 Jasmin Knopf
Jasmin Knopf Moritz Leppkes
Moritz Leppkes Georg Schett
Georg Schett Martin Herrmann
Martin Herrmann Luis E. Muñoz
Luis E. Muñoz- 1Department of Medicine 3 – Rheumatology and Immunology, Universitätsklinikum Erlangen, Erlangen, Germany
- 2Department of Medicine 1 – Gastroenterology, Pneumology and Endocrinology, Universitätsklinikum Erlangen, Erlangen, Germany
In response to various infectious and sterile stimuli neutrophils release chromatin decorated with bactericidal proteins, referred to as NETs. Their scaffolds are formed from chromatin fibers which display an apparent diameter of 15–17 nm and mainly consist from DNA (2 nm) and DNA-associated histones (11 nm). The NET-forming strands are thus not naked DNA but higher ordered chromatin structures. The histones may be released from the NET, especially if their tail arginines have been citrullinated. Several studies indicate that extracellular histones are toxic for mammalian epithelia and endothelia and contribute to the microvascular dysfunction observed e.g., in patients suffering from autoimmune diseases or sepsis. NETs formed at sites of very high neutrophil densities tend to clump and form fairly stable enzymatically active aggregates, referred to as aggNETs. The latter are endowed with a bunch of enzymes that cleave, bind, and/or modify autologous as well as foreign macromolecules. The tight binding of the serine proteases to the matrix precludes the spread of these toxic enzymes into the tissue but still allows the access of soluble inflammatory mediators to the enzymatic active internal surfaces of the NETs where they are degraded. Here, we describe that externally added histones are removed from culture supernatants of aggNETs. We will address the fate of the histones and discuss the feature on the background of neutrophil-driven diseases and the resolution of inflammation.
Introduction
Histones are a major part of nucleosomes, the basic structural unit of chromatin in the nuclei of eukaryotic cells (1). These nucleosomes each consist of two copies of the histones H2A, H2B, H3, and H4 forming an octamer with 140–150 base pairs of superhelical DNA wrapped around the histone core (2). The linker histone H1 assembles the repeating nucleosome cores into higher-order structures (3). Biosynthesis of histones takes places in the cytoplasm and many histones transiently remain there (4, 5); some reportedly accumulate on the plasma membranes (6). Extra-nuclear localization of histones is also found associated with the DNA structures in neutrophil extracellular traps (NETs), first described in 2004 as bactericidal mechanism (7). Furthermore, histones display anti-microbial activity reviewed in Hoeksema et al. (8), and have been implicated in tissue destruction, sepsis (9), and thrombosis (10). Mechanistically, histones reportedly display direct cytotoxic effect on eukaryotic cells (11), may directly activate phagocytes (12) and platelets (13).
Since their first description in 2004, NETs are now known to play a role in physiology and pathology (14). In high densities these NETs tend to aggregate; these aggNETs, first described to be induced by monosodium urate crystals (MSU) orchestrate the resolution of inflammation in gout by the degradation of inflammatory cytokines (15). With increasing cell densities the proteolytic degradation of cytokines/chemokines outweighs their release (16). The granular neutrophil elastase, a major protein of NETs and aggNETs, degrades various proteins of the extracellular matrix or immunoglobulins (17). Here, we describe that histones are sequestered and detoxified by aggNETs and that this increased viability of epithelial cells in contact to extracellular histones.
Materials and Methods
Preparation of AggNETs
We isolated polymorphonuclear cells (PMN) from healthy donors (permit #193/13B from the local ethical committee; written informed consent of participants) by Ficoll density gradient (Lymphoflot, Bio-Rad Laboratories, Inc.) as described previously (18). The granulocytes were then incubated with 50 pg/cell monosodium urate crystals (MSU) for 18 h at 37°C. Successful formation of aggNETs is visible without magnification as depicted in Figure S1E in bright-field as well as under UV (~312 nm excitation) after staining with 1 mg/ml propidium iodide (Sigma-Aldrich) for 2 h. Macrophotographs were taken using a Nikon D700.
Biotinylation of Histones
We biotinylated calf thymus histones (Sigma-Aldrich) using the EZ-Link™ Sulfo-NHS-LC-Biotinylation Kit (Thermo Fisher Scientific) according to manufacturer's instructions.
Treatment of Histones With AggNETs/Neutrophil Elastase/Proteinase3
We incubated 1 mg/ml of biotinylated histones with or without (1) aggNETs, (2) 5 mU Neutrophil Elastase (Sigma-Aldrich) or, (3) 5 mU Proteinase3 (Elastin Products Company) in RPMI 1640 medium (Thermo Fisher Scientific) for 24 h at 37°C. If indicated we added the neutrophil elastase inhibitors Sivelestat (6.6 μM) or Elafin (166 μM) (both Sigma-Aldrich).
SDS-PAGE and Western Blot Analysis
We added 5x PAGE-buffer (2% SDS, 10% glycerol, 5% β-mercaptoethanol, 0.01% bromophenol blue in 60 mM Tris-Cl pH 6.8) to the samples and denatured them at 95°C for 10 min. SDS-PAGE was performed using SERVAGelTM TG PRiMETM 4–20% gels (SERVA Electrophoresis GmbH) for 2.5 h at 100 V. Gels were either transferred onto an Immobilon-PSQ PVDF membrane (Merck Millipore Ltd.) using a Trans-Blot® SD Semi-Dry Transfer Cell (Bio-Rad Laboratories, Inc.) for 1 h at 350 mA or stained with 0.1% Coomassie Brilliant-Blue-G250 (Sigma-Aldrich). Macrophotographs of the Coomassie gels were taken using a Nikon D700. Membranes were blocked with 5% powdered milk (Carl Roth) in Tris-buffered saline (TBS) for 2 h at RT. We detected histone H1 employing rabbit anti-human histone H1.0 antibody [EPR6536] (ab134914, Abcam) overnight at 4°C followed by goat anti-rabbit IgG HRP Antibody (4030-05, Southern Biotech) for 1 h at RT. Biotinylation was detected with Pierce™ High Sensitivity Streptavidin-HRP (21130, Thermo Fisher Scientific). We developed Blots using Celvin® S-320+ (Biostep).
Prediction and Visualization of Neutrophil Elastase Cleaving Sites
We used the sequence of bovine histone H1.3 (A7MAZ5, UniProtKB) to model its structure with SWISS-MODEL (19). Neutrophil Elastase cleavage sites on histone H1.3 were predicted using the ExPASy PeptideCutter tool (20) and were visualized using the RasMol Molecular Graphics Visualization Tool V2.7.5 (21).
In vitro Histone Cytotoxicity Assay
Analyses by flow cytometry of HeLa cells treated with soluble histones or aggNET pre-treated histones was performed using the four color staining method adapted from Janko et al. (22) and Munoz et al. (23). Briefly, 24 h after seeding of HeLa cells into CELLSTAR® 24-well plates (Greiner Bio-One GmbH), the cells were treated for 1 h with 500 μg of histones, histones pre-incubated with aggNETs or aggNET supernatant in serum-free medium. Mock-treated cells served as controls. After removal of the media to fresh tubes, we washed the cells with DPBS (Thermo Fisher Scientific), detached them using Trypsin/EDTA (Merck) and combined them with the original media. After centrifugation, cells were resuspended in Ringer's solution (Fresenius Kabi) containing 1 μg/ml AnnexinA5, 1 μg/ml propidium iodide, 1 μg/ml Hoechst33342, and 10 nM 1,1′-dimethyl-3,3,3′,3′-tetramethylindodicarbocyanine iodide. Flow cytometry was performed using a Gallios Flow Cytometer (Beckman-Coulter) and Kaluza Analysis Software V2.1 (Beckman-Coulter). Graphs were created using Prism® V5.03 (GraphPad Software). Pictures of cells were taken using a Canon Eos 6D, the Eos Utility3 software (both Canon) in combination with an Axiovert 25 inverted microscope (Carl Zeiss) and the Adobe Photoshop CS5 V12.0.1 (Adobe Systems).
Results
AggNETs Proteolytically Degrade Histones
Incubation of calf thymus histones with aggNETs for 24 h results in a complete degradation of histone H1 (Figure 1A) as shown by Coomassie staining of protein. Histone H1 was only detected by Western Blot analysis in the untreated sample, but neither in the aggNET-treated sample nor in the aggNET itself. We biotinylated the histone samples to exclude that the epitope recognized by the antibody was cleaved and therefore not recognized by Western Blotting. The biotinylation was again only detected in the untreated sample but neither in the aggNET-treated ones nor in the aggNETs. Proteinase3 (PR3) and Neutrophil Elastase (NE) are hallmark proteases located in the azurophilic granula of viable neutrophils and on the surfaces of aggNETs. As shown in Figures 1B,C, PR3 and NE degrade histone H1; the reaction is prevented by the specific inhibitors Elafin and Sivelestat, respectively. Prediction by ExPASy PeptideCutter shows that bovine histone H1.3 (amino acids 39–119) exhibits various cleavage sites for NE (Figure 1D). Importantly, this degradation favors histone over bovine serum albumin (BSA) or human immunoglobulin G (IgG) (Figure 1E). Only NE and aggNETs but not PR3 slightly decrease the amount of full-size BSA and IgG. For NE, this was prevented by its specific inhibitor Sivelestat. Surprisingly, neither the addition of Sivelestat nor of Elafin nor a combination of both blocked the degradation of histones by aggNETs at any given time point and concentration.
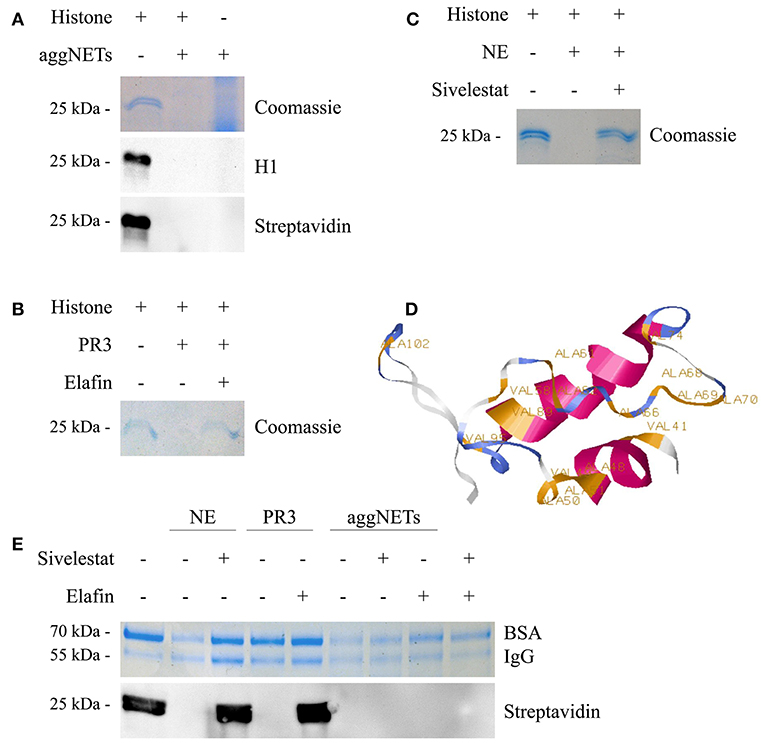
Figure 1. aggNETs degrade histones. (A) Histones incubated with aggNETs are degraded as seen in Coomassie and staining and anti-histone H1 Blot. The biotinylationed histones are not bound by the aggNETs. (B) Proteinase3 (PR3) degrades histones. This degradation is inhibited by Elafin as seen in the Coomassie staining. (C) Neutrophil Elastase (NE) degrades histones, specifically inhibited by Sivelestat as shown in the Coomassie staining. (D) SWISS-MODEL of histone H1 (amino acids 39–119) with the cleavage sites for NE as predicted by ExPASy PeptideCutter. (E) NE and aggNETs favor histone over bovine serum albumin (BSA) and human Immunoglobulin G (IgG) for degradation; whereas PR3 can only degrade histones. Degradation of biotinylated histones by aggNETs is not inhibited by Sivelestat or Elafin or a combination of both as seen by the detection of Streptavidin HRP in Western Blot analysis. SDS-PAGE, Western Blot Analysis and Coomassie staining in (A–C) were performed after incubation of the samples for 24 h at 37°C. For (E) the incubation time was 8 h at 37°C. All images shown are representative images of at least three independent experiments. The full-sized images are shown in Figures S1A–D. The successful formation of an aggNET is shown in the macrophotographs in Figure S1E in bright-field and under UV after staining with propidium iodide.
AggNET-Treatment of Histones Attenuates Cellular Cytotoxicity
As soon as 1 h after treatment with 500 μg/ml histone mix HeLa cells are in a bad shape, increase clustering and apparently die as displayed in the bright-field microscopic images (Figure 2A). Pre-treatment of histones with aggNETs prevented this fate. The supernatants of aggNET (aggNET-SN) did not affect the viability of the cells. Flow cytometry revealed that culture in the presence of histones markedly reduced viability and increased apoptosis and necrosis in HeLa cells (Figure 2B). This histone-mediated cytotoxicity is attenuated by pre-treatment with aggNETs. Detailed analyses of the different forms of cell death is depicted in Figure 2C and showed that the pre-treatment with aggNETs significantly decreased early apoptotic, apoptotic and primary necrotic cells; the population of secondary necrotic HeLa cells was only slightly increased. HeLa cells co-cultured with aggNET-SN show comparable viability as medium controls. Therefore, we can exclude that the incomplete rescue in aggNET pre-treated histones is caused by toxic aggNET- derived mediators.
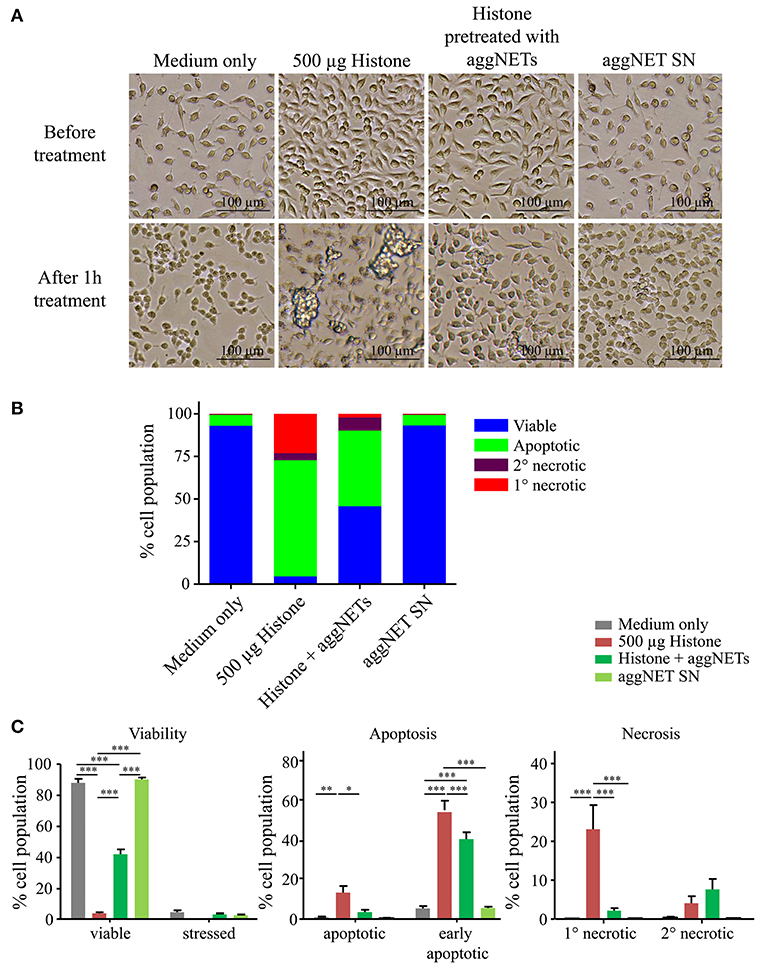
Figure 2. Pre-treatment of histones with aggNETs attenuates histone-mediated cellular cytotoxicity. (A) Light microscope images in 10x magnification of HeLa cells before and after treatment with (1) 500 μg histones, (2) histones pre-treated with aggNETs for 24 h or (3) aggNET supernatant (SN). (B) Overview of different forms of cell death of HeLa cells after treatment assessed by flow cytometry. Pre-treatment of histones with aggNETs increases viability of HeLa cells. (C) Detailed analysis of the different forms of cell death. Viability of HeLa cells incubated with aggNET-treated histones is significantly increased compared to histone treatment due to a reduction in early apoptosis, apoptosis and primary (1°) necrosis. Standard error of mean was calculated from three independent experiments. ***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05 as determined by Two-way ANOVA with Bonferroni post testing. The gating strategy for flow cytometer analysis is depicted in Figure S2.
Discussion
Here, we show for the first time that aggNETs sequester and degrade histones, and thus attenuate their cytotoxic effect on epithelial cells. This process was executed by at least two aggNET-borne serine proteases, NE and PR3. We already have demonstrated the ability of aggNETs to resolve inflammation by the proteolytical degradation of inflammatory cytokines and chemokines (15, 16). NE is established to degrade various proteins, such as immunoglobulins and extracellular matrix components (17, 24). The degradation of histones by NE and PR3 was inhibited by Sivelestat or Elafin, respectively. Importantly, the degradation of histones by aggNETs was resistant to the two inhibitors. Interestingly, a decreased inhibitory capacity of the natural proteinous inhibitors α-1 anti-trypsin and ß2-macroglobulin for membrane-associated NE was already reported before the first description of NET formation (25).
The cytotoxic effect of histones on epithelial cells described here, confirms already existing literature (9, 12). This is especially true when the histone release is exaggerated and not properly controlled. Here, we analyze this cytotoxic effect in more detail using a four color staining method to discriminate between different states of apoptosis and necrosis as described previously (22, 23). The addition of histones to HeLa cells induced profound apoptosis and necrosis (>90% of the cells). This can be partially rescued pre-treating the histones with aggNETs. This procedure increased viability to 50%. To examine if toxic proteins/peptides are released from the aggNETs, we also co-cultured the cells with aggNET-SN only; the cell viability did not differ from medium controls. It is conceivable that some of the small histone-derived peptides, too small to be detected in PAGE, retain residual cytotoxic activities.
Extracellular histones are described as major mediators of death in sepsis due to their contribution to endothelial injury and dysfunction, hemorrhage, thrombosis and organ failure (9). Released histones potentially act as damage-associated molecular pattern molecules (DAMPs) (26) and signal through toll-like receptors (TLR) 2 and 4 leading to a massive pro-inflammatory cytokine production (27). Moreover, histones are shown to enhance plasma thrombin generation and the blood clotting process by involvement of the platelet TLR2 and TLR4 (13). Histones are further released during trauma or severe cellular stress mediating their cytotoxicity by triggering an increased calcium flux in immune and endothelial cells (12).
Histones, released during NET formation, were described in the circulation of patients suffering from autoimmune diseases such as systemic lupus erythematosus (28) or rheumatoid arthritis (29) and were discussed to cause NET-associated tissue destruction. Histones not only act as autoantigens but also prevented the degradation of DNA by the formation of DNA-histone complexes (28). Here we describe that aggNETs degrade and detoxify histones and thus contribute to the resolution of histone-induced inflammatory reactions. If this also takes place in vivo and how it can be further enhanced to completely rescue cells from histone-mediated cytotoxicity, needs further investigation. We conclude that histones are targeted by aggNETs for degradation. This leads to a decreased cytotoxicity of histones and, therefore, fosters the resolution of inflammation.
Ethics Statement
All analyses of human material were performed in full agreement with institutional guidelines and with the approval of the Ethical committee of the Universitätsklinikum Erlangen (permit # 193 13B). Participants gave written informed consent.
Author Contributions
JK planned and performed experiments, conducted data analysis, and wrote the manuscript. ML, GS, MH, and LM supervised the project, planned experiments, performed data analysis, and wrote the manuscript. All authors read and approved the manuscript.
Funding
This work was partially supported by the German Research Foundation (DFG) to MH (CRC1181-C03, TRR241-B04), GS (CRC1181-A01, -Z02), and ML (TRR241-B04), the Volkswagen Foundation project 90361 (MH) and by the doctoral training program GK1660 of the DFG to JK. The authors acknowledge support by DFG and Friedrich-Alexander-Universität Erlangen-Nürnberg (FAU) within the funding program Open Access Publishing.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.02176/full#supplementary-material
References
1. Kornberg RD. Structure of chromatin. Annu Rev Biochem. (1977) 46:931–54. doi: 10.1146/annurev.bi.46.070177.004435
2. Mcghee JD, Felsenfeld G. Nucleosome structure. Annu Rev Biochem. (1980) 49:1115–56. doi: 10.1146/annurev.bi.49.070180.005343
3. Allan J, Hartman PG, Crane-Robinson C, Aviles FX. The structure of histone H1 and its location in chromatin. Nature. (1980) 288:675–9. doi: 10.1038/288675a0
4. Zlatanova JS, Srebreva LN, Banchev TB, Tasheva BT, Tsanev RG. Cytoplasmic pool of histone H1 in mammalian cells. J Cell Sci. (1990) 96(Pt 3):461–8.
5. Chang L, Loranger SS, Mizzen C, Ernst SG, Allis CD, Annunziato AT. Histones in transit: cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry. (1997) 36:469–80. doi: 10.1021/bi962069i
6. Watson K, Edwards RJ, Shaunak S, Parmelee DC, Sarraf C, Gooderham NJ, et al. Extra-nuclear location of histones in activated human peripheral blood lymphocytes and cultured T-cells. Biochem Pharmacol. (1995) 50:299–309. doi: 10.1016/0006-2952(95)00142-M
7. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. (2004) 303:1532–5. doi: 10.1126/science.1092385
8. Hoeksema M, Van Eijk M, Haagsman HP, Hartshorn KL. Histones as mediators of host defense, inflammation and thrombosis. Future Microbiol. (2016) 11:441–53. doi: 10.2217/fmb.15.151
9. Xu J, Zhang X, Pelayo R, Monestier M, Ammollo CT, Semeraro F, et al. Extracellular histones are major mediators of death in sepsis. Nat Med. (2009) 15:1318–21. doi: 10.1038/nm.2053
10. Michels A, Albanez S, Mewburn J, Nesbitt K, Gould TJ, Liaw PC, et al. Histones link inflammation and thrombosis through the induction of Weibel-Palade body exocytosis. J Thromb Haemost. (2016) 14:2274–86. doi: 10.1111/jth.13493
11. Wildhagen KC, Garcia De Frutos P, Reutelingsperger CP, Schrijver R, Areste C, Ortega-Gomez A, et al. Nonanticoagulant heparin prevents histone-mediated cytotoxicity in vitro and improves survival in sepsis. Blood. (2014) 123:1098–101. doi: 10.1182/blood-2013-07-514984
12. Abrams ST, Zhang N, Manson J, Liu T, Dart C, Baluwa F, et al. Circulating histones are mediators of trauma-associated lung injury. Am J Respir Crit Care Med. (2013) 187:160–9. doi: 10.1164/rccm.201206-1037OC
13. Semeraro F, Ammollo CT, Morrissey JH, Dale GL, Friese P, Esmon NL, et al. Extracellular histones promote thrombin generation through platelet-dependent mechanisms: involvement of platelet TLR2 and TLR4. Blood. (2011) 118:1952–61. doi: 10.1182/blood-2011-03-343061
14. Manda A, Pruchniak MP, Arazna M, Demkow UA. Neutrophil extracellular traps in physiology and pathology. Cent Eur J Immunol. (2014) 39:116–21. doi: 10.5114/ceji.2014.42136
15. Schauer C, Janko C, Munoz LE, Zhao Y, Kienhofer D, Frey B, et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat Med. (2014) 20:511–7. doi: 10.1038/nm.3547
16. Hahn J, Schauer C, Czegley C, Kling L, Petru L, Schmid B, et al. Aggregated neutrophil extracellular traps resolve inflammation by proteolysis of cytokines and chemokines and protection from antiproteases. FASEB J. (2018) 2018:fj201800752R. doi: 10.1096/fj.201800752R
17. Shapiro SD. Neutrophil elastase: path clearer, pathogen killer, or just pathologic? Am J Respir Cell Mol Biol. (2002) 26:266–8. doi: 10.1165/ajrcmb.26.3.f233
18. Biermann MH, Podolska MJ, Knopf J, Reinwald C, Weidner D, Maueroder C, et al. Oxidative burst-dependent NETosis is implicated in the resolution of necrosis-associated sterile inflammation. Front Immunol. (2016) 7:557. doi: 10.3389/fimmu.2016.00557
19. Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. (2018) 46:W296–303. doi: 10.1093/nar/gky427
20. Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins MR, Appel RD, et al. Protein identification and analysis tools on the ExPASy server. Proteomics Protocols Handbook. (2005) 571–607. doi: 10.1385/1-59259-890-0:571
21. Bernstein HJ. Recent changes to RasMol, recombining the variants. Trends Biochem Sci. (2000) 25:453–5. doi: 10.1016/S0968-0004(00)01606-6
22. Janko C, Munoz L, Chaurio R, Maueroder C, Berens C, Lauber K, et al. Navigation to the graveyard-induction of various pathways of necrosis and their classification by flow cytometry. Methods Mol Biol. (2013) 1004:3–15. doi: 10.1007/978-1-62703-383-1_1
23. Munoz LE, Maueroder C, Chaurio R, Berens C, Herrmann M, Janko C. Colourful death: six-parameter classification of cell death by flow cytometry–dead cells tell tales. Autoimmunity. (2013) 46:336–41. doi: 10.3109/08916934.2012.755960
24. Eckle I, Kolb G, Havemann K. Inhibition of neutrophil oxidative burst by elastase-generated IgG fragments. Biol Chem Hoppe Seyler. (1990) 371:69–77. doi: 10.1515/bchm3.1990.371.1.69
25. Owen CA, Campbell MA, Sannes PL, Boukedes SS, Campbell EJ. Cell surface-bound elastase and cathepsin G on human neutrophils: a novel, non-oxidative mechanism by which neutrophils focus and preserve catalytic activity of serine proteinases. J Cell Biol. (1995) 131:775–89. doi: 10.1083/jcb.131.3.775
26. Chen R, Kang R, Fan XG, Tang D. Release and activity of histone in diseases. Cell Death Dis. (2014) 5:e1370. doi: 10.1038/cddis.2014.337
27. Xu J, Zhang X, Monestier M, Esmon NL, Esmon CT. Extracellular histones are mediators of death through TLR2 and TLR4 in mouse fatal liver injury. J Immunol. (2011) 187:2626–31. doi: 10.4049/jimmunol.1003930
28. Hakkim A, Furnrohr BG, Amann K, Laube B, Abed UA, Brinkmann V, et al. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc Natl Acad Sci USA. (2010) 107:9813–8. doi: 10.1073/pnas.0909927107
Keywords: histones, NET formation, aggNETs, proteolysis, autoimmunity, sepsis
Citation: Knopf J, Leppkes M, Schett G, Herrmann M and Muñoz LE (2019) Aggregated NETs Sequester and Detoxify Extracellular Histones. Front. Immunol. 10:2176. doi: 10.3389/fimmu.2019.02176
Received: 14 December 2018; Accepted: 28 August 2019;
Published: 11 September 2019.
Edited by:
Reinhard Edmund Voll, Freiburg University Medical Center, GermanyReviewed by:
Stamatis-Nick C. Liossis, University of Patras, GreeceAngelo A. Manfredi, Vita-Salute San Raffaele University, Italy
Copyright © 2019 Knopf, Leppkes, Schett, Herrmann and Muñoz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Herrmann, bWFydGluLmhlcnJtYW5uQHVrLWVybGFuZ2VuLmRl