
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 06 June 2019
Sec. Cancer Immunity and Immunotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fimmu.2019.01126
This article is part of the Research Topic NKT Cells in Cancer Immunotherapy View all 18 articles
 Yingting Zhang†Ryan Springfield†Siyang Chen†Xin LiXiaotian FengRosa Moshirian
Yingting Zhang†Ryan Springfield†Siyang Chen†Xin LiXiaotian FengRosa Moshirian Rirong Yang‡
Rirong Yang‡ Weiming Yuan*
Weiming Yuan*NKT cells are CD1d-restricted innate-like T cells expressing both T cell receptor and NK cell markers. The major group of NKT cells in both human and mice is the invariant NKT (iNKT) cells and the best-known function of iNKT cells is their potent anti-tumor function in mice. Since its discovery 25 years ago, the prototype ligand of iNKT cells, α-galactosylceramide (α-GalCer) has been used in over 30 anti-tumor clinical trials with mostly suboptimal outcomes. To realize its therapeutic potential, numerous preclinical models have been developed to optimize the scheme and strategies for α-GalCer-based cancer immunotherapies. Nevertheless, since there is no standard protocol for α-GalCer delivery, we reviewed the preclinical studies with a focus on B16 melanoma model in the goal of identifying the best treatment schemes for α-GalCer treatment. We then reviewed the current progress in developing more clinically relevant mouse models for these preclinical studies, most notably the generation of new mouse models with a humanized CD1d/iNKT cell system. With ever-emerging novel iNKT cell ligands, invention of novel α-GalCer delivery strategies and significantly improved preclinical models for optimizing these new strategies, one can be hopeful that the full potential of anti-tumor potential for α-GalCer will be realized in the not too distant future.
Natural Killer T (NKT) cells are CD1d-restricted innate-like T cells expressing both T cell receptor and NK cell markers (1). Invariant NKT (iNKT) cells are the major group of NKT cells in both human and mice. They express the invariant Vα24-Jα18 chains and Vα14-Jα18 TCRα chains in human and mice, respectively (2–4). Since their discovery in the early 1990s, the best-studied function of iNKT cells has been their anti-tumor function. Activated iNKT cells rapidly secrete both Th1 and Th2 cytokines and activate NK and other immune cells to stimulate anti-tumor immune responses (5, 6). The prototypical iNKT cell ligand, α-galactosylceramide (α-GalCer), is a sphingolipid that was first isolated from the marine sponge Agelas mauritianas in 1994 by chloroform extraction and HPLC purification techniques (7). Mice injected with free α-GalCer demonstrated potent anti-tumor activity against metastatic B16 melanoma cells (7, 8). Furthermore, α-GalCer demonstrated synergistic anti-tumor effects when co-administered with another chemotherapy agent, adriamycin (8), suggesting α-GalCer has a different target other than the tumor cells themselves. Interestingly, early researchers believed that α-GalCer was a non-specific immunostimulatory agent (8). However, by 1998, studies using knockout mice had concluded that α-GalCer's anti-tumor properties were mediated by CD1d-restricted iNKT cells (9, 10). Several excellent reviews have been published recently on the anti-tumor function of α-GalCer and iNKT cells (5, 6, 11–15). Here we aim to review commonly used preclinical mouse models for α-GalCer and iNKT cell-based cancer immunotherapy to compare and contrast the different approaches in mobilizing iNKT cells for anti-tumor therapies. We specifically focus on the syngeneic mouse B16 melanoma model, a well-established model for human melanoma (16). For a comprehensive review of preclinical modeling of α-GalCer-based cancer therapy targeting diverse tumor types, readers are referred to a recent review by Nair and Dhodapkar (6).
Despite extensive literature on the anti-tumor function of α-GalCer and iNKT cells, no standard procedure has been established in delivering the iNKT cell ligands. Procedures vary widely between different research groups. The glycolipid ligands can be administered prior to, simultaneously or after the inoculation of tumor cells at different time points, via intravenous, intraperitoneal or subcutaneous routes, and in free or vehicled forms. Clearly, the scheme, time points, routes and forms for glycolipid deliveries all have a significant impact on the ensuing immune response and therefore the interpretation of the results.
In initial reports on the anti-tumor function of α-GalCer [KRN7000, a close analog of original AGL9b (8)], the glycolipid was injected post B16 cell inoculation. For example, Morita et al. (8) reported that α-GalCer injected 1, 5, and 9 days after subcutaneous inoculation of B16 cells suppressed the tumor volume growth by about 50%. Glycolipid treatment before tumor inoculation represents a prophylactic treatment and may be particularly applicable for future tumor vaccination with specific neoantigens, while the post-tumor inoculation studies are more clinically relevant for anti-cancer therapies.
While several reports have demonstrated that pre-treatment with α-GalCer can lead to an anti-tumor response in mice, one study found that injecting α-GalCer immediately before tumor inoculation does not show an anti-tumor effect (17) (Table 1). However, pre-administration of a single-dose α-GalCer 2 days prior to B16 cell inoculation leads to powerful anti-metastatic effect (20). This has been confirmed by ours and other studies (14, 21–23). It is unclear how long the anti-tumor response can last, but it is unlikely to last too long, for example 30 days, as the NKT cells will become anergic by then (17, 25). More kinetic experiments are warranted to determine the duration of this anti-tumor response before the anergy induction because the information will be important for future repetitive administration of α-GalCer and its analogs in clinics.
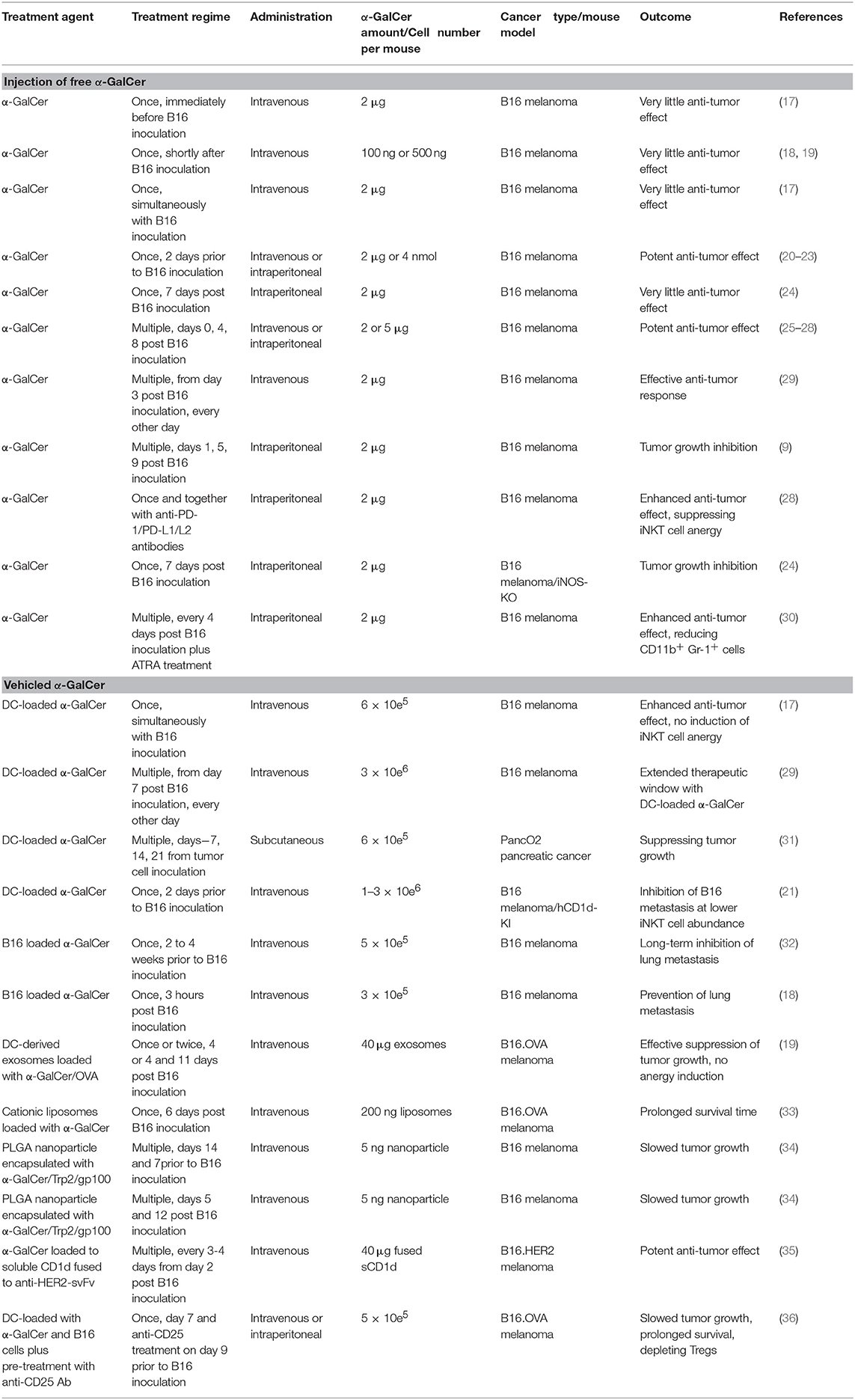
Table 1. Preclinical studies of α-GalCer and iNKT cell-mediated anti-tumor therapies.
Several reports showed that one single injection of α-GalCer either simultaneously or shortly after the B16 melanoma inoculation does not inhibit tumor growth (17–19, 24). Similarly, a single α-GalCer treatment 4 days (19), or seven days after B16 cell inoculation (24) had little beneficial effect on suppressing tumor growth or mouse survival. Therefore, most reports investigating anti-B16 function of α-GalCer have utilized multiple dosages of α-GalCer, typically in a three-dose scheme at days 0, 4, and 8 post B16 inoculation (25–28). In one study, repetitive administration of α-GalCer was initiated at different time points post B16 inoculation (29). α-GalCer was administrated every other day until the end of the experiment on day 14. The free α-GalCer glycolipid demonstrated anti-B16 function as late as 3 days after tumor inoculation, but not beyond 5 days after (29). This may be due to immune-suppression by the established B16 tumors as reported (30). On the other hand, DC-vehicled α-GalCer clearly can extend this treatment window to at least seven days after B16 inoculation (29), suggesting that the vehicled α-GalCer is more efficient in boosting immune response and/or overcoming tumor-led immune suppression.
Many possible mechanisms have been proposed for the suboptimal efficacies of α-GalCer in anti-tumor clinical trials (5, 6, 11), such as the induction of anergy, the secretion of both Th1 and Th2 cytokines by iNKT cells and immune suppression by the tumors in the microenvironment (30). Many novel α-GalCer analogs have been designed to increase the Th1/Th2 ratio and enhance the anti-tumor immunity (22, 37, 38). While we focus on the anti-tumor function of the prototypic α-GalCer, the chemistry and anti-tumor efficacy and mechanism for these novel α-GalCer analogs have been elegantly reviewed elsewhere (38).
Pioneering work from Fujii and Van Kaer groups demonstrated the induction of long-lasting anergy post α-GalCer activation of iNKT cells (17, 25). The anergy induction not only makes further activation of iNKT cells inefficient, anergic iNKT cells can actually exacerbate tumorigenesis upon further stimulation by glycolipids (25).
The arguably best approach by far to overcome iNKT cell anergy is to load the α-GalCer to dendritic cells (17). Although the absolute amounts of Th1/Th2 cytokines secreted post DC-loaded α-GalCer were not as high as that of free α-GalCer and the cytokines were secreted at a delayed kinetics, the DC-vehicled α-GalCer stimulated higher numbers of cytokine-secreting splenocytes. Importantly, DC-loaded α-GalCer does not lead to iNKT cell anergy (17). More importantly, the DC-vehicled α-GalCer showed more potent anti-tumor activity than free α-GalCer in the B16 melanoma model (17). Interestingly, in this study, both the free α-GalCer and DC-loaded α-GalCer were administered simultaneously with the B16 melanoma cells. While co-injected α-GalCer does not induce immediate anti-tumor activity as discussed above, DC-vehicled α-GalCer can immediately induce anti-tumor activity. Free α-GalCer takes 2 days to induce an anti-tumor response in mice (20), suggesting that these two approaches boosted different downstream effectors. It is particularly important to note that NK cells are only responsible for approximately half of the anti-tumor effect for DC-vehicled α-GalCer (17), while they account for almost all of free α-GalCer mediated anti-tumor function (26, 39, 40). Given the fact that DC-loaded α-GalCer has been widely used in anti-tumor clinical trials (31, 41–43), it is important to further delineate the exact anti-tumor mechanism of DC-vehicled α-GalCer.
The second reported approach to suppress NKT cell anergy is to use exosomes loaded with α-GalCer (19). While in early clinical trials, exosomes loaded with tumor antigens have mostly been tolerated and had little immunostimulatory effects (44, 45), exosomes loaded with α-GalCer as an immune-stimulatory adjuvant led to an effective anti-tumor responses in mice (19). Using a subcutaneous B16 melanoma model, Gehrmann et al. (19) demonstrated that dendritic cells-derived exosomes loaded with α-GalCer administered 4 days after tumor inoculation could effectively suppress tumor growth and extend mouse survival. More importantly, a second injection of loaded exosomes 1 week after the first one can further inhibit tumor growth, suggesting that the first injection with α-GalCer-loaded exosomes did not induce anergy.
Rejuvenating anergic NKT cells at molecular levels is the third approach for suppressing NKT cell anergy. Expression of inhibitory co-stimulatory molecules including PD-1 and PD-L1/L2 is partially responsible for the anergy of NKT cells (28). Three injections of anti-PD-L1/L2 or anti-PD-1 antibodies post α-GalCer activation of iNKT cells could maintain the iNKT cells response for at least 30 days after the α-GalCer treatment (28). This allowed the recovery of iNKT cells to a responsive state and repeated activation of iNKT cells with α-GalCer extended the anti-B16 metastatic function (28). Considering the recent success of anti-PD-L1/2 and anti-PD-1 antibodies in rejuvenating tumor-specific T cells in clinics, future combination treatment with these antibodies and α-GalCer may synergize their anti-tumor functions.
IL-2 has shown anti-anergy function to iNKT cells. In light of its function in breaking anergy of conventional T cells (46), Parekh et al. (25) demonstrated that IL-2, but not IL-12, IFN-γ or IL-4 could re-stimulate the anergic iNKT cells to proliferate both in vitro and in vivo.
One major innovation in the field pioneered by the Fujii group is to load α-GalCer to the tumor cells for immunization (18, 32). Even for low immunogenicity tumor cells including B16 melanoma cells, one single vaccination with α-GalCer-loaded tumor cells could stimulate potent tumor-specific CD8+ T cell responses. Memory CD4 and CD8 T cells could protect the immunized mice from tumor re-challenge for as long as 6–12 months (32). It was also demonstrated that CD1d expression significantly improved the efficacy of iNKT cell-based therapies, presumably due to increased efficiency of direct killing by iNKT cells. Therefore, CD1d expression on tumor cells can be a positive biomarker for future iNKT cell therapies in clinics, as suggested by another report (47). Importantly, the tumor protection from vaccination in this study is tumor-specific. The mice were only immune to the specific tumor that was used for vaccination (32). On the other hand, α-GalCer-loaded dendritic cells induce short-term tumor resistance against different types of tumors, including melanoma (29), multiple myeloma (48), pancreatic cancer (31) and B cell lymphoma (49). These studies suggested that dendritic cells loaded with α-GalCer induce mostly innate immunity-based non-specific anti-tumor responses including activated NK cells, whereas tumor cells loaded with α-GalCer induce more specific long-term adaptive immunity-based anti-tumor responses.
Several other groups have explored delivering tumor cells or specific tumor antigens with α-GalCer using vehicles such as dendritic cells (36, 50, 51), dendritic cells-derived exosomes (19), dendritic cells loaded with tumor-derived exosomes (52), PLGA nanoparticles (34), cationic liposomes (33), chemically conjugated α-GalCer-tumor peptide antigen compound vaccine (53), or α-GalCer-loaded recombinant soluble CD1d protein fused with single chain antibodies against neoantigen (35, 54). In vivo, the tumor antigens are either directly or cross-presented by endogenous dendritic cells to CD8+ T cells while the co-delivered α-GalCer is presented to iNKT cells. As expected, all of these approaches have shown enhanced tumor antigen-specific CTL responses and increased IFN-γ secretion in these T cells. These approaches have demonstrated both prophylactic (36), or therapeutic effects (19, 34, 50–53) to challenges by vaccinated tumors. One Phase I trial has been completed using dendritic cells loaded with α-GalCer and the well-established neoantigen NY-ESO-1 (51). It is encouraging that there were increases in NKT cell proliferation, NKT cell-associated cytokine secretion and more importantly, the circulating NY-ESO-1-specific T cells in most (7 out 8) patients (51).
It has been well-established that CD4+CD25+ Treg cells suppress anti-tumor immunity (55, 56). On the other hand, several reports showed that α-GalCer-activated NKT cells secret IL-2 leading to the expansion of Treg cells (57, 58). Pre-administration of depleting anti-CD25 monoclonal antibody (PC61) 2 days prior to α-GalCer vaccination increased the α-GalCer-induced prophylactic anti-tumor function in a subcutaneous challenge model with B16 melanoma cells (36). However, pre-administration with the same PC61 antibody prior to α-GalCer treatment did not enhance the anti-tumor function of α-GalCer in a therapeutic tumor challenge model with a lung tumor cell line TC1 (59). Interestingly, in the Petersen report (36), α-GalCer challenge and NKT cell activation did not induce an expansion of Treg cells as previously reported (57). This difference is likely due to the different routes of α-GalCer delivery. While in the previous report, delivery of free α-GalCer led to Treg expansion (57), the α-GalCer delivered in dendritic cell-vehicled form in the later study did not (36). More studies are needed to delineate the interaction between iNKT cells and Tregs in order to manipulate Tregs for the benefit of iNKT cell-mediated cancer therapies.
IFN-γ is one of the major cytokine effectors after α-GalCer administration (1). The high amount of IFN-γ induces immuno-suppressive factors including the iNOS enzyme, which produces nitric oxide and inhibits anti-tumor immunity (60). In iNOS-knockout mice or wild-type mice treated with an iNOS inhibitor, L-NAME, the B16 metastasis was more efficiently suppressed by a suboptimal treatment of α-GalCer (one single treatment seven days after B16 melanoma inoculation) (24). Another study demonstrated that lung metastasis of B16 melanoma was also significantly inhibited by a suboptimal treatment of α-GalCer when the mice were simultaneously treated with all-trans-retinoic acid (ATRA) (30). ATRA, a derivative of vitamin A, can induce the differentiation of CD11b+Gr-1+ immature myeloid cells and reduce this major nitric oxide-producing population (30).
There are more innovative approaches of enhancing anti-tumor activity of α-GalCer, such as adoptive iNKT cell transfer (61, 62), using artificial antigen-presenting cells to expand iNKT cells in vitro (63), co-administration of NK cell activator, IL-18 (64). Altogether, all the reported approaches could increase α-GalCer function. Clearly more research is required to realize their therapeutic potential and achieve the optimal therapeutic efficacy by combining these novel approaches.
The sharp difference between mouse and human immune systems, including the difference in the CD1d/iNKT cell system, urgently demand better mouse models with improved predictive powers for clinics. In addition to the significantly lower affinities of the human CD1d and iNKT TCR to α-GalCer compared to that of mice (65, 66), human iNKT cells are present at a much lower abundance with very different subset compositions (21, 67, 68). The journey from the original discovery of α-GalCer's anti-tumor function in mice to current clinical trials also suggests that preclinical modeling with more relevant mouse strains is warranted before translating α-GalCer and its analogs into clinics.
One attractive direction to improve the preclinical modeling of α-GalCer-based immunotherapies is to develop mouse models with a human-like CD1d/iNKT TCR system. The first mouse model with a humanized CD1d/NKT cell system is from the Wang group in which human CD1d is expressed under a mouse MHC class I (Kb) promoter (69). Human CD1d is highly expressed in all nucleated cells as a MHC class I expression pattern. It is not clear how NKT cells are developed in this strain. However, it was clear that the exogenous human CD1d can function as a strong transplantation antigen (69). The second mouse model generated by the Casorati group expressed human CD1d using Lck or CD11c promoters to direct specific human CD1d expression in thymocytes or dendritic cells, respectively (70). By breeding to CD1d-knockout mice, the authors demonstrated that thymocyte-specific expression of human CD1d alone is sufficient to support iNKT cell development (70). However, because of no human CD1d expression on dendritic cells in these pLck-hCD1dTg mice, in vivo α-GalCer treatment is not feasible to test the α-GalCer-based cancer immunotherapy. For a human-like iNKT cell population, the Casorati group generated a pre-arranged human invariant Vα24-Jα18 TCRα chain and expressed it as a transgene under the human CD2 promoter (71). In the Jα18-knockout background, which eliminates the expression of mouse iNKT TCRα chain (Vα14), the human Vα24-Jα18 TCRα chain could support the development of human-like Vα24 iNKT cells. The Gumperz group has utilized the humanized SCID mice to generate mice with a humanized CD1d/iNKT cell system. Immune-deficient mice were engrafted with human fetal thymus, liver and CD34+ hematopoietic cells. Four surface CD1 gene family members, CD1a, CD1b, CD1c, and CD1d were all expressed in vivo. Furthermore, T cell responses have been detected for all the CD1 family members. In addition, α-GalCer can stimulate IFN-γ secretion in the mouse serum, suggesting the NKT cells are developed and functional in vivo (72). Nevertheless, more investigation on the immune cell development and adaptive immune responses may be needed before this engrafted system can be widely used for modeling NKT cell-based cancer immunotherapies.
Our group has been working on yet another approach to humanize the CD1d/iNKT cell system. By homologous recombination, we generated a human CD1d knock-in mouse, in which human CD1d is under the endogenous mouse CD1d promoter (21). Consistent with the previous report (70), thymic expression of human CD1d supports NKT cell development. Importantly, this new human CD1d-knock in mouse possesses an iNKT cell population with human-like abundance and similar subset composition in terms of co-receptor expression pattern (21), making this strain a particularly useful tool for modeling in vivo human iNKT cell responses to α-GalCer or its analogs. By expressing the pre-arranged human Vα24/Jα18 TCRα chain (23), this further improved mouse strain can be particularly instrumental to test and optimize the glycolipid ligands for anti-tumor therapies. However, since the human Vα24/Jα18 TCRα is a transgene, the current mouse strain is not optimal for investigating the antigen-specific T cell responses during anti-tumor immunotherapies. Nevertheless, since the NK cells and other innate immune cells are not affected by the transgene, this strain can still be used to investigate the innate immunity-mediated anti-tumor function of α-GalCer. To further improve this model, future “knock-in” of human Vα24/Jα18 and Vβ11 genomic regions will be necessary. The continuous improvement of current gene-editing techniques, including CRISPR-Cas9 (73), may make the knock-in more feasible. For preclinical modeling of α-GalCer-mediated anti-tumor therapy, we have demonstrated that prophylactic treatment with α-GalCer in the two CD1d-humanized mouse strains can suppress B16 metastasis (21, 23). Nevertheless, it will be most interesting to investigate whether α-GalCer can suppress B16 melanoma in these humanized mice under therapeutic settings, and if not, how the treatment regimes can be improved for an optimal anti-tumor effect.
In summary, joint efforts from researchers in chemistry, pharmaceutics and immunology fields will bring about more potent α-GalCer analogs, optimized delivery and treatment schemes and much-improved preclinical models. We envision that the α-GalCer-based cancer immunotherapy will be reaching its full potential in clinics in the near future.
WY, YZ, RS, and SC participated in conceptualization and drafting of the article as well as critical revision of the article for important intellectual content. All authors participated in writing and revision of the manuscript and gave final approval of the submitted publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work has been supported by NIH R01 AI 0919878, NIH U01 GM 111849 and supported in part by NIH P30CA014089 grant to the University of Southern California Norris Comprehensive Cancer Center from the National Cancer Institute. We thank Miss Christine Quach for critically reading our manuscript and apologize to the investigators whose work can not be included in this review due to space limit.
1. Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. (2007) 25:297–336. doi: 10.1146/annurev.immunol.25.022106.141711
2. Porcelli S, Yockey CE, Brenner MB, Balk SP. Analysis of T cell antigen receptor (TCR) expression by human peripheral blood CD4-8− alpha/beta T cells demonstrates preferential use of several V beta genes and an invariant TCR alpha chain. J Exp Med. (1993) 178:1–16.
3. Lantz O, Bendelac A. An invariant T cell receptor alpha chain is used by a unique subset of major histocompatibility complex class I-specific CD4+ and CD4-8− T cells in mice and humans. J Exp Med. (1994) 180:1097–106.
4. Porcelli S, Gerdes D, Fertig AM, Balk SP. Human T cells expressing an invariant V alpha 24-J alpha Q TCR alpha are CD4− and heterogeneous with respect to TCR beta expression. Hum Immunol. (1996) 48:63–7.
5. Bedard M, Salio M, Cerundolo V. Harnessing the power of invariant natural killer T cells in cancer immunotherapy. Front Immunol. (2017) 8:1829. doi: 10.3389/fimmu.2017.01829
6. Nair S, Dhodapkar MV. Natural killer T cells in cancer immunotherapy. Front Immunol. (2017) 8:1178. doi: 10.3389/fimmu.2017.01178
7. Natori T, Morita M, Akimoto K, Koezuka Y. Agelasphins, novel antitumor and immunostimulatory cerebrosides from the marine sponge agelas-mauritianus. Tetrahedron. (1994) 50:2771–84. doi: 10.1016/S0040-4020(01)86991-X
8. Morita M, Motoki K, Akimoto K, Natori T, Sakai T, Sawa E, et al. Structure-activity relationship of alpha-galactosylceramides against B16-bearing mice. J Med Chem. (1995) 38:2176–87.
9. Kawano T, Cui J, Koezuka Y, Toura I, Kaneko Y, Sato H, et al. Natural killer-like nonspecific tumor cell lysis mediated by specific ligand-activated Valpha14 NKT cells. Proc Natl Acad Sci USA. (1998) 95:5690–3.
10. Brossay L, Chioda M, Burdin N, Koezuka Y, Casorati G, Dellabona P, et al. CD1d-mediated recognition of an alpha-galactosylceramide by natural killer T cells is highly conserved through mammalian evolution. J Exp Med. (1998) 188:1521–8.
11. Wolf BJ, Choi JE, Exley MA. Novel approaches to exploiting invariant NKT cells in cancer immunotherapy. Front Immunol. (2018) 9:384. doi: 10.3389/fimmu.2018.00384
12. Teyton L. New directions for natural killer T cells in the immunotherapy of cancer. Front Immunol. (2017) 8:1480. doi: 10.3389/fimmu.2017.01480
13. Shissler SC, Lee MS, Webb TJ. Mixed signals: co-stimulation in invariant natural killer T cell-mediated cancer immunotherapy. Front Immunol. (2017) 8:1447. doi: 10.3389/fimmu.2017.01447
14. King LA, Lameris R, de Gruijl TD, van der Vliet HJ. CD1d-invariant natural killer T cell-based cancer immunotherapy: α-galactosylceramide and beyond. Front Immunol. (2018) 9:1519. doi: 10.3389/fimmu.2018.01519
15. Fujii S, Shimizu K, Okamoto Y, Kunii N, Nakayama T, Motohashi S, et al. NKT cells as an ideal anti-tumor immunotherapeutic. Front Immunol. (2013) 4:409. doi: 10.3389/fimmu.2013.00409
16. Overwijk WW, Restifo NP. B16 as a mouse model for human melanoma. Curr Protoc Immunol. (2001) Chapter 20:Unit 20.21. doi: 10.1002/0471142735.im2001s39
17. Fujii S, Shimizu K, Kronenberg M, Steinman RM. Prolonged IFN-gamma-producing NKT response induced with alpha-galactosylceramide-loaded DCs. Nat Immunol. (2002) 3:867–74. doi: 10.1038/ni827
18. Shimizu K, Goto A, Fukui M, Taniguchi M, Fujii S. Tumor cells loaded with alpha-galactosylceramide induce innate NKT and NK cell-dependent resistance to tumor implantation in mice. J Immunol. (2007) 178:2853–61. doi: 10.4049/jimmunol.178.5.2853
19. Gehrmann U, Hiltbrunner S, Georgoudaki AM, Karlsson MC, Näslund TI, Gabrielsson S. Synergistic induction of adaptive antitumor immunity by codelivery of antigen with alpha-galactosylceramide on exosomes. Cancer Res. (2013) 73:3865–76. doi: 10.1158/0008-5472.can-12-3918
20. Schmieg J, Yang G, Franck RW, Tsuji M. Superior protection against malaria and melanoma metastases by a C-glycoside analogue of the natural killer T cell ligand alpha-Galactosylceramide. J Exp Med. (2003) 198:1631–41. doi: 10.1084/jem.20031192jem.20031192
21. Wen X, Rao P, Carreño LJ, Kim S, Lawrenczyk A, Porcelli SA, et al. Human CD1d knock-in mouse model demonstrates potent antitumor potential of human CD1d-restricted invariant natural killer T cells. Proc Natl Acad Sci USA. (2013) 110:2963–8. doi: 10.1073/pnas.1300200110
22. Chennamadhavuni D, Saavedra-Avila NA, Carreño LJ, Guberman-Pfeffer MJ, Arora P, Yongqing T, et al. Dual modifications of alpha-galactosylceramide synergize to promote activation of human invariant natural killer T cells and stimulate anti-tumor immunity. Cell Chem Biol. (2018) 25:571–84.e578. doi: 10.1016/j.chembiol.2018.02.009
23. Wen X, Kim S, Xiong R, Li M, Lawrenczyk A, Huang X, et al. A subset of CD8alphabeta+ invariant NKT cells in a humanized mouse model. J Immunol. (2015) 195:1459–69. doi: 10.4049/jimmunol.1500574
24. Ito H, Ando T, Seishima M. Inhibition of iNOS activity enhances the anti-tumor effects of alpha-galactosylceramide in established murine cancer model. Oncotarget. (2015) 6:41863–74. doi: 10.18632/oncotarget.6172
25. Parekh VV, Wilson MT, Olivares-Villagómez D, Singh AK, Wu L, Wang CR, et al. Glycolipid antigen induces long-term natural killer T cell anergy in mice. J Clin Invest. (2005) 115:2572–83. doi: 10.1172/JCI24762
26. Smyth MJ, Crowe NY, Pellicci DG, Kyparissoudis K, Kelly JM, Takeda K, et al. Sequential production of interferon-gamma by NK1.1+ T cells and natural killer cells is essential for the antimetastatic effect of alpha-galactosylceramide. Blood. (2002) 99:1259–66. doi: 10.1182/blood.v99.4.1259
27. Okajo J, Kaneko Y, Murata Y, Tomizawa T, Okuzawa C, Saito Y, et al. Regulation by Src homology 2 domain-containing protein tyrosine phosphatase substrate-1 of alpha-galactosylceramide-induced antimetastatic activity and Th1 and Th2 responses of NKT cells. J Immunol. (2007) 178:6164–72. doi: 10.4049/jimmunol.178.10.6164
28. Parekh VV, Lalani S, Kim S, Halder R, Azuma M, Yagita H, et al. PD-1/PD-L blockade prevents anergy induction and enhances the anti-tumor activities of glycolipid-activated invariant NKT cells. J Immunol. (2009) 182:2816–26. doi: 10.4049/jimmunol.0803648
29. Toura I, Kawano T, Akutsu Y, Nakayama T, Ochiai T, Taniguchi M. Cutting edge: inhibition of experimental tumor metastasis by dendritic cells pulsed with alpha-galactosylceramide. J Immunol. (1999) 163:2387–91.
30. Yanagisawa K, Exley MA, Jiang X, Ohkochi N, Taniguchi M, Seino K. Hyporesponsiveness to natural killer T-cell ligand alpha-galactosylceramide in cancer-bearing state mediated by CD11b+ Gr-1+ cells producing nitric oxide. Cancer Res. (2006) 66:11441–6. doi: 10.1158/0008-5472.can-06-0944
31. Nagaraj S, Ziske C, Strehl J, Messmer D, Sauerbruch T, Schmidt-Wolf IG. Dendritic cells pulsed with alpha-galactosylceramide induce anti-tumor immunity against pancreatic cancer in vivo. Int Immunol. (2006) 18:1279–83. doi: 10.1093/intimm/dxl059
32. Shimizu K, Kurosawa Y, Taniguchi M, Steinman RM, Fujii S. Cross-presentation of glycolipid from tumor cells loaded with alpha-galactosylceramide leads to potent and long-lived T cell mediated immunity via dendritic cells. J Exp Med. (2007) 204:2641–53. doi: 10.1084/jem.20070458
33. Neumann S, Young K, Compton B, Anderson R, Painter G, Hook S. Synthetic TRP2 long-peptide and alpha-galactosylceramide formulated into cationic liposomes elicit CD8+ T-cell responses and prevent tumour progression. Vaccine. (2015) 33:5838–44. doi: 10.1016/j.vaccine.2015.08.083
34. Ghinnagow R, De Meester J, Cruz LJ, Aspord C, Corgnac S, Macho-Fernandez E, et al. Co-delivery of the NKT agonist α-galactosylceramide and tumor antigens to cross-priming dendritic cells breaks tolerance to self-antigens and promotes antitumor responses. Oncoimmunology. (2017) 6:e1339855. doi: 10.1080/2162402X.2017.1339855
35. Corgnac S, Perret R, Derré L, Zhang L, Stirnemann K, Zauderer M, et al. CD1d-antibody fusion proteins target iNKT cells to the tumor and trigger long-term therapeutic responses. Cancer Immunol Immunother. (2013) 62:747–60. doi: 10.1007/s00262-012-1381-7
36. Petersen TR, Sika-Paotonu D, Knight DA, Dickgreber N, Farrand KJ, Ronchese F, et al. Potent anti-tumor responses to immunization with dendritic cells loaded with tumor tissue and an NKT cell ligand. Immunol Cell Biol. (2010) 88:596–604. doi: 10.1038/icb.2010.9
37. Aspeslagh S, Li Y, Yu ED, Pauwels N, Trappeniers M, Girardi E, et al. Galactose-modified iNKT cell agonists stabilized by an induced fit of CD1d prevent tumour metastasis. EMBO J. (2011) 30:2294–305. doi: 10.1038/emboj.2011.145
38. Banchet-Cadeddu A, Hénon E, Dauchez M, Renault JH, Monneaux F, Haudrechy A. The stimulating adventure of KRN 7000. Org Biomol Chem. (2011) 9:3080–104. doi: 10.1039/c0ob00975j
39. Hayakawa Y, Takeda K, Yagita H, Kakuta S, Iwakura Y, Van Kaer L, et al. Critical contribution of IFN-gamma and NK cells, but not perforin-mediated cytotoxicity, to anti-metastatic effect of alpha-galactosylceramide. Eur J Immunol. (2001) 31:1720–7. doi: 10.1002/1521-4141(200106)31:6<1720::aid-immu1720>3.3.co;2-l
40. Nakagawa R, Nagafune I, Tazunoki Y, Ehara H, Tomura H, Iijima R, et al. Mechanisms of the antimetastatic effect in the liver and of the hepatocyte injury induced by alpha-galactosylceramide in mice. J Immunol. (2001) 166:6578–84. doi: 10.4049/jimmunol.166.11.6578
41. Richter J, Neparidze N, Zhang L, Nair S, Monesmith T, Sundaram R, et al. Clinical regressions and broad immune activation following combination therapy targeting human NKT cells in myeloma. Blood. (2013) 121:423–30. doi: 10.1182/blood-2012-06-435503
42. Chang DH, Osman K, Connolly J, Kukreja A, Krasovsky J, Pack M, et al. Sustained expansion of NKT cells and antigen-specific T cells after injection of alpha-galactosyl-ceramide loaded mature dendritic cells in cancer patients. J Exp Med. (2005) 201:1503–17. doi: 10.1084/jem.20042592
43. Exley MA, Nakayama T. NKT-cell-based immunotherapies in clinical trials. Clin Immunol. (2011) 140:117–8. doi: 10.1016/j.clim.2011.04.015
44. Escudier B, Dorval T, Chaput N, André F, Caby MP, Novault S, et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of thefirst phase I clinical trial. J Transl Med. (2005) 3:10. doi: 10.1186/1479-5876-3-10
45. Morse MA, Garst J, Osada T, Khan S, Hobeika A, Clay TM, et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J Transl Med. (2005) 3:9. doi: 10.1186/1479-5876-3-9
46. Schwartz RH. T cell anergy. Annu Rev Immunol. (2003) 21:305–34. doi: 10.1146/annurev.immunol.21.120601.141110
47. Haraguchi K, Takahashi T, Nakahara F, Matsumoto A, Kurokawa M, Ogawa S, et al. CD1d expression level in tumor cells is an important determinant for anti-tumor immunity by natural killer T cells. Leuk Lymphoma. (2006) 47:2218–23. doi: 10.1080/10428190600682688
48. Nur H, Rao L, Frassanito MA, De Raeve H, Ribatti D, Mfopou JK, et al. Stimulation of invariant natural killer T cells by alpha-Galactosylceramide activates the JAK-STAT pathway in endothelial cells and reduces angiogenesis in the 5T33 multiple myeloma model. Br J Haematol. (2014) 167:651–63. doi: 10.1111/bjh.13092
49. Escribà-Garcia L, Alvarez-Fernández C, Tellez-Gabriel M, Sierra J, Briones J. Dendritic cells combined with tumor cells and alpha-galactosylceramide induce a potent, therapeutic and NK-cell dependent antitumor immunity in B cell lymphoma. J Transl Med. (2017) 15:115. doi: 10.1186/s12967-017-1219-3
50. Matsuyoshi H, Hirata S, Yoshitake Y, Motomura Y, Fukuma D, Kurisaki A, et al. Therapeutic effect of alpha-galactosylceramide-loaded dendritic cells genetically engineered to express SLC/CCL21 along with tumor antigen against peritoneally disseminated tumor cells. Cancer Sci. (2005) 96:889–96. doi: 10.1111/j.1349-7006.2005.00123.x
51. Gasser O, Sharples KJ, Barrow C, Williams GM, Bauer E, Wood CE, et al. A phase I vaccination study with dendritic cells loaded with NY-ESO-1 and alpha-galactosylceramide: induction of polyfunctional T cells in high-risk melanoma patients. Cancer Immunol Immunother. (2018) 67:285–98. doi: 10.1007/s00262-017-2085-9
52. Liu H, Chen L, Liu J, Meng H, Zhang R, Ma L, et al. Co-delivery of tumor-derived exosomes with alpha-galactosylceramide on dendritic cell-based immunotherapy for glioblastoma. Cancer Lett. (2017) 411:182–90. doi: 10.1016/j.canlet.2017.09.022
53. Anderson RJ, Compton BJ, Tang CW, Authier-Hall A, Hayman CM, Swinerd GW, et al. NKT cell-dependent glycolipid-peptide vaccines with potent anti-tumour activity. Chem Sci. (2015) 6:5120–7. doi: 10.1039/c4sc03599b
54. Stirnemann K, Romero JF, Baldi L, Robert B, Cesson V, Besra GS, et al. Sustained activation and tumor targeting of NKT cells using a CD1d-anti-HER2-scFv fusion protein induce antitumor effects in mice. J Clin Invest. (2008) 118:994–1005. doi: 10.1172/jci33249
55. Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. (2017) 27:109–18. doi: 10.1038/cr.2016.151
56. Whiteside TL. FOXP3+ Treg as a therapeutic target for promoting anti-tumor immunity. Expert Opin Ther Targets. (2018) 22:353–63. doi: 10.1080/14728222.2018.1451514
57. Liu R, La Cava A, Bai XF, Jee Y, Price M, Campagnolo DI, et al. Cooperation of invariant NKT cells and CD4+CD25+ T regulatory cells in the prevention of autoimmune myasthenia. J Immunol. (2005) 175:7898–904. doi: 10.4049/jimmunol.175.12.7898
58. Jiang S, Game DS, Davies D, Lombardi G, Lechler RI. Activated CD1d-restricted natural killer T cells secrete IL-2: innate help for CD4+CD25+ regulatory T cells? Eur J Immunol. (2005) 35:1193–200. doi: 10.1002/eji.200425899
59. Rosalia RA, Štěpánek I, Polláková V, Šímová J, Bieblová J, Indrová M, et al. Administration of anti-CD25 mAb leads to impaired alpha-galactosylceramide-mediated induction of IFN-gamma production in a murine model. Immunobiology. (2013) 218:851–9. doi: 10.1016/j.imbio.2012.10.012
60. Ekmekcioglu S, Grimm EA, Roszik J. Targeting iNOS to increase efficacy of immunotherapies. Human Vaccines Immunotherapeut. (2017) 13:1105–8. doi: 10.1080/21645515.2016.1276682
61. Exley MA, Friedlander P, Alatrakchi N, Vriend L, Yue S, Sasada T, et al. Adoptive transfer of invariant NKT cells as immunotherapy for advanced melanoma: a phase I clinical trial. Clin Cancer Res. (2017) 23:3510–9. doi: 10.1158/1078-0432.ccr-16-0600
62. Smith DJ, Liu S, Ji S, Li B, McLaughlin J, Cheng D, et al. Genetic engineering of hematopoietic stem cells to generate invariant natural killer T cells. Proc Natl Acad Sci USA. (2015) 112:1523–8. doi: 10.1073/pnas.1424877112
63. Webb TJ, Bieler JG, Schneck JP, Oelke M. Ex vivo induction and expansion of natural killer T cells by CD1d1-Ig coated artificial antigen presenting cells. J Immunol Methods. (2009) 346:38–44. doi: 10.1016/j.jim.2009.05.003
64. Nishio S, Yamada N, Ohyama H, Yamanegi K, Nakasho K, Hata M, et al. Enhanced suppression of pulmonary metastasis of malignant melanoma cells by combined administration of alpha-galactosylceramide and interleukin-18. Cancer Sci. (2008) 99:113–20. doi: 10.1111/j.1349-7006.2007.00636.x
65. Pellicci DG, Patel O, Kjer-Nielsen L, Pang SS, Sullivan LC, Kyparissoudis K, et al. Differential recognition of CD1d-alpha-galactosyl ceramide by the V beta 8.2 and V beta 7 semi-invariant NKT T cell receptors. Immunity. (2009) 31:47–59. doi: 10.1016/j.immuni.2009.04.018
66. Koch M, Stronge VS, Shepherd D, Gadola SD, Mathew B, Ritter G, et al. The crystal structure of human CD1d with and without alpha-galactosylceramide. Nat Immunol. (2005) 6:819–26. doi: 10.1038/ni1225
67. Berzins SP, Smyth MJ, Baxter AG. Presumed guilty: natural killer T cell defects and human disease. Nat Rev Immunol. (2011) 11:131–42. doi: 10.1038/nri2904
68. Wen X, Xiong R, Dai Z, Kim S, Lawrenczyk A, Yuan W. Humanizing mice for the identification of novel anticancer lipids targeting iNKT cells. Oncoimmunology. (2013) 2:e25475. doi: 10.4161/onci.25475
69. Wang B, Chun T, Rulifson IC, Exley M, Balk SP, Wang CR. Human CD1d functions as a transplantation antigen and a restriction element in mice. J Immunol. (2001) 166:3829–36. doi: 10.4049/jimmunol.166.6.3829
70. Schümann J, Pittoni P, Tonti E, Macdonald HR, Dellabona P, Casorati G. Targeted expression of human CD1d in transgenic mice reveals independent roles for thymocytes and thymic APCs in positive and negative selection of Valpha14i NKT cells. J Immunol. (2005) 175:7303–10. doi: 10.4049/jimmunol.175.11.7303
71. Capone M, Cantarella D, Schümann J, Naidenko OV, Garavaglia C, Beermann F, et al. Human invariant V alpha 24-J alpha Q TCR supports the development of CD1d-dependent NK1.1+ and NK1.1− T cells in transgenic mice. J Immunol. (2003) 170:2390–8. doi: 10.4049/jimmunol.170.5.2390
72. Lockridge JL, Chen X, Zhou Y, Rajesh D, Roenneburg DA, Hegde S, et al. Analysis of the CD1 antigen presenting system in humanized SCID mice. PLoS ONE. (2011) 6:e21701. doi: 10.1371/journal.pone.0021701
Keywords: iNKT cell, cancer immonotherapy, preclinical modeling, humanized mice, α-GalCer
Citation: Zhang Y, Springfield R, Chen S, Li X, Feng X, Moshirian R, Yang R and Yuan W (2019) α-GalCer and iNKT Cell-Based Cancer Immunotherapy: Realizing the Therapeutic Potentials. Front. Immunol. 10:1126. doi: 10.3389/fimmu.2019.01126
Received: 22 January 2019; Accepted: 03 May 2019;
Published: 06 June 2019.
Edited by:
Jin S. Im, University of Texas MD Anderson Cancer Center, United StatesReviewed by:
Moriya Tsuji, Aaron Diamond AIDS Research Center, United StatesCopyright © 2019 Zhang, Springfield, Chen, Li, Feng, Moshirian, Yang and Yuan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Weiming Yuan, d2VpbWluZy55dWFuQHVzYy5lZHU=
†These authors have contributed equally to this work
‡Present Address: Rirong Yang, Department of Immunology, School of Preclinical Medicine, Center for Genomic and Personalized Medicine, Guangxi Medical University, Guangxi, China
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.