T, Castaman G, Schiavone L, et al. Characterization of the neutralizing anti-emicizumab antibody in a patient with hemophilia a and inhibitor. J Thromb Haemost (2021) 19:711–8. doi: 10.1111/jth.15226
106. Jiménez-Yuste V PF, Klamroth R, Castaman G, Shanmukhaiah C, Rangarajan S, García Chavez J, et al. Final analysis of the STASEY trial: A single-arm, multicenter, open-label, phase III clinical trial evaluating the safety and tolerability of emicizumab prophylaxis in persons with hemophilia a (PwHA) with factor (F)VIII inhibitors [abstract]. Res Pract Thromb Haemost (2021) 5(Suppl 2):e12589. doi: 10.1002/rth2.12589
107. Harkins Druzgal C, Kizilocak H, Brown J, Sennett M, Young G. Neutralizing antidrug antibody to emicizumab in a patient with severe hemophilia a with inhibitors: New case with detailed laboratory evaluation. J Thromb Haemost (2020) 18:2205–8. doi: 10.1111/jth.14957
108. Hassan E, Jonathan L, Jayashree M. Real-world experience on the tolerability and safety of emicizumab prophylaxis in paediatric patients with severe haemophilia a with and without FVIII inhibitors. Haemophilia (2021) 27:e698–703. doi: 10.1111/hae.14432
109. Callaghan MU, Negrier C, Paz-Priel I, Chang T, Chebon S, Lehle M, et al. Long-term outcomes with emicizumab prophylaxis for hemophilia a with or without FVIII inhibitors from the HAVEN 1-4 studies. Blood (2021) 137:2231–42. doi: 10.1182/blood.2020009217
110. Ostergaard H, Lund J, Greisen PJ, Kjellev S, Henriksen A, Lorenzen N, et al. A factor VIIIa-mimetic bispecific antibody, Mim8, ameliorates bleeding upon severe vascular challenge in hemophilia a mice. Blood (2021) 138:1258–68. doi: 10.1182/blood.2020010331
111. Lauritzen B, Bjelke M, Bjorkdahl O, Bloem E, Keane K, Kjalke M, et al. A novel next-generation FVIIIa mimetic, Mim8, has a favorable safety profile and displays potent pharmacodynamic effects: Results from safety studies in cynomolgus monkeys. J Thromb Haemost (2022) 20:1312–24. doi: 10.1111/jth.15682
112. Peterson JA, Maroney SA, Mast AE. Targeting TFPI for hemophilia treatment. Thromb Res (2016) 141 Suppl 2:S28–30. doi: 10.1016/s0049-3848(16)30359-0
113. Mahlangu JN. Progress in the development of anti-tissue factor pathway inhibitors for haemophilia management. Front Med (Lausanne) (2021) 8:670526. doi: 10.3389/fmed.2021.670526
114. Mancuso ME, Ingham SJM, Kunze M. Befovacimab, an anti-tissue factor pathway inhibitor antibody: Early termination of the multiple-dose, dose-escalating phase 2 study due to thrombosis. Haemophilia (2022) 28:702–12. doi: 10.1111/hae.14595
115. Chowdary P, Lethagen S, Friedrich U, Brand B, Hay C, Abdul Karim F, et al. Safety and pharmacokinetics of anti-TFPI antibody (concizumab) in healthy volunteers and patients with hemophilia: a randomized first human dose trial. J Thromb Haemost (2015) 13:743–54. doi: 10.1111/jth.12864
116. Shapiro AD, Angchaisuksiri P, Astermark J, Benson G, Castaman G, Chowdary P, et al. Subcutaneous concizumab prophylaxis in hemophilia a and hemophilia A/B with inhibitors: Phase 2 trial results. Blood (2019) 134:1973–82. doi: 10.1182/blood.2019001542
117. Shapiro AD. Concizumab: a novel anti-TFPI therapeutic for hemophilia. Blood Adv (2021) 5:279. doi: 10.1182/bloodadvances.2019001140
118. Forest R. Novo Nordisk resumes phase 3 trials of concizumab in hemophilia a and b. Hemophilia News Today (2021).
119. Shapiro AD, Angchaisuksiri P, Astermark J, Benson G, Castaman G, Eichler H, et al. Long-term efficacy and safety of subcutaneous concizumab prophylaxis in hemophilia a and hemophilia A/B with inhibitors. Blood Adv (2022) 6:3422–32. doi: 10.1182/bloodadvances.2021006403
120. Mahlangu JN, Lamas JL, Morales JC, Malan DR, Salek SZ, Wang M, et al. A phase 1b/2 clinical study of marstacimab, targeting human tissue factor pathway inhibitor, in haemophilia. Br J Haematol (2022). doi: 10.1111/bjh.18420
121. Sehgal A, Barros S, Ivanciu L, Cooley B, Qin J, Racie T, et al. An RNAi therapeutic targeting antithrombin to rebalance the coagulation system and promote hemostasis in hemophilia. Nat Med (2015) 21:492–7. doi: 10.1038/nm.3847
122. Pasi KJ, Rangarajan S, Georgiev P, Mant T, Creagh MD, Lissitchkov T, et al. Targeting of antithrombin in hemophilia a or b with RNAi therapy. N Engl J Med (2017) 377:819–28. doi: 10.1056/NEJMoa1616569
123. Swan D, Mahlangu J, Thachil J. Non-factor therapies for bleeding disorders: A primer for the general haematologist. EJHaem (2022) 3:584–95. doi: 10.1002/jha2.442
124. Ragni MV, Georgiev P, Mant T, Creagh M, Lissitchkov T, Bevan D, et al. Fitusiran, an investigational RNAi therapeutic targeting antithrombin for the treatment of hemophilia: Updated results from a phase 1 and phase 1/2 extension study in patients without inhibitors. Blood (2016) 128:2572. doi: 10.1182/blood.V128.22.2572.2572
125. Pasi KJ, Lissitchkov T, Mamonov V, Mant T, Timofeeva M, Bagot C, et al. Targeting of antithrombin in hemophilia a or b with investigational siRNA therapeutic fitusiran-results of the phase 1 inhibitor cohort. J Thromb Haemost (2021) 19:1436–46. doi: 10.1111/jth.15270
126. Machin N, Ragni MV. An investigational RNAi therapeutic targeting antithrombin for the treatment of hemophilia a and b. J Blood Med (2018) 9:135–40. doi: 10.2147/JBM.S159297
127. Mancuso ME, Apte S, Hermans C. Managing invasive procedures in haemophilia patients with limited resources, extended half-life concentrates or non-replacement therapies in 2022. Haemophilia (2022) 28 Suppl 4:93–102. doi: 10.1111/hae.14551
128. Aymonnier K, Kawecki C, Venisse L, Boulaftali Y, Christophe OD, Lenting PJ, et al. Targeting protease nexin-1, a natural anticoagulant serpin, to control bleeding and improve hemostasis in hemophilia. Blood (2019) 134:1632–44. doi: 10.1182/blood.2019000281
129. Sanrattana W, Smits S, Barendrecht AD, van Kleef ND, El Otmani H, Zivkovic M, et al. (TaSER): A dual-action antithrombotic agent that targets platelets for SERPIN delivery. J Thromb Haemost (2022) 20:353–65. doi: 10.1111/jth.15554
130. Sanrattana W, Sefiane T, Smits S, van Kleef ND, Fens MH, Lenting PJ, et al. A reactive center loop-based prediction platform to enhance the design of therapeutic SERPINs. Proc Natl Acad Sci USA (2021) 118:e2108458118. doi: 10.1073/pnas.2108458118
131. Polderdijk SG, Adams TE, Ivanciu L, Camire RM, Baglin TP, Huntington JA. Design and characterization of an APC-specific serpin for the treatment of hemophilia. Blood (2017) 129:105–13. doi: 10.1182/blood-2016-05-718635
132. Aymonnier K, Kawecki C, Arocas V, Boulaftali Y, Bouton MC. Serpins, new therapeutic targets for hemophilia. Thromb Haemost (2021) 121:261–9. doi: 10.1055/s-0040-1716751
133. Zhao XY, Wilmen A, Wang D, Wang X, Bauzon M, Kim JY, et al. Targeted inhibition of activated protein c by a non-active-site inhibitory antibody to treat hemophilia. Nat Commun (2020) 11:2992. doi: 10.1038/s41467-020-16720-9
134. Magisetty J, Kondreddy V, Keshava S, Das K, Esmon CT, Pendurthi UR, et al. Selective inhibition of activated protein c anticoagulant activity protects against hemophilic arthropathy in mice. Blood (2022) 139:2830–41. doi: 10.1182/blood.2021013119
135. Shi Q, Weiler H. Blocking hemophilic arthropathy. Blood (2022) 139:2734–5. doi: 10.1182/blood.2022015776
136. Park CY, Lee DR, Sung JJ, Kim DW. Genome-editing technologies for gene correction of hemophilia. Hum Genet (2016) 135:977–81. doi: 10.1007/s00439-016-1699-x
137. Rangarajan S, Walsh L, Lester W, Perry D, Madan B, Laffan M, et al. AAV5-factor VIII gene transfer in severe hemophilia a. N Engl J Med (2017) 377:2519–30. doi: 10.1056/NEJMoa1708483
138. Ozelo MC, Wong WY. Gene therapy for hemophilia a. reply. N Engl J Med (2022) 386:2247. doi: 10.1056/NEJMc2204934
139. Pasi KJ, Rangarajan S, Mitchell N, Lester W, Symington E, Madan B, et al. Multiyear follow-up of AAV5-hFVIII-SQ gene therapy for hemophilia a. N Engl J Med (2020) 382:29–40. doi: 10.1056/NEJMoa1908490
140. Fassel H, McGuinn C. Haemophilia: factoring in new therapies. Br J Haematol (2021) 194:835–50. doi: 10.1111/bjh.17580
141. Butterfield JSS, Hege KM, Herzog RW, Kaczmarek R. A molecular revolution in the treatment of hemophilia. Mol Ther (2020) 28:997–1015. doi: 10.1016/j.ymthe.2019.11.006
142. Xue F, Li H, Wu X, Liu W, Zhang F, Tang D, et al. Safety and activity of an engineered, liver-tropic adeno-associated virus vector expressing a hyperactive padua factor IX administered with prophylactic glucocorticoids in patients with haemophilia b: a single-centre, single-arm, phase 1, pilot trial. Lancet Haematol (2022) 9:e504–13. doi: 10.1016/S2352-3026(22)00113-2
143. Samelson-Jones BJ, George LA. Adeno-associated virus gene therapy for hemophilia. Annu Rev Med (2022). doi: 10.1146/annurev-med-043021-033013
144. Samelson-Jones BJ, Arruda VR. Translational potential of immune tolerance induction by AAV liver-directed factor VIII gene therapy for hemophilia a. Front Immunol (2020) 11:618. doi: 10.3389/fimmu.2020.00618
145. Finn JD, Ozelo MC, Sabatino DE, Franck HW, Merricks EP, Crudele JM, et al. Eradication of neutralizing antibodies to factor VIII in canine hemophilia a after liver gene therapy. Blood (2010) 116:5842–8. doi: 10.1182/blood-2010-06-288001
146. Arruda VR, Samelson-Jones BJ. Gene therapy for immune tolerance induction in hemophilia with inhibitors. J Thromb Haemost (2016) 14:1121–34. doi: 10.1111/jth.13331
147. Sack BK, Merchant S, Markusic DM, Nathwani AC, Davidoff AM, Byrne BJ, et al. Transient b cell depletion or improved transgene expression by codon optimization promote tolerance to factor VIII in gene therapy. PLoS One (2012) 7:e37671. doi: 10.1371/journal.pone.0037671
148. Liu CL, Ye P, Lin J, Butts CL, Miao CH. Anti-CD20 as the b-cell targeting agent in a combined therapy to modulate anti-factor VIII immune responses in hemophilia a inhibitor mice. Front Immunol (2014) 4:502. doi: 10.3389/fimmu.2013.00502
149. Liu CL, Lyle MJ, Shin SC, Miao CH. Strategies to target long-lived plasma cells for treating hemophilia a inhibitors. Cell Immunol (2016) 301:65–73. doi: 10.1016/j.cellimm.2016.01.005
150. Biswas M, Palaschak B, Kumar SRP, Rana J, Markusic DM. B cell depletion eliminates FVIII memory b cells and enhances AAV8-coF8 immune tolerance induction when combined with rapamycin. Front Immunol (2020) 11:1293. doi: 10.3389/fimmu.2020.01293
151. Doshi BS, Raffini LJ, George LA. Combined anti-CD20 and mTOR inhibition with factor VIII for immune tolerance induction in hemophilia a patients with refractory. J Thromb Haemost (2020) 18:848–52. doi: 10.111/jth.14740
152. Shi Q. Platelet-targeted gene therapy for hemophilia. Mol Ther Methods Clin Dev (2018) 9:100–8. doi: 10.1016/j.omtm.2018.01.011
153. Chen Y, Schroeder JA, Gao C, Li J, Hu J, Shi Q. In vivo enrichment of genetically manipulated platelets for murine hemophilia b gene therapy. J Cell Physiol (2021) 236:354–65. doi: 10.1002/jcp.29861
154. Chen Y, Schroeder JA, Kuether EL, Zhang G, Shi Q. Platelet gene therapy by lentiviral gene delivery to hematopoietic stem cells restores hemostasis and induces humoral immune tolerance in FIX(null) mice. Mol Ther (2014) 22:169–77. doi: 10.1038/mt.2013.197
155. Kuether EL, Schroeder JA, Fahs SA, Cooley BC, Chen Y, Montgomery RR, et al. Lentivirus-mediated platelet gene therapy of murine hemophilia a with pre-existing anti-factor VIII immunity. J Thromb Haemost (2012) 10:1570–80. doi: 10.1111/j.1538-7836.2012.04791.x
156. Miao CH. Hemophilia a gene therapy via intraosseous delivery of factor VIII-lentiviral vectors. Thromb J (2016) 14:41. doi: 10.1186/s12959-016-0105-1
157. Shi Q, Fahs SA, Wilcox DA, Kuether EL, Morateck PA, Mareno N, et al. Syngeneic transplantation of hematopoietic stem cells that are genetically modified to express factor VIII in platelets restores hemostasis to hemophilia a mice with preexisting FVIII immunity. Blood (2008) 112:2713–21. doi: 10.1182/blood-2008-02-138214
158. Du LM, Nurden P, Nurden AT, Nichols TC, Bellinger DA, Jensen ES, et al. Platelet-targeted gene therapy with human factor VIII establishes haemostasis in dogs with haemophilia a. Nat Commun (2013) 4:2773. doi: 10.1038/ncomms3773
159. Sherman A, Su J, Lin S, Wang X, Herzog RW, Daniell H. Suppression of inhibitor formation against FVIII in a murine model of hemophilia a by oral delivery of antigens bioencapsulated in plant cells. Blood (2014) 124:1659–68. doi: 10.1182/blood-2013-10-528737
160. Li LY, Han J, Wu L, Fang C, Li WG, Gu JM, et al. Alterations of gut microbiota diversity, composition and metabonomics in testosterone-induced benign prostatic hyperplasia rats. Mil Med Res (2022) 9:12. doi: 10.1186/s40779-022-00373-4
161. Kumar SRP, Wang X, Avuthu N, Bertolini TB, Terhorst C, Guda C, et al. Role of small intestine and gut microbiome in plant-based oral tolerance for hemophilia. Front Immunol (2020) 11:844. doi: 10.3389/fimmu.2020.00844
162. Landwehr-Kenzel S, Muller-Jensen L, Kuehl JS, Abou-El-Enein M, Hoffmann H, Muench S, et al. Adoptive transfer of ex vivo expanded regulatory T cells improves immune cell engraftment and therapy-refractory chronic GvHD. Mol Ther (2022) 30:2298–314. doi: 10.1016/j.ymthe.2022.02.025
163. Good Z, Spiegel JY, Sahaf B, Malipatlolla MB, Ehlinger ZJ, Kurra S, et al. Post-infusion CAR TReg cells identify patients resistant to CD19-CAR therapy. Nat Med (2022) 28:1860–71. doi: 10.1038/s41591-022-01960-7
164. Yoon J, Schmidt A, Zhang AH, Konigs C, Kim YC, Scott DW. FVIII-specific human chimeric antigen receptor T-regulatory cells suppress T- and b-cell responses to FVIII. Blood (2017) 129:238–45. doi: 10.1182/blood-2016-07-727834
165. Zhang Q, Lu W, Liang CL, Chen Y, Liu H, Qiu F, et al. Chimeric antigen receptor (CAR) treg: A promising approach to inducing immunological tolerance. Front Immunol (2018) 9:2359. doi: 10.3389/fimmu.2018.02359
Keywords: Hemophilia A, inhibitor, immune tolerance, pathogenesis, factor VIII
Citation: Luo L, Zheng Q, Chen Z, Huang M, Fu L, Hu J, Shi Q and Chen Y (2022) Hemophilia a patients with inhibitors: Mechanistic insights and novel therapeutic implications. Front. Immunol. 13:1019275. doi: 10.3389/fimmu.2022.1019275
Received: 14 August 2022; Accepted: 09 November 2022;
Published: 08 December 2022.
Edited by:
Moanaro Biswas, Indiana University, United StatesReviewed by:
Radek Kaczmarek, Indiana University Bloomington, United StatesBhavya Doshi, Children’s Hospital of Philadelphia, United States
Copyright © 2022 Luo, Zheng, Chen, Huang, Fu, Hu, Shi and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yingyu Chen, Y2hlbnlpbmd5dUB5YWhvby5jb20=; Qizhen Shi, cXNoaUB2ZXJzaXRpLm9yZw==
†These authors have contributed equally to this work
‡ORCID: Qizhen Shi, orcid.org/0000-0003-1548-0708
Yingyu Chen, orcid.org/0000-0003-1243-3607