 Mariame Mohamed Ahamada
Mariame Mohamed Ahamada Yang Jia
Yang Jia Xiaochuan Wu*
Xiaochuan Wu*
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 20 December 2021
Sec. Autoimmune and Autoinflammatory Disorders
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.734008
Systemic lupus erythematosus (SLE) is an autoimmune disease that attacks almost every organ. The condition mostly happens to adults but is also found in children, and the latter have the most severe manifestations. Among adults, females, especially non-Caucasian, are mostly affected. Even if the etiology of SLE remains unclear, studies show a close relation between this disease and both genetics and environment. Despite the large number of published articles about SLE, we still do not have a clear picture of its pathogenesis, and no specific drug has been found to treat this condition effectively. The implication of macrophages in SLE development is gaining ground, and studying it could answer these gaps. Indeed, both in vivo and in vitro studies increasingly report a strong link between this disease and macrophages. Hence, this review aims to explore the role of macrophages polarization and plasticity in SLE development. Understanding this role is of paramount importance because in-depth knowledge of the connection between macrophages and this systemic disease could clarify its pathogenesis and provide a foundation for macrophage-centered therapeutic approaches.
Systemic lupus erythematosus belongs to a group of autoimmune diseases commonly called lupus. Lupus means “wolf” in Latin and describes a facial rash resembling a wolf bite (1). Initially, the disease was simply called lupus because it was thought only to involve the skin. Later, when other systemic manifestations were reported, the name Systemic lupus erythematosus (SLE) was introduced (2). So far, there is no known direct cause of this disease. However, its higher distribution in adults, female gender, and certain ethnicities such as non-Caucasian, makes genetics and environment two crucial determinants of disease development (3, 4). SLE affects almost every organ and presents with a high range of clinical manifestations (5). The damage to various organs can be explained by a disturbance of innate and adaptive immune responses leading to a production of autoantibodies, immune complexes, and a loss of immune tolerance to autoantigens (6, 7). It is well known that innate and adaptive immunity work together to defend an individual against pathogens. However, in some disorders such as SLE, this function is deregulated. Consequently, the immune system attacks the individual’s self-organs instead of protecting them. Among the immune components, macrophages are believed to play a significant role in the pathogenesis of SLE. Indeed, it has been established that the polarization of macrophages into M1-/M2-like macrophages affects the development of lupus (8). A significant influence on the disease process is also exerted by macrophage plasticity. This unique property of macrophages could enable the pursuit of these immune cells for therapeutic goals by inducing a phenotype switch between M1-/M2-like macrophages (9). Given the crucial role macrophages play in the development of SLE, studies on this autoimmune disease are increasingly focusing on its relationship with these immune cells. Therefore, the purpose of this review is to provide an up-to-date summary of the connection between Systemic lupus erythematosus and macrophages.
One of the indispensable arms of the immune system is macrophages. These large multitasking and plastic cells polarize into different phenotypes and ultimately carry different functions depending on the microenvironment. Here, polarization can be understood as the spatiotemporal activation of macrophages at a given point (10, 11). In other words, polarization is the induction of functionally distinct macrophages with regard to the dominant factors in the inflammatory zone. Based on their polarization and function, macrophages were traditionally categorized into two main phenotypes. However, this long-held view is obsolete and is nowadays considered to be an oversimplified approach. Indeed, accumulating evidence affirms the existence of other macrophage populations in vivo (12, 13) and reveals different behaviors between in vitro and in vivo macrophages. More interestingly, it is suggested that macrophages express different genes in vitro and in vivo (14). Therefore, it is evident that the nomenclature of macrophages is not as simple as it was thought to be, and it would be erroneous to identify them as M1/M2 macrophages.
While macrophage activation and polarization allow macrophages cells to acquire a specific phenotype, macrophage plasticity, on the other hand, enables these immune cells to switch from one phenotype to another (15). In other words, these plastic cells have the unique ability to re-polarize in response to environmental factors and adopt a new phenotype. In diseases like SLE, where deregulation of macrophage phenotypes is known to play a pathogenic role, this functional adaptability has a tremendous therapeutic value because it could be exploited to restore the balance between different macrophages subtypes. Thus, we must review the diversity of macrophage phenotypes in SLE development and the impact of the environment on the behavior of these immune cells.
Macrophages play an essential role in inflammatory reactions. Based on how they have been polarized, they can either exert a pro-inflammatory effect or an anti-inflammatory effect (16). Such a feature, among others, makes macrophages a potential participant in the development of inflammatory and autoimmune diseases (17). Hence, in the following sections, we explore the link between macrophage subsets and SLE development. This connection is also summarized in Figure 1.
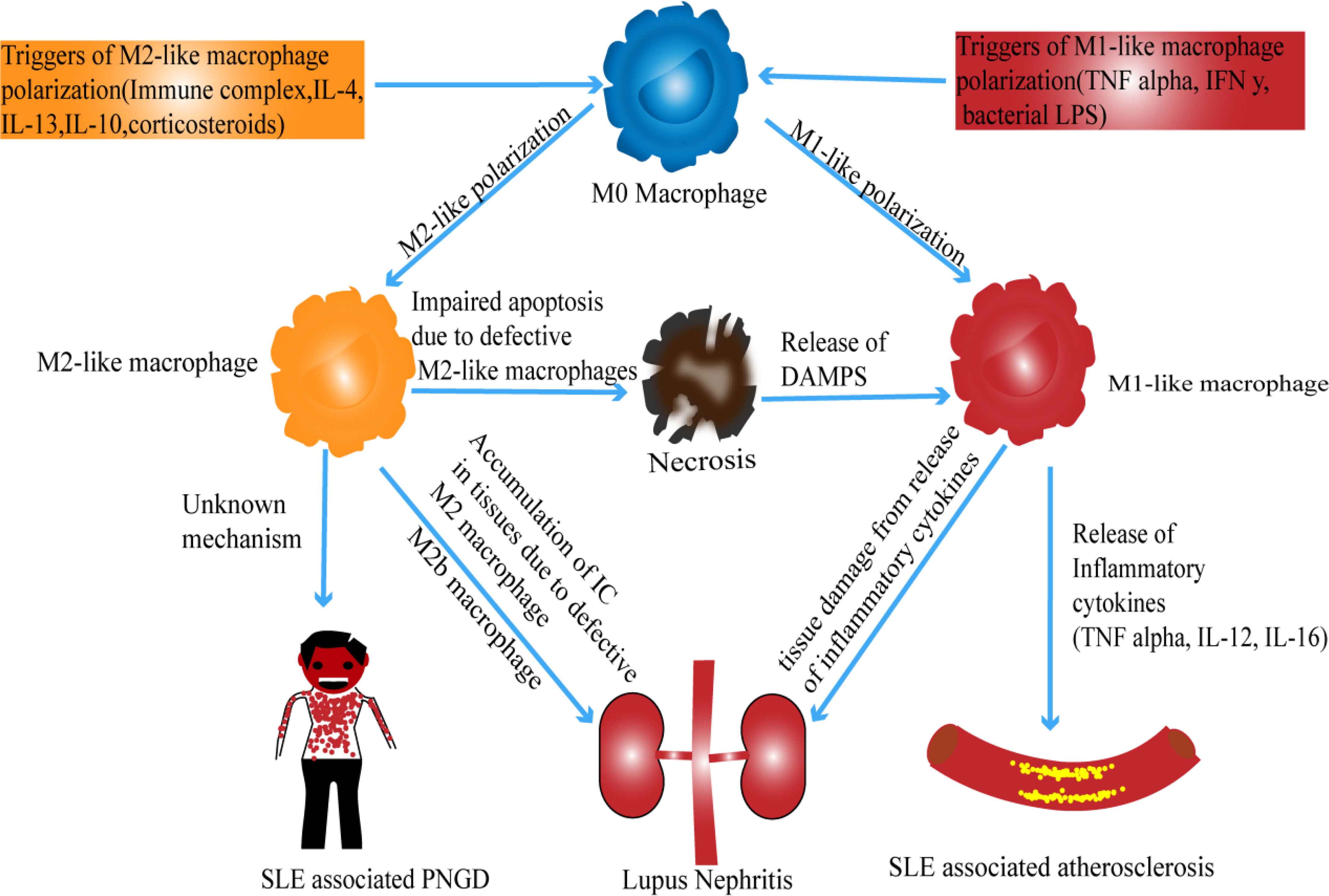
Figure 1 Possible mechanism of macrophage polarization in SLE.
Previously, macrophages were assumed to have an M1 phenotype when their polarization was mediated by T helper one cytokines such as INF-γ. However, this hypothesis is now deemed erroneous because in vitro polarization can be achieved with bacterial lipopolysaccharide (LPS) without the intervention of T lymphocytes (9, 18). Functionally, these types of macrophages are known to have pro-inflammatory activity mainly. Their ability to produce inflammatory cytokines makes them, on the one hand, an essential participant in the elimination of pathogens; however, on the other hand, they interfere with wound healing and tissue repair (9, 19). More interestingly, M1-like macrophage-derived cytokines are believed to mediate autoimmune and chronic inflammatory diseases (20).
The role of M1-like inflammatory macrophages in SLE development is reported in many articles (17, 21, 22). Recently, clinical research has further demonstrated a positive correlation between the number of monocytes expressing M1 macrophage-like markers (CD163-CD14+) in peripheral blood of children with lupus and the severity of childhood-onset SLE (23). Even though CD14 and CD163 are not specific for macrophages, CD163-CD14+ cells are considered M1-like cells (24), and therefore, there is a possibility that the disease activity observed in this study is associated with M1-like macrophages. The connection between M1-like macrophages and SLE can also be appreciated in Lupus nephritis (LN). LN, one of SLE complications, is believed to be characterized by a deregulation of both M1-and M2-like macrophages. Evidence suggesting the involvement of the M1 phenotype showed that LN could be mediated in vivo by type I interferon signature, to which M1 macrophages are very responsive (25). This implication of M1 macrophages is additionally supported by their involvement in the development of atherosclerosis. Atherosclerosis poses a significant threat to global health, and its incidence is high in young patients with SLE. In these subjects, various immune cells, specifically macrophages, are thought to be involved in developing this vascular disease. Whereas the exact role of macrophages in the progression of SLE-associated atherosclerosis is not extensively investigated, the increased serum neopterin concentration in patients with SLE-atherosclerosis indicates a possible association of macrophages with atherogenic mediators of inflammation (26, 27). Additionally, it has been reported that M1-like macrophage-derived cytokines such as TNFα, IFN γ, IL-6, and IL-12 are pro-atherosclerotic and found in patients with SLE-induced atherosclerosis (28). Also, oxidative stress in the plaque seems to be worsened by nitrogen species and reactive oxygen. These molecules are generated from M1-like macrophages and could be seen in patients with SLE-associated atherosclerosis (28). Such information could allow researchers to find therapeutic targets that may improve atherosclerosis in SLE patients.
Likewise, factors that can promote the polarization of M1 macrophages can exacerbate inflammatory conditions such as lupus. Among these factors are Microparticles (MP), remarkably immune complex-forming microparticles (MP-IC). In an in vitro study, these extracellular vesicles were seen to favor the polarization of M1-like inflammatory macrophages and thus amplify inflammation and autoimmunity in diseases such as SLE (29). High-mobility-group box chromosomal protein (HMGB1) might be another facilitator of M1-like inflammatory macrophages (8, 30, 31). In vitro and in vivo studies showed that These proteins can induce SLE by promoting macrophage inflammatory response (32). In conformity with this theory, in vitro study found that the serum level of HMGB1 increased with the severity of neuropsychiatric SLE (NPSLE) (33). However, because evidence showed that inflammation could be mediated by other types of macrophage other than M1-like macrophages (14), we cannot confidently conclude that these inflammatory macrophages are indeed M1-like macrophages. Further research is needed to clarify this point.
These findings address the association of macrophages with SLE and point out the implication of M1-like macrophages in the pathogenesis of the disease. Nevertheless, it should be noted that non-negligible differences can be appreciated within macrophages if observed in different environments (in vivo/in vitro). For example, differences in biomarkers were found between in vivo and in vitro M1-like macrophages (14). This means in vitro results can not necessarily be translated into in vivo and vice versa. Such a concept needs to be taken into consideration when interpreting results from studies conducted in different environments. Immunological biomarkers that can help identify in vivo and in vitro M1-like macrophages are summarized in Table 1.
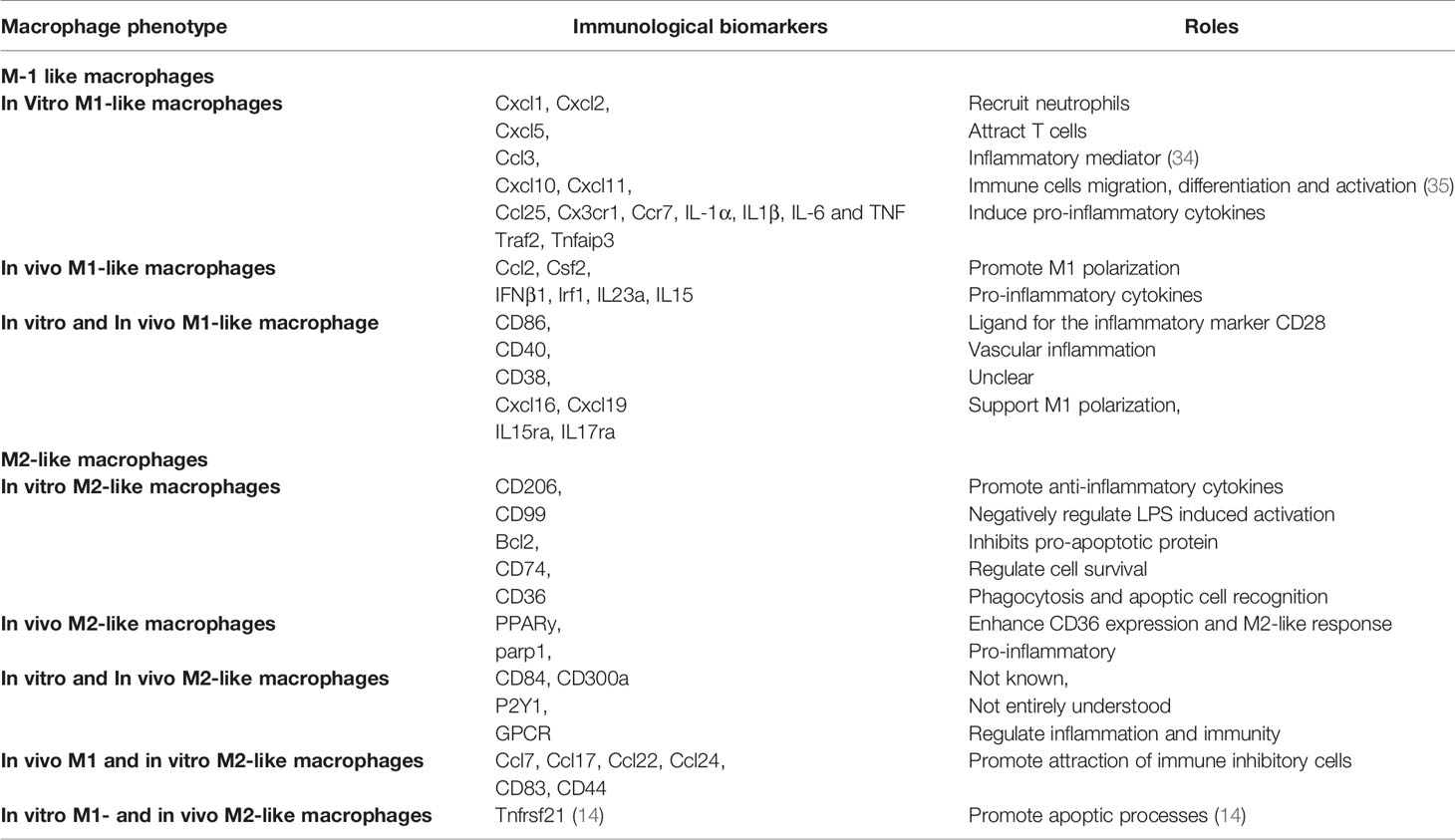
Table 1 In Vivo/in Vitro macrophage markers and their corresponding roles.
M2-like macrophages mainly have anti-inflammatory properties (17). However, recent studies showed that these cells might also exert a pro-inflammatory function. Indeed, poly (ADP-ribose)-polymerase1 (parp1), which represents the majority of poly (ADP-ribose)-polymerase (PARPS), do have pro-inflammatory functions even though they are present in M2-like macrophages in vivo (14, 36). This statement highlights the significant influence that the environment (in vitro/in vivo) has on the behavior of macrophages and why much attention needs to be paid when defining macrophage polarization. It is well accepted that the polarization of M2-like macrophages is induced by Th2 cytokines IL-4 and IL-13. Besides IL-4 and IL-13, other cytokines such as IL-10, IL-33, and IL-21 can also drive M2 polarization (9). Depending on the stimulating factor, M2-like macrophages can further be classified into different subtypes with distinct functions. M2a subtypes are triggered by IL-4 and IL-13 and participate in tissue repair (37). M2b macrophages are stimulated by fcyR/TLR and immune complexes and contribute to tumors and infections. As for M2c macrophages, they respond to glucocorticoid, TGF-β, and IL-10 and exert anti-inflammatory effects (38). M2d macrophages, on the other hand, represent a new subset and take part in angiogenesis and cancer metastasis (39–41). However, in vivo translation of these M2 subtypes is not that simple because some macrophages are found to express mixed phenotypes (42), and others do not meet the M1/M2 model (43). For this reason, the M2 classification is believed to be more about the macrophages’ activation stimuli rather than the resulting functions of these macrophages (39). Table 1 summarizes in vivo/in vitro M2-like macrophages biomarkers.
M2-like macrophages have been reported to play a vital role in the development of SLE. Although both M1-and M2-like macrophages contribute to the pathogenesis of lupus nephritis, several studies suggest that the M2 phenotype is the dominant subpopulation (21, 44, 45). It was reported that defective M2-like macrophages could uncontrollably produce cytokines that contribute to the development of SLE (46, 47). Similarly, the inability to clear immune complexes (ICs) by defective M2-like macrophages results in organ damage by allowing ICs to accumulate in different tissues (48). Mechanically, apoptotic cells are eliminated by M2-like macrophages in a normally non-inflammatory response named efferocytosis (49). Such response gives rise to an increase of anti-inflammatory cytokines and a reduction of pro-inflammatory cytokines (50). Therefore, the inflammation in LN could be caused by nonfunctional M2-like macrophages, which have lost their anti-inflammatory property. This assumption is supported by evidence suggesting that M2-like macrophages found in LN lacked heme oxygenase-1 (HO-1) expression and that supplementation of HO-1 could ameliorate LN (51). In contrast, another research has demonstrated that Granulin (GRN), a protein linked to inflammation, can worsen LN by enhancing M2-like macrophage polarization, specifically M2b polarization (52). Although the M2b macrophages-induced LN mechanism is not clarified in this study (52), it is reported elsewhere that M2b macrophages can produce both pro-and anti-inflammatory cytokines (9, 11). Thus, further study is required to clarify whether these M2b macrophages do indeed mediate LN via the release of inflammatory cytokines or they are just nonfunctional macrophages that have lost their anti-inflammatory activity.
A recent study has further revealed another possible role played by M2-like macrophages in the pathogenesis of palisaded neutrophilic and granulomatous dermatitis (PNGD), a skin condition found in systemic autoimmune diseases like SLE. The study examined two cases of SLE patients with PNGD and revealed CD163+ M2- like macrophages to be the primary phenotype (53). Even if a similar hypothesis was previously deduced from evidence showing a higher expression of CD163 in SLE skin lesions (54), it is still premature to confidently speculate that M2-like macrophages are undoubtedly responsible for the pathogenesis of PNGD firstly because CD163 is not a specific marker to macrophages (55–57). Secondly, because the study was conducted on only two patients, and finally, the pathogenesis is not fully understood. Further investigations are therefore needed.
Interestingly, MicroRNAs (miRNAs), a class of small non-coding RNAs, are reported to contribute to SLE development by promoting both M1-and M2-like macrophages. Indeed, evidence showed that miRNAs are involved in SLE progression by inducing M2-like macrophages polarization through activation of lymphocyte-derived DNA (ALD-DNA) (58). Conversely, other articles reported that miRNAs induce inflammation by favoring M1 polarization (59). Besides indexing the complexities of the behavior of macrophages in diseases and health, these data also highlight why macrophages cannot simply be categorized as builders or killers. Despite the oversimplified classification of these immune cells, we cannot simply speculate that M1-like macrophages only contribute to SLE development while M2-like macrophages alleviate the disease.
Macrophage activation syndrome (MAS) is a rare but severe complication seen in rheumatic disorders. Although it occurs in many rheumatic diseases, it is most common in idiopathic juvenile arthritis, SLE, and Kawasaki disease (60, 61). Recent studies have further revealed that MAS is most likely in SLE when the disease is diagnosed during childhood (62, 63). And even more likely when the child presents with lupus pancreatitis (64, 65). Clinically, the syndrome is represented by hepatic dysfunction, hepatosplenomegaly, clotting disorder, hyperferritinemia, pancytopenia, and high fever (66). Even though there is no published article explaining the role of macrophage polarization in SLE-MAS, hematologic and organ alterations in MAS are believed to result from the uncontrolled release of pro-inflammatory cytokines by abnormally activated T lymphocytes and macrophages (67). Given this implication of macrophages in the development of MAS and its association with SLE, it is reasonable that we consider this syndrome to be a part of this review.
Although the etiology of SLE-MAS remains a mystery, several factors have been reported to trigger this syndrome. The most common ones are infection and SLE flare. Other reported factors include abortion and parturition (68, 69). While some articles have described that change of therapy and malignancy could trigger SLE-MAS (70), another study suggests that no link was found between these two factors and the development of SLE-MAS (69). Among these triggering factors, infection is the one that can probably explain the implication of macrophages in the development of SLE-MAS. When infection cannot be eliminated, the immune system becomes inappropriately stimulated, leading to hyperinflammation via an uncontrolled release of cytokines from immune cells such as macrophages. The resulting cytokine storm is believed to be responsible for the development of MAS (71). So far, The most reported infective triggers include Epstein-Barr virus (EBV) and cytomegalovirus (CMV) (72).
Until now, the exact mechanism that leads to the development of MAS in SLE patients is not well established. Nevertheless, some pathways are believed to contribute to the pathogenesis of SLE-MAS. Exaggerated hypercytokinemia due to dysregulation of the macrophage-lymphocyte interaction is one way to understand the occurrence of SLE-MAS (71). Figure 2. Among cytokines, TNF-α is of particular significance as it is characteristic of SLE-MAS rather than other inflammatory diseases (71, 73). The importance of tumor necrosis factor is also reported in a recent study comparing cytokine levels in SLE patients with and without MAS. The same study has additionally shown that CXCL9 is likewise significantly elevated in SLE-MAS (74).
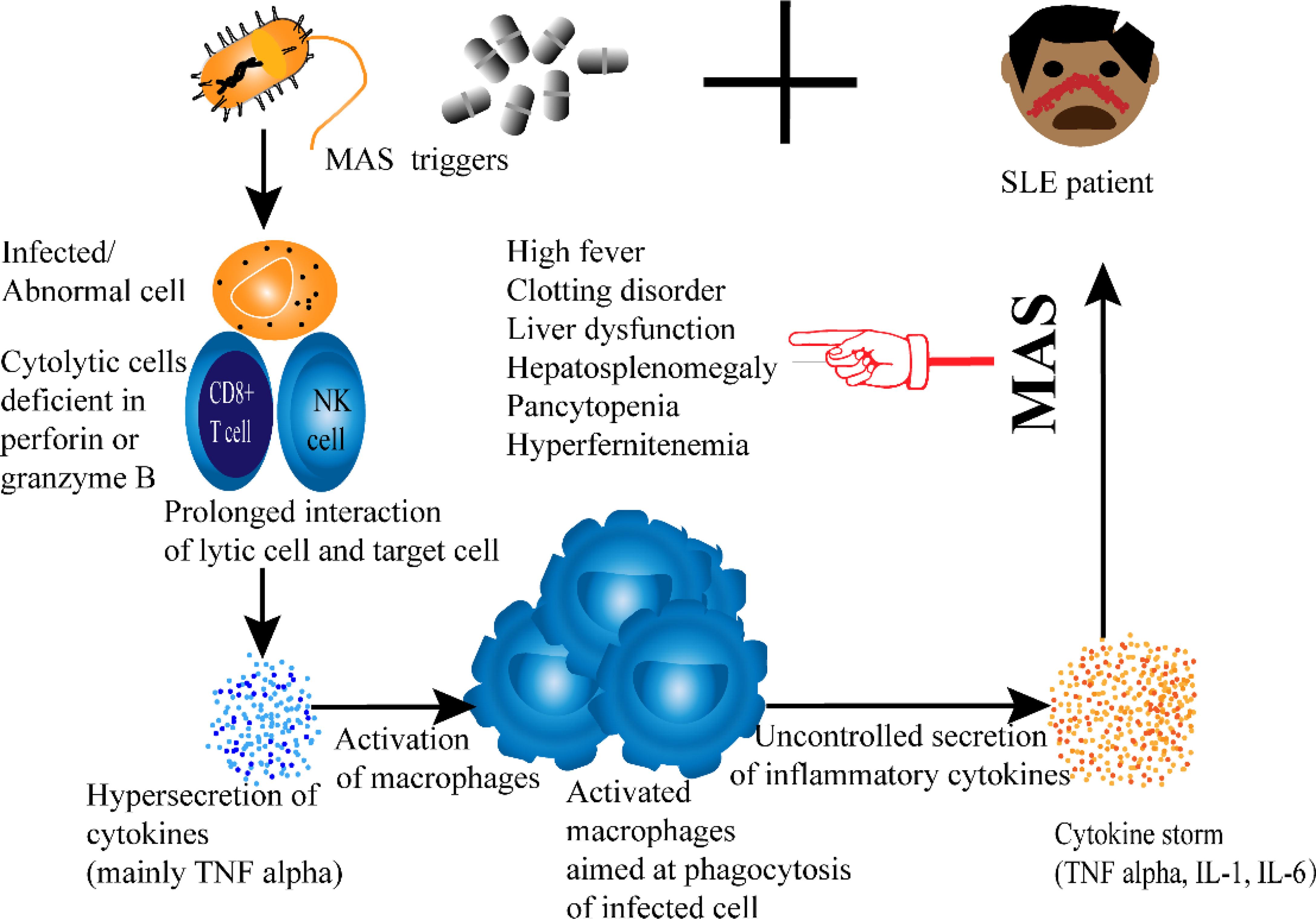
Figure 2 Proposed Pathogenesis of SLE-associated MAS.
The mechanism of SLE-MAS could also be explained by IgM type antilymphocyte antibody (ALAB) and a mutation on the MEFV gene. The Presence of ALAB on lymphocytes is reported to be proportional to disease severity. In fact, in a young patient diagnosed with SLE-MAS, IgM-ALAB could be observed on the surface of lymphocytes during the acute phase but not with disease improvement (71). Even if we still do not understand the exact mechanism of the syndrome, this indicates a strong association between ALAB and SLE-MAS development. Validating this theory with a large number of patients would be beneficial since ALAB could be used to assess the activity of the disease.
Similarly, a retrospective study has demonstrated that serum sCD163 level is proportional to SLE-MAS activity (75). While these data suggest the possible usefulness of CD163 as a significant biomarker for SLE-MAS, further investigation is necessary to address the limitation of the current study. Apart from being retrospective, some patients in this study were treated with dexamethasone before sample collection. Since corticosteroids are known to upregulate the expression of CD163, this could have affected the serum sCD163 levels of some subjects.
In a nutshell, despite progress in clarifying the occurrence of SLE-MAS, studies on this syndrome are mostly conducted in a very limited number of patients. Possibly due to the rareness of MAS. This limitation emphasizes the need for extended investigations, especially since the syndrome can be fatal.
The use of corticosteroids and immunosuppressants has considerably improved the prognosis of SLE (76, 77). However, despite being the cornerstones treatment for SLE, these drugs are associated with unwanted side effects, especially in children (78). Such issues bring up the great need for specific agents with fewer side effects. In order to reduce corticosteroids doses, several biological agents such as rituximab, belimumab have been developed, and some have yielded favorable results. Among the commonly used SLE medications, some are reported to impact macrophage polarization and therefore are summarized in Table 2.
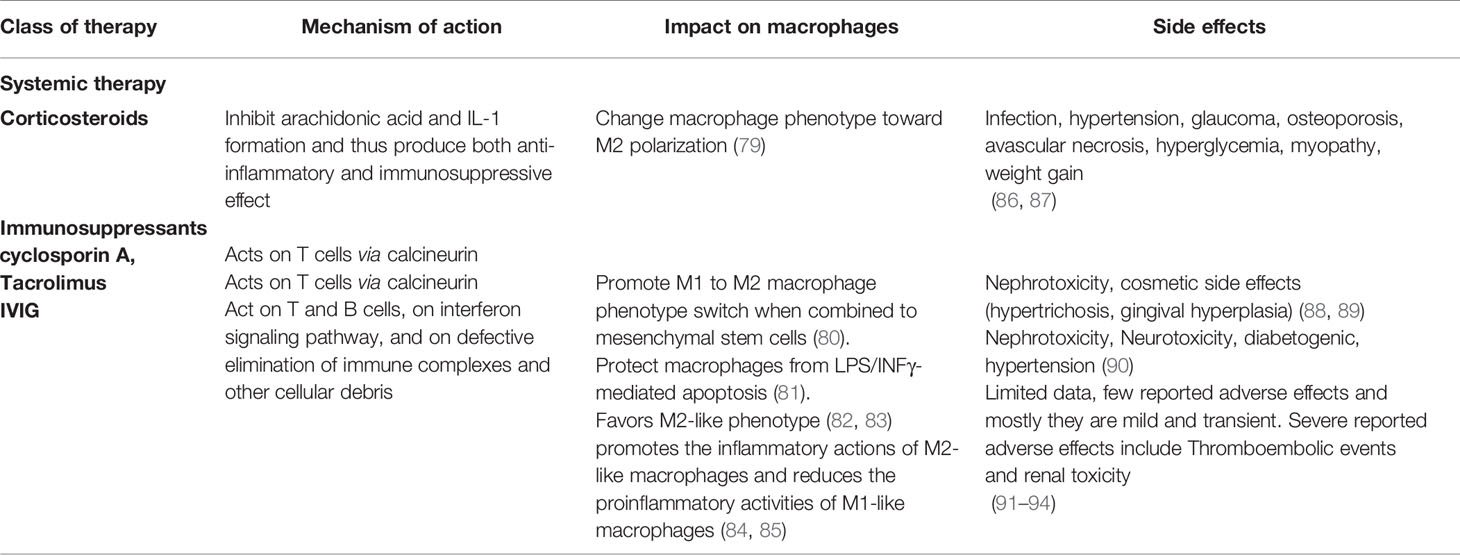
Table 2 Current SLE medications with an impact on macrophage polarization.
Many articles discuss the treatment options of systemic lupus erythematosus. However, to remain true to our topic, the treatment section of this review mainly focuses on two aspects. On the one hand, we discuss the management of patients with SLE-associated MAS, and on the other hand, we review the reported therapeutic options that target M1- and M2- like macrophages.
For rheumatic diseases induced-MAS such as SLE-MAS, therapy is directed toward controlling the hyperinflammation state. Usually, this can be achieved through a combination of high-dose corticosteroids and immunosuppressants (95–97). Interestingly, hydroxychloroquine has been reported to decrease the probability of developing MAS in SLE patients. The mechanism is not fully understood, but it is hypothesized that hydroxychloroquine decreases the production of IL-1, IL-6, and TNF-α, which inhibits toll-like receptor (TLR) 3 and 7 and consequently lowers the likelihood of developing SLE-MAS (98).
Among immunosuppressants, some agents are reported to be more effective than others. Indeed, even if cyclosporin is a good immunosuppressant (77, 99), for maintenance of remission, the combination of tacrolimus and corticosteroids has been reported to provide a better outcome. In the same study, intravenous immunoglobulin (IVIG) is proposed to be more effective when combined with oral tacrolimus as compared to cyclosporin A (100). Reported biological agents that have been used to treat SLE-MAS include Rituximab (101) and the IL-1 receptor antagonist anakinra (96).
Several ex vivo, in vivo, and in vitro studies have reported various experimental therapeutic approaches that affect the expression of M1-and M2-like macrophages. One of the investigated compounds is the Aryl hydrocarbon receptor (AhR) agonist indole-3-carbinol (I3C). AhR is a cytoplasmic receptor that exhibits numerous physiological functions in immune cells such as B cells, T cells, neutrophils, and macrophages (102). Emerging evidence suggests that I3C can be targeted to ameliorate diverse inflammatory and autoimmune conditions (103–106). Furthermore, it is reported that I3C can immunoregulate macrophages by activating AhR (107). An ex vivo study in SLE patients demonstrated that AhR activation could be mediated by I3C, resulting in a downregulation of M1 markers (CD86) and overexpression of M2 markers (CD163) (46). Meanwhile, an increase of anti-inflammatory cytokines and a decrease of inflammatory cytokines were induced, and consequently, inflammation was controlled (46). However, this study is not without limitations. Firstly, because CD163 can also be expressed by other cells and secondly because CD86 can be expressed by M2b macrophages (40, 108). Hence, the assessment of macrophages cannot be accurately achieved with these biomarkers alone. Similar ex vivo investigations demonstrated that I3C could assuredly alleviate lupus flares via macrophages regulation (109). The researchers have considered the environment’s influence on macrophage plasticity and used autologous plasma instead of M-CSF and GM-CSF synthetic growth factors so that in vivo conditions could be mimicked and bias avoided. Unfortunately, the small number of patients in this study and the characterization of M2-like macrophages using CD163+ minimizes its power.
Consistent with this theory, it is also suggested that sodium valproate alleviates inflammation in SLE patients. In fact, in an ex vivo study, sodium valproate was seen to upregulate the expression of anti-inflammatory cytokines from M2 phenotype macrophages (CD163) and to downregulate the expression of pro-inflammatory cytokines from M1 phenotype macrophages (CD86) (110). Nevertheless, the use of sodium valproate in neuropsychiatric lupus needs to be cautiously evaluated because the drug is reported to induce hyperammonemia which leads to a metabolic brain insult and the worsening of neuropsychiatric symptoms (111). In addition, previous studies have suggested a possible link between this drug and SLE development (112, 113). Such conflicting findings make the use of valproate in SLE controversial, and future studies need to look into this contradiction. Moreover, it is important to highlight again that CD163 alone cannot be used to characterize macrophages.
HMGB1 is another investigated molecule. As mentioned above, HMGB1 can trigger inflammation via inflammatory macrophages. However, an ex vivo study has revealed that the cooperation of C1q with HMGB1 can suppress inflammation by stimulating the polarization of M2-like macrophages (114). Such a finding is undoubtedly promising, but it should be noted that the study was conducted on murine lupus, and it is still unclear whether the same effect can be seen in human beings. Various preclinical studies have also explored the possible therapeutic effects of several inhibitors of HMGB1 (115) as well as HMGB1-specific antagonists (116), and the results are favorable.
Tuberous sclerosis complex 1 (TSC1) is an additional regulator of macrophage polarization. An in vivo study proved that the TSC1/2 complex helps to maintain macrophage homeostasis by regulating macrophage polarization through different signaling pathways (117). Via the Ras GTPase pathway, the TSC1/2 complex inhibits the M1 phenotype and contributes to autoimmune diseases’ amelioration. On the other hand, inhibition of the mammalian target of rapamycin (mTOR) pathway by TSC1/2 complex enhances M2 polarization (117). This makes TSC1 a potential regulator of macrophage polarization, and identifying its master role in macrophage activation could provide therapeutic strategies for autoimmune diseases. While the current study has demonstrated that TSC1/2 could prevent autoimmune diseases by controlling macrophage polarization, it does not address any specific autoimmune disease. Thus, validation in SLE needs to be determined.
Another compound that has been experimentally used to control SLE is pioglitazone. Pioglitazone corresponds to a peroxisome proliferator-activated receptor (PPAR)-γ agonist that is widely used to treat diabetes. In addition to its anti-diabetic effect, this drug also has anti-inflammatory properties (118). Such anti-inflammatory effect has been reported to benefit inflammatory diseases, including SLE. Indeed, an ex vivo study of newly diagnosed lupus patients showed that activation of PPAR-γ by pioglitazone could induce anti-inflammatory properties by promoting the expression of M2-like macrophages and decreasing the expression of M1-like macrophage (119). Likewise, an in vivo study has acknowledged this contribution of pioglitazone in alleviating murine LN. Even though the study has not mentioned macrophages’ involvement, it confirmed that the use of pioglitazone induces a protective effect of PPAR-γ against LN (120).
Clinically, Virgin olive oil (VOO) has been successfully used to ease symptoms in patients with lupus. A study conducted in humans proved that the phenolic fraction of extra virgin oil provided a beneficial effect on immune-inflammatory diseases such as SLE (121). Although this particular article does not state that this immunoregulation is achieved by macrophages, a more recent study has revealed that VOO dampens TLR4, which plays a role in both macrophage activation and polarization (122). Moreover, a comparison of the use of virgin olive oil and sunflower oil in murine lupus showed that the former could block the expression of M1 subtype macrophage while enhancing the M2 phenotype (122). Despite accumulated evidence showing the role of VOO in alleviating chronic immune-mediated diseases such as SLE, the exact mechanism is still unclear, and this limits its clinical use.
One of the processes that determine cell fate and cellular development is Notch signaling. Four Notch receptors (Notch1-4) have been identified in mammals, and impaired regulation of Notch1 signaling is associated with many diseases (123). The association of Notch signaling with SLE is not a new concept as a defective expression of Notch 1 in SLE T cells has been reported previously (124, 125). However, the involvement of Notch 1 in SLE macrophages has been newly established. Indeed, both in vitro and in vivo studies demonstrated that blockage of Notch 1 signaling could alleviate murine lupus by blunting M2b polarization (126). Although blocking Notch 1 could also suppress M1-like macrophage polarization and attenuate inflammation in cardiovascular diseases (127), data about Notch 1 signaling and M1 polarization in SLE is lacking. Further studies are required to explore this possible connection. Another investigated approach that could also improve SLE is the administration of PAM3CSK4 (PAM3), a TLR 2/1 agonist. By using PAM3, researchers were able to stimulate the polarization of monocytes from SLE patients toward M2-like macrophages in vitro. Moreover, PAM3 treatment delayed the disease progression and prolonged survival in murine models (128, 129). Similar to the inducers of M2 polarization mentioned above, the use of artemisinin in vivo has offered a therapeutic effect on female mice with lupus by stimulating the secretion of anti-inflammatory cytokines from M2-like macrophages (130). Another compound that could help patients with lupus is Liver x-receptor Alpha (LXRα). LXRα is one of the two isoforms of liver X receptors, a nuclear receptor superfamily member. Besides being an essential receptor for cholesterol regulation, this protein also regulates the activation of macrophages, and its therapeutic effects on autoimmune diseases are gaining much attention (131, 132). Recently, an in vivo study has shown that LXR α could promote M2-like macrophages polarization in murine lupus and subsequently prevent the occurrence of diffuse alveolar hemorrhage, a deadly complication of lupus (133).
Clinically, azithromycin, a macrolide antibiotic useful for controlling inflammation in various diseases (134), has been proven to ease lupus flare via the regulation of M2-like macrophages. In vivo and in vitro studies in SLE patients revealed that azithromycin promotes M2 polarization via the pl3k/Akt pathway, resulting in the suppression of inflammation and SLE remission (135). Albeit its reported side effects (136), azithromycin does have a good record of safety, and exploring it could offer a new therapeutic strategy for lupus patients. Another drug that favors the M2 phenotype is steroid. A recent study has demonstrated that the clinical use of steroids can change macrophage phenotype toward M2 polarization and consequently suppress inflammation in lupus patients (79). However, this phenotype switch is also reported to worsen LN. The study showed that by promoting M2 polarization, steroids also induce interstitial fibrosis and exacerbate chronic glomerular lesions (79). This finding highlights the great need to evaluate the possible side effects of inducing a macrophage phenotype change.
The appreciation of macrophage polarization and plasticity in SLE has allowed scientists to gain a better insight into the development of this autoimmune disease. Even though it is still yet to be determined whether the deregulation of macrophages phenotypes observed in SLE is a cause or consequence of the pathogenic process, numerous studies have reported an amelioration of the disease activity after restoring the balance between M1/M2 phenotypes. Indeed, exposure of M1-like macrophages to M2 modulators or vice versa was seen to induce re-polarization of macrophages and consequently control lupus flares both in vitro and in vivo. Currently, various new and exciting macrophages-based therapeutic strategies are being exploited, and they appear to be promising. These therapeutic approaches bring a glimmer of hope for the lupus community as they provide us with a new pathway toward the treatment of SLE. However, there is still work to be done, and several gaps need to be addressed in the future in order to get more clarification about these macrophages-centered therapies. First of all, scientists have established the effect of various compounds on macrophages polarization, but their possible physiological effects on other cells are poorly studied. SLE is a systemic disease with numerous manifestations in different organs. While inducing a macrophage phenotype switch has been proven to ameliorate some aspects of lupus flares, it could also worsen other manifestations of the disease, as it is seen with corticosteroids (79). Such possible outcome needs to be carefully evaluated and enlightened in future work. Secondly, the influence exerted by the environment on the plasticity of macrophages is non-negligible, and this needs to be taken into consideration. In vitro findings cannot directly be translated into in vivo. Therefore, we still do not know how some of these macrophages-centered therapies would behave in the human body. Paying closer attention to this fact is necessary in order to exploit these immune cells better. Finally, the field could also benefit from more studies in children because even though they do not represent the majority of the SLE community, they do have the most severe manifestations, and children are known to function differently from adults.
YJ was resident and provided the main plan and conceptual ideas. MA was resident and wrote the manuscript. XW was professor and supervised the work. All authors contributed to the article and approved the submitted version.
This work was supported by the Natural Science Foundation of Hunan Province (No.2021JJ30960) and Natural Science Foundation of Hunan Province (No.2021JJ40836).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Holubar K. Terminology and Iconography of Lupus Erythematosus. A Historical Vignette. Am J Dermatopathol (1980) 2(3):239–42. doi: 10.1097/00000372-198000230-00010
2. Felten R, Lipsker D, Sibilia J, Chasset F, Arnaud L. The History of Lupus Throughout the Ages. J Am Acad Dermatol (2020). doi: 10.1016/j.jaad.2020.04.150
3. Pons-Estel GJ, Ugarte-Gil MF, Alarcón GS. Epidemiology of Systemic Lupus Erythematosus. Expert Rev Clin Immunol (2017) 13(8):799–814. doi: 10.1080/1744666X.2017.1327352
4. Lau CS, Yin G, Mok MY. Ethnic and Geographical Differences in Systemic Lupus Erythematosus: An Overview. Lupus (2006) 15(11):715–9. doi: 10.1177/0961203306072311
5. Levy DM, Kamphuis S. Systemic Lupus Erythematosus in Children and Adolescents. Pediatr Clin North Am (2012) 59(2):345–64. doi: 10.1016/j.pcl.2012.03.007
6. Sawada T, Fujimori D, Yamamoto Y. Systemic Lupus Erythematosus and Immunodeficiency. Immunol Med (2019) 42(1):1–9. doi: 10.1080/25785826.2019.1628466
7. Tsokos GC, Lo MS, Costa Reis P, Sullivan KE. New Insights Into the Immunopathogenesis of Systemic Lupus Erythematosus. Nat Rev Rheumatol (2016) 12(12):716–30. doi: 10.1038/nrrheum.2016.186
8. Li F, Yang Y, Zhu X, Huang L, Xu J. Macrophage Polarization Modulates Development of Systemic Lupus Erythematosus. Cell Physiol Biochem (2015) 37(4):1279–88. doi: 10.1159/000430251
9. Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F, et al. Macrophage Plasticity, Polarization, and Function in Health and Disease. J Cell Physiol (2018) 233(9):6425–40. doi: 10.1002/jcp.26429
10. Murray PJ. Macrophage Polarization. Annu Rev Physiol (2017) 79:541–66. doi: 10.1146/annurev-physiol-022516-034339
11. Martinez FO, Sica A, Mantovani A, Locati M. Macrophage Activation and Polarization. Front Biosci (2008) 13:453–61. doi: 10.2741/2692
12. Ruytinx P, Proost P, Van Damme J, Struyf S. Chemokine-Induced Macrophage Polarization in Inflammatory Conditions. Front Immunol (2018) 9. doi: 10.3389/fimmu.2018.01930
13. Deng H, Sun Y, Zeng W, Li H, Guo M, Yang L, et al. New Classification of Macrophages in Plaques: A Revolution. Curr Atheroscler Rep (2020) 22(8):31. doi: 10.1007/s11883-020-00850-y
14. Orecchioni M, Ghosheh Y, Pramod AB, Ley K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front Immunol (2019) 10. doi: 10.3389/fimmu.2019.01084
15. Jain N, Moeller J, Vogel V. Mechanobiology of Macrophages: How Physical Factors Coregulate Macrophage Plasticity and Phagocytosis. Annu Rev BioMed Eng (2019) 21:267–97. doi: 10.1146/annurev-bioeng-062117-121224
16. Labonte AC, Tosello-Trampont AC, Hahn YS. The Role of Macrophage Polarization in Infectious and Inflammatory Diseases. Mol Cells (2014) 37(4):275–85. doi: 10.14348/molcells.2014.2374
17. Funes SC, Rios M, Escobar-Vera J, Kalergis AM. Implications of Macrophage Polarization in Autoimmunity. Immunology (2018) 154(2):186–95. doi: 10.1111/imm.12910
18. Italiani P, Boraschi D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front Immunol (2014) 5:514. doi: 10.3389/fimmu.2014.00514
19. Atri C, Guerfali FZ, Laouini D. Role of Human Macrophage Polarization in Inflammation During Infectious Diseases. Int J Mol Sci (2018) 19(6). doi: 10.3390/ijms19061801
20. Cuda CM, Pope RM, Perlman H. The Inflammatory Role of Phagocyte Apoptotic Pathways in Rheumatic Diseases. Nat Rev Rheumatol (2016) 12(9):543–58. doi: 10.1038/nrrheum.2016.132
21. Orme J, Mohan C. Macrophage Subpopulations in Systemic Lupus Erythematosus. Discov Med (2012) 13(69):151–8.
22. Labonte AC, Kegerreis B, Geraci NS, Bachali P, Madamanchi S, Robl R, et al. Identification of Alterations in Macrophage Activation Associated With Disease Activity in Systemic Lupus Erythematosus. PloS One (2018) 13(12):e0208132. doi: 10.1371/journal.pone.0208132
23. Niu X-L, Feng D, Hao S, Kuang X-Y, Wu Y, Zhu G-H, et al. The Significance of M1/M2 Macrophage-Like Monocytes in Children With Systemic Lupus Erythematosus. Eur J Inflamm (2019) 17. doi: 10.1177/2058739218824463
24. Hou J, Zhang M, Ding Y, Wang X, Li T, Gao P, et al. Circulating CD14(+)CD163(+)CD206(+) M2 Monocytes Are Increased in Patients With Early Stage of Idiopathic Membranous Nephropathy. Mediators Inflamm (2018) 2018:5270657. doi: 10.1155/2018/5270657
25. Han S, Zhuang H, Lee PY, Li M, Yang L, Nigrovic PA, et al. NF-E2-Related Factor 2 Regulates Interferon Receptor Expression and Alters Macrophage Polarization in Lupus. Arthritis Rheumatol (2020) 72(10):1707–20. doi: 10.1002/art.41383
26. Al Gadban MM, Alwan MM, Smith KJ, Hammad SM. Accelerated Vascular Disease in Systemic Lupus Erythematosus: Role of Macrophage. Clin Immunol (2015) 157(2):133–44. doi: 10.1016/j.clim.2015.01.008
27. Rho YH, Solus J, Raggi P, Oeser A, Gebretsadik T, Shintani A, et al. Macrophage Activation and Coronary Atherosclerosis in Systemic Lupus Erythematosus and Rheumatoid Arthritis. Arthritis Care Res (Hoboken) (2011) 63(4):535–41. doi: 10.1002/acr.20365
28. Jinnouchi H, Guo L, Sakamoto A, Torii S, Sato Y, Cornelissen A, et al. Diversity of Macrophage Phenotypes and Responses in Atherosclerosis. Cell Mol Life Sci (2020) 77(10):1919–32. doi: 10.1007/s00018-019-03371-3
29. Burbano C, Villar-Vesga J, Vásquez G, Muñoz-Vahos C, Rojas M, Castaño D. Proinflammatory Differentiation of Macrophages Through Microparticles That Form Immune Complexes Leads to T- and B-Cell Activation in Systemic Autoimmune Diseases. Front Immunol (2019) 10:2058. doi: 10.3389/fimmu.2019.02058
30. Ivanov S, Dragoi AM, Wang X, Dallacosta C, Louten J, Musco G, et al. A Novel Role for HMGB1 in TLR9-Mediated Inflammatory Responses to Cpg-DNA. Blood (2007) 110(6):1970–81. doi: 10.1182/blood-2006-09-044776
31. Liu T, Son M, Diamond B. HMGB1 in Systemic Lupus Erythematosus. Front Immunol (2020) 11:1057. doi: 10.3389/fimmu.2020.01057
32. Lu M, Yu S, Xu W, Gao B, Xiong S. HMGB1 Promotes Systemic Lupus Erythematosus by Enhancing Macrophage Inflammatory Response. J Immunol Res (2015) 2015:946748. doi: 10.1155/2015/946748
33. Huang Q, Shen S, Qu H, Huang Y, Wu D, Jiang H, et al. Expression of HMGB1 and TLR4 in Neuropsychiatric Systemic Lupus Erythematosus Patients With Seizure Disorders. Ann Trans Med (2020) 8(1):9–9. doi: 10.21037/atm.2019.12.44
34. Baba T, Mukaida N. Role of Macrophage Inflammatory Protein (MIP)-1α/CCL3 in Leukemogenesis. Mol Cell Oncol (2014) 1(1):e29899. doi: 10.4161/mco.29899
35. Tokunaga R, Zhang W, Naseem M, Puccini A, Berger MD, Soni S, et al. CXCL9, CXCL10, CXCL11/CXCR3 Axis for Immune Activation - a Target for Novel Cancer Therapy. Cancer Treat Rev (2018) 63:40–7. doi: 10.1016/j.ctrv.2017.11.007
36. Gongol B, Marin T, Peng IC, Woo B, Martin M, King S, et al. Ampkα2 Exerts Its Anti-Inflammatory Effects Through PARP-1 and Bcl-6. Proc Natl Acad Sci USA (2013) 110(8):3161–6. doi: 10.1073/pnas.1222051110
37. Hesketh M, Sahin KB, West ZE, Murray RZ. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int J Mol Sci (2017) 18(7). doi: 10.3390/ijms18071545
38. Arora S, Dev K, Agarwal B, Das P, Syed MA. Macrophages: Their Role, Activation and Polarization in Pulmonary Diseases. Immunobiology (2018) 223(4-5):383–96. doi: 10.1016/j.imbio.2017.11.001
39. Rőszer T. Understanding the Mysterious M2 Macrophage Through Activation Markers and Effector Mechanisms. Mediators Inflamm (2015) 2015:816460. doi: 10.3389/fimmu.2019.01084
40. Wang LX, Zhang SX, Wu HJ, Rong XL, Guo J. M2b Macrophage Polarization and Its Roles in Diseases. J Leukoc Biol (2019) 106(2):345–58. doi: 10.1002/JLB.3RU1018-378RR
41. Shrivastava R, Shukla N. Attributes of Alternatively Activated (M2) Macrophages. Life Sci (2019) 224:222–31. doi: 10.1016/j.lfs.2019.03.062
42. Sica A, Mantovani A. Macrophage Plasticity and Polarization: In Vivo Veritas. J Clin Invest (2012) 122(3):787–95. doi: 10.1172/JCI59643
43. Colin S, Chinetti-Gbaguidi G, Staels B. Macrophage Phenotypes in Atherosclerosis. Immunol Rev (2014) 262(1):153–66. doi: 10.1111/imr.12218
44. Olmes G, Büttner-Herold M, Ferrazzi F, Distel L, Amann K, Daniel C. CD163+ M2c-Like Macrophages Predominate in Renal Biopsies From Patients With Lupus Nephritis. Arthritis Res Ther (2016) 18:90. doi: 10.1186/s13075-016-0989-y
45. Sung SJ, Ge Y, Dai C, Wang H, Fu SM, Sharma R, et al. Dependence of Glomerulonephritis Induction on Novel Intraglomerular Alternatively Activated Bone Marrow-Derived Macrophages and Mac-1 and PD-L1 in Lupus-Prone NZM2328 Mice. J Immunol (2017) 198(7):2589–601. doi: 10.4049/jimmunol.1601565
46. Mohammadi S, Memarian A, Sedighi S, Behnampour N, Yazdani Y. Immunoregulatory Effects of Indole-3-Carbinol on Monocyte-Derived Macrophages in Systemic Lupus Erythematosus: A Crucial Role for Aryl Hydrocarbon Receptor. Autoimmunity (2018) 51(5):199–209. doi: 10.1080/08916934.2018.1494161
47. Ma C, Xia Y, Yang Q, Zhao Y. The Contribution of Macrophages to Systemic Lupus Erythematosus. Clin Immunol (2019) 207:1–9. doi: 10.1016/j.clim.2019.06.009
48. Katsiari CG, Liossis SN, Sfikakis PP. The Pathophysiologic Role of Monocytes and Macrophages in Systemic Lupus Erythematosus: A Reappraisal. Semin Arthritis Rheum (2010) 39(6):491–503. doi: 10.1016/j.semarthrit.2008.11.002
49. Boeltz S, Hagen M, Knopf J, Mahajan A, Schick M, Zhao Y, et al. Towards a Pro-Resolving Concept in Systemic Lupus Erythematosus. Semin Immunopathol (2019) 41(6):681–97. doi: 10.1007/s00281-019-00760-5
50. Tajbakhsh A, Rezaee M, Barreto GE, Moallem SA, Henney NC, Sahebkar A. The Role of Nuclear Factors as “Find-Me”/Alarmin Signals and Immunostimulation in Defective Efferocytosis and Related Disorders. Int Immunopharmacol (2020) 80:106134. doi: 10.1016/j.intimp.2019.106134
51. Kishimoto D, Kirino Y, Tamura M, Takeno M, Kunishita Y, Takase-Minegishi K, et al. Dysregulated Heme Oxygenase-1(Low) M2-Like Macrophages Augment Lupus Nephritis via Bach1 Induced by Type I Interferons. Arthritis Res Ther (2018) 20(1):64. doi: 10.1186/s13075-018-1568-1
52. Kanellopoulos J, Chen X, Wen Z, Xu W, Xiong S. Granulin Exacerbates Lupus Nephritis via Enhancing Macrophage M2b Polarization. PloS One (2013) 8(6). doi: 10.1371/journal.pone.0065542
53. Terai S, Ueda-Hayakawa I, Nguyen CTH, Ly NTM, Yamazaki F, Kambe N, et al. Palisaded Neutrophilic and Granulomatous Dermatitis Associated With Systemic Lupus Erythematosus: Possible Involvement of CD163(+) M2 Macrophages in Two Cases, and a Review of Published Works. Lupus (2018) 27(14):2220–7. doi: 10.1177/0961203318809892
54. Nakayama W, Jinnin M, Makino K, Kajihara I, Makino T, Fukushima S, et al. CD163 Expression is Increased in the Involved Skin and Sera of Patients With Systemic Lupus Erythematosus. Eur J Dermatol (2012) 22(4):512–7. doi: 10.1684/ejd.2012.1756
55. Sulahian TH, Högger P, Wahner AE, Wardwell K, Goulding NJ, Sorg C, et al. Human Monocytes Express CD163, Which is Upregulated by IL-10 and Identical to P155. Cytokine (2000) 12(9):1312–21. doi: 10.1006/cyto.2000.0720
56. Barros MH, Hauck F, Dreyer JH, Kempkes B, Niedobitek G. Macrophage Polarisation: An Immunohistochemical Approach for Identifying M1 and M2 Macrophages. PloS One (2013) 8(11):e80908. doi: 10.1371/journal.pone.0080908
57. Bourdely P, Anselmi G, Vaivode K, Ramos RN, Missolo-Koussou Y, Hidalgo S, et al. Transcriptional and Functional Analysis of CD1c(+) Human Dendritic Cells Identifies a CD163(+) Subset Priming CD8(+)CD103(+) T Cells. Immunity (2020) 53(2):335–52.e8. doi: 10.1016/j.immuni.2020.06.002
58. Xiao P, Dong C, Yue Y, Xiong S. Dynamic Expression of MicroRNAs in M2b Polarized Macrophages Associated With Systemic Lupus Erythematosus. Gene (2014) 547(2):300–9. doi: 10.1016/j.gene.2014.06.065
59. Essandoh K, Li Y, Huo J, Fan GC. MiRNA-Mediated Macrophage Polarization and Its Potential Role in the Regulation of Inflammatory Response. Shock (2016) 46(2):122–31. doi: 10.1097/SHK.0000000000000604
60. Abdul Jalal M, Mohammad Iqbal KM, Mahadevan P, Neethu S. Systemic Lupus Erythematosus Presenting as Macrophage Activation Syndrome in an Adult Male. Curr Med Issues (2020) 18(1). doi: 10.4103/cmi.cmi_50_19
61. Sen ES, Clarke SL, Ramanan AV. Macrophage Activation Syndrome. Indian J Pediatr (2016) 83(3):248–53. doi: 10.1007/s12098-015-1877-1
62. Borgia RE, Gerstein M, Levy DM, Silverman ED, Hiraki LT. Features, Treatment, and Outcomes of Macrophage Activation Syndrome in Childhood-Onset Systemic Lupus Erythematosus. Arthritis Rheumatol (2018) 70(4):616–24. doi: 10.1002/art.40417
63. Poudel P, Swe T, Rayancha S. A Rare Case of Macrophage Activation Syndrome Presenting as the First Manifestation of Systemic Lupus Erythematosus. J Investig Med High Impact Case Rep (2018) 6:2324709618812196. doi: 10.1177/2324709618812196
64. Lin Q, Zhang M, Tang H, Shen Y, Zhu Y, Xu Q, et al. Acute Pancreatitis and Macrophage Activation Syndrome in Pediatric Systemic Lupus Erythematosus: Case-Based Review. Rheumatol Int (2020) 40(5):811–9. doi: 10.1007/s00296-019-04388-4
65. Campos LM, Omori CH, Lotito AP, Jesus AA, Porta G, Silva CA. Acute Pancreatitis in Juvenile Systemic Lupus Erythematosus: A Manifestation of Macrophage Activation Syndrome? Lupus (2010) 19(14):1654–8. doi: 10.1177/0961203310378863
66. Alongi A, Naddei R, De Miglio L, Natoli V, Ravelli A. Macrophage Activation Syndrome in Pediatrics. Pediatr Allergy Immunol (2020) 31(Suppl 24):13–5. doi: 10.1111/pai.13158
67. Strippoli R, Caiello I, De Benedetti F. Reaching the Threshold: A Multilayer Pathogenesis of Macrophage Activation Syndrome. J Rheumatol (2013) 40(6):761–7. doi: 10.3899/jrheum.121233
68. Bracaglia C, Prencipe G, De Benedetti F. Macrophage Activation Syndrome: Different Mechanisms Leading to a One Clinical Syndrome. Pediatr Rheumatol Online J (2017) 15(1):5. doi: 10.1186/s12969-016-0130-4
69. Liu AC, Yang Y, Li MT, Jia Y, Chen S, Ye S, et al. Macrophage Activation Syndrome in Systemic Lupus Erythematosus: A Multicenter, Case-Control Study in China. Clin Rheumatol (2018) 37(1):93–100. doi: 10.1007/s10067-017-3625-6
70. Szulawski R, Kourlas PJ, Antonchak M. Macrophage Activation Syndrome (MAS) in a Recently Released Prisoner With Systemic Lupus Erythematosus (SLE). Am J Case Rep (2018) 19:734–8. doi: 10.12659/AJCR.906154
71. Shimizu M, Yokoyama T, Tokuhisa Y, Ishikawa S, Sakakibara Y, Ueno K, et al. Distinct Cytokine Profile in Juvenile Systemic Lupus Erythematosus-Associated Macrophage Activation Syndrome. Clin Immunol (2013) 146(2):73–6. doi: 10.1016/j.clim.2012.11.004
72. Awasthi S, Upreti S. Macrophage Activation Syndrome in a Patient With Systemic Lupus Erythematosus (SLE) and Dual Viremia. J Community Hosp Intern Med Perspect (2020) 10(5):470–4. doi: 10.1080/20009666.2020.1787811
73. Mizuta M, Shimizu M, Irabu H, Usami M, Inoue N, Nakagishi Y, et al. Comparison of Serum Cytokine Profiles in Macrophage Activation Syndrome Complicating Different Background Rheumatic Diseases in Children. Rheumatol (Oxford) (2021) 60(1):231–8. doi: 10.1093/rheumatology/keaa299
74. Usami M, Shimizu M, Mizuta M, Inoue N, Irabu H, Sakumura N, et al. Extensive Serum Biomarker Analysis in Patients With Macrophage Activation Syndrome Associated With Systemic Lupus Erythematosus. Clin Immunol (2019) 208:108255. doi: 10.1016/j.clim.2019.108255
75. Nishino A, Katsumata Y, Kawasumi H, Hirahara S, Kawaguchi Y, Yamanaka H. Usefulness of Soluble CD163 as a Biomarker for Macrophage Activation Syndrome Associated With Systemic Lupus Erythematosus. Lupus (2019) 28(8):986–94. doi: 10.1177/0961203319860201
76. Tanaka Y. State-of-the-Art Treatment of Systemic Lupus Erythematosus. Int J Rheum Dis (2020) 23(4):465–71. doi: 10.1111/1756-185X.13817
77. Fava A, Petri M. Systemic Lupus Erythematosus: Diagnosis and Clinical Management. J Autoimmun (2019) 96:1–13. doi: 10.1016/j.jaut.2018.11.001
78. Deng J, Chalhoub NE, Sherwin CM, Li C, Brunner HI. Glucocorticoids Pharmacology and Their Application in the Treatment of Childhood-Onset Systemic Lupus Erythematosus. Semin Arthritis Rheum (2019) 49(2):251–9. doi: 10.1016/j.semarthrit.2019.03.010
79. Ikezumi Y, Kondoh T, Matsumoto Y, Kumagai N, Kaneko M, Hasegawa H, et al. Steroid Treatment Promotes an M2 Anti-Inflammatory Macrophage Phenotype in Childhood Lupus Nephritis. Pediatr Nephrol (2021) 36(2):349–59. doi: 10.1007/s00467-020-04734-w
80. Hajkova M, Javorkova E, Zajicova A, Trosan P, Holan V, Krulova M. A Local Application of Mesenchymal Stem Cells and Cyclosporine a Attenuates Immune Response by a Switch in Macrophage Phenotype. J Tissue Eng Regener Med (2017) 11(5):1456–65. doi: 10.1002/term.2044
81. Hortelano S, López-Collazo E, Boscá L. Protective Effect of Cyclosporin a and FK506 From Nitric Oxide-Dependent Apoptosis in Activated Macrophages. Br J Pharmacol (1999) 126(5):1139–46. doi: 10.1038/sj.bjp.0702422
82. Bai L, Gabriels K, Wijnands E, Rousch M, Daemen MJ, Tervaert JW, et al. Low- But Not High-Dose FK506 Treatment Confers Atheroprotection Due to Alternative Macrophage Activation and Unaffected Cholesterol Levels. Thromb Haemost (2010) 104(1):143–50. doi: 10.1160/th09-07-0502
83. Kannegieter NM, Hesselink DA, Dieterich M, Kraaijeveld R, Rowshani AT, Leenen PJ, et al. The Effect of Tacrolimus and Mycophenolic Acid on CD14+ Monocyte Activation and Function. PloS One (2017) 12(1):e0170806. doi: 10.1371/journal.pone.0170806
84. Domínguez-Soto A, de las Casas-Engel M, Bragado R, Medina-Echeverz J, Aragoneses-Fenoll L, Martin-Gayo E, et al. Intravenous Immunoglobulin Promotes Antitumor Responses by Modulating Macrophage Polarization. J Immunol (2014) 193(10):5181–9. doi: 10.4049/jimmunol.1303375
85. Saha C, Kothapalli P, Patil V, ManjunathaReddy GB, Kaveri SV, Bayry J. Intravenous Immunoglobulin Suppresses the Polarization of Both Classically and Alternatively Activated Macrophages. Hum Vaccin Immunother (2020) 16(2):233–9. doi: 10.1080/21645515.2019.1602434
86. Kragballe K. Topical Corticosteroids: Mechanisms of Action. Acta Derm Venereol Suppl (Stockh) (1989) 151:7–10; discussion 47-52.
87. Kasturi S, Sammaritano LR. Corticosteroids in Lupus. Rheum Dis Clin North Am (2016) 42(1):47–62, viii. doi: 10.1016/j.rdc.2015.08.007
88. Ho S, Clipstone N, Timmermann L, Northrop J, Graef I, Fiorentino D, et al. The Mechanism of Action of Cyclosporin a and FK506. Clin Immunol Immunopathol (1996) 80(3 Pt 2):S40–5. doi: 10.1006/clin.1996.0140
89. Mihatsch MJ, Kyo M, Morozumi K, Yamaguchi Y, Nickeleit V, Ryffel B. The Side-Effects of Ciclosporine-a and Tacrolimus. Clin Nephrol (1998) 49(6):356–63.
90. Yu M, Liu M, Zhang W, Ming Y. Pharmacokinetics, Pharmacodynamics and Pharmacogenetics of Tacrolimus in Kidney Transplantation. Curr Drug Metab (2018) 19(6):513–22. doi: 10.2174/1389200219666180129151948
91. Heyneman CA, Gudger CA, Beckwith JV. Intravenous Immune Globulin for Inducing Remissions in Systemic Lupus Erythematosus. Ann Pharmacother (1997) 31(2):242–4. doi: 10.1177/106002809703100218
92. Zandman-Goddard G, Krauthammer A, Levy Y, Langevitz P, Shoenfeld Y. Long-Term Therapy With Intravenous Immunoglobulin is Beneficial in Patients With Autoimmune Diseases. Clin Rev Allergy Immunol (2012) 42(2):247–55. doi: 10.1007/s12016-011-8278-7
93. Chasset F, Francès C. Current Concepts and Future Approaches in the Treatment of Cutaneous Lupus Erythematosus: A Comprehensive Review. Drugs (2019) 79(11):1199–215. doi: 10.1007/s40265-019-01151-8
94. Martínez T, Garcia-Robledo JE, Plata I, Urbano MA, Posso-Osorio I, Rios-Serna LJ, et al. Mechanisms of Action and Historical Facts on the Use of Intravenous Immunoglobulins in Systemic Lupus Erythematosus. Autoimmun Rev (2019) 18(3):279–86. doi: 10.1016/j.autrev.2018.10.002
95. Bennett TD, Fluchel M, Hersh AO, Hayward KN, Hersh AL, Brogan TV, et al. Macrophage Activation Syndrome in Children With Systemic Lupus Erythematosus and Children With Juvenile Idiopathic Arthritis. Arthritis Rheum (2012) 64(12):4135–42. doi: 10.1002/art.34661
96. Kubler L, Bittmann I, Kuipers JG. Macrophage Activation Syndrome Triggered by Active Systemic Lupus Erythematosus: Successful Treatment by Interleukin-1 Inhibition (Anakinra). Z Rheumatol (2020) 79(10):1040–5. doi: 10.1007/s00393-020-00836-w
97. Dall’Ara F, Cavazzana I, Frassi M, Taraborelli M, Fredi M, Franceschini F, et al. Macrophage Activation Syndrome in Adult Systemic Lupus Erythematosus: Report of Seven Adult Cases From a Single Italian Rheumatology Center. Reumatismo (2018) 70(2):100–5. doi: 10.4081/reumatismo.2018.1023
98. Cohen EM, D'Silva K, Kreps D, Son MB, Costenbader KH. Arthritis and Use of Hydroxychloroquine Associated With a Decreased Risk of Macrophage Activation Syndrome Among Adult Patients Hospitalized With Systemic Lupus Erythematosus. Lupus (2018) 27(7):1065–71. doi: 10.1177/0961203318759428
99. Cruz-Perez F, Vila S, Rios G, Vila LM. Efficacy of Cyclosporine in the Induction and Maintenance of Remission in a Systemic Lupus Erythematosus Patient Presenting With Macrophage-Activating Syndrome. Case Rep Rheumatol (2018) 2018:1961585. doi: 10.1155/2018/1961585
100. Aoyama-Maeda N, Horino T, Ichii O, Terada Y. Macrophage Activation Syndrome Associated With Systemic Lupus Erythematosus Treated Successfully With the Combination of Steroid Pulse, Immunoglobulin and Tacrolimus. Rom J Intern Med (2018) 56(2):117–21. doi: 10.1515/rjim-2017-0043
101. Junga Z, Stitt R, Tracy C, Keith M. Novel Use of Rituximab in Macrophage Activation Syndrome Secondary to Systemic Lupus Erythematosus. BMJ Case Rep (2017) 2017. doi: 10.1136/bcr-2017-221347
102. Hao N, Whitelaw ML. The Emerging Roles of Ahr in Physiology and Immunity. Biochem Pharmacol (2013) 86(5):561–70. doi: 10.1016/j.bcp.2013.07.004
103. Khan AS, Langmann T. Indole-3-Carbinol Regulates Microglia Homeostasis and Protects the Retina From Degeneration. J Neuroinflamm (2020) 17(1):327. doi: 10.1186/s12974-020-01999-8
104. Busbee PB, Menzel L, Alrafas HR, Dopkins N, Becker W, Miranda K, et al. Indole-3-Carbinol Prevents Colitis and Associated Microbial Dysbiosis in an IL-22-Dependent Manner. JCI Insight (2020) 5(1). doi: 10.1172/jci.insight.127551
105. Alkarkoushi RR, Hui Y, Tavakoli AS, Singh U, Nagarkatti P, Nagarkatti M, et al. Immune and MicroRNA Responses to Helicobacter Muridarum Infection and Indole-3-Carbinol During Colitis. World J Gastroenterol (2020) 26(32):4763–85. doi: 10.3748/wjg.v26.i32.4763
106. Hasan H, Ismail H, El-Orfali Y, Khawaja G. Therapeutic Benefits of Indole-3-Carbinol in Adjuvant-Induced Arthritis and Its Protective Effect Against Methotrexate Induced-Hepatic Toxicity. BMC Complement Altern Med (2018) 18(1):337. doi: 10.1186/s12906-018-2408-1
107. Nolan LS, Mihi B, Agrawal P, Gong Q, Rimer JM, Bidani SS, et al. Indole-3-Carbinol-Dependent Aryl Hydrocarbon Receptor Signaling Attenuates the Inflammatory Response in Experimental Necrotizing Enterocolitis. Immunohorizons (2021) 5(4):193–209. doi: 10.4049/immunohorizons.2100018
108. Zhang B, Bailey WM, Braun KJ, Gensel JC. Age Decreases Macrophage IL-10 Expression: Implications for Functional Recovery and Tissue Repair in Spinal Cord Injury. Exp Neurol (2015) 273:83–91. doi: 10.1016/j.expneurol.2015.08.001
109. Eghbalpour F, Aghaei M, Ebrahimi M, Tahsili MR, Golalipour M, Mohammadi S, et al. Effect of Indole-3-Carbinol on Transcriptional Profiling of Wound-Healing Genes in Macrophages of Systemic Lupus Erythematosus Patients: An RNA Sequencing Assay. Lupus (2020) 29(8):954–63. doi: 10.1177/0961203320929746
110. Mohammadi S, Saghaeian-Jazi M, Sedighi S, Memarian A. Sodium Valproate Modulates Immune Response by Alternative Activation of Monocyte-Derived Macrophages in Systemic Lupus Erythematosus. Clin Rheumatol (2018) 37(3):719–27. doi: 10.1007/s10067-017-3922-0
111. Chan E, McQueen F. Valproate-Induced Hyperammonaemia Superimposed Upon Severe Neuropsychiatric Lupus: A Case Report and Review of the Literature. Clin Rheumatol (2013) 32(3):403–7. doi: 10.1007/s10067-012-2150-x
112. Bleck TP, Smith MC. Possible Induction of Systemic Lupus Erythematosus by Valproate. Epilepsia (1990) 31(3):343–5. doi: 10.1111/j.1528-1157.1990.tb05386.x
113. Asconapé JJ, Manning KR, Lancman ME. Systemic Lupus Erythematosus Associated With Use of Valproate. Epilepsia (1994) 35(1):162–3. doi: 10.1111/j.1528-1157.1994.tb02927.x
114. Son M, Porat A, He M, Suurmond J, Santiago-Schwarz F, Andersson U, et al. C1q and HMGB1 Reciprocally Regulate Human Macrophage Polarization. Blood (2016) 128(18):2218–28. doi: 10.1182/blood-2016-05-719757
115. Musumeci D, Roviello GN, Montesarchio D. An Overview on HMGB1 Inhibitors as Potential Therapeutic Agents in HMGB1-Related Pathologies. Pharmacol Ther (2014) 141(3):347–57. doi: 10.1016/j.pharmthera.2013.11.001
116. Andersson U, Yang H, Harris H. Extracellular HMGB1 as a Therapeutic Target in Inflammatory Diseases. Expert Opin Ther Targets (2018) 22(3):263–77. doi: 10.1080/14728222.2018.1439924
117. Zhu L, Yang T, Li L, Sun L, Hou Y, Hu X, et al. TSC1 Controls Macrophage Polarization to Prevent Inflammatory Disease. Nat Commun (2014) 5(1). doi: 10.1038/ncomms5696
118. Katsiki N, Ferrannini E. Anti-Inflammatory Properties of Antidiabetic Drugs: A “Promised Land” in the COVID-19 Era? J Diabetes Complications (2020) 34(12):107723. doi: 10.1016/j.jdiacomp.2020.107723
119. Mohammadi S, Saghaeian-Jazi M, Sedighi S, Memarian A. Immunomodulation in Systemic Lupus Erythematosus: Induction of M2 Population in Monocyte-Derived Macrophages by Pioglitazone. Lupus (2017) 26(12):1318–27. doi: 10.1177/0961203317701842
120. Liu D, Zhang W. Pioglitazone Attenuates Lupus Nephritis Symptoms in Mice by Modulating Mir-21-5p/TIMP3 Axis: The Key Role of the Activation of Peroxisome Proliferator-Activated Receptor-γ. Inflammation (2021) 44(4):1416–25. doi: 10.1007/s10753-021-01426-x
121. Aparicio-Soto M, Sánchéz-Hidalgo M, Cárdeno A, Lucena JM, Gonzáléz-Escribano F, Castillo MJ, et al. The Phenolic Fraction of Extra Virgin Olive Oil Modulates the Activation and the Inflammatory Response of T Cells From Patients With Systemic Lupus Erythematosus and Healthy Donors. Mol Nutr Food Res (2017) 61(8). doi: 10.1002/mnfr.201601080
122. Aparicio-Soto M, Montserrat-de la Paz S, Sanchez-Hidalgo M, Cardeno A, Bermudez B, Muriana FJG, et al. Virgin Olive Oil and Its Phenol Fraction Modulate Monocyte/Macrophage Functionality: A Potential Therapeutic Strategy in the Treatment of Systemic Lupus Erythematosus. Br J Nutr (2018) 120(6):681–92. doi: 10.1017/S0007114518001976
123. Lee HJ, Kim MY, Park HS. Phosphorylation-Dependent Regulation of Notch1 Signaling: The Fulcrum of Notch1 Signaling. BMB Rep (2015) 48(8):431–7. doi: 10.5483/BMBRep.2015.48.8.107
124. Rauen T, Grammatikos AP, Hedrich CM, Floege J, Tenbrock K, Ohl K, et al. Camp-Responsive Element Modulator α (Cremα) Contributes to Decreased Notch-1 Expression in T Cells From Patients With Active Systemic Lupus Erythematosus (SLE). J Biol Chem (2012) 287(51):42525–32. doi: 10.1074/jbc.M112.425371
125. Sodsai P, Hirankarn N, Avihingsanon Y, Palaga T. Defects in Notch1 Upregulation Upon Activation of T Cells From Patients With Systemic Lupus Erythematosus Are Related to Lupus Disease Activity. Lupus (2008) 17(7):645–53. doi: 10.1177/0961203308089406
126. Zhang W, Xu W, Xiong S. Blockade of Notch1 Signaling Alleviates Murine Lupus via Blunting Macrophage Activation and M2b Polarization. J Immunol (2010) 184(11):6465–78. doi: 10.4049/jimmunol.0904016
127. Zhou P, Li Q, Su S, Dong W, Zong S, Ma Q, et al. Interleukin 37 Suppresses M1 Macrophage Polarization Through Inhibition of the Notch1 and Nuclear Factor Kappa B Pathways. Front Cell Dev Biol (2020) 8:56. doi: 10.3389/fcell.2020.00056
128. Horuluoglu B, Bayik D, Kayraklioglu N, Goguet E, Kaplan MJ, Klinman DM. PAM3 Supports the Generation of M2-Like Macrophages From Lupus Patient Monocytes and Improves Disease Outcome in Murine Lupus. J Autoimmun (2019) 99:24–32. doi: 10.1016/j.jaut.2019.01.004
129. Liu Y, Luo S, Zhan Y, Wang J, Zhao R, Li Y, et al. Increased Expression of PPAR-γ Modulates Monocytes Into a M2-Like Phenotype in SLE Patients: An Implicative Protective Mechanism and Potential Therapeutic Strategy of Systemic Lupus Erythematosus. Front Immunol (2020) 11:579372. doi: 10.3389/fimmu.2020.579372
130. Hou LF, He SJ, Li X, Wan CP, Yang Y, Zhang XH, et al. SM934 Treated Lupus-Prone NZB X NZW F1 Mice by Enhancing Macrophage Interleukin-10 Production and Suppressing Pathogenic T Cell Development. PloS One (2012) 7(2):e32424. doi: 10.1371/journal.pone.0032424
131. Xu N, Li J, Gao Y, Zhou N, Ma Q, Wu M, et al. Apoptotic Cell-Mimicking Gold Nanocages Loaded With LXR Agonist for Attenuating the Progression of Murine Systemic Lupus Erythematosus. Biomaterials (2019) 197:380–92. doi: 10.1016/j.biomaterials.2019.01.034
132. A-Gonzalez N, Bensinger SJ, Hong C, Beceiro S, Bradley MN, Zelcer N, et al. Apoptotic Cells Promote Their Own Clearance and Immune Tolerance Through Activation of the Nuclear Receptor LXR. Immunity (2009) 31(2):245–58. doi: 10.1016/j.immuni.2009.06.018
133. Han S, Zhuang H, Shumyak S, Wu J, Xie C, Li H, et al. Liver X Receptor Agonist Therapy Prevents Diffuse Alveolar Hemorrhage in Murine Lupus by Repolarizing Macrophages. Front Immunol (2018) 9:135. doi: 10.3389/fimmu.2018.00135
134. Bleyzac N, Goutelle S, Bourguignon L, Tod M. Azithromycin for COVID-19: More Than Just an Antimicrobial? Clin Drug Investig (2020) 40(8):683–6. doi: 10.1007/s40261-020-00933-3
135. Wang J, Xie L, Wang S, Lin J, Liang J, Xu J. Azithromycin Promotes Alternatively Activated Macrophage Phenotype in Systematic Lupus Erythematosus via PI3K/Akt Signaling Pathway. Cell Death Dis (2018) 9(11):1080. doi: 10.1038/s41419-018-1097-5
Keywords: systemic lupus erythematosus, macrophage activation syndrome (MAS), M1 macrophage polarization, M2 macrophage polarization, macrophage plasticity and polarization
Citation: Ahamada MM, Jia Y and Wu X (2021) Macrophage Polarization and Plasticity in Systemic Lupus Erythematosus. Front. Immunol. 12:734008. doi: 10.3389/fimmu.2021.734008
Received: 30 June 2021; Accepted: 08 November 2021;
Published: 20 December 2021.
Edited by:
Michele Marie Kosiewicz, University of Louisville, United StatesReviewed by:
Carla Marie Cuda, Northwestern University, United StatesCopyright © 2021 Ahamada, Jia and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaochuan Wu, eGlhb2NodWFud3VAY3N1LmVkdS5jbg==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.