 Eveline M. Delemarre1,2†
Eveline M. Delemarre1,2† Laura van Hoorn1,3†
Laura van Hoorn1,3† Aik W. J. Bossink4
Aik W. J. Bossink4 Julia Drylewicz1
Julia Drylewicz1 Simone A. Joosten5
Simone A. Joosten5 Tom H. M. Ottenhoff5Onno W. Akkerman6
Tom H. M. Ottenhoff5Onno W. Akkerman6 Delia Goletti7
Delia Goletti7 Elisa Petruccioli7
Elisa Petruccioli7 Assunta Navarra7Brigitte T. A. van den Broek1Sanne P. A. Paardekooper1Ineke van Haeften8
Assunta Navarra7Brigitte T. A. van den Broek1Sanne P. A. Paardekooper1Ineke van Haeften8 Leo Koenderman1,3Jan-Willem J. Lammers3Steven F. T. Thijsen9Regina W. Hofland3,4‡
Leo Koenderman1,3Jan-Willem J. Lammers3Steven F. T. Thijsen9Regina W. Hofland3,4‡ Stefan Nierkens1,2*‡
Stefan Nierkens1,2*‡- 1Center for Translational Immunology (CTI), University Medical Center Utrecht, Utrecht, Netherlands
- 2Platform Immune Monitoring (PIM), University Medical Center Utrecht, Utrecht, Netherlands
- 3Department of Respiratory Medicine and Tuberculosis, University Medical Center Utrecht, Utrecht, Netherlands
- 4Department of Respiratory Medicine and Tuberculosis, Diakonessenhuis, Utrecht, Netherlands
- 5Department of Infectious Diseases, Leiden University Medical Center, Leiden, Netherlands
- 6Department of Respiratory Medicine and Tuberculosis, University Medical Center Groningen, Groningen, Netherlands
- 7Translational Research Unit, Department of Epidemiology and Preclinical Research, National Institute for Infectious Diseases-IRCCS L. Spallanzani, Rome, Italy
- 8Department of Tuberculosis, Municipal Public Health Service, Utrecht, Netherlands
- 9Department of Medical Microbiology and Immunology, Diakonessenhuis, Utrecht, Netherlands
Introduction: There is an urgent medical need to differentiate active tuberculosis (ATB) from latent tuberculosis infection (LTBI) and prevent undertreatment and overtreatment. The aim of this study was to identify biomarker profiles that may support the differentiation between ATB and LTBI and to validate these signatures.
Materials and Methods: The discovery cohort included adult individuals classified in four groups: ATB (n = 20), LTBI without prophylaxis (untreated LTBI; n = 20), LTBI after completion of prophylaxis (treated LTBI; n = 20), and healthy controls (HC; n = 20). Their sera were analyzed for 40 cytokines/chemokines and activity of adenosine deaminase (ADA) isozymes. A prediction model was designed to differentiate ATB from untreated LTBI using sparse partial least squares (sPLS) and logistic regression analyses. Serum samples of two independent cohorts (national and international) were used for validation.
Results: sPLS regression analyses identified C-C motif chemokine ligand 1 (CCL1), C-reactive protein (CRP), C-X-C motif chemokine ligand 10 (CXCL10), and vascular endothelial growth factor (VEGF) as the most discriminating biomarkers. These markers and ADA(2) activity were significantly increased in ATB compared to untreated LTBI (p ≤ 0.007). Combining CCL1, CXCL10, VEGF, and ADA2 activity yielded a sensitivity and specificity of 95% and 90%, respectively, in differentiating ATB from untreated LTBI. These findings were confirmed in the validation cohort including remotely acquired untreated LTBI participants.
Conclusion: The biomarker signature of CCL1, CXCL10, VEGF, and ADA2 activity provides a promising tool for differentiating patients with ATB from non-treated LTBI individuals.
Introduction
Tuberculosis (TB) is an infectious disease caused by a species of the Mycobacterium tuberculosis (Mtb) complex, most commonly the bacillus Mtb (1). TB represents a major global health problem with an estimated incidence of 10 million and a mortality of 1.4 million persons in 2019 (2). Patients with TB can infect 10–15 close contacts within 1 year, and approximately 45% of infected patients will die without proper treatment (2). It is estimated that one-fourth of the world’s population suffers from a latent TB infection (LTBI) (3–5) that (in the absence of a gold standard) is defined by the World Health Organization as “a state of persistent immune response to Mtb without clinical evidence of active TB disease” (2). This definition includes a continuum of stages ranging from clearance of the pathogen to dormant, but live tuberculous mycobacteria (6, 7). The lack of evidence for the presence of live tuberculous mycobacteria in LTBI generates clinical unanswered questions in general practice and research. The lifetime risk of reactivation and progression from LTBI to active TB (ATB) is 5%–10% and mostly occurs in the first years following infection (8). In low-burden countries, reactivation accounts for approximately 80% of new ATB cases, which could be averted by timely diagnosis and preventive treatment with an efficacy of 60%–90% (8). The tuberculin skin test (TST) and interferon gamma release assay (IGRA) are currently used for LTBI diagnosis, although they do not allow discriminating ATB from LTBI and predict with low accuracy the risk of progression to ATB (2, 9–13).
The clinical presentation of ATB is diverse; definite diagnosis can be challenging because of limited sensitivity of the nucleic acid amplification test and Mtb cultures, especially in extrapulmonary forms of ATB. The GeneXpert® MTB/RIF sputum test has a higher sensitivity compared to that of Mtb cultures (14, 15). The newer Xpert Ultra® test increased the sensitivity for paucibacillary disease. However, both tests are sputum based, which causes problems for patients who are not able to produce sufficient sputum production, which is more often seen in children and patients with extrapulmonary disease (16). As a result, discrimination between ATB and LTBI can be difficult in clinical practice, leading to undertreatment of patients with ATB and overtreatment of patients with LTBI (4, 6). Despite optimal diagnostic facilities, microbiological confirmation is lacking in about 30%–35% of the patients with ATB in the Netherlands (17). Therefore, the aim of this study was to identify novel biomarker profiles in serum to differentiate between ATB and LTBI.
Infection with Mtb causes a cascade of immune responses, including production of cytokines/chemokines and generation of enzyme activities important to limit bacterial growth, regulate inflammation, and seal off the infection by creating a granuloma (18). These mechanisms may result in systemic fingerprints indicating active Mtb replication and therefore potentially useful in distinguishing ATB from LTBI. Activity measurement of the enzyme adenosine deaminase (ADA) and its two isoforms (ADA1 and ADA2) in a variety of biological fluids has been suggested to aid in the diagnosis of ATB; however, ADA activity in serum has never been used to distinguish ATB from LTBI (19–23). Several studies exploring the use of inflammatory serum cytokines involved in Mtb infection have been performed (24–29), with inconsistent conclusions for cytokine production in Mtb-specific stimulated CD4+ and CD8+ T cells (30–45). Most of these studies showed that the discriminative value of a single marker is limited. Here, we report that a specific combination of serum markers has promising diagnostic potential to distinguish ATB from untreated LTBI.
Materials and Methods
Study Participants
In the discovery cohort, 80 HIV-negative adults were enrolled in this cross-sectional study from December 2015 until January 2017 through the Public Health Service of Utrecht and the Diakonessenhuis (Utrecht, Netherlands). Individuals were classified into four groups: ATB (n = 20) with pulmonary and/or extrapulmonary disease, latent TB without prophylactic treatment (untreated LTBI; n = 20), latent TB individuals after completion of prophylactic treatment (treated LTBI; n = 20), and healthy controls (HCs; n = 20). ATB patients were microbiologically diagnosed according to the American Thoracic Society guidelines and were included before or within 8 days after start of treatment (46). Participants were included in the LTBI group in case of a positive TST and/or IGRA in the absence of clinical evidence of TB and a likely recent infection in the last 2 years, as determined by documented contact with an infectious case and absence of documented TB contact previously. Prophylactic treatment entailed 3 months of rifampicin and isoniazid or 6 months of isoniazid. LTBI individuals who completed prophylactic therapy were indicated as “LTBI-cured,” here referred to as treated LTBI. HCs were included as a reference group and consisted of healthcare workers with no history of or contact with TB who were screened annually for TB and they all had a recent negative IGRA. All enrolled participants signed a written informed consent and completed a questionnaire regarding their medical history, history of TB, smoking status, and demographics (Table 1). This study was approved by the Medical Research Ethics Committees-United (NL53628.100.15) and the Board of Directors of the Public Health and Diakonessenhuis, Utrecht, Netherlands.
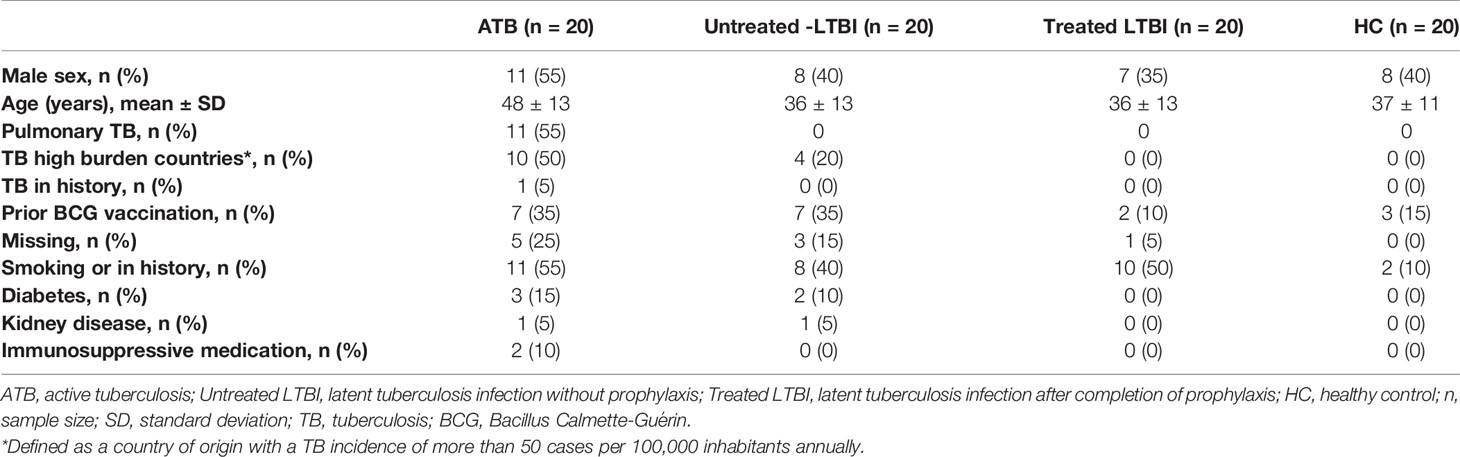
Table 1 Baseline characteristics of the discovery cohort.
For the validation, two independent cohorts were included. One cohort, collected by the Leiden University Medical Center (LUMC, Leiden, Netherlands), consisted of 12 ATB patients (treated in a specialized TB clinic, Groningen, Netherlands) and 20 untreated LTBI individuals who all had a positive IGRA. The untreated LTBI individuals within this cohort have been previously described (47). A second cohort (National Institute of Infectious Diseases, Rome, Italy) included 31 ATB patients and 20 untreated LTBI individuals with positive IGRA who were sharing their households with ATB patients no more than 3 months before inclusion in the study. These Italian untreated LTBI individuals are different compared to the Dutch untreated LTBI individuals because the Dutch individuals were the result of a contact investigation, employment verification, or periodic screening and probably had a recent infection in the last 2 years. Therefore, we will refer to the Italian untreated LTBI individuals who all shared their households with ATB patients before inclusion in the study as HH-LTBI. This cohort has been previously described (35). All participants signed a written informed consent before participation. For the Leiden cohort, the study was approved by the Medical Ethics Committee of LUMC (METC project number P07.048). The Italian study was approved by the Ethical Committee of the L. Spallanzani National Institute of Infectious diseases (INMI approval numbers 02/2007 and 72/2015). Inclusion strategies for both ATB and untreated LTBI groups of the validation cohorts were similar as those for the discovery cohort.
Measurement of Serum Mediators
Levels of 40 cytokines/chemokines were measured in serum samples of the discovery cohort using an in-house developed and validated (ISO9001) multiplex immunoassay based on xMAP technology (Luminex Corporation, Austin, USA) (Supplementary Table S1). The assay was performed as previously described (48, 49). In short, a specific heterophilic immunoglobulins were preabsorbed from all samples with heteroblock (Omega Biologicals, Bozeman, MT, USA). Next, samples were incubated with antibody-conjugated MagPlex microspheres for 1 h at room temperature with continuous shaking, followed by 1-h incubation with biotinylated antibodies and 10-min incubation with phycoerythrin-conjugated streptavidin diluted in high-performance ELISA buffer (HPE, Sanquin, Netherlands). Acquisition was performed with the Bio-Rad FlexMAP3D (Bio-Rad Laboratories, Hercules, CA, USA) in combination with xPONENT software version 4.2 (Luminex). Data were analyzed by five-parametric curve fitting using Bio-Plex Manager software, version 6.1.1 (Bio-Rad). Serum samples were diluted to measure S100A8, SAA1, Angiotensin Converting Enzyme (ACE), and C-reactive protein (CRP) in detectable range: 1:10, 1:100, 1:100, 1:1,000, respectively. Imputation of the lower limit of quantification (LLOQ) divided by 2 was performed for samples with concentrations under the detection limit. Samples with concentrations above the detection limit, the upper limit of quantification (ULOQ) multiplied by 2 was imputed, except for the diluted cytokines. For the diluted cytokines, the highest result of the extrapolated values was used to impute for results above the detection limit.
Total ADA and ADA2 activities were determined using an ADA assay kit (Diazyme Laboratories, Poway, CA, USA) according to manufacturer’s instructions with minor modifications. In short, to measure ADA2 activity, 0.1 μM erythro-9 (2-hydroxy-3-nonyl) adenine (EHNA) was added to inhibit ADA1 activity (50). Activity measurements were performed using the SpectraMax M2e Plate Reader (Molecular Devices, Sunnyvale, CA, USA). The activity of ADA1 was calculated by subtracting the ADA2 activity from the total ADA activity. ADA activity was expressed in U/L.
Statistical Analysis
Baseline characteristics were reported as mean and standard deviation (SD) for continuous variables and as absolute numbers and percentages for binary variables. Comparisons between patient groups in the discovery cohort were performed using the Kruskal–Wallis test for continuous variables and Fisher–Freeman–Halton exact test for binary variables. Within the validation cohorts, comparisons were made between the ATB and LTBI groups using the Mann–Whitney U test for continuous variables and Fisher’s exact test for binary variables. Sparse partial least squares regression (sPLS) analysis was applied on the dataset including the 40 measured serum mediators obtained for the discovery cohort to identify serum markers that could be used to differentiate ATB from untreated LTBI (R package MixOmics) (51). The hereby selected markers, total ADA, ADA1, and ADA2 activities were compared between the four patient groups of the discovery cohort (ATB, untreated LTBI, treated LTBI, and HC) and the two groups in the validation cohorts (ATB vs. untreated LTBI/HH-LTBI) using Mann–Whitney tests. Logistic regression analyses including backward stepwise selection were used to acquire a model able to classify individuals into ATB from untreated LTBI in the discovery cohort (R package stats) (52). The model (i.e., formula based on the selected biomarkers) with the lowest Akaike information criterion (AIC) was kept for further analyses. All variables measured by Luminex were logarithmically transformed (log10). Receiver operating characteristic (ROC) analyses and area under the curve (AUC) were obtained for all markers separately and the final models. The two biomarker signatures were applied on the datasets obtained for the two independent validation cohorts using the logistic regression coefficients and optimal cutoff found for the discovery cohort (R package InformationValue) (53). The optimal cutoff is obtained when the most participants are correctly identified, thereby achieving the highest sensitivity and specificity. For all three patient cohorts, sensitivity and specificity in discriminating ATB from untreated LTBI were calculated using the biomarker combinations. Results for the 40 measured cytokines/chemokines per subgroup of the discovery cohort were reported as median and interquartile range (IQR), and raw data are available (Supplementary Data 1). SPSS software (version 22.0) and R Project Software (version 3.2.0) were used for statistical analysis. p-values <0.05 were considered statistically significant.
Results
Study Participants
Within the discovery cohort, participants in the ATB group were significantly older compared to the untreated LTBI, treated LTBI, and HC groups (p < 0.005; Table 1) and more often originating from a TB-endemic country (defined as a country of origin with a TB incidence of more than 50 cases per 100,000 inhabitants annually) compared to individuals in the treated LTBI and HC groups (p = 0.0001). Smoking was more frequent in the ATB and treated LTBI groups compared to HCs (p = 0.006 and p = 0.014, respectively). Other baseline characteristics showed no significant differences between the four groups (Table 1). In the Leiden and Italy cohorts, age did not significantly differ between the ATB and untreated LTBI/HH-LTBI individuals (Table 2). Within the Italian cohort, more males were included in the ATB group compared to the HH-LTBI group (p = 0.046) and more ATB patients originated from a TB-endemic country (p = 0.004).
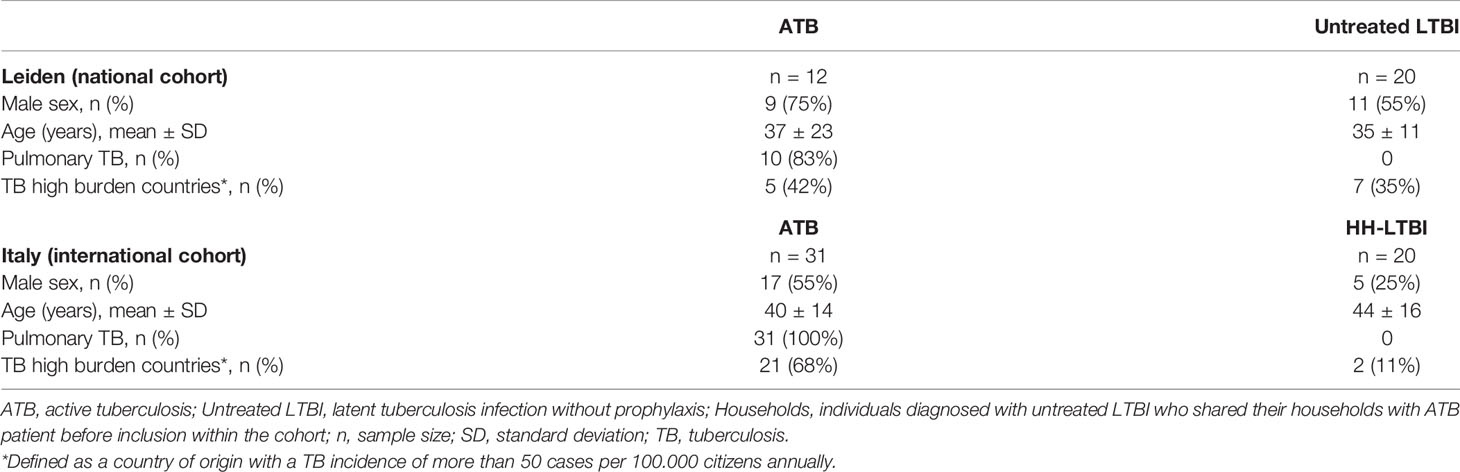
Table 2 Baseline characteristics of the validation cohorts.
Identification of Serum Biomarkers for Distinguishing ATB From Untreated LTBI
To identify biomarkers that may aid in differentiating ATB from untreated LTBI, 40 cytokines/chemokines and ADA activity including isoforms were measured in serum samples of participants included in the discovery cohort. sPLS analyses and Mann–Whitney tests demonstrated that C-C motif chemokine ligand 1 (CCL1), CRP, C-X-C motif chemokine ligand 10 (CXCL10), and vascular endothelial growth factor (VEGF) were the strongest differentiating markers between ATB and untreated LTBI individuals (Figures 1, 2; p = 0.0001, p = 0.007, p = 0.0001, and p = 0.0001, respectively). Moreover, these markers were significantly increased in the sera of ATB patients compared to treated LTBI individuals and HCs (Figure 2). Within the untreated LTBI, for each marker, three amples seem to have higher levels of each protein compared to the other samples. These samples do not belong to the same participants. Comparing cytokine expression between pulmonary and extrapulmonary TB in a subgroup analysis showed no differences (Supplementary Figure S1). Results for all measured cytokines/chemokines are shown in Supplementary Table S1.
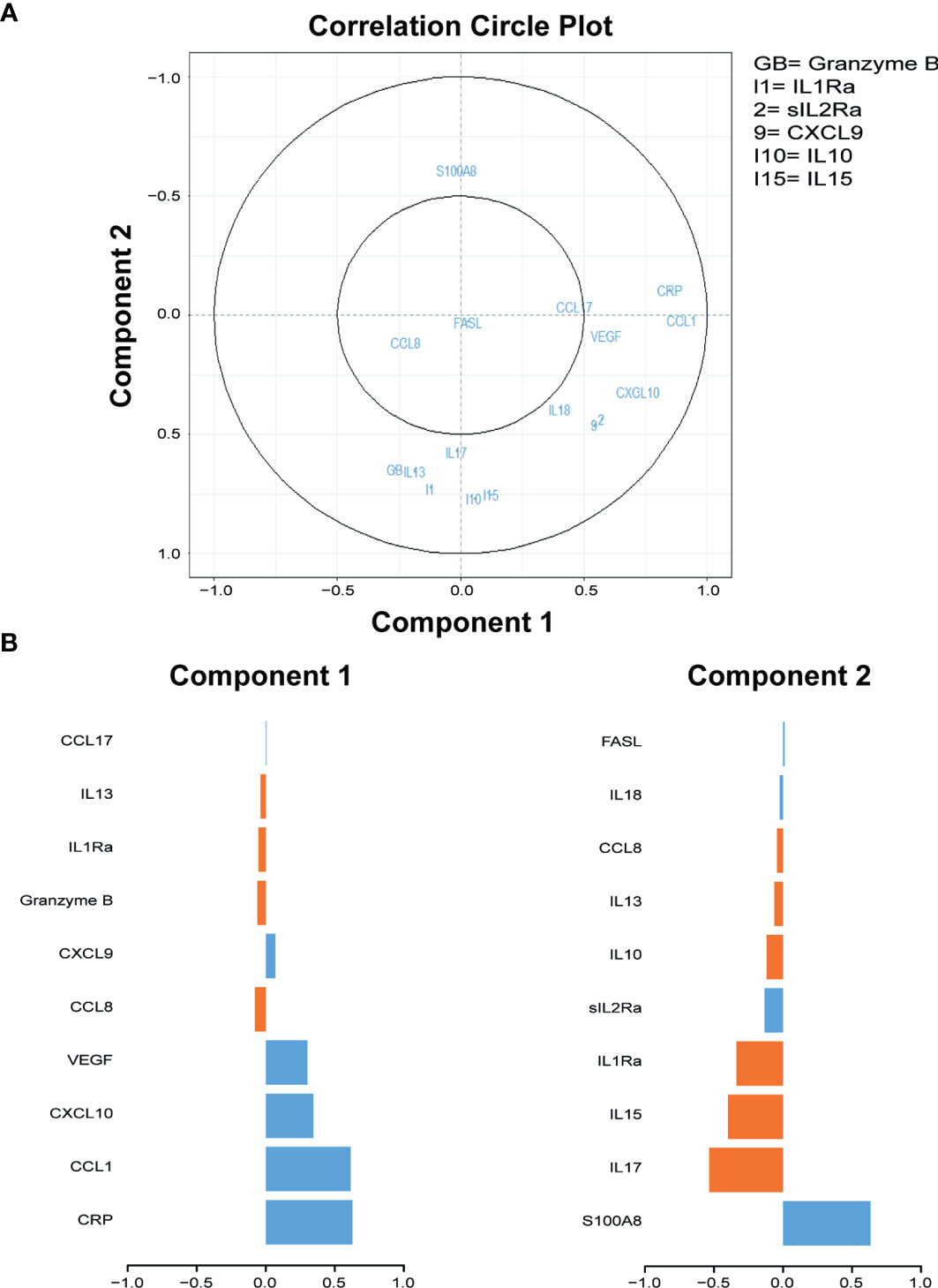
Figure 1 Sparse partial least squares regression analyses of 40 cytokines measured in serum samples of individuals diagnosed with ATB and untreated LTBI. (A) Correlation circle plots representing the correlations between the cytokines and the first two components of the sPLS. A correlation circle is a two-dimensional plot helping the interpretation of the correlations between the initial variables (i.e., the cytokines) and the new variables (i.e., the components) that are combinations of the first ones: the closer a variable is to an axis and to the circle, the higher the correlation with the corresponding component will be. It is also a way to plot the correlations between the cytokines: if two cytokines are in opposite quarters, they are negatively correlated; if two cytokines are perpendicular, then their correlation is close to 0; if two variables are close, their correlation is close to 1. Top 10 cytokines of the first two components are shown in the graph. GB, Granzyme B; I1, Il1Ra; 2, sIL2Ra; 9, CXCL9; I10, IL-10; I15, IL-15. (B) The contribution (i.e., the correlation) of the top 10 cytokines for components 1 and 2. The blue bars represent ATB, and the orange bars represent untreated LTBI. ATB, active tuberculosis; untreated LTBI, latent tuberculosis infection without prophylaxis; sPLS, sparse partial least squares.
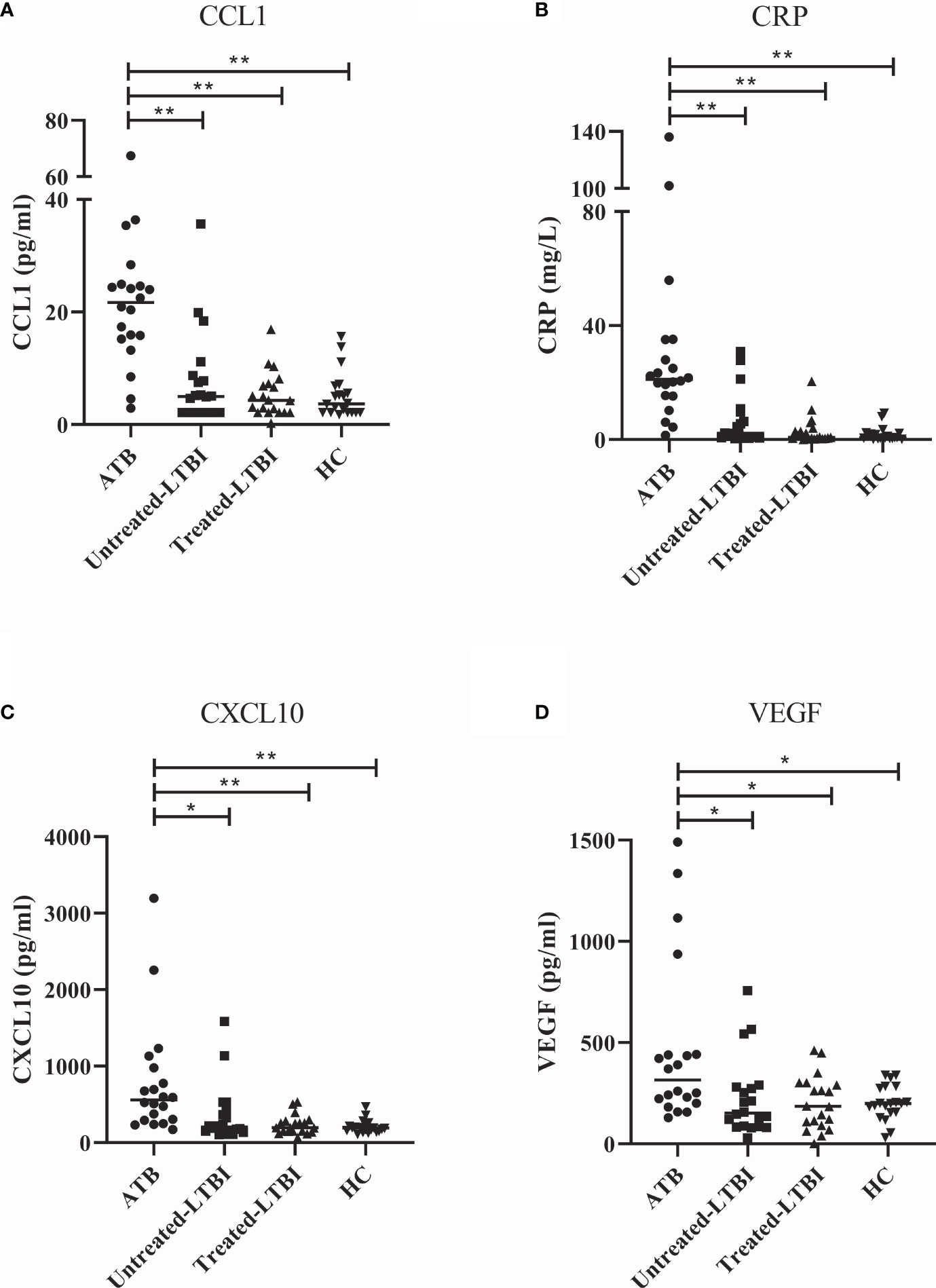
Figure 2 Serum levels of CCL1 (A), CRP (B), CXCL10 (C), and VEGF (D) in individuals with ATB, untreated LTBI, and treated LTBI and HCs. All groups were compared for each marker using Mann–Whitney U tests. In panels (A–D), the filled dots represent participants with ATB, the filled squares represent the participants with untreated LTBI, the filled triangles represent the participants with treated LTBI, and the filled inversed triangles represent the HCs. The horizontal line in each study group represents the median concentration. ATB, active tuberculosis; CCL1, C-C motif chemokine ligand 1; CRP, C-reactive protein; CXCL10, C-X-C motif chemokine ligand 10; HC, healthy controls; untreated LTBI, latent tuberculosis infection without prophylaxis; treated LTBI, latent tuberculosis infection after completion of prophylaxis; VEGF, vascular endothelial growth factor. * 0.001 < p < 0.05, ** p < 0.001.
Furthermore, ADA activity was significantly increased in the ATB group compared to untreated LTBI, treated LTBI, and HC groups (p ≤ 0.001; Figure 3). The isoform ADA2 was responsible for this increase (ADA1 showed no significant between-group differences).
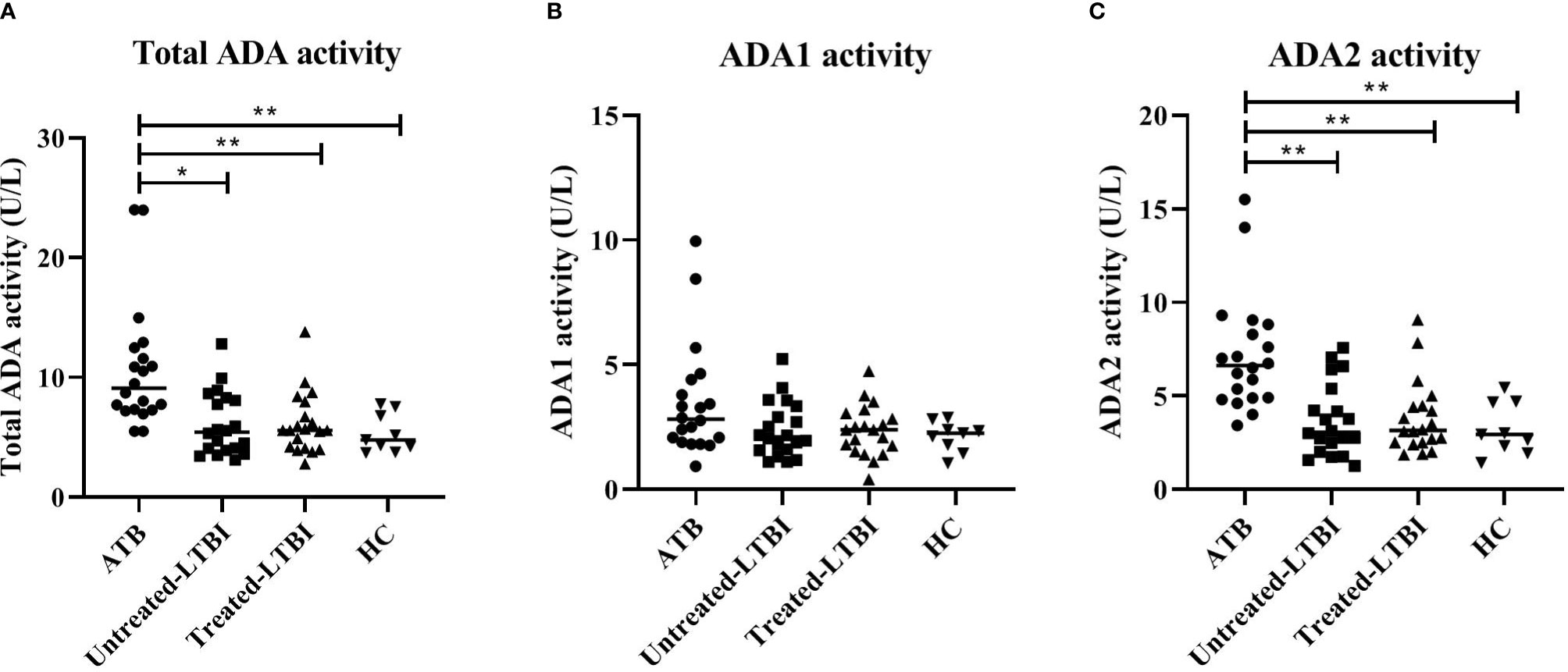
Figure 3 ADA activity levels in serum of individuals with ATB, untreated LTBI, and treated LTBI and HCs. Serum levels for total ADA (A), ADA1 (B), and ADA2 (C) activity levels are shown. All groups were compared for each marker using Mann–Whitney U tests. In panels (A–C), the filled dots represent participants with ATB, the filled squares represent the participants with untreated LTBI, the filled triangles represent the participants with treated-LTBI, and the filled inversed triangles represent the HCs. The horizontal line in each study group represents the median concentration. ADA, adenosine deaminase; ADA1, adenosine deaminase-1; ADA2, adenosine deaminase-2; ATB, active tuberculosis; LTBI, latent tuberculosis infection. * 0.001 < p < 0.05, ** p < 0.001.
To investigate whether CCL1, CXCL10, CRP, VEGF, and ADA(2) activity are potential biomarkers, these markers were measured in two independent validation cohorts. For one patient within the “Italian” ATB group, ADA activity measurement was not performed. In both cohorts, CCL1, CXCL10, CRP, and VEGF levels were significantly increased in the sera of ATB patients compared to untreated LTBI or HH-LTBI individuals (Figure 4A). In addition, total ADA activity was also significantly increased in the ATB patients due to increased activity of ADA2 (Figure 4B). Both validation cohorts confirmed the different expression patterns of CCL1, CRP, CXCL10, VEGF, and ADA activity in sera of ATB patients compared to untreated LTBI/HH-LTBI individuals. Hence, we identified five potential biomarkers that may help discriminate between ATB and untreated LTBI individuals.
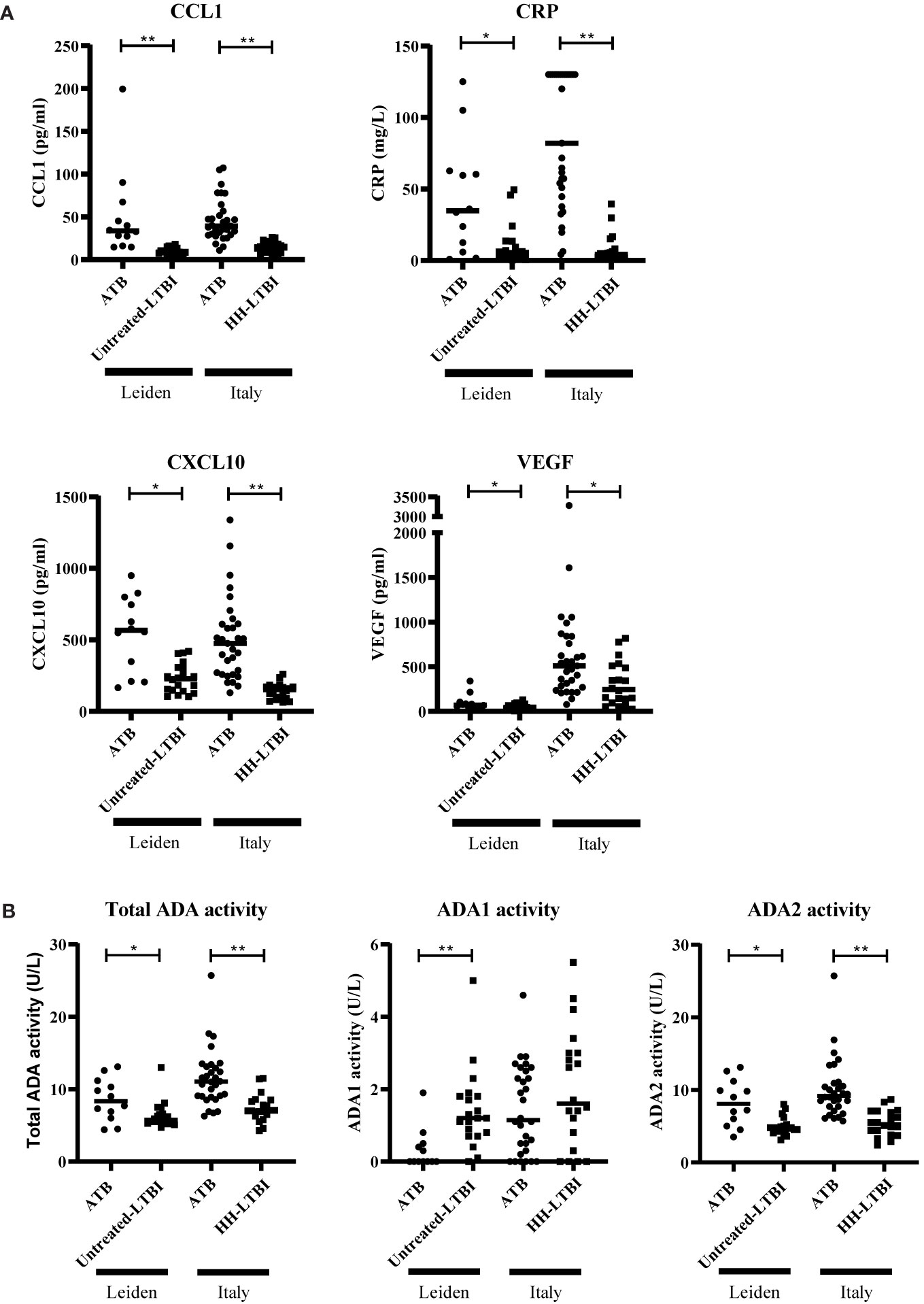
Figure 4 Serum levels of CCL1, CRP, CXCL10, VEGF, and ADA activity differently expressed in patients with ATB compared to untreated LTBI individuals in two independent cohorts. Serum levels of CCL1, CRP, CXCL10, VEGF (A), and ADA activity (B) are shown. The two patient groups per cohort were compared for each marker using Mann–Whitney U tests. Filled dots represent patients with ATB, and filled squares represent individuals with untreated LTBI or households of ATB patients diagnosed with untreated LTBI/untreated HH-LTBI. Median concentration is depicted with horizontal line in each group. ADA, adenosine deaminase; ATB, active tuberculosis; CCL1, C-C motif chemokine ligand 1; CRP, C-reactive protein; CXCL10, C-X-C motif chemokine ligand 10; LTBI, latent tuberculosis infection; VEGF, vascular endothelial growth factor. * 0.001 < p < 0.05, ** p < 0.001.
Prediction Model to Discriminate ATB From Untreated LTBI
Subsequently, we explored whether these biomarkers could be used for differentiating between ATB and untreated LTBI individuals. Logistic regression analyses with the discovery cohort dataset showed that the combination of CCL1, CXCL10, VEGF, and ADA2 (without CRP) yielded the lowest AIC (29.78) with area under the curve (AUC = 0.9525). The following formula was produced: with optimal cutoff value set on 0.50998. Adding the values obtained for each marker per participant within the formula obtained a new value. If this value was greater than the optimal cutoff, the participant was classified as ATB. This model with optimal cutoff value yielded a sensitivity of 95% and specificity of 90% in distinguishing ATB patients from untreated LTBI individuals in the discovery cohort (Table 3, Supplementary Figure S2). For total ADA activity, the same marker combination gave the lowest AIC (AUC = 0.9475; Table 3). With the optimal cutoff set on 0.44997, this biomarker combination yielded a sensitivity of 100% and specificity of 90% in the discovery cohort. Patients with extrapulmonary disease were correctly identified as ATB by both biomarker signatures (data not shown). Together, these data indicate that a combination of markers may be a promising tool to distinguish ATB from untreated LTBI.
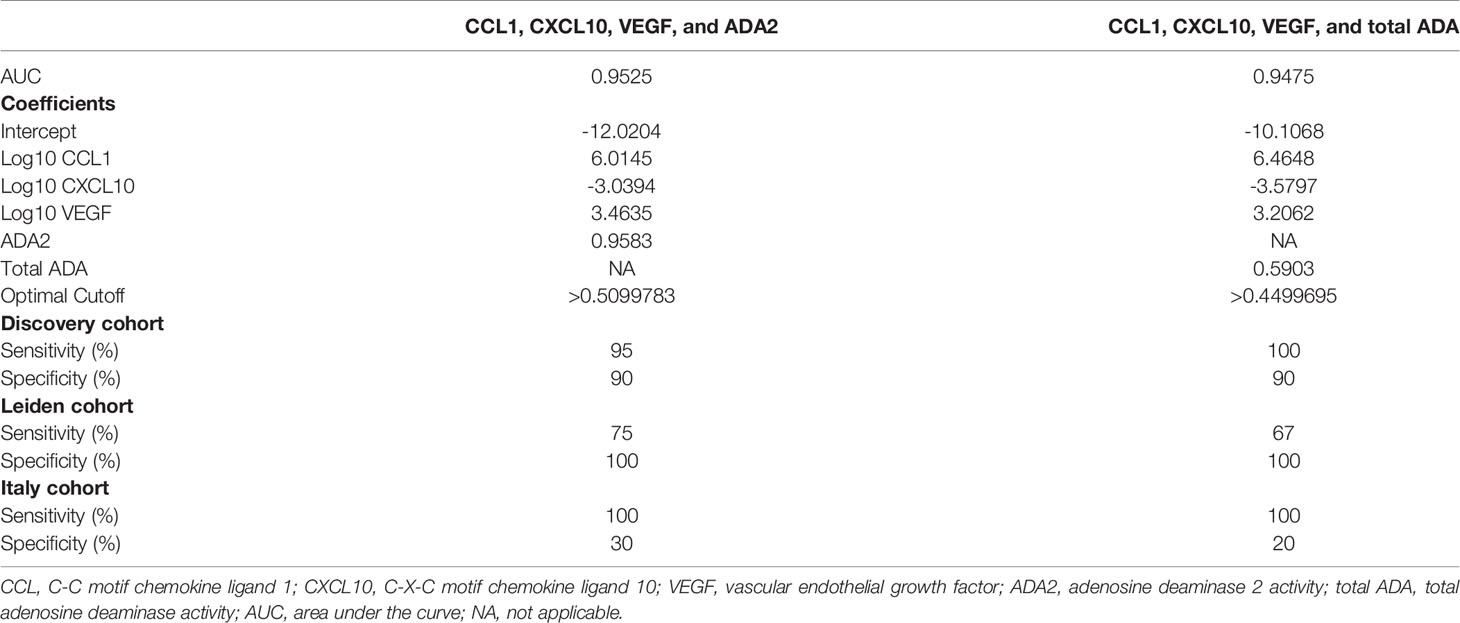
Table 3 Evaluation biomarker signature (CCL1, CXCL10, VEGF, and ADA2 or total ADA) in distinguishing ATB from untreated LTBI.
Validation of Serum Biomarker Profiles for the Identification of ATB and Untreated LTBI
To confirm the usefulness of these biomarker signatures in discriminating ATB patients from untreated LTBI individuals, the signatures and corresponding optimal cutoff values were first applied on the dataset obtained with the samples originating from the Leiden cohort. In this cohort, the included participants in the untreated LTBI group were similar to the untreated LTBI group of the discovery cohort. Using the biomarker signature including ADA2 activity, the accuracy in distinguishing ATB from untreated LTBI was 90.6%, with sensitivity of 75% and specificity of 100%. The biomarker profile with total ADA activity yielded an accuracy of 87.5%, with sensitivity of 67% and specificity of 100%. Results are shown in Table 3 and Supplementary Table S3.
In the Italian cohort, where untreated HH-LTBI individuals were sharing their households with ATB patients, we obtained an accuracy of 72% for the biomarker signature with ADA2 activity, a sensitivity of 100%, and specificity of 30%. The profile including total ADA activity yielded the same sensitivity; however, a specificity of 20% (Table 3, Supplementary Table S3).
Results obtained for both the discovery cohort and validation cohort (Leiden) indicated that the combinations of CCL1, CXCL10, VEGF, and ADA(2) activity could be promising tools for discriminating between ATB patients and untreated LTBI individuals. However, the model lacks sufficient specificity to discriminate ATB patients and untreated HH-LTBI individuals sharing a household with an ATB patient before inclusion in the study.
Discussion
With currently available routine sputum-based tests, it may be difficult to differentiate ATB from LTBI. In daily practice, this means that ATB patients might be undertreated, possibly leading to severe morbidity and continued transmission due to untreated disease, whereas LTBI individuals might be overtreated with an unnecessary drug exposure and toxicity (2, 8). Therefore, the aim of this study was to identify novel biomarker profiles to differentiate ATB from LTBI using serum. Such a tool may help clinicians in deciding whether patients should receive full medical treatment in ATB patients without microbiological confirmation or prophylactic treatment in case of LTBI.
To our knowledge, this is the first study evaluating the diagnostic accuracy of activities of serum ADA and its isozymes in combination with other serum biomarkers to differentiate ATB from LTBI. In ATB patients, CCL1, VEGF, CRP, CXCL10, and ADA(2) activity were significantly elevated compared to untreated LTBI individuals. Increased concentrations of CCL1, CRP, VEGF, and CXCL10 in ATB compared to LTBI are in line with previous reports (30, 31, 38–41, 54) and may reflect the activity of infected macrophages, innate immune cells such as natural killer (NK) cells and innate lymphoid cells (ILCs), as well as activated T lymphocytes. CCL1, also known as I-309, is a glycoprotein secreted by activated T lymphocytes and stimulates chemotaxis of monocytes (55). VEGF and CXCL10 are produced by activated macrophages and antigen-presenting cells and regulate cell growth and chemotaxis and may be the driving forces for stimulated angiogenesis as observed in ATB lesions (56–58). CXCL10 may also limit Mtb replication, as recently observed (59, 60). CRP is synthesized in the liver and is secreted following interleukin (IL)-6 stimulation by macrophages and T lymphocytes (61). CRP levels have been shown to depend on the virulence of the Mtb strain and the location of the infection, for example, pulmonary TB and miliary TB show higher levels of CRP (62). Activation of monocytes during TB infection is further emphasized by increased ADA activity and in particular of ADA2. ADA is an enzyme that protects cells from apoptosis by eliminating intracellular toxic derivatives of both adenosine and deoxyadenosine (63). The isoform ADA2 is exclusively produced by monocytes and macrophages when exposed to, or infected by, intracellular bacterial pathogens, such as Mtb (64). Secretion of ADA2 promotes CD4+ T-cell proliferation and differentiation of monocytes into macrophages followed by proliferation (63). The diagnostic utility of total ADA(2) activity has been studied in various TB (sub)populations, and increased total ADA activity was shown in serum from ATB patients when compared with HCs or cured TB subjects combined with positive TST (20, 65, 66). However, in these studies, no comparison was made between ATB patients and LTBI individuals.
Validating the combination of ADA(2) activity with CCL1, CXCL10, and VEGF in two independent cohorts led to confirmatory results, with some differences observed as well. The obtained sensitivity in identifying ATB was high in both validation cohorts. However, the specificity obtained within the Leiden cohort was high, but in the Italian cohort, it was low. As discussed before, within this Italian cohort, untreated HH-LTBI participants were individuals sharing their households with patients diagnosed with ATB not more than 3 months before inclusion in the study. We expect that close proximity with an ATB patient increases the exposure to Mtb. Therefore, we hypothesize that the Italian untreated LTBI individuals may have an active Mtb replication being in an intermediate stage between ATB and untreated LTBI. This difference may have led to incorrect identification of the Italian untreated LTBI patients using the biomarker profiles identified.
Wawrocki et al. (67) described serum IL18, IL37, and CXCL10 as potential biomarker signatures for distinguishing ATB from LTBI. In the current study, we did not measure IL37; however, IL18 was present in the top 10 markers that contribute to principal component 2 (Figure 1), suggesting that this marker may play a role in distinguishing ATB from untreated LTBI. However, in our cohort, other markers were found to be more promising in discriminating ATB from LTBI, including CXCL10.
A limitation of our designed prediction model is the lack of samples from patients with different diagnoses, such as pneumonia, lung cancer, sarcoidosis, vasculitis, non-tuberculous mycobacterial infections, and aspergillosis. Others reported ADA(2) activity in populations of ATB, pneumonia, lung cancer, and pleural effusions of any origin with sensitivity and specificity values of ADA(2) ranging from 35% to 94% and 55% to 97%, respectively (19, 20, 68–70). In addition, both CXCL10 and VEGF have been described to be potential biomarkers for various diagnoses such as chronic obstructive pulmonary disease (COPD) and non-small cell lung cancer and for exacerbations of autoimmune diseases. The absence of a subanalysis for endemic background might be another limitation of this study. However, when comparing individuals from a high-burden country to participants from a low-burden country in the ATB and untreated LTBI groups of the discovery cohort, ADA and cytokine/chemokine measurements are not significantly different (p ≥ 0.18, data not shown).
The lack of a gold standard for LTBI is a known limitation. Until diagnostic tools are available to detect the presence of live Mtb in LTBI, we are dependent on the WHO definition for LTBI (2). To get better insight in LTBI individuals, we included two LTBI subgroups in the discovery cohort, which provides added value compared to several other studies (33, 42, 56, 71, 72). Treated LTBI individuals are assumed to be “cured,” and any difference between both LTBI groups possibly could indicate which of the untreated LTBI individuals represent persistent infection. Unfortunately, results of untreated LTBI and treated LTBI were highly overlapping in our cohorts and did not show any significant differences.
In conclusion, the combination of CCL1, CXCL10, VEGF, and ADA(2) activity in serum resulted in promising signatures able to differentiate between ATB and LTBI. These obtained biomarker signatures could facilitate the ongoing challenge of diagnosing TB and distinguishing ATB from LTBI. More research is necessary to optimize the prediction models in prospective clinical studies and including geographically different populations.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by Medical Research Ethics Committees-United (NL53628.100.15) and the Board of Directors of the Public Health and Diakonessenhuis, Utrecht, Netherlands. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
ED and LH processed the experimental data, drafted the article, and designed the figures. AB, LK, J-WL, ST, RH, and SN designed the study. ED, LH, and JD performed data analyses. SJ, TO, OA, DG, EP, AN, IH, and RH provided resources. BB, SP, and RH contributed to sample preparation and data analyses. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Italian Ministry of Health, Ricerca Corrente, Linea 4 and Italian Ministry of Health, GR-2018-12367178.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer HD declared a past co-authorship with the authors SJ and TO to the handling editor.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We kindly thank the nurses from Public Health Service of Utrecht for their assistance with the inclusion of participants, the nurses of INMI, Corine Prins, Krista van Meijgaarden, and colleagues for long-standing efforts in recruiting and organizing the Leiden Flow Up study samples. Lydia Kool and colleagues from the Medical Microbiological Laboratory Diakonessenhuis for technical assistance. Lysette Ebskamp-van Raaij, Karin van Veghel, and Amelia Lacna and the Multiplex Core Facility from Platform Immune Monitoring, University Medical Center Utrecht. In addition, we gratefully acknowledge all the participants of the different cohorts.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.725447/full#supplementary-material.
References
1. Sinha P, Gupta A, Prakash P, Anupurba S, Tripathi R, Srivastava GN. Differentiation of Mycobacterium Tuberculosis Complex From Non-Tubercular Mycobacteria by Nested Multiplex PCR Targeting IS6110, MTP40 and 32kd Alpha Antigen Encoding Gene Fragments. BMC Infect Dis (2016) 16:1–10. doi: 10.1186/s12879-016-1450-1
2. WHO. WHO | Global Tuberculosis Report 2019. (2020). Available at: https://www.who.int/teams/global-tuberculosis-programme/tb-reports/global-report-2019.
3. Cohen A, Mathiasen VD, Schön T, Wejse C. The Global Prevalence of Latent Tuberculosis: A Systematic Review and Meta-Analysis. Eur Respir J (2019) 54:1–4. doi: 10.1183/13993003.00655-2019
4. Sotgiu G, Goletti D, Matteelli A. Global Tuberculosis Prevention: Should We Start From the Beginning? Eur Respir J (2019) 54:1–4. doi: 10.1183/13993003.01394-2019
5. Houben RMGJ, Dodd PJ. The Global Burden of Latent Tuberculosis Infection: A Re-Estimation Using Mathematical Modelling. PloS Med (2016) 13:1–13. doi: 10.1371/journal.pmed.1002152
6. Cadena AM, Fortune SM, Flynn JL. Heterogeneity in Tuberculosis. Nat Rev Immunol (2017) 17:691–702. doi: 10.1038/nri.2017.69
7. Delogu G, Goletti D. The Spectrum of Tuberculosis Infection: New Perspectives in the Era of Biologics. J Rheumatol (2014) 91:11–16. doi: 10.3899/jrheum.140097
8. World Health Organization (WHO). Guidelines on the Management of Latent Tuberculosis Infection. (2015). Available at: https://www.who.int/publications/who-guidelines.
9. Goletti D, Lee MR, Wang JY, Walter N, Ottenhoff THM. Update on Tuberculosis Biomarkers: From Correlates of Risk, to Correlates of Active Disease and of Cure From Disease. Respirology (2018) 23:455–66. doi: 10.1111/resp.13272
10. Petruccioli E, Scriba TJ, Petrone L, Hatherill M, Cirillo DM, Joosten SA, et al. Correlates of Tuberculosis Risk: Predictive Biomarkers for Progression to Active Tuberculosis. Eur Respir J (2016) 48:1751–63. doi: 10.1183/13993003.01012-2016
11. Hofland RW, Bossink AWJ, Nierkens S, Paardekooper SPA, van den Broek BTA, Lammers JJ, et al. QuantiFERON-Plus Does Not Discriminate Between Active and Latent Tuberculosis QuantiFERON-Plus Does Not Discriminate Between Active and Latent Tuberculosis. Infect Dis (Auckl) (2018) 0:1–4. doi: 10.1080/23744235.2018.1425550
12. Esmail H, Cobelens F, Goletti D. Transcriptional Biomarkers for Predicting Development of Tuberculosis: Progress and Clinical Considerations. Eur Respir J (2020) 55:1–5. doi: 10.1183/13993003.01957-2019
13. Carranza C, Pedraza-Sanchez S, de Oyarzabal-Mendez E, Torres M. Diagnosis for Latent Tuberculosis Infection: New Alternatives. Front Immunol (2020) 11. doi: 10.3389/fimmu.2020.02006
14. Trébucq A, Enarson DA, Chiang CY, Van Deun A, Harries AD, Boillot F, et al. Xpert® MTB/RIF for National Tuberculosis Programmes in Low-Income Countries: When, Where and How? Int J Tuberc Lung Dis (2011) 15:1567–72. doi: 10.5588/ijtld.11.0392
15. Steingart KR, Schiller I, Horne DJ, Pai M, Boehme CC, Dendukuri N. Xpert® MTB/RIF Assay for Pulmonary Tuberculosis and Rifampicin Resistance in Adults. Cochrane Database Syst Rev (2014) 1:1–168. doi: 10.1002/14651858.CD009593.pub3
16. Marais BJ, Pai M. New Approaches and Emerging Technologies in the Diagnosis of Childhood Tuberculosis. Paediatr Respir Rev (2007) 8:124–33. doi: 10.1016/j.prrv.2007.04.002
17. Slump E, van Beurden KM, Erkens CGM, Schimmel HJ, van Soolingen D de VG. Tuberculose in Nederland 2019: Surveillancerapport Inclusief Rapportage Monitoring Van Interventies. (2020). Available at: https://www.rivm.nl/publicaties/tuberculose-in-nederland-2019-surveillancerapport-inclusief-rapportage-monitoring-van.
18. Torrado E, Cooper A. Cytokines in the Balance of Protection and Pathology During Mycobacterial Infections. Adv Exp Med Biol (2013) 783:121–40. doi: 10.1007/978-1-4614-6111-1
19. Yurt S, Kucukergin C, Yigitbas BA, Seckin S, Tigin HC, Kosar AF. Diagnostic Utility of Serum and Pleural Levels of Adenosine Deaminase 1-2, and Interferon-Gamma in the Diagnosis of Pleural Tuberculosis. Multidiscip Respir Med (2014) 9:1–7. doi: 10.1186/2049-6958-9-12
20. Conde MB, Marinho SR, Pereira MDF, Silva JRLE, Saad MHF, Sales CL, et al. The Usefulness of Serum Adenosine Deaminase 2 (ADA2) Activity in Adults for the Diagnosis of Pulmonary Tuberculosis. Respir Med (2002) 96:607–10. doi: 10.1053/rmed.2001.1273
21. Xie DL, Cheng B, Sheng Y, Jin J. Diagnostic Accuracy of Adenosine Deaminase for Tuberculous Pericarditis: A Meta-Analysis. Eur Rev Med Pharmacol Sci (2015) 19:4411–8.
22. Schutte C-M, Ungerer JPJ, Plessis H. Meyden CH Van Der. Significance of Cerebrospinal Fluid Adenosine Deaminase Isoenzymes in Tuberculous (TB) Meningitis. J Clin Lab Anal (2001) 15:236–8. doi: 10.1002/jcla.1033
23. Tao L, Ning HJ, Nie HM, Guo XY, Qin SY, Jiang HX. Diagnostic Value of Adenosine Deaminase in Ascites for Tuberculosis Ascites: A Meta-Analysis. Diagn Microbiol Infect Dis (2014) 79:102–7. doi: 10.1016/j.diagmicrobio.2013.12.010
24. Penn-Nicholson A, Hraha T, Thompson EG, Sterling D, Mbandi SK, Wall KM, et al. Discovery and Validation of a Prognostic Proteomic Signature for Tuberculosis Progression: A Prospective Cohort Study. PloS Med (2019) 16:1–21. doi: 10.1371/journal.pmed.1002781
25. Lubbers R, Sutherland JS, Goletti D, De Paus RA, Van Moorsel CHM, Veltkamp M, et al. Complement Component C1q as Serum Biomarker to Detect Active Tuberculosis. Front Immunol (2018) 23. doi: 10.3389/fimmu.2018.02427
26. Ahmad R, Xie L, Pyle M, Suarez MF, Broger T, Steinberg D, et al. A Rapid Triage Test for Active Pulmonary Tuberculosis in Adult Patients With Persistent Cough. Sci Transl Med (2019) 11:1–14. doi: 10.1126/scitranslmed.aaw8287
27. Petrone L, Cannas A, Vanini V, Cuzzi G, Aloi F, Nsubuga M, et al. Blood and Urine Inducible Protein 10 as Potential Markers of Disease Activity. Int J Tuberc Lung Dis (2016) 20:1554–61. doi: 10.5588/ijtld.16.0342
28. Morris TC, Hoggart CJ, Chegou NN, Kidd M, Oni T, Goliath R, et al. Evaluation of Host Serum Protein Biomarkers of Tuberculosis in Sub-Saharan Africa. Front Immunol (2021) 12. doi: 10.3389/fimmu.2021.639174
29. Mutavhatsindi H, van der Spuy GD, Malherbe ST, Sutherland JS, Geluk A, Mayanja-Kizza H, et al. Validation and Optimization of Host Immunological Bio-Signatures for a Point-Of-Care Test for TB Disease. Front Immunol (2021) 12. doi: 10.3389/fimmu.2021.607827
30. El-emiry FA, Attia GA, Ahmad AY, Sakr BM. Diagnostic Value of Inducible Protein-10 in Pulmonary Tuberculosis. Egypt J Chest Dis Tuberc (2016) 65:219–25. doi: 10.1016/j.ejcdt.2015.08.012
31. Chen T, Li Z, Yu L, Li H, Lin J, Guo H, et al. Profiling the Human Immune Response to Mycobacterium Tuberculosis by Human Cytokine Array. Tuberculosis (2016) 97:108–17. doi: 10.1016/j.tube.2015.12.007
32. Rozot V, Patrizia A, Vigano S, Mazza-Stalder J, Idrizi E, Day CL, et al. Combined Use of Mycobacterium Tuberculosis-Specific CD4 and CD8 T-Cell Responses Is a Powerful Diagnostic Tool of Active Tuberculosis. Clin Infect Dis (2015) 60:432–7. doi: 10.1093/cid/ciu795
33. Marín ND, París SC, Rojas M, García LF. Functional Profile of CD4+ and CD8+ T Cells in Latently Infected Individuals and Patients With Active TB. Tuberculosis (2013) 93:155–66. doi: 10.1016/j.tube.2012.12.002
34. Chiacchio T, Petruccioli E, Vanini V, Cuzzi G, Pinnetti C, Sampaolesi A, et al. Polyfunctional T-Cells and Effector Memory Phenotype Are Associated With Active TB in HIV-Infected Patients. J Infect (2014) 69:533–45. doi: 10.1016/j.jinf.2014.06.009
35. Joosten SA, van Meijgaarden KE, del Nonno F, Baiocchini A, Petrone L, Vanini V, et al. Patients With Tuberculosis Have a Dysfunctional Circulating B-Cell Compartment, Which Normalizes Following Successful Treatment. PloS Pathog (2016) 12:1–24. doi: 10.1371/journal.ppat.1005687
36. Petruccioli E, Navarra A, Petrone L, Vanini V, Cuzzi G, Gualano G, et al. Use of Several Immunological Markers to Model the Probability of Active Tuberculosis. Diagn Microbiol Infect Dis (2016) 86:169–71. doi: 10.1016/j.diagmicrobio.2016.06.007
37. Chegou NN, Sutherland JS, Malherbe S, Crampin AC, Corstjens PLAM, Geluk A, et al. Diagnostic Performance of a Seven-Marker Serum Protein Biosignature for the Diagnosis of Active TB Disease in African Primary Healthcare Clinic Attendees With Signs and Symptoms Suggestive of TB. Thorax (2016) 71:785–794. doi: 10.1136/thoraxjnl-2015-207999
38. Chen T, Lin J, Wang W, Fleming J, Chen L, Wang Y, et al. Cytokine and Antibody Based Diagnostic Algorithms for Sputum Culture-Positive Pulmonary Tuberculosis. PloS One (2015) 10:1–16. doi: 10.1371/journal.pone.0144705
39. Clifford V, Tebruegge M, Zufferey C, Denholm J, Curtis N. Serum IP-10 in the Diagnosis of Latent and Active Tubeculosis. J Infect (2015) 71:696–8. doi: 10.1016/j.jinf.2015.08.001
40. Hur Y, Ae Y, Jang S, Young J, Kim A, Lee SA, et al. Adjunctive Biomarkers for Improving Diagnosis of Tuberculosis and Monitoring Therapeutic Effects. J Infect (2015) 70:346–55. doi: 10.1016/j.jinf.2014.10.019
41. Hong JY, Jung GS, Kim H, Kim YM, Lee HJ, Cho SN, et al. Efficacy of Inducible Protein 10 as a Biomarker for the Diagnosis of Tuberculosis. Int J Infect Dis (2012) 16:855–9. doi: 10.1016/j.ijid.2012.07.013
42. Harari A, Rozot V, Enders FB, Perreau M, Stalder JM, Nicod LP, et al. Dominant TNF-α+ Mycobacterium Tuberculosis-Specific CD4+ T Cell Responses Discriminate Between Latent Infection and Active Disease. Nat Med (2011) 17:372–7. doi: 10.1038/nm.2299
43. Rozot V, Vigano S, Mazza-stalder J, Idrizi E, Day CL, Perreau M, et al. Mycobacterium Tuberculosis -Specific CD8+ T Cells Are Functionally and Phenotypically Different Between Latent Infection and Active Disease. Eur J Immunol (2013) 43:1568–77. doi: 10.1002/eji.201243262
44. Petruccioli E, Petrone L, Vanini V, Sampaolesi A, Gualano G, Girardi E, et al. Ifnγ/Tnfα Specific-Cells and Effector Memory Phenotype Associate With Active Tuberculosis. J Infect (2013) 66:475–86. doi: 10.1016/j.jinf.2013.02.004
45. Caccamo N, Guggino G, Joosten SA, Gelsomino G, Di CP, Titone L, et al. Multifunctional CD4+ T Cells Correlate With Active Mycobacterium Tuberculosis Infection. Eur J Immunol (2010) 40:2211–20. doi: 10.1002/eji.201040455
46. Lewinsohn DM, Leonard MK, Lobue PA, Cohn DL, Daley CL, Desmond E, et al. Official American Thoracic Society/Infectious Diseases Society of America/Centers for Disease Control and Prevention Clinical Practice Guidelines: Diagnosis of Tuberculosis in Adults and Children. Clin Infect Dis (2017) 64:111–5. doi: 10.1093/cid/ciw778
47. de Paus RA, van Meijgaarden KE, Prins C, Kamphorst MH, Arend SM, Ottenhoff THM, et al. Immunological Characterization of Latent Tuberculosis Infection in a Low Endemic Country. Tuberculosis (2017) 106:62–72. doi: 10.1016/j.tube.2017.07.001
48. De JW, Prakken BJ, Bijlsma JWJ, Kuis W, Rijkers GT. Improved Multiplex Immunoassay Performance in Human Plasma and Synovial Fluid Following Removal of Interfering Heterophilic Antibodies. J Immunol Methods (2005) 300:124–35. doi: 10.1016/j.jim.2005.03.009
49. de Jager W, te Velthuis H, Prakken BJ, Kuis W, Rijkers GT. Simultaneous Detection of 15 Human Cytokines in a Single Sample of Stimulated Peripheral Blood Mononuclear Cells. Clin Diagn Lab Immunol (2003) 10:133–9. doi: 10.1128/CDLI.10.1.133-139.2003
50. Muraoka T, Katsuramarki T, Shiraishi H, Yokoyama MM. Automated Enzymatic Measurement of Adenosine Deaminase Isoenzyme Activities in Serum. Anal Biochem (1990) 187:268–72. doi: 10.1016/0003-2697(90)90455-I
51. Le Cao K-A, Rohart F, Gonzalez I, Dejean S, with key contributors Benoit Gautier, Francois Bartolo, Contributions from Pierre Monget, Jeff Coquery FY and BL. Mixomics: Omics Data Integration Project. (2016). Available at: http://mixomics.org/.
52. Team RC. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing (2013). Available at: http://lib.stat.cmu.edu/R/CRAN/doc/manuals/r-devel/fullrefman.pdf.
53. Prabhakaran S. R Package ‘InformationValue (2016). Available at: http://r-statistics.co/Information-Value-With-R.html.
54. Yoon C, Semitala FC, Atuhumuza E, Katende J, Mwebe S, Asege L, et al. Point-Of-Care C-Reactive Protein-Based Tuberculosis Screening for People Living With HIV: A Diagnostic Accuracy Study. Lancet Infect Dis (2017) 17:1285–92. doi: 10.1016/S1473-3099(17)30488-7
55. Miller MD, Kranger MS. The Human Cytokine I-309 Is a Monocyte Chemoattractant. Proc Natl Acad Sci USA (1992) 89:2950–4. doi: 10.1073/pnas.89.7.2950
56. Chegou NN, Heyckendorf J, Walzl G, Lange C, Ruhwald M. Beyond the IFN-γ Horizon: Biomarkers for Immunodiagnosis of Infection With Mycobacterium Tuberculosis. Eur Respir J (2014) 43:1472–86. doi: 10.1183/09031936.00151413
57. Matsuyama W, Hashiguchi T, Matsumuro K, Iwami F, Hirotsu Y, Kawabata M, et al. Increased Serum Level of Vascular Endothelial Growth Factor in Pulmonary Tuberculosis. Am J Respir Crit Care Med (2000) 162:1120–2. doi: 10.1164/ajrccm.162.3.9911010
58. Ferrara N, Gerber HP, LeCoutier J. The Biology of VEGF and Its Receptors. Nat Med (2003) 9:669–76. doi: 10.1038/nm0603-669
59. Joosten SA, Van Meijgaarden KE, Arend SM, Prins C, Oftung F, Korsvold GE, et al. Mycobacterial Growth Inhibition Is Associated With Trained Innate Immunity. J Clin Invest (2018) 128:1837–51. doi: 10.1172/JCI97508
60. Palucci I, Battah B, Salustri A, De Maio F, Petrone L, Ciccosanti F, et al. IP-10 Contributes to the Inhibition of Mycobacterial Growth in an Ex Vivo Whole Blood Assay. Int J Med Microbiol (2019) 309:299–306. doi: 10.1016/j.ijmm.2019.05.005
61. Kishimoto T. INTERLEUKIN-6: From Basic Science to Medicine—40 Years in Immunology. Annu Rev Immunol (2005) 23:1–21. doi: 10.1146/annurev.immunol.23.021704.115806
62. Brown J, Clark K, Smith C, Hopwood J, Lynard O, Toolan M, et al. Variation in C - Reactive Protein Response According to Host and Mycobacterial Characteristics in Active Tuberculosis. BMC Infect Dis (2016) 16:1–8. doi: 10.1186/s12879-016-1612-1
63. Zavialov AV, Gracia E, Glaichenhaus N, Franco R, Zavialov AV, Lauvau G. Human Adenosine Deaminase 2 Induces Differentiation of Monocytes Into Macrophages and Stimulates Proliferation of T Helper Cells and Macrophages. J Leukoc Biol (2010) 88:279–90. doi: 10.1189/jlb.1109764
64. Bae MJ, Ryu S, Kim HJ, Cha SI, Kim CH, Lee J. Mycobacterium Tuberculosis ESAT6 and CPF10 Induce Adenosine Deaminase 2 mRNA Expression in Monocyte-Derived Macrophages. Tuberc Respir Dis (Seoul) (2017) 80:77–82. doi: 10.4046/trd.2017.80.1.77
65. Canbolat O, Ulusdoyuran S, Özgen G, Ceyhan I, Gümüşlü F, Akbay A. The Comparison of Adenosine Deaminase Activity Values With Polymerase Chain Reaction Results in Patients With Tuberculosis. J Clin Lab Anal (1999) 13:209–12. doi: 10.1002/(SICI)1098-2825(1999)13:5<209::AID-JCLA3>3.0.CO;2-F
66. Afrasiabian S, Mohsenpour B, Bagheri KH, Sigari N, Aftabi K. Diagnostic Value of Serum Adenosine Deaminase Level in Pulmonary Tuberculosis. J Res Med Sci India (2013) 18:252–4.
67. Wawrocki S, Seweryn M, Kielnierowski G, Rudnicka W, Wlodarczyk M, Druszczynska M. IL-18/IL-37/IP-10 Signalling Complex as a Potential Biomarker for Discriminating Active and Latent TB. PloS One (2019) 14:1–19. doi: 10.1371/journal.pone.0225556
68. Salmanzadeh S, Tavakkol H, Bavieh K, Alavi SM. Diagnostic Value of Serum Adenosine Deaminase (ADA) Level for Pulmonary Tuberculosis. Jundishapur J Microbiol (2015) 8:1–5. doi: 10.5812/jjm.21760
69. Kanchan SK, Santosh VG, Vishal SM, Leela AG, Niyogi DG, Joshi D. Study of Adenosine Deaminase Levels in Patients of Pulmonary Tuberculosis With and Without Pleural Effusion. IOSR J Dent Med Sci e-ISSN (2014) 13:30–7. doi: 10.9790/0853-13193037
70. Gorguner M, Cerci M, Gorguner I. Determination of Adenosine Deaminase Activity and Its Isoenzymes for Diagnosis of Pleural Effusions. Respirology (2000) 5:321–4.
71. Petruccioli E, Petrone L, Vanini V, Cuzzi G, Lauri FN, Goletti D. Different Polyfunctional Characterstics of RD1 Specific CD4 T Cells in Active TB and LTBI. Eur Respir J (2012) 40:Supplement 56.
Keywords: active tuberculosis (ATB), biomarker, diagnosis, adenosine deaminase activity (ADA), CCL1, vascular endothelial growth factor (VEGF), CXCL10 (IP-10), latent tuberculosis infection (LTBI)
Citation: Delemarre EM, van Hoorn L, Bossink AWJ, Drylewicz J, Joosten SA, Ottenhoff THM, Akkerman OW, Goletti D, Petruccioli E, Navarra A, van den Broek BTA, Paardekooper SPA, van Haeften I, Koenderman L, Lammers J-WJ, Thijsen SFT, Hofland RW and Nierkens S (2021) Serum Biomarker Profile Including CCL1, CXCL10, VEGF, and Adenosine Deaminase Activity Distinguishes Active From Remotely Acquired Latent Tuberculosis. Front. Immunol. 12:725447. doi: 10.3389/fimmu.2021.725447
Received: 15 June 2021; Accepted: 15 September 2021;
Published: 07 October 2021.
Edited by:
Annalisa Del Prete, University of Brescia, ItalyReviewed by:
Hazel Marguerite Dockrell, University of London, United KingdomNoyal Mariya Joseph, Jawaharlal Institute of Postgraduate Medical Education and Research, India
Damien Portevin, Swiss Tropical and Public Health Institute, Switzerland
Copyright © 2021 Delemarre, van Hoorn, Bossink, Drylewicz, Joosten, Ottenhoff, Akkerman, Goletti, Petruccioli, Navarra, van den Broek, Paardekooper, van Haeften, Koenderman, Lammers, Thijsen, Hofland and Nierkens. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stefan Nierkens, cy5uaWVya2Vuc0B1bWN1dHJlY2h0Lm5s
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share last authorship