 Fátima Ferragut
Fátima Ferragut Gonzalo R. Acevedo
Gonzalo R. Acevedo Karina A. Gómez
Karina A. Gómez- Laboratorio de Inmunología de las Infecciones por Tripanosomátidos, Instituto de Investigaciones en Ingeniería Genética y Biología Molecular (INGEBI), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Buenos Aires, Argentina
The CD4+ and CD8+ T cell immune response against T. cruzi, the parasite causing Chagas disease, are relevant for both parasite control and disease pathogenesis. Several studies have been focused on their phenotype and functionally, but only a few have drilled down to identify the parasite proteins that are processed and presented to these cells, especially to CD4+ T lymphocytes. Although approximately 10,000 proteins are encoded per haploid T. cruzi genome, fewer than 200 T cell epitopes from 49 T. cruzi proteins have been identified so far. In this context, a detailed knowledge of the specific targets of T cell memory response emerges as a prime tool for the conceptualization and development of prophylactic or therapeutic vaccines, an approach with great potential to prevent and treat this chronic disease. Here, we review the available information about this topic in a comprehensive manner and discuss the future challenges in the field.
Introduction
Chagas disease, caused by the infection with the protozoan Trypanosoma cruzi, is a neglected tropical disease from the American continent that has spread from the limits established by vector ecology due to human migration (1) to non-endemic places such Canada, USA, Europe, Australia and Japan (2). Last estimates calculate that about 6-8 million people are infected in the world (1), with more than 70 million people living in areas at risk for infection (3).
The parasite can be transmitted through other routes than vectorial spread and its life cycle includes different intermediate hosts and developmental stages (Figure 1). When vectorially acquired, Chagas disease shows two major phases. The acute phase typically lasts for about 2 months and presents a high number of parasites circulating in the blood, but in most cases symptoms are absent, mild or unspecified (4) -for instance prolonged fever, headache, myalgia, lymphadenitis, hepatomegaly, and splenomegaly-. However, visible signs of infection such as skin lesion (chagoma) or a swollen eyelid (the so-called Romaña sign) can help in the diagnosis of new cases (1). If patients are not treated, a chronic phase follows, during which most people remain asymptomatic but infected for life. Nonetheless, it is estimated that up to 40% of chronically infected patients can develop organ involvement, being cardiomyopathy and megaviscera (megaoesophagus, megacolon, or both) the most prevailing (5). Based on these phases, some diagnostic tests are better suited than others. In the acute phase, the high parasitemia makes it possible to identify the presence of circulating parasites by the direct microscopic visualization of blood samples or by polymerase chain reaction (PCR)-based diagnostics methods. In contrast, serologic testing is preferred in the chronic phase when parasitemia is low. Since no single standard reference test is available, diagnosis should be based on the presence of IgG against various T. cruzi antigens by the use of at least two serological assays based on different principles, such as indirect fluorescent assay, indirect hemagglutination, and ELISA. And, a third assay is then indicated to clarify infection status when serology results are discordant and samples can yield persistent inconclusive results (5).
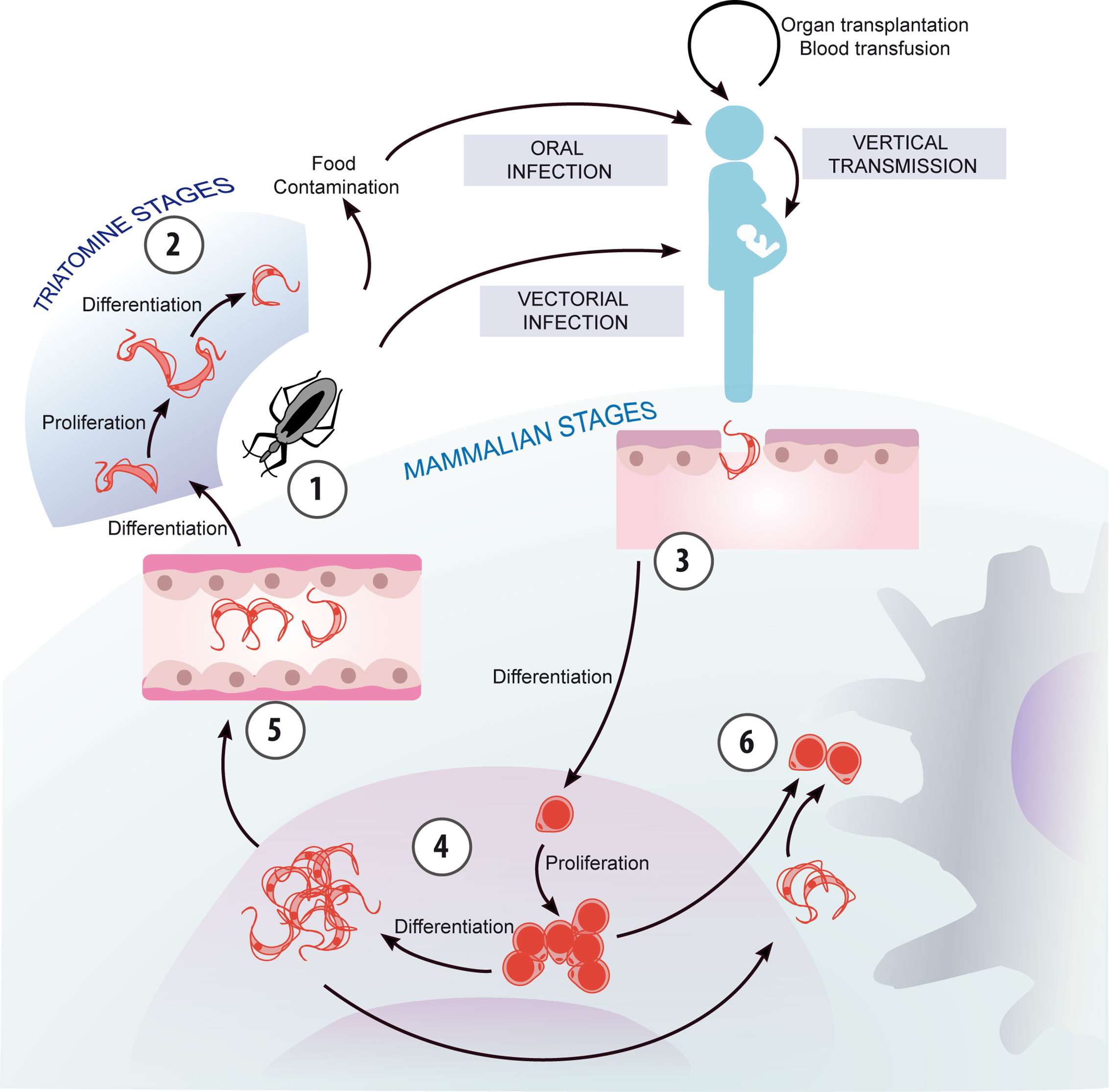
Figure 1 Chagas disease transmission and parasite life cycle. The life cycle of T. cruzi can be conceptualized with its beginning at moment when the triatomine vector ingests bloodstream trypomastigotes from an infected mammalian host, which differentiate into replicative epimastigotes (1). Epimastigotes migrate to the hindgut and differentiate into non-replicative, infectious metacyclic trypomastigotes (2), which are excreted with the feces of the vector and are ready to infect a new mammalian host. Additionally, T. cruzi can be transmitted through other routes than vectorial spread, such as mother-to-child and oral transmission besides blood transfusion and organ transplantation. Trypomastigotes invade host cells and differentiate into the intracellular, replicative amastigote form (3), which can also transform into trypomastigotes (4). These are released when the host cells break down, and may invade new cells, migrate through the bloodstream, or be ingested by a vector (5). In addition, extracellular amastigotes, originated by the premature rupture of the host cell or by extracellular trypomastigote differentiation, can be engulfed by macrophages (6).
With regard to Chagas disease treatment, two drugs -benznidazole and nifurtimox- are currently available. Despite their effectiveness in the acute phase, there is no consensus for their use in the chronic phase. Furthermore, the drugs prolonged duration and the side effects, which include gastric intolerance, skin rashes or neuromuscular problems, can lead to treatment discontinuation and failure (6, 7). Thus, the recent clinical trial BENDITA (Benznidazole New Doses Improved Treatment & Therapeutic Associations) aimed to find effective regimes that produce fewer adverse effects and improve patients´ adherence to treatment (8). This work demonstrated the efficacy of the treatment protocol duration of 2 weeks with a daily dose of 300 mg/day of benznidazole after the 12-month follow-up (8). In summary, in chronic phase, generally a symptomatic supportive treatment is performed. Chagas disease patients with cardiomyopathy are treated with specific medication to control cardiac damage or with an implantable cardioverter defibrillator (9). Heart transplantation is the only course of action in case of advanced tissue failure, which is an expensive procedure, limited by the lack of donors and with a risk of reactivation of parasitaemia due to the use of immunosuppressive drugs. In patients with digestive involvement, conservative or even surgical treatment is indicated depending on the stage of the disease (2).
Specific T Cell Response in Chagas Disease
Upon infection with the parasite, mammal hosts develop both innate and adaptive immune responses that play a major role during the acute and chronic phases of the disease (10). Regarding T cells, CD8+ cytotoxic T lymphocytes have been predominantly studied (11), given that the replicative phase of T. cruzi life cycle within mammals host takes place in the intracellular environment (Figure 2). The activation of these T cells results in the acquisition of the molecular machinery for the elimination of the target cell, which includes cytoplasmic granules containing proteins such as perforin and granzymes and cytokine production, like IFN-γ, TNF-α and IL-2. In chronic Chagas disease, several types of dysfunction have been described. Thus, concomitant with terminal differentiation, CD8+ T cells from patients with severe disease present a higher frequency of cells coexpressing inhibitory receptors, and a lower frequency of polyfunctional parasite-specific CD8+ T cells (11). Besides its role in the elimination of the parasite, some evidence suggests that CD8+ T cells are involved in tissue damage and inflammatory processes linked to the clinical manifestations of Chagas disease (10).
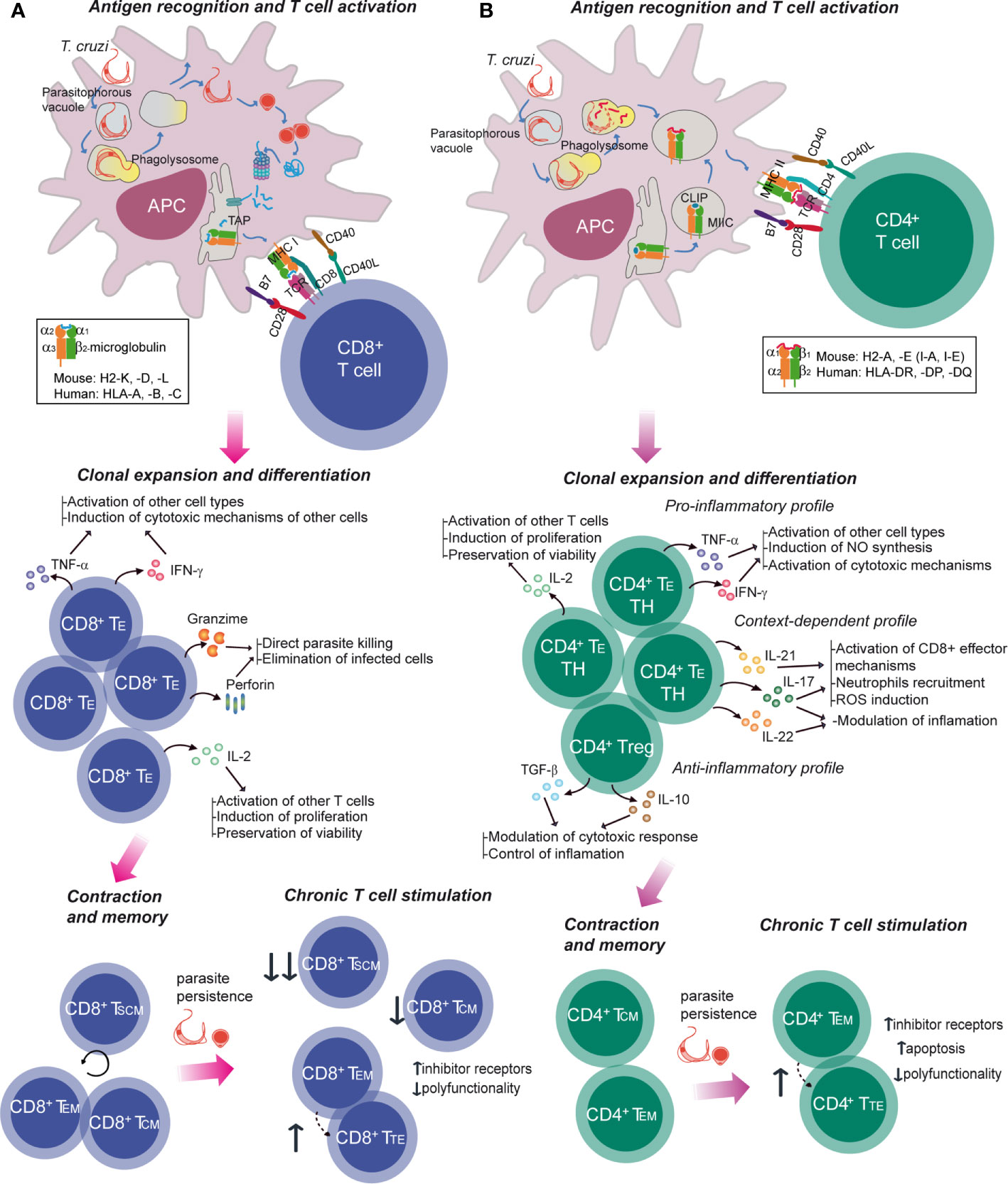
Figure 2 Specific T cell response in Chagas disease. The activation of T-cell immune responses is mediated by APCs, which deliver antigen-specific and costimulatory signals. Trypanosoma cruzi trypomastigote invades the host cell or is passively internalized, leading to the formation of the parasitophorous vacuole which fuses with a lysosome resulting in a phagolysosome. The parasite can escape to the cytoplasm (A) where it differentiates into the amastigote form and its proteins are targeted for proteosomal degradation. Peptides generated are then imported into the endoplasmic reticulum (ER) to be presented to CD8+ T cells in the context of MHC class I molecules. These molecules consists of a β2-microglobulin and a polymorphic alpha chain which contains the peptide binding groove and it is encoded by one of the mouse H2-K, H2-D and H2-L class I MHC genes and human HLA-A, -B or –C class I MHC genes. (B) Class II MHC is made up of polymorphic α and β chains that form the peptide binding groove, each encoding in mouse H2-A and H2-E class II MHC loci and in human HLA-DR, -DP and -DQ class II MHC loci. In the class II MHC pathway, proteins from parasite are degraded and antigenic peptides are loaded onto the binding cleft from the MHC. Peptide-MHC complexes are then presented in the host cell surface to the CD4+ T cell. In both CD8+ and CD4+ T cells, activation generates the expansion and differentiation into effector specific T cells (TE), their survival and function being sustained by cytokines and soluble mediators. Following antigen clearance, contraction takes place with a numerical reduction of antigen-specific cells. A subset of the activated T cells survives beyond this phase and differentiates into long-lasting memory T cells. During chronic infections, which entail persistent antigen exposure and/or inflammation, different degrees of dysfunctionality can be observed, including terminal differentiation and T cell exhaustion with progressive loss of effector functions and co-expression of inhibitor receptors. IL, interleukin; IFN, interferon; TNF, tumor necrosis factor; TH, T helper; Treg, regulatory T cell; TSCM, stem cell memory T cell.
Much less is known about CD4+ T cells involvement (Figure 2), possibly because their ultimate action requires the collaboration with other cells (10). These cells are characterized by the expression of surface molecules and the secretion of cytokines that modulate the activity of other cells, mainly macrophages, DCs and other lymphocytes. The majority of proliferating activated CD4+ T cells differentiate into one of the multiple TH profiles, acquiring a diversity of effector properties. It has been suggested that a coordinated response between TH1 and TH2 profiles is desirable, with predominance of TH1 effector mechanisms for the control/elimination of the parasite (10). In addition, the subset of regulatory T cells (Treg), which operate tolerance mechanisms, is increased in asymptomatic patients compared to those with cardiac or digestive symptoms. Furthermore, CD4+ T cells develop some signatures of T cell response exhaustion in the chronic phase of the disease (10).
The specific responses of both types of T cells depend on the processing and presentation of T. cruzi peptides by antigen presenting cells (APC) and their recognition by the T cell receptor (TCR). For this reason, studies on new therapeutic approaches (12–15) attempt to understand the mechanisms underlying the interaction between peptide-MHC (pMHC, MHC: major histocompatibility complex) complexes and TCR, and also functional and phenotypic characteristics of antigen-specific T cells, which can be investigated by diverse strategies (16–23). In Chagas disease, different T. cruzi targets that can be processed and presented to T cells have been evaluated. Extensive research is mainly focused on members of the T. cruzi trans-sialidase (TS) superfamily, the largest T. cruzi gene family. These proteins are distributed along the cell body, flagellum and flagellar pocket of the parasite. This superfamily includes not only active trans-sialidases that transfer sialic acid from host glycoconjugates to the parasite membrane proteins, but also several proteins with unknown function, among others. In addition, non-trans-sialidase (non-TS) T cell activating antigens are also analyzed.
Taking into consideration the central role that T cell response plays in Chagas disease and the potential use of antigen-specific T cell response data in disease prevention or treatment, this review revisits the results of published research on the specific response against T. cruzi epitopes recognized by CD8+ and CD4+ T cells. To this end, reports about a variety of T. cruzi antigens are discussed, with particular emphasis on peptides that trigger T cell response in mouse models or human samples. Additionally, we comment on what we consider the greatest challenges faced by scientists today in the study of T-cell specificity in the immune response against this parasite.
Models for Chagas Disease Specific T Cell Response
Despite the translational challenges of animal models in Chagas disease (24, 25), they are still crucial for increasing the knowledge of the disease and the interaction between the host and the parasite (26–29). Indeed, mouse models (wild type mice with H2 –mouse MHC- molecules or transgenic ones expressing human MHC) are broadly utilized under different strategies to evaluate the specificity of T cell response against T. cruzi infection (Figure 3). Importantly, human samples, especially blood, are often used to validate results obtained from animal models and to analyze the particularities of human T cell responses against the parasite (Figure 3). In this regard, the HLA (Human Leukocyte Antigen, human MHC molecules) restriction ultimately determines the repertoire of peptides that are presented for interaction with the TCR, limiting the usefulness of non-human models to evaluate T cell specificity at the epitope level.
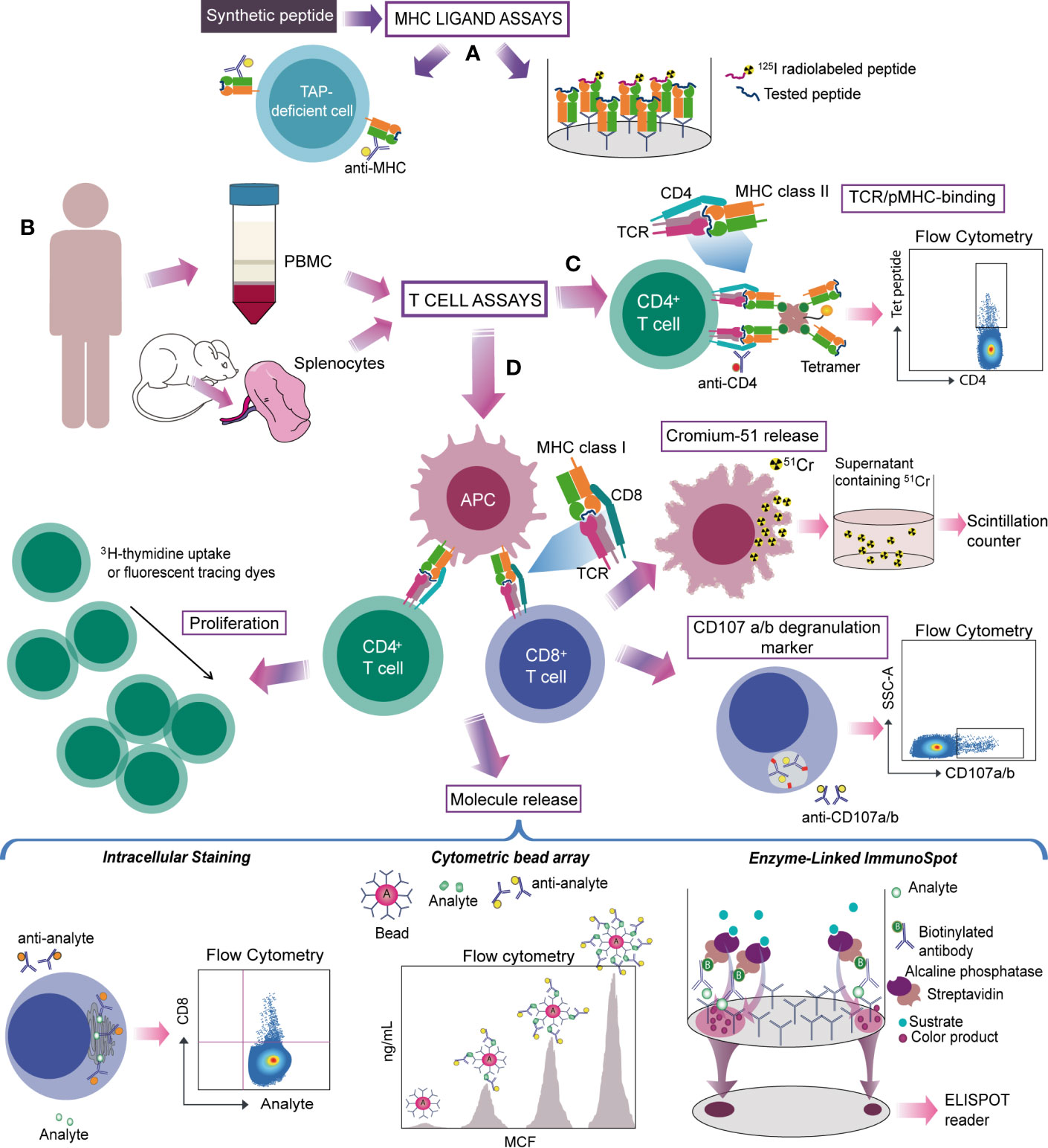
Figure 3 Strategies for studying antigen-specific T cells in Chagas disease. (A) Experimentally identified or predicted candidate target peptides are synthesized and tested by MHC ligand assays. MHC-peptide binding can be evaluated by TAP deficient cells (cells lacking the transporter associated with antigen-processing that fail to transport peptides into the endoplasmic reticulum, reducing MHC class I stability and surface expression) loaded with the peptide and stained for the MHC, or by the ability to inhibit the binding of a radiolabeled probe peptide. (B) Splenocytes from mouse, PBMC from human or CD4+- or CD8+-depleted PBMC are used to detect antigen-specific T cells trough T cell assays. (C) MHC-peptide multimer/tetramer is based on the ability of TCR to recognize the pMHC complex from CD8+ or CD4+ T cells. (D) Other strategies evaluate the specific T-cell response on the basis of a functional readout. Activated cytotoxic CD8+ T cells are commonly detected by measuring chromium in the supernatant released by killed target cells. Assays that determine secretion of cytolytic granule contents (granzyme or perforin) and/or labelling responding cells with antibodies to CD107a and b provide an alternative way to measure cytolytic activity. On CD4+ T cells activation, the amplification of antigen-specific populations is tested through proliferation assays. Molecule release (cytokines or cytolytic granule contents) from activated CD8+ or CD4+ T cells can be evaluated by different methods. Intracellular staining of induced molecules detects specific T-cells on a single cell level whereas CBA measures secreted proteins produced by bulk cell populations. ELISPOT detects secretion “spots” of a specific mediator and quantifies the magnitude of this response. Monoclonal antibodies for phenotyping can be used to characterize specific T cells.
CD8+ T Cell Specificity in Chagas Disease
Several T CD8+ epitopes have been described and confirmed as immunogens in Chagas disease patients, being those derived from the trans-sialidase proteins family widely assessed (30–33). Most studies focus on MHC class I presentation restricted to HLA-A*02 alleles because of its high prevalence in individuals of various ethnic backgrounds, including those living in areas where Chagas disease is endemic (34, 35).
Within the TS family, immunogenic CD8+ T cell epitopes identified in HLA-A*02:01+ patients belong predominantly to trypomastigote surface antigen (TSA)-1, amastigote surface proteins (ASP)-1 and -2 and flagellum-associated protein FL-160 (Table 1). The first studies on the role of these antigens in Chagas disease were performed in mouse models and showed protective responses against T. cruzi infection after vaccination with antigen-encoding plasmids (53–55). Additionally, CD8+ cytotoxic T cells epitopes from those proteins (Table 2), restricted to the mouse H-2Kb class I molecule, sensitized RMA-S target cells (a TAP deficient cell line derived from the Rauscher leukemia virus-induced mouse T-cell lymphoma RBL-5) for lysis by specific spleen cells from T. cruzi-infected mice (55–57). The antigen-specific CD8+ cytotoxic T cells response was also demonstrated in T. cruzi-infected or peptide-immunized A2.1/Kb transgenic mice (expressing a chimeric class I molecule consisting of the human HLA-A2.1 α1 and α2 domains ligated to the mouse α3, transmembrane and cytoplasmic H2-Kb domains). Splenocytes from these animals were submitted to cytotoxicity assays, in which Jurkat A2.1/Kb or lymphoblastoid 721.221 A2.1/Kb targets cells were sensitized with TSA-1-, ASP-1- and ASP-2-derived peptides (36). Furthermore, specific CD8+ T cell response against peptides from these antigens has been also observed in peripheral blood mononuclear cells (PBMC) from HLA-A*02:01+ donors with chronic T. cruzi infection (32, 36) (Table 1). Interestingly, the cytotoxic activity induced by selected epitopes from ASP-1, ASP-2 and TSA-1 was reduced by the depletion of CD8+ T cells, but not CD4+ T cells, in samples from individuals living in endemic regions in Guatemala (36). Additionally, assays using MHC tetramers bound to some of those peptides demonstrated a low frequency of peptide-specific CD8+ T cells in HLA-A*02:01+ T. cruzi-infected patients residing in non-endemic areas of Argentina at the time of sample collection (32).
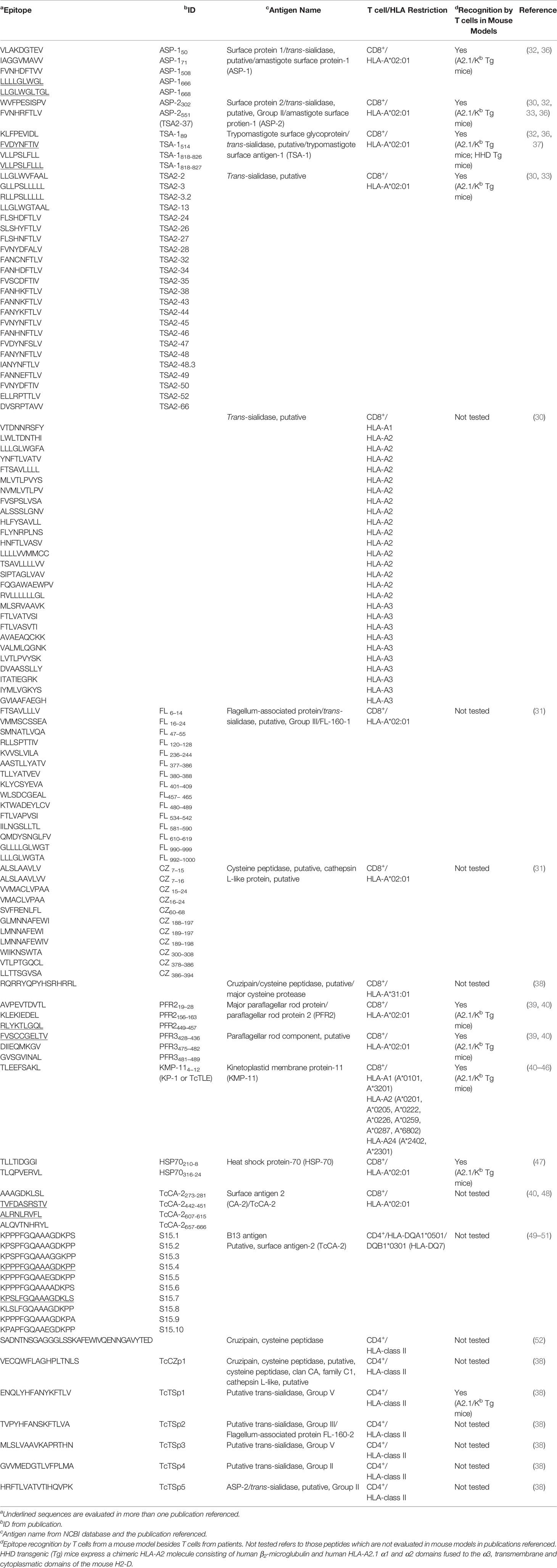
Table 1 T. cruzi epitopes recognized by T cells from Chagas disease patients.
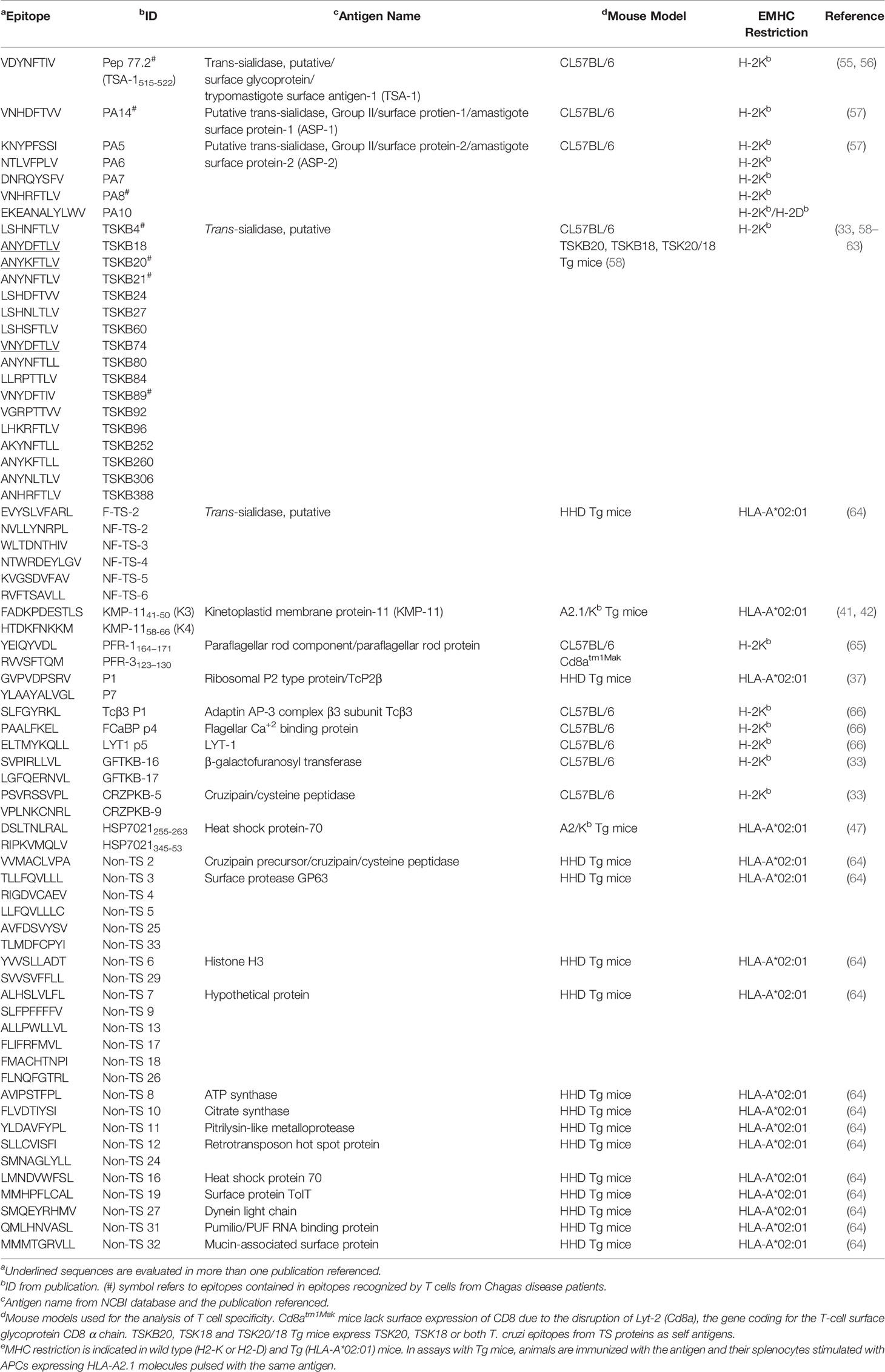
Table 2 T. cruzi epitopes recognized by CD8+ T cell from mice (not tested in Chagas disease patients).
Studies based on peptides predicted to bind H-2Kb MHC molecules showed a CD8+ T cell response that is dominated by epitopes encoded by trans-sialidase family genes (33) (Table 2). Frequencies of IFN-γ-producing cells were lower against non-TS epitopes, and target splenocytes from naïve mice loaded with non-TS epitopes were poorly recognized in in vivo cytotoxic T lymphocyte assays (33). Similarly, the immunization of C57BL/6 mice with pools of trans-sialidase but not mucin genes provided protection against an otherwise lethal T. cruzi challenge (66). Moreover, the CD8+ T cell response focused on a few epitopes (33, 59–61) was interpreted in favor of the hypothesis of immunodominance against certain peptides as mechanism to evade the immune response (62). Nonetheless, experimental approaches with mouse models later demonstrated that the immune control of T. cruzi infection was independent of the recognition of the therein called “dominant” TSKB20 and “subdominant” TSKB18 epitopes (58, 63). In fact, despite the widespread idea of TS broad dominance of the T cell response against T. cruzi, there is insufficient evidence of a correlate in the context of human infection.
A similar approach to that described for the prediction and selection of H-2Kb–binding peptides allowed the determination of TS peptides (Table 1) that induced diverse levels of reactivity in IFN-γ ELISPOT experiments for HLA-A*02:01+ chronic Chagas disease patients (33). Interestingly, many TS peptides (Table 1), which were found by in silico prediction of their binding affinity to MHC from different HLA allele supertypes (HLA-A1, -A2, -A3, -A24, -B7 and -B44), showed a positive outcome in IFN-γ ELISPOT assays, being A2 supertype peptides the most frequently recognized (30).
Additional CD8+ T cell targets were identified from enzymatically active TS (F-TS), non-functional TS (NF-TS) and non-TS proteins (Table 2) after vaccination of HHD transgenic mice with dendritic cells pulsed with peptide pools (64). Furthermore, HLA-A*02:01-restricted epitopes from cruzipain (also known as GP 57/51 or cruzain, a cysteine peptidase) and the FL-160 protein (renamed FL-160-1, the 160 kDa flagellum-associated surface protein of trypomastigotes) were described (31). Peptides from those proteins (Table 1) were recognized by PBMC from chronic Chagas disease patients in IFN-γ ELISPOT and tetramer assays (31). Another cruzipain epitope was described in a recent publication of our group using an in silico guided approach (38). Sequences of T. cruzi proteins characterized as T cell immunogens in the Immune Epitope Database [IEDB-, www.iedb.org (67)] were screened and the binding to a set of HLA-A and HLA-B or HLA-DRB1 molecules were predicted. One out of the four novel peptides reported (namely TcCZp2, Table 1), revealed predominant CD8+ T cell activation in IFN-γ ELISPOT assays with PBMC depleted of CD4+ T cells (38). In addition, the predicted nested core of this peptide (TcCZp2me) allowed the identification of epitope-specific CD8+ T cells in HLA-A*31:01+ patients, with a predominant effector memory TEMRA (terminal effector T cells re-expressing CD45RA) or TEM (effector memory) cell phenotype (38).
Potential CD8+ T cells targets were also described in the paraflagellar rod proteins (known as PFR or TcPRP), located at the T. cruzi flagellum and specific to the kinetoplastids. Mice receiving injections of these proteins (namely PAR1 or PFR-3 and PAR2) had an immune response capable of reducing the level of circulating parasites, and improved survival (68). Another research on the subject showed that cytotoxic T lymphocyte lines generated from chronically T. cruzi-infected mice specific for peptides PFR-1164−171 and PFR-3123–130 (Table 2) had high levels of lytic activity and secreted IFN-γ in response to parasite-infected target cells. Moreover, CD8 knockout (Cd8atm1Mak) mice deficient in functional cytotoxic T cells, immunized with PFR and challenged with T. cruzi, showed higher parasitemia and lower survival than PFR-immunized control mice (65). DNA immunization with the PFR2/heat shock protein 70 (HSP70) fused genes not only triggered the cytotoxic activity of antigen-specific CD8+ T cells but also provided a protective response against a T. cruzi infection (69). Furthermore, the immunization of C57BL/6-A2/Kb transgenic mice with plasmids encoding the fusion proteins PFR2-HSP70 and PFR3-HSP70 induced a specific CD8+ cytotoxic T cells response against epitopes PFR2449–457 and PFR3481–489 (39). Regarding the response in the human host, peptides containing potential HLA-A*02:01-binding sites were identified by the analysis of T. cruzi PFR2 and PFR3 using the algorithms SYFPEITHI [www.syfpeithi.de, (70)] and BIMAS [www-bimas.cit.nih.gov, (71)] (39). Peptides from each protein (Table 1) were recognized by PBMC from chronic Chagas disease patients and peptide-specific CD8+ T cells exhibited a pro-inflammatory cytokine secretion profile (IFN-γ, TNF-α and IL-6). Remarkably, cytotoxic activity was observed in CD8+ T cells from asymptomatic patients but not in chronic patients with cardiomyopathy (39). Interestingly, some epitopes from PFR and other paraflagellar rod component proteins were described in a recent in silico study based on the identification of conserved epitopes (72).
Another vastly described T. cruzi protein containing CD8+ T cell epitopes is the kinetoplastid membrane protein-11 (KMP-11), which has been reported to be mainly located in the flagellar pocket. The first investigations into CD8+ T cell response against this antigen were carried out in mouse models. Immunization of A2.1/Kb transgenic mice with fused T. cruzi KMP-11/HSP70 genes generated a population of reactive CD8+ cytotoxic T cells against KMP-11 peptides (named K1, K3 and K4) (Table 2) containing predicted HLA-A*02:01 binding motifs (41, 42). With regard to humans, HLA-A*02:01+ chronic Chagas disease patients from endemic zones exerted a CD8+ T cell specific response towards the TcTLE peptide [previously known as K1 peptide, Table 1, (43)]. This peptide was also recognized by infected patients carrying HLA-A2 supertype alleles other than HLA-A*02:01 (A*0205, A*0222, A*0226, A*0259 and A*0287). Both HLA-A*02:01+ and HLA-A*02:01- infected patients had TcTLE-specific CD8+ T cells displaying a phenotype compatible with differentiated TEM cells (44). Moreover, the introduction of specific mutations in TcTLE showed limited effect on the functional activity of CD8+ T cells from HLA-A*02:01+ chronic Chagas disease patients (45). TcTLE also exhibited a promiscuous recognition in patients with Chagas disease expressing different HLA-A supertypes (HLA-A1, -A2 and -A24). Remarkably, the evaluation of IFN-γ, TNF-α, IL-2, perforin, GrB and CD107a/b after stimulation with this peptide demonstrated that HLA-A2- patients had mono- and polyfunctional (simultaneously expressing multiple cytokines and exhibiting cytotoxic activity at the single-cell level) CD8+ T cells and their frequency could be associated with disease severity (46). Similar results had been previously described when PBMC from patients at different stages of chronic Chagas disease were stimulated with KMP-11 recombinant protein. In fact, patients with less severe disease showed a higher frequency of polyfunctional KMP11-specific CD8+ T cells whereas patients at advanced stages of the disease demonstrated a higher frequency of monofunctional CD8+ T cells (73).
Class-I epitopes recognized by patient cells were identified in other T. cruzi proteins, such as HSP70 and the putative surface antigen TcCA-2. Two T. cruzi HSP70 CD8+ epitopes were shown to be processed and presented during natural infection. Peptides predicted with the algorithms SYFPEITHI and RANKPEP [http://immunax.dfci.harvard.edu/Tools/rankpep.html (74),] were recognized by cells from A2.1/Kb transgenic animals immunized with the HSP70 recombinant protein or infected with the parasite. Two of those peptides (HSP70210-8 and HSP70316-24,Table 1) recognized by CD8+ T cells of HLA-A*02:01+ Chagas disease patients from endemic areas residing in a non-endemic zone induced peptide-specific cytotoxic activity and pro-inflammatory cytokine (IFN-γ and TNF-α) secretion (47). With regard to the T. cruzi carbonic anhydrase protein TcCA-2, four CD8+ epitopes (TcCA-2273-281, TcCA-2442-451, TcCA-2607-615 and TcCA-2657-666) (Table 1) were found to be processed and presented during Chagas disease in a multiplexed cytokine secretion assay (48). CD8+ T cell specific for these epitopes were functionally active in both asymptomatic and symptomatic cardiomyopathy chronic Chagas disease patients (from endemic areas but residing in a non-endemic zone) (48). In addition, TcCA-2442-451 and TcCA-2607-615 peptides allowed the characterization of different phenotypical profiles in cells from asymptomatic versus cardiac Chagas disease patients. Thus, the TcCA-2-specific CD8+ T cells from patients with cardiac symptoms were mainly effector memory cells (TEM or TEMRA) whereas those present in the asymptomatic phase were predominantly naïve T cells (48). In a recent study, the same two peptides from TcCA-2, and those from PFR2 (PFR2449-457), PFR3 (PFR3428-436) and KMP-11 (K1) (Table 1), were used to evaluate the antigen-specific CD8+ T cell response in asymptomatic chronic Chagas disease patients, both before and after benznidazole treatment (40). Although different outcomes for memory phenotype, senescence and antigen experience were found depending on the peptide analyzed, CD8+ T cells specific to all the peptides showed a high percentage of these cells at an advanced stage of differentiation (CD45RA+CD127-) both before and after treatment (40). In a prior report, the same group had characterized the functional capacity of antigen-specific CD8+ T cells (based on the expression of IFN-γ, IL-2, TNF-α, granzyme B and perforin) before and after drug treatment but, in this case, they used recombinant proteins instead of peptides from those T. cruzi antigens (75). Patients showing therapeutic efficacy (based on serological response) presented polyfunctional antigen-specific CD8+ T cells whereas cells from patients with therapeutic failure were monofunctional (75).
Other targets for the CD8+ T-cell response were described in mouse models, but without confirmed immunogenicity in humans (Table 2). The sequences for flagellar Ca2+ binding protein (FCaBP), LYT-1 and AP-3 were scanned for peptides that matched the mouse H-2Kb allele-specific class I peptide binding motif. LYT-1, a protein predicted to be secreted by trypomastigotes, was found to be a target of cytotoxic T cell responses in chronically infected mice, and the immunization with a plasmid encoding a peptide from this protein protected mice from lethal T. cruzi challenge (66). Peptides from FCaBP and a homologue of the adaptin AP-3 complex β3 subunit (Tcβ3) were recognized by spleen cells from infected mice, and these lysed peptide-pulsed RMA-S target cells (66). With regard to ribosomal P2β protein (TcP2β), two peptides (TcP2-P7 and TcP2-P1, Table 2), containing known binding motifs for MHC molecule encoded by the HLA-A*02:01, elicited lytic activity when HHD transgenic mice were immunized with them individually (37).
CD4+ T Cell Specificity in Chagas Disease
Only a few proteins have been demonstrated to induce a recall response on CD4+ T cells from T. cruzi infected-individuals in the chronic phase of the disease (38, 49, 52, 76, 77).
Cruzipain elicited proliferative responses of PBMC from chronic Chagas disease patients both in its recombinant and isolated (from T. cruzi epimastigotes) forms. Moreover, T cell lines generated from PBMC using those antigens consisted of CD4+ T cells, and their stimulation with recombinant or isolated cruzipain induced their production of IFN-γ. Furthermore, anti-cruzipain T cell lines showed a strong proliferative response against one out 11 peptides spanning portions of the catalytic domain of the cruzipain protein [Table 1 (52)].
The recognition of the tandemly repetitive T. cruzi B13 protein (a homolog of TcCA-2) was also characterized by T cell lines, clones and PBMC from chronic Chagas disease patients (49). The PBMC proliferative response to recombinant B13 protein was similar between cardiac and asymptomatic patients and among non-infected control individuals. Likewise, when the proliferative response to B13-derived peptides (Table 1) was analyzed in HLA-DQ7+ subjects, peptide S15.4 was mainly recognized by patients with cardiomyopathy whereas S15.1 and S15.3 peptides more frequently targeted by cells from asymptomatic donors, and peptide S15.9 triggered a response in cells from both groups of Chagas disease patients (49). Additionally, a CD4+ T cell clone from an HLA-DQ7+ individual responder to the peptide S15.4 presented cross-reactivity against peptides from human β-cardiac myosin heavy chain according to the central HLA-DQ7 binding motif (50). Interestingly, proliferation assays on PBMC from S15.7-responder HLA-DQ7+ individuals revealed that peptide S15.7 was recognized by PBMC from all subjects tested whereas none bound to its highly resistant to proteolytic digestion analogues (51).
Concerning KMP-11, a specific CD4+ T cell response was shown as INF-γ production upon the stimulation of PBMC from chronic Chagas disease patients with the recombinant antigen (76). KMP-11 responders were mostly patients with severe cardiomyopathy (abnormal ECG and cardiomegaly in chest radiography) whereas only one subject was determined as responder out of eight evaluated donors in patients with normal findings on ECG, or abnormal ECG and no cardiac enlargement (76).
On the other hand, TSA-1 and Tc24 (flagellar calcium binding protein of 24 kDa) were evaluated for the ability to recall CD4+ T cell response in chronic Chagas disease patients (77). Results demonstrated a higher proportion of CD4+ T cells that proliferated after stimulation with rTc24, whereas CD3+ T cells responded when rTSA-1 was used as stimulus (77).
A recent in silico approach also enabled the identification of potential T. cruzi-specific CD4+ epitopes from a co-chaperone GrpE and from a hypothetical protein (72). And more recently, another predictive approach developed by our group led to the discovery of peptides demonstrated to induce CD4+ T cell response in Chagas disease patients (38). In fact, these cells were the leading producers of IFN-γ in response to the novel T. cruzi peptides TcTSp3, TcTSp4 and TcTSp5 (Table 1). Notably, the IFN-γ responses against TcTSp1, TcTSp2 and TcCZp1 (Table 1) were also predominantly attributed to CD4+ T cell activation, although they comprised sequences from cruzipain and TS proteins previously reported to induce CD8+ T cell response in Chagas disease or to bind HLA-A*02:01 (31, 33). Regarding phenotype characterization, CD4+ T cells showed heterogeneous degree of maturation between chronic Chagas´ disease patient population, although central memory (TCM) T cells seemed to be preponderant in patients with IFN-γ response against TcTSp4 (38).
Concluding Remarks and Future Perspectives
Experimental animal models and studies with clinical samples have shown that CD4+ and CD8+ T cells play a crucial role in the control of Chagas disease. For this reason, elucidating their specificity in relation with their functional features represents an opportunity for a deeper understanding of the immune response against T. cruzi infection. Up to now, almost 200 T cell epitopes from 49 T. cruzi antigens have been described (IEDB, accessed February 2021) whereas the haploid T. cruzi genome has more than 10,000 protein coding genes (78). Thus, continued efforts are required to explore the role of the majority of T. cruzi proteome in the host immune response.
Taking into account the disadvantages of treatments with current drugs (benznidazole and nifurtimox), which have an unclear efficacy at chronic stage (79–81), the identification of T. cruzi antigens could contribute not only to discover putative biomarkers for the follow-up of Chagas disease patients, but also to develop vaccines based on the induction of antigen-specific T cells. These could be an attractive and cost effective alternative (or complement) to current drug-based therapies. Therefore, it is crucial to consider the high biological complexity of T. cruzi and the huge diversity of HLA haplotypes, the major factors determining the repertoire of an individual´s T cell specificities. Targeting as many antigens as possible in a vaccine design could provide a way to overcome the limitations imposed by host diversity.
In this regard, in silico analyses to identify proteins conserved across diverse T. cruzi strains and expressed in stages of the parasite life cycle relevant to human infection, constitute useful approaches to narrow down the universe of T. cruzi antigens to a set of candidates addressable by laboratory scale immunoassays. However, these methods come with caveats and pitfalls of their own. Up to date, reports about antigen-specific T cell response against T. cruzi infection have focused on predicting peptide binding to MHC. Despite this prediction is very useful to limit the number of potential T cell epitopes to test experimentally, not all MHC presented peptides are immunogenic. Consequently, the TCR:pMHC interaction should be better understood in order predict MHC restricted peptides that actually become T cell epitopes. Remarkably, this issue may result challenging due to the promiscuity of TCRs (82), the limited information about the patterns that govern T-cell recognition and the ability of TCRs to cross-recognize structurally related elements (83). In general, extensive data on peptides-MHC binding is available, whereas there is less information linking specific TCRs to their cognate target (84). In this context, different approaches have been proposed to gain knowledge on TCR:pMHC interaction (84–89). Nonetheless, none of them has so far been used to analyze specific T cell response in Chagas disease.
In parallel with immunobioinformatics methods which have powered the search for epitope-based vaccines candidates against T. cruzi (72, 90), other promising and newer tools may be leveraged to detect MHC-bound peptides relevant to the parasite-specific T cell response. These include pipelines used for neoantigen discovery (91) and immunopeptidomics approaches (92–94), which consist in the isolation of peptides from the pMHC complexes purified from the APC surface and their identification by mass spectrometry and bioinformatics.
Taken together, our review highlights the importance of the specific T cell response in Chagas disease. Relevant information about peptides involved in this response has been accumulating across decades of research. Nonetheless, the available data is so far insufficient to effectively enable the translation of this knowledge into treatment strategies for this neglected disease. In this regard, attempts to successfully reach optimal solutions for the millions of people affected are still needed and emerging techniques employed in the study of other diseases are highly promising to this end.
Author Contributions
FF, GA, and KG contributed to conception of the revision. FF organized the data and wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET; PIP Number 112-2015010-0547).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
ASP, amastigote surface protein; APC, antigen presenting cell; CZ, cruzipain; ELISPOT, enzyme-linked immunospot; FCaBP, flagellar Ca2+ binding protein; FL-160, 160 kDa flagellum-associated surface protein; HLA, human leukocyte antigen; HSP70, heat shock protein 70; IEDB, Immune Epitope Database; KMP-11, kinetoplastid membrane protein-11; MHC, major histocompatibility complex; PBMC, peripheral blood mononuclear cell; PFR or PAR, paraflagellar rod protein; pMHC, peptide-MHC complex; TCR, T cell receptor; Tc24, flagellar calcium binding protein of 24 kDa; TcCA-2, surface antigen-2; TEM, effector memory T cell; TEMRA, terminal effector T cells re-expressing CD45RA; TH, helper T cell; Treg, regulatory T cell; Tscm, stem cell memory T cell; TS, trans-sialidase; TSA, trypomastigote surface antigen.
References
1. World Health Organization. Integrating Neglected Tropical Diseases in Global Health and Development. In: Fourth WHO Report on Neglected Tropical Diseases (2017).
2. Lidani KCF, Andrade FA, Bavia L, Damasceno FS, Beltrame MH, Messias-Reason IJ, et al. Chagas Disease: From Discovery to a Worldwide Health Problem. Front Public Health (2019) 7:166. doi: 10.3389/fpubh.2019.00166
3. PAHO. Estimación Cuantitativa De La Enfermedad De Chagas En Las Americas. Panamerican Health Organization (2006). Ops/Hdm/Cd/425-06.
5. Perez-Molina JA, Molina I. Chagas Disease. Lancet (2018) 391(10115):82–94. doi: 10.1016/S0140-6736(17)31612-4
6. Aldasoro E, Posada E, Requena-Mendez A, Calvo-Cano A, Serret N, Casellas A, et al. What to Expect and When: Benznidazole Toxicity in Chronic Chagas’ Disease Treatment. J Antimicrob Chemother (2018) 73(4):1060–7. doi: 10.1093/jac/dkx516
7. Jackson Y, Alirol E, Getaz L, Wolff H, Combescure C, Chappuis F. Tolerance and Safety of Nifurtimox in Patients With Chronic Chagas Disease. Clin Infect Dis (2010) 51(10):e69–75. doi: 10.1086/656917
8. DNDi. The BENDITA Study: A Phase II Study to Assess Safety, Tolerability, and Efficacy of Different Benznidazole Regimens, Alone and in Combination With Fosravuconazole (2019). Available at: https://www.dndial.org/wp-content/uploads/2019/03/2page_BenditatStudyOverview_ENG.pdf.
9. Muratore CA, Batista Sa LA, Chiale PA, Eloy R, Tentori MC, Escudero J, et al. Implantable Cardioverter Defibrillators and Chagas’ Disease: Results of the ICD Registry Latin America. Europace (2009) 11(2):164–8. doi: 10.1093/europace/eun325
10. Acevedo GR, Girard MC, Gomez KA. The Unsolved Jigsaw Puzzle of the Immune Response in Chagas Disease. Front Immunol (2018) 9:1929. doi: 10.3389/fimmu.2018.01929
11. Acosta Rodriguez EV, Araujo Furlan CL, Fiocca Vernengo F, Montes CL, Gruppi A. Understanding CD8(+) T Cell Immunity to Trypanosoma Cruzi and How to Improve it. Trends Parasitol (2019) 35(11):899–917. doi: 10.1016/j.pt.2019.08.006
12. June CH, O’Connor RS, Kawalekar OU, Ghassemi S, Milone MC. Car T Cell Immunotherapy for Human Cancer. Science (2018) 359(6382):1361–5. doi: 10.1126/science.aar6711
13. Kar T, Narsaria U, Basak S, Deb D, Castiglione F, Mueller DM, et al. A Candidate Multi-Epitope Vaccine Against SARS-Cov-2. Sci Rep (2020) 10(1):10895. doi: 10.1038/s41598-020-67749-1
14. Met O, Jensen KM, Chamberlain CA, Donia M, Svane IM. Principles of Adoptive T Cell Therapy in Cancer. Semin Immunopathol (2019) 41(1):49–58. doi: 10.1007/s00281-018-0703-z
15. Zawawi A, Forman R, Smith H, Mair I, Jibril M, Albaqshi MH, et al. In Silico Design of a T-Cell Epitope Vaccine Candidate for Parasitic Helminth Infection. PloS Pathog (2020) 16(3):e1008243. doi: 10.1371/journal.ppat.1008243
16. Bentzen AK, Hadrup SR. Evolution of MHC-Based Technologies Used for Detection of Antigen-Responsive T Cells. Cancer Immunol Immunother (2017) 66(5):657–66. doi: 10.1007/s00262-017-1971-5
17. Bentzen AK, Marquard AM, Lyngaa R, Saini SK, Ramskov S, Donia M, et al. Large-Scale Detection of Antigen-Specific T Cells Using Peptide-MHC-I Multimers Labeled With DNA Barcodes. Nat Biotechnol (2016) 34(10):1037–45. doi: 10.1038/nbt.3662
18. Betts MR, Koup RA. Detection of T-Cell Degranulation: CD107a and B. Methods Cell Biol (2004) 75:497–512. doi: 10.1016/s0091-679x(04)75020-7
19. Grifoni A, Weiskopf D, Ramirez SI, Mateus J, Dan JM, Moderbacher CR, et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans With COVID-19 Disease and Unexposed Individuals. Cell (2020) 181(7):1489–501:e1415. doi: 10.1016/j.cell.2020.05.015
20. Hadrup SR, Newell EW. Determining T-Cell Specificity to Understand and Treat Disease. Nat BioMed Eng (2017) 1(10):784–95. doi: 10.1038/s41551-017-0143-4
21. Herati RS, Muselman A, Vella L, Bengsch B, Parkhouse K, Del Alcazar D, et al. Successive Annual Influenza Vaccination Induces a Recurrent Oligoclonotypic Memory Response in Circulating T Follicular Helper Cells. Sci Immunol (2017) 2(8):eaag2152. doi: 10.1126/sciimmunol.aag2152
22. Lin D, Maecker HT. Mass Cytometry Assays for Antigen-Specific T Cells Using Cytof. Methods Mol Biol (2018) 1678:37–47. doi: 10.1007/978-1-4939-7346-0_3
23. Reiss S, Baxter AE, Cirelli KM, Dan JM, Morou A, Daigneault A, et al. Comparative Analysis of Activation Induced Marker (AIM) Assays for Sensitive Identification of Antigen-Specific CD4 T Cells. PloS One (2017) 12(10):e0186998. doi: 10.1371/journal.pone.0186998
24. Chatelain E, Konar N. Translational Challenges of Animal Models in Chagas Disease Drug Development: A Review. Drug Des Devel Ther (2015) 9:4807–23. doi: 10.2147/DDDT.S90208
25. Chatelain E, Scandale I. Animal Models of Chagas Disease and Their Translational Value to Drug Development. Expert Opin Drug Discovery (2020) 15(12):1381–402. doi: 10.1080/17460441.2020.1806233
26. Akle V, Agudelo-Duenas N, Molina-Rodriguez MA, Kartchner LB, Ruth AM, Gonzalez JM, et al. Establishment of Larval Zebrafish as an Animal Model to Investigate Trypanosoma Cruzi Motility In Vivo. J Vis Exp (2017) (127):56238. doi: 10.3791/56238
27. Barry MA, Versteeg L, Wang Q, Pollet J, Zhan B, Gusovsky F, et al. A Therapeutic Vaccine Prototype Induces Protective Immunity and Reduces Cardiac Fibrosis in a Mouse Model of Chronic Trypanosoma Cruzi Infection. PloS Negl Trop Dis (2019) 13(5):e0007413. doi: 10.1371/journal.pntd.0007413
28. Caldas IS, Menezes APJ, Diniz LF, Nascimento A, Novaes RD, Caldas S, et al. Parasitaemia and Parasitic Load Are Limited Targets of the Aetiological Treatment to Control the Progression of Cardiac Fibrosis and Chronic Cardiomyopathy in Trypanosoma Cruzi-Infected Dogs. Acta Trop (2019) 189:30–8. doi: 10.1016/j.actatropica.2018.09.015
29. Vitelli-Avelar DM, Sathler-Avelar R, Mattoso-Barbosa AM, Gouin N, Perdigao-de-Oliveira M, Valerio-Dos-Reis L, et al. Cynomolgus Macaques Naturally Infected With Trypanosoma Cruzi-I Exhibit an Overall Mixed Pro-Inflammatory/Modulated Cytokine Signature Characteristic of Human Chagas Disease. PloS Negl Trop Dis (2017) 11(2):e0005233. doi: 10.1371/journal.pntd.0005233
30. Alvarez MG, Postan M, Weatherly DB, Albareda MC, Sidney J, Sette A, et al. Hla Class I-T Cell Epitopes From Trans-Sialidase Proteins Reveal Functionally Distinct Subsets of CD8+ T Cells in Chronic Chagas Disease. PloS Negl Trop Dis (2008) 2(9):e288. doi: 10.1371/journal.pntd.0000288
31. Fonseca SG, Moins-Teisserenc H, Clave E, Ianni B, Nunes VL, Mady C, et al. Identification of Multiple HLA-A*0201-restricted Cruzipain and FL-160 CD8+ Epitopes Recognized by T Cells From Chronically Trypanosoma Cruzi-Infected Patients. Microbes Infect (2005) 7(4):688–97. doi: 10.1016/j.micinf.2005.01.001
32. Laucella SA, Postan M, Martin D, Hubby Fralish B, Albareda MC, Alvarez MG, et al. Frequency of Interferon- Gamma -Producing T Cells Specific for Trypanosoma Cruzi Inversely Correlates With Disease Severity in Chronic Human Chagas Disease. J Infect Dis (2004) 189(5):909–18. doi: 10.1086/381682
33. Martin DL, Weatherly DB, Laucella SA, Cabinian MA, Crim MT, Sullivan S, et al. Cd8+ T-Cell Responses to Trypanosoma Cruzi Are Highly Focused on Strain-Variant Trans-Sialidase Epitopes. PloS Pathog (2006) 2(8):e77. doi: 10.1371/journal.ppat.0020077
34. Rosales T, Guilherme L, Chiarella J, Marin ML, Rosales C, Melo CP, et al. Human Leukocyte A and B Antigen, Gene and Haplotype Frequencies in the Population of the City of Sao Paulo in Brazil. Braz J Med Biol Res (1992) 25(1):39–47.
35. Santiago-Delpin EA. Histocompatibility Profile of Selected Latin American Countries. Transplant Proc (1991) 23(2):1861–4.
36. Wizel B, Palmieri M, Mendoza C, Arana B, Sidney J, Sette A, et al. Human Infection With Trypanosoma Cruzi Induces Parasite Antigen-Specific Cytotoxic T Lymphocyte Responses. J Clin Invest (1998) 102(5):1062–71. doi: 10.1172/JCI3835
37. Garcia F, Sepulveda P, Liegeard P, Gregoire J, Hermann E, Lemonnier F, et al. Identification of HLA-A*0201-restricted Cytotoxic T-Cell Epitopes of Trypanosoma Cruzi TcP2beta Protein in HLA-Transgenic Mice and Patients. Microbes Infect (2003) 5(5):351–9. doi: 10.1016/s1286-4579(03)00044-3
38. Acevedo GR, Juiz NA, Ziblat A, Perez Perri L, Girard MC, Ossowski MS, et al. In Silico Guided Discovery of Novel Class I and II Trypanosoma Cruzi Epitopes Recognized by T Cells From Chagas’ Disease Patients. J Immunol (2020) 204(6):1571–81. doi: 10.4049/jimmunol.1900873
39. Egui A, Thomas MC, Morell M, Maranon C, Carrilero B, Segovia M, et al. Trypanosoma Cruzi Paraflagellar Rod Proteins 2 and 3 Contain Immunodominant CD8(+) T-Cell Epitopes That are Recognized by Cytotoxic T Cells From Chagas Disease Patients. Mol Immunol (2012) 52(3-4):289–98. doi: 10.1016/j.molimm.2012.05.021
40. Egui A, Lopez MC, Gomez I, Simon M, Segovia M, Thomas MC. Differential Phenotypic and Functional Profile of Epitope-Specific Cytotoxic CD8(+) T Cells in Benznidazole-Treated Chronic Asymptomatic Chagas Disease Patients. Biochim Biophys Acta Mol Basis Dis (2020) 1866(3):165629. doi: 10.1016/j.bbadis.2019.165629
41. Planelles L, Thomas MC, Alonso C, Lopez MC. DNA Immunization With Trypanosoma Cruzi HSP70 Fused to the KMP11 Protein Elicits a Cytotoxic and Humoral Immune Response Against the Antigen and Leads to Protection. Infect Immun (2001) 69(10):6558–63. doi: 10.1128/IAI.69.10.6558-6563.2001
42. Maranon C, Thomas MC, Planelles L, Lopez MC. The Immunization of A2/K(b) Transgenic Mice With the KMP11-HSP70 Fusion Protein Induces CTL Response Against Human Cells Expressing the T. Cruzi KMP11 Antigen: Identification of A2-Restricted Epitopes. Mol Immunol (2001) 38(4):279–87. doi: 10.1016/s0161-5890(01)00059-1
43. Diez H, Lopez MC, Del Carmen Thomas M, Guzman F, Rosas F, Velazco V, et al. Evaluation of IFN-Gamma Production by CD8 T Lymphocytes in Response to the K1 Peptide From KMP-11 Protein in Patients Infected With Trypanosoma Cruzi. Parasite Immunol (2006) 28(3):101–5. doi: 10.1111/j.1365-3024.2005.00815.x
44. Lasso P, Mesa D, Cuellar A, Guzman F, Bolanos N, Rosas F, et al. Frequency of Specific CD8+ T Cells for a Promiscuous Epitope Derived From Trypanosoma Cruzi KMP-11 Protein in Chagasic Patients. Parasite Immunol (2010) 32(7):494–502. doi: 10.1111/j.1365-3024.2010.01206.x
45. Lasso P, Cardenas C, Guzman F, Rosas F, Thomas MC, Lopez MC, et al. Effect of Secondary Anchor Amino Acid Substitutions on the Immunogenic Properties of an HLA-a*0201-Restricted T Cell Epitope Derived From the Trypanosoma Cruzi KMP-11 Protein. Peptides (2016) 78:68–76. doi: 10.1016/j.peptides.2016.02.002
46. Lasso P, Beltran L, Guzman F, Rosas F, Thomas MC, Lopez MC, et al. Promiscuous Recognition of a Trypanosoma Cruzi CD8+ T Cell Epitope Among HLA-A2, Hla-A24 and HLA-A1 Supertypes in Chagasic Patients. PloS One (2016) 11(3):e0150996. doi: 10.1371/journal.pone.0150996
47. Maranon C, Egui A, Carrilero B, Thomas MC, Pinazo MJ, Gascon J, et al. Identification of HLA-A *02:01-Restricted CTL Epitopes in Trypanosoma Cruzi Heat Shock protein-70 Recognized by Chagas Disease Patients. Microbes Infect (2011) 13(12-13):1025–32. doi: 10.1016/j.micinf.2011.05.010
48. Egui A, Thomas MC, Carrilero B, Segovia M, Alonso C, Maranon C, et al. Differential Phenotypic and Functional Profiles of TcCA-2 -Specific Cytotoxic CD8+ T Cells in the Asymptomatic Versus Cardiac Phase in Chagasic Patients. PloS One (2015) 10(3):e0122115. doi: 10.1371/journal.pone.0122115
49. Abel LC, Iwai LK, Viviani W, Bilate AM, Fae KC, Ferreira RC, et al. T Cell Epitope Characterization in Tandemly Repetitive Trypanosoma Cruzi B13 Protein. Microbes Infect (2005) 7(11-12):1184–95. doi: 10.1016/j.micinf.2005.03.033
50. Iwai LK, Juliano MA, Juliano L, Kalil J, Cunha-Neto E. T-Cell Molecular Mimicry in Chagas Disease: Identification and Partial Structural Analysis of Multiple Cross-Reactive Epitopes Between Trypanosoma Cruzi B13 and Cardiac Myosin Heavy Chain. J Autoimmun (2005) 24(2):111–7. doi: 10.1016/j.jaut.2005.01.006
51. Iwai LK, Duranti MA, Abel LC, Juliano MA, Kalil J, Juliano L, et al. Retro-Inverso Peptide Analogues of Trypanosoma Cruzi B13 Protein Epitopes Fail to be Recognized by Human Sera and Peripheral Blood Mononuclear Cells. Peptides (2001) 22(6):853–60. doi: 10.1016/s0196-9781(01)00409-0
52. Arnholdt AC, Piuvezam MR, Russo DM, Lima AP, Pedrosa RC, Reed SG, et al. Analysis and Partial Epitope Mapping of Human T Cell Responses to Trypanosoma Cruzi Cysteinyl Proteinase. J Immunol (1993) 151(6):3171–9.
53. Boscardin SB, Kinoshita SS, Fujimura AE, Rodrigues MM. Immunization With cDNA Expressed by Amastigotes of Trypanosoma Cruzi Elicits Protective Immune Response Against Experimental Infection. Infect Immun (2003) 71(5):2744–57. doi: 10.1128/iai.71.5.2744-2757.2003
54. Garg N, Tarleton RL. Genetic Immunization Elicits Antigen-Specific Protective Immune Responses and Decreases Disease Severity in Trypanosoma Cruzi Infection. Infect Immun (2002) 70(10):5547–55. doi: 10.1128/iai.70.10.5547-5555.2002
55. Wizel B, Garg N, Tarleton RL. Vaccination With Trypomastigote Surface Antigen 1-Encoding Plasmid DNA Confers Protection Against Lethal Trypanosoma Cruzi Infection. Infect Immun (1998) 66(11):5073–81. doi: 10.1128/IAI.66.11.5073-5081.1998
56. Wizel B, Nunes M, Tarleton RL. Identification of Trypanosoma Cruzi Trans-Sialidase Family Members as Targets of Protective CD8+ TC1 Responses. J Immunol (1997) 159(12):6120–30.
57. Low HP, Santos MA, Wizel B, Tarleton RL. Amastigote Surface Proteins of Trypanosoma Cruzi are Targets for CD8+ Ctl. J Immunol (1998) 160(4):1817–23.
58. Rosenberg CS, Zhang W, Bustamante JM, Tarleton RL. Long-Term Immunity to Trypanosoma Cruzi in the Absence of Immunodominant trans-Sialidase-Specific Cd8+ T Cells. Infect Immun (2016) 84(9):2627–38. doi: 10.1128/IAI.00241-16
59. Bixby LM, Tarleton RL. Stable CD8+ T Cell Memory During Persistent Trypanosoma Cruzi Infection. J Immunol (2008) 181(4):2644–50. doi: 10.4049/jimmunol.181.4.2644
60. Bustamante JM, Bixby LM, Tarleton RL. Drug-Induced Cure Drives Conversion to a Stable and Protective CD8+ T Central Memory Response in Chronic Chagas Disease. Nat Med (2008) 14(5):542–50. doi: 10.1038/nm1744
61. Padilla A, Xu D, Martin D, Tarleton R. Limited Role for CD4+ T-Cell Help in the Initial Priming of Trypanosoma Cruzi-Specific CD8+ T Cells. Infect Immun (2007) 75(1):231–5. doi: 10.1128/IAI.01245-06
62. Tzelepis F, de Alencar BC, Penido ML, Claser C, Machado AV, Bruna-Romero O, et al. Infection With Trypanosoma Cruzi Restricts the Repertoire of Parasite-Specific CD8+ T Cells Leading to Immunodominance. J Immunol (2008) 180(3):1737–48. doi: 10.4049/jimmunol.180.3.1737
63. Rosenberg CS, Martin DL, Tarleton RL. Cd8+ T Cells Specific for Immunodominant Trans-Sialidase Epitopes Contribute to Control of Trypanosoma Cruzi Infection But Are Not Required for Resistance. J Immunol (2010) 185(1):560–8. doi: 10.4049/jimmunol.1000432
64. Eickhoff CS, Van Aartsen D, Terry FE, Meymandi SK, Traina MM, Hernandez S, et al. An Immunoinformatic Approach for Identification of Trypanosoma Cruzi HLA-A2-restricted Cd8(+) T Cell Epitopes. Hum Vaccin Immunother (2015) 11(9):2322–8. doi: 10.1080/21645515.2015.1061160
65. Wrightsman RA, Luhrs KA, Fouts D, Manning JE. Paraflagellar Rod Protein-Specific CD8+ Cytotoxic T Lymphocytes Target Trypanosoma Cruzi-Infected Host Cells. Parasite Immunol (2002) 24(8):401–12. doi: 10.1046/j.1365-3024.2002.00479.x
66. Fralish BH, Tarleton RL. Genetic Immunization With LYT1 or a Pool of Trans-Sialidase Genes Protects Mice From Lethal Trypanosoma Cruzi Infection. Vaccine (2003) 21(21-22):3070–80. doi: 10.1016/s0264-410x(03)00121-x
67. Vita R, Zarebski L, Greenbaum JA, Emami H, Hoof I, Salimi N, et al. The Immune Epitope Database 2.0. Nucleic Acids Res (2010) 38(Database issue):D854–862. doi: 10.1093/nar/gkp1004
68. Wrightsman RA, Miller MJ, Saborio JL, Manning JE. Pure Paraflagellar Rod Protein Protects Mice Against Trypanosoma Cruzi Infection. Infect Immun (1995) 63(1):122–5. doi: 10.1128/IAI.63.1.122-125.1995
69. Morell M, Thomas MC, Caballero T, Alonso C, Lopez MC. The Genetic Immunization With Paraflagellar Rod Protein-2 Fused to the HSP70 Confers Protection Against Late Trypanosoma Cruzi Infection. Vaccine (2006) 24(49-50):7046–55. doi: 10.1016/j.vaccine.2006.07.006
70. Rammensee H, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: Database for MHC Ligands and Peptide Motifs. Immunogenetics (1999) 50(3-4):213–9. doi: 10.1007/s002510050595
71. Parker KC, Bednarek MA, Coligan JE. Scheme for Ranking Potential HLA-A2 Binding Peptides Based on Independent Binding of Individual Peptide Side-Chains. J Immunol (1994) 152(1):163–75.
72. Michel-Todo L, Reche PA, Bigey P, Pinazo MJ, Gascon J, Alonso-Padilla J. Corrigendum: In Silico Design of an Epitope-Based Vaccine Ensemble for Chagas Disease. Front Immunol (2019) 10:3124. doi: 10.3389/fimmu.2019.03124
73. Lasso P, Mateus J, Pavia P, Rosas F, Roa N, Thomas MC, et al. Inhibitory Receptor Expression on CD8+ T Cells Is Linked to Functional Responses Against Trypanosoma Cruzi Antigens in Chronic Chagasic Patients. J Immunol (2015) 195(8):3748–58. doi: 10.4049/jimmunol.1500459
74. Reche PA, Glutting JP, Zhang H, Reinherz EL. Enhancement to the RANKPEP Resource for the Prediction of Peptide Binding to MHC Molecules Using Profiles. Immunogenetics (2004) 56(6):405–19. doi: 10.1007/s00251-004-0709-7
75. Egui A, Thomas MC, Fernandez-Villegas A, Perez-Anton E, Gomez I, Carrilero B, et al. A Parasite Biomarker Set for Evaluating Benznidazole Treatment Efficacy in Patients With Chronic Asymptomatic Trypanosoma Cruzi Infection. Antimicrob Agents Chemother (2019) 63(10):e02436-18. doi: 10.1128/AAC.02436-18
76. Cuellar A, Rojas F, Bolanos N, Diez H, Del Carmen Thomas M, Rosas F, et al. Natural CD4(+) T-Cell Responses Against Trypanosoma Cruzi KMP-11 Protein in Chronic Chagasic Patients. Immunol Cell Biol (2009) 87(2):149–53. doi: 10.1038/icb.2008.76
77. Villanueva-Lizama LE, Cruz-Chan JV, Aguilar-Cetina ADC, Herrera-Sanchez LF, Rodriguez-Perez JM, Rosado-Vallado ME, et al. Trypanosoma Cruzi Vaccine Candidate Antigens Tc24 and TSA-1 Recall Memory Immune Response Associated With HLA-A and -B Supertypes in Chagasic Chronic Patients From Mexico. PloS Negl Trop Dis (2018) 12(1):e0006240. doi: 10.1371/journal.pntd.0006240
78. El-Sayed NM, Myler PJ, Bartholomeu DC, Nilsson D, Aggarwal G, Tran AN, et al. The Genome Sequence of Trypanosoma Cruzi, Etiologic Agent of Chagas Disease. Science (2005) 309(5733):409–15. doi: 10.1126/science.1112631
79. Marin-Neto JA, Rassi A Jr, Avezum A Jr, Mattos AC, Rassi A, Morillo CA, et al. The BENEFIT Trial: Testing the Hypothesis That Trypanocidal Therapy Is Beneficial for Patients With Chronic Chagas Heart Disease. Mem Inst Oswaldo Cruz (2009) 104(Suppl 1):319–24. doi: 10.1590/s0074-02762009000900042
80. Morillo CA, Marin-Neto JA, Avezum A, Sosa-Estani S, Rassi A Jr, Rosas F, et al. Randomized Trial of Benznidazole for Chronic Chagas’ Cardiomyopathy. N Engl J Med (2015) 373(14):1295–306. doi: 10.1056/NEJMoa1507574
81. Silva ACC, Brelaz-de-Castro MCA, Leite ACL, Pereira VRA, Hernandes MZ. Chagas Disease Treatment and Rational Drug Discovery: A Challenge That Remains. Front Pharmacol (2019) 10:873. doi: 10.3389/fphar.2019.00873
82. Sewell AK. Why Must T Cells be Cross-Reactive? Nat Rev Immunol (2012) 12:669–77. doi: 10.1038/nri3279
83. Bentzen AK, Such L, Jensen KK, Marquard AM, Jessen LE, Miller NJ, et al. T Cell Receptor Fingerprinting Enables In-Depth Characterization of the Interactions Governing Recognition of Peptide-MHC Complexes. Nat Biotechnol (2018) 36:1191–6. doi: 10.1038/nbt.4303
84. Jurtz VI, Jessen LE, Bentzen AK, Jespersen MC, Mahajan S, Vita R, et al. NetTCR: Sequence-Based Prediction of TCR Binding to peptide-MHC Complexes Using Convolutional Neural Networks. bioRxiv (2018) 433706. doi: 10.1101/433706
85. Dash P, Fiore-Gartland AJ, Hertz T, Wang GC, Sharma S, Souquette A, et al. Quantifiable Predictive Features Define Epitope-Specific T Cell Receptor Repertoires. Nature (2017) 547(7661):89–93. doi: 10.1038/nature22383
86. Glanville J, Huang H, Nau A, Hatton O, Wagar LE, Rubelt F, et al. Identifying Specificity Groups in the T Cell Receptor Repertoire. Nature (2017) 547(7661):94–8. doi: 10.1038/nature22976
87. Jensen KK, Rantos V, Jappe EC, Olsen TH, Jespersen MC, Jurtz V, et al. Tcrpmhcmodels: Structural Modelling of TCR-pMHC Class I Complexes. Sci Rep (2019) 9(1):14530. doi: 10.1038/s41598-019-50932-4
88. Lanzarotti E, Marcatili P, Nielsen M. Identification of the Cognate peptide-MHC Target of T Cell Receptors Using Molecular Modeling and Force Field Scoring. Mol Immunol (2018) 94:91–7. doi: 10.1016/j.molimm.2017.12.019
89. Lanzarotti E, Marcatili P, Nielsen M. T-Cell Receptor Cognate Target Prediction Based on Paired Alpha and Beta Chain Sequence and Structural Cdr Loop Similarities. Front Immunol (2019) 10:2080. doi: 10.3389/fimmu.2019.02080
90. Khatoon N, Ojha R, Mishra A, Prajapati VK. Examination of Antigenic Proteins of Trypanosoma Cruzi to Fabricate an Epitope-Based Subunit Vaccine by Exploiting Epitope Mapping Mechanism. Vaccine (2018) 36(42):6290–300. doi: 10.1016/j.vaccine.2018.09.004
91. Richters MM, Xia H, Campbell KM, Gillanders WE, Griffith OL, Griffith M. Best Practices for Bioinformatic Characterization of Neoantigens for Clinical Utility. Genome Med (2019) 11(1):56. doi: 10.1186/s13073-019-0666-2
92. Bettencourt P, Muller J, Nicastri A, Cantillon D, Madhavan M, Charles PD, et al. Identification of Antigens Presented by MHC for Vaccines Against Tuberculosis. NPJ Vaccines (2020) 5:2. doi: 10.1038/s41541-019-0148-y
93. Mou Z, Li J, Boussoffara T, Kishi H, Hamana H, Ezzati P, et al. Identification of Broadly Conserved Cross-Species Protective Leishmania Antigen and its Responding CD4+ T Cells. Sci Transl Med (2015) 7(310):310ra167. doi: 10.1126/scitranslmed.aac5477
Keywords: Chagas disease, Trypanosoma cruzi, T cell specificity, mouse model, human model
Citation: Ferragut F, Acevedo GR and Gómez KA (2021) T Cell Specificity: A Great Challenge in Chagas Disease. Front. Immunol. 12:674078. doi: 10.3389/fimmu.2021.674078
Received: 28 February 2021; Accepted: 09 June 2021;
Published: 29 June 2021.
Edited by:
Ricardo Fujiwara, Federal University of Minas Gerais, BrazilReviewed by:
Xiao He, The University of Utah, United StatesSinéad Marian Smith, Trinity College Dublin, Ireland
Copyright © 2021 Ferragut, Acevedo and Gómez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karina A. Gómez, Z29tZXpAZG5hLnViYS5hcg==