
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 04 May 2021
Sec. Vaccines and Molecular Therapeutics
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.671325
 Francesca Micoli1
Francesca Micoli1 Omar Rossi1
Omar Rossi1 Valentino Conti1Odile Launay2,3
Valentino Conti1Odile Launay2,3 Antonella Silvia Sciré1Maria Grazia Aruta1Usman Nasir Nakakana1
Antonella Silvia Sciré1Maria Grazia Aruta1Usman Nasir Nakakana1 Elisa Marchetti1
Elisa Marchetti1 Rino Rappuoli1Allan Saul1†
Rino Rappuoli1Allan Saul1† Laura B. Martin1Francesca Necchi1
Laura B. Martin1Francesca Necchi1 Audino Podda1*
Audino Podda1*Shigella is the second most deadly diarrheal disease among children under five years of age, after rotavirus, with high morbidity and mortality in developing countries. Currently, no vaccine is widely available, and the increasing levels of multidrug resistance make Shigella a high priority for vaccine development. The single-component candidate vaccine against Shigella sonnei (1790GAHB), developed using the GMMA technology, contains the O antigen (OAg) portion of lipopolysaccharide (LPS) as active moiety. The vaccine was well tolerated and immunogenic in early-phase clinical trials. In a phase 1 placebo-controlled dose escalation trial in France (NCT02017899), three doses of five different vaccine formulations (0.06/1, 0.3/5, 1.5/25, 3/50, 6/100 µg of OAg/protein) were administered to healthy adults. In the phase 1 extension trial (NCT03089879), conducted 2–3 years following the parent study, primed individuals who had undetectable antibody levels before the primary series received a 1790GAHB booster dose (1.5/25 µg OAg/protein). Controls were unprimed participants immunized with one 1790GAHB dose. The current analysis assessed the functionality of sera collected from both studies using a high-throughput luminescence-based serum bactericidal activity (SBA) assay optimized for testing human sera. Antibodies with complement-mediated bactericidal activity were detected in vaccinees but not in placebo recipients. SBA titers increased with OAg dose, with a persistent response up to six months after the primary vaccination with at least 1.5/25 µg of OAg/protein. The booster dose induced a strong increase of SBA titers in most primed participants. Correlation between SBA titers and anti-S. sonnei LPS serum immunoglobulin G levels was observed. Results suggest that GMMA is a promising OAg delivery system for the generation of functional antibody responses and persistent immunological memory.
Diarrheal diseases continue to be a major cause of death worldwide, with approximately 1.6 million fatalities estimated in 2017 (1). Although mortality rates from diarrheal diseases have decreased since 1990 (1), diarrhea morbidity remains high, particularly in low- and middle-income countries, lacking relevant microbiological diagnostics, water quality, and sanitation, and adequate health-care facilities and treatment interventions are not easily accessible (2, 3). Complications from repeated infections are especially common in malnourished children, with possible long-term consequences including stunting and intellectual deficit (4–6). Shigella is the second leading cause of diarrheal disease after rotavirus and is the main pathogen associated with diarrhea in children under five years of age in developing countries (7–9). Shigella has also been associated with diarrhea in adults, with increased disease incidence in the elderly (10). Moreover, it is a leading cause of diarrhea in travelers and military personnel (11–13).
The genus Shigella comprises four species (S. flexneri, S. sonnei, S. dysenteriae, and S. boydii), with more than 50 serotypes differentiated based on the structure of the somatic O antigen (OAg), the polysaccharide moiety of the lipopolysaccharide (LPS) anchored in the outer membrane of the bacteria (14). OAg is the component involved in many pathogen-host interactions and is a key antigen recognized by the immune system following natural infection (14). It has been shown that individuals infected with Shigella acquire natural immunity that prevents or reduces severity of recurrent infections caused by the same serotype (15–19). This protective immunity was associated with the level of LPS-specific serum immunoglobulin G (IgG) and immunoglobulin A (IgA) antibodies during several disease outbreaks (11, 19). Individuals with a high level of anti-LPS serum antibodies showed significantly reduced disease severity (20). Therefore, current vaccine development strategies against Shigella mostly target the serotype-specific OAg of the bacteria. Several vaccine candidates, developed using different techniques, are under investigation (21–25), but no vaccine is widely available. However, the morbidity of the disease coupled with the rise of antimicrobial resistance (26, 27) urges for the introduction of an effective vaccine.
Shigella-specific antibody response following both natural infection and vaccination has traditionally been evaluated using antigen-specific enzyme-linked immunosorbent assay (ELISA). This serological method relies on antibody binding to an immobilized antigen and does not provide information on the functionality of the antibody. Although protection against Shigella is mediated by multiple mechanisms that are not yet fully elucidated, antibodies against OAg can fix complement and kill Shigella (28, 29). Bactericidal antibody activity has been identified in adults from endemic regions who mounted immunity after natural exposure (30). Even if an immunological correlate of protection is not established for Shigella, antibodies with bactericidal activity are expected to be a relevant indicator of protective immunity facilitating the development of vaccines against Shigella (31–33). A recent study reported a strong association between S. flexneri 2a-specific serum bactericidal activity (SBA) titers in human adult volunteers and reduced clinical disease following challenge with wild-type bacteria (34), thus supporting the value of bactericidal antibodies to potentially predict vaccine efficacy.
1790GAHB is a single-component S. sonnei candidate vaccine, developed using the GMMA-technology (23, 35) as a delivery system for OAg. The vaccine has been demonstrated to induce anti-LPS serum IgG antibodies and to have an acceptable safety profile in phase 1 trials conducted in adults from France and the United Kingdom (36) and in a phase 2a trial in an adult population from Kenya, where Shigella is endemic (37). A booster dose of 1790GAHB administered in an extension trial 2–3 years following the primary vaccination was well tolerated and induced an anamnestic response in adult French participants (38).
Here we report bactericidal activity of sera collected from participants of the phase 1 and extension studies using a complement-mediated high-throughput luminescence-based SBA assay developed at the GSK Vaccines Institute for Global Health (39, 40). We also assessed the impact of baseline ELISA antibody levels on SBA titer levels and correlation between ELISA IgG levels and SBA titers. A summary contextualizing the results and potential clinical relevance and impact of the research is provided in the Plain Language Summary (Figure 1).

Figure 1 Plain Language Summary.
The parent study was a phase 1, observer-blind, randomized, placebo-controlled, dose-escalation study (NCT02017899) conducted in France between February 2014 and March 2015. Detailed study design and eligibility criteria are described in the primary publication (36). Fifty healthy adult volunteers (aged 18–45 years old) were enrolled and received three doses of either one of the following 1790GAHB vaccine formulations or placebo, one month apart: 1790GAHB dose containing 0.06 µg OAg and 1 µg protein (0.06/1 group, 8 participants), 1790GAHB dose containing 0.3 µg OAg and 5 µg protein (0.3/5 group, 9 participants), 1790GAHB dose containing 1.5 µg OAg and 25 µg protein (1.5/25 group, 8 participants), 1790GAHB dose containing 3 µg OAg and 50 µg protein (3/50 group, 8 participants), or 1790GAHB dose containing 6 µg OAg and 100 µg protein (6/100 group, 9 participants). The Placebo group (8 participants) received three doses of Alhydrogel (0.7 mg Al3+/dose) in tris-buffered saline.
The extension of the parent trial was a phase 1, open label, non-randomized study (NCT03089879) conducted between March 2017 and August 2017 and designed to evaluate the memory response elicited by a booster dose of 1790GAHB (1.5/25 µg OAg/protein) in adults who had undetectable ELISA antibody levels at baseline and were primed with three vaccine doses containing different amount of OAg (four participants received 0.06/1 µg, one participant 0.3/5 µg, and two participants 3/50 µg OAg/protein dose) 2–3 years earlier in the parent study (Booster group, 7 participants). These participants were compared to vaccine-naïve individuals receiving one 1.5/25 µg 1790GAHB dose (Control group) (38). The Control group consisted of adults who received placebo during the parent study (2 participants) and newly enrolled volunteers (26 participants).
Trials were designed and conducted in accordance with the Good Clinical Practice Guidelines and the Declaration of Helsinki. Written informed consent was obtained before enrollment from each individual. The relevant ethical and regulatory approval was obtained from the respective institutional and national ethics review committees to reuse human serum pool.
Blood samples used in this analysis were collected before the first primary dose (baseline, D1), 28 days after the first (D29), second (D57), and third (D85) primary vaccination, 168 days after the third primary dose (D225) in the parent study, as well as before (pre-booster) and 28 days (D29) and 84 days (D85) after the booster dose in the extension study. Sera were stored at -80°C until further analysis.
The bactericidal activity of antibodies induced by 1790GAHB against S. sonnei 53G virG::cat, a strain with stabilized major virulence plasmid to ensure stable OAg expression, was evaluated using the high-throughput luminescence-based SBA assay, developed at GSK Vaccines Institute for Global Health (GVGH) and optimized for testing human sera (40). All tested samples were heat inactivated at 56°C for 30 minutes to remove endogenous complement activity. The assay was performed in 96-well round-bottom sterile plates (Corning). Sera were serially diluted, starting from 1:30 dilution, by 2-fold dilution steps up to seven dilution points in Phosphate Buffer Saline at pH 7 (PBS) in the SBA plates (10 µL/well). A reaction mixture of target bacterial cells (20,000 cells/well) and exogenous baby rabbit complement (at 20% final concentration) in PBS (60 µL/well) was added to each well of the SBA plate, mixed and incubated for three hours at 37°C. At the end of the incubation period, the SBA plate was centrifuged at room temperature for 10 minutes at 4000 × g. The supernatant was removed, and the remaining live bacterial pellets were resuspended in PBS, transferred to a white round-bottom 96-well plate (Greiner) and mixed 1:1 (v:v) with BacTiter-Glo Reagent (Promega). The reaction was incubated for five minutes at room temperature on an orbital shaker, and the luminescence signal was measured by a luminometer (Synergy HT, Biotek). The level of luminescence detected is directly proportional to the number of living bacteria present in the wells, which is inversely proportional to the level of functional antibodies present in the serum (39). The results of the assay are expressed as the IC50, represented by the reciprocal of the serum dilution that results in a 50% reduction of luminescence (correspondent to reciprocal of the serum dilution killing half of the bacteria present in the assay). IC50 were calculated by direct fitting of the raw luminescence data in a 4-parameter logistic regression analysis versus the log-transformed serum dilutions. Antibody titers below the lower limit of quantification (LLOQ) (IC50 of 100) were set to half that limit (IC50 of 50) for the purpose of statistical analysis.
The SBA analysis of sera from the parent study was performed in the modified full analysis set (FAS) including participants who received the study vaccine and whose sera were assayed by ELISA at all five time points. Participants were analyzed as treated and those who did not receive a study vaccine dose but provided a blood sample at a certain time point were excluded from all subsequent analysis. The analysis of sera from the extension study was performed in the FAS comprising participants who received the booster dose and provided at least one evaluable serum sample at a relevant time point.
For each group in both studies, unadjusted SBA geometric mean titers (GMTs) were calculated with their two-sided 95% confidence intervals (CIs) at each time point by exponentiating (base 10) the means and the lower and upper limit of the 95% CIs of the log-transformed titers. Anti-S. sonnei LPS serum IgG antibody geometric mean concentrations were calculated with their 95% CIs by exponentiating the mean and 95% CIs of the log-transformed (base 10) ELISA units (EUs). Subjects with missing values were excluded from analyses. Within-group geometric mean ratios and associated 95% CIs were calculated for titers/concentrations at post-vaccination time points versus baseline/pre-booster level. Detailed description of anti-S. sonnei LPS serum IgG antibody responses were provided in the original publications (36, 38). A post-hoc sub-analysis of SBA GMTs and anti-S. sonnei LPS serum IgG antibody concentrations was performed for participants with baseline antibody levels below or above the LLOQ for ELISA. For ELISA testing, the minimum measurable antibody level was determined in each assay and varied from 3.1 to 4.1 EU/mL in the parent study and 5.5 and 7.4 EU/mL in the extension study (36, 38). Antibody concentrations below this level were set at half the minimum measurable level. The number and percentage of participants with a 4-fold increase in SBA titers post-vaccination as compared to pre-vaccination were also computed with 95% CIs. Correlation of SBA GMTs and the anti-S. sonnei LPS serum IgG antibody concentrations were assessed by calculating the Pearson correlation coefficients on the log-transformed (base 10) values, for all samples pooled together within studies and by antibodies below and above the ELISA LLOQ at baseline in the parent study.
Sera from 45 participants of the parent study (38 in 1790GAHB group and seven in Placebo group) were included in the modified FAS of the current analysis. From the extension study, sera from seven participants in 1790GAHB group and 28 participants in Control group were analyzed. Demographic characteristics are presented in Figure 2.
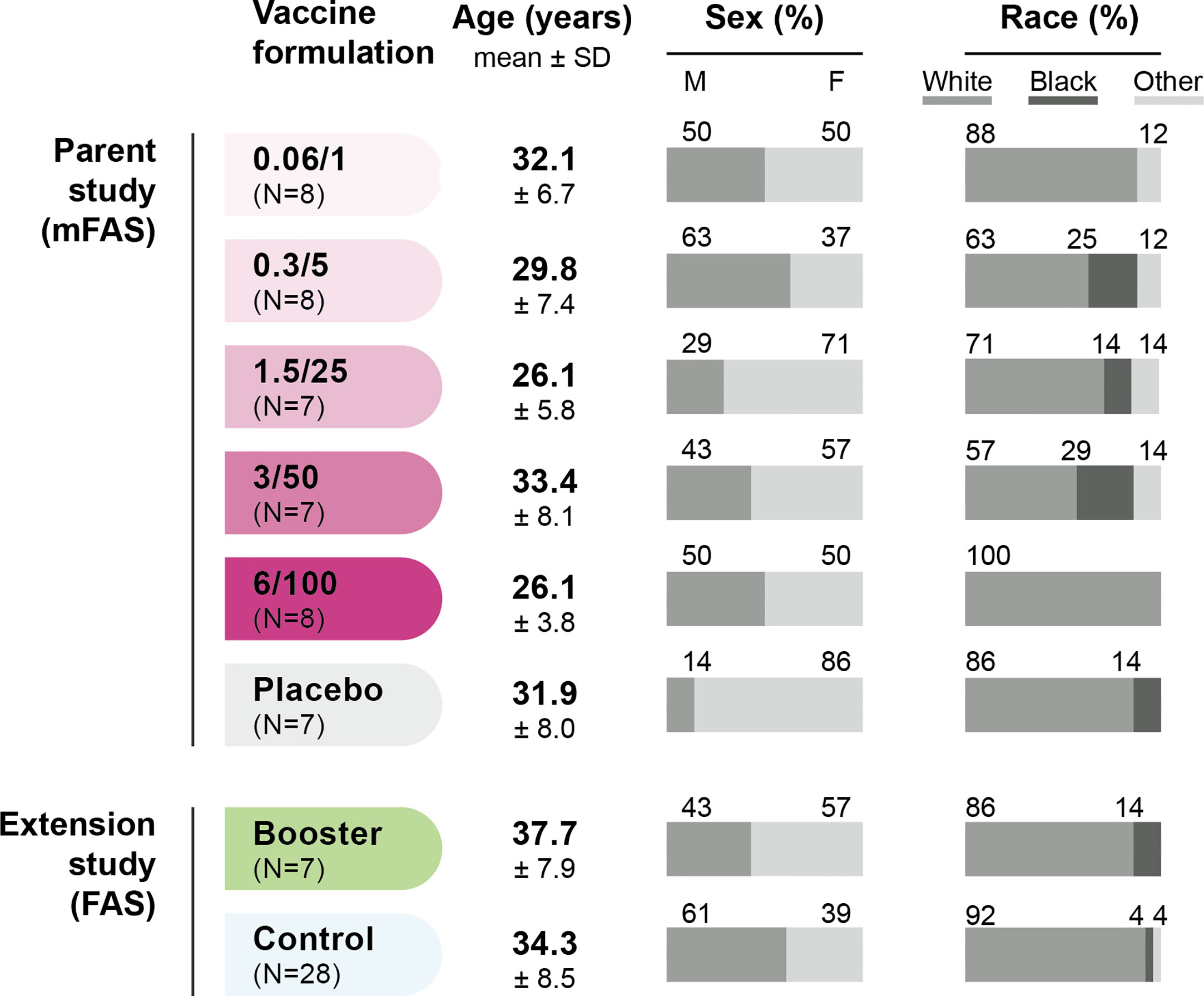
Figure 2 Demographic characteristics of participants. FAS, full analysis set; mFAS, modified FAS; SD, standard deviation; N, number of participants; 0.06/1, group receiving 1790GAHB formulation with 0.06 µg O antigen (OAg) and 1 µg protein; 0.3/5, group receiving 1790GAHB formulation with 0.3 µg OAg and 5 µg protein; 1.5/25, group receiving 1790GAHB formulation with 1.5 µg OAg and 25 µg protein; 3/50, group receiving 1790GAHB formulation with 3 µg OAg and 50 µg protein; 6/100, group receiving 1790GAHB formulation with 6 µg OAg and 100 µg protein; Placebo, group receiving placebo; Booster, group receiving a booster 1790GAHB dose (1.5/25 µg OAg/protein) 2–3 years after primary vaccination; Control, placebo recipients (from the parent study) and vaccine-naïve participants (newly enrolled in the extension study) receiving one dose of 1790GAHB (1.5/25 µg OAg/protein).
Baseline SBA GMTs were below the LLOQ in the 0.06/1 and 0.3/5 groups and were 55 (95% CI: 44–70) and 58 (95% CI: 41–83) at D29, respectively. Among participants who received 1790GAHB formulations with high OAg/protein doses, baseline SBA GMTs were 60 (95% CI: 39–93) in the 1.5/25 group, 80 (95% CI: 38–167) in the 3/50 group, and 57 (95% CI: 41–79) in the 6/100 group (Figure 3A). At D29, GMTs increased to 174 (95% CI: 51–600) in the 1.5/25 group, 255 (95% CI: 58–1117) in the 3/50 group, and 218 (95% CI: 49–975) in the 6/100 group. At six months following the third dose (D225), GMTs were below the LLOQ in the 0.06/1 group, 61 (95% CI: 38–99) in the 0.3/5 group, 276 (95% CI: 59–1293) in the 1.5/25 group, 167 (95% CI: 39–711) in the 3/50 group, and 189 (95% CI: 69–519) in the 6/100 group. When post-vaccination SBA GMTs were compared with baseline values, fold-increases remained below 1.8 at all time points in the 0.06/1 and 0.3/5 groups (Figure 3B). Conversely, SBA GMTs were 2.9-fold [1.5/25 group], 3.2-fold [3/50 group], and 3.8-fold [6/100 group] at D29 and were still 4.5-fold [1.5/25 group], 2.6-fold [3/50 group], and 3.3-fold [6/100 group] above the baseline level at D225 (Figure 3B). As expected, no SBA level increase in SBA titers was observed in the Placebo group; antibody levels remained below the LLOQ at all time points. Participants who reached 4-fold increase in bactericidal titers by D29 or D225 were only identified in groups receiving higher OAg/protein doses (Supplementary Table 1). Both SBA GMTs and anti-S. sonnei LPS serum IgG geometric mean concentrations were highest in groups receiving vaccine doses with higher OAg/protein content, irrespective of baseline antibody levels (Table 1 and Supplementary Table 2).
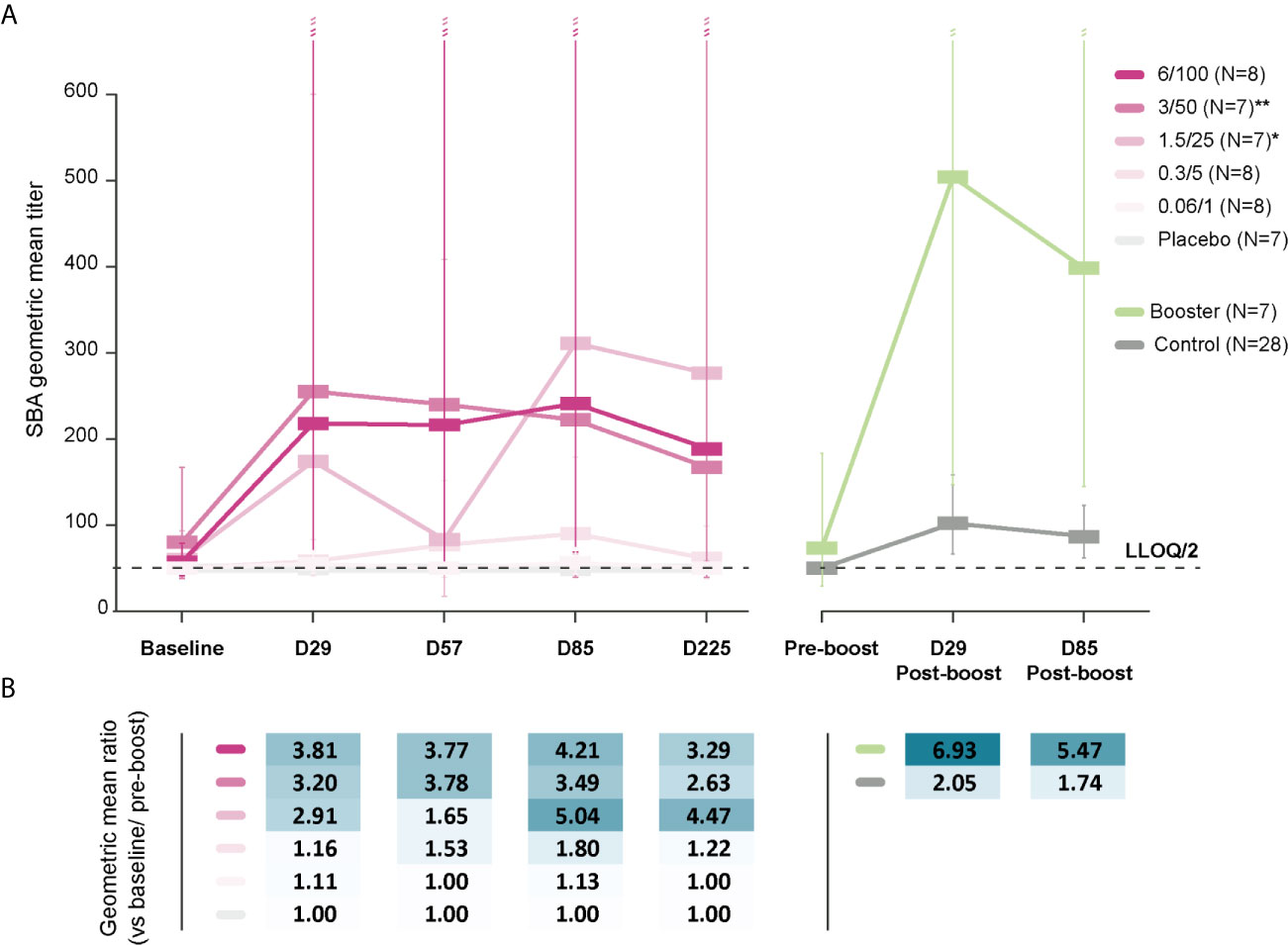
Figure 3 SBA geometric mean titers (A) and within-group geometric mean ratios (B). LLOQ, lower limit of quantification; SBA, serum bactericidal activity; Baseline, day of administration of the first dose in the parent study; D29, 28 days post-dose 1; D57, 28 days post-dose 2; D85, 28 days post-dose 3; D225, 6 months post-dose 3; Pre-boost, day of administration of the booster dose (Booster)/vaccine dose (Control) in the extension study; D29 Post-boost; 28 days following booster dose (Booster)/vaccine dose (Control); D85 post-boost, 3 months after the booster dose (Booster)/vaccine dose (Control); N, maximum number of participants with available results; 0.06/1, group receiving 1790GAHB formulation with 0.06 µg O antigen (OAg) and 1 µg protein; 0.3/5, group receiving 1790GAHB formulation with 0.3 µg OAg and 5 µg protein; 1.5/25, group receiving 1790GAHB formulation with 1.5 µg OAg and 25 µg protein; 3/50, group receiving 1790GAHB formulation with 3 µg OAg and 50 µg protein; 6/100, group receiving 1790GAHB formulation with 6 µg OAg and 100 µg protein; Placebo, group receiving placebo; Booster, group receiving a booster 1790GAHB dose (1.5/25 µg OAg/protein) 2–3 years after primary vaccination; Control, placebo recipients (from the parent study) and vaccine-naïve participants (newly enrolled in extension study) receiving one dose of 1790GAHB (1.5/25 µg OAg/protein). *N=4 at D57 and N=6 at D85, D225. **N=6 at D57, D85, D225. Error bars depict 95% confidence intervals.
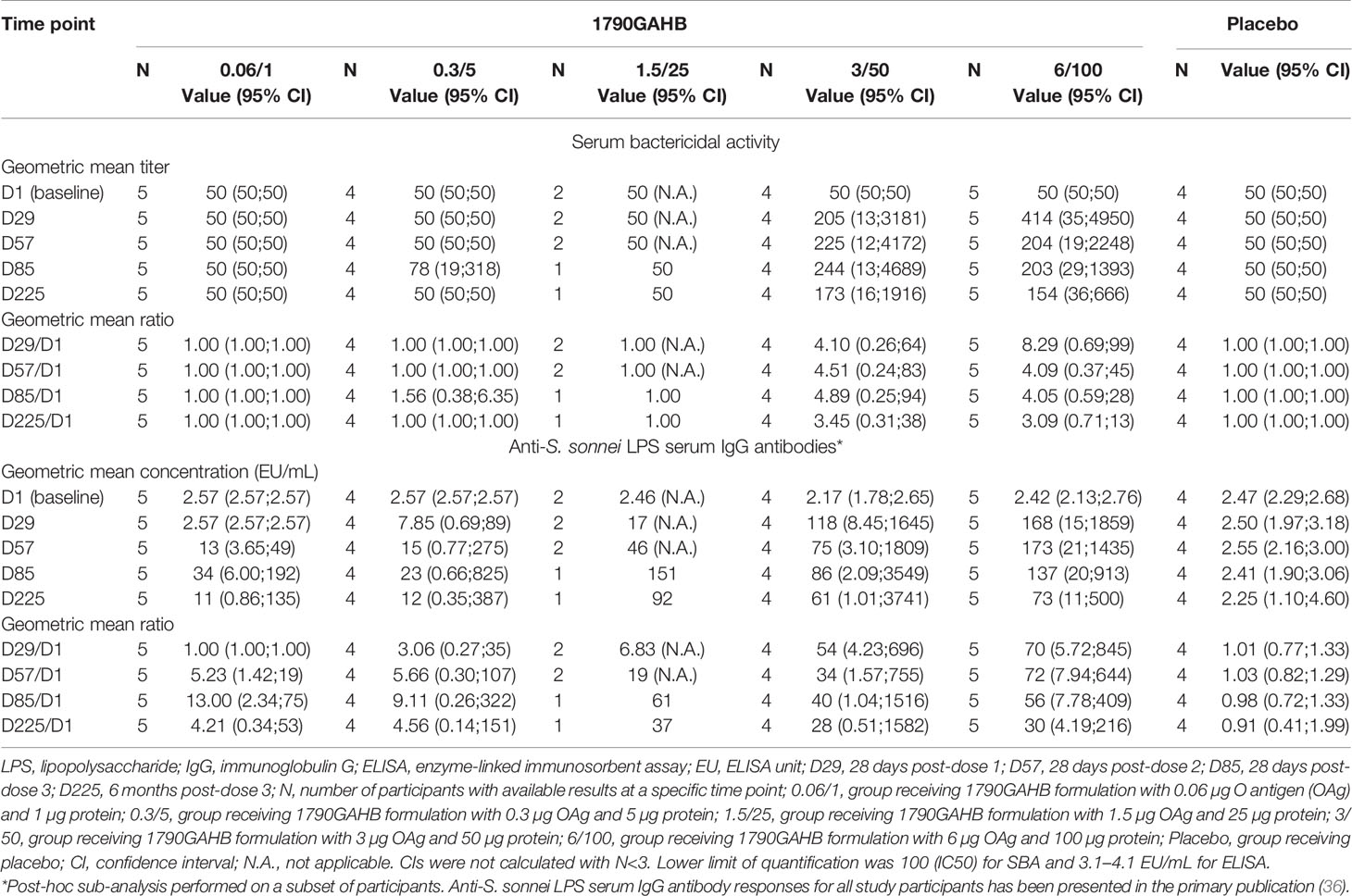
Table 1 Serum bactericidal activity geometric mean titers, anti-S. sonnei LPS serum IgG geometric mean concentrations, and within-group geometric mean ratios in participants of the parent study who had baseline antibody levels below the lower limit of quantification for ELISA (modified full analysis set).
At pre-booster, SBA GMT was 73 (95% CI: 29–183) in the Booster group and below the LLOQ in the Control group (Figure 3). In the Booster group, SBA GMTs increased 6.9-fold to 504 (95% CI: 147–1729) by D29 post-booster and remained as high as 398 (95% CI: 145–1090) at D85 post-booster vaccination. In the Control group, SBA GMTs increased 2.1-fold to 102 (95% CI: 66–158) by D29, subsequently falling to 87 (95% CI: 62–123) by D85 after vaccination. Among participants in the Control group who had pre-vaccination antibody levels above or equal to the LLOQ of the ELISA, SBA GMTs were 131 (95% CI: 72–238) at D29 and 106 (95% CI: 66–172) at D85. By contrast, lower SBA titers (62 [95% CI: 37–105] at D29 and 58 [95% CI: 40–84] at D85) were observed in Control participants with pre-vaccination levels below the LLOQ of the ELISA.
Correlation between SBA GMTs and anti-S. sonnei LPS IgG antibody concentrations was observed for both studies, the Pearson correlation coefficients being 0.748 in the parent and 0.758 in the extension study (Figure 4). In the parent study, similarly high correlation coefficients were calculated for participants with baseline antibody level either below or above the ELISA LLOQ (Supplementary Figure 1).
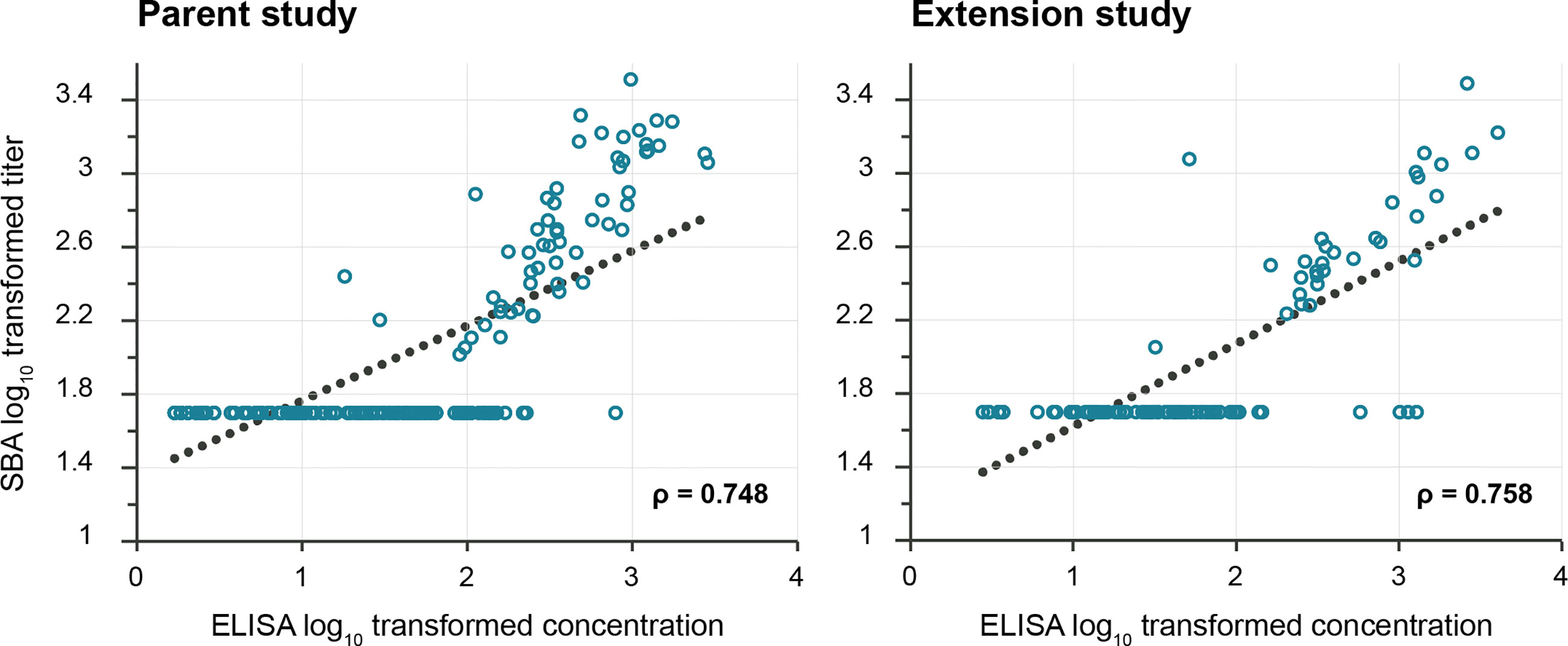
Figure 4 Pearson correlation between anti-S. sonnei LPS serum IgG antibody concentrations and SBA titers. LPS, lipopolysaccharide; IgG, immunoglobulin G; SBA, serum bactericidal activity; ELISA, enzyme-linked immunosorbent assay; EU, ELISA unit. The lower limit of quantification was 100 (IC50) for SBA and 3.1–4.1 EU/mL (parent study) and 5.5–7.4 EU/mL (extension study) for ELISA. The p-values were <0.0001 for both coefficients.
In phase 1 and phase 2 studies, immunogenicity of 1790GAHB has been evaluated in terms of anti-S. sonnei LPS serum IgG antibody response (36–38). The current analysis further characterized the immune response elicited by the vaccine and showed the ability of serum antibodies to kill Shigella in the SBA assay.
Antibodies with bactericidal activity were detected in sera from 1790GAHB vaccinees and their levels increased with antigen doses. Participants in the 0.06/1 and 0.3/5 groups did not show increase in SBA GMTs at D29 and showed a slight increase at subsequent time points only in the 0.3/5 group. In the 1.5/25, 3/50, and 6/100 groups, however, titers increased at least 2.9-fold by D29 and remained ≥2.6-fold higher by D225 than the SBA titers detected at baseline, which demonstrated the persistence of functional antibodies up to at least six months post-vaccination. The differences among groups clearly indicated an association between the OAg dose in the vaccine and the functional antibody levels elicited by this vaccine. This was also detected in the anti-S. sonnei LPS serum IgG antibody concentrations. No bactericidal activity was measured in any participants who had antibody levels below the ELISA LLOQ, demonstrating the specificity of the SBA assay toward this OAg. Bactericidal antibody responses appeared to peak at D29; overall, no further increase in SBA titers was observed after subsequent doses. A booster 1790GAHB dose administered to primed individuals during the extension study induced a robust increase in SBA titers at D29 post-booster, which slightly declined by D85 post-booster, but remained notably higher than post-primary titers. The same trend was observed in the Control group, but at a lower magnitude than in participants in the Booster group. Results obtained in SBA are in line with the observed increase in ELISA concentrations shortly after the booster dose (38). These data suggest that the one-month interval between primary doses is not adequate to further increase immune responses and an interval longer than 28 days between doses might induce higher functional antibody levels. Bactericidal activity titers correlated with anti-S. sonnei LPS serum IgG antibody concentrations in both the parent and extension studies as demonstrated by the Pearson correlation coefficients.
In the parent study, a 4-fold increase in functional antibodies, as compared to baseline titers, was detected in participants who received OAg/protein doses of at least 1.5/25 µg and the percentage of participants with a 4-fold increase tended to increase with the OAg dosage. In the extension study, almost half of the vaccinees mounted 4-fold increase post-booster. However, these data should be interpreted with caution. The use of the 4-fold criterion might lead to contradictory results due to its sensitivity to the characteristics and precision of the assay (for continuous assay readouts) or the step size used for serum dilution (for ordinal assay readouts) (41).
Our analysis is affected by the high LLOQ of the luminescence-based assay used to produce this dataset that detracts from the accurate definition of baseline titers. Additionally, this does not allow to obtain precise titers for individuals with low antibody levels, which might consequently result in a less clear dose response and lower correlation between SBA titers and ELISA concentrations. By starting the SBA assay at serum dilution of 1:4 (instead of 1:30), the LLOQ could be reduced to 33 (40), which might allow the analysis of sera with higher sensitivity and better discrimination of participants with pre-existing functional antibodies in future studies. The small sample size can be considered as another potential limitation of this study, which did not allow the assessment of statistical differences.
Other Shigella live-attenuated and conjugate vaccine candidates also elicited robust SBA titers in healthy adults following vaccination (21, 24), comparable with titers of individuals from endemic regions who acquire immunity following natural exposure (42). In a recent controlled human infection model, a challenge dose of S. sonnei 53G induced LPS-specific serum IgG and IgA antibodies with S. sonnei-specific bactericidal activity (43). In contrast, SBA against S. sonnei was not detected in adults from endemic settings vaccinated with the live oral S. sonnei vaccine (WRSS1), while in children, functional antibody response was significantly higher after three doses of WRSS1 as compared to placebo (44).
SBA reflects the functionality of the antibody response induced and may detect the contribution of multiple immune markers, indicative of a robust response (43). Our high-throughput luminescence-based SBA assay allows the analysis of a high number of samples in a relatively short time (39, 40), thus, can be suitable to perform clinical analysis in future challenge studies or field trials in Shigella-endemic regions, and to evaluate the nature of the immune response induced by Shigella vaccines.
Functional activities of vaccine-induced Shigella-specific antibodies, including both complement-mediated SBA and opsonophagocytosis, may be indicative of clinical protection against shigellosis in humans, as it has been demonstrated for other bacterial pathogens (31). SBA and opsonophagocytic killing antibody (OPKA) titers significantly correlated with reduced illness in patients who received the Shigella oral vaccine candidate EcSf2a-2 and were challenged with virulent S. flexneri 2a (34). As compared to OPKA, the SBA assay was more sensitive in detecting responses post-challenge and post-vaccination and had more potential to predict the absence of severe disease after challenge (34).
The results indicated that the serum IgG antibodies elicited by 1790GAHB were functional, with bactericidal activity detectable only in sera from vaccinees, but not from placebo recipients. SBA titers increased with the OAg dosage and sustained responses were detected after the first 1790GAHB dose with OAg/protein content ≥1.5/25 µg. An additional booster dose administered 2–3 years following primary vaccination induced a strong anamnestic response in SBA titers in all primed participants, irrespective of the OAg antigen dose they received during the primary study. SBA titers were found to strongly correlate with anti-S. sonnei LPS serum IgG antibodies. The dose response shown in the parent trial, which peaked after the first primary dose, and the strong booster response suggest that high OAg dose and longer interval between primary vaccinations might further increase immunogenicity of this GMMA-based vaccine.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The studies involving human participants were reviewed and approved by the National Ethic Committee (CPP EST1) assigned according to the pilot phase of the European Union Regulation No. 536/2014 for clinical trial applications in France (for the protocol of the extension trial H03_01E1TP) and CPP Ile-de-France III. Hopital Cochin, Paris, France (for the parent trial H03_01TP). The patients/participants provided their written informed consent to participate in this study.
FM, LM, FN, OR, AP, ASS, EM, VC, UN, and OL were involved in the design of the study. FM, FN, OR, EM, and OL performed the study. FM, LM, FN, OR, AP, MA, ASS, EM, VC, OL participated in the collection or generation of study data. FM, LM, FN, OR, and MA contributed with material and reagent tools. FM, LM, FN, OR, AP, ASS, VC, UN, and RR were involved in the analyses and interpretation of the data. LM conceived and coordinated SBA implementation in the overall strategy of the 1790GAHB development. All authors contributed to the article and approved the submitted version.
GlaxoSmithKline Biologicals SA funded this analysis and took responsibility for all costs associated with the development of the present manuscript.
FM, OR, VC, ASS, MA, UN, EM, RR, LM, FN, and AP are employees of the GSK group of companies and RR, LM, and AP hold shares in the GSK group of companies. AS was an employee of the GSK group of companies at the time of the study and holds shares in the GSK group of companies. FM and AS report grant from the Bill and Melinda Gates Foundation during the conduct of the study. LM reports grant from EU FP7 STOPENTERICS during the conduct of the study and from the Bill and Melinda Gates Foundation and Wellcome Trust outside the submitted work. LM and AS are inventors of patents owned by the GSK group of companies and relevant to Shigella vaccine. OL’s institution received grant from the GSK group of companies for conducting the study. All authors have no non-financial relationships and activities.
The authors would like to thank the current and former GVGH staff members for enabling this study. They also thank the Modis platform, on behalf of GSK, for editorial assistance and manuscript coordination. Medical writing services were provided by Botond Nagy and editorial assistance and publication coordination were provided by Camille Turlure.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.671325/full#supplementary-material
OAg, O antigen; LPS, lipopolysaccharide; SBA, serum bactericidal activity; ELISA, enzyme-linked immunosorbent assay; LLOQ, lower limit of quantification; GVGH, GSK Vaccine Institute for Global Health; IgA, immunoglobulin A; IgG, immunoglobulin G; OPKA, opsonophagocytic killing antibody; PBS, phosphate buffer saline; FAS, full analysis set, GMT, geometric mean titer; CI, confidence interval; EU, ELISA unit.
1. GBD 2017 Causes of Death Collaborators. Global, Regional, and National Age-Sex-Specific Mortality for 282 Causes of Death in 195 Countries and Territories, 1980-2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet (2018) 392(10159):1736–88. doi: 10.1016/s0140-6736(18)32203-7
2. Anderson M, Sansonetti PJ, Marteyn BS. Shigella Diversity and Changing Landscape: Insights for the Twenty-First Century. Front Cell Infect Microbiol (2016) 6:45. doi: 10.3389/fcimb.2016.00045
3. Ahs JW, Tao W, Lofgren J, Forsberg BC. Diarrheal Diseases in Low- and Middle-Income Countries: Incidence, Prevention and Management. Open Infect Dis J (2010) 4:113–24. doi: 10.2174/1874279301004010113
4. Malnutrition and Enteric Disease Study (MAL-ED). The Significant Role of Malnutrition, Intestinal Infection and Disease on Child Development (2017). Available at: http://fnih.org/news/announcements/mal-ed-study-results (Accessed on 11 January 2021).
5. Khatun F, Faruque AS, Koeck JL, Olliaro P, Millet P, Paris N, et al. Changing Species Distribution and Antimicrobial Susceptibility Pattern of Shigella Over a 29-Year Period (1980-2008). Epidemiol Infect (2011) 139(3):446–52. doi: 10.1017/s0950268810001093
6. Mahbub MM, Ahsan CR, Yasmin M, Nessa J. Analysis of Different Prognostic Indicators for Malnutrition and Shigella flexneri Infection Among the Children in Bangladesh. Indian J Microbiol (2012) 52(3):400–5. doi: 10.1007/s12088-012-0247-1
7. Platts-Mills JA, Babji S, Bodhidatta L, Gratz J, Haque R, Havt A, et al. Pathogen-Specific Burdens of Community Diarrhoea in Developing Countries: A Multisite Birth Cohort Study (MAL-ED). Lancet Glob Health (2015) 3(9):e564–75. doi: 10.1016/S2214-109X(15)00151-5
8. Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, et al. Burden and Aetiology of Diarrhoeal Disease in Infants and Young Children in Developing Countries (the Global Enteric Multicenter Study, GEMS): A Prospective, Case-Control Study. Lancet (2013) 382(9888):209–22. doi: 10.1016/s0140-6736(13)60844-2
9. GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the Global, Regional, and National Morbidity, Mortality, and Aetiologies of Diarrhoea in 195 Countries: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Infect Dis (2018) 18(11):1211–28. doi: 10.1016/S1473-3099(18)30362-1
10. Khalil IA, Troeger C, Blacker BF, Rao PC, Brown A, Atherly DE, et al. Morbidity and Mortality Due to Shigella and Enterotoxigenic Escherichia coli Diarrhoea: The Global Burden of Disease Study 1990-2016. Lancet Infect Dis (2018) 18(11):1229–40. doi: 10.1016/s1473-3099(18)30475-4
11. Cohen D, Block C, Green MS, Lowell G, Ofek I, et al. Immunoglobulin M, A, and G Antibody Response to Lipopolysaccharide O Antigen in Symptomatic and Asymptomatic Shigella Infections. J Clin Microbiol (1989) 27(1):162–7. doi: 10.1128/JCM.27.1.162-167.1989
12. Ekdahl K, Andersson Y. The Epidemiology of Travel-Associated Shigellosis-Regional Risks, Seasonality and Serogroups. J Infect (2005) 51:222–9. doi: 10.1016/j.jinf.2005.02.002
13. Riddle MS. Is a Shigella Vaccine Needed for Travellers and the Military? J Travel Med (2018) 25(1):tay49. doi: 10.1093/jtm/tay049
14. Mattock E, Blocker AJ. How do the Virulence Factors of Shigella Work Together to Cause Disease? Front Cell Infect Microbiol (2017) 7:64. doi: 10.3389/fcimb.2017.00064
15. Raqib R, Qadri F, SarkEr P, Mia SMS, Sansonnetti PJ, Albert MJ, et al. Delayed and Reduced Adaptive Humoral Immune Responses in Children With Shigellosis Compared With in Adults. Scand J Immunol (2002) 55(4):414–23. doi: 10.1046/j.1365-3083.2002.01079.x
16. Cohen D, Green MS, Block C, Slepon R, Ofek I. Prospective Study of the Association Between Serum Antibodies to Lipopolysaccharide O Antigen and the Attack Rate of Shigellosis. J Clin Microbiol (1991) 29(2):386–9. doi: 10.1128/JCM.29.2.386-389.1991
17. Robin G, Cohen D, Orr N, Markus I, Slepon R, Ashkenazi S, et al. Characterization and Quantitative Analysis of Serum IgG Class and Subclass Response to Shigella sonnei and Shigella flexneri 2a Lipopolysaccharide Following Natural Shigella Infection. J Infect Dis (1997) 175(5):1128–33. doi: 10.1086/516452
18. Ferreccio C, Prado V, Ojeda A, Cayyazo M, Abrego P, Guers L, et al. Epidemiologic Patterns of Acute Diarrhea and Endemic Shigella Infections in Children in a Poor Periurban Setting in Santiago, Chile. Am J Epidemiol (1991) 134(6):614–27. doi: 10.1093/oxfordjournals.aje.a116134
19. Cohen D, Green MS, Block C, Rouach T, Ofek I. Serum Antibodies to Lipopolysaccharide and Natural Immunity to Shigellosis in an Israeli Military Population. J Infect Dis (1988) 157(5):1068–71. doi: 10.1093/infdis/157.5.1068
20. Wahid R, Simon JK, Picking WL, Kotloff KL, Levine MM, Sztein MB. Shigella Antigen-Specific B Memory Cells are Associated With Decreased Disease Severity in Subjects Challenged With Wild-Type Shigella flexneri 2a. Clin Immunol (2013) 148(1):35–43. doi: 10.1016/j.clim.2013.03.009
21. Cohen D, Atsmon J, Artaud C, Meron-Sudai S, Gougeon M-L, Bialik A, et al. Safety and Immunogenicity of a Synthetic Carbohydrate Conjugate Vaccine Against Shigella flexneri 2a in Healthy Adult Volunteers: A Phase 1, Dose-Escalating, Single-Blind, Randomised, Placebo-Controlled Study. Lancet Infect Dis (2021) 21(4):546–58. doi: 10.1016/S1473-3099(20)30488-6
22. Frenck RW Jr, Baqar S, Alexander W, Dickey M, McNeal M, El-Khorazaty J, et al. A Phase I Trial to Evaluate the Safety and Immunogenicity of WRSs2 and WRSs3; Two Live Oral Candidate Vaccines Against Shigella sonnei. Vaccine (2018) 36(32 Pt B):4880–9. doi: 10.1016/j.vaccine.2018.06.063
23. Gerke C, Colucci AM, Giannelli C, Sanzone S, Vitali CG, Sollai L, et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790gahb. PloS One (2015) 10(8):e0134478. doi: 10.1371/journal.pone.0134478
24. Riddle MS, Kaminski RW, Di Paolo C, Porter CK, Gutierrez RL, Clarkson KA, et al. Safety and Immunogenicity of a Candidate Bioconjugate Vaccine Against Shigella flexneri 2a Administered to Healthy Adults: A Single-Blind, Randomized Phase I Study. Clin Vaccine Immunol (2016) 23(12):908–17. doi: 10.1128/cvi.00224-16
25. van der Put RMF, Kim TH, Guerreiro C, Thouron F, Hoogerhout P, Sansonetti PJ, et al. A Synthetic Carbohydrate Conjugate Vaccine Candidate Against Shigellosis: Improved Bioconjugation and Impact of Alum on Immunogenicity. Bioconjug Chem (2016) 27(4):883–92. doi: 10.1021/acs.bioconjchem.5b00617
26. Puzari M, Sharma M, Chetia P. Emergence of Antibiotic Resistant Shigella Species: A Matter of Concern. J Infect Public Health (2018) 11(4):451–4. doi: 10.1016/j.jiph.2017.09.025
27. Rahman M, Haque AF, Deeba I, Ahmed D, Zahidi T, Rimu A, et al. Emergence of Extensively Drug-Resistant Shigella sonnei in Bangladesh. Immunol Infect Dis (2017) 5:1–9. doi: 10.13189/iid.2017.050101
28. Okamura N, Nakaya R, Suzuki K, Kondo S, Hisatsune K, Imagawa Y, et al. Differences Among Shigella spp. in Susceptibility to the Bactericidal Activity of Human Serum. J Gen Microbiol (1988) 134(7):2057–65. doi: 10.1099/00221287-134-7-2057
29. Roantree RJ, Rantz LA. A Study of the Relationship of the Normal Bactericidal Activity of Human Serum to Bacterial Infection. J Clin Invest (1960) 39(1):72–81. doi: 10.1172/JCI104029
30. Sayem MA, Ahmad SM, Rekha RS, Sarker P, Agerberth B, Talukder KA, et al. Differential Host Immune Responses to Epidemic and Endemic Strains of Shigella dysenteriae Type I. J Health Popul Nutr (2011) 29(5):429–37. doi: 10.3329/jhpn.v29i5.8896
31. Ndungo E, Pasetti MF. Functional Antibodies as Immunological Endpoints to Evaluate Protective Immunity Against Shigella. Hum Vaccin Immunother (2020) 16(1):197–205. doi: 10.1080/21645515.2019.1640427
32. Nahm MH, Yu J, Weerts HP, Wenzel H, Tamilselvi CS, Chandrasekaran L, et al. Development, Interlaboratory Evaluations, and Application of a Simple, High-Throughput Shigella Serum Bactericidal Assay. mSphere (2018) 3(3):e00146–00118. doi: 10.1128/mSphere.00146-18
33. Weerts HP, Yu J, Kaminski RW, Nahm MH. A High-throughput Shigella-Specific Bactericidal Assay. J Vis Exp (2019) 144:e59164. doi: 10.3791/59164
34. Shimanovich AA, Buskirk AD, Heine SJ, Blackwelder WC, Wahid R, Kotloff KL, et al. Functional and Antigen-Specific Serum Antibody Levels as Correlates of Protection Against Shigellosis in a Controlled Human Challenge Study. Clin Vaccine Immunol (2017) 24(2):e00412–00416. doi: 10.1128/CVI.00412-16
35. Micoli F, MacLennan CA. Outer Membrane Vesicle Vaccines. Semin Immunol (2020) 50:101433. doi: 10.1016/j.smim.2020.101433
36. Launay O, Lewis DJM, Anemona A, Loulergue P, Leahy J, Sciré AS, et al. Safety Profile and Immunologic Responses of a Novel Vaccine Against Shigella sonnei Administered Intramuscularly, Intradermally and Intranasally: Results From Two Parallel Randomized Phase 1 Clinical Studies in Healthy Adult Volunteers in Europe. EBioMedicine (2017) 22:164–72. doi: 10.1016/j.ebiom.2017.07.013
37. Obiero CW, Ndiaye AGW, Sciré AS, Kaunyangi BM, Marchetti E, Gone AM, et al. A Phase 2a Randomized Study to Evaluate the Safety and Immunogenicity of the 1790GAHB Generalized Modules for Membrane Antigen Vaccine Against Shigella sonnei Administered Intramuscularly to Adults From a Shigellosis-Endemic Country. Front Immunol (2017) 8:1884. doi: 10.3389/fimmu.2017.01884
38. Launay O, Ndiaye AGW, Conti V, Loulergue P, Scire AS, Landre AM, et al. Booster Vaccination With GVGH Shigella sonnei 1790GAHB GMMA Vaccine Compared to Single Vaccination in Unvaccinated Healthy European Adults: Results From a Phase 1 Clinical Trial. Front Immunol (2019) 10:335. doi: 10.3389/fimmu.2019.00335
39. Necchi F, Saul A, Rondini S. Development of a High-Throughput Method to Evaluate Serum Bactericidal Activity Using Bacterial ATP Measurement as Survival Readout. PloS One (2017) 12(2):e0172163. doi: 10.1371/journal.pone.0172163
40. Rossi O, Molesti E, Saul A, Giannelli C, Micoli F, Necchi F. Intra-Laboratory Evaluation of Luminescence Based High-Throughput Serum Bactericidal Assay (L-SBA) to Determine Bactericidal Activity of Human Sera Against Shigella. High Throughput (2020) 9(2):33. doi: 10.3390/ht9020014
41. Saul A, Podda A, Rappuoli R. The Use and Abuse of a 4-Fold Increase in Antibody Response to Assess Immunogenicity in Early Stage Vaccine Clinical Trials. Vaccine (2020) 38(5):951–3. doi: 10.1016/j.vaccine.2019.11.067
42. Chitradevi STS, Heine SJ, Mani S, Kotloff KL, Harro C, Chakraborty S, et al. Shigella-specific Serum Bactericidal and Opsonophagocytic Killing Aantibodies Induced by Oral S. flexneri 2a Whole Cell Killed and Live Attenuated Vaccines. Vaccines Against Shigella and ETEC (VASE) Conference. Mexico City, Mexico, Abstract CL079 (2018). Available at: https://custom.cvent.com/6E6C241B944D48C3BEC9C4EC34A7E314/files/4f4cf8f90c4e4b37b707b2e29fc24866.pdf.
43. Clarkson KA, Frenck RW Jr, Dickey M, Suvarnapunya AE, Chandrasekaran L, Weerts HP, et al. Immune Response Characterization After Controlled Infection With Lyophilized Shigella sonnei 53G. mSphere (2020) 5(5):e00988–00919. doi: 10.1128/mSphere.00988-19
44. Sarker P, Mily A, Walker R, Wierzba T, Venkatesan M, Raqib R. Functional Antibodies and Cytokine Responses to Live Oral Shigella sonnei Vaccine Strain WRSS1 in Bangladeshi Adults and Children. Vaccines Against Shigella and ETEC (VASE) Conference. Mexico City, Mexico, Abstract CL033 (2018). Available at: https://custom.cvent.com/6E6C241B944D48C3BEC9C4EC34A7E314/files/4f4cf8f90c4e4b37b707b2e29fc24866.pdf.
Keywords: Shigella sonnei, GMMA, serum bactericidal activity, dose escalation, booster response, 1790GAHB
Citation: Micoli F, Rossi O, Conti V, Launay O, Sciré AS, Aruta MG, Nakakana UN, Marchetti E, Rappuoli R, Saul A, Martin LB, Necchi F and Podda A (2021) Antibodies Elicited by the Shigella sonnei GMMA Vaccine in Adults Trigger Complement-Mediated Serum Bactericidal Activity: Results From a Phase 1 Dose Escalation Trial Followed by a Booster Extension. Front. Immunol. 12:671325. doi: 10.3389/fimmu.2021.671325
Received: 23 February 2021; Accepted: 20 April 2021;
Published: 04 May 2021.
Edited by:
Susu M. Zughaier, Qatar University, QatarReviewed by:
Sanjay Ram, University of Massachusetts Medical School, United StatesCopyright © 2021 Micoli, Rossi, Conti, Launay, Sciré, Aruta, Nakakana, Marchetti, Rappuoli, Saul, Martin, Necchi and Podda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Audino Podda, YXVkaW5vLnAucG9kZGFAZ3NrLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.