 Simon Le Gallou1,2†Faustine Lhomme1,3†Jonathan M. Irish3,4Anna Mingam1Celine Pangault1,2Celine Monvoisin2
Simon Le Gallou1,2†Faustine Lhomme1,3†Jonathan M. Irish3,4Anna Mingam1Celine Pangault1,2Celine Monvoisin2 Juliette Ferrant2Imane Azzaoui1Delphine Rossille1,2Krimo Bouabdallah5Gandhi Damaj6Guillaume Cartron7Pascal Godmer8Steven Le Gouill9René-Olivier Casasnovas10Thierry Jo Molina11Roch Houot2,12Thierry Lamy2,12
Juliette Ferrant2Imane Azzaoui1Delphine Rossille1,2Krimo Bouabdallah5Gandhi Damaj6Guillaume Cartron7Pascal Godmer8Steven Le Gouill9René-Olivier Casasnovas10Thierry Jo Molina11Roch Houot2,12Thierry Lamy2,12 Karin Tarte1,2
Karin Tarte1,2 Thierry Fest1,2
Thierry Fest1,2 Mikael Roussel1,2*
Mikael Roussel1,2*- 1Centre Hospitalier Universitaire de Rennes, Pôle Biologie, Rennes, France
- 2Institut National de la Santé et de la Recherche Médicale, Unité Mixte de Recherche U1236, Université Rennes 1, Etablissement Français du Sang Bretagne, Rennes, France
- 3Department of Cell and Developmental Biology, Vanderbilt University School of Medicine, Nashville, TN, United States
- 4Department of Pathology, Microbiology and Immunology, Vanderbilt University School of Medicine, Nashville, TN, United States
- 5Centre Hospitalier Universitaire de Bordeaux, Service d’Hématologie Clinique, Bordeaux, France
- 6Centre Hospitalier Universitaire de Caen, Service d’Hématologie Clinique, Caen, France
- 7Centre Hospitalier Universitaire de Montpellier, Service d’Hématologie Clinique, Montpellier, France
- 8Centre Hospitalier de Bretagne Atlantique, Unité d’Hématologie Clinique, Vannes, France
- 9Centre Hospitalier Universitaire de Nantes, Service d’Hématologie Clinique, Institut National de la Sante et de la Recherche Medicale, Centre de Recherche en Cancérologie et Immunologie Nantes Angers (INSERM CCRCINA) Nantes-Angers, NeXT Université de Nantes, Nantes, France
- 10Centre Hospitalier Universitaire de Dijon, Service d’Hématologie Clinique, Dijon, France
- 11Asistance Publique, Hopitaux de Paris (APHP), Necker, Service d’Anatomopathologie, Sorbonne Université, Paris, France
- 12Centre Hospitalier Universitaire de Rennes, Service d’Hématologie Clinique, Rennes, France
Absolute count of circulating monocytes has been proposed as an independent prognostic factor in diffuse large B-cell lymphoma (DLBCL). However, monocyte nomenclature includes various subsets with pro-, anti-inflammatory, or suppressive functions, and their clinical relevance in DLBCL has been poorly explored. Herein, we broadly assessed circulating monocyte heterogeneity in 91 DLBCL patients. Classical- (cMO, CD14pos CD16neg) and intermediate- (iMO, CD14pos CD16pos) monocytes accumulated in DLBCL peripheral blood and exhibited an inflammatory phenotype. On the opposite, nonclassical monocytes (ncMOSlanpos, CD14low CD16pos Slanneg and ncMOSlanneg, CD14low CD16pos, Slanneg) were decreased in peripheral blood. Tumor-conditioned monocytes presented similarities with ncMO phenotype from DLBCL and were prone to migrate in response to CCL5 and CXCL12, and presented similarities with DLBCL-infiltrated myeloid cells, as defined by mass cytometry. Finally, we demonstrated the adverse value of an accumulation of nonclassical monocytes in 2 independent cohorts of DLBCL.
Introduction
Circulating monocytes are classified by their CD14 and CD16 expression as classical- (cMO, CD14pos CD16neg), intermediate- (iMO, CD14pos CD16pos), and nonclassical- monocytes (ncMO, CD14low CD16pos) (1). In addition, Slan expression [6-Sulfo LacNac, which is a carbohydrate modification of P-selectin glycoprotein ligand-1 (PSGL-1)], allows the sub-classification of ncMO Slanpos (CD14low CD16pos Slanpos) and ncMO Slanneg (CD14low CD16pos Slanneg) (2, 3). Lastly monocytic myeloid derived suppressor cells (M-MDSC, CD14pos HLA-DRlow) found in acute or chronic inflammatory context, including cancers, are defined by an impairment of T- and NK- effector functions (4). This nomenclature reflects pro-inflammatory, anti-inflammatory, or suppressive functions described for monocytes (5, 6).
In diffuse large B-cell lymphoma (DLBCL), tumor microenvironment (TME), myeloid cells are supportive of the neoplastic process (7–10). In blood from DLBCL patients, an increase in circulating monocytes is a marker of adverse prognosis (11–15). However, so far monocytes were considered as a whole, and few studies analyzed the monocyte subsets and their clinical relevance even if their intrinsic functions are known to be different. Among monocyte subsets: i) Slanpos monocytes were increased and displayed high rituximab mediated antibody-dependent cellular cytotoxicity (16); ii) an increase in CD16pos or CD11bposCX3CR1pos monocytes predicted poor progression free- and overall- survival (17, 18); iii) CD14posCD163posPD-L1pos monocytes were increased (19); and finally iv) functional M-MDSCs were enriched in peripheral blood and predicted poor event-free survival (20–22). In DLBCL tumor, the myeloid compartment heterogeneity was recently approached by high dimensional analysis revealing distinct macrophage phenotype across lymphoma subtypes (23).
In light with the observation that various monocyte subsets are involved in the biology of DLBCL, we investigated the canonical cMO, iMO, ncMO Slanpos, and ncMO Slanneg subsets in two large cohorts of patients. We quantified these subsets, analyzed their phenotype and functions as well as the clinical relevance of these cells. We found here that in DLBCL, ncMO are prone to migrate into tissues and that their increase in peripheral blood is associated with an adverse prognosis.
Methods
Samples
A cohort of 91 DLBCL patients at diagnosis from the BMS-LyTRANS clinical trial (ClinicalTrials.gov Identifier: NCT01287923) was used in this study. Clinical characteristics of DLBCL patients enrolled in this training cohort are listed in Table 1. Patients with previous corticosteroid treatment were excluded from this study. As controls, age-matched heathy donors (HD, n = 49), follicular lymphomas (n = 9), mantle cell lymphomas (n = 9), chronic lymphocytic leukemias (n = 11), and marginal zone lymphomas (n = 10) were included. Part of these samples (DLBCL and HD) were used in a previous work (22). Prognosis scores were validated in a second cohort of 155 DLBCL patients from the recently published GAINED trial (ClinicalTrials.gov Identifier: NCT01659099) (24). Clinical characteristics of DLBCL patients enrolled in this validation cohort are listed in Table 1. The research protocol was conducted under French legal guidelines and fulfilled the requirements of the local institutional ethics committee and biosecurity procedures.
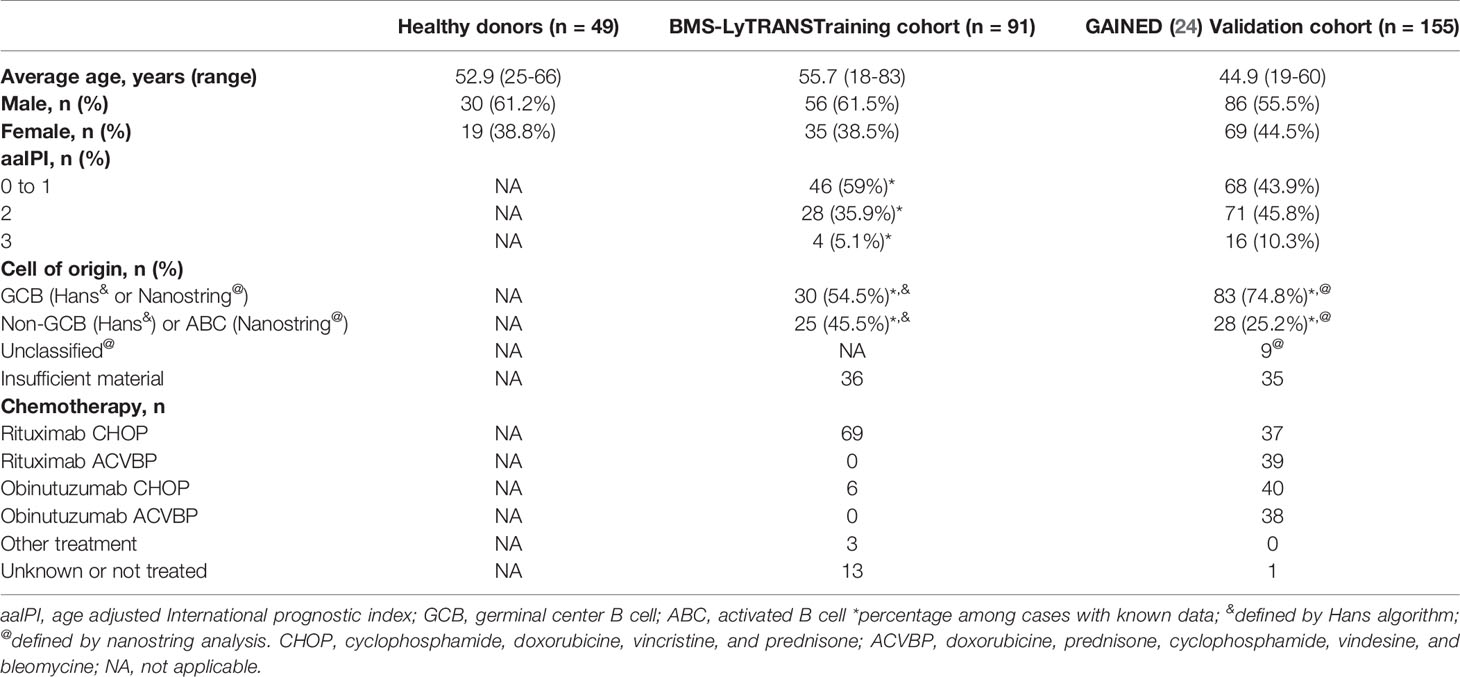
Table 1 Patient’s characteristics at baseline.
Fluorescent Flow Cytometry Analysis
Blood samples were collected on heparin tubes. Flow cytometry analysis of M-MDSCs, cMO, iMO, and ncMO were performed on whole blood (300 µL/tube) with the antibody panel shown Table S1 and the gating strategy defined Figure S1. Absolute counts were obtained by using Flow-Count beads (Beckman Coulter, Brea, CA). An erythrocytes lysis (Uti-Lyse Dako, Carpinteria, CA) was performed before analysis by flow cytometry (Navios, Beckman Coulter). Analyses were performed using Kaluza software (Beckman Coulter).
In Vitro Culture
OCI-Ly3 and OCI-Ly19 cell lines were cultured in OCI-Ly medium (IMDM supplemented with 10% human AB serum,1% penicillin–streptomycin (Invitrogen) and 50 µM of β-Mercaptoethanol). For supernatants, OCI-Ly 3 and 19 were seeded at 3 x106 cells/ml in OCI-Ly medium for 24h (37°C, 5% CO2). Supernatants were obtained after centrifugation and were frozen at -80°C until migration assay. For control of migration assay, OCI-Ly medium were incubated at (37°C, 5% CO2) without cells for 24h before centrifugation and freezing.
Monocytes were obtained from PBMCs by elutriation before cryopreservation (plate-forme DTC; CIC Biotherapie, Nantes, France). Monocytes were thawed and resuspended at 4 x106 cells/mL in RPMI 1640 (Invitrogen, Carlsband, CA, USA) supplemented with 10% FCS and antibiotics (Invitrogen) and then diluted at 2 x106 cells/mL by adding, as control, the OCI-Ly medium (IMDM supplemented with 10% human AB serum,1% penicillin–streptomycin and 50 µM of β-Mercaptoethanol, or OCI-Ly3 or OCI-Ly19 supernatant. Two mL of cell suspension were seeded in a 6-well plate during 4 days before mass cytometry analysis or migration assay.
Mass Cytometry Analysis
Cell labeling and mass-cytometry analysis were performed as previously described. (25–27) Briefly, cells were incubated with 25 µM cisplatin (Fluidigm San Francisco, CA, USA). Then, 5 x106 cells were washed in PBS (HyClone Laboratories, Logan, UT, USA) containing 1% BSA (Thermo Fisher Scientific) and stained in 100 µL PBS and BSA 1%-containing Antibody cocktail. Cells were stained for 30 min at RT with the antibodies (Table S2) Cells were washed twice in PBS - BSA 1% before fixation in 1.6% PFA, and permeabilization with methanol (Electron Microscopy Sciences, Hatfield, PA, USA). After incubating overnight at -20°C in MeOH, cells were washed twice with PBS -BSA 1% and stained 20 min with iridium intercalator (Fluidigm, Sunnyvale, CA, US). Finally, cells were washed twice with PBS and twice with diH2O before acquisition a CyTOF 2.0 mass cytometer (Fluidigm). Mass cytometry raw data were deposited in Flow Repository (http://flowrepository.org/id/FR-FCM-Z4N6).
Data Processing and Analysis
Data analysis was performed using the workflow previously developed (23). Briefly, after acquisition, intrafile signal drift was normalized and.fcs files were obtained using CyTOF software. To diminish batch effects, all files were normalized on EQ Beads (Fluidigm) using the premessa R package (https://github.com/ParkerICI/premessa). Raw median intensity values were transformed to a hyperbolic arcsine (arcsinh) and then analysis was performed using Cytobank software (Beckman Coulter, Brea, CA, USA).
Migration Assay
At day 4 of monocyte culture with OCI-Ly3, or OCI-Ly19 supernatant, or control culture medium, cells were collected and washed twice in PBS before starvation during 1 hour (37°C, 5% CO2) at 106 cells/mL in RPMI 1% HSA. Then, cells were washed once, counted and 100 µL of cells at 106 cells/mL were added to the upper compartment of Transwell chambers with 5 µM pore filters (Corning Incorporated, Kennebunk, ME, USA). The lower chamber contained a chemiokine among CCL2 (R&D Systems, 30 ng/ml), CCL3 (R&D Systems, 20 ng/ml), CCL5 (R&D Systems, 30 ng/ml), CCL22 (R&D Systems, 20 ng/ml), CXCL5 (R&D Systems, 20 ng/ml), CXCL12 (R&D Systems, 20 ng/ml), or RPMI 1640 1% HSA as control (corresponding to Basal migration). Cells in the lower chamber were collected after 5h (37°C, 5% CO2) and the absolute number of viable (DAPI negative) monocytes was quantified by flow cytometry using Precision Count Beads™ (Biolegend Inc, San Diego, CA, USA). The absolute number of cells (Total Cells) added to the upper compartment at the beginning of migration assay was also determined for each condition (control, OCI-Ly3 and OCI-Ly19) with Precision Count Beads™. Percentage of cells that have specifically migrated against a chemokine was calculated as follow: Specific migration = (Lower chamber - Basal migration)/Total cells x 100. The migration represents the variation of this percentage with OCI-Ly supernatants compared to control culture medium (% of specific migration with supernatant – % of specific migration with medium) for each chemokine.
Cell Sorting
cMO (CD19neg CD3neg CD335neg CD45pos CD14high CD16neg), iMO (CD19neg CD3neg CD335neg CD45pos CD14high CD16pos), ncMO Slanpos (CD19neg CD3neg CD335neg CD45pos CD14low CD16pos Slanpos), and ncMO Slanneg (CD19neg CD3neg CD335neg CD45pos CD14low CD16pos Slanneg) were sorted from thawed PBMC of DLBCL patients and HD using an ARIA II (FACSAria, BD Biosciences).
Quantitative Real-Time PCR
Total RNA was extracted usingNucleospin® RNA XS kit (Macherey-Nagel, Duren, Germany). cDNA was then generated using Fluidigm Reverse Transcription Master Mix (Fluidigm). The qPCR were performed in triplicate using 96.96 Dynamic Array™ IFCs and the BioMark™ HD System from Fluidigm. For each sample, the mean CT value for the gene of interest was calculated, normalized to the geometric mean value of the 2 housekeeping genes (CDKN1B and ELF1) (Table S3), and compared to the median value obtained from the reference population (HD cMO or iMO, and DLBCL ncMO) using the 2-ddCT method. Results were expressed as the ratio of sample mean to reference mean for each gene.
Statistical Analysis
Statistical analyses were performed with GraphPad Prism 8.4.3 software (GraphPad Software, San Diego, CA, USA) using Pearson correlation, Wilcoxon, Mann-Whitney, Ordinary one-way ANOVA with Tukey’s multiple comparisons test, and Fishers’s exact tests as appropriate. Optimal thresholds were defined with the maxstat package, log-rank tests were performed with the survminer package, cox model for univariate and multivariate analysis were performed with the survival package. Analyses were generated with R v4.0.3, using Rstudio v1.3.1093.
Data Sharing Statement
For original data, please contact the corresponding author. Mass cytometry raw data were deposited in Flow Repository (http://flowrepository.org/id/FR-FCM-Z4N6. QPCR raw data are included in Table S4.
Results
cMO and iMO Are Increased in DLBCL
We have previously shown that M-MDSCs accumulated in DLBCL peripheral blood (22). However, this increase accounts for only a part of the total monocyte accumulation suggesting that additional monocyte subsets are also increased in DLBCL samples (Figure 1A). We quantified the absolute count of the 4 circulating monocyte subsets M-MDSC, cMO, iMO, and ncMO. M-MDSC, cMO, and iMO were increased in DLBCL when compared to HDs (P <.05, median: 5.75 x106 cells/L vs 2.8 x106 cells/L, 348.8 x106 cells/L vs 274.1 x106 cells/L, and 34.4 x106 cells/L vs 26.1 x106 cells/L; respectively). Conversely, ncMO were significantly decreased in DLBCL when compared to HD (P <.0001, median: 17.1 x106 cells/L vs 36.1 x106 cells/L; Figure 1B). Noteworthy, whereas the increase of cMO and iMO was also found in other B cell lymphomas, this decrease in ncMO was specific of DLBCL (Figure S2). Then, we wondered in which monocyte subset M-MDSCs were included. Of note M-MDSC count was correlated with total monocyte (R = .57, P <.0001), cMO (R = .61, P <.0001), but was not correlated with iMO and ncMO (Figure 1C) No correlation were observed between MO subsets in HD samples (data not shown). Regarding CD14 and CD16 expression, M-MDSCs were essentially aligned with the cMO phenotype and to a lesser extent to iMO (Figure 1D). Altogether, these results confirmed that in addition to MDSCs, cMO and iMO were also involved in the monocyte increase observed in DLBCL patients.
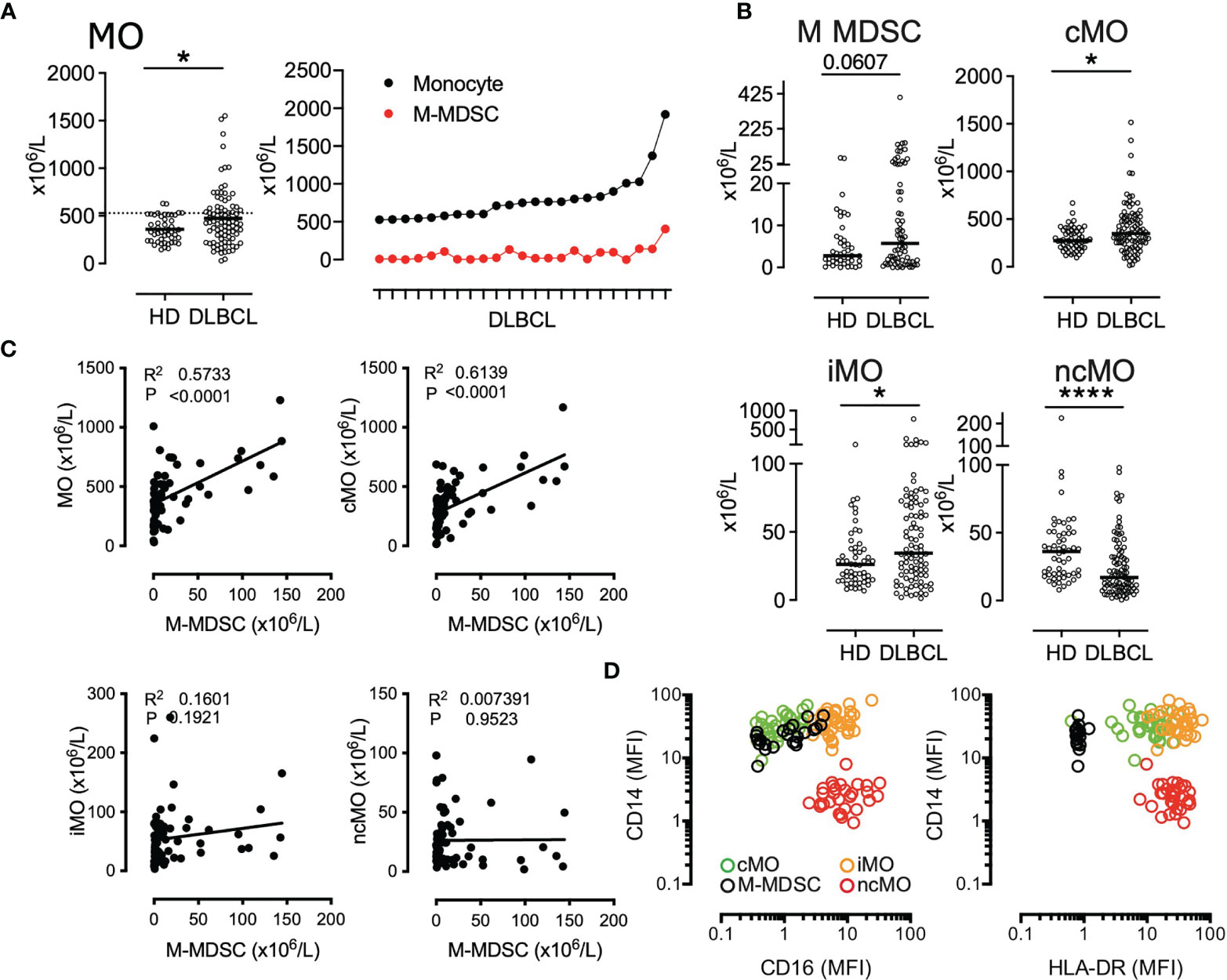
Figure 1 cMO and iMO are increased in peripheral blood from DLBCL. (A) Monocyte absolute counts in HD (n = 49) and DLBCL (n = 88). For the 23 DLBCL samples with monocyte above 528 x106 monocytes/L (corresponding to the 90th percentile of HD), proportion of M-MDSC within monocyte (B) M-MDSC in HD (n = 43) and DLBCL (n = 69) and monocyte subset counts [classical- (cMO), intermediate- (iMO), and nonclassical- (ncMO)] in peripheral blood from HD (n = 55) and DLBCL patients (n = 91). (C) Pearson correlation between M-MDSC and monocyte (MO), cMO, iMO, and ncMO (n = 68). (D) Mean fluorescent intensity (MFI) for CD14, CD16, and HLA-DR. Each dot represents a DLBCL sample (n = 33) colored by monocyte subset (MDSC, cMO, iMO, and ncMO). Mann-Whitney test were performed. *P <. 05, ****P < .0001.
DLBCL cMO and iMO Share a Common Inflammatory Phenotype
To further identify the immune properties of monocyte subsets, we sorted cMO and iMO from DLBCL (n=7) and HD (n=4) samples. Gene expression was assessed by high throughput qPCR on 71 genes involved in myeloid biology (22) (Tables S3, S4, and Figure S3). Of note, 6 cMO and 5 iMO out of 7 DLBCL were clustered (Figure 2A and Figure S3). DLBCL cMO and DLBCL iMO were significantly enriched for inflammatory genes (FCGR3A, CD36, FCGR1A, CYBB, AIM2, STAT6, FCGR2A, CCR2, NLRC4, S100A8, and CD14 genes), when compared to the corresponding subsets in HDs (P <.05, ∣log2FC∣>1) (Figure 2B). In addition, S100A9 and CD163 were also increased in DLBCL cMO, whereas CD33 and ITGAM were enriched only in DLBCL iMO. For both subsets, SLC7A11, CD274, and CXCL1 were expressed at lower levels in DLBCLs (Figure 2B). Biological processes enriched in DLBCL cMO and iMO included apoptosis, production of ROS, immune response, and phagocytosis (Figure 2C). By flow cytometry, we showed that cMO and iMO from DLBCL displayed a higher expression of CD64 and CCR2 (P <.05), without variation in HLA-DR and CD163 (Figure 2D). Altogether, these results suggested that, in DLBCL, cMO and iMO share a common deregulated inflammatory phenotype.
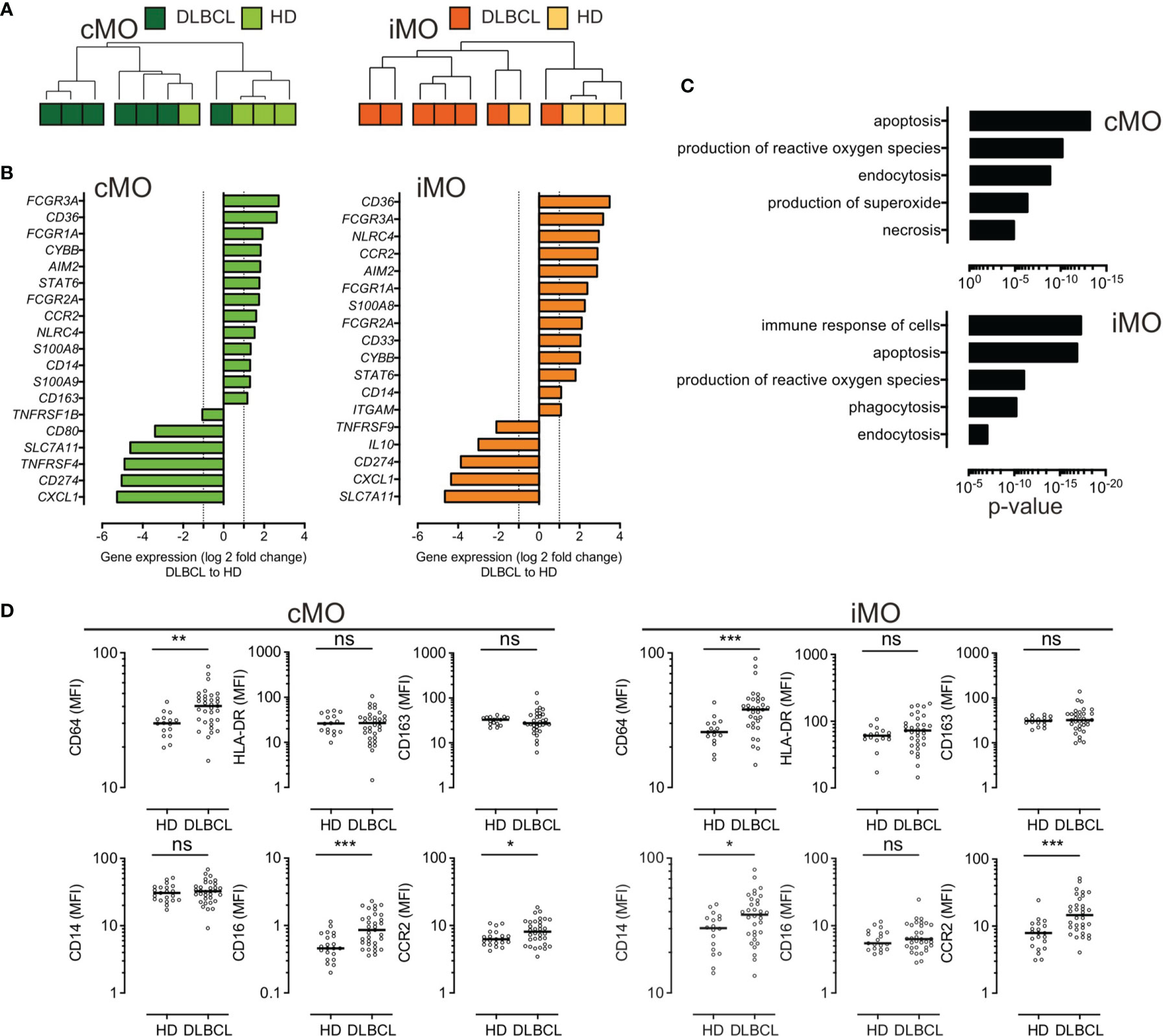
Figure 2 Circulating cMO and iMO share a common inflammatory phenotype, in DLBCL. (A) Unsupervised hierarchical clustering of classical- (cMO) and intermediate- (iMO) monocytes, from HD (n = 4) and DLBCL (n = 7) samples. See Table S3 for a list of genes analyzed on monocyte subsets after cell sorting. Pearson’s correlation and complete linkage was employed. (B) Transcripts differentially expressed (P <.05; ∣log2FC∣ > 1) between DLBCLs and HDs, for cMO and iMO. (C) Predicted top 5 biological processes increased for cMO and iMO from DLBCL (Ingenuity Pathway Analysis, z-score > 2.5, ranked by p-value). (D) Mean fluorescence (MFI) for CD64, HLA-DR, CD163, CD14, CD16, and CCR2 for HD (n = 16) and DLBCL (n = 33) samples. Mann-Whitney test were performed. *P < .05, **P < .01, ***P < .001, ns, non-significant.
DLBCL ncMO Are Decreased in Peripheral Blood and Exhibit an Inflammatory- and Tolerogenic-Like- Phenotype
We then focused on ncMO in DLBCL samples and found that both subsets of ncMO (ncMO Slanpos and ncMO Slanneg) were decreased in DLBCLs when compared to HDs [ncMO Slanpos median at 3.3 x106 cells/L vs 9.1 x106 cells/L (P <.001) and ncMO Slanneg median at 19 x106 cells/L vs 23.7 x106 cells/L (P <.05), respectively] (Figure 3A). No increase of apoptosis was detected in ncMO from DLBCL patients (data not shown). Then, sorted ncMO Slanpos and ncMO Slanneg from DLBCL and HD samples were analyzed by high-throughput Q-PCR (Figure S4). To explore the similarities between ncMO Slanpos, ncMO Slanneg, iMO, and cMO, we performed an unsupervised hierarchical clustering on the DLBCL samples. For 6 out of 7 patients, cMO and iMO were separated from ncMO independently of Slan expression (Figure 3B and Figure S5). In addition, ncMO Slanpos and ncMO Slanneg exhibited tolerogenic genes (PDCD1LG2, IL10, IDO, CD274, AGER, TNFAIP6) (Figure 3C and Figure S5), most of these genes were not expressed in HD ncMO (Figure S6). In DLBCL, cMO and iMO in one hand and ncMO Slanpos and ncMO Slanneg in the other hand shared similar gene expression (Figure 2A and Figure S5), thus we compared the gene expression between ncMO, irrespectively of the Slan status, and both cMO and iMO. DLBCL ncMO were enriched for both inflammatory (CXCL10, AIM2, IL12A) and tolerogenic (PDCD1LG2, IL10, IDO, CD274, AGER) genes (P <.05, ∣log2FC∣ > 1) compared to cMO and iMO (Figure 3C). Biological processes involved by genes enriched in ncMO from DLBCL patients were growth of tumor, inhibition of cells, and chemotaxis (Figures 3C, D).
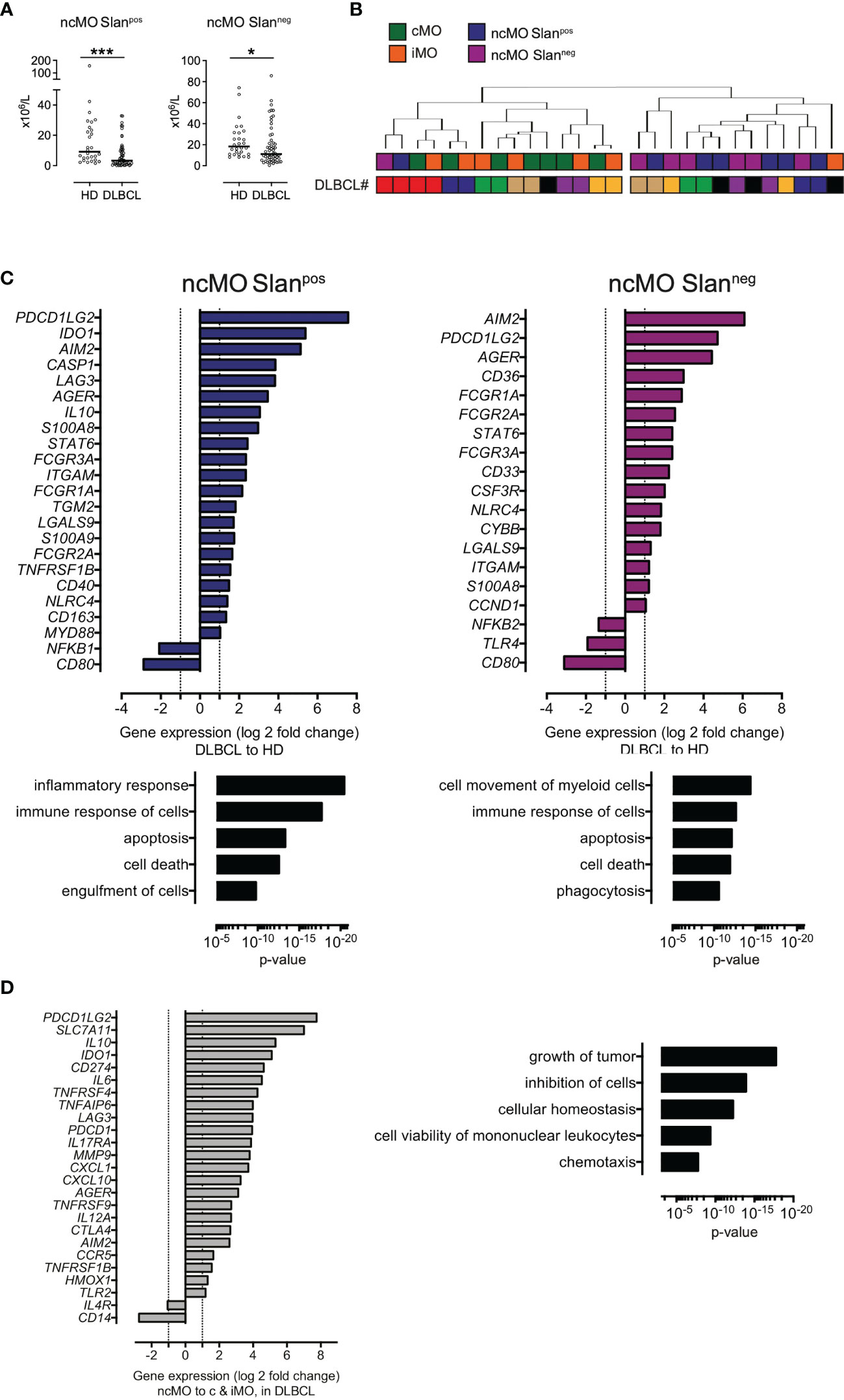
Figure 3 ncMO are decreased in peripheral blood but exhibit an inflammatory- and tolerogenic- phenotype. (A) Nonclassical Slanpos or Slanneg (ncMOSlanpos and ncMoSlanneg) monocytes from HDs (n = 28) and DLBCLs (n = 56). Mann-Whitney test were performed. *P <.05, ***P <.001. (B) Unsupervised hierarchical clustering of classical- (cMO), intermediate- (iMO), nonclassical Slanpos- or Slanneg- (ncMOSlanpos and ncMoSlanneg) monocytes from DLBCL (n = 7) samples. DLBCL identity (#). List of genes analyzed on monocyte subsets after cell sorting is on Table S3. Pearson’s correlation and complete linkage was employed. (C) Transcripts differentially expressed (P <.05; ∣log2FC∣ > 1) between DLBCLs (n=7) and HDs (n=4), for ncMO Slanpos and ncMO Slanneg. Predicted top 5 biological processes increased for ncMO Slanpos and ncMO Slanneg from DLBCL when compared to HD samples (Ingenuity Pathway Analysis, z-score > 2.5, ranked by p-value). (D) Transcripts (P <.05; ∣log2FC∣ > 1) enriched in ncMO compared to cMO and iMO for DLBCL. Predicted top 5 biological processes increased for ncMO compared to cMO and iMO, from DLBCL (Ingenuity Pathway Analysis, z-score > 2.5, ranked by p-value).
Tumor Conditioned Monocytes Give Rise to MO Prone to Migrate in Response to CCL5 and CXCL12
In order to evaluate how tumor B cells contribute directly to the phenotype of DLBCL monocytes, we cultured monocytes from HD with supernatants from the DLBCL cell lines OCI-Ly3 and OCI-Ly19. After coculture, we analyzed the monocyte phenotype by mass cytometry (23, 25) and noticed an increased expression of CD16, Slan, CD64, CD163. We concluded that tumor conditioned monocytes triggered the cMO to ncMO transition (Figure 4A). Then we wondered if these cells were prone to migrate into tissue. Tumor-conditioned monocytes demonstrated an increase in in vitro migration in response to CCL5 and CXCL12 when compared to non-conditioned monocytes (Figure 4B).
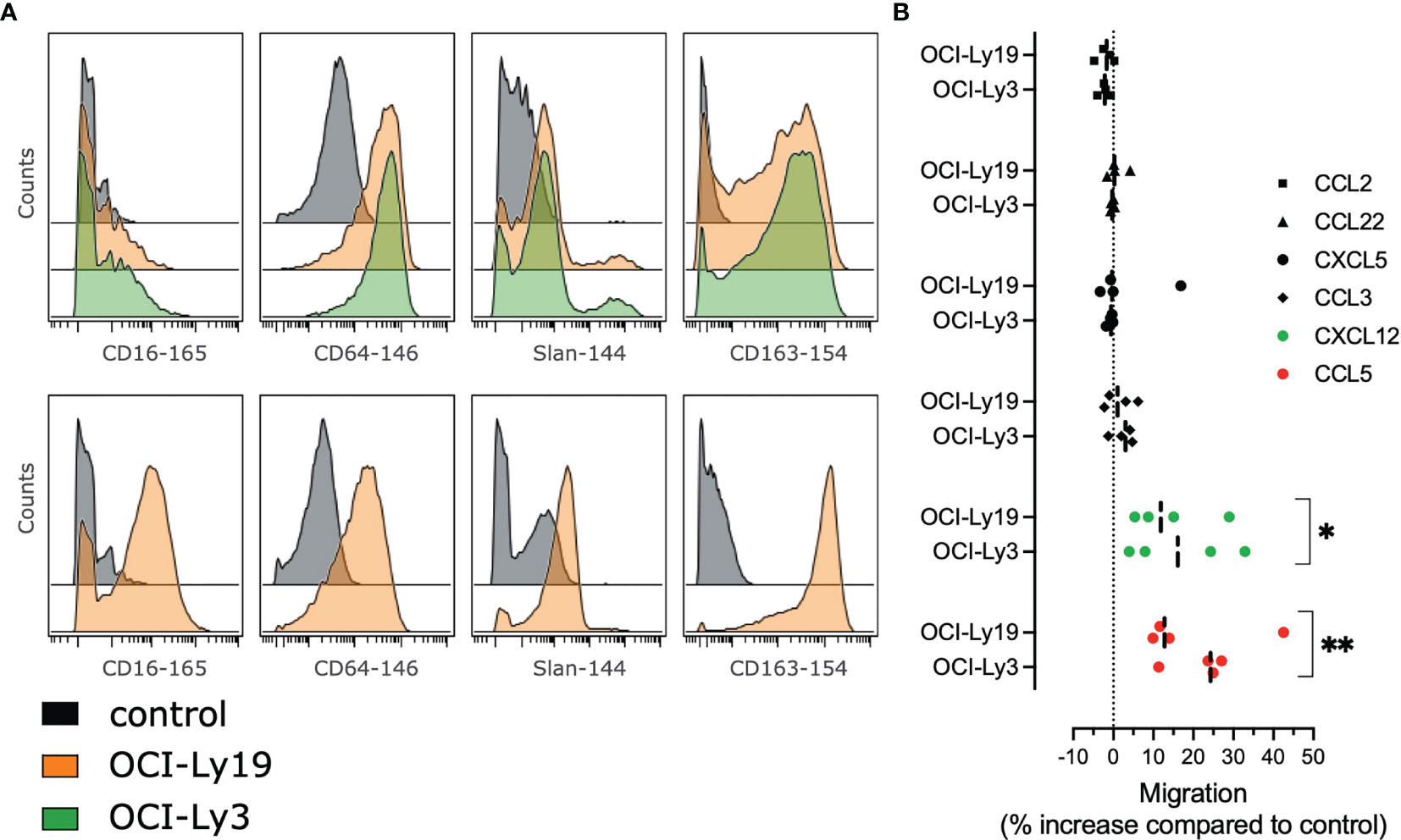
Figure 4 Tumor cells supernatants polarize monocytes with higher migratory abilities. (A) Monocytes from healthy donors (n = 2) were treated with OCI-Ly3, OCI-Ly19 supernatant, or vehicle as control. Overlay histograms represent mean marker intensities for selected markers. Monocytes treated with vehicle (grey), OCI-Ly19 supernatant (orange), or OCI-Ly19 supernatant (green). Top: experiment 1. Bottom: experiment 2. (B) Migration assay for HD monocytes (n = 4) cultured or not with OCI-Ly3 and OCI-Ly19 supernatant in response to CCL2, CCL3, CCL5, CCL22, CXCL5, and CXCL12. Wilcoxon matched-pairs signed rank test were used to compare tumor-conditioned monocytes migration to the control condition (OCI-Ly medium). *P < .05, **P < .01 (22).
High Level of Circulating ncMO Is Correlated With an Adverse Prognosis in DLBCL
Then, we evaluated the prognosis value of cMO, iMO, and ncMO in DLBCL. We used i) the proportion of ncMO to other monocytes (ratio ncMO to sum of cMO and iMO) and ii) the absolute count of circulating cMO, iMO, and ncMO. Analysis was performed on 52 patients for which clinical data were available. cMO and iMO were not associated with prognosis (data not shown). By contrast, patients with high proportion of ncMO and high absolute count of circulating ncMO were associated with a lower event-free survival probability (P = .043 and P = .0061, respectively) using thresholds (ratio at 0.06 and ncMO at 20.58 x106 cells/L) defined with the maxstat package (Figure 5A and Figure S7). To validate the prognosis value of ncMO obtained on this training cohort, we analyzed by flow cytometry the proportion of monocyte subsets in an independent cohort of 155 DLBCL samples from the recently published GAINED trial (NCT01659099) (24). With the previously calculated thresholds, high proportion of ncMO and high absolute count of ncMO was associated with a lower overall survival (P = .017 and P = .011, respectively) (Figure 5A). A univariate analysis on the validation cohort showed that Ann Arbor Stage III-IV, ECOG status >1, elevated LDH, PET4 positivity, and increase in circulating ncMO were associated with lower OS (Table 2). In a multivariable analysis Ann Arbor Stage III-IV, PET4 positivity, increase in circulating ncMO remained statistically significant (Table 2). We previously demonstrated the accumulation of M-MDSC in DLBCL (22) and since no phenotypic overlap existed between M-MDSC and ncMO (Figure 1D), we wondered if patients’ characteristics were different between M-MDSChigh and ncMOhigh DLBCLs. Both M-MDSC and ncMO were infrequently increased together [11 cases out of 155 (7.1%)]; ncMO were increased alone in 51 cases (32.9%), and M-MDSC were increased alone in 28 cases (18.1%) (Figure 5B). Interestingly, ncMOhigh and M-MDSChigh patients corresponded to different types of patients. In particular when compared to ncMOlow, ncMOhigh were enriched in ABC DLBCL subtypes [37.5 vs 15.9% (P = .014)] and in older patients [median age at 50 vs 46 years (P = .044)]. On the other hand, when compared to ncMOhigh, M-MDSChigh patients were younger [median age at 42 vs 50 years (P = .0027)] and had higher levels of soluble PD-L1 [sPD-L1 at 1849 vs 1142 pg/mL (P = .008)] (Figure 5B).
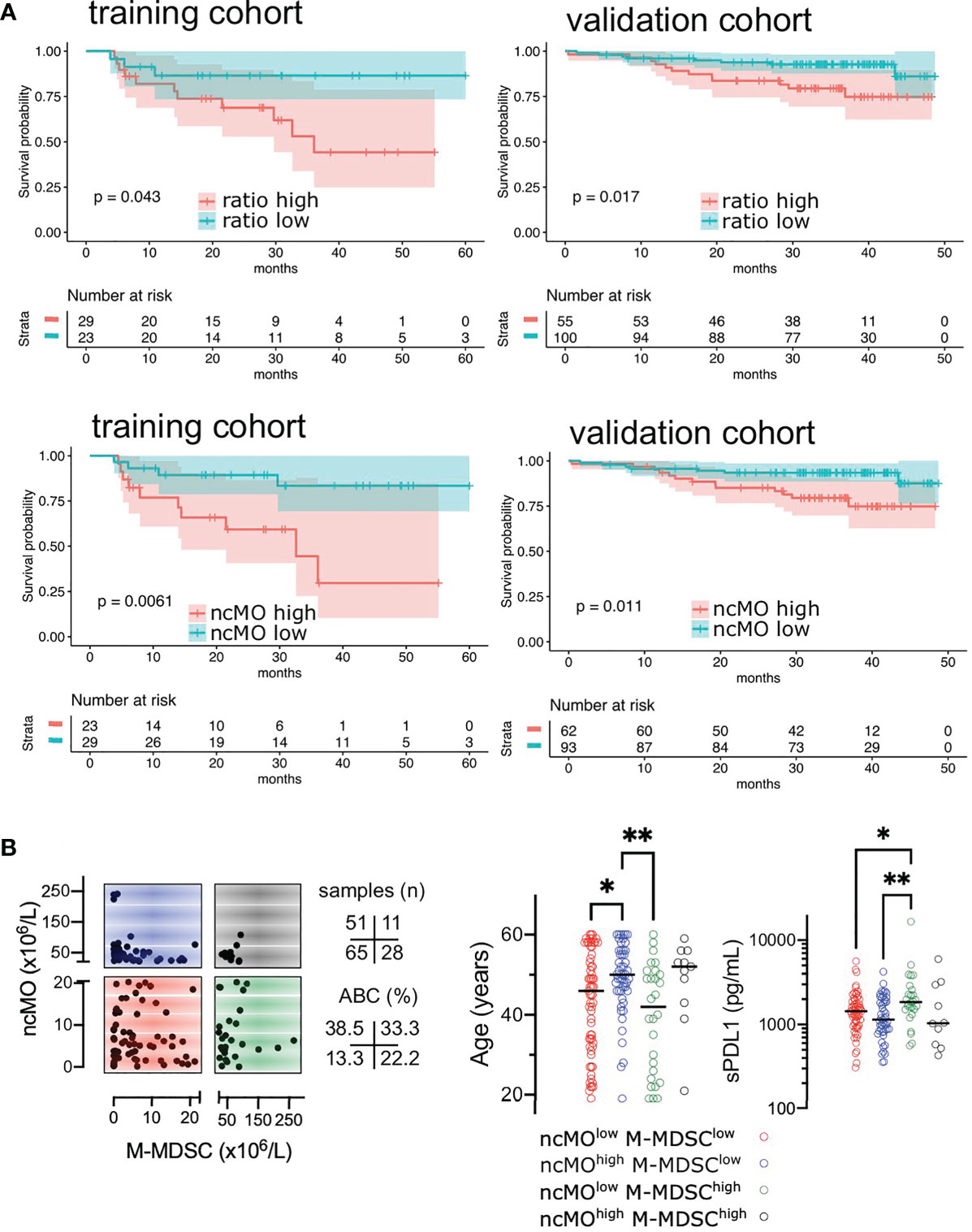
Figure 5 High levels of circulating ncMO is correlated to adverse prognosis in DLBCL. (A) Event-free survival (EFS) in training cohorts (NCT01287923) and overall survival (OS) in validation cohorts (NCT01659099) (24). Patients were stratified on the ratio of ncMO to other monocytes (cMO and iMO) and on the absolute count of circulating ncMO. Threshold was defined on the training cohort using the maxstat package (Figure S7). Survival probability was calculated for both groups with a log-rank test. (B) Absolute count for ncMO (threshold at 20.58 x106 cells/L) and M-MDSC (threshold at 22.51 x106 cells/L) (22), and distribution of age and soluble PD-L1 (sPD-L1), *P < .05, **P < .01.
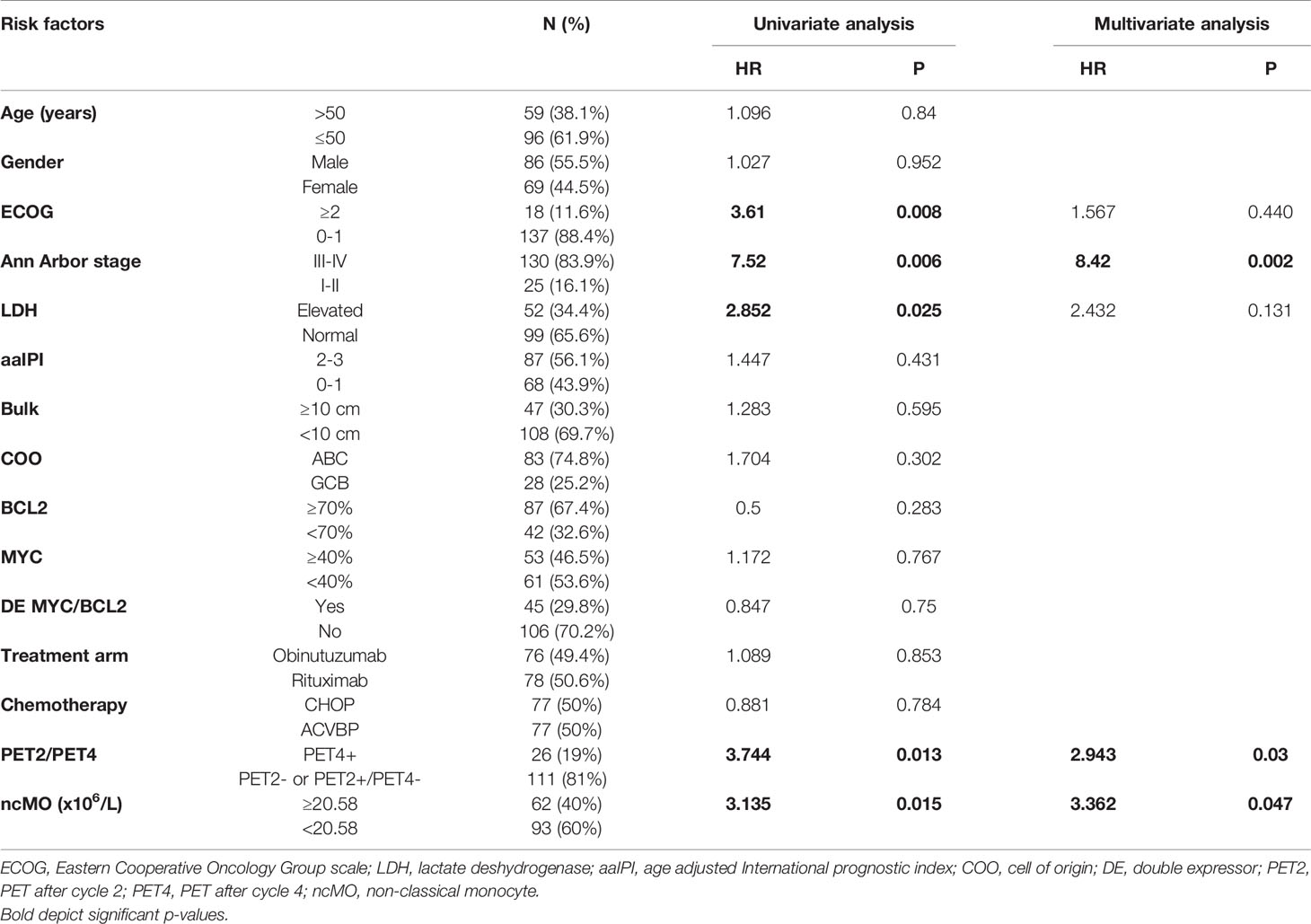
Table 2 Factors influencing overall survival in validation cohort.
Discussion
Although the prognostic relevance of total monocyte count has been described in large cohorts of DLBCL in the last decade (11–15), few studies evaluated which particular monocyte subset was involved. In a previous work, we have shown an accumulation of M-MDSC (CD14pos HLA-DRlow) in peripheral blood from DLBCL patients (22). Because M-MDSCs were not responsible for the whole increase in monocytes in our cohort, we explored cMO, iMO, and ncMO subsets. We demonstrated an increase in cMOs and iMOs in DLBCL, as in other lymphomas subtypes tested (CLL, MCL, MZL, and FL). In DLBCL, these MO subsets shared an inflammatory phenotype. By contrast, ncMOs were decreased in peripheral blood only in DLBCLs when compared to HDs or other B cell lymphomas. Interestingly, high number of circulating ncMO was an adverse prognosis in 2 independent cohorts of DLBCL patients. Finally, we found that tumor-conditioned monocytes shared a common phenotype with ncMOs and were prone to migrate in response to chemokines.
Surprisingly cMO and iMO from DLBCL showed common deregulated pathways with an enrichment for FCGR3A, CD36, FCGR1A, CYBB, AIM2, STAT6, FCGR2A, CCR2, NLRC4, S100A8, and CD14. These genes are broadly expressed in cMO in healthy samples (2, 5) and our results suggest that iMO and cMO are tumor-educated and polarized to a common inflammatory phenotype in DLBCL. In our study we found a decrease in both circulating ncMO Slanneg and ncMO Slanpos when compared to HDs, whereas an increase in ncMO Slanpos was previously described in DLBCL (16). This discrepancy might be explained by differences in patient characteristics between both studies. In particular patients were older in the study from Verni et coll [63.9 years (range: 31-86) vs 50 years (range:18-83)] and at higher grade (clinical stage III-IV at 80.5% vs 70% and IPI ≥3 at 55.6% vs 40%) (16). In CLL, an increase of ncMO correlates with high cytogenetic risk (deletion 11q, 17p, or trisomy 12) (28). In our study, an increase of the proportion of circulating ncMO was a worse prognosis factor in 2 independent cohorts. This was previously suggested on 45 DLBCLs where the decrease of CD16neg monocyte to CD16pos monocyte ratio predicted poor prognosis, however conclusions were limited because iMO and cMO were analyzed conjointly (17). ncMO abundance also predicted patient survival of pediatric and adult B acute lymphoblastic leukemia (29). Interestingly, in a pre-clinical mouse model of B cell lymphomas, Ly6Clow monocytes (corresponding to the ncMO) (30) accumulated and showed high levels of immunosuppressive genes (PD-L1, PD-L2, Arg1, IDO1, and CD163) associated with suppression of T cell proliferation (31). In colorectal cancer Ly6Clow monocyte mediated immunosuppression by IL-10 production (32). Finally, ncMO were increased in gastric cancer (33). Conversely, in a lung cancer model, LyClow monocytes recruited NK cells to prevent cancer metastasis (34). In DLBCL, we and others focused on total monocyte and on M-MDSC and few attentions were given to other monocyte subsets. Interestingly, ncMO and M-MDSC have non-overlapping phenotype regarding HLA-DR expression and these cells infrequently correlated in patients suggesting different mechanism of myelopoiesis dysregulation. Patients that were enriched in circulating MDSCs were younger and presented high amount of sPD-L1, a pejorative marker (35). Interestingly, release of PD-L1 was a mechanism of immune suppression suggested in DLBCL (22). Nonclassical monocytes have been associated age in healthy patients (36), however, no correlation was found in DLBCL samples between age and ncMO (data not shown).
Beside immunosuppression, gene enriched in ncMO were related to chemotaxis. Circulating ncMO are diminished in DLBCL, on the contrary there were enriched in other B cell lymphomas or solid tumor (37), thus we hypothesized that these cells might migrate into tissue to contribute to the tumor-associated macrophage compartment. CCL2, CCL3, CCL5, CCL22, CXCL5, and CXCL12 are involved in monocyte, MDSC, and macrophage recruitment into the TME (38). Tumor-conditioned monocyte shown an increased migration in response to CXCL5, CXCL12, CCL3, and CCL5. In our previous study, CXCL5 expression was increased in peripheral blood from DLBCL compared to healthy donors and its expression was related to a worse event-free survival (22). CCL3 is also increased in DLBCL when compared to HD and high level correlates with shorter survival (39, 40). CCL5 is involved in macrophage recruitment in DLBCL (41)
In DLBCL, TAM are heterogenous (23), in particular a Slanpos macrophage subset is involved in rituximab mediated antibody dependent cellular cytotoxicity (16). In agreement, we found in DLBCL a compartment of cells expressing Slan at high level with CD14, CD32, and HLA-DR. However, DLBCL clusters that correlated with tumor-conditioned monocytes highly expressed CD64, CD36, and S100A9 and thus presented similarities with IFNγ in-vitro polarized macrophages (25). Few studies compared paired samples from circulating and in situ myeloid cells. In melanoma patient, myeloid cells obtained from the blood, but not from the tumor, were suppressive (42). In lung adenocarcinoma, macrophages phenotype detected in tumor were not present in peripheral blood (43). Currently, there is no model of lymphoma that allows tracking the myeloid cell from the blood to the tissues. Future studies entailing a prospective collection of paired blood and tumor samples are needed to confirm these observations on ncMO and to put in perspective the myeloid compartment with the T/NK compartment. Also, it would be interested to test the prognosis value in cohort of DLBCL treated with other immunotherapies and correlate with responders vs non-responders.
Our study as some limitations, in particular the lack of extensive functional studies due to the low number of circulating ncMO in DLBCL samples precluding large cell sorting. Data on gene expression in the monocyte subsets need confirmation with cell preparations, for which a high purity is documented. Even if we previously identified CD16 macrophages subsets in DLBCL in dissociated tissue (23), additional studies with high dimensional spatial positioning of macrophage subsets will enrich our study. Finally, functional assays would be required to definitively address the suppressive activity of DLBCL ncMO for instance using mice models allowing to track the cMO to ncMO transition and the migration to tissue during the lymphoma course will also be valuable. Taken together, our results show that ncMO are involved in the DLBCL physiopathology and impact the prognosis of the disease. Given the current and our previous data, we propose that cMO and iMO are reflecting the inflammatory status in DLBCL, whereas M-MDSC are responsible of a systemic suppressive response, and ncMO are involved in suppressive response and migration to tissue.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: http://flowrepository.org/id/FR-FCM-Z4N6.
Ethics Statement
The studies involving human participants were reviewed and approved by CHU Rennes. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
SGa, FL, AM, CM, and IA designed and performed experiments, analyzed data. JI, DR, JF, and TM analyzed data. CP, KB, GD, GC, PG, SGo, R-OC, RH, and TL provided samples. TF and KT raised the funds and analyzed data. MR designed and supervised research, analyzed data, and wrote the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by a fellowship from the Nuovo-Soldati Foundation (Switzerland) (MR), from the Ligue contre le Cancer (MR), from the COmite de la REcherche Clinique et Translationnelle, CHU of Rennes (FL), from the Association pour le Développement de l’Hématologie Oncologie (FL), from the National Institute of Cancer (INCa Recherche Translationnelle 2010) (TF), from the Groupe Ouest-Est des Leucémies et des Autres Maladies du Sang (GOELAMS) (TF), from the Agence Nationale de la Recherche (ANR-17-CE15-0015 StroMAC) (KT), and from the Fondation ARC (PGA1 RF20190208534) (KT and MR).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are indebted to the clinicians of the BREHAT network and to the French Blood Bank (EFS) of Rennes for providing samples. The authors acknowledge the Centre de Ressources Biologiques (CRB-santé) of Rennes (BB-0033-00056) for managing samples and the Flow Cytometry facility, Biosit, University of Rennes 1 for cell sorting. This article was submitted as preprint (https://www.biorxiv.org/content/10.1101/2021.04.10.439292v1) (44).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.755623/full#supplementary-material
Figure S1 | Gating strategy.
Figure S2 | All monocyte subsets are increased in B-cell lymphomas A- Monocyte subsets counts in peripheral blood from HD (n = 6), follicular lymphomas (n = 9), mantle cell lymphomas (MCL, n = 9), chronic lymphocytic leukemias (CLL, n = 11), and marginal zone lymphomas (SMZL, n = 10). On these patients, flow cytometry was performed on cryopreserved cells. B- Ratio between MO (classical and intermediate) and ncMO in B-cell lymphomas.
Figure S3 | Q-PCR analysis, unsupervised hierarchical clustering for cMO and iMO from DLBCL and HD patients cMO and iMO were sorted from 7 DLBCL and 4 HD before analysis by high-throughput Q-PCR for genes listed in Table S3. Pearson's correlation and complete linkage were employed.
Figure S4 | Q-PCR analysis, unsupervised hierarchical clustering for ncMO Slanpos and ncMO Slanneg from DLBCL and HD patients ncMO Slanpos and ncMO Slanneg were sorted from 7 DLBCL and 4 HD before analysis by high-throughput Q-PCR for genes listed in Table S3. Pearson's correlation and complete linkage were employed.
Figure S5 | Q-PCR analysis, unsupervised hierarchical clustering for cMO, iMO, ncMO Slanpos and ncMO Slanneg from DLBCL patients cMO, iMO, ncMO Slanpos, and ncMO Slanneg were sorted from 7 DLBCL before analysis by high-throughput Q-PCR for genes listed in Table S3. Pearson's correlation and complete linkage were employed.
Figure S6 | Q-PCR analysis, unsupervised hierarchical clustering for cMO, iMO, ncMO Slanpos and ncMO Slanneg from DLBCL and HD patients cMO, iMO, ncMO Slanpos, and ncMO Slanneg were sorted from 7 DLBCL and 4 HD before analysis by high-throughput Q-PCR for genes listed in Table S3. Pearson's correlation and complete linkage were employed.
Figure S7 | High levels of circulating ncMO is an adverse event in DLBCL.Event-free survival (EFS) in training cohorts (NCT01287923) Threshold of the ratio of ncMO to other monocytes parameter was defined on the training cohort using the maxstat package.
Table S1 | Antibodies for fluorescent flow cytometry analysis.
Table S2 | Antibodies or parameters used for mass cytometry analysis.
Table S3 | List of genes evaluated by high-throughput Q-PCR on monocyte subsets.
Table S4 | QPCR data.
References
1. Ziegler-Heitbrock L, Ancuta P, Crowe S, Dalod M, Grau V, Hart DN, et al. Nomenclature of Monocytes and Dendritic Cells in Blood. Blood (2010) 116:e74–80. doi: 10.1182/blood-2010-02-258558
2. Cros J, Cagnard N, Woollard K, Patey N, Zhang S-Y, Senechal B, et al. Human CD14dim Monocytes Patrol and Sense Nucleic Acids and Viruses via TLR7 and TLR8 Receptors. Immunity (2010) 33:375–86. doi: 10.1016/j.immuni.2010.08.012
3. Ahmad F, Döbel T, Schmitz M, Schäkel K. Current Concepts on 6-Sulfo LacNAc Expressing Monocytes (slanMo). Front Immunol (2019) 10:948. doi: 10.3389/fimmu.2019.00948
4. Bronte V, Brandau S, Chen S, Colombo MP, Frey AB, Greten TF, et al. Recommendations for Myeloid-Derived Suppressor Cell Nomenclature and Characterization Standards. Nat Commun (2016) 7:12150. doi: 10.1038/ncomms12150
5. Wong KL, Tai JJ-Y, Wong W-C, Han H, Sem X, Yeap W-H, et al. Gene Expression Profiling Reveals the Defining Features of the Classical, Intermediate, and Nonclassical Human Monocyte Subsets. Blood (2011) 118:e16–31. doi: 10.1182/blood-2010-12-326355
6. Guilliams M, Mildner A, Yona S. Developmental and Functional Heterogeneity of Monocytes. Immunity (2018) 49:595–613. doi: 10.1016/j.immuni.2018.10.005
7. Lenz G, Wright G, Dave SS, Xiao W, Powell J, Zhao H, et al. Stromal Gene Signatures in Large-B-Cell Lymphomas. N Engl J Med (2008) 359:2313–23. doi: 10.1056/nejmoa0802885
8. Riihijarvi S, Fiskvik I, Taskinen M, Vajavaara H, Tikkala M, Yri O, et al. Prognostic Influence of Macrophages in Patients With Diffuse Large B-Cell Lymphoma: A Correlative Study From a Nordic Phase II Trial. Haematologica (2015) 100:238–45. doi: 10.3324/haematol.2014.113472
9. Scott DW, Gascoyne RD. The Tumour Microenvironment in B Cell Lymphomas. Nat Rev Cancer (2014) 14:517–34. doi: 10.1038/nrc3774
10. Nam SJ, Kim S, Kwon D, Kim H, Kim S, Lee E, et al. Prognostic Implications of Tumor-Infiltrating Macrophages, M2 Macrophages, Regulatory T-Cells, and Indoleamine 2,3-Dioxygenase-Positive Cells in Primary Diffuse Large B-Cell Lymphoma of the Central Nervous System. Oncoimmunology (2018) 7:1–39. doi: 10.1080/2162402x.2018.1442164
11. Tadmor T, Bari A, Sacchi S, Marcheselli L, Liardo EV, Avivi I, et al. Monocyte Count at Diagnosis Is a Prognostic Parameter in Diffuse Large B-Cell Lymphoma: Results From a Large Multicenter Study Involving 1191 Patients in the Pre- and Post-Rituximab Era. Haematologica (2014) 99:125–30. doi: 10.3324/haematol.2013.088161
12. Nitta H, Terui Y, Yokoyama M, Mishima Y, Nishimura N, Ueda K, et al. Absolute Peripheral Monocyte Count at Diagnosis Predicts Central Nervous System Relapse in Diffuse Large B-Cell Lymphoma. Haematologica (2015) 100:87–90. doi: 10.3324/haematol.2014.114934
13. Troppan K, Deutsch A, Gerger A, Stojakovic T, Beham-Schmid C, Wenzl K, et al. The Derived Neutrophil to Lymphocyte Ratio Is an Independent Prognostic Factor in Patients With Diffuse Large B-Cell Lymphoma. Br J Cancer (2014) 110:369–74. doi: 10.1038/bjc.2013.763
14. Wilcox RA, Ristow K, Habermann TM, Inwards DJ, Micallef INM, Johnston PB, et al. The Absolute Monocyte and Lymphocyte Prognostic Score Predicts Survival and Identifies High-Risk Patients in Diffuse Large-B-Cell Lymphoma. Leukemia (2011) 25:1502–9. doi: 10.1038/leu.2011.112
15. Maurer MJ, Jais J-P, Ghesquières H, Witzig TE, Hong F, Haioun C, et al. Personalized Risk Prediction for Event-Free Survival at 24 Months in Patients With Diffuse Large B-Cell Lymphoma. Am J Hematol (2016) 91:179–84. doi: 10.1002/ajh.24223
16. Vermi W, Micheletti A, Finotti G, Tecchio C, Calzetti F, Costa S, et al. Slan+ Monocytes and Macrophages Mediate CD20-Dependent B-Cell Lymphoma Elimination via ADCC and ADCP. Cancer Res (2018) 78:3544–59. doi: 10.1158/0008-5472.can-17-2344
17. Han X, Ruan J, Zhang W, Zhou D, Xu D, Pei Q, et al. Prognostic Implication of Leucocyte Subpopulations in Diffuse Large B-Cell Lymphoma. Oncotarget (2017) 8:47790–800. doi: 10.18632/oncotarget.17830
18. Yhim H-Y, Kim J-A, Ko S-H, Park Y, Yim E, Kim HS, et al. The Prognostic Significance of CD11b+CX3CR1+ Monocytes in Patients With Newly Diagnosed Diffuse Large B-Cell Lymphoma. Oncotarget (2017) 8:92289–99. doi: 10.18632/oncotarget.21241
19. Vari F, Arpon D, Keane C, Hertzberg MS, Talaulikar D, Jain S, et al. Immune Evasion via PD-1/PD-L1 on NK Cells and Monocyte/Macrophages Is More Prominent in Hodgkin Lymphoma Than DLBCL. Blood (2018) 131:1809–19. doi: 10.1182/blood-2017-07-796342
20. Lin Y, Gustafson MP, Bulur PA, Gastineau DA, Witzig TE, Dietz AB. Immunosuppressive CD14+HLA-DRlow/- Monocytes in B-Cell Non-Hodgkin Lymphoma. Blood (2011) 117:872–81. doi: 10.1182/blood-2010-05-283820
21. Xiu B, Lin Y, Grote DM, Ziesmer SC, Gustafson MP, Maas ML, et al. IL-10 Induces the Development of Immunosuppressive CD14(+)HLA-DR(low/-) Monocytes in B-Cell Non-Hodgkin Lymphoma. Blood Cancer J (2015) 5:e328. doi: 10.1038/bcj.2015.56
22. Azzaoui I, Uhel F, Rossille D, Pangault C, Dulong J, Priol JL, et al. T-Cell Defect in Diffuse Large B-Cell Lymphomas Involves Expansion of Myeloid-Derived Suppressor Cells. Blood (2016) 128:1081–92. doi: 10.1182/blood-2015-08-662783
23. Roussel M, Lhomme F, Roe CE, Bartkowiak T, Gravelle P, Laurent C, et al. Mass Cytometry Defines Distinct Immune Profile in Germinal Center B-Cell Lymphomas. Cancer Immunol Immunother (2020) 69:407–20. doi: 10.1007/s00262-019-02464-z
24. Gouill SL, Ghesquières H, Oberic L, Morschhauser F, Tilly H, Ribrag V, et al. Obinutuzumab vs Rituximab for Advanced DLBCL: A PET-Guided and Randomized Phase 3 Study by LYSA. Blood (2021) 137:2307–20. doi: 10.1182/blood.2020008750
25. Roussel M, Ferrell PB, Greenplate AR, Lhomme F, Gallou SL, Diggins KE, et al. Mass Cytometry Deep Phenotyping of Human Mononuclear Phagocytes and Myeloid-Derived Suppressor Cells From Human Blood and Bone Marrow. J Leukocyte Biol (2017) 102:437–47. doi: 10.1189/jlb.5ma1116-457r
26. Roussel M, Bartkowiak T, Irish JM. Picturing Polarized Myeloid Phagocytes and Regulatory Cells by Mass Cytometry. Methods Mol Biol (2019) 1989:217–26. doi: 10.1007/978-1-4939-9454-0_14
27. Ferrant J, Lhomme F, Gallou SL, Irish JM, Roussel M. Circulating Myeloid Regulatory Cells: Promising Biomarkers in B-Cell Lymphomas. Front Immunol (2021) 11:623993. doi: 10.3389/fimmu.2020.623993
28. Maffei R, Bulgarelli J, Fiorcari S, Bertoncelli L, Martinelli S, Guarnotta C, et al. The Monocytic Population in Chronic Lymphocytic Leukemia Shows Altered Composition and Deregulation of Genes Involved in Phagocytosis and Inflammation. Haematologica (2013) 98:1115–23. doi: 10.3324/haematol.2012.073080
29. Witkowski MT, Dolgalev I, Evensen NA, Ma C, Chambers T, Roberts KG, et al. Extensive Remodeling of the Immune Microenvironment in B Cell Acute Lymphoblastic Leukemia. Cancer Cell (2020) 37:867–82.e12. doi: 10.1016/j.ccell.2020.04.015
30. Ginhoux F, Jung S. Monocytes and Macrophages: Developmental Pathways and Tissue Homeostasis. Nat Rev Immunol (2014) 14:392–404. doi: 10.1038/nri3671
31. McKee SJ, Tuong ZK, Kobayashi T, Doff BL, Soon MS, Nissen M, et al. B Cell Lymphoma Progression Promotes the Accumulation of Circulating Ly6Clo Monocytes With Immunosuppressive Activity. Oncoimmunology (2018) 7:e1393599. doi: 10.1080/2162402x.2017.1393599
32. Jung K, Heishi T, Khan OF, Kowalski PS, Incio J, Rahbari NN, et al. Ly6Clo Monocytes Drive Immunosuppression and Confer Resistance to Anti-VEGFR2 Cancer Therapy. J Clin Invest (2017) 127:3039–51. doi: 10.1172/jci93182
33. Eljaszewicz A, Jankowski M, Gackowska L, Helmin-Basa A, Wiese M, Kubiszewska I, et al. Gastric Cancer Increase the Percentage of Intermediate (CD14++CD16+) and Nonclassical (CD14+CD16+) Monocytes. Clin Immunol (2012) 4:355–61. doi: 10.5114/ceji.2012.32725
34. Hanna RN, Cekic C, Sag D, Tacke R, Thomas GD, Nowyhed H, et al. Patrolling Monocytes Control Tumor Metastasis to the Lung. Science (2015) 350:985–90. doi: 10.1126/science.aac9407
35. Rossille D, Azzaoui I, Feldman AL, Maurer MJ, Labouré G, Parrens M, et al. Soluble Programmed Death-Ligand 1 as a Prognostic Biomarker for Overall Survival in Patients With Diffuse Large B-Cell Lymphoma: A Replication Study and Combined Analysis of 508 Patients. Leukemia (2017) 31:988–91. doi: 10.1038/leu.2016.385
36. Ong SM, Hadadi E, Dang TM, Yeap W-H, Tan CT-Y, Ng T-P, et al. The Pro-Inflammatory Phenotype of the Human Non-Classical Monocyte Subset Is Attributed to Senescence. Cell Death Dis (2018) 9(3):266. doi: 10.1038/s41419-018-0327-1
37. Cassetta L, Fragkogianni S, Sims AH, Swierczak A, Forrester LM, Zhang H, et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell (2019) 35:588–602.e10. doi: 10.1016/j.ccell.2019.02.009
38. Nagarsheth N, Wicha MS, Zou W. Chemokines in the Cancer Microenvironment and Their Relevance in Cancer Immunotherapy. Nat Rev Immunol (2017) 17:559–72. doi: 10.1038/nri.2017.49
39. Charbonneau B, Maurer MJ, Ansell SM, Slager SL, Fredericksen ZS, Ziesmer SC, et al. Pretreatment Circulating Serum Cytokines Associated With Follicular and Diffuse Large B-Cell Lymphoma: A Clinic-Based Case-Control Study. Cytokine (2012) 60:882–9. doi: 10.1016/j.cyto.2012.08.028
40. Takahashi K, Sivina M, Hoellenriegel J, Oki Y, Hagemeister FB, Fayad L, et al. CCL3 and CCL4 Are Biomarkers for B Cell Receptor Pathway Activation and Prognostic Serum Markers in Diffuse Large B Cell Lymphoma. Brit J Haematol (2015) 171:726–35. doi: 10.1111/bjh.13659
41. Manfroi B, Grandis MD, Moreaux J, Tabruyn SP, Mayol J-F, Quintero M, et al. The Microenvironment of DLBCL Is Characterized by Non-Canonical Macrophages Recruited by Tumor-Derived CCL5. Blood Adv (2021) 5(21):4338–51. doi: 10.1182/bloodadvances.2021004203
42. Gros A, Turcotte S, Wunderlich JR, Ahmadzadeh M, Dudley ME, Rosenberg SA. Myeloid Cells Obtained From the Blood But Not From the Tumor Can Suppress T-Cell Proliferation in Patients With Melanoma. Clin Cancer Res (2012) 18:5212–23. doi: 10.1158/1078-0432.ccr-12-1108
43. Lavin Y, Kobayashi S, Leader A, Amir ED, Elefant N, Bigenwald C, et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired Single-Cell Analyses. Cell (2017) 169:750–7.e15. doi: 10.1016/j.cell.2017.04.014
Keywords: B cell lymphoma, DLBCL, biomarker, monocyte, immune suppression
Citation: Le Gallou S, Lhomme F, Irish JM, Mingam A, Pangault C, Monvoisin C, Ferrant J, Azzaoui I, Rossille D, Bouabdallah K, Damaj G, Cartron G, Godmer P, Le Gouill S, Casasnovas R-O, Molina TJ, Houot R, Lamy T, Tarte K, Fest T and Roussel M (2021) Nonclassical Monocytes Are Prone to Migrate Into Tumor in Diffuse Large B-Cell Lymphoma. Front. Immunol. 12:755623. doi: 10.3389/fimmu.2021.755623
Received: 09 August 2021; Accepted: 24 November 2021;
Published: 16 December 2021.
Edited by:
Sven Brandau, University of Duisburg-Essen, GermanyReviewed by:
Marc Seifert, Essen University Hospital, GermanyGero Knittel, Essen University Hospital, Germany
Loems Ziegler-Heitbrock, Independent Researcher, Munich, Germany
Copyright © 2021 Le Gallou, Lhomme, Irish, Mingam, Pangault, Monvoisin, Ferrant, Azzaoui, Rossille, Bouabdallah, Damaj, Cartron, Godmer, Le Gouill, Casasnovas, Molina, Houot, Lamy, Tarte, Fest and Roussel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mikael Roussel, bWlrYWVsLnJvdXNzZWxAdW5pdi1yZW5uZXMxLmZy
†These authors have contributed equally to this work