 Prathapan Ayyappan1
Prathapan Ayyappan1 Robert Z. Harms1
Robert Z. Harms1 Jennifer A. Seifert2Elizabeth A. Bemis3Marie L. Feser2Kevin D. Deane2
Jennifer A. Seifert2Elizabeth A. Bemis3Marie L. Feser2Kevin D. Deane2 M. Kristen Demoruelle2Ted R. Mikuls4V. Michael Holers2
M. Kristen Demoruelle2Ted R. Mikuls4V. Michael Holers2 Nora E. Sarvetnick1,5*
Nora E. Sarvetnick1,5*- 1Department of Surgery-Transplant, University of Nebraska Medical Center, Omaha, NE, United States
- 2Division of Rheumatology, University of Colorado-Denver, Aurora, CO, United States
- 3Department of Epidemiology, Colorado School of Public Health, University of Colorado Anschutz Medical Campus, Aurora, CO, United States
- 4Division of Rheumatology, University of Nebraska Medical Center, Omaha, NE, United States
- 5Mary and Dick Holland Regenerative Medicine Program, University of Nebraska Medical Center, Omaha, NE, United States
Rheumatoid arthritis (RA) is a chronic progressive autoimmune disease leading to considerable disability over time. The disease can be characterized by the presence of multiple autoantibodies in the serum and synovial fluid. Microbial dysbiosis is proposed to play a role in the pathogenesis of RA. Increased systemic bacterial exposure leads to elevated levels of antimicrobial response factors (ARFs) in the circulation. In the present study, we tested whether RA patients have increased levels of ARFs by analyzing the levels of multiple ARFs in serum from RA patients and healthy age and sex-matched controls. The levels of soluble CD14 (sCD14), lysozyme, and CXCL16 were significantly elevated in RA patients compared to healthy controls. Lipopolysaccharide binding protein (LBP) levels remained unchanged in RA patients compared to healthy controls. A positive correlation of LBP with rheumatoid factor (RF) was also found in RA subjects. Interestingly, the levels of anti-endotoxin core antibodies (EndoCAb) IgM, total IgM, EndoCAb IgA, and total IgA were significantly elevated in RA patients compared to healthy controls. No significant changes in the levels of EndoCAb IgG and total IgG were observed in RA patients compared to healthy controls. Furthermore, lysozyme and CXCL16 levels were positively correlated with disease severity among RA subjects. Increases in the levels of several ARFs and their correlations with clinical indices suggest systemic microbial exposure in the RA cohort. Modulation of microbial exposure may play an important role in disease pathogenesis in individuals with RA.
Introduction
Rheumatoid arthritis (RA) is a chronic progressive autoimmune disease leading to severe disability. Genetic, environmental, and epigenetic factors instigate the production of autoantibodies and the loss of tissue tolerance in RA (1–4). These autoantibodies recognize cartilage components, cellular chaperonins, IgG molecules, and citrullinated proteins (5). Similar to other autoimmune diseases, the disease predominantly occurs in females (4, 6, 7). The disease perturbs the synovial joint lining, which undergoes hyperplasia and inflammation leading to irreversible destruction of articular cartilage, ligaments, and bone (8–10). Frequent involvement of extra-articular tissues including the heart, lungs, skin, eyes, and nervous system is associated with very high levels of autoantibodies and circulating immune complexes (11, 12). Early diagnosis can greatly improve the outcome of RA, but the disease prediction remains a challenge (4).
Recently it was hypothesized that microbial dysbiosis plays a role in the pathogenesis of RA (3, 13–16). Patients with classified RA showed alterations in the gut microbiome with a relative increase in the abundance of Prevotella copri and decrease in Haemophilus spp. compared to healthy controls (14, 17). Alterations in lung microbiota, including increased levels of members of Pseudonocardia suggest that distal airway dysbiosis is also associated with RA (18). A pathogenic role for Porphyromonas gingivalis, an oral commensal was also reported (19, 20). These changes in the gut, oral and lung microbiome could cause the leakage of bacterial products into circulation, promoting inflammation and aggravating disease (20–24).
Constant exposure of microbes in the circulation elicits an antibody response to bacteria and thus can act as a measure of microbial exposure (25). Antibodies directed against multiple bacteria have been found to be elevated in RA patients. For example, circulating antibodies directed against the periodontal bacteria Prevotella intermedia, P. gingivalis, and Bacteroides forsythus were reported (19, 26, 27). Elevated levels of IgA and IgM antibodies directed against Proteus mirabilis were also found in RA patients and were positively correlated with total IgA and total IgM levels (28). Antibodies against members of Enterobacteriaceae and bacterial nucleic acids from P. gingivalis and P. copri were detected in synovial fluid from RA patients (15, 29–31). A role of Aggregatibacter actinomycetemcomitans as a factor in the pathogenesis of RA has also been proposed (32, 33). Persistence of microbial products and elevated levels of antimicrobial antibodies in RA patients further suggests the role of systemic bacterial exposure in the pathogenesis and progression of the disease.
In response to microbial exposure, antimicrobial response factors (ARFs) are released into the circulation to neutralize microbial products. ARFs are diverse pleiotropic molecules that include cytokines, chemokines, anti-endotoxin core antibodies (EndoCAb), peptides, and proteases (34, 35). The bactericidal activity of many ARFs is based on their ability to disrupt the bacterial cell envelope, opsonize targets, and/or inhibit intracellular functions of bacteria. The bacterial functions disrupted by ARFs include respiration, enzyme activation, and protein and nucleic acid synthesis. ARFs also modulate immune responses. For example, ARFs can activate innate immunity by recruiting and/or activating immune cells. Furthermore, some ARFs can regulate Toll-like receptor (TLR) recognition of microbial products (36). These immunomodulatory ARFs can lead to inflammation and tissue damage in the host (37).
In the present study, we tested whether RA patients have increased levels of ARFs by analyzing the levels of multiple ARFs in serum from RA patients and healthy age- and sex-matched controls. Increased levels of ARFs may indicate an increase in systemic bacterial exposure. The ARFs tested include soluble CD14 (sCD14), lipopolysaccharide-binding protein (LBP), lysozyme, CXCL16, EndoCAb IgG, EndoCAb IgA, and EndoCAb IgM. Our results revealed a marked elevation of several ARFs in RA patients. These significant elevations of ARFs may be clinically relevant since they correlate with clinical indices. Our results point to systemic microbial exposure as a common stimulus in RA, which could perpetuate the disease.
Materials and Methods
Study Subjects
Subjects were recruited for the Studies of the Etiology of Rheumatoid Arthritis (SERA), a prospective longitudinal study designed to evaluate the contributions of environmental and genetic factors to the development of RA. Recruitment of RA population has been described in detail previously (38). Healthy control subjects included in this study were recruited via local advertisement from the general population and tested negative for RA related autoantibodies at their baseline visit. For both the RA and healthy control populations, the base line visit was selected for this study and the duration of the study entry would be time=0 since this was their first visit. Ethical approval for this study was obtained from University of Colorado's Institutional Review Board (COMIRB#01-675) in compliance with Declaration of Helsinki. Informed consents were obtained from each participant prior to including them in the study. Our study included 50 RA subjects (39 females and 11 males), all fulfilling the revised criteria of 1987 American Rheumatism Association (39) and 50 age- and sex-matched healthy control subjects. All but three of the RA subjects reported being currently or previously on immunosuppressive and/or immunomodulatory drugs at the time of their research study visit. Health assessment, pain index, and disease activity index were collected for RA group at their study visit. The health assessment disability questionnaire index (HAQ; range 0–3) is considered the benchmark for measuring the functional status in adults with RA (40). HAQ Total assesses the hierarchy of patient outcomes by analyzing activity index, disability index, and pain index collected in 100 mm visual analog scale. Demographics and smoking history were obtained by questionnaire. Patient data is provided in Table 1. Individual ARF values per patient are included in Supplementary Table 1.
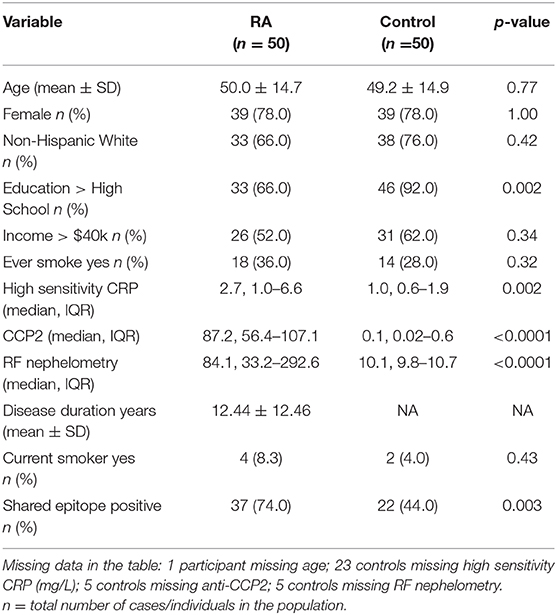
Table 1. Demographic and descriptive characteristics of rheumatoid arthritis and control population.
Sample Collection
Venous blood was drawn in BD Vacutainer® serum separator tubes (Franklin Lakes, NJ, USA) from both RA patients and healthy controls. After clotting, the whole blood collected was centrifuged (for 10 minutes at 3,000 × g and 20°C) and the serum layer was removed. Measurements of rheumatoid factor (RF), high sensitivity C-reactive protein (CRP), and anti-cyclic citrullinated protein antibodies (anti-CCP) in the serum were measured using previously described methodologies (38–41). Multiple aliquots were made from all the serum samples and stored at −80°C until analysis.
Measurement of Analytes in the Serum
sCD14 and LBP were measured using sandwich ELISA kits procured from R&D systems (Minneapolis, USA) and Hycult Biotech (Pennsylvania, USA), respectively. EndoCAb IgG, EndoCAb IgA, and EndoCAb IgM were measured using direct ELISA kits procured from Hycult Biotech. CXCL16 was analyzed using a sandwich ELISA kit procured from Thermo Scientific (Frederick, MD, USA). Lysozyme levels were measured using sandwich ELISA kit procured from MBL (Massachusetts, USA). Total IgG, IgA, and IgM were measured using sandwich ELISA kits procured from Invitrogen (Carlsbad, CA, USA). To block non-specific antibodies that may interfere with the assay, the samples were diluted in appropriate buffers, which contained 50 μg/ml of HeteroBlock (Omega Biologicals, Bozeman, MT, USA) and kept for 30 minutes before adding into the ELISA plate. All the analyses were performed blinded to case/control and clinical status. In order to maintain the test quality and reproducibility, an internal control was included in all the assays and the coefficient of variation (CV) of replicates was set at ≤ 10%.
Statistical Analysis
We transformed all the data into base-10 logarithm values for statistical analysis and correlation studies. Square root transformation was employed for disease indices, which included true zeros. For testing statistical significance, the unpaired t-test was used. For correlation analysis, Pearson product-moment correlation coefficient (Pearson's r) analysis was performed. Gender stratification was also done to detect sex-related changes in the levels of ARFs in RA subjects compared to their respective control subjects. For all statistical tests, P < 0.05 was considered to be statistically significant. All the statistical tests were done with GraphPad Prism 7 (GraphPad Software, Inc., San Deigo, USA). Descriptive statistics of all the analyte levels are given in Supplementary Table 2.
Results
sCD14 Levels Are Increased in RA Patients
Since CD14 acts a co-receptor for LPS, elevated levels of sCD14 are considered to reflect LPS exposure and subsequent monocyte/macrophage activation (42–44). Interestingly, Gram-positive bacterial cell wall components can also bind with CD14 (44–46). Binding of endotoxins to CD14 activates TLRs and promotes the release of proinflammatory cytokines (47, 48). We found a significant increase in the levels of sCD14 in RA patients (P = 0.004) compared to healthy controls (Figure 1A). Gender stratification showed that sCD14 levels in the male RA cohort were significantly elevated compared to control males (P = 0.009). RA females showed a trend toward an increase in sCD14 levels when compared to control females (P = 0.075).
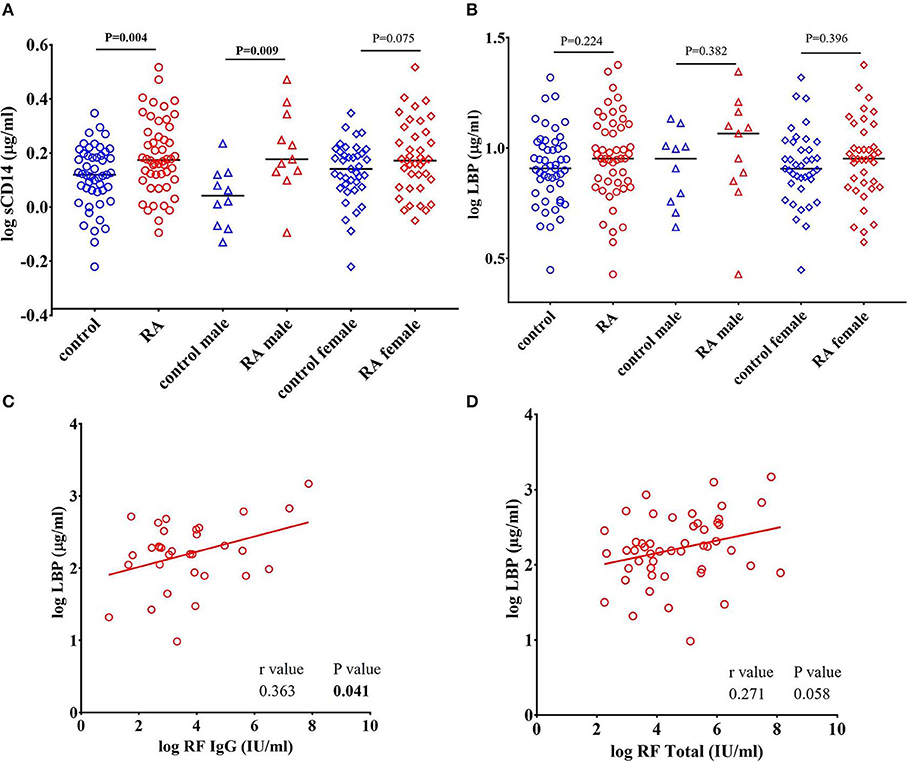
Figure 1. Levels of sCD14 are increased in RA patients. (A) Circulating levels of sCD14 in RA patients showed a significant elevation compared to healthy controls. Gender stratification revealed a significant elevation of sCD14 only in RA males compared to healthy control males while RA females showed a trend toward increase in the levels of sCD14 compared to healthy control females. (B) Concentration of LBP in healthy controls and RA patients. LBP levels were not significantly different in RA patients compared to healthy controls. Bars represent median analyte levels. (C) LBP is correlated with rheumatoid factor IgG (RF IgG). Correlation analysis revealed a significant positive correlation of LBP with RF IgG (C) and a trend toward significant positive correlation with total rheumatoid factor (RF Total) (D).
LBP Levels Positively Correlate With Rheumatoid Factor (RF) in RA Subjects
LBP is an acute phase protein synthesized by hepatocytes involved in the transfer of LPS to CD14, which partners with TLR4 expressed on innate immune cells (49, 50). We determined that LBP levels were increased in our RA cohort, however the results did not achieve statistical difference (P = 0.224) (Figure 1B). A positive correlation between LBP and CRP (r = 0.335, P = 0.017) in RA patients is observed in our study (Supplementary Figure 1). However, we did not find a correlation between CRP with any other ARFs that we measured.
RF was the first described autoantibody in RA and is directed against the Fc region of IgG. RF is also a valuable biomarker in terms of disease severity, diagnosis and prognosis in RA (5, 51). We found that levels of RF IgG were positively correlated with LBP (r = 0.363, P = 0.041) (Figure 1C). Furthermore, total antibody levels for all RF isotypes were also positively, though weakly, correlated with LBP (r = 0.271, P = 0.058) (Figure 1D). We did not find any correlation between RF and other ARFs that we measured (Data not shown).
Levels of Lysozyme Are Increased in RA Patients
Lysozyme is an important ARF that is secreted by monocytes, macrophages, neutrophils, glandular cells, and dendritic cells. Lysozyme kills bacteria by hydrolyzing the peptidoglycan component of the bacterial cell wall. Lysozyme also possess bactericidal activity against Gram-negative bacteria (52, 53). We observed a significant increase in the levels of lysozyme in RA patients (P = 0.033) compared to healthy controls (Figure 2A). However, gender stratification of RA subjects showed that neither RA males (P = 0.118) nor RA females (P = 0.141) (Figure 2A) were significantly different than their respective healthy controls.
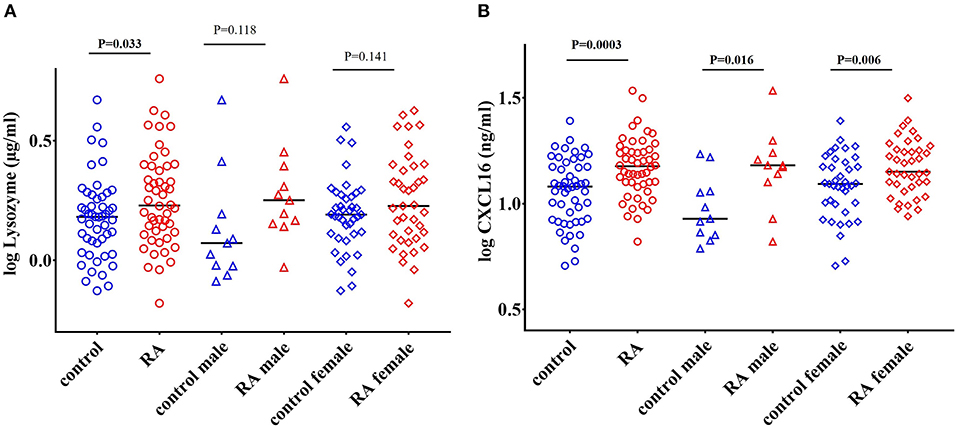
Figure 2. Elevated levels of lysozyme and CXCL16 in RA subjects. (A) Circulating levels of lysozyme are significantly elevated in RA patients compared to healthy controls. Following gender stratification, the lysozyme levels were similar in both the male and female RA cohorts compared to respective healthy controls. (B) RA patients have elevated levels of CXCL16 in the circulation compared to healthy controls. A significant increase in the levels of CXCL16 was also observed in both the male and female RA patients compared to respective healthy controls. Bars represent median analyte levels.
Levels of CXCL16 Are Increased in RA Patients
CXCL16 is an important chemokine that acts as a mediator of the innate immune response (54). CXCL16 mediates adhesion and phagocytosis of both Gram-negative and Gram-positive bacteria and acts as a strong chemoattractant for CXCR6+ T cells (55–57). CXCL16 levels are also affected by alterations in the gut microbiome (58). We observed a significant elevation in the levels of CXCL16 in RA patients (P = 0.0003) compared to healthy controls (Figure 2B). Both RA males (P = 0.016) and RA females (P = 0.006) showed a significant increase in the levels of CXCL16 compared to their respective controls (Figure 2B).
sCD14 Levels Positively Correlate With LBP, Lysozyme, and CXCL16 in RA Subjects and Healthy Controls
Pearson's r analysis showed a significant positive correlation of sCD14 with LBP (r = 0.669, P < 0.0001 in RA subjects and r = 0.521, P = 0.0001 in healthy controls), lysozyme (r = 0.708, P < 0.0001 in RA subjects and r = 0.480, P = 0.0005 in healthy controls), and CXCL16 (r = 0.618, P < 0.0001 in RA subjects and r = 0.759, P < 0.0001 in healthy controls) in both RA patients and in healthy controls (Figures 3A–C). Moreover, a significant positive correlation between CXCL16 and lysozyme (r = 0.501, P = 0.0002 in RA subjects and r = 0.507, P = 0.0002 in healthy controls) was also observed in both RA subjects and in healthy controls (Figure 3D).
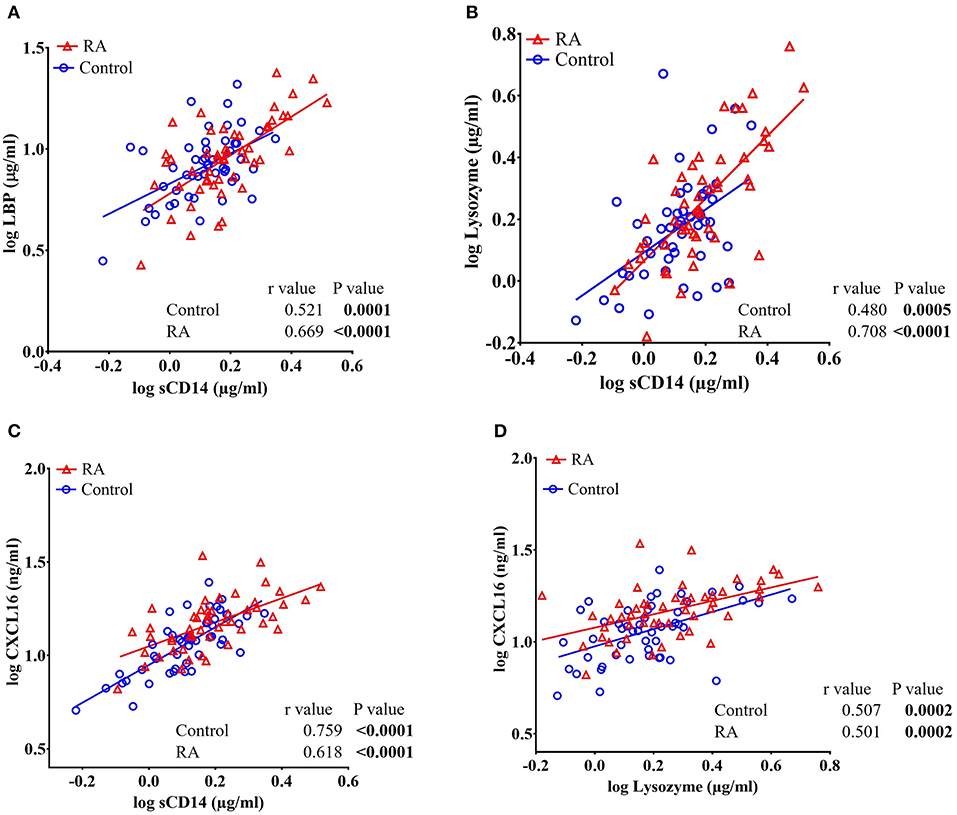
Figure 3. Circulating levels of sCD14 are positively and significantly correlated with LBP, lysozyme, and CXCL16 in RA patients and healthy controls. (A–C) Analysis showing a significant positive correlation of sCD14 with LBP, lysozyme, and CXCL16 in both RA patients and healthy controls. (D) Analysis showing a significant positive correlation between CXCL16 and lysozyme in RA patients and healthy controls.
Total IgA and IgM levels, Including EndoCAb-Specific IgA and IgM, Are Increased in RA Patients
EndoCAbs are antibodies directed against the endotoxin core of LPS. They bind and neutralize LPS activity (59–61). We did not find significant changes in the levels of EndoCAb IgG in RA subjects compared to healthy controls (Figure 4A). Interestingly, levels of EndoCAb IgA (P = 0.001) and EndoCAb IgM (P = 0.011) were elevated in our RA cohort compared to healthy controls (Figures 4B,C). RA females showed a significant increase in the levels of EndoCAb IgA compared to control females (P = 0.007) whereas males showed only a trend toward an increase (P = 0.06) (Figure 4B). EndoCAb IgM levels were found to be significantly elevated in RA females (P = 0.032) compared to control females. RA males did not show any significant difference in EndoCAb IgM levels (P = 0.156) compared to control males (Figure 4C).
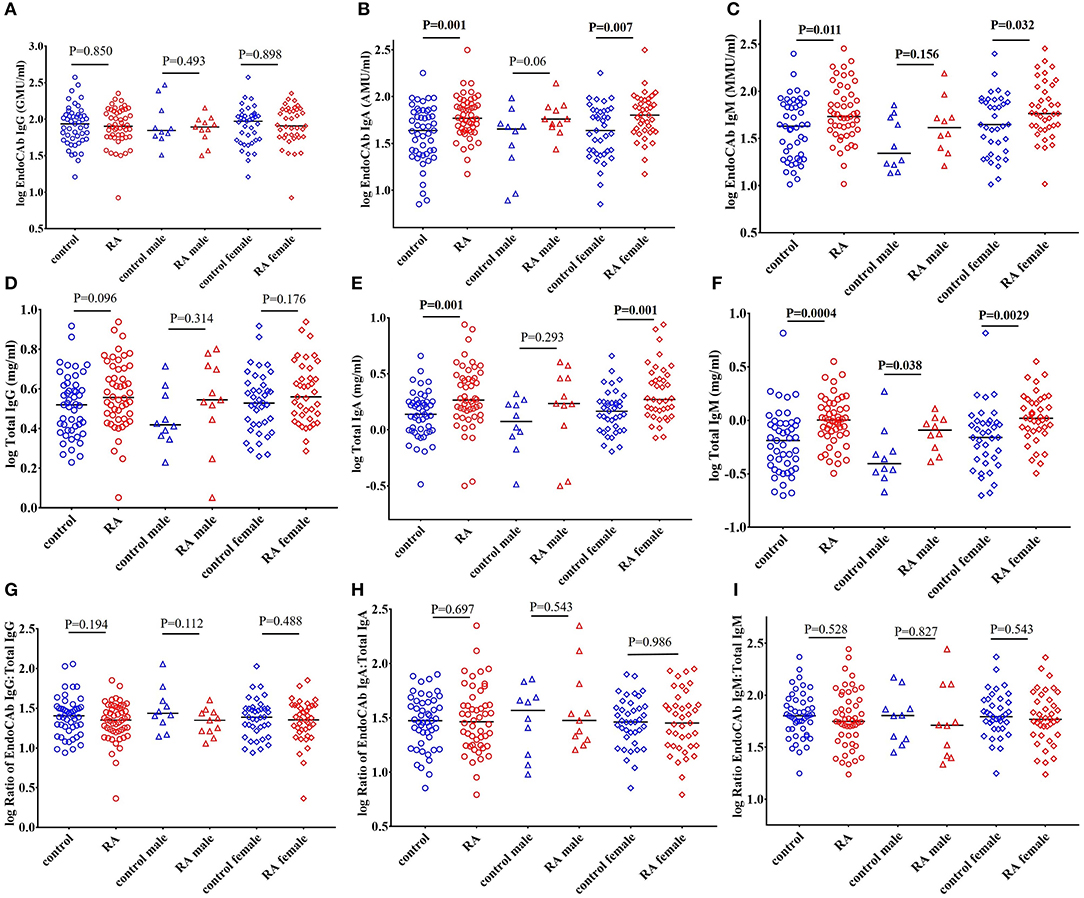
Figure 4. Elevated levels of total IgA and total IgM, including EndoCAb-specific IgA and IgM, in RA patients. (A) Circulating EndoCAb IgG levels were not significantly different in RA patients compared to healthy controls with or without gender stratification. (B) Circulating levels of EndoCAb IgA were significantly elevated in RA patients compared to healthy controls. RA females showed a significant elevation of EndoCAb IgA whereas RA males showed a trend toward significance compared to their respective healthy controls. (C) Levels of circulating EndoCAb IgM were significantly elevated in RA patients compared to healthy controls. Gender stratification revealed a significant increase of EndoCAb IgM in RA females whereas RA males did not show any significant change compared to their respective healthy controls. (D) Circulating levels of total IgG were similar in RA and controls, with or without gender stratification. (E) Total IgA levels were significantly increased in RA patients compared to healthy controls. RA females showed a significant elevation of total IgA whereas in RA males the changes did not achieve any statistical significance (F) Levels of circulating total IgM were significantly elevated in RA patients compared to healthy controls. Both the RA males and RA females showed a significant elevation in the levels of total IgM compared to their respective healthy controls. (G–I) Ratio of EndoCAbs:total Igs were not significantly different in RA patients than the healthy controls. Gender stratification also showed no significant changes in the ratios of EndoCAbs:total Igs compared to their respective healthy controls. For all figures, bars represent median analyte levels.
Analysis of total immunoglobulins (Igs) in RA cohort showed that IgA (P = 0.001) and IgM (P = 0.0004) were significantly elevated in RA patients compared to healthy controls, confirming earlier reports (62–64). The levels of IgG were also elevated in the RA cohort but did not achieve statistical significance (P = 0.096) (Figure 4D). RA females showed a significant elevation of IgA (P = 0.001) whereas RA males showed a trend toward significant increase (P = 0.293). IgM levels of both RA males (P = 0.038) and RA females (P = 0.003) were elevated compared to respective control subjects (Figures 4E,F).
We then assessed whether the proportion of EndoCAbs was elevated by analyzing the ratios of EndoCAbs:total Igs in all the groups. We did not find any significant difference between the ratios of EndoCAb IgG:total IgG (P = 0.194), EndoCAb IgA:total IgA (P = 0.697), and EndoCAb IgM:total IgM (P = 0.528) in RA subjects compared to healthy controls (Figures 4G–I).
Correlations of EndoCAbs With sCD14, and Lysozyme in RA Subjects
In order to determine whether the levels of EndoCAbs were associated with other factors, we analyzed the correlations of EndoCAbs with other ARFs and RF. We observed a negative correlation between the ratio of EndoCAb IgA:total IgA with sCD14 in RA subjects (r = −0.268, P = 0.059). Healthy controls did not show any significant correlation between these factors (r = 0.176, P = 0.226) (Figure 5A). We found a significant positive correlation between lysozyme and EndoCAb IgG (r = 0.301, P = 0.033 in RA subjects; r = 0.329, P = 0.021 in healthy controls) and EndoCAb IgA (r = 0.291, P = 0.040 in RA subjects; r = 0.420. P = 0.002 in healthy controls) in both RA subjects and healthy controls (Figures 5B,C). Moreover, lysozyme levels were also found to be positively correlated with total IgG (r = 0.497, P = 0.0002 in RA subjects; r = 0.389, P = 0.0057 in healthy controls) and total IgA levels (r = 0.392, P = 0.0049 in RA subjects; r = 0.319, P = 0.025 in healthy controls) in both RA patients and healthy controls (Figures 5D,E).
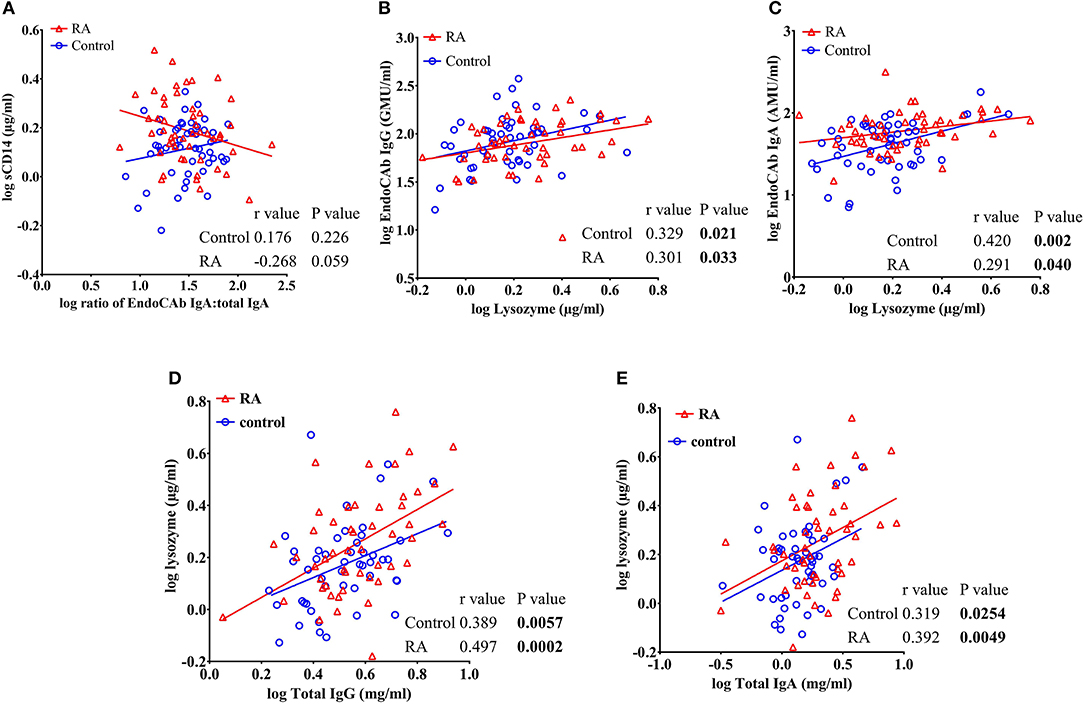
Figure 5. sCD14 levels correlate with the ratio of EndoCAb IgA:total IgA in RA patients. (A) Analysis showing a trend toward negative correlation between sCD14 and the ratio of EndoCAb IgA:total IgA in RA patients whereas healthy controls did not show any significant correlation between these values. (B–E) Analysis showing a significant positive correlation of circulating levels of lysozyme with EndoCAb IgG, EndoCAb IgA, total IgG, and total IgA in both RA patients and healthy controls.
Lysozyme Levels Positively Correlate With Total Health Assessment Disability Questionnaire Index (HAQ Total) Values in RA Subjects
We found a significant positive correlation between lysozyme levels and the HAQ Total index (r = 0.308, P = 0.032) (Figure 6A). A trend toward a positive correlation was observed in the pain index vs. CXCL16 (r = 0.280, P = 0.051) and the activity index vs. CXCL16 (r = 0.283, P = 0.054) in RA subjects (Figures 6B,C). Furthermore, a significant positive correlation between pain index and total IgA was also observed (r = 0.336, P = 0.019) (Figure 6D). Thus, the levels of these analytes parallel clinical measurements of disease severity.
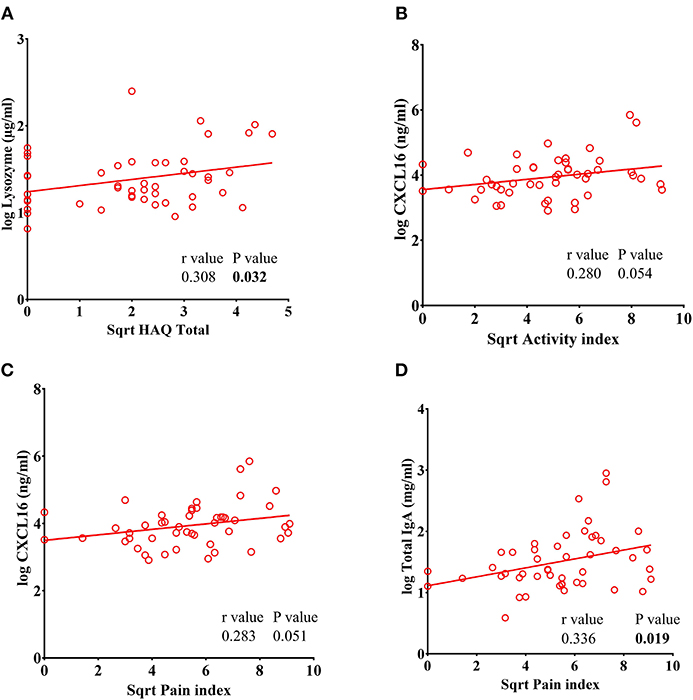
Figure 6. Lysozyme levels correlate with Total Health Assessment Quality questionnaire disability index (HAQ Total) in RA patients. (A) Analysis showing a significant positive correlation of lysozyme with HAQ total in RA patients. (B–C) CXCL16 levels showed a trend toward significant positive correlation of CXCL16 with pain index and disease activity index in RA patients. (D) Analysis showing a significant positive correlation between pain index and total IgA levels in RA patients.
No other ARFs showed a significant association with disease activity measures. In addition, we did not find any significant correlation between the levels of anti-CCP antibodies with ARFs in RA patients (data not shown).
Discussion
Systemic exposure to microbial products has been hypothesized to trigger and/or potentiate several autoimmune diseases including RA (65–67). In response to microbial products, multiple ARFs are released into circulation as a protective mechanism to clear microbes and reduce inflammation (68, 69). Interestingly, increased circulatory levels of several ARFs in response to bacterial infection such as in sepsis also indicates that these ARFs may be specific for infection (70–73). In this study, we found increased levels of multiple ARFs in RA patients.
We observed a significant elevation of sCD14 levels in RA patients compared to healthy controls confirming previous reports (74–76). sCD14 acts as a co-receptor for endotoxin and facilitates the activation of those cells which are devoid of membrane bound CD14 via TLR4 transmembrane signaling (77–80). Elevated levels of sCD14 could be caused by bacterial exposure, alterations in the microbiome, compromised gut integrity and increased levels of cytokines. This would induce monocyte/macrophage activation and elevate the circulating concentrations of sCD14 (43, 81–86). Release of sCD14 by synovial macrophages was also suggested to contribute to elevated levels in RA patients (74). Overproduction of sCD14 by macrophages may act as a death associated molecular pattern (DAMP) and induce the production of proinflammatory cytokines (87). Thus, the elevated levels of sCD14 in circulation contributes to the maintenance of tissue inflammation by increasing the responsiveness against endotoxins (82, 87). Alternatively, elevated levels of sCD14 were reported to reduce the interaction between LPS and monocytes thereby reducing the adverse effects of monocyte/macrophage activation (42, 88). Due to the ambiguities in these experimental outcomes, further investigations are required to define the pathophysiological role of elevated levels of sCD14 in the circulation.
Similar to sCD14, LBP is a critical circulatory molecule involved in endotoxin clearance (89). We did not observe any significant difference in the levels of LBP in RA subjects compared to healthy controls. Our findings are in contrast to a recent report of elevated levels of LBP in RA patients (90). From their results, it is proposed that LBP is a specific and sensitive biomarker for RA (90). The discrepancies with our results could reflect differences in assay methodology or patient populations. Regarding methodology, while the assays are identical, the inclusion of HeteroBlock in our study could make a difference. Autoantibodies produced in RA such as RF cause interference in some immunoassays (91). RF can generate false signals by bridging capture and detection antibodies in sandwich ELISAs (92), an effect that can be mitigated through HeteroBlock (91, 93). A lack of corrective measures to block the RF interference in the study reported by Wen et al. (90) could explain their results. Differences in the two patient populations could also affect the observed differences in LBP levels. It should be noted that the LBP values for RA subjects reported by Wen et al. (90) were comparable with those reported for sepsis and other severe illnesses (73, 94, 95). Furthermore, their reported CRP values were substantially higher than those for our cohort.
Levels of lysozyme were significantly elevated in RA patients, confirming previous observations (96). Lysozyme is an important bacteriolytic enzyme produced by monocytes, macrophages, neutrophils, dendritic cells and glandular cells (52, 97). The antimicrobial potential of lysozyme is derived from its ability to hydrolyze the glycosidic bond of peptidoglycan, which is found in the cell walls of both Gram-positive and Gram-negative bacteria (94). In circulation, lysozyme facilitates the degradation of bacterial peptidoglycan into peptidoglycan monomers. This leads to the activation of myeloid cells via various pattern recognition receptors (52). Bacterial exposure can elevate the levels of lysozyme by increasing the activation of monocytes/macrophages and neutrophils (97–100). Moreover, proinflammatory cytokines released by activated macrophages can elevate the production and/or release of lysozyme (96, 97, 101).
We found that CXCL16 was elevated in our RA subjects compared to healthy controls, confirming earlier results (9, 102). CXCL16 is recognized as an antimicrobial protein involved in the adhesion and phagocytosis of bacteria (54, 55, 103, 104). Moreover, CXCL16 serves as a chemoattractant that mediates the recruitment of CXCR6-expressing immune cells and mediates inflammation (105–107). The binding of LPS with CD14 triggers the activation of NF-κB, inducing the release of CXCL16 (108, 109). Elevated circulatory levels of CXCL16 in RA patients may reflect systemic inflammation. Interestingly, we also observed a positive correlation of CXCL16 with disease severity (pain index and activity index). Li and colleagues (9) also reported a significant positive correlation of CXCL16 and RA disease activity.
B cells play an important role in the pathogenesis of RA by secreting autoantibodies, presenting antigens and producing cytokines. We observed increased levels of total IgA and total IgM in RA patients compared to healthy controls. Increased levels of total IgA and IgM in RA patients reflects activation of the immune system (61–63). EndoCAbs are endotoxin core antibodies, which can bind and neutralize circulating LPS (58). Similar to total Igs, EndoCAb IgA and EndoCAb IgM levels were significantly increased in RA patients compared to healthy controls. Elevated levels of IgM and IgA specific to some bacterial species were also observed in RA patients (28, 110). However, we found that the ratios of EndoCAb Igs:total Igs were not different from controls. This indicates that elevated levels of IgA and IgM may be due to polyclonal B cell activation. Systemic exposure of microbial products leads to polyclonal B cell hyperactivation and elevated levels of Igs (111–113). From our study, it appears that monocyte/macrophage activation is the likely cause of B cell activation and subsequent increase of total Igs and EndoCAbs in RA patients. Apart from microbial products, sCD14 was found to activate B cells (114). In addition, Ig secretion could be stimulated by cytokines released by activated monocytes/macrophages and dendritic cells (115–117).
Similar to other autoimmune conditions, RA mainly affects females (6). We observed gender differences in the levels of some ARFs. Levels of sCD14 in RA males were significantly elevated compared to control males and not females. Conversely, EndoCAb IgA, EndoCAb IgM, and total IgA levels were higher in RA females compared to control females, but were unaffected in males. This could reflect gender-specific changes in the microbiome, which were found to modulate the immune response distinctly in males and females (118–120). These gender-specific changes in gut microbiota could drive gender-biased autoimmunity (121, 122).
Multiple studies suggest the potential role of microbes as triggering and/or accelerating factors in autoimmunity (66, 123–126). Our previous studies with systemic lupus erythematosus (SLE) samples showed elevated levels of ARFs similar to what we observed in this RA study. Similar to SLE, RA subjects also showed elevated levels of total IgA, sCD14, lysozyme, and CXCL16 compared to healthy controls (127). In RA, elevated levels of IgA and total IgM along with EndoCAb IgA and EndoCAb IgM in RA indicate a global immune response. Differences in the microbiome could be a determining factor in the changes in the levels of ARFs (128).
Elevated levels of ARFs in our study support the role of myeloid cell activation in disease pathogenesis, possibly via systemic microbial exposure in RA cohort (Supplementary Figure 2). The gastrointestinal and lung microbiomes could contribute to the modulation of ARF levels. Longitudinal studies in human RA subjects are required to understand the significance of these ARFs as biomarkers for future RA development. A deeper understanding of the connection between antimicrobial responses and autoimmunity in RA could help to establish therapeutic strategies for the effective disease management in highly susceptible populations.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
The studies involving human participants were reviewed and approved by University of Colorado's Institutional Review Board (COMIRB#01-675). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
PA: experimental design and execution, data analysis, and manuscript preparation. RH and NS: experimental design, data analysis and reviewed the manuscript. JS, EB, MF, KD, MD, TM, and VH: substantial contribution by acquiring patient samples and clinical data and manuscript revision and review. All the authors approved the final version of the manuscript.
Funding
This work was supported by grants to NS from NIAD (UO1AI130841, UO1AI102012), and the grants to VH from the NIH Autoimmunity Prevention Center (U01 AI101981, R01 AR051394). Funding support from the Leona M. and Harry B. Helmsley Charitable Trust (2018PG-T1D048), the Juvenile Diabetes Research Foundation (2-SRA-2016-288-S-B, 1-PNF-2014-165-A-V), and the J. W. Kieckhefer Foundation to NS is also acknowledged.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the individuals who participated in the study, coordinators of the recruitment sites and those involved in sample collection and processing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.00427/full#supplementary-material
References
1. Liao KP, Alfredsson L, Karlson EW. Environmental influences on risk for rheumatoid arthritis. Curr Opin Rheumatol. (2009) 21:279–83. doi: 10.1097/BOR.0b013e32832a2e16
2. Raychaudhuri S, Sandor C, Stahl EA, Freudenberg J, Lee HS, Jia X, et al. Five amino acids in three HLA proteins explain most of the association between MHC and seropositive rheumatoid arthritis. Nat Genet. (2012) 44:291–6. doi: 10.1038/ng.1076
3. Deane KD, Demoruelle MK, Kelmenson LB, Kuhn KA, Norris JM, Holers VM. Genetic and environmental risk factors for rheumatoid arthritis. Best Pract Res Clin Rheumatol. (2017) 31:3–18. doi: 10.1016/j.berh.2017.08.003
4. Guo Q, Wang Y, Xu D, Nossent J, Pavlos NJ, Xu J. Rheumatoid arthritis: pathological mechanisms and modern pharmacologic therapies. Bone Res. (2018) 6:15. doi: 10.1038/s41413-018-0016-9
5. Song YW, Kang EH. Autoantibodies in rheumatoid arthritis: rheumatoid factors and anticitrullinated protein antibodies. QJM. (2010) 103:139–46. doi: 10.1093/qjmed/hcp165
6. van Vollenhoven RF. Sex differences in rheumatoid arthritis: more than meets the eye. BMC Med. (2009) 7:12. doi: 10.1186/1741-7015-7-12
7. Akhlaghi S, Sahebari M, Mahmoodi M, Yaseri M, Mansournia MA, Zeraati H. Casual effect of methotrexate+etanercept/infliximab on survival of patients with rheumatoid arthritis. Pragmat Obs Res. (2019) 10:23–8. doi: 10.2147/POR.S194408
8. Choy E. Understanding the dynamics: pathways involved in the pathogenesis of rheumatoid arthritis. Rheumatology. (2012) 51:v3–11. doi: 10.1093/rheumatology/kes113
9. Li CH, Xu LL, Zhao JX, Sun L, Yao ZQ, Deng XL, et al. CXCL16 upregulates RANKL expression in rheumatoid arthritis synovial fibroblasts through the JAK2/STAT3 and p38/MAPK signaling pathway. Inflamm Res. (2016) 65:193–202. doi: 10.1007/s00011-015-0905-y
10. Bemis EA, Norris JM, Seifert J, Frazer-Abel A, Okamoto Y, Feser ML, et al. Complement and its environmental determinants in the progression of human rheumatoid arthritis. Mol Immunol. (2019) 112:256–65. doi: 10.1016/j.molimm.2019.05.012
11. Das S, Padhan P. An overview of the extraarticular involvement in rheumatoid arthritis and its management. J Pharmacol Pharmacother. (2017) 8:81–6. doi: 10.4103/jpp.JPP_194_16
12. Marcucci E, Bartoloni E, Alunno A, Leone MC, Cafaro G, Luccioli F, et al. Extra-articular rheumatoid arthritis. Reumatismo. (2018) 70:212–24. doi: 10.4081/reumatismo.2018.1106
13. Liu X, Zou Q, Zeng B, Fang Y, Wei H. (2013) Analysis of fecal Lactobacillus community structure in patients with early rheumatoid arthritis. Curr Microbiol. (2013) 67:170–6. doi: 10.1007/s00284-013-0338-1
14. Scher JU, Sczesnak A, Longman RS, Segata N, Ubeda C, Bielski C, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife. (2013) 2:e01202. doi: 10.7554/eLife.01202
15. Sandhya P, Danda D, Sharma D, Scaria V. Does the buck stop with the bugs?: an overview of microbial dysbiosis in rheumatoid arthritis. Int J Rheum Dis. (2016) 19:8–20. doi: 10.1111/1756-185X.12728
16. Mikuls TR, Payne JB, Deane KD, Thiele GM. Autoimmunity of the lung and oral mucosa in a multisystem inflammatory disease: The spark that lights the fire in rheumatoid arthritis? J Allergy Clin Immunol. (2016) 137:28–34. doi: 10.1016/j.jaci.2015.10.024
17. Zhang X, Zhang D, Jia H, Feng Q, Wang D, Liang D. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med. (2015) 21:895–905. doi: 10.1038/nm.3914
18. Scher JU, Joshua V, Artacho A, Abdollahi-Roodsaz S, Ockinger J, Kullberg S, et al. The lung microbiota in early rheumatoid arthritis and autoimmunity. Microbiome. (2016) 4:60. doi: 10.1186/s40168-016-0206-x
19. Ogrendik M, Kokino S, Ozdemir F, Bird PS, Hamlet S. Serum antibodies to oral anaerobic bacteria in patients with rheumatoid arthritis. Med Gen Med. (2005) 7:2. Available online at: https://www.medscape.com/viewarticle/505458_1
20. Scher JU, Ubeda C, Equinda M, Khanin R, Buischi Y, Viale A, et al. Periodontal disease and the oral microbiota in new-onset rheumatoid arthritis. Arthritis Rheum. (2012) 64:3083–94. doi: 10.1002/art.34539
21. Wu HJ, Ivanov II, Darce J, Hattori K, Shima T, Umesaki Y, et al. Gut residing segmented filamentous bacteria drive autoimmune arthritis via T helper 17 cells. Immunity. (2010) 32:815–27. doi: 10.1016/j.immuni.2010.06.001
22. Teng F, Klinger CN, Felix KM, Bradley CP, Wu E, et al. Gut microbiota drive autoimmune arthritis by promoting differentiation and migration of Peyer's patch T follicular helper cells. Immunity. (2016) 44:875–88. doi: 10.1016/j.immuni.2016.03.013
23. Picchianti-Diamanti A, Panebianco C, Salemi S, Sorgi ML, Di Rosa R, Tropea A, et al. Analysis of gut microbiota in rheumatoid arthritis patients: disease-related dysbiosis and modifications induced by etanercept. Int J Mol Sci. (2018) 19:E2938. doi: 10.3390/ijms19102938
24. Rogier R, Evans-Marin H, Manasson J, van der Kraan PM, Walgreen B, Helsen MM, et al. Alteration of the intestinal microbiome characterizes preclinical inflammatory arthritis in mice and its modulation attenuates established arthritis. Sci Rep. (2017) 7:15613. doi: 10.1038/s41598-017-15802-x
25. Zhang P, Minardi LM, Kuenstner JT, Zekan SM, Kruzelock R. Anti-microbial antibodies, host immunity, and autoimmune disease. Front Med. (2018) 5:153. doi: 10.3389/fmed.2018.00153
26. Moen K, Brun JG, Madland TM, Tynning T, Jonsson R. Immunoglobulin G and A antibody responses to Bacteroides forsythus and Prevotella intermedia in sera and synovial fluids of arthritis patients. Clin Diagn Lab Immunol. (2003) 10:1043–50. doi: 10.1128/CDLI.10.6.1043-1050.2003
27. Mikuls TR, Payne JB, Reinhardt RA, Thiele GM, Maziarz E, Cannella AC, et al. Antibody responses to Porphyromonas gingivalis (P. gingivalis) in subjects with rheumatoid arthritis and periodontitis. Int Immunopharmacol. (2009) 9:38–42. doi: 10.1016/j.intimp.2008.09.008
28. Newkirk MM, Goldbach-Mansky R, Senior BW, Klippel J, Schumacher HR Jr, El-Gabalawy HS. Elevated levels of IgM and IgA antibodies to Proteus mirabilis and IgM antibodies to Escherichia coli are associated with early rheumatoid factor (RF)-positive rheumatoid arthritis. Rheumatology. (2005) 44:1433–41. doi: 10.1093/rheumatology/kei036
29. Aoki S, Yoshikawa K, Yokoyama T, Monogaki T, Iwasaki S, Mitsui T, Niwa S. Role of enteric bacteria in the pathogenesis of rheumatoid arthritis: evidence for antibodies to enterobacterial common antigens in rheumatoid sera and synovial fluids. Ann Rheum Dis. (1996) 55:363–9. doi: 10.1136/ard.55.6.363
30. Moen K, Brun JG, Valen M, Skartveit L, Eribe EK, Olsen I, et al. Synovial inflammation in active rheumatoid arthritis and psoriatic arthritis facilitates trapping of a variety of oral bacterial DNAs. Clin Exp Rheumatol. (2006) 24:656–63. Available online at: https://www.clinexprheumatol.org/abstract.asp?a=2980
31. Zhao Y, Chen B, Li S, Yang L, Zhu D, Wang Y, et al. Detection and characterization of bacterial nucleic acids in culture-negative synovial tissue and fluid samples from rheumatoid arthritis or osteoarthritis patients. Sci Rep. (2018) 8:14305. doi: 10.1038/s41598-018-32675-w
32. Konig MF, Abusleme L, Reinholdt J, Palmer RJ, Teles RP, Sampson K, et al. Aggregatibacter actinomycetemcomitans-induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci Transl Med. (2016) 8:369ra176. doi: 10.1126/scitranslmed.aaj1921
33. Mukherjee A, Jantsch V, Khan R, Hartung W, Fischer R, Jantsch J, et al. Rheumatoid arthritis-associated autoimmunity due to Aggregatibacter actinomycetemcomitans and its resolution with antibiotic therapy. Front Immunol. (2018) 9:2352. doi: 10.3389/fimmu.2018.02352
34. Stapels DA, Geist BV, Rooijakkers SH. Neutrophil serine proteases in antibacterial defense. Curr Opin Microbiol. (2015) 23:42–8. doi: 10.1016/j.mib.2014.11.002
35. Lee EY, Lee MW, Wong GCL. Modulation of toll-like receptor signaling by antimicrobial peptides. Semin Cell Dev Biol. (2019) 88:173–84. doi: 10.1016/j.semcdb.2018.02.002
36. Zhang L, Gallo R. Antimicrobial peptides. Current Biol. (2016) 26:R14–9. doi: 10.1016/j.cub.2015.11.017
37. Lazar V, Ditu LM, Pircalabioru GG, Gheorghe I, Curutiu C, Holban AM, et al. Aspects of gut microbiota and immune system interactions in infectious diseases, immunopathology, and cancer. Front Immunol. (2018) 9:1830. doi: 10.3389/fimmu.2018.01830
38. Kolfenbach JR, Deane KD, Derber LA, O'Donnell C, Weisman MH, Buckner JH, et al. A prospective approach to investigating the natural history of preclinical rheumatoid arthritis (RA) using first-degree relatives of probands with RA. Arthritis Rheum. (2009) 61:1735–42. doi: 10.1002/art.24833
39. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. (1988) 31:315–24. doi: 10.1002/art.1780310302
40. Fries JF, Spitz P, Kraines RG, Holman HR. Measurement of patient outcome in arthritis. Arthritis Rheum. (1980) 23:137–45. doi: 10.1002/art.1780230202
41. Berens HM, Polinski KJ, Mikuls TR, Khatter S, August J, Visser A, et al. Anticyclic citrullinated peptide antibodies 3.1 and Anti-CCP-IgA are associated with increasing age in individuals without rheumatoid arthritis. J Rheumatol. (2019) 46, 1556–1559. doi: 10.3899/jrheum.180897
42. Landmann R, Knopf HP, Link S, Sansano S, Schumann R, Zimmerli W. Human monocyte CD14 is upregulated by lipopolysaccharide. Infect Immun. (1996) 64:1762–9. doi: 10.1128/IAI.64.5.1762-1769.1996
43. Kitchens RL, Thompson PA, Viriyakosol S, O'Keefe GE, Munford RS. Plasma CD14 decreases monocyte responses to LPS by transferring cell-bound LPS to plasma lipoproteins. J Clin Invest. (2001) 108:485–93. doi: 10.1172/JCI200113139
44. Shive CL, Jiang W, Anthony DD, Lederman MM. Soluble CD14 is a nonspecific marker of monocyte activation. AIDS. (2015) 29:1263–5. doi: 10.1097/QAD.0000000000000735
45. Kol A, Lichtman AH, Finberg RW, Libby P, Kurt-Jones EA. Cutting edge: heat shock protein (HSP) 60 activates the innate immune response: CD14 is an essential receptor for HSP60 activation of mononuclear cells. J Immunol. (2000) 164:13–7. doi: 10.4049/jimmunol.164.1.13
46. Moreillon P, Majcherczyk PA. Proinflammatory activity of cell-wall constituents from Gram-positive bacteria. Scand J Infect Dis. (2003) 35:632–41. doi: 10.1080/00365540310016259
47. Dentener MA, Bazil V, Vonasmuth EJU, Ceska M, Buurman WA. Involvement of CD14 in lipopolysaccharide-induced tumor-necrosis-factor-alpha, IL-6 and IL-8 release by human monocytes and alveolar macrophages. J Immunol. (1993) 150:2885–91.
48. Pugin J, Schurer-Maly CC, Leturcq D, Moriarty A, Ulevitch RJ, Tobias PS. Lipopolysaccharide activation of human endothelial and epithelial cells is mediated by lipopolysaccharide-binding protein and soluble CD14. Proc Natl Acad Sci USA. (1993) 90:2744. doi: 10.1073/pnas.90.7.2744
49. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC. CD14, a receptor for complexes of lipopolysaccharide (LPS) and LPS binding protein. Science. (1990) 249:1431–3. doi: 10.1126/science.1698311
50. Park BS, Lee J-O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp Mol Med. (2013) 45:e66. doi: 10.1038/emm.2013.97
51. Ingegnoli F, Castelli R, Gualtierotti R. Rheumatoid factors: clinical applications. Dis Mark. (2013) 35:727–734. doi: 10.1155/2013/726598
52. Nash JA, Ballard TN, Weaver TE, Akinbi HT. The peptidoglycan-degrading property of lysozyme is not required for bactericidal activity in vivo. J Immunol. (2006) 177:519–26. doi: 10.4049/jimmunol.177.1.519
53. Ragland SA, Criss AK. From bacterial killing to immune modulation: recent insights into the functions of lysozyme. PLoS Pathog. (2017) 13:e1006512. doi: 10.1371/journal.ppat.1006512
54. Tohyama M, Sayama K, Komatsuzawa H, Hanakawa Y, Shirakata Y, Dai X, et al. CXCL16 is a novel mediator of the innate immunity of epidermal keratinocytes. Int Immunol. (2007) 19:1095–102. doi: 10.1093/intimm/dxm083
55. Shimaoka T, Nakayama T, Kume N, Takahashi S, Yamaguchi J, Minami M, et al. Cutting edge: SR-PSOX/CXC chemokine ligand 16 mediates bacterial phagocytosis by APCs through its chemokine domain. J Immunol. (2003) 171:1647–51. doi: 10.4049/jimmunol.171.4.1647
56. Uza N, Nakase H, Yamamoto S, Yoshino T, Takeda Y, Ueno S, et al. SR-PSOX/CXCL16 plays a critical role in the progression of colonic inflammation. Gut. (2011) 60:1494–505. doi: 10.1136/gut.2010.221879
57. Abed M, Towhid ST, Pakladok T, Alesutan I, Gotz F, Gulbins E, et al. Effect of bacterial peptidoglycan on erythrocyte death and adhesion to endothelial cells. Int J Med Microbiol. (2013) 303:182–9. doi: 10.1016/j.ijmm.2013.01.004
58. Ma C, Han M, Heinrich B, Fu Q, Zhang Q, Sandhu M, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. (2018) 360:eaan5931. doi: 10.1126/science.aan5931
59. Barclay GR. Endogenous endotoxin-core antibody (EndoCAb) as a marker of endotoxin exposure and a prognostic indicator: a review. Prog Clin Biol Res. (1995) 392:263–72.
60. Bennett-Guerrero E, Barclay GR, Weng PL, Bodian CA, Feierman DE, Vela-Cantos F, et al. Endotoxin-neutralizing capacity of serum from cardiac surgical patients. J Cardiothorac Vasc Anesth. (2001) 15:451–4. doi: 10.1053/jcan.2001.24980
61. Aravindhan V, Mohan V, Arunkumar N, Sandhya S, Babu S. Chronic endotoxemia in subjects with type-1 diabetes is seen much before the onset of microvascular complications. PLoS ONE. (2015) 10:e0137618. doi: 10.1371/journal.pone.0137618
62. Barden J, Mulinax F, Waller M. Immunoglobulin levels in rheumatoid arthritis: comparison with rheumatoid factor titers, clinical stage and disease duration. Arthritis Rheum. (1967) 10:228–34. doi: 10.1002/art.1780100308
63. Veys EM, Claessens HE. Serum levels of IgG, IgM, and IgA in rheumatoid arthritis. Ann Rheum Dis. (1968) 27:431−440. doi: 10.1136/ard.27.5.431
64. Jorgensen CH, Anaya JM, Cognot C, Sany J. Rheumatoid arthritis associated with high levels of immunoglobulin A: clinical and biological characteristics. Clin Exp Rheumatol. (1992) 10:571–5.
66. Mu Q, Kirby J, Reilly CM, Luo XM. Leaky Gut As a Danger Signal for Autoimmune Diseases. Front Immunol. (2017) 8:598. doi: 10.3389/fimmu.2017.00598
67. Pretorius E, Akeredolu OO, Soma P, Kell DB. Major involvement of bacterial components in rheumatoid arthritis and its accompanying oxidative stress, systemic inflammation and hypercoagulability. Exp Biol Med. (2017) 242:355–73. doi: 10.1177/1535370216681549
68. Hancock RE, Sahl HG. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol. (2006) 16:1551–7. doi: 10.1038/nbt1267
69. Sun Y, Shang D. Inhibitory effects of antimicrobial peptides on lipopolysaccharide-induced inflammation. Med Inflammation. (2015) 2015:167572. doi: 10.1155/2015/167572
70. Korpelainen S, Intke C, Hamalainen S, Jantunen E, Juutilainen A, Pulkki K. Soluble CD14 as a diagnostic and prognostic biomarker in hematological patients with febrile neutropenia. Dis Mark. (2017) 2017:9805609. doi: 10.1155/2017/9805609
71. Jones JW Jr, Su S, Jones MB, Heniford BT, McIntyre K, Granger DK. Serum lysozyme activity can differentiate infection from rejection in organ transplant recipients. J Surg Res. (1999) 84:134–7. doi: 10.1006/jsre.1999.5628
72. Wittel UA, Schmidt AI, Poxleitner PJ, Seifert GJ, Chikhladze S, Puolakkainen P, et al. The chemokine ligand CXCL16 is an indicator of bacterial infection in necrotizing pancreatitis. Pancreatology. (2015) 15:124–30. doi: 10.1016/j.pan.2015.01.004
73. Zweigner J, Gramm HJ, Singer OC, Wegscheider K, Schumann RR. High concentrations of lipopolysaccharide-binding protein in serum of patients with severe sepsis or septic shock inhibit the lipopolysaccharide response in human monocytes. Blood. (2001) 15:98:3800–8. doi: 10.1182/blood.V98.13.3800
74. Yu S, Nakashima N, Xu BH, MatsudaA T, zumihara I, Sunahara N, et al. Pathological significance of elevated soluble CD14 production in rheumatoid arthritis: in the presence of soluble CD14, lipopolysaccharides at low concentrations activate RA synovial fibroblasts. Rheumatol Int. (1998) 17:237–43. doi: 10.1007/s002960050041
75. Bas S, Gauthier BR, Spenato U, Stingelin S, Gabay C. CD14 is an acute-phase protein. J Immunol. (2004) 172:4470–9. doi: 10.4049/jimmunol.172.7.4470
76. Mikuls TR, LeVan TD, Sayles H, Yu F, Caplan L, Cannon GW, et al. Soluble CD14 and CD14 polymorphisms in rheumatoid arthritis. J Rheumatol. (2011) 38:2509–16. doi: 10.3899/jrheum.110378
77. Haziot A, Rong GW, Silver J, Goyert SM. Recombinant soluble CD14 mediates the activation of endothelial cells by lipopolysaccharide. J Immunol. (1993) 151:1500–7.
78. Loppnow H, Stelter F, Schonbeck U, Schluter C, Ernst M, Schutt C, et al. Endotoxin activates human vascular smooth muscle cells despite lack of expression of CD14 mRNA or endogenous membrane CD14. Infect Immun. (1995) 63:1020–6 doi: 10.1128/IAI.63.3.1020-1026.1995
79. Arditi MJ, Zhou R, Dorio GW, Rong SM, Goyert Kim KS. Endotoxin-mediated endothelial cell injury and activation: role of soluble CD14. Infect Immun. (1993) 61:3149–56. doi: 10.1128/IAI.61.8.3149-3156.1993
80. Arditi MJ, Zhou M, Torres DL, Durden M, Stins Kim KS. Lipopolysaccharide stimulates the tyrosine phosphorylation of mitogen-activated protein kinases p44, p42, and p41 in vascular endothelial cells in a soluble CD14-dependent manner. Role of protein tyrosine phosphorylationin lipopolysaccharide-induced stimulation of endothelial cells. J Immunol. (1995) 155:3994–4003.
81. Burgmann HS, Winkler GJ, Locker Presterl E, Laczika K, Staudinger T, et al. Increased serum concentration of soluble CD14 is a prognostic marker in gram-positive sepsis. Clin Immunol Immunopathol. (1996) 80:307–10. doi: 10.1006/clin.1996.0128
82. Hayashi J, Masaka T, Ishikawa I. Increased levels of soluble CD14 in sera of periodontitis patients. Infect Immun. (1999) 67:417–20. doi: 10.1128/IAI.67.1.417-420.1999
83. Lin B, Noring R, Steere AC, Klempner MS, Hu LT. Soluble CD14 levels in the serum, synovial fluid, and cerebrospinal fluid of patients with various stages of Lyme disease. J Infect Dis. (2000) 181:1185–8. doi: 10.1086/315357
84. Marcos V, Latzin P, Hector A, Sonanini S, Hoffmann F, Lacher M, et al. Expression, regulation and clinical significance of soluble and membrane CD14 receptors in pediatric inflammatory lung diseases. Respir Res. (2010) 11:32. doi: 10.1186/1465-9921-11-32
85. Giloteaux L, Goodrich JK, Walters WA, Levine SM, Ley RE, Hanson MR. Reduced diversity and altered composition of the gut microbiome in individuals with myalgic encephalomyelitis/chronic fatigue syndrome. Microbiome. (2016) 4:30. doi: 10.1186/s40168-016-0171-4
86. Tabung FK, Birmann BM, Epstein MM, Martínez-Maza O, Breen EC, Wu K, et al. Influence of dietary patterns on plasma soluble CD14, a surrogate marker of gut barrier dysfunction. Curr Dev Nutr. (2017) 1:e001396. doi: 10.3945/cdn.117.001396
87. Leveque M, Simonin-Le Jeune K, Jouneau S, Moulis S, Desrues B, Belleguic C, et al. Soluble CD14 acts as a DAMP in human macrophages: origin and involvement in inflammatory cytokine/chemokine production. FASEB J. (2017) 31:1891–902. doi: 10.1096/fj.201600772R
88. Jacque B, Stephan K, Smirnova I, Kim B, Gilling D, Poltorak A. Mice expressing high levels of soluble CD14 retain LPS in the circulation and are resistant to LPS-induced lethality. Eur J Immunol. (2006) 36:3007–16. doi: 10.1002/eji.200636038
89. Gallay P, Barras C, Tobias PS, Calandra T, Glauser MP, Heumann D. Lipopolysaccharide (LPS)-binding protein in human serum determines the tumor necrosis factor response of monocytes to LPS. J Infect Dis. (1994) 170:1319–22. doi: 10.1093/infdis/170.5.1319
90. Wen W, Li Y, Cheng Y, He J, Jia R, Li C, et al. Lipopolysaccharide-binding protein is a sensitive disease activity biomarker for rheumatoid arthritis. Clin Exp Rheumatol. (2018) 36:233–40. Available online at: https://www.clinexprheumatol.org/abstract.asp?a=11764
91. Todd DJ, Knowlton N, Amato M, Frank MB, Schur PH, Izmailova ES, et al. Erroneous augmentation of multiplex assay measurements in patients with rheumatoid arthritis due to heterophilic binding by serum rheumatoid factor. Arthritis Rheum. (2011) 63:894–903. doi: 10.1002/art.30213
92. Bartels EM, Watjen FI, Andersen EL, Danneskiold-Samsoe B, Bliddal H, Ribel-Madsen S. Rheumatoid factor and its interference with cytokine measurements: problems and solutions. Arthritis. (2011) 2011:741071. doi: 10.1155/2011/741071
93. Olsson P, Theander E, Bergström U, Jovinge S, Jacobsson L, Turesson C. Multiplex cytokine analyses in patients with rheumatoid arthritis require use of agents blocking heterophilic antibody activity. Scand J Rheumatol. (2017) 46:1–10. doi: 10.3109/03009742.2016.1161070
94. Myc A, Buck J, Gonin J, Reynolds B, Hammerling U, Emanuel D. The level of lipopolysaccharide-binding protein is significantly increased in plasma in patients with the systemic inflammatory response syndrome. Clin Diagn Lab Immunol. (1997) 4:113–6. doi: 10.1128/CDLI.4.2.113-116.1997
95. Blairon L, Wittebole X, Laterre PF. Lipopolysaccharide-binding protein serum levels in patients with severe sepsis due to gram-positive and fungal infections. J Infect Dis. (2003) 187:287–91. doi: 10.1086/346046
96. Torsteinsdottir I, Hakansson L, Hallgren R, Gudbjornsson B, Arvidson NG, Venge P. Serum lysozyme: a potential marker of monocyte/macrophage activity in rheumatoid arthritis. Rheumatology (1999) 38:1249–54. doi: 10.1093/rheumatology/38.12.1249
97. Kluter T, Fitschen-Oestern S, Lippross S, Weuster M, Mentlein R, Steubesand N, et al. The antimicrobial peptide lysozyme is induced after multiple trauma. Med Inflamm. (2014) 2014:303106. doi: 10.1155/2014/303106
98. Meng F, Lowell CA. Lipopolysaccharide (LPS)-induced macrophage activation and signal transduction in the absence of Src-family kinases Hck, Fgr, and Lyn. J Exp Med. (1997) 185:1661–70. doi: 10.1084/jem.185.9.1661
99. Soler-Rodriguez AM, Zhang H, Lichenstein HS, Qureshi N, Niesel DW, Crowe SE, et al. Neutrophil activation by bacterial lipoprotein versus lipopolysaccharide: differential requirements for serum and CD14. J Immunol. (2000) 164:2674–83. doi: 10.4049/jimmunol.164.5.2674
100. Rubio C. Increased production of lysozyme associated with bacterial proliferation in barrett's esophagitis, chronic gastritis, gluten-induced atrophic duodenitis (celiac disease), lymphocyte colitis, collagenous colitis, ulcerative colitis and crohn's colitis. Anticancer Res. (2015) 35:6365–72. Available online at: http://ar.iiarjournals.org/content/35/12/6365.full.pdf+html
101. Arango Duque G, Descoteaux A. Macrophage cytokines: involvement in immunity and infectious diseases. Front Immunol. (2014) 5:491. doi: 10.3389/fimmu.2014.00491
102. Kageyama Y, Torikai E, Nagano A. Anti-tumor necrosis factor-alpha antibody treatment reduces serum CXCL16 levels inpatients with rheumatoid arthritis. Rheumatol Int. (2007) 27:467–72. doi: 10.1007/s00296-006-0241-1
103. Shimaoka T, Nakayama T, Hieshima K, Kume N, Fukumoto N, Minami M, et al. Chemokines generally exhibit scavenger receptor activity through their receptor-binding domain. J Biol Chem. (2004) 279:26807–10. doi: 10.1074/jbc.C400163200
104. Gutwein P, Abdel-Bakky MS, Schramme A, et al. CXCL16 is expressed in podocytes and acts as a scavenger receptor for oxidized low-density lipoprotein. Am J Pathol. (2009) 174:2061–72. doi: 10.2353/ajpath.2009.080960
105. Jiang X, Shimaoka T, Kojo S, Harada M, Watarai H, Wakao H, et al. Cutting edge: critical role of CXCL16/CXCR6 in NKT cell trafficking in allograft tolerance. J Immunol. (2005) 175:2051–5. doi: 10.4049/jimmunol.175.4.2051
106. van der Voort R, van Lieshout AW, Toonen LW, Sloetjes AW, van den Berg WB, Figdor CG, et al. Elevated CXCL16 expression by synovial macrophages recruits memory T cells into rheumatoid joints. Arthritis Rheum. (2005) 52:1381–91. doi: 10.1002/art.21004
107. Sheikine Y, Sirsjo A. CXCL16/SR-PSOX-a friend or a foe in atherosclerosis? Atherosclerosis. (2008) 197:487–95. doi: 10.1016/j.atherosclerosis.2007.11.034
108. Lehrke M, Millington SC, Lefterova M, Cumaranatunge RG, Szapary P, Wilensky R, et al. CXCL16 is a marker of inflammation, atherosclerosis, and acute coronary syndromes in humans. J Am Coll Cardiol. (2007) 49:442–9. doi: 10.1016/j.jacc.2006.09.034
109. Kang S, Yang C, Luo R. LysoPtdOH enhances CXCL16 production stimulated by LPS from macrophages and regulates T cell migration. Lipids. (2008) 43:1075–83. doi: 10.1007/s11745-008-3238-6
110. Rashid T, Jayakumar KS, Binder A, Ellis S, Cunningham P, Ebringer A. Rheumatoid arthritis patients have elevated antibodies to cross-reactive and non cross-reactive antigens from Proteus microbes. Clin Exp Rheumatol. (2007) 25:259–67. Available online at: https://www.clinexprheumatol.org/abstract.asp?a=3060
111. Nagase H, Agematsu K, Kitano K, Takamoto M, Okubo Y, Komiyama A, et al. Mechanism of hypergammaglobulinemia by HIV infection: circulating memory B-cell reduction with plasmacytosis. Clin Immunol. (2001) 100:250–9. doi: 10.1006/clim.2001.5054
112. De Milito A, Nilsson A, Titanji K, Thorstensson R, Reizenstein E, Narita M, et al. Mechanisms of hypergammaglobulinemia and impaired antigen-specific humoral immunity in HIV-1 infection. Blood. (2004) 103:2180–6. doi: 10.1182/blood-2003-07-2375
113. Xu W, Luo Z, Alekseyenko AV, Martin L, Wan Z, Ling B, et al. Distinct systemic microbiome and microbial translocation are associated with plasma level of anti-CD4 autoantibody in HIV infection. Sci Rep. (2018) 8:12863. doi: 10.1038/s41598-018-31116-y
114. Filipp D, Alizadeh-Khiavi K, Richardson C, et al. Soluble CD14 enriched in colostrum and milk induces B cell growth and differentiation. Proc Natl Acad Sci USA. (2001) 98:603–8. doi: 10.1073/pnas.98.2.603
115. Riva S, Nolli ML, Lutz MB, Citterio S, Girolomoni G, Winzler C, et al. Bacteria and bacterial cell wall constituents induce the product ion of regulatory cytokines in dendritic cell clones. J Inflamm. (1996) 46:98–105.
116. Aiba S, Tagami H. Dendritic cell activation induced by various stimuli, e.g. exposure to microorganisms, their products, cytokines, and simple chemicals as well as adhesion to extracellular matrix. J Dermatol Sci. (1998) 20:1–13. doi: 10.1016/S0923-1811(99)00005-5
117. Vazquez MI, Catalan-Dibene J, Zlotnik A. B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine. (2015) 74:318–26. doi: 10.1016/j.cyto.2015.02.007
118. Markle JGM, Frank DN, Mortin-Toth S, Robertson CE, Feazel LM, Rolle-Kampczyk U, et al. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science. (2013) 339:1084–8. doi: 10.1126/science.1233521
119. Yurkovetskiy L, Burrows M, Khan AA, Graham L, Volchkov P, Becker L, et al. Gender bias in autoimmunity is influenced by microbiota. Immunity. (2013) 39:400–12. doi: 10.1016/j.immuni.2013.08.013
120. Elderman M, Hugenholtz F, Belzer C, Boekschoten M, van Beek A, de Haan B, et al. Sex and strain dependent differences in mucosal immunology and microbiota composition in mice. Biol Sex Differ. (2018) 9:26. doi: 10.1186/s13293-018-0186-6
121. Rubtsova K, Marrack P, Rubtsov AV. Sexual dimorphism in autoimmunity. J Clin Invest. (2015) 125:2187–93. doi: 10.1172/JCI78082
122. Taneja V. Sex hormones determine immune response. Front Immunol. (2018) 9:1931. doi: 10.3389/fimmu.2018.01931
123. Li B, Selmi C, Tang R, Gershwin ME, Ma X. The microbiome and autoimmunity: a paradigm from the gut-liver axis. Cell Mol Immunol. (2018) 15:595–609. doi: 10.1038/cmi.2018.7
124. Chervonsky AV. Microbiota and Autoimmunity. Cold Spring Harb Perspect Biol. (2013) 5:a007294. doi: 10.1101/cshperspect.a007294
125. Manfredo Vieira S, Hiltensperger M, Kumar V, Zegarra-Ruiz D, Dehner C, Khan N, et al. Translocation of a gut pathobiont drives autoimmunity in mice and humans. Science. (2018) 9:1156–61. doi: 10.1126/science.aar7201
126. Brown EM, Kenny DJ, Xavier RJ. Gut microbiota regulation of T cells during inflammation and autoimmunity. Ann Rev Immunol. (2019) 37:599–624. doi: 10.1146/annurev-immunol-042718-041841
127. Ayyappan P, Harms RZ, Buckner JH, Sarvetnick NE. Coordinated induction of antimicrobial response factors in systemic lupus erythematosus. Front Immunol. (2019) 10:658. doi: 10.3389/fimmu.2019.00658
Keywords: rheumatoid arthritis, antimicrobial proteins, EndoCAbs, sCD14, CXCL16, lysozyme
Citation: Ayyappan P, Harms RZ, Seifert JA, Bemis EA, Feser ML, Deane KD, Demoruelle MK, Mikuls TR, Holers VM and Sarvetnick NE (2020) Heightened Levels of Antimicrobial Response Factors in Patients With Rheumatoid Arthritis. Front. Immunol. 11:427. doi: 10.3389/fimmu.2020.00427
Received: 22 November 2019; Accepted: 25 February 2020;
Published: 20 March 2020.
Edited by:
Kutty Selva Nandakumar, Southern Medical University, ChinaReviewed by:
Allen C. Steere, Harvard Medical School, United StatesLars Klareskog, Karolinska Institutet, Sweden
Copyright © 2020 Ayyappan, Harms, Seifert, Bemis, Feser, Deane, Demoruelle, Mikuls, Holers and Sarvetnick. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nora E. Sarvetnick, bm9yYXNAdW5tYy5lZHU=