 Iris Paola Guzmán-Guzmán1*
Iris Paola Guzmán-Guzmán1* Claudia Isabel Ramírez-Vélez1
Claudia Isabel Ramírez-Vélez1 Ramcés Falfán-Valencia2
Ramcés Falfán-Valencia2 José Eduardo Navarro-Zarza3
José Eduardo Navarro-Zarza3 Ilse Adriana Gutiérrez-Pérez1
Ilse Adriana Gutiérrez-Pérez1 Oscar Zaragoza-García1Mónica Ramírez4Natividad Castro-Alarcón1
Oscar Zaragoza-García1Mónica Ramírez4Natividad Castro-Alarcón1 Isela Parra-Rojas1
Isela Parra-Rojas1- 1Faculty of Chemical-Biological Sciences, Universidad Autónoma de Guerrero, Chilpancingo, Mexico
- 2HLA Laboratory, Instituto Nacional de Enfermedades Respiratorias Ismael Cosío Villegas, Mexico City, Mexico
- 3Department of Rheumatology and Internal Medicine, Hospital General de Chilpancingo Dr. Raymundo Abarca Alarcón, Guerrero, Mexico
- 4Consejo Nacional de Ciencia y Tecnología, Universidad Autónoma de Guerrero, Chilpancingo, Mexico
The enzymes of the family peptidylarginine deiminases (PADs) have an important role in the pathogenesis of rheumatoid arthritis (RA) due to their association with the anti-citrullinated protein antibodies (ACPA) production. To evaluate the association between single-nucleotide polymorphisms (SNPs) in the PADI2 gene and RA susceptibility, related clinical parameters, and the serologic status of autoantibodies in a women population with RA from southern Mexico, a case-control study was conducted (case n=229; control n=333). Sociodemographic characteristics were evaluated, along with clinical parameters, inflammation markers, the levels of ACPAs as anti-cyclic citrullinated peptides (anti-CCPs), anti-modified citrullinated vimentin (anti-MCV), and rheumatoid factor (RF). Genomic DNA was extracted from peripheral blood, and three SNPs of the PADI2 gene (rs1005753, rs2057094, and rs2235926) were performed by qPCR using TaqMan probes. The data analysis reveals that the carriers of the T allele for rs2057094 and rs2235926 presented an earlier onset of the disease (β= -3.26; p = 0.03 and β = -4.13; p = 0.015, respectively) while the carriers of the T allele for rs1005753 presented higher levels of anti-CCPs (β= 68.3; p = 0.015). Additionally, the T allele of rs2235926 was associated with a positive RF (OR = 2.90; p = 0.04), anti-MCV (OR = 2.92; p = 0.05), and with the serologic status anti-CCP+/anti-MCV+ (OR = 3.02; p = 0.03), and anti-CCP+/anti-MCV+/RF+ (OR = 3.79; p = 0.004). The haplotypes GTT (OR =1.52; p = 0.027) and TTT (OR = 1.32; p = 0.025) were associated with the presence of RA. In addition, in this study the haplotype TTT is linked to the presence of radiographic joint damage defined by a Sharp-van der Heijde score (SHS) ≥2 (OR = 1.97; p = 0.0021) and SHS ≥3 (OR = 1.94; p = 0.011). The haplotype TTT of SNPs rs1005753, rs2057094, and rs2235926 of the PADI2 gene confers genetic susceptibility to RA and radiographic joint damage in women from southern Mexico. The evidence reveals that SNPs of the PADI2 gene favors the presence of a positive serologic status in multiple autoantibodies and the clinical manifestations of RA at an early onset age.
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease with a variable prevalence amongst populations (1, 2). Clinical manifestations for RA can be from a mild self-limiting arthritis to a progressive multisystemic inflammatory arthritis with high morbidity and mortality (3, 4). In the RA physiopathology, enzymes of family peptidylarginine deiminases (PADs) have an important role in the citrullination of proteins that promote the antibody synthesis against citrullinated proteins (ACPAs) (5–8). The ACPAs positive status has been described as a predictive marker for the severity, the radiological degree (9, 10), joint damage, and the functional disability in patients with RA (11, 12). The growth of ACPAs aimed against citrullinated proteins’ epitopes –histones, vimentin and enolase-derived peptides– and fibrinogen is mainly identified during pre-clinical stages of RA. Furthermore, the levels of ACPAs are correlated to the increase of proinflammatory cytokines (TNF-α, IL-6, IL-12p70, IFN-γ, IL-2, and IL-15) and high sensitivity C-reactive protein (hsCRP) (13). In addition, it has been stated that its positive status, along with the rheumatoid factor (RF), promotes the inflammatory process in RA (14).
The expression of the PAD2 and PAD4 isoenzymes have been identified in synovial tissue and fluid in RA patients (15, 16). Although PADs exhibit a limited expression in some tissues, PAD2 is considered the broader expressed isoform (17), and the specificity of its substratum could be related to the clinic phenotype and serologic variability observed in RA patients. It was determined that PAD2 possesses specificity against β/γ-actin, myelin basic protein, histones (H3R26), vimentin, and the glial fibrillary acidic protein (6, 18–21). Meanwhile, for PAD4, the main protein substrates are histones (H2A, H3R2, H3R8, H3R17, H3R26, and H4R3), nuclear lamin C, nucleophosmin/B23, p300/CBP, p21, and inhibitor of growth 4 (6, 22–27). Particularly, high levels of PAD2 have been reported in synovial fluid of RA patients (28–31), as well as their correlation with inflammation markers, the disease’s clinical activity, and the anti-CCPs levels (32).
On the other hand, the influence of genetic factors on the modulation of the expression and function of PADs has been proven (8), and, even though the role of single-nucleotide polymorphisms (SNPs) and of a functional haplotype in PADI4 has been established, few studies that have determined the role of SNPs in PADI2 in the genetic susceptibility to RA or its association with the serologic status and the clinical parameters that are related to the disease. Lee et al. reported that PADI2 could be a candidate-like gene for RA (33) and, in some populations, it was determined that SNPs rs2076596 (34), rs1005753 (35), rs2057094, and rs2235926 (29) in the PADI2 gene are associated with the presence of RA.
This study aims to evaluate the association between SNPs, rs1005753, rs2057094, and rs2235926 of the PADI2 gene and RA susceptibility as the relation with the clinical parameters, inflammation markers, and the serologic status for antibodies in a women population from southern Mexico.
Materials and Methods
Subject Selection
A case-control study was carried out in RA patients (n = 229 women). These patients were diagnosed with RA according to the ACR/EULAR criteria 2010 (36). The control group (n = 333 women) was treated for external causes to RA, autoimmune disease, musculoskeletal disease, or cancer in the General Hospital Dr. Raymundo Abarca Alarcón, in Chilpancingo, in the state of Guerrero, Mexico. The case and control subjects were recruited during the period from December 2017 to December 2019. The study was approved by the Research Ethics Committee of the Autonomous University of Guerrero, Mexico (approval code CB-004/2017). All patients agreed to participate and gave their informed consent in writing.
Clinical Assessment
All of the patients were surveyed to obtain sociodemographic data. The clinical and treatment characteristics were evaluated during the consultation and from the clinical file. In this study “Patients of recent diagnostic” refers to those patients without pharmacological prescription for anti-rheumatic treatment to the date of the sample obtainment.
The rheumatologist performed a clinical evaluation and counted the number of inflamed and painful joints. The patient indicated the level of perception of health status and level of pain perception through a visual analog scale. Rheumatoid arthritis disease activity and the disability level were evaluated through the Disease Activity Score 28 (DAS28), and the Spanish version of the Health Assessment Questionnaire (HAQ-DI), respectively.
To determine joint damage, a radiographic evaluation was performed to observe features of destructive and proliferative changes, and radiological damage. According to Sharp-van der Heijde Score (SHS), this method reviews plain films of 8 proximal interphalangeal joints, 2 interphalangeal thumb joints, 10 metacarpophalangeal joints, and both wrists. The method defines 5 categories: 0 = normal; 1 = asymmetrical or minimal narrowing up to a maximum of 25%; 2 = definite narrowing with loss of up to 50% of the normal space; 3 = definite narrowing with loss of 50-99% of the normal space or subluxation; and, 4 = absence of joint space, presumptive evidence of ankylosis, or complete luxation (37).
Assay of RA-Related Antibodies and Inflammation Markers
Using a venous blood sample, the erythrocyte sedimentation rate (ESR) was analyzed by the Wintrobe method. Serum samples were used to determine the levels of high sensitivity C- reactive protein (hsCRP) and rheumatoid factor (RF) isotype IgG, using the immunoturbidimetry technique in the automatized reader (COBAS C311, Roche Diagnostics GmbH, Germany). In addition, the anti-cyclic citrullinated peptide (anti-CCPs) and anti-modified citrullinated vimentin (anti-MCV) autoantibodies were measured with a second-generation ELISA kit (DIASTAT anti-CCP Axis-Shield, Dundee, United Kindom; and ORGENTEC Diagnostika GmbH, Mainz, Germany, respectively). Values >20 UI/mL were considered positive for RF, values >5 U/mL for anti-CCPs, and >20 U/mL for anti-MCV, according to the manufacturer instructions.
DNA Extraction and TaqMan Genotyping
Genomic DNA was extracted from peripheral blood and the SNPs were genotyped by allele discrimination using commercial TaqMan probes (Applied Biosystems, San Francisco, CA, USA). The evaluated SNPs were C_2190445_20 (rs1005753/Intron, cat. 4351379), C_11647256_20 (rs2057094/Intron, Cat.4351379), and C_2190476_1_ (rs2235926/3´UTR, Cat.4351379). These were evaluated using quantitative polymerase chain reaction (qPCR) in a 7300 Real-Time PCR System (Applied Biosystems/Thermo Fisher Scientific Inc., Singapore), following the instructions by the manufacturer. The thermal cycling was performed by denaturation at 60°C for 30 sec, followed by 40 cycles of 95°C for 10 min and 95°C for 15 sec, and alignment and extension at 60°C 1 min and 4°C. Genotype analysis was performed through the sequence detection software (SDS) version 2.3 (Applied Biosystems, CA, United States).
Statistical Analysis
The categorical variables were expressed as numbers and proportions, and they were compared using a Chi-squared test. For quantitative variables, median and percentiles p5-p95th were used according to the Mann Whitney test to compare groups. Allele and genotype frequencies were calculated by direct counting. The differences in the distributions of allele and genotype frequencies between cases and controls, and the associations between clinical characteristics in patients with RA, were performed using a Chi-square test and a logistic regression. The Hardy-Weinberg equilibrium (HWE) was assessed in both groups. The effect of polymorphisms on clinical assessment and serum autoantibodies levels and other clinical parameters was tested using β coefficient and their standard errors, adjusted by age, treatment with disease-modifying anti-rheumatic drugs (DMARDs) and wood smoke exposure. Statistical analysis was carried out using the Stata version 13.0 (StataCorp, College Station, TX, USA). The association and pairwise measure of linkage disequilibrium of the SNPs rs1005753, rs2057094, and rs2235926 of PADI2 was calculated using SHEsis software (38) (Figure 1). The association between SNPs and clinical parameters and serologic status was determined using logistic regression models, settling odds ratios (ORs), and 95% confidence intervals (95% CI). Results were considered significant at p <0.05.
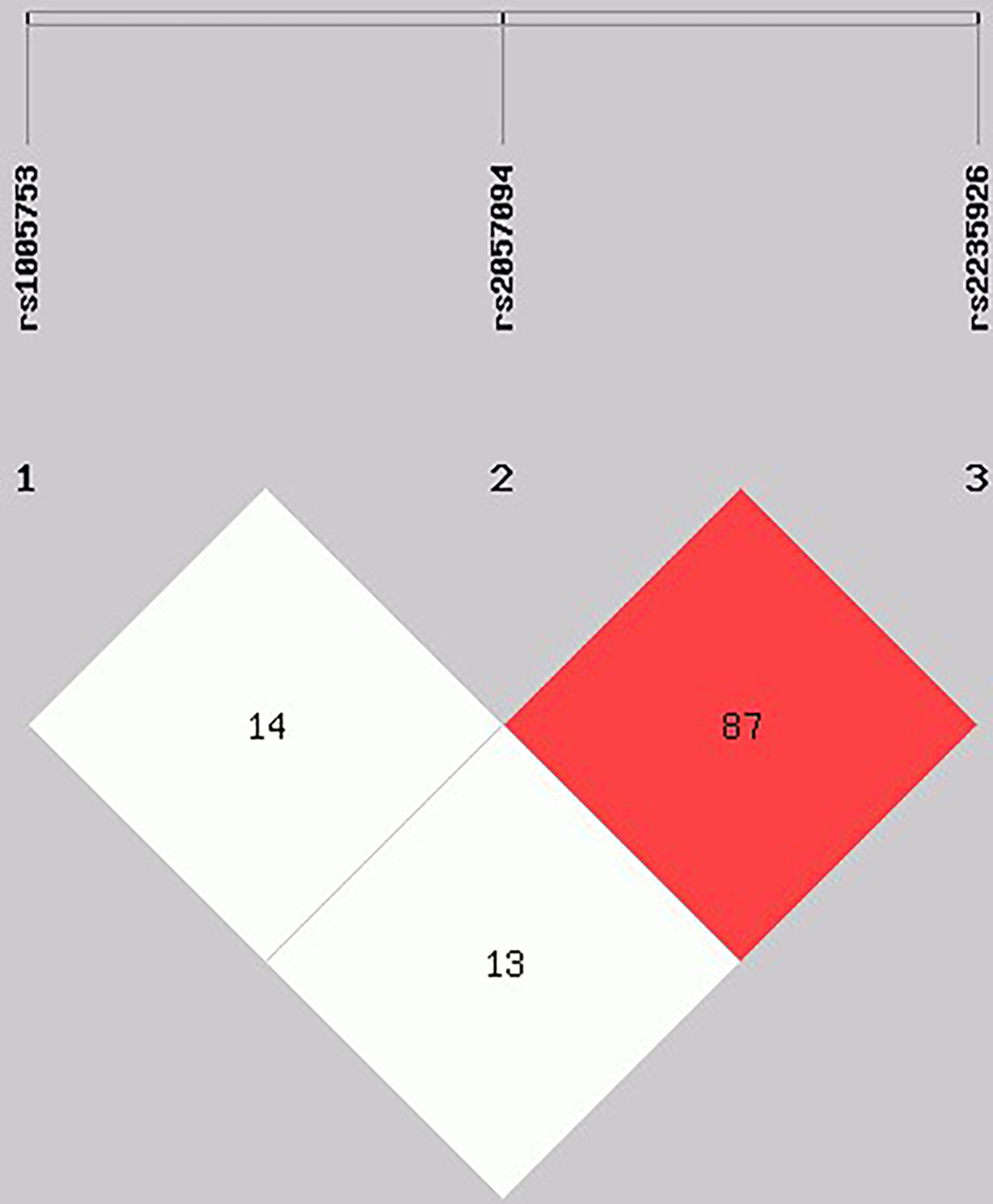
Figure 1 Linkage disequilibrium (LD) test of PADI2 gene SNPs in RA patients. Haplotype frequencies and LD were calculated using SHEsis software. Red area represents higher levels of LD. A D´ value of 100 indicates a complete LD between two markers and a D´ value of 0 indicates complete linkage equilibrium.
Results
Demographic Data, Clinical Features, and Serologic Status
The sociodemographic and clinical characteristics between the case and control groups are presented in Table 1. The participants with RA displayed a period of evolution of the disease with a median of 7 years, along with a low functional disability HAQ-DI score. At the same time, they also presented a moderate clinical activity and a radiographic joint damage index that refers to the presence of the narrowing of the intra-articular space and bone erosion, even though most of them were pharmacologically treated with DMARDs. Furthermore, the autoantibodies’ levels (RF, anti-CCPs, and anti-MCV) were elevated. When comparing the positive serologic status to the antibodies, more than 85% of the subjects were positive, with a predominant positive status for anti-MCV (89.1%). After evaluating the serologic status of combined positivity, it was found that 83.8% were anti-CCPs+/anti-MCV+ and that 75.1% were positive for ACPAs and RF (anti-CCP+/anti-MCV+/RF+) (Table 2).
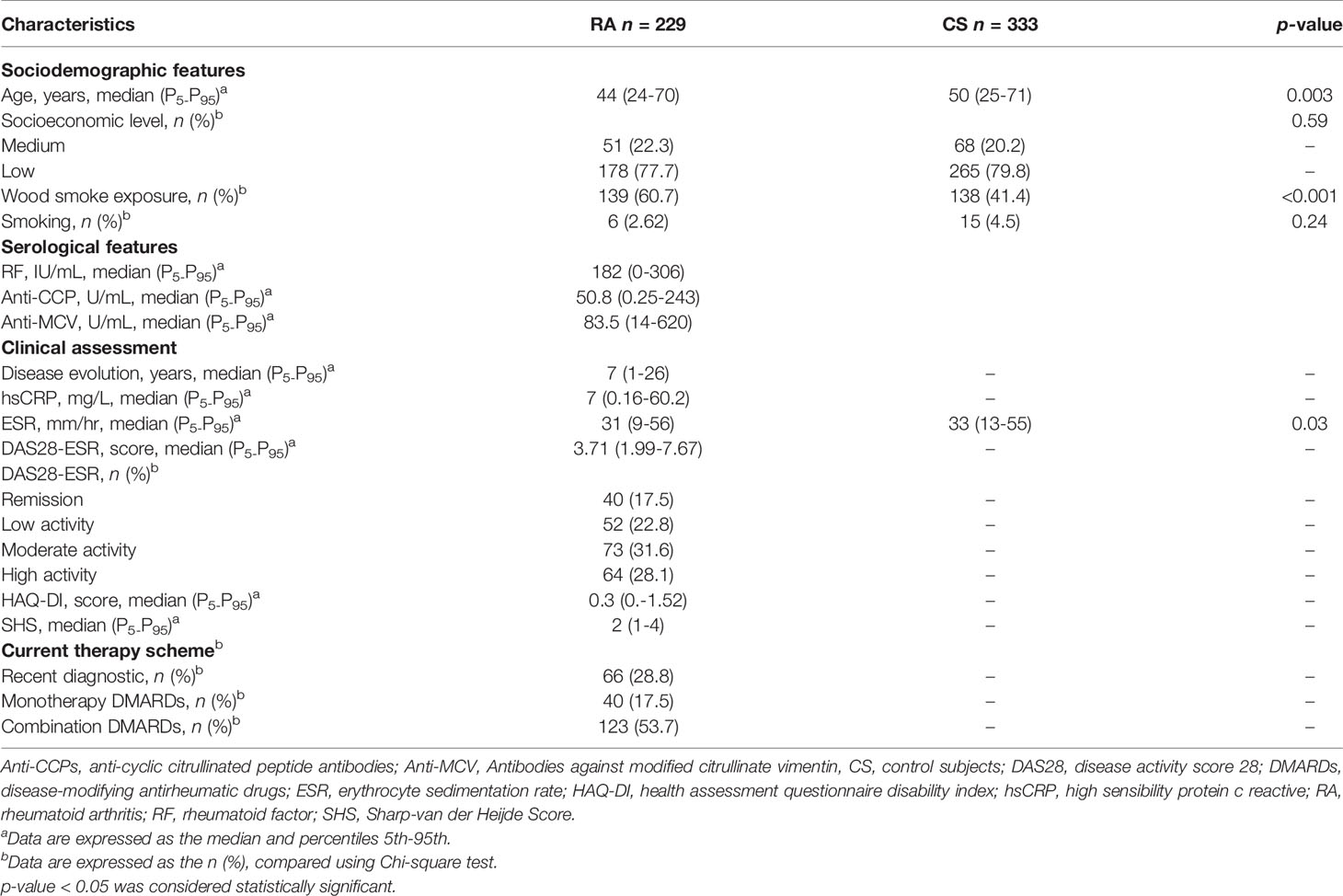
Table 1 Demographics and clinical characteristics in the study population.
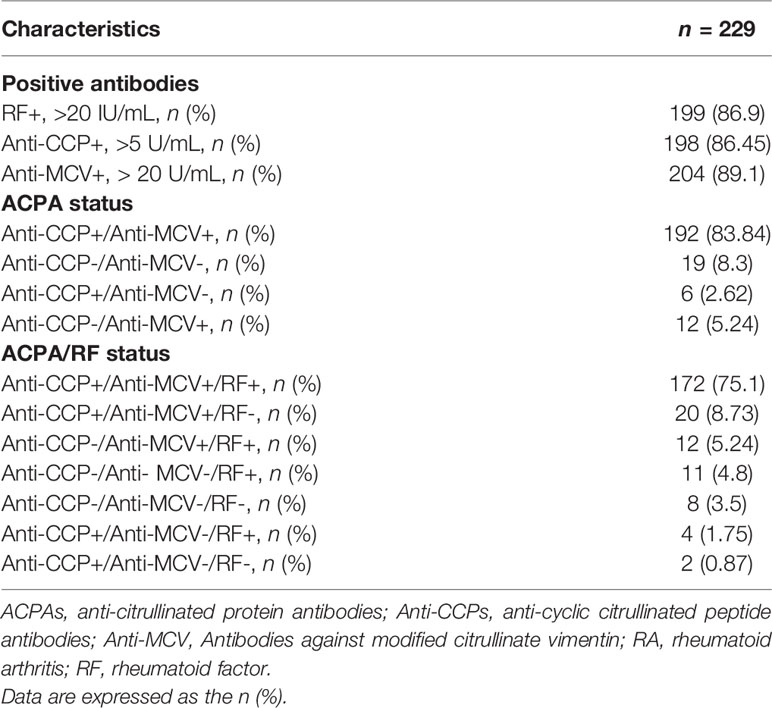
Table 2 Serologic pattern status in RA patients.
Frequency of PADI2 Gene SNPs and Haplotype Analysis
The genotypical distribution of the SNPs rs1005753 (T>G), rs2057094 (T>C), and rs2235926 (T>C) of the PADI2 gene was in Hardy-Weinberg genetic equilibrium in both studied groups (p > 0.5). The genotypical and allele frequencies are shown in Table 3. The T allele of rs2057094 was marginally associated with RA susceptibility (OR = 1.27; 95% CI, 0.99-1.65; p = 0.05), however, the haplotypes TTT (OR=1.32; 95% CI, 1.03-1.67; p = 0.025) and GTT (OR = 1.52; 95% CI, 1.04-2.19; p = 0.027) were significantly associated with RA susceptibility (Table 4).
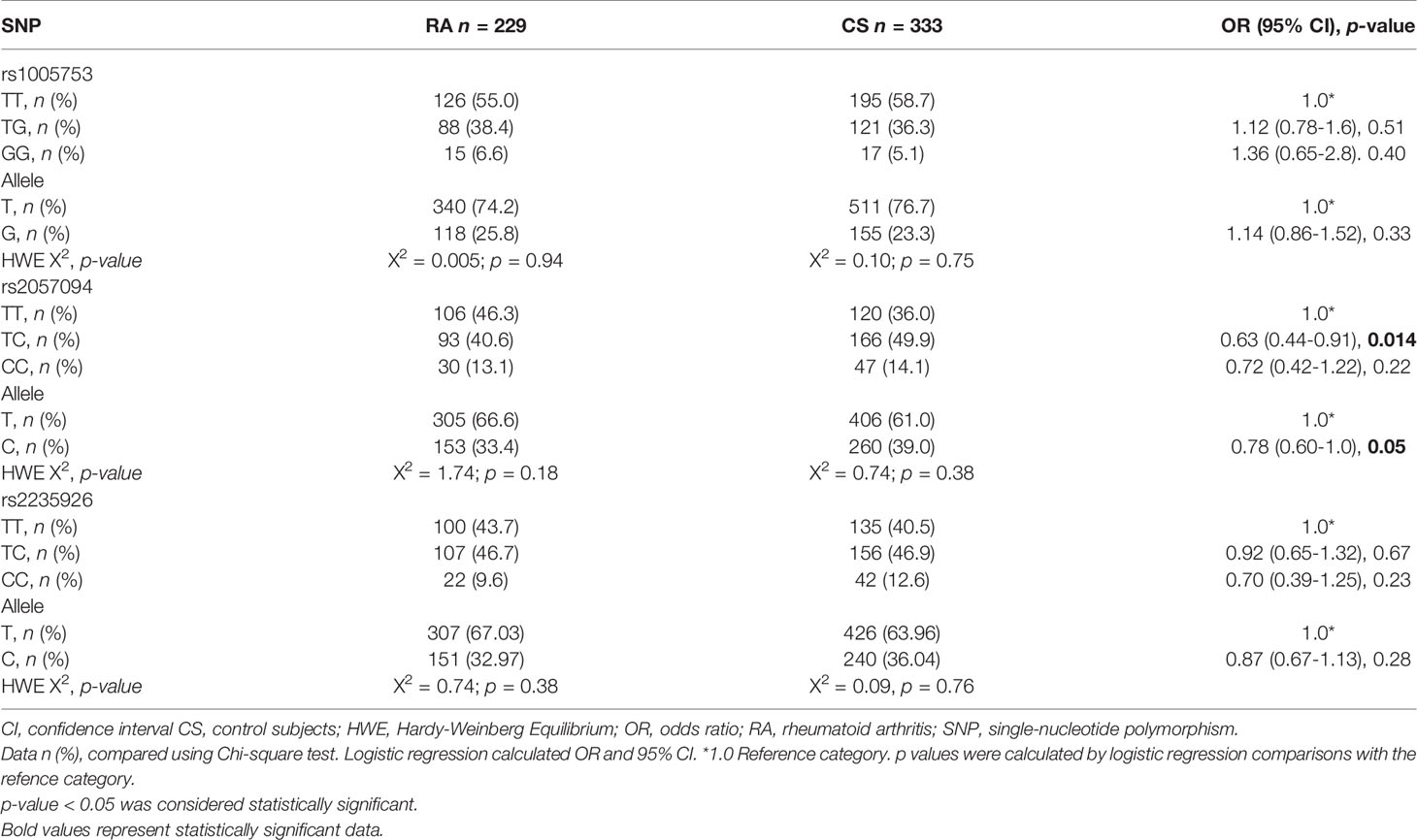
Table 3 Genotypic and allele frequencies of polymorphisms in PADI2 gene.

Table 4 Haplotype frequencies of three PADI2 SNPs in RA and CS.
Association of Clinical Parameters and Serologic Status According to the SNPs and Haplotype of the PADI2 Gene in RA Patients
In this study, we evaluated the potential effect and the association between PADI2’s SNPs and clinical parameters, inflammation markers, and the serologic status in opposition to autoantibodies in RA patients. The presence of the T allele of SNPs rs2057094 and rs2235926 was associated with the presence of the disease’s clinical signs at a younger age (β = -3.26; p = 0.03 and β = -4.13; p = 0.02, years respectively), while the presence of the T allele of rs1005753 was associated with the increase of the anti-CCPs levels (β = 68.3; p = 0.01) (Table 5).
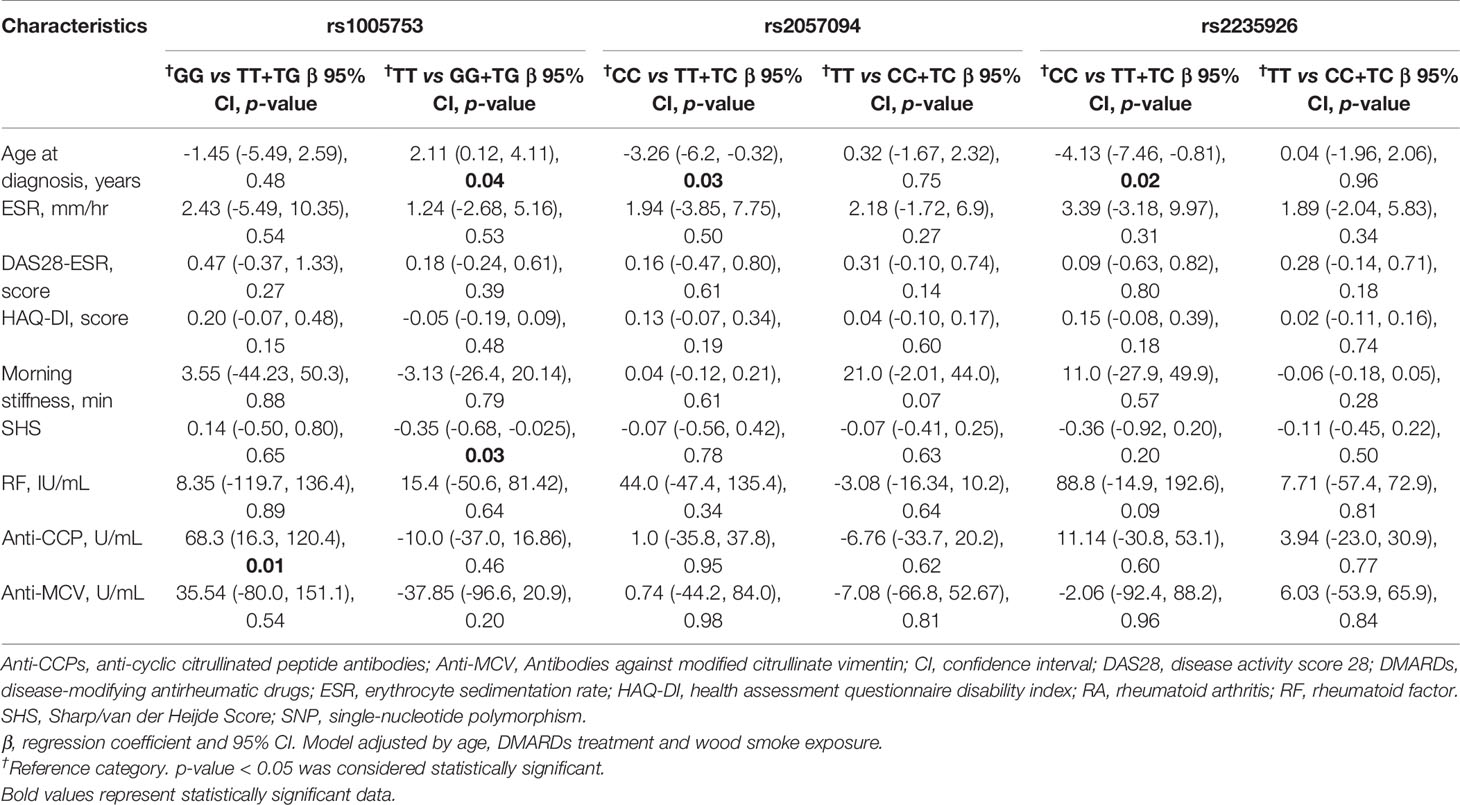
Table 5 Effect of PADI2 SNPs on the clinical characteristics in RA patients.
When analyzing the association between SNPs and the autoantibodies’ serologic status, the carriers of the T allele of SNP rs2057094 exhibited a tendency to associate with the presence of positive RF (OR = 2.5; 95% CI, 0.93-6.6; p = 0.06) and with the positive status for ACPAs and RF (anti-CCP+/anti-MCV+/RF+, OR = 2.23; 95% CI, 0.96-5.18; p = 0.06). In addition, the genotypes TT+TC of rs2235926 were related to the individual positivity to RF (OR = 2.90; 95% CI, 1.02-8.26; p = 0.04), and to anti-MCV (OR = 2.92; 95% CI, 0.96-8.9; p = 0.05), as well as with the positive status for anti-CCP+/anti-MCV+ (OR = 3.02; 95% CI, 1.11-8.23; p = 0.03) and anti-CCP+/anti-MCV+/RF+ (OR = 3.79; 95% CI, 1.51-9.5; p = 0.004) (Table 6).
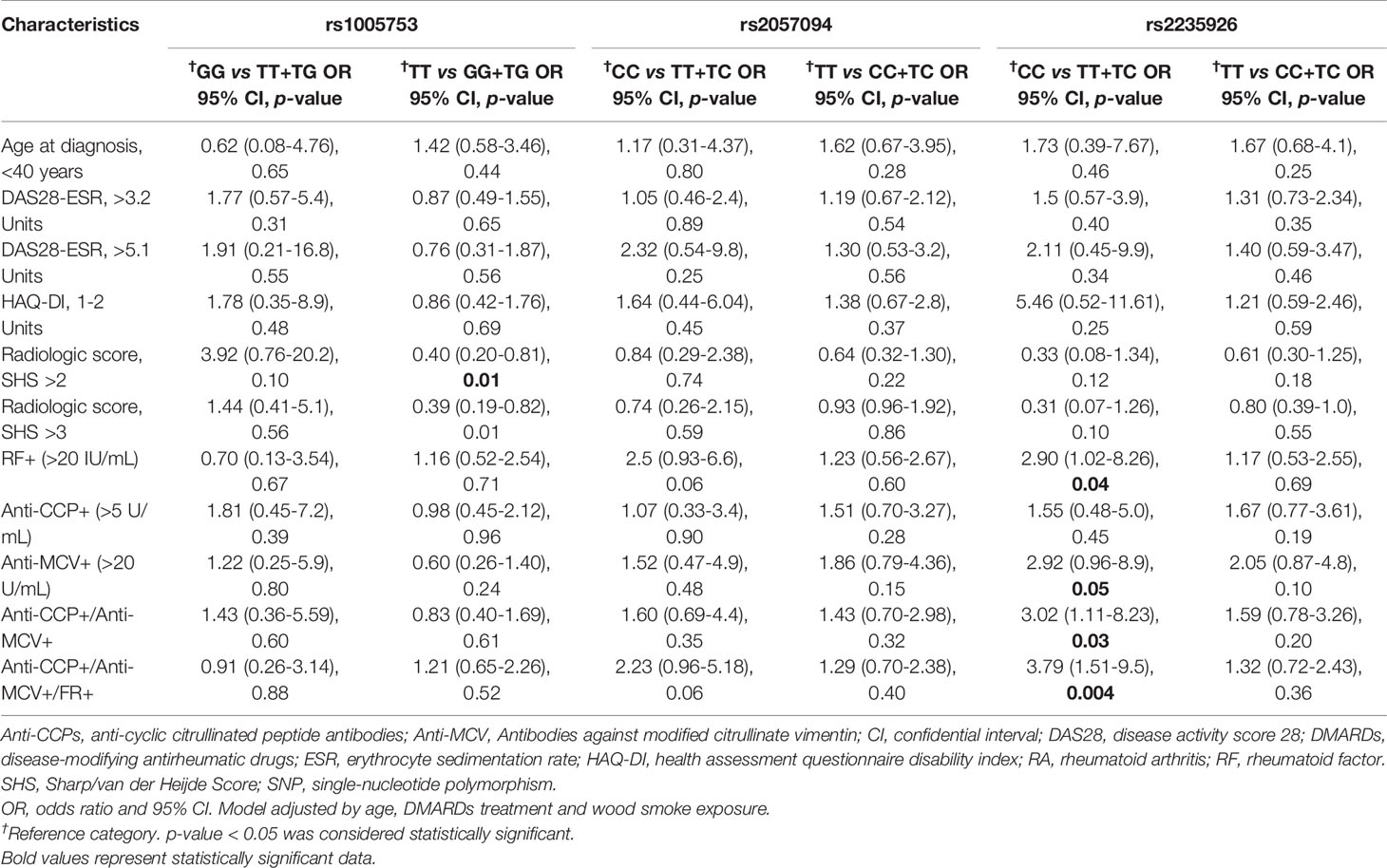
Table 6 Association of PADI2 SNPs on the clinical characteristics in RA patients.
Finally, in the association analysis among haplotypes of PADI2 gene and the clinical characteristics of the disease, the haplotype TTT was significantly related with the presence of joint damage defined by a SHS score ≥ 2 (OR = 1.97; 95% CI, 1.27-3.05; p = 0.002) and SHS ≥3 (OR = 1.94; 95% CI, 1.15-3.19; p = 0.011).
Discussion
Our finds prove the potential role of PADI2 single-nucleotide variants in the clinical heterogeneity of RA in a women population of southern Mexico. Chang et al. studied a Chinese population and proved that SNPs rs2235926 (OR = 1.57; p <0.001) and rs2057094 (OR = 1.36; p = 0.003) conferred RA susceptibility (29). In our study, these SNPs were not associated individually with RA, but they were associated with the presence of high levels of anti-CCPs and a positive status for anti-CCPs+/anti-MCV+/RF+. In the PADI4 gene, the presence of SNPs and a functional haplotype has been described as a factor for genetic susceptibility to RA in different populations, including the Mexican (39–41). In the Mexican population, the GTG haplotype in PADI4 was associated with the RA emergence in ages ≤40 years old and with elevated anti-CCPs levels (40), as well as with anti-MCV antibodies, which are related with the increase of inflammatory cytokines levels and the RA DAS28 score (42). In this study, we found that the T allele of rs2235926 and rs2057094 in PADI2 was associated with the rise of RA in ages younger than 40 years old. Even though the age of onset for RA is variable, in the Mexican population, the highest point of most incidence fluctuates between 56-65 years old (2), similar to what was described in a Chinese population where it fluctuates between 60-70 years of age (43). It has been described that the citrullination process precedes the appearance of the disease’s clinical signs (44, 45), therefore the genetic susceptibility attributed to the PADIs genes involved in the protein citrullination process and of its interaction with other genetic and environmental factors could determine the clinical appearance of RA in ages younger than the largest incidence peak of the disease.
Particularly for our population, the observed seropositivity for ACPAs (>80%), as well as the combined positivity for anti-CCP+/anti-MCV+RF+ (75.1%) is high, especially when compared to that described in other populations where seropositivity oscillates between 38% and 53.9% (42, 46). Anti-CCPs positivity is considered a predictive marker for structural joint damage in RA patients (47), and for higher radiographic and inflammation progression (48–50). In this study, the TTT haplotype of the studied SNPs of the PADI2 gene was associated with a representative score for joint damage, and particularly, the SNP rs2235926 with the ACPAs positivity (anti-CCPs+/anti-MCV+) and ACPAs/RF (anti-CCPs+/anti-MCV+/RF+). Furthermore, the seropositivity to multiple autoantibodies of the ACPAs type and RF isotopes is associated with the radiological progression and erosive RA (51), just as with the high levels of C-reactive protein (CRP) and proinflammatory cytokines in RA patients (13, 52–54). In an in vitro model, it was proven that the RF from isotype IgM favors the increase of the anti-CCPs levels and the cytokine production, thus suggesting that a positive serologic status for RF and anti-CCPs contributes to the pathogenesis of RA (14), as well as to the clinical diversification of the disease (12). On the other hand, the anti-MCV antibodies are considered better predictors of the disease’s severity (55), high clinical activity, and radiographic joint damage and progression (53, 56), given that it increases up to 7.3 times the risk of radiographic progression (10) when compared to anti-CCPs. In our study, the common allele of SNP rs2235926 was associated with the positivity for anti-MCV.
The peptide repertoire that is susceptible to citrullination by PAD2 is more extensive in comparison to PAD4. Assohou-Luty et al. characterized 320 citrullination sites for PAD2 and 178 sites for PAD4, thus proving the specificity variability of these enzymes to their substrate (57). Two main regions have been described in PADs that are involved in the substratum selection, the N-terminal domain, and the cleft of the active site. In PAD4, the Arg-374 contributes to recognizing the substratum and its structural formation, while in PAD2 and PAD3, the amino acid is Gly-374 (58). In this study, we found a relation between positivity to anti-CCPs, anti-MCV and RF, and SNP rs2235926 of PADI2. This polymorphism is found in a 3´untranslated region, however, even though it hasn’t been defined, this could influence the post-transcriptional expression of PAD2 and the citrullination of peptides. Nevertheless, PAD2 can citrullinate transcript factors that determine the Th0 cell lineage differentiation, such as GATA3 and RORγt. Citrullination of R300 of GATA3 and 4 arginine residues (R56, R59, R77, and R90) from RORγt, proves it has an effect on the gene regulation and on cell functions (59). This is important because the presence of polymorphisms in PADI2 could modulate the expression and enzymatic function, therefore relating not only with the serologic positivity to a wide repertoire of antibodies, but also with the activation of naive T cells in response to antigen or “self” antigens, and their subsequent proliferation and differentiation, involved in the pathogenesis and clinical diversification of RA.
Conversely, the enzymatic activity of PADs is mediated by the Ca2+ concentrations. In PAD4 five binding sites for Ca2+ have been identified. The binding of this element to their sites induces conformational changes that create the active site cleft of the enzyme (60), while in PAD2, the Ca2+ binding happens in 6 sites. The activation of the binding sites 3, 4, and 5 control the dependency on Ca2+ and on regulatory elements that include a Ca2+ switch (61). The citrullination by PAD2 and PAD4 requires Ca2+ concentrations in a range of 0.35 to 1.85 mM (30). Vossenar et al. reported that in samples of synovial fluid from RA patients that vimentin can be citrullinated by PAD2 and PAD4 isotypes (28). However, using mass spectrometry, it was found that of 47 sites in fibrinogen that are susceptible to citrullination, 46 of them were citrullinated by PAD2 in vitro (62). Furthermore, PAD2 could have more protein citrullination activity such as fibrin, and also it could increase the proinflammatory cytokines’ expression (63), indicating the leading role of PAD2 in the pathogenesis of RA related not only with the production of ACPAs but also with the modulation of the inflammatory process. We consider that the identification of the susceptibility polymorphisms in PADI2 is important from the clinical viewpoint; however, the clinical perspective of our findings related to the presence of a positive serologic status for multiple autoantibodies and the clinical manifestations of RA at an early onset age deserves replication studies amongst populations.
Citrullination of vimentin in joints is crucial during the pathogenesis of RA (64). In an in vitro model, it was observed that citrullinated vimentin increases the secretion of TNF-α and IL-1, as well as the expression of PAD4 and RANKL (65). Moreover, during the differentiation process from monocyte to macrophage, the mRNA and PAD2 protein levels increase (66, 67). Likewise, it was observed that the expression of the mRNA of PADI2 is high in CD34+ cells and that its expression levels are correlated to the Sp1 transcription factor (68), which, interestingly, along with transcription factor Sp3 has an influence over the PADI2 transcription (69). This proves the important role of PAD2 in the regulation of the expression of proinflammatory cytokines, of genes that promote citrullination, and of differentiation processes and activation of cells involved in joint bone resorption. In this study the haplotype TTT was significantly related with the presence of joint damage defined by a SHS, however other studies are required to clarify the functional role of SNPs in the PADI2 gene on expression of the protein.
It was proven that the expression of PAD2 is elevated in samples of synovial tissue from RA patients (28–31). Damagaard et al. reported that the levels of PAD2 were high in synovial fluid in RA and that these are higher in patients that are positive to anti-CCPs. Additionally, the PAD2 levels in synovial fluid correlated with clinical activity, the levels of CRP, anti-CCPs, and leukocyte count, as well as cytokines such as IL-6, IL-8, and IL-10 (32). In an in vitro model, it was observed that PAD2 could promote IL-1β, IL-6, and TNF-α production in macrophages, and apoptosis induction when activating the caspases 2, 3, and 9, and, at the same time, when activating cell adhesion by FAK, paxillin, and PAK1 ways, which leads to the increase of inflammation (70). Meanwhile, the presence of anti-PAD2 antibodies is associated with a small number of inflamed joints, with a low prevalence of interstitial lung disease, and slower progression of joint damage (71). However, the current study is not exempt from limitations, including the lack of validation in a second population. Moreover, other studies are required to clarify the functional role of SNPs in the PADI2 gene.
In conclusion, in a women population from southern Mexico, the TTT haplotype in the PADI2 gene confers genetic susceptibility to RA and radiographic joint damage related to a positive status to autoantibodies anti-CCP+/anti-MCV+/RF+ and the clinical manifestations of RA at an early onset age.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by CB-004/2017. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
IG-G Designed the study and wrote the manuscript. OZ-G, CR-V, and JN-Z recruited the patients, control subjects, and obtained the samples. IG-P and OZ-G data collection. CR-V, IG-P, and RF-V performed the experiments. MR, NC-A, and IP-R provided relevant opinions in the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We especially thank the patients for participating in the present study. Thanks to the group of collaborators in the multidisciplinary team who contributed with their knowledge and support in the management and integral approach to the patients.
References
1. Tobón GJ, Youinou P, Saraux A. The Environment, Geo-Epidemiology, and Autoimmune Disease: Rheumatoid Arthritis. J Autoimmun (2010) 35(1):10–4. doi: 10.1016/j.jaut.2009.12.009
2. Moreno-Montoya J, Alvarez-Nemegyei J, Sanin LH, Pérez-Barbosa L, Trejo-Valdivia B, Santana N, et al. Association of Regional and Cultural Factors With the Prevalence of Rheumatoid Arthritis in the Mexican Population: A Multilevel Analysis. J Clin Rheumatol (2015) 21(2):57–62. doi: 10.1097/RHU.0000000000000223
3. McInnes IB, Schett G. The Pathogenesis of Rheumatoid Arthritis. N Engl J Med (2011) 365(23):2205–19. doi: 10.1056/NEJMra1004965
4. Pukšič S, Morovič-Vergles J. Comorbidities in Patients With Rheumatoid Arthritis. Reumatizam (2014) 61(2):31–6.
5. Yamada R, Suzuki A, Chang X, Yamamoto K. Citrullinated Proteins in Rheumatoid Arthritis. Front Biosci (2005) 10:54–64. doi: 10.2741/1506
6. Darrah E, Rosen A, Giles JT, Andrade F. Peptidylargine Deiminase 2, 3 and 4 Have Distinct Specificities Against Cellular Substrates: Novel Insights Into Autoantigen Selection in Rheumatoid Arthritis. Ann Rheum Dis (2012) 71(1):92–8. doi: 10.1136/ard.2011.151712
7. Romero V, Darrah E, Andrade F. Generation of Distinct Patterns of Rheumatoid Arthritis Autoantigens by Peptidylarginine Deiminase Types 2 and 4 During Perforin-Induced Cell Damage. Arthritis Rheumatol (2020) 72(6):912–8. doi: 10.1002/art.41196
8. Curran AM, Naik P, Giles JT, Darrah E. PAD Enzymes in Rheumatoid Arthritis: Pathogenic Effectors and Autoimmune Targets. Nat Rev Rheumatol (2020) 16(6):301–15. doi: 10.1038/s41584-020-0409-1
9. Syversen SW, Gaarder PI, Goll GL, Ødegård S, Haavardsholm EA, Mowinckel P, et al. High Anti-Cyclic Citrullinated Peptide Levels and an Algorithm of Four Variables Predict Radiographic Progression in Patients With Rheumatoid Arthritis: Results From a 10-Year Longitudinal Study. Ann Rheum Dis (2008) 67(2):212–7. doi: 10.1136/ard.2006.068247
10. Syversen SW, Goll GL, van der Heijde D, Landewé R, Lie BA, Odegård S, et al. Prediction of Radiographic Progression in Rheumatoid Arthritis and the Role of Antibodies Against Mutated Citrullinated Vimentin: Results From a 10-Year Prospective Study. Ann Rheum Dis (2010) 69(2):345–51. doi: 10.1136/ard.2009.113092
11. Scott DL. Prognostic Factors in Early Rheumatoid Arthritis. Rheumatol (Oxford) (2000) 39(Suppl 1):24–9. doi: 10.1093/oxfordjournals.rheumatology.a031490
12. Katchamart W, Koolvisoot A, Aromdee E, Chiowchanwesawakit P, Muengchan C. Associations of Rheumatoid Factor and Anti-Citrullinated Peptide Antibody With Disease Progression and Treatment Outcomes in Patients With Rheumatoid Arthritis. Rheumatol Int (2015) 35(10):1693–9. doi: 10.1007/s00296-015-3271-8
13. Sokolove J, Bromberg R, Deane KD, Lahey LJ, Derber LA, Chandra PE, et al. Autoantibody Epitope Spreading in the Pre-Clinical Phase Predicts Progression to Rheumatoid Arthritis. PLoS ONE (2012) 7(5):e35296. doi: 10.1371/journal.pone.0035296
14. Sokolove J, Johnson DS, Lahey LJ, Wagner CA, Cheng D, Thiele GM, et al. Rheumatoid Factor as a Potentiator of Anti-Citrullinated Protein Antibody-Mediated Inflammation in Rheumatoid Arthritis. Arthritis Rheumatol (2014) 66(4):813–21. doi: 10.1002/art.38307
15. Foulquier C, Sebbag M, Clavel C, Chapuy-Regaud S, Al Badine R, Méchin MC, et al. Peptidyl Arginine Deiminase Type 2 (PAD-2) and PAD-4 But Not PAD-1, PAD-3, and PAD-6 are Expressed in Rheumatoid Arthritis Synovium in Close Association With Tissue Inflammation. Arthritis Rheum (2007) 56(11):3541–53. doi: 10.1002/art.22983
16. Kinloch A, Lundberg K, Wait R, Wegner N, Lim NH, Zendman AJ, et al. Synovial Fluid is a Site of Citrullination of Autoantigens in Inflammatory Arthritis. Arthritis Rheum (2008) 58(8):2287–95. doi: 10.1002/art.23618
17. Beato M, Sharma P. Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer. Int J Mol Sci (2020) 21(4):1351. doi: 10.3390/ijms21041351
18. Lamensa JW, Moscarello MA. Deimination of Human Myelin Basic Protein by a Peptidylarginine Deiminase From Bovine Brain. J Nuerochem (1993) 61(3):987–96. doi: 10.1111/j.1471-4159.1993.tb03612.x
19. Zhang X, Bolt M, Guertin MJ, Chen W, Zhang S, Cherrington BD, et al. Peptidylarginine Deiminase 2-Catalyzed Histone H3 Arginine 26 Citrullination Facilitates Estrogen Receptor α Target Gene Activation. Proc Natl Acad Sci USA (2012) 109(33):13331–6. doi: 10.1073/pnas.1203280109
20. Hsu PC, Liao YF, Lin CL, Lin WH, Liu GY, Hung HC. Vimentin is Involved in Peptidylarginine Deiminase 2-Induced Apoptosis of Activated Jurkat Cells. Mol Cells (2014) 37(5):426–34. doi: 10.14348/molcells.2014.2359
21. Kim SE, Park JW, Kim MJ, Jang B, Jeon YC, Kim HJ, et al. Accumulation of Citrullinated Glial Fibrillary Acidic Protein in a Mouse Model of Bile Duct Ligation-Induced Hepatic Fibrosis. PLoS ONE (2018) 13(8):e0201744. doi: 10.1371/journal.pone.0201744
22. Kan R, Jin M, Subramanian V, Causey CP, Thompson PR, Coonrod SA. Potential Role for PADI-Mediated Histone Citrullination in Preimplantation Development. BMC Dev Biol (2012) 12:19. doi: 10.1186/1471-213X-12-19
23. Tanikawa C, Espinosa M, Suzuki A, Masuda K, Yamamoto K, Tsuchiya E, et al. Regulation of Histone Modification and Chromatin Structure by the P53-PADI4 Pathway. Nat Commun (2012) 3:676. doi: 10.1038/ncomms1676
24. Hagiwara T, Nakashima K, Hirano H, Senshu T, Yamada M. Deimination of Arginine Residues in Nucleophosmin/B23 and Histones in HL-60 Granulocytes. Biochem Biophys Res Commun (2002) 290(3):979–83. doi: 10.1006/bbrc.2001.6303
25. Lee YH, Coonrod SA, Kraus WL, Jelinek MA, Stallcup MR. Regulation of Coactivator Complex Assembly and Function by Protein Arginine Methylation and Demethylimination. Proc Natl Acad Sci USA (2005) 102(10):3611–6. doi: 10.1073/pnas.0407159102
26. Li P, Yao H, Zhang Z, Li M, Luo Y, Thompson PR, et al. Regulation of P53 Target Gene Expression by Peptidylarginine Deiminase 4. Mol Cell Biol (2008) 28(15):4745–58. doi: 10.1128/MCB.01747-07
27. Guo Q, Fast W. Citrullination of Inhibitor of Growth 4 (ING4) by Peptidylarginine Deiminase 4 (PAD4) Disrupts the Interaction Between ING4 and P53. J Biol Chem (2011) 286(19):17069–78. doi: 10.1074/jbc.M111.230961
28. Vossenaar ER, Radstake TR, van der Heijden A, van Mansum MA, Dieteren C, de Rooij DJ, et al. Expression and Activity of Citrullinating Peptidylarginine Deiminase Enzymes in Monocytes and Macrophages. Ann Rheum Dis (2004) 63(4):373–81. doi: 10.1136/ard.2003.012211
29. Chang X, Xia Y, Pan J, Meng Q, Zhao Y, Yan X. PADI2 is Significantly Associated With Rheumatoid Arthritis. PLoS ONE (2013) 8(12):e81259. doi: 10.1371/journal.pone.0081259
30. Damgaard D, Senolt L, Nielsen MF, Prujin GJ, Nielsen CH. Demonstration of Extracellular Peptidylarginine Deiminase (PAD) Activity in Synovial Fluid of Patients With Rheumatoid Arthritis Using a Novel Assay for Citrullination of Fibrinogen. Arthritis Res Ther (2014) 16(6):498. doi: 10.1186/s13075-014-0498-9
31. Olivares-Martínez E, Hernández-Ramírez DF, Núñez-Álvarez CA, Cabral AR, Llorente L. The Amount of Citrullinated Proteins in Synovial Tissue is Related to Serum Anti-Cyclic Citrullinated Peptide (Anti-CCP) Antibody Levels. Clin Rheumatol (2016) 35(1):55–61. doi: 10.1007/s10067-015-3047-2
32. Damgaard D, Senolt L, Nielsen CH. Increased Levels of Peptidylarginine Deiminase 2 in Synovial Fluid From Anti-CCP-Positive Rheumatoid Arthritis Patients: Association With Disease Activity and Inflammatory Markers. Rheumatol (Oxford) (2016) 55(5):918–27. doi: 10.1093/rheumatology/kev440
33. Lee YH, Bae SC, Choi SJ, Ji JD, Song GG. Genome-Wide Pathway Analysis of Genome-Wide Association Studies on Systemic Lupus Erythematosus and Rheumatoid Arthritis. Mol Biol Rep (2012) 39(12):10627–35. doi: 10.1007/s11033-012-1952-x
34. Freudenberg J, Lee HS, Han BG, Shin HD, Kang YM, Sung YK, et al. Genome-Wide Association Study of Rheumatoid Arthritis in Koreans: Population-Specific Loci as Well as Overlap With European Susceptibility Loci. Arthritis Rheum (2011) 63(4):884–93. doi: 10.1002/art.30235
35. Too CL, Murad S, Dhaliwal JS, Larsson P, Jiang X, Ding B, et al. Polymorphism in Peptidylarginine Deiminase Associate With Rheumatoid Arthritis in Diverse Asian Populations: Evidence From MyEIRA Study and Meta-Analysis. Arthritis Res Ther (2012) 14(6):R250. doi: 10.1186/ar4093
36. Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham CO, et al. 2010 Rheumatoid Arthritis Classification Criteria: An American College of Rheumatology/European League Against Rheumatism Collaborative Initiative. Arthritis Rheum (2010) 62(9):2569–81. doi: 10.1002/art.27584
37. vand der Heijde D. How to Read Radiographs According to the Sharp/van Der Heijde Method. J Rheumatol (2000) 27(1):261–3.
38. Shi YY, He L. SHEsis, a Powerful Software Platform for Analyses of Linkage Disequilibrium, Haplotype Construction, and Genetic Association at Polymorphism Loci. Cell Res (2005) 15(2):97–8. doi: 10.1038/sj.cr.7290272
39. Zavala-Cerna MG, Gonzalez-Montoya NG, Nava A, Gamez-Nava JI, Moran-Moguel MC, Rosales-Gomez RC, et al. PADI4 Haplotypes in Association With RA Mexican Patients, a New Prospect for Antigen Modulation. Clin Dev Immunol (2013) 2013:383681. doi: 10.1155/2013/383681
40. Guzmán Guzmán IP, Reyes-Castillo Z, Muñoz-Barrioss S, Ruiz-Noa Y, Martínez-Bonilla GE, Parra-Rojas I, et al. Polymorphisms and Functional Haplotype in PADI4: Further Evidence for Contribution on Rheumatoid Arthritis Susceptibility and Anti-Cyclic Citrullinated Peptide Antibodies in a Western Mexican Population. Immunol Lett (2015) 163(2):214–20. doi: 10.1016/j.imlet.2014.10.029
41. Baños-Hernández CJ, Navarro-Zarza JE, Parra-Rojas I, Vázquez-Villamar M, Padilla-Gutiérrez JR, Valle Y, et al. PADI4 Polymorphisms and the Functional Haplotype are Associated With Increased Rheumatoid Arthritis Susceptibility: A Replication Study in a Southern Mexican Population. Hum Immunol (2017) 78(9):553–8. doi: 10.1016/j.humimm.2017.05.005
42. Reyes-Castillo Z, Palafox-Sánchez CA, Parra-Rojas I, Martínez-Bonilla GE, del Toro-Arreola S, Ramírez-Dueñas MG, et al. Comparative Analysis of Antibodies Targeting Peptidylarginine Deiminase Type 4, Mutated Citrullinated Vimentin and Cyclic Citrullinated Peptides in Rheumatoid Arthritis: Associations With Cytokine Profiles, Clinical and Genetic Features. Clin Exp Immunol (2015) 182(2):119–31. doi: 10.1111/cei.12677
43. Zhang Q, Liu Q, Lin C, Baima Y, Li H, Gong H, et al. The Prevalence of Rheumatoid Arthritis in Middle-Aged and Elderly People Living in Naqu City, Tibet, Autonomous Region of China. J Orthop Surg Res (2020) 15(1):338. doi: 10.1186/s13018-020-01883-4
44. Kokkonen H, Mullazehi M, Berglin E, Hallmans G, Wadel G, Rönnelid J, et al. Antibodies of IgG, IgA and IgM Isotypes Against Cyclic Citrullinated Peptide Precede the Development of Rheumatoid Arthritis. Arthritis Res Ther (2011) 13(1):R13. doi: 10.1186/ar3237
45. Svärd A, Kastbom A, Söderlin MK, Reckner-Olsson Å, Skogh T. A Comparison Between IgG- and IgA-Class Antibodies to Cyclic Citrullinated Peptides and to Modified Citrullinated Vimentin in Early Rheumatoid Arthritis and Very Early Arthritis. J Rheumatol (2011) 38(7):1265–72. doi: 10.3899/jrheum.101086
46. González-Lopez L, Rocha-Muñoz AD, Ponce-Guarneros M, Flores-Chavez A, Salazar-Paramo M, Nava A, et al. Anti-Cyclic Citrullinated Peptide (Anti-CCP) and Anti-Mutated Citrullinated Vimentin (Anti-MCV) Relation With Extra-Articular Manifestations in Rheumatoid Arthritis. J Immunol Res (2014) 2014:536050. doi: 10.1155/2014/536050
47. Kim HH, Kim J, Park SH, Kim SK, Kim OD, Cho JY. Correlation of Anti-Cyclic Citrullinated Antibody With Hand Joint Erosion Score in Rheumatoid Arthritis Patients. Korean J Intern Med (2010) 25(2):201–6. doi: 10.3904/kjim.2010.25.2.201
48. Kirino Y, Hama M, Takase-Minegishi K, Kunishita Y, Kishimoto D, Yoshimi R, et al. Predicting Joint Destruction in Rheumatoid Arthritis With Power Doppler, Anti-Citrullinated Peptide Antibody, and Joint Swelling. Mod Rheumatol (2015) 25(6):842–8. doi: 10.3109/14397595.2015.1026025
49. Hetland ML, Stengaard-Pedersen K, Junker P, Østergaard M, Ejbjerg BJ, Jacobsen S, et al. Radiographic Progression and Remission Rates in Early Rheumatoid Arthritis-MRI Bone Oedema and Anti-CCP Predicted Radiographic Progression in the 5-Year Extension of the Double-Blind Randomized CIMESTRA Trial. Ann Rheum Dis (2010) 69(10):1789–95. doi: 10.1136/ard.2009.125534
50. Joo YB, Park YJ, Park KS, Kim KJ. Association of Cumulative Anti-Cyclic Citrullinated Protein Antibodies With Radiographic Progression in Patients With Rheumatoid Arthritis. Clin Rheumatol (2019) 38(9):2423–32. doi: 10.1007/s10067-019-04554-w
51. Vencovský J, Machácek S, Sedová L, Kafková J, Gatterová J, Pesákova V, et al. Autoantibodies can be Prognostic Markers O Fan Erosive Disease in Early Rheumatoid Arthritis. Ann Rheum Dis (2003) 62(5):427–30. doi: 10.1136/ard.62.5.427
52. Lu MC, Lai NS, Yu HC, Huang HB, Hsieh SC, Yu CL. Anti-Citrullinated Protein Antibodies Bind Surface-Expressed Citrullinated Grp78 on Monocyte/Macrophages and Stimulate Tumor Necrosis Factor Alpha Production. Arthritis Rheum (2010) 62(5):1213–23. doi: 10.1002/art.27386
53. Mansour HE, Metwaly KM, Hassan IA, Elshamy HA, Elbeblawy MM. Antibodies to Mutated Citrullinated Vimentin in Rheumatoid Arthritis: Diagnostic Value, Association With Radiological Damage and Axial Skeleton Affection. Clin Med Insights Arthritis Musculoskelet Disord (2010) 3:33–42. doi: 10.4137/cmamd.s4827
54. Salafí F, Carotti M, Ciapetti A, Gasparini S, Filippucci E, Grassi W. Relationship Between Time-Integrate Disease Activity Estimated by DAS28-CRP and Radiographic Progression of Anatomical Damage in Patients With Early Rheumatoid Arthritis. BMC Musculoskelet Disord (2011) 12:120. doi: 10.1186/1471-2474-12-120
55. Jilani AA, Mackworth-Young CG. The Role of Citrullinated Protein Antibodies in Redicting Erosive Disease in Rheumatoid Arthritis: A Systematic Literature Review and Meta-Analysis. Int J Rheumatol (2015) 2015:728610. doi: 10.1155/2015/728610
56. Barouta G, Katsiari CG, Alexiou I, Liaskos C, Varna A, Bogdanos DP, et al. Anti-MCV Antibodies Predict Radiographic Progression in Greek Patients With Very Early (<3 Months Duration) Rheumatoid Arthritis. Clin Rheumatol (2017) 36(4):885–94. doi: 10.1007/s10067-016-3494-4
57. Assohou-Luty C, Raijmakers R, Benckhuijsen WE, Stammen-Vogelzangs J, de Ru A, vann Veelen PA, et al. The Human Peptidylarginine Deiminases Type 2 and Type 4 Have Distinct Substrate Specificities. Biochim Biophys Acta (2014) 1844(4):829–36. doi: 10.1016/j.bbapap.2014.02.019
58. Arita K, Shimizu T, Hashimoto H, Hidaka Y, Yamada M, Sato M. Structural Basis for Histone N-Terminal Recognition by Human Peptidylarginine Deiminase 4. Proc Natl Acad Sci USA (2006) 103(14):5291–6. doi: 10.1073/pnas.0509639103
59. Sun B, Chang HH, Salinger A, Tomita B, Bawadekar M, Holmes CL, et al. Reciprocal Regulation of Th2 and Th17 Cells by PAD2-Mediated Citrullination. JCI Insight (2019) 4(22):e129687. doi: 10.1172/jci.insight.129687
60. Arita K, Hashimoto H, Shimizu T, Nakashima K, Yamada M, Sato M. Structural Basis for Ca(2+)-Induced Activation of Human PAD4. Nat Struct Mol Biol (2004) 11(8):777–83. doi: 10.1038/nsmb799
61. Slade DJ, Fang P, Dreyton CJ, Zhang Y, Fuhrmann J, Rempel D, et al. Protein Arginine Deiminase 2 Binds Calcium in an Ordered Fashion: Implications for Inhibitor Design. ACS Chem Biol (2015) 10(4):1043–53. doi: 10.1021/cb500933j
62. Sharma M, Damgaard D, Senolt L, Svensson B, Bay-Jensen AC, Nielsen CH, et al. Expanding the Citrullinome of Synovial Fibrinogen From Rheumatoid Arthritis Patients. J Proteomics (2019) 208:103484. doi: 10.1016/j.jprot.2019.103484
63. Sanchez-Pernaute O, Filkova M, Gabucio A, Klein M, Maciejewska-Rodrigues H, Ospelt C, et al. Citrullination Enhances the Pro-Inflammatory Response to Fibrin in Rheumatoid Arthritis Synovial Fibroblasts. Ann Rheum Dis (2013) 72(8):1400–6. doi: 10.1136/annrheumdis-2012-201906
64. Steendam KV, Tilleman K, Deforce D. The Relevance of Citrullinated Vimentin in the Production of Antibodies Against Citrullinated Proteins and the Pathogenesis of Rheumatoid Arthritis. Rheumatol (Oxford) (2011) 50(5):830–7. doi: 10.1093/rheumatology/keq419
65. Fan LY, He DY, Wang Q, Zong M, Zhang H, Yang H, et al. Citrullinated Vimentin Stimulates Proliferation, Proinflammatory Cytokine Secretion, and PADI4 and RANKL Expression of Fibroblast-Like Synoviocytes in Rheumatoid Arthritis. Scand J Rheumatol (2012) 41(5):354–8. doi: 10.3109/03009742.2012.670263
66. Hojo-Nakashima I, Sato R, Nakashima K, Hagiwara T, Yamada M. Dynamic Expression of Peptidylarginine Deiminase 2 in Human Monocytic Leukaemia THP-1 Cells During Macrophage Differentiation. J Biochem (2009) 146(4):471–9. doi: 10.1093/jb/mvp097
67. Lai NS, Yu HC, Tung CH, Huang KY, Huang HB, Lu MC. Increased Peptidylarginine Deiminases Expression During the Macrophage Differentiation and Participated Inflammatory Responses. Arthritis Res Ther (2019) 21(1):108. doi: 10.1186/s13075-019-1896-9
68. Nagia T, Matsueda Y, Tomita T, Yoshikawa H, Hirohata S. The Expression of mRNA for Peptidylarginine Deiminase Type 2 and Type 4 in Bone Marrow CD34+ Cells in Rheumatoid Arthritis. Clin Exp Rheumatol (2018) 36(2):248–53.
69. Dong S, Kojima T, Shiraiwa M, Méchin MC, Chavanas S, Serre G, et al. Regulation of the Expression of Peptidylarginine Deiminase Type II Gene (PADI2) in Human Keratinocytes Involves Sp1 and Sp3 Transcription Factors. J Invest Dermatol (2005) 124(5):1026–33. doi: 10.1111/j.0022-202X.2005.23690.x
70. Yu HC, Tung CH, Huang KY, Huang HB, Lu MC. The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage. Int J Mol Sci (2020) 21(16):5720. doi: 10.3390/ijms21165720
Keywords: PADI2, polymorphisms, autoantibodies, radiologic damage, rheumatoid arthritis
Citation: Guzmán-Guzmán IP, Ramírez-Vélez CI, Falfán-Valencia R, Navarro-Zarza JE, Gutiérrez-Pérez IA, Zaragoza-García O, Ramírez M, Castro-Alarcón N and Parra-Rojas I (2021) PADI2 Polymorphisms Are Significantly Associated With Rheumatoid Arthritis, Autoantibodies Serologic Status and Joint Damage in Women from Southern Mexico. Front. Immunol. 12:718246. doi: 10.3389/fimmu.2021.718246
Received: 31 May 2021; Accepted: 19 July 2021;
Published: 04 August 2021.
Edited by:
Andrea Picchianti-Diamanti, Sapienza University, ItalyReviewed by:
Erika Darrah, Johns Hopkins University, United StatesChristelle Darrieutort-Laffite, University of Pennsylvania, United States
Copyright © 2021 Guzmán-Guzmán, Ramírez-Vélez, Falfán-Valencia, Navarro-Zarza, Gutiérrez-Pérez, Zaragoza-García, Ramírez, Castro-Alarcón and Parra-Rojas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Iris Paola Guzmán-Guzmán, cGFvX25raWxsZXJAeWFob28uY29tLm14