 Qingqing Jiao1,2,3†
Qingqing Jiao1,2,3† Ying Luo2†
Ying Luo2† Jörg Scheffel2
Jörg Scheffel2 Peng Geng1,4
Peng Geng1,4 Yuhan Wang1,4
Yuhan Wang1,4 Stefan Frischbutter2
Stefan Frischbutter2 Ruoyu Li1,4
Ruoyu Li1,4 Marcus Maurer2*†
Marcus Maurer2*† Zuotao Zhao1,4*†
Zuotao Zhao1,4*†- 1Department of Dermatology, First Hospital, Peking University, Beijing, China
- 2Department of Dermatology and Allergy, Charite–Universitätsmeidzin Berlin, Berlin, Germany
- 3Department of Dermatology, The First Affiliated Hospital of Soochow University, Suzhou, China
- 4National Clinical Research Center for Skin and Immune Diseases, Department of Dermatology, Peking University First Hospital, Beijing, China
Background: Sporothrix schenckii (S. schenckii), a dimorphic fungus, causes sporotrichosis. Mast cells (MCs) have been described to be involved in skin fungal infections. The role of MCs in cutaneous sporotrichosis remains largely unknown.
Objectives: To characterize the role and relevance of MCs in cutaneous sporotrichosis.
Methods: We analyzed cutaneous sporotrichosis in wild-type (WT) mice and two different MC-deficient strains. In vitro, MCs were assessed for S. schenckii-induced cytokine production and degranulation after incubation with S. schenckii. We also explored the role of MCs in human cutaneous sporotrichosis.
Results: WT mice developed markedly larger skin lesions than MC-deficient mice (> 1.5 fold) after infection with S. schenckii, with significantly increased fungal burden. S. schenckii induced the release of tumor necrosis factor alpha (TNF), interleukin (IL)-6, IL-10, and IL-1β by MCs, but not degranulation. S. schenckii induced larger skin lesions and higher release of IL-6 and TNF by MCs as compared to the less virulent S. albicans. In patients with sporotrichosis, TNF and IL-6 were increased in skin lesions, and markedly elevated levels in the serum were linked to disease activity.
Conclusions: These findings suggest that cutaneous MCs contribute to skin sporotrichosis by releasing cytokines such as TNF and IL-6.
What Is Already Known About This Topic?
• Sporotrichosis is an infection caused by Sporothrix schenckii (S. schenckii) and can be particularly harmful in immunocompromised patients.
• In addition to their critical role in allergic disorders, Mast cells (MCs) have been recognized for their complex role in fungal infections.
What Does This Study Add?
• Wild type (WT) mice that are infected with S. schenckii develop larger sporotrichosis lesions than MC-deficient mice. This is associated with the production of the proinflammatory cytokines IL-6 and TNF by MCs.
• The severity of S. schenckii infections in humans correlates with IL-6 and TNF levels.
• MCs contribute to the progress of skin sporotrichosis.
What Is the Translational Message?
Mast cells and mast cell-derived cytokines should be further explored for their role in sporotrichosis.
Introduction
Sporotrichosis is a subacute or chronic infection caused by the dimorphic fungus Sporothrix schenckii (S. schenckii), which has a worldwide distribution (1–3). During the last 10 years, the overall incidence of sporotrichosis has continued to increase, especially in the northeast of China. In humans, this mycosis mainly affects the skin of infected patients. Although less common, other organs may also be affected, such as the lungs, joints, bones, and even the brain, especially in immunocompromised individuals. Treatment usually involves the use of antifungal agents and/or surgical excision. However, patients are frequently resistant to treatment, there have been reports of relapse or progression during therapy, and problems due to a lack of tolerability of antifungal drugs occur often (4). Thus, research is necessary to identify and characterize the underlying mechanisms of Sporothrix infection, which may lead to the development of new and better therapeutic options.
Mast cells (MCs) are strategically located in tissues at the interface with the environment such as the lung, the gut, and the skin. Therefore, MCs are perfectly positioned to orchestrate defense mechanisms for invading pathogens including fungi (5, 6). Our recent review of the literature on the role of MCs during innate immune responses against invading fungi revealed that MCs function as positive or negative immunoregulatory cells depending on the situation (7). Examples of beneficial host responses against fungal infections by MCs include their effects in promoting the restriction of infections or the resolution of inflammation (8–12). Negative MC responses to fungal infections include the promotion of a wider dissemination of antigens, increasing the severity of the infection and inflammation, and promoting the development of atopic disease (13–25). These findings challenge and extend the current notion of MCs primarily serving the host rather than the pathogen when immune responses are raised at sites of infections.
As of now, there is only limited information on the contribution of MCs to immune responses against S. schenckii infection (19, 22). Using an experimental mouse model of systemic S. schenckii infection, Romo-Lozano et al. found that mice without functional peritoneal MCs presented with a decreased fungal load in organs and a reduced severity of clinical manifestations (26). This suggests that MCs may promote the dissemination of S. schenckii and increase the severity of the infection. However, the major clinical manifestations in most cases of sporotrichosis occur in the skin, after traumatic inoculation or zoonotic transmission, and infections with S. schenckii primarily results in cutaneous sporotrichosis (fixed cutaneous, lymphocutaneous, or disseminated forms) (27). As of now, the role of MCs in sporotrichosis of the skin has not been investigated and, thus, remains to be characterized and defined (28–31). Consequently, the purpose of the current study was to investigate the in vitro response of skin MCs to S. schenckii involvement in a murine model of experimental cutaneous sporotrichosis, and their role in human skin infections with Sporothrix.
Materials and Methods
Mice
All mice used in the experiments were on a C57BL/6 background. Wild-type (WT) C57BL/6-Kit+/+ and MC-deficient C57BL/6-KitW/KitW−v mice were bred and housed at our facilities, and wild-type (WT) Cpa3-Cre/Mcl-1+/+ and MC-deficient Cpa3-Cre/Mcl-1fl/fl (Hello Kitty, HK) mice were also obtained from breeding colonies of the animal facilities of the Charite - Universitätsmedizin Berlin. All mice were kept under specific pathogen-free conditions, and all experiments were conducted according to institutional regulations.
Fungal Strains
The S. schenckii wild-type strain M-64 (ATCC MYA 4822) was a donation from Prof. Sandro Rogerio de Almeida's laboratory (Department of Clinical and Toxicological Analyses, School of Pharmaceutical Sciences, University of São Paulo, SP, Brazil). The Sporothrix albicans wild-type strain (S. albicans, ATCC® 201162TM) was purchased from ATCC. Initially, the isolates were subcultured from conidia until complete differentiation into the yeast form. Yeast cells were grown in brain-heart infusion broth (BHI, HB8297-1, Hopebio, China) for 7 days at 37°C with constant rotary shaking at 150 cycles/min, in order to yield a high percentage of yeast conversion. Then, yeasts were harvested from the BHI culture by centrifugation, washed twice and adjusted to the required concentration in sterile phosphate buffered saline (PBS), pH 7.4, and stored at 2–8°C until use.
Murine Model of Skin Sporotrichosis
To study the role of MCs in skin infections with Sporothrix, we infected MC-deficient mice (HK) and WT littermates (C57BL/6) with S. schenckii subcutaneously by injecting 30 μl of PBS containing 3 × 107 cells of S. schenckii yeast into the right foot pad, and PBS vehicle into the left foot pad as autologous control. Mice were then examined every 3 days during 5 weeks by evaluating the thickness, width, and length of foot inflammation with an electronic micrometer and the presence of skin ulceration or scarring in foot pad by photographing. The lesion volume was calculated (in mm3) as ellipsoids [(a/2×b/2×c/2)×4/3×π].
Determination of Fungal Burden
The fungal burden of infected skin sites of mice was measured by counting colony-forming units (CFU). Briefly, the organs were separated, weighed, and homogenized in sterile PBS with a tissue grinder. Samples (100 μL) of each homogenate were seeded on Petri dishes containing BHI agar and incubated at 37°C. The material was incubated for 7 days at room temperature, and the number of colonies formed on each plate (CFU) was counted, and the mean of viable fungi was then calculated for each group. The results were expressed as CFU/g tissue.
Histology
Tissue samples from infected mice were fixed in 10% formalin for 16 h, dehydrated in alcohol, and embedded in paraffin. Histological analyses of foot, spleen, lungs, liver, and lymph node were performed with a Zeiss Axioplan 2 Imaging microscope using a Zeiss AxioCam camera run by AxioVision Rel. 4.8 software. Giemsa stain was used to identify degranulated MCs.
For immunohistochemistry staining, sections were deparaffinized in xylene for 10 min and then rehydrated in graded alcohols and water. TNF staining was performed according to the manufacturer's instructions (DakoEnVision+ System- HRP Labeled PolymerAnti-Rabbit). Briefly, antigen retrieval was executed in citrate buffer (pH 6.0) in 95°C for 30 min. After blocking of non-specific binding with Block Dako for 10 min at room temperature, sections were incubated with primary antibodies against human and mouse TNF (ab9739, Abcam) in a humid chamber at 4°C overnight. Then sections were treated with H2O2 for 5 min at room temperature (RT). After three washes with TBS, sections were incubated with polymer secondary anti-rabbit secondary antibodies for 30 min, washed three times in TBS, and incubated with substrate AEC for 5 min. Slides were rinsed in TBS and counterstained with Mayer's hematoxylin (ab128990, Abcam; negative control = omission of the primary antibody). For IL-6 staining (Dako REAL™ Detection System, Alkaline Phosphatase/RED, Rabbit/Mouse, K5005), antigen retrieval was executed in citrate buffer (pH 6.0) in 95°Cfor 30 min. After blocking with avidin block, biotin block and 5% goat serum for 15 min each at room temperature, sections were incubated with primary antibodies against mouse IL-6 (sc-1265, Santa Cruz Biotechnology) and human IL-6 (sc-130326, Santa Cruz Biotechnology) in a humid chamber at 4°C overnight. After three washes with TBS, sections were incubated with biotinylated goat anti-rabbit secondary antibodies for 30 min, washed three times in TBS and incubated with ABC and substrate. Slides were rinsed in TBS and counterstained with Mayer's hematoxylin as described before. The expression of TNF and IL-6 was assessed by histomorphometry (Zeiss Axioplan 2 Imaging microscope using Zeiss AxioCam camera run by AxioVision Rel. 4.8 software).
BMCMC Cultures and Reconstitution of Mast Cell-Deficient KitW/KitW-v Mice
Femoral and tibial bone marrow from 4 to 6-week old C57BL/6 mice was cultured in DMEM plus recombinant stem cell factor (SCF, 20 ng/mL, Biolegend) and recombinant interleukin-3 (IL-3, 20 ng/mL, Biolegend) containing medium for 4–8 weeks to generate populations of bone marrow-derived cultured mast cells (BMCMCs) that were >95% pure as assessed by flow cytometry counting CD117 and FceRI double positive cells.
BMCMCs (106 in 50 μl 0.9% NaCl) were injected intradermally and mice were used for experiments, together with sex- and age-matched MC-deficient KitW/KitW−v and Kit+/+ mice, 4 wk after adoptive transfer. Reconstitution of cutaneous MC populations was confirmed by histomorphometric analyses of paraffin-embedded, Giemsa-stained sections of injected skin (Supplementary Figure 8).
Assessment of degranulation by flow cytometry, Wild-type BMCMCs were incubated with different concentrations of S. schenkii yeasts, medium alone as a negative control, or PMA/ionomycin as a positive control, respectively. After 0.5 h of incubation with S. schenkii yeasts, BMCMCs were stained with APC anti-mouse CD63 antibody (143906, Biolegend). APC Rat IgG2a, κ- antibody (400512, Biolegend) was used as isotype control. Cells were analyzed with a Coulter Epics XL Flow cytometer or a Coulter FC 500 ANALYZER (Beckman Coulter). The relevant data were obtained and analyzed using FlowJo software, version 7.6.
Cytokine Release Assay
BMCMCs (5 × 106) were incubated with or without yeasts (9.4 × 107, 4.7 × 107, or 2.35 × 107) of S. schenkii or S. albicans at 37°C in supplemented RPMI 1640 medium under an atmosphere of 5% CO2 for 24 h. Cell-free supernatants were collected and stored at −80°C until assayed for TNF and IL-6. Then, we used ELISA kits (Biolegend) to determine the levels of TNF, IL-6, IL-1β, IL-10, and MCP-1 in cell culture media according to the manufacturer's instructions. Serum levels of TNF and IL-6 in sporotrichosis patients were determined using human TNF or IL-6 enzyme-linked immunosorbent assay kits, respectively (R&D Systems). The results were expressed in pg/ml.
Patients and Control Subjects
Ethical approval from the Ethics Committee of Department of Dermatology, First Hospital, Peking University (Beijing, China) was obtained prior to the study. A total of 15 sporotrichosis patients in Han Chinese population (n = 15, male: 6, female: 9; aged 30–50 years, n = 4, mean 43.75; aged 50–70 years, n = 7, mean 61;aged 70-90 years, n = 4, mean 78;) were included after informed consent (Table 1). The definitive diagnosis of sporotrichosis was established by isolation of the fungus from skin specimens (exudate, scales, and skin biopsy), which were sent to the Laboratory of Mycology. The definitive diagnosis was made by isolation of Sprothrix species as previously described. The control group consisted of 15 plastic surgery-derived skin samples of healthy individuals.
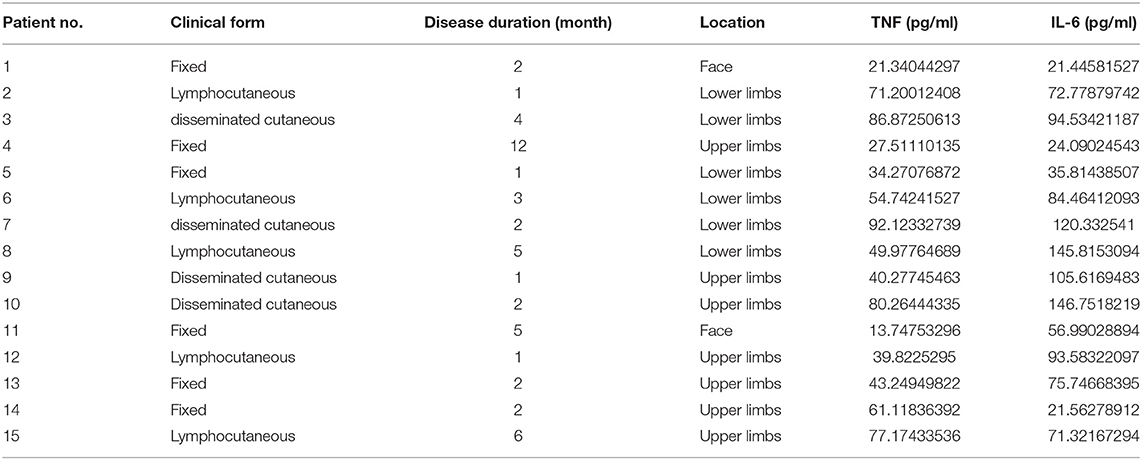
Table 1. Clinical data for the 15 sporotrichosis cases in the present study.
Statistical Analysis
All results are presented as means ± SEM unless specified otherwise. The two-tailed t-test and one-way or two-way ANOVA were used to determine significance between two groups or multiple groups. A P-value ≤ 0.05 was considered to reflect statistical significance.
Results
Mast Cell-Competent Mice Are More Susceptible to S. schenckii Infection Than Mast Cell-Deficient Mice
MC-deficient HK (Hello Kitty) mice and WT (Wild type) mice both developed typical skin sporotrichosis lesions with ulceration and scarring after subcutaneous inoculation with S. schenckii (Figure 1A). However, WT mice infected with S. schenckii developed markedly larger skin lesions compared with HK mice (Figures 1A,B). Differences were most pronounced on day 4 (260 ± 35 vs. 199 ± 42.1 mm3, p = 0.03) and on day 7 post-infection (280 ± 46 vs. 211 ± 12 mm3, p = 0.02), with lesion volumes decreased some 25% in HK mice vs. WT mice (Figure 1B). WT mice also exhibited increased lesional fungal burden, more than double as compared to HK mice on day 7 (Figure 1C; 49 ± 13 vs. 20 ± 6 CFU/mm3, p = 0.002). No inflammation or spores were found in the lungs, spleens, or livers of any of the mice (Supplementary Figure 6A). WT and HK mice showed no spores and similar levels of immune cell infiltration in the draining lymph nodes of sites of infection (Supplementary Figure 6B).
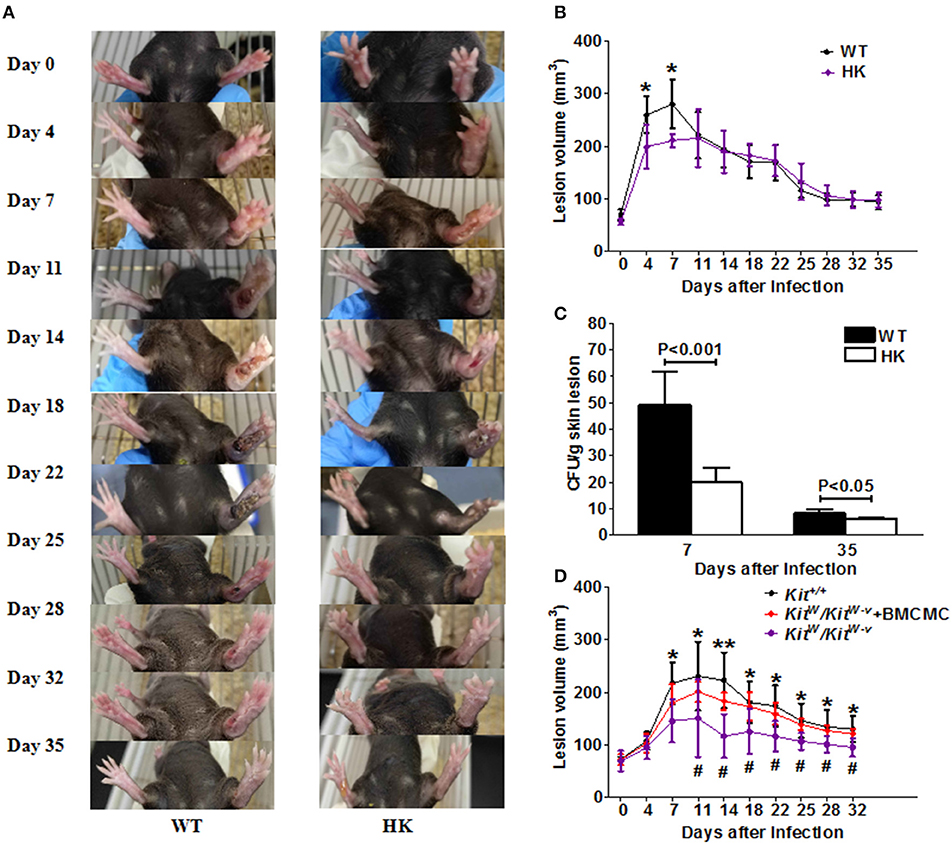
Figure 1. S. schenkii infection in MC-deficient mice. (A–C) On day 0, 3 × 107 yeasts of S. schenkii were injected into the right foot pad of MC-deficient (Hello Kitty, HK, Cpa3-Cre/Mcl-1fl/fl) mice and WT littermate control mice. Lesion development was monitored 3-dimensionally over the course of 35 days and calculated as an ellipsoid. (A) Representative pictures of S. schenkii infection in WT and HK mice during the first 35 days. (B) Lesional volume and (C) fungal burden of skin lesions were assessed at different time points in infected HK and WT mice. One of 3 experiments with six to eight mice each and similar results is shown. *P < 0.05. (D) On day 0, 3 × 106 conidias of S. schenkii were injected into the right foot pad of MC-deficient C57BL/6-KitW/KitW−v mice and WT (Kit+/+) littermate control mice. Locally MC-reconstituted KitW/KitW−v mice were also used for infections, and the volume of skin lesions was estimated at different time points after infection. Data are expressed as mean ± SEM. Kit+/+ vs. KitW/KitW−v, *P < 0.05, **P < 0.01; KitW/KitW−v + BMCMC vs. KitW/KitW−v, #P < 0.05, ##P < 0.01. Three independent experiments with six to eight mice per group were performed.
MC-competent WT mice also developed larger sporotrichosis lesions as compared to MC-deficient C57BL/6-KitW/KitW−v mice infected with S. schenckii (Figure 1D). Reconstitution of skin MCs by local adoptive transfer to C57BL/6-KitW/KitW−v mice 4 weeks prior to infection resulted in skin lesions comparable to those of WT mice (Figure 1D). In WT mice, infection with S. schenckii resulted in 2-fold increased numbers of MCs at sites of infection, but not non-infected skin sites (Supplementary Figures 1A,B).
S. schenckii Induces MC Cytokine Release, but Not Degranulation
To investigate how MCs contribute to S. schenckii infections, we tested if MCs degranulate in response to S. schenckii, in vitro and in situ. MCs did not degranulate in vitro as assessed by β-hexosaminidase release and CD63 surface expression by flow cytometry in BMCMCs (Figures 2A,B and Supplementary Figure 2A). Also, MCs did not exhibit signs of degranulation in response to S. schenckii in situ, as assessed by quantitative histomorphometric analyses of infected skin sites (Supplementary Figure 1C). In contrast, MCs dose-dependently released substantial amounts of the early response cytokines IL-6, TNF, IL-10, and IL-1β, but not MCP-1, in response to S. schenckii (Figures 2C–F and Supplementary Figure 2B). MC-deficient mice showed markedly reduced expression of TNF and IL-6 at sites of S. schenckii infection as compared to WT mice (Figure 3). In addition, in WT mice, most cutaneous MCs at sites of S. schenckii infection were positive for IL-6 and/or TNF, and most IL-6/TNF positive cells were MCs (Figures 4, 5 and Supplementary Figure 7A).
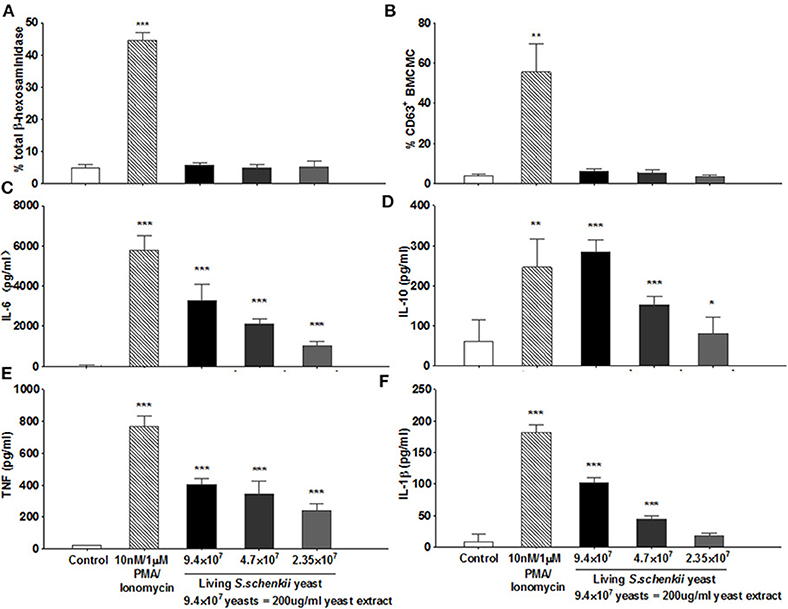
Figure 2. Analysis of MC degranulation and activation following exposure to yeasts from S. schenckii. (A) β hexosaminidase release by MCs following incubation with supplemented RPMI medium (control), PMA/Ionomycin (10 nM/1 μM), or yeasts of S. schenckii for 30 min at 37°C. Values of mediators released from cells incubated with RPMI medium and with PMA/Ionomycin were indicative of spontaneous and degranulation-induced secretion, respectively. N = 4. (B) WT BMCMCs were incubated with increasing concentrations of S. schenkii yeasts, medium alone as negative control, or PMA/Ionomycin as positive control. Expression of CD63 on BMCMCs after 0.5 h of incubation was measured by Flow cytometry, N = 6. The release of (C) IL-6, (D) IL-10, (E) TNF, and (F) IL-1β into the supernatants after 24 h of incubation was measured by ELISA, N = 3. Data expressed as Mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001 were compared with the control group.
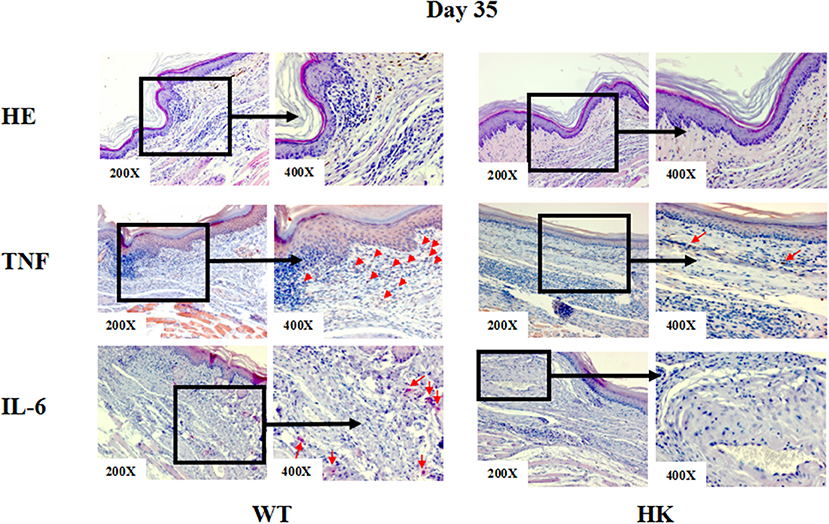
Figure 3. Representative immunohistochemical staining of skin lesions from S. schenckii infected WT and HK mice at day 35. Representative immunohistochemical staining of TNF and IL-6 and with HE of skin lesions from S. schenckii and S. albicans-infected mice at day 35. Red arrows: Positive cells.
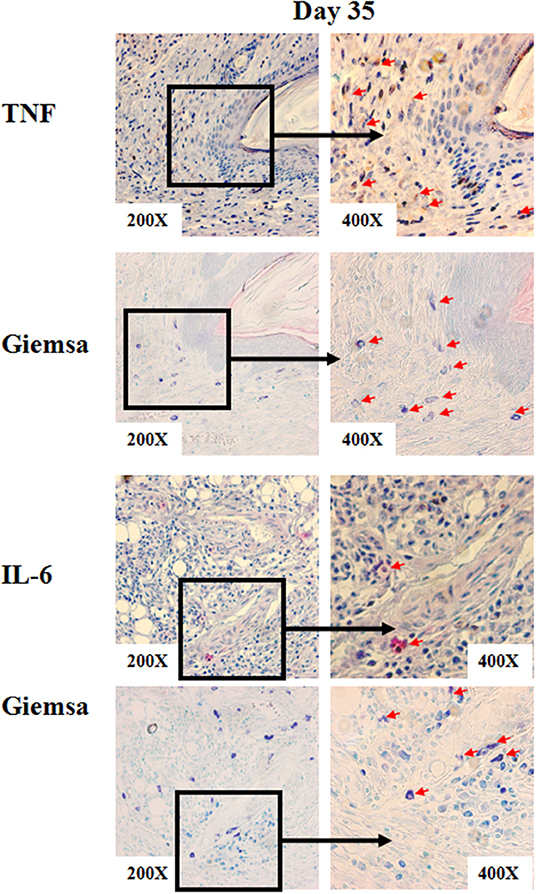
Figure 4. Representative immunohistochemical staining of skin lesions from S. schenckii-infected WT mice at day 35. Representative immunohistochemical staining of TNF and IL-6 and with Giemsa of skin lesions from S.schenckii-infected WT mice at day 35. Red arrows: Positive cells.
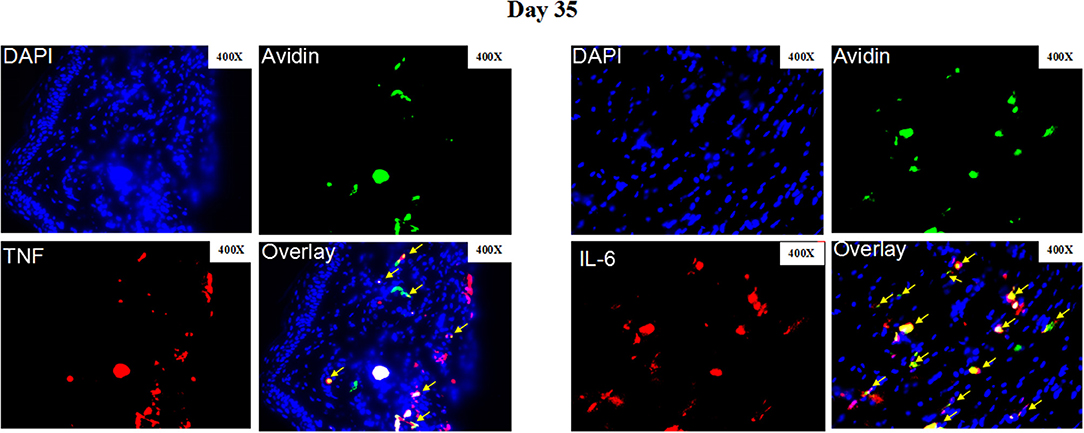
Figure 5. Representative immunofluorescence staining of skin lesions from S. schenckii-infected WT mice at day 35. Representative immunofluorescence staining TNF (Red), IL-6 (Red), mast cells (Avidin, green), and DAPI (Blue, nuclei) of skin lesions from S. schenckii-infected WT mice at day 35. Red arrows: Double positive cells. Original magnification X40.
Sporothrix Virulence Is Linked to the Induction of MC Cytokine Release
To assess the relevance of MC cytokine production in sporotrichosis, we used S. schenckii and the less virulent Sporothrix species S. albicans. We first compared skin responses of mice and then the release of cytokines of MCs exposed to these two Sporothrix species. Mice infected with S. schenckii developed markedly larger skin lesions than mice infected with S. albicans (Figure 6A, 175 ± 38 vs. 112 ± 45 mm3 at day 7, p = 0.02). In addition, S. schenckii-infected mice, but not S. albicans-infected mice developed necrosis, with scabbing and ulcerations at sites of infection (Supplementary Figure 3).

Figure 6. The response of MCs to S. schenkii and S. albicans. (A) S. schenkii and S. albicans infection in WT mice. 3 × 107 yeasts of S. schenkii or S. albicans were injected into the right foot pad of WT control mice. Lesion development was monitored 3-dimensionally over the course of 35 days and calculated as an ellipsoid. Data are expressed as mean ± SEM. WT BMCMCs were incubated with increasing concentrations of S. schenkii or S. albicans yeasts, respectively. Release of (B) TNF and (C) IL-6 into the supernatants after 24 h of incubation was measured by ELISA, N = 3. The numbers on top of the bar mean the fold increase of cytokines induced by S. schenckii vs. S. albicans. (D) Representative immunohistochemical staining TNF and IL-6 of skin lesions from S. schenckii and S. albicans infected mice at day 35. Red arrows: Staining positive cells. Three independent experiments with six to eight mice per group were performed. *P < 0.05.
When compared for their effects on MC cytokine release, S. schenkii induced more than 11-fold and 8-fold higher release of IL-6 and TNF, respectively, as compared to S. albicans (Figures 6B,C), whereas IL-10 and IL-1ß release were similar (Supplementary Figures 4A,B). Immunohistochemical analyses showed that TNF and IL-6 were also increased in the skin lesions of S.schenckii-infected mice as compared to S. albicans-infected mice (Figure 6D and Supplementary Figure 5A).
TNF and IL-6 Are Increased and Linked to Disease Activity in Patients With Sporotrichosis
Patients with sporotrichosis showed markedly higher serum levels of TNF and IL-6 than healthy controls (HC; Figure 7; TNF-α: 53 ± 24 vs. 6 ± 10 pg/ml, P < 0.001; IL-6: 80 ± 42 vs. 25 ± 10 pg/ml, P < 0.001). Serum levels of TNF and IL-6 were highest in patients of disseminated type sporotrichosis, followed by patients with lymphocutaneous sporotrichosis, and lowest in fixed cutaneous sporotrichosis patients (Figures 7A,B). Expression levels of TNF and IL-6 in the lesional skin of sporotrichosis patients were markedly higher as compared to healthy control skin (Figure 7C and Supplementary Figure 5B). Some cutaneous IL-6/TNF-positive cells were MCs, and most MCs were IL-6/TNF-positive (Figures 7C, 8 and Supplementary Figure 7B).
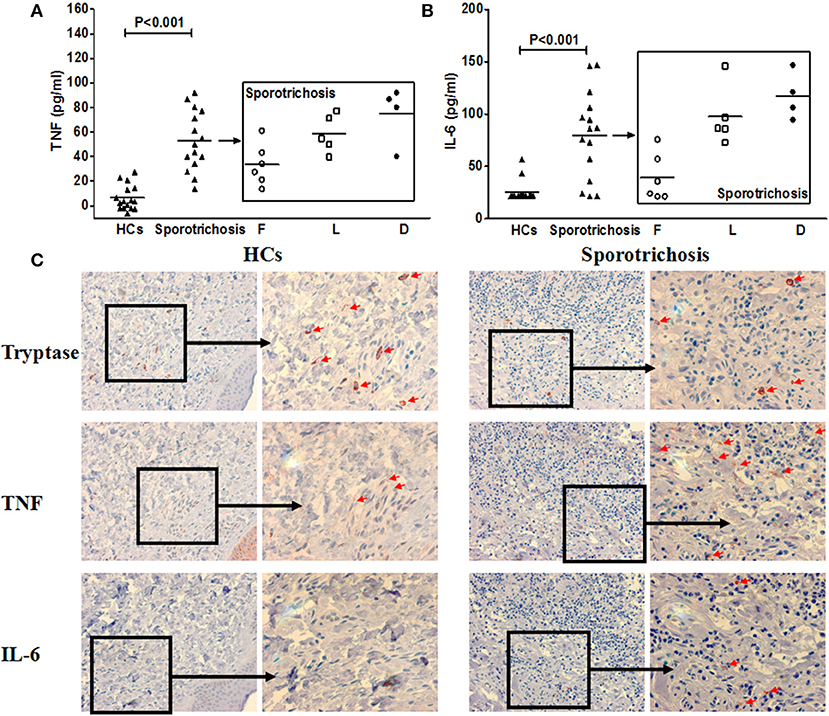
Figure 7. TNF and IL-6 expression in patients with sporotrichosis. Clinical forms of Sporotrichosis: F, fixed; L, lymphocutaneous; D, disseminated cutaneous. Serum concentrations of TNF (A) and IL-6 (B) in sporotrichosis patients. (C) Representative immunohistochemical staining of tryptase, TNF and IL-6 positive cells in the lesional skin of sporotrichosis patients and healthy control skin.
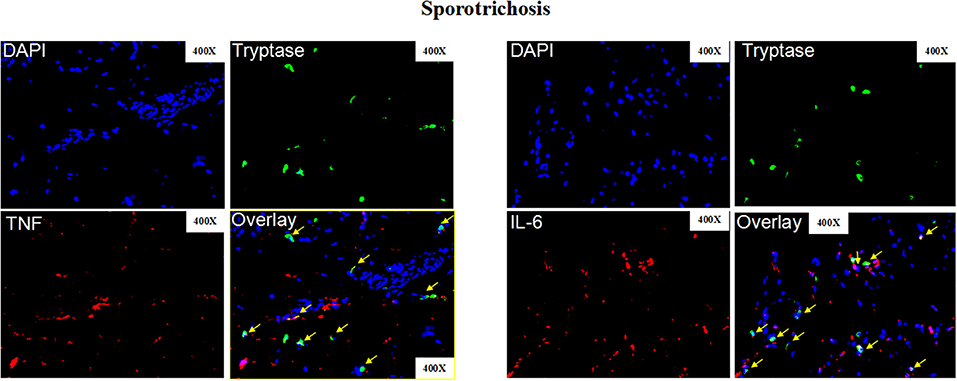
Figure 8. Representative immunofluorescence staining of skin lesions from patients with sporotrichosis. Representative immunofluorescence staining TNF (Red), IL-6 (Red), mast cells (Tryptase, green), and DAPI (Blue, nuclei) of skin lesions from patients with sporotrichosis. Red arrows: Double positive cells. Original magnification X40.
Discussion
Our study is the first to show that MCs contribute to the pathogenesis of skin sporotrichosis. Mice with normal MC populations that are infected with S. schenckii develop larger sporotrichosis lesions than MC-deficient mice. This is, at least in part, due to the production of the proinflammatory cytokines IL-6 and TNF by MCs. MCs showed pronounced production of these cytokines when exposed to S. schenckii in vitro or in vivo, and the severity of S. schenckii infections correlates, in mice and humans with IL-6 and TNF levels.
To investigate the role of MCs in sporotrichosis, we used a modified mouse model with subcutaneous injections of S. schenckii into the foot pad of mouse (31, 32). This model shows many features of skin sporotrichosis in patients including ulceration, scarring, and crusty lesions, as well as nodules at infected skin sites (23, 33). By using this model, we found that MC-deficient mice are significantly less susceptible to S. schenckii infections: They developed smaller lesions at the site of infection and had lower fungal burden at the peak of infection development. And the differences between MC-deficient mice and WT controls are relatively small at most time points, this may be related to the development process of S. schenckii infection. Of importance, our model appears to be well-suited to study acute S. schenckii infections of the skin, whereas studies on chronic Sporothrix infections, which are common in patients, may require the development of a different model.
Notably, we used two independent MC-deficient mouse models to investigate the role of MCs in sporotrichosis. One was the KIT-independent HK mouse, the other was the Classical KIT-dependent KitW/KitW−v mouse. Both models showed very similar results. As KitW/KitW−v mice that had been repaired for their skin MC deficiency by adoptive transfer before S. schenckii infection showed significantly larger lesions than the non-repaired mice, the effects observed in the KitW/KitW−v mouse model appear to be MC-dependent. Our results are supported by an earlier study that showed that the depletion of MCs in mice prior to being subjected to sporotrichosis decreases the fungal load in organs and significantly reduces the severity of infections (19).
Our study does not address through which receptors MCs are activated by S. schenckii. MCs have repeatedly been shown to respond to various pathogens through interacting with their toll like receptor (TLR) ligands by secreting cytokines, chemokines, and lipid mediators, while little or no degranulation occurs (34). In addition, fungus-specific IgE-mediated cross linking of the high affinity IgE receptor, FcεRI, can also result in MC degranulation and the release of preformed mediators as well as the de novo synthesis of lipid mediators and cytokines (7). Recently, it has been shown that both yeasts and hyphae of C. albica activate MCs through TLR and dectin-1 (12). Thus, MCs may detect S. schenckii by TLRs (e.g., TLR-2 and/or TLR-4) or C-type lectin receptors, but further studies are needed to confirm this (35–38).
How do MCs contribute to S. schenkii injections? Our findings, together with those of a previous study on peritoneal MCs (22), suggest that the de novo synthesis and secretion of proinflammatory cytokines including TNF, IL-6, IL-10, and IL-1β by skin MCs activated by S. schenckii promotes skin inflammation, thereby aggravating skin sporotrichosis. This notion is based, in part, on the results of our comparisons of sporotrichosis and MC responses to S. schenckii and the less virulent sporothrix species S. albicans. First, S. schenkii yeasts induced more TNF and IL-6 release from MCs compared with S. albicans at the same density. Second, MC-deficient mice showed markedly reduced expression of TNF and IL-6 at sites of S. schenckii infection as compared to WT mice. Third, most cutaneous MCs at sites of skin sporotrichosis infection are positive for IL-6 and/or TNF. How exactly do TNF and IL-6 released from MCs make sprorotrichosis worse? The molecular mechanisms remain to be investigated and identified. Importantly, other innate immune cells such as macrophages, neutrophils, and dendritic cells have also been described to release TNF and IL-6 in response to S. schenckii infection (32, 39). In addition, both HK and KitW/Wv mice have reduced basophil numbers. Basophils have been shown in several studies to become activated in an IgE-dependent and -independent manner to various fungi (40–42). Therefore, a contribution of basophils cannot be excluded and requires further investigation. The role of basophils in S. schenckii infection is still worthy of further investigation.
Are MCs, TNF, and IL-6 involved in human sporotrichosis? The results of our analyses of TNF and IL-6 expression in sporotrichosis patients suggest that this is so. We found the levels of both cytokines to be higher in the serum and skin lesions of affected patients. This is in line with earlier reports that suggest that high levels of TNF, IL-6, and other cytokines can contribute to the pathology of S. schenckii infection (43–45). On the other hand, these studies also suggest that very low levels of these cytokines can be harmful in S. schenckii infections (43–45). Also, two case reports describe the development of sporotrichosis in patients treated with TNF antagonists (46, 47), whereas there are no reported cases of treatment for sporotrichosis with a TNF antagonist or anti-IL-6. Finally, TNF and IL-6, in the context of sporotrichosis, may also be produced by cells other than MCs, for example T cells and macrophages (43, 48, 49). Taken together, our results suggest, but do not prove, that MC-derived TNF and IL-6 contribute to the pathology of sporotrichosis. The use of drugs that target these cytokines may be promising treatment strategies in patients with sporotrichosis who fail standard therapy.
Our study has several strengths and some limitations. On the plus side, we used two different mouse models of mast cell-deficiency, one KIT-independent, the other KIT-dependent, the latter one complemented by adoptively transferring MCs to MC-deficient recipient mice. Also, we complemented the results from our mouse experiments with human studies. As for limitations, we relied on formalin-fixed skin for our histomorphometric analyses, and not all mast cells, especially in inflamed skin, may be visualized with this fixation. Also, what our current report does not do is provide insights on how MCs are activated by S. schenckii, how MCs and the release of cytokines make sporotrichosis worse, and on whether or not MC-targeted treatment or the use of antagonists of MC-derived cytokines can improve the course of sporotrichosis. These questions will have to be answered by ongoing and future studies, including studies that make use of TNF or IL-6 knockout mice or siRNA technology.
Taken together, our present study provides evidence that MCs exacerbate mouse and human skin S. schenckii infection and sporotrichosis by releasing TNF and IL-6. Both of these cytokines should be explored for their value as therapeutic targets in the treatment of patients with sporotrichosis. Also, our findings support the notion that the role of MCs in innate immunity to fungal and other pathogens may be more complex than previously thought and that the characterization of MC responses in skin fungal infections merits further investigation.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
The studies involving human participants were reviewed and approved by Department of Dermatology, First Hospital, Peking University, Beijing 100034, China. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by Charite - Universitätsmedizin Berlin.
Author Contributions
Conceptualization and funding acquisition: MM, ZZ, and QJ. Methodology: ZZ, JS, and PG. Software: YL and YW. Validation, formal analysis, and investigation: YL and QJ. Resources: MM, ZZ, JS, and PG. Data curation: QJ, YL, and YW. Writing: QJ. Writing (review and editing): SF, ZZ, RL, and MM. Visualization: QJ, SF, and MM. Supervision: MM, ZZ, and JS. Project administration: MM and ZZ.
Funding
This work was supported by a joint Sino-German research project grant (No. GZ901) to MM (DFG MA1909/17-1) and ZZ as well as by the Jiangsu Provincial Medical Youth Talent programme to QJ (No. QNRC2016736).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank José Roberto Fogaça de Almeida for his assistance in the preparation of S. schenckii. We thank the Sino-German Center for Research Promotion (SGC) for continued help and support. This work was supported by a joint Sino-German research project grant (No. GZ901) to MM (DFG MA1909/17-1) and ZZ as well as by the Jiangsu Provincial Medical Youth Talent programme to QJ (No. QNRC2016736). We also acknowledge support from the Open Access Publication Fund of Charité-Universitätsmedizin Berlin.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.00469/full#supplementary-material
Supplementary Figure 1. MCs in skin tissues of mice. Giemsa staining was used to detect MCs in skin tissues of mice. (A) Number [mean (S. D.)] of Giemsa-positive cells in skin tissues of left and right foot pad of S. schenkii infected WT mice, respectively. (B) Representative immunohistological pictures of Giemsa staining in mouse foot pads from S. schenkii infected WT mice. (C) Skin tissue from the back of WT mice was incubated with increasing concentrations of S. schenkii yeasts, medium alone and PMA/Ionomycin as positive control, respectively. Giemsa staining was used to detect the degranulation of MCs in mouse skin tissue. Shown are representative images from three independent experiments.
Supplementary Figure 2. Analysis of CD63 expression and cytokines release of MC in response to yeasts of S. schenckii. WT BMCMCs were incubated with increasing concentrations of S. schenkii yeasts, medium alone as negative control, or PMA/ionomycin as positive control, respectively. (A) Representative flow cytometry analysis of CD63 on BMCMCs after 0.5 h of incubation, N = 6. (B) Release MCP-1 into the supernatants after 24 h of incubation was measured by ELISA, N = 3. ***P < 0.001 was compared with the control group.
Supplementary Figure 3. S. schenkii and S. albicans infection in WT mice. 3 × 107 yeasts of S. schenkii or S. albicans were injected into the right foot pad of WT control mice. Representative pictures of S. schenkii or S. albicans infection in WT mice during the 35 days.
Supplementary Figure 4. The response of MCs to S. schenkii and S. albicans. WT BMCMCs were incubated with increasing concentrations of S. schenkii or S. albicans, respectively. Release of (A) IL-1β and (B) IL-10 into the supernatants after 24 h of incubation was measured by ELISA, N = 3. The numbers on top of the bar mean the fold increase of cytokines induced by S. schenckii vs. S. albicans.
Supplementary Figure 5. TNF and IL-6 expression in skin tissues. (A) Average of TNF or IL-6 positive stained area percentage in skin tissues of right foot pad of S. schenkii or S. albicans infected WT mice, respectively. (B) Average of Tryptase, TNF or IL-6 positive stained area percentage in the lesional skin of sporotrichosis patients and healthy control skin, respectively.
Supplementary Figure 6. Representative HE staining of internal organs in mice with S. schenkii infection. (A) Representative immunohistological pictures of HE staining in mice from S. schenkii infected WT and HK mice. (B) Representative HE staining pictures from skin tissues of WT mice with injection of S. schenkii yeasts. Shown are representative images from three independent experiments.
Supplementary Figure 7. Co-localization of mast cells and IL-6 or TNF expression in skin tissues. (A) Average percentage of IL-6 or TNF positive stained cells on mast cells (Avidin positive) or Avidin positive stained cells (Mast cells) on IL-6 or TNF positive stained cells in skin tissues of right foot pad of S. schenkii infected WT mice at day 35, respectively. (B) Average percentage of IL-6 or TNF positive stained cells on mast cells (Tryptase positive) or Tryptase positive stained cells (Mast cells) on IL-6 or TNF positive stained cells in the lesional skin of sporotrichosis patients, respectively.
Supplementary Figure 8. Number of MCs in skin tissues of mice. Mast cell was counted in randomly selected fields (X200) of baseline control after 4 wk in KitW/KitW−v reconstituted with wild type MCs, There is no difference in the mast cell number in skin of WT mice and KitW/KitW−v mice reconstituted with wild type mast cells 4 wk after adoptive transfer. Three independent experiments with 6 mice per group were performed for each experiment.
References
1. Bonifaz A, Vazquez-Gonzalez D. Sporotrichosis: an update. G Ital Dermatol Venereol. (2010) 145:659–73.
4. Koga T, Matsuda T, Matsumoto T, Furue M. Therapeutic approaches to subcutaneous mycoses. Am J Clin Dermatol. (2003) 4:537–43. doi: 10.2165/00128071-200304080-00003
5. Abraham SN, St John AL. Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol. (2010) 10:440–52. doi: 10.1038/nri2782
6. St John AL, Abraham SN. Innate immunity and its regulation by mast cells. J Immunol. (2013) 190:4458–63. doi: 10.4049/jimmunol.1203420
7. Jiao Q, Luo Y, Scheffel J, Zhao Z, Maurer M. The complex role of mast cells in fungal infections. Exp Dermatol. (2019) 28:749–55. doi: 10.1111/exd.13907
8. Nosal R, Novotny J, Sikl D. The effect of glycoprotein from Candida albicans on isolated rat mast cells. Toxicon. (1974) 12:103–8. doi: 10.1016/0041-0101(74)90233-5
9. Trevisan E, Vita F, Medic N, Soranzo MR, Zabucchi G, Borelli V. Mast cells kill Candida albicans in the extracellular environment but spare ingested fungi from death. Inflammation. (2014) 37:2174–89. doi: 10.1007/s10753-014-9951-9
10. Yan Y, Zhao Z, Wan H, Wu R, Fang J, Liu H. A novel fungus concentration-dependent rat model for acute invasive fungal rhinosinusitis: an experimental study. BMC Infect Dis. (2014) 14:3856. doi: 10.1186/s12879-014-0713-y
11. Lopes JP, Stylianou M, Nilsson G, Urban CF. Opportunistic pathogen Candida albicans elicits a temporal response in primary human mast cells. Sci Rep. (2015) 5:12287. doi: 10.1038/srep12287
12. Pinke KH, Lima HG, Cunha FQ, Lara VS. Mast cells phagocyte Candida albicans and produce nitric oxide by mechanisms involving TLR2 and Dectin-1. Immunobiology. (2016) 221:220–7. doi: 10.1016/j.imbio.2015.09.004
13. Kauffman HF. Immunopathogenesis of allergic bronchopulmonary aspergillosis and airway remodeling. Front Biosci. (2003) 8:e190–196. doi: 10.2741/990
14. Noverr MC, Noggle RM, Toews GB, Huffnagle GB. Role of antibiotics and fungal microbiota in driving pulmonary allergic responses. Infect Immun. (2004) 72:4996–5003. doi: 10.1128/IAI.72.9.4996-5003.2004
15. Batista AC, Soares CT, Lara VS. Failure of nitric oxide production by macrophages and decrease in CD4+ T cells in oral paracoccidioidomycosis: possible mechanisms that permit local fungal multiplication. Rev Inst Med Trop Sáo Paulo. (2005) 47:267–73. doi: 10.1590/S0036-46652005000500006
16. Pagliari C, Fernandes ER, Guedes F, Alves C, Sotto MN. Role of mast cells as IL10 producing cells in paracoccidioidomycosis skin lesions. Mycopathologia. (2006) 162:331–5. doi: 10.1007/s11046-006-0069-y
17. Selander C, Engblom C, Nilsson G, Scheynius A, Andersson CL. TLR2/MyD88-dependent and -independent activation of mast cell IgE responses by the skin commensal yeast Malassezia sympodialis. J Immunol. (2009) 182:4208–16. doi: 10.4049/jimmunol.0800885
18. Ribbing C, Engblom C, Lappalainen J, Lindstedt K, Kovanen PT, Karlsson MA, et al. Mast cells generated from patients with atopic eczema have enhanced levels of granule mediators and an impaired Dectin-1 expression. Allergy. (2011) 66:110–9. doi: 10.1111/j.1398-9995.2010.02437.x
19. Romo-Lozano Y, Hernandez-Hernandez F, Salinas E. Mast cell activation by conidia of Sporothrix schenckii: role in the severity of infection. Scand J Immunol. (2012) 76:11–20. doi: 10.1111/j.1365-3083.2012.02706.x
20. Hiragun T, Ishii K, Hiragun M, Suzuki H, Kan T, Mihara S, et al. Fungal protein MGL_1304 in sweat is an allergen for atopic dermatitis patients. J Allergy Clin Immunol. (2013) 132:608–15.e4. doi: 10.1016/j.jaci.2013.03.047
21. Niranjan R, Mavi P, Rayapudi M, Dynda S, Mishra A. Pathogenic role of mast cells in experimental eosinophilic esophagitis. Am J Physiol Gastrointest Liver Physiol. (2013) 304:G1087–94. doi: 10.1152/ajpgi.00070.2013
22. Romo-Lozano Y, Hernandez-Hernandez F, Salinas E. Sporothrix schenckii yeasts induce ERK pathway activation and secretion of IL-6 and TNF-alpha in rat mast cells, but no degranulation. Med Mycol. (2014) 52:862–8. doi: 10.1093/mmy/myu055
23. Ishii K, Hiragun M, Hiragun T, Kan T, Kawaguchi T, Yanase Y, et al. A human monoclonal IgE antibody that binds to MGL_1304, a major allergen in human sweat, without activation of mast cells and basophils. Biochem Biophys Res Commun. (2015) 468:99–104. doi: 10.1016/j.bbrc.2015.10.154
24. Barbosa-Lorenzi VC, Peyda S, Scheynius A, Nilsson G, Lunderius-Andersson C. Curdlan induces selective mast cell degranulation without concomitant release of LTC4, IL-6 or CCL2. Immunobiology. (2017) 222:647–50. doi: 10.1016/j.imbio.2016.12.001
25. Hiragun T, Hiragun M, Ishii K, Kan T, Hide M. Sweat allergy: extrinsic or intrinsic? J Dermatol Sci. (2017) 87:3–9. doi: 10.1016/j.jdermsci.2017.03.002
26. Saluja R, Metz M, Maurer M. Role and relevance of mast cells in fungal infections. Front Immunol. (2012) 3:146. doi: 10.3389/fimmu.2012.00146
27. Kauffman CA, Bustamante B, Chapman SW, Pappas PG, Infectious Diseases Society Of A. Clinical practice guidelines for the management of sporotrichosis: 2007 update by the Infectious Diseases Society of America. Clin Infect Dis. (2007) 45:1255–65. doi: 10.1086/522765
28. Sassa MF, Saturi AE, Souza LF, Ribeiro LC, Sgarbi DB, Carlos IZ. Response of macrophage Toll-like receptor 4 to a Sporothrix schenckii lipid extract during experimental sporotrichosis. Immunology. (2009) 128:301–9. doi: 10.1111/j.1365-2567.2009.03118.x
29. Fernandes GF, Dos Santos PO, Rodrigues AM, Sasaki AA, Burger E, De Camargo ZP. Characterization of virulence profile, protein secretion and immunogenicity of different Sporothrix schenckii sensu stricto isolates compared with S. globosa and S. brasiliensis species. Virulence. (2013) 4:241–9. doi: 10.4161/viru.23112
30. Alba-Fierro CA, Perez-Torres A, Toriello C, Romo-Lozano Y, Lopez-Romero E, Ruiz-Baca E. Molecular components of the Sporothrix schenckii complex that induce immune response. Curr Microbiol. (2016) 73:292–300. doi: 10.1007/s00284-016-1045-5
31. De Almeida JRF, Jannuzzi GP, Kaihami GH, Breda LCD, Ferreira KS, De Almeida SR. An immunoproteomic approach revealing peptides from Sporothrix brasiliensis that induce a cellular immune response in subcutaneous sporotrichosis. Sci Rep. (2018) 8:4192. doi: 10.1038/s41598-018-22709-8
32. Alba-Fierro CA, Perez-Torres A, Toriello C, Pulido-Camarillo E, Lopez-Romero E, Romo-Lozano Y, et al. Immune response induced by an immunodominant 60 kDa glycoprotein of the cell wall of Sporothrix schenckii in two mice strains with experimental sporotrichosis. J Immunol Res. (2016) 2016:6525831. doi: 10.1155/2016/6525831
33. Mahajan VK. Sporotrichosis: an overview and therapeutic options. Dermatol Res Pract. (2014) 2014:272376. doi: 10.1155/2014/272376
34. Silver R, Curley JP. Mast cells on the mind: new insights and opportunities. Trends Neurosci. (2013) 36:513–21. doi: 10.1016/j.tins.2013.06.001
35. Sandig H, Bulfone-Paus S. TLR signaling in mast cells: common and unique features. Front Immunol. (2012) 3:185. doi: 10.3389/fimmu.2012.00185
36. Sassa MF, Ferreira LS, Ribeiro LC, Carlos IZ. Immune response against Sporothrix schenckii in TLR-4-deficient mice. Mycopathologia. (2012) 174:21–30. doi: 10.1007/s11046-012-9523-1
37. De CNT, Ferreira LS, Arthur RA, Alegranci P, Placeres MC, Spolidorio LC, et al. Influence of TLR-2 in the immune response in the infection induced by fungus Sporothrix schenckii. Immunol Invest. (2014) 43:370–90. doi: 10.3109/08820139.2013.879174
38. Agier J, Zelechowska P, Kozlowska E, Brzezinska-Blaszczyk E. Expression of surface and intracellular Toll-like receptors by mature mast cells. Cent Eur J Immunol. (2016) 41:333–8. doi: 10.5114/ceji.2016.65131
39. Carlos IZ, Sassa MF, Da Graca Sgarbi DB, Placeres MC, Maia DC. Current research on the immune response to experimental sporotrichosis. Mycopathologia. (2009) 168:1–10. doi: 10.1007/s11046-009-9190-z
40. Reber LL, Marichal T, Galli SJ. New models for analyzing mast cell functions in vivo. Trends Immunol. (2012) 33:613–25. doi: 10.1016/j.it.2012.09.008
41. Mukai K, Karasuyama H, Kabashima K, Kubo M, Galli SJ. Differences in the importance of mast cells, basophils, IgE, and IgG versus that of CD4(+) T cells and ILC2 cells in primary and secondary immunity to strongyloides venezuelensis. Infect Immun. (2017) 85:e00053–17. doi: 10.1128/IAI.00053-17
42. Shen SS, Qu XY, Zhang WZ, Li J, Lv ZY. Infection against infection: parasite antagonism against parasites, viruses and bacteria. Infect Dis Poverty. (2019) 8:49. doi: 10.1186/s40249-019-0560-6
43. Carlos IZ, Zini MM, Sgarbi DB, Angluster J, Alviano CS, Silva CL. Disturbances in the production of interleukin-1 and tumor necrosis factor in disseminated murine sporotrichosis. Mycopathologia. (1994) 127:189–94. doi: 10.1007/BF01102920
44. Da Silva AC, Bezerra LM, Aguiar TS, Tavares D, Araujo LM, Pinto CE, et al. Effect of genetic modifications by selection for immunological tolerance on fungus infection in mice. Microbes Infect. (2001) 3:215–22. doi: 10.1016/S1286-4579(01)01373-9
45. Fernandes KS, Neto EH, Brito MM, Silva JS, Cunha FQ, Barja-Fidalgo C. Detrimental role of endogenous nitric oxide in host defence against Sporothrix schenckii. Immunology. (2008) 123:469–79. doi: 10.1111/j.1365-2567.2007.02712.x
46. Gottlieb GS, Lesser CF, Holmes KK, Wald A. Disseminated sporotrichosis associated with treatment with immunosuppressants and tumor necrosis factor-alpha antagonists. Clin Infect Dis. (2003) 37:838–40. doi: 10.1086/377235
47. Ursini F, Russo E, Leporini C, Calabria M, Bruno C, Tripolino C, et al. Lymphocutaneous sporotrichosis during treatment with anti-TNF-alpha monotherapy. Case Rep Rheumatol. (2015) 2015:614504. doi: 10.1155/2015/614504
48. Tachibana T, Matsuyama T, Mitsuyama M. Involvement of CD4+ T cells and macrophages in acquired protection against infection with Sporothrix schenckii in mice. Med Mycol. (1999) 37:397–404. doi: 10.1046/j.1365-280X.1999.00239.x
Keywords: Sporothrix schenckii, sporotrichosis, skin, mast cells, TNF, IL-6
Citation: Jiao Q, Luo Y, Scheffel J, Geng P, Wang Y, Frischbutter S, Li R, Maurer M and Zhao Z (2020) Skin Mast Cells Contribute to Sporothrix schenckii Infection. Front. Immunol. 11:469. doi: 10.3389/fimmu.2020.00469
Received: 05 October 2019; Accepted: 28 February 2020;
Published: 19 March 2020.
Edited by:
Jagadeesh Bayry, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Jean Sylvia Marshall, Dalhousie University, CanadaLaurent Reber, INSERM U1043 Centre de Physiopathologie de Toulouse Purpan, France
Copyright © 2020 Jiao, Luo, Scheffel, Geng, Wang, Frischbutter, Li, Maurer and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcus Maurer, bWFyY3VzLm1hdXJlckBjaGFyaXRlLmRl; Zuotao Zhao, emhhb3p1b3Rhb3Rhb0AxNjMuY29t
†These authors have contributed equally to this work