
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 03 May 2019
Sec. Cancer Immunity and Immunotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fimmu.2019.00865
This article is part of the Research Topic The Role of Complement in Tumors View all 10 articles
 Alessandro Mangogna1
Alessandro Mangogna1 Chiara Agostinis2*
Chiara Agostinis2* Deborah Bonazza3
Deborah Bonazza3 Beatrice Belmonte4Paola Zacchi1Gabriella Zito2Andrea Romano3Fabrizio Zanconati3Giuseppe Ricci2,3
Beatrice Belmonte4Paola Zacchi1Gabriella Zito2Andrea Romano3Fabrizio Zanconati3Giuseppe Ricci2,3 Uday Kishore5
Uday Kishore5 Roberta Bulla1
Roberta Bulla1C1q is the first subcomponent of the classical pathway of the complement system and belongs to the C1q/Tumor Necrosis Factor superfamily. C1q can perform a diverse range of immune and non-immune functions in a complement-dependent as well as -independent manner. Being a pattern recognition molecule of the innate immunity, C1q can recognize a number of self, non-self and altered-self ligands and bring about effector mechanisms designed to clear pathogens via opsonisation and inflammatory response. C1q is locally synthesized by macrophages and dendritic cells, and thus, can get involved in a range of biological processes, such as angiogenesis and tissue remodeling, immune modulation, and immunologic tolerance. The notion of C1q involvement in the pathogenesis of cancer is still evolving. C1q appears to have a dual role in cancer: tumor promoting as well as tumor-protective, depending on the context of the disease. In the current study, we performed a bioinformatics analysis to investigate whether C1q can serve as a potential prognostic marker for human carcinoma. We used the Oncomine database and the survival analysis platforms Kaplan-Meier plotter. Our results showed that high levels of C1q have a favorable prognostic index in basal-like breast cancer for disease-free survival, and in HER2-positive breast cancer for overall survival, while it showed a pro-tumorigenic role of C1q in lung adenocarcinoma, and in clear cell renal cell carcinoma. This in silico study, if validated via a retrospective study, can be a step forward in establishing C1q as a new tool as a prognostic biomarker for various carcinoma.

Graphical Abstract. Summary of the conclusions of the study.
C1q is the first recognition subcomponent of the complement classical pathway, which when associated with C1r and C1s, forms a C1 complex, allowing the activation of the complement cascade (1). By virtue of its ability to bind to IgG and IgM containing immune complexes and activating the classical pathway, C1q acts as prototypical link between innate and adaptive immune wings of the immune system (2). C1q can bind to a range of non-self-target ligands (pathogens), altered self (β-amyloid peptide, prion protein, apoptotic and necrotic cells via phosphatidylserine and DNA, respectively), and cell surface receptors (such as calreticulin and gC1qR) (3). Several features of the C1q render it a versatile molecular sensor of damage-modified self or non-self antigens (4). C1q, unlike most of the complement proteins which are exclusively produced by hepatocytes, can also be synthesized in a local environment by a wide range of cell types including macrophages and dendritic cells (5). Local synthesis, therefore, offers an additional avenue to C1q in order to exert specific functions in situ that are strictly connected to its site of production without involving complement activation (6).
C1q is an hexametric glycoprotein of about 460 kDa, resembling a “bouquet of tulips” being composed by three polypeptide chains: A (28 kDa), B (25 kDa), C (24 kDa), which are the product of three distinct genes clustered in the same orientation, and in the order A–C–B, on a 24 kb stretch of DNA on chromosome 1p (7). Each chain consists of a C-terminal globular head (gC1q) domain and an N-terminal triple-helical collagen-like (cC1q) domain (8). C1q associates with the Ca2+-dependent C1r2-C1s2 tetramer, of about 360 kDa, to form the soluble pentameric C1 complex (9). The C-terminal ends of A, B and C chains assemble together to form a heterotrimeric gC1q domain, which by virtue of its modular organization, can work independently and engage with a diverse range of target ligands (3). While the gC1q domain latches on to the charge patterns on the ligands, the cC1q domain can interact with effector mechanism inducers, such as C1r, C1s, cell surface receptors, etc. Thus, a combination of a highly versatile and modular gC1q domain and a cell surface interacting cC1q domain, together with its local synthesis, makes C1q a potent orchestrator of molecular pathways. C1q is involved not only in innate and adaptive immune mechanisms, but also in a wide range of physiological and pathological processes, such as placental development (10, 11), pre-eclampsia (12, 13), wound healing (14) and cancer (15–18).
Markiewski et al. provided evidence that C1q is present in syngeneic mouse tumors. Indeed, they found that the activation of the classical pathway is the major contributor to complement-mediated tumor progression (19). Subsequently, we showed that locally expressed C1q had important effects in the tumor microenvironment (TME) (17). C1q expressed in the stroma and vascular endothelium of several human malignant tumors acted as a tumor-promoting factor by favoring adhesion, migration and proliferation of cancer cells as well as angiogenesis and metastasis. C1q-deficient (C1qa−/−) mice, bearing a syngeneic B16 melanoma, exhibited slower tumor growth and prolonged survival, compared to C3 or C5 deficient mice although it has been shown that C3/C5 deficiency may also create microenvironment suboptimal for tumor growth (20, 21). Recently, we demonstrated that C1q is abundantly present in malignant pleural mesothelioma (MPM), where it can combine with hyaluronic acid (HA), which is a principal component of the TME, and enhance the tumor growth by promoting cell adhesion and proliferation (18). However, other have shown a pro-apoptotic effect of C1q on prostate (15) and ovarian cancer cells in vitro (16). These rather two set of contradicting studies warranted a systematic analysis of the context of the disease and TME that can render C1q protective or pathogenic in cancer.
In the current study, we performed a bioinformatics analysis, using Oncomine database and the survival analysis platforms Kaplan-Meier plotter, in order to investigate whether C1q can serve as a potential prognostic marker for human carcinoma, i.e., tumors of epithelial origin. Our results showed that high levels of C1q have a favorable prognostic index in basal-like breast cancer (BLBC) and in HER-2 positive breast cancer. However, we found a pro-tumorigenic role of C1q in lung adenocarcinoma, and in clear cell renal cell carcinoma (CCRCC). This study is an important step forward in highlighting C1q as a new prognostic candidate biomarker for a range of carcinomas.
The expression levels of C1QA, C1QB, and C1QC genes in various carcinomas were analyzed using Oncomine (www.oncomine.org), a cancer microarray database and web-based data mining platform from genome-wide expression analyses (22, 23). We compared the differences in mRNA level between normal tissue and carcinoma. The mRNA expression levels in neoplastic tissues compared to the healthy tissues were obtained as the parameters of p-value < 0.05, fold change >2, and gene ranking in the top 10%. Information about the dataset used in this study is summarized in Supplementary Table 1.
A Kaplan-Meier plotter database can be used to assess the effect of 54,675 genes on survival using 10,461 carcinoma samples (5,143 breast, 1,816 ovarian, 2,437 lung, and 1,065 gastric cancer patients with a mean follow-up of 69/40/49/33 months) using probe sets on the HGU133 Plus 2.0 array from Gene Expression Omnibus (GEO). For other human carcinoma, a total of 3,439 patients with RNA HiSeq data from The Cancer Genome Atlas (TCGA) cohort were collected. The prognostic significance of C1QA, C1QB, and C1QC expression and survival in several carcinomas was analyzed by Kaplan-Meier plotter (www.kmplot.com/analysis/) (24). The hazard ratio with 95% confidence intervals and logrank p-value was also computed.
Normal and neoplastic human tissues, including breast, kidney and lung, were selected from the archives of the Department of Pathology, University of Trieste. Immunohistochemistry (IHC) was performed using a polymer detection method. Briefly, tissue samples were fixed in 10% v/v buffered formalin and then paraffin embedded. Four μm-thick tissue sections were deparaffinized and rehydrated. The antigen unmasking technique was carried out using Novocastra Epitope Retrieval Solutions, pH 9 (Leica Biosystems) in a PT Link pre-treatment module (Dako) at 98°C for 30 min. Sections were then brought to RT and washed in PBS. After neutralization of the endogenous peroxidase with 3% v/v H2O2 and Fc blocking by a specific protein block (Novocastra, Leica Biosystems), samples were incubated overnight at 4°C with rabbit polyclonal anti-human C1q (dilution 1:200) antibodies (Dako). Staining was carried out via polymer detection kit (Novocastra, Leica Biosystems) and DAB (3,3′-Diaminobenzidine; Dako, Denmark) substrate-chromogen. Slides were counterstained with Harris Haematoxylin (Novocastra, Leica Biosystems). Sections were analyzed under the Axio Scope A1 optical microscope (Zeiss) and microphotographs were collected through the Axiocam 503 color digital camera (Zeiss) using the Zen2 software.
Survival curves were generated by the Kaplan-Meier plotter. All results are displayed with p-values from a log-rank test. P-values < 0.05 were considered significant. Similarly, with Oncomine, the statistical significance of data (p-values) was provided by the program.
The expression of C1QA, C1QB, and C1QC genes was analyzed between different carcinoma and normal tissue counterparts using the Oncomine database. The threshold was determined as the following values: p-value < 0.05, fold change >2, and gene ranking in the top 10%. Carcinomas included in this analysis were: bladder carcinoma, breast cancer, cervical squamous cell carcinoma, esophageal carcinoma, head-neck squamous cell carcinoma, clear cell renal cell carcinoma (CCRCC), papillary renal cell carcinoma (PRCC), liver hepatocellular carcinoma, lung adenocarcinoma, lung squamous cell carcinoma, ovarian cancer, pancreatic ductal adenocarcinoma, rectum adenocarcinoma, gastric carcinoma, and uterine corpus endometrial carcinoma. We only investigated carcinomas in which all the three C1q chains showed a significant prognostic effect by Kaplan-Meier plotter analysis. The C1QA, C1QB, and C1QC genes were either over-expressed, or downregulated depending on the type of carcinoma investigated, as compared to their normal tissue counterparts. All the three C1q chains showed a differential prognostic significance. These data appear to suggest that C1q can have pro- or anti-tumorigenic implications, depending on the carcinoma types (Table 1). Thus, detailed analyses of the expression profiles of all three C1q chains were performed.
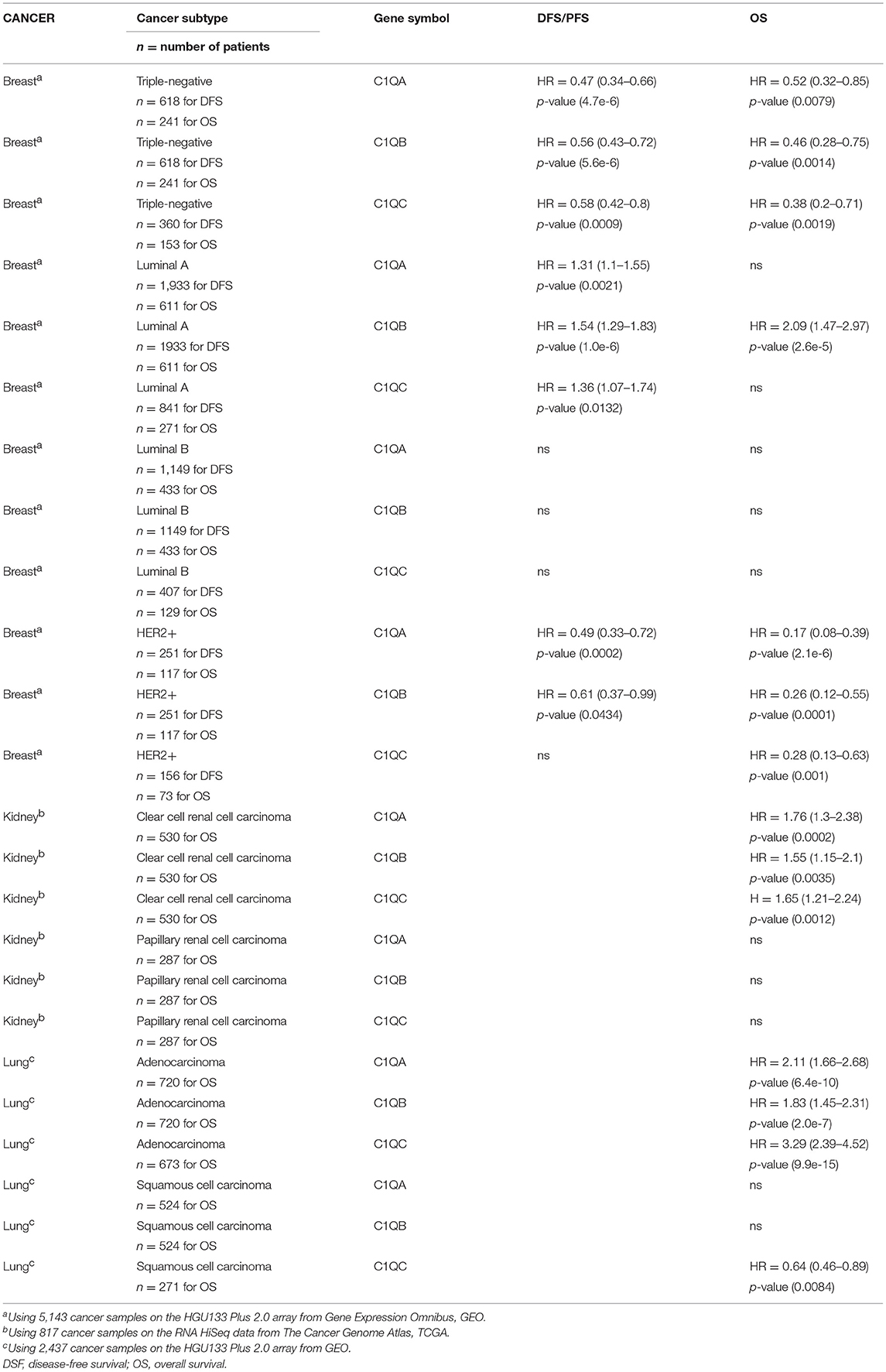
Table 1. Prognostic significance of C1q in patients with carcinomas.
Bioinformatics analysis of C1QA, C1QB, and C1QC mRNA expression was performed in the context of the breast cancer using Karnoub's, Finak's, Curtis's, and Perou's datasets. A higher expression level of the three chains of C1q was detected as compared to normal breast tissue (Figure 1A, p < 0.05). When breast cancer was stratified into different histological subtypes, C1QA, C1QB, and C1QC mRNA expression achieved a statistical significance only in medullary carcinoma (Figure 1B, p < 0.05). To evaluate the prognostic significance of C1q in all breast cancers, we considered their molecular classification, such as luminal-A, luminal-B, HER-2 positive, and basal-like cancers (BLBC) (Supplementary Table 2).
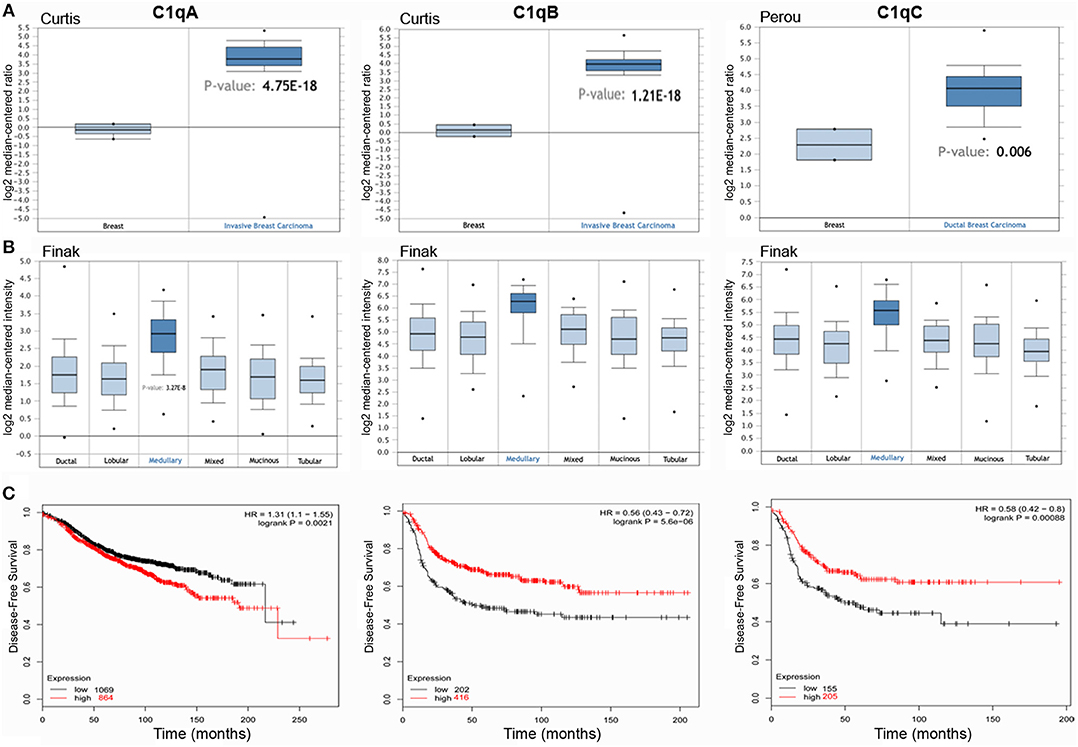
Figure 1. C1QA, C1QB, and C1QC expression in invasive breast carcinoma. Curtis's datasets were used for bioinformatics analysis to explore C1QA and C1QB mRNAs expression in the breast cancer, whereas Perou's datasets was used for bioinformatics analysis to evaluate C1QC mRNA. A higher C1q mRNAs expression was detectable in invasive breast carcinoma compared to normal breast tissue (A). The analysis of the different breast carcinoma histotypes by Finak's dataset revealed how the medullary breast cancer presented the major intensity of C1q mRNA expression (B). According to the data Kaplan-Meir plotter, C1QA, C1QB, and C1QC mRNA expressions was positively linked to a disease-free survival (DFS) rate in patients with basal-like cancers (C) (p<0.05) and to an overall survival (OS) rate with HER-2 positive cancers (Supplementary Table 2). HR, hazard ratio.
According to Kaplan-Meir plotter data, C1QA, C1QB, and C1QC mRNA expression was positively associated with a disease-free survival (DFS) rate in patients with BLBC (Figure 1C, p < 0.05) and with an overall survival (OS) rate with HER-2 positive cancers (Table 1). This correlation was not evident in luminal-A and luminal-B patients. Only C1QB mRNA expression was negatively associated with high DFS and OS rates in the breast cancer patients with luminal-A, and to a DFS rate with all breast cancer.
The IHC analysis within the BLBC microenvironment revealed that C1q was diffusely present in the tumor stroma and was expressed by macrophage-like cells, suggestive of tumor-infiltrating myeloid elements (Figure 2A).
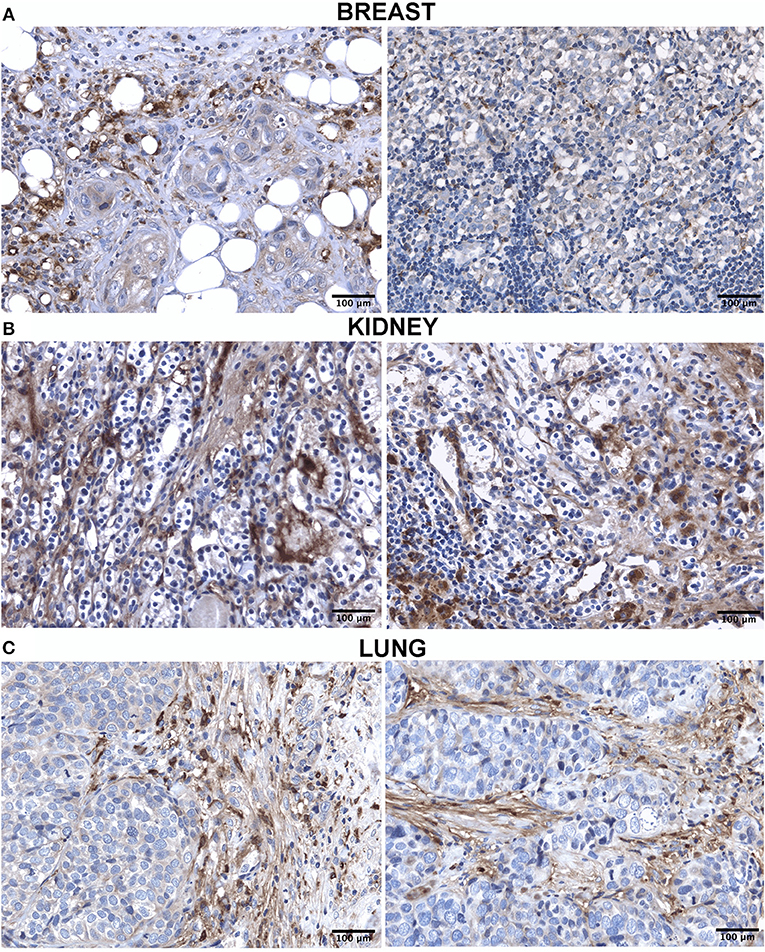
Figure 2. Immunohistochemistry analysis for C1q in breast (A), kidney (B) and lung carcinoma (C). Representative microphotographs showing expression of C1q in different carcinoma. The expression of C1q in carcinoma was observed in all tissues with differential distribution in the TME, as described in the result section. DAB (brown) chromogen was used to visualize the binding of anti-human C1q antibodies; scale bars, 50μm.
The C1QA, C1QB, and C1QC mRNA expression was evaluated in kidney cancer based on the results obtained from different datasets. In CCRCC, the expression of the three C1q chains was higher as compared to normal kidney (Figure 3A, p < 0.05). However, in the case of PRCC, this trend was evident only for C1QA and C1QB mRNA expression (data not shown). The data obtained from Kaplan-Meier plotter showed a negative relationship between C1QA, C1QB, and C1QC mRNA expression and OS rate of patients with CCRCC (Figure 3B, p < 0.05). No correlation was observed between C1QA, C1QB, and C1QC mRNA expression and OS in the PRCC patients (Table 1).
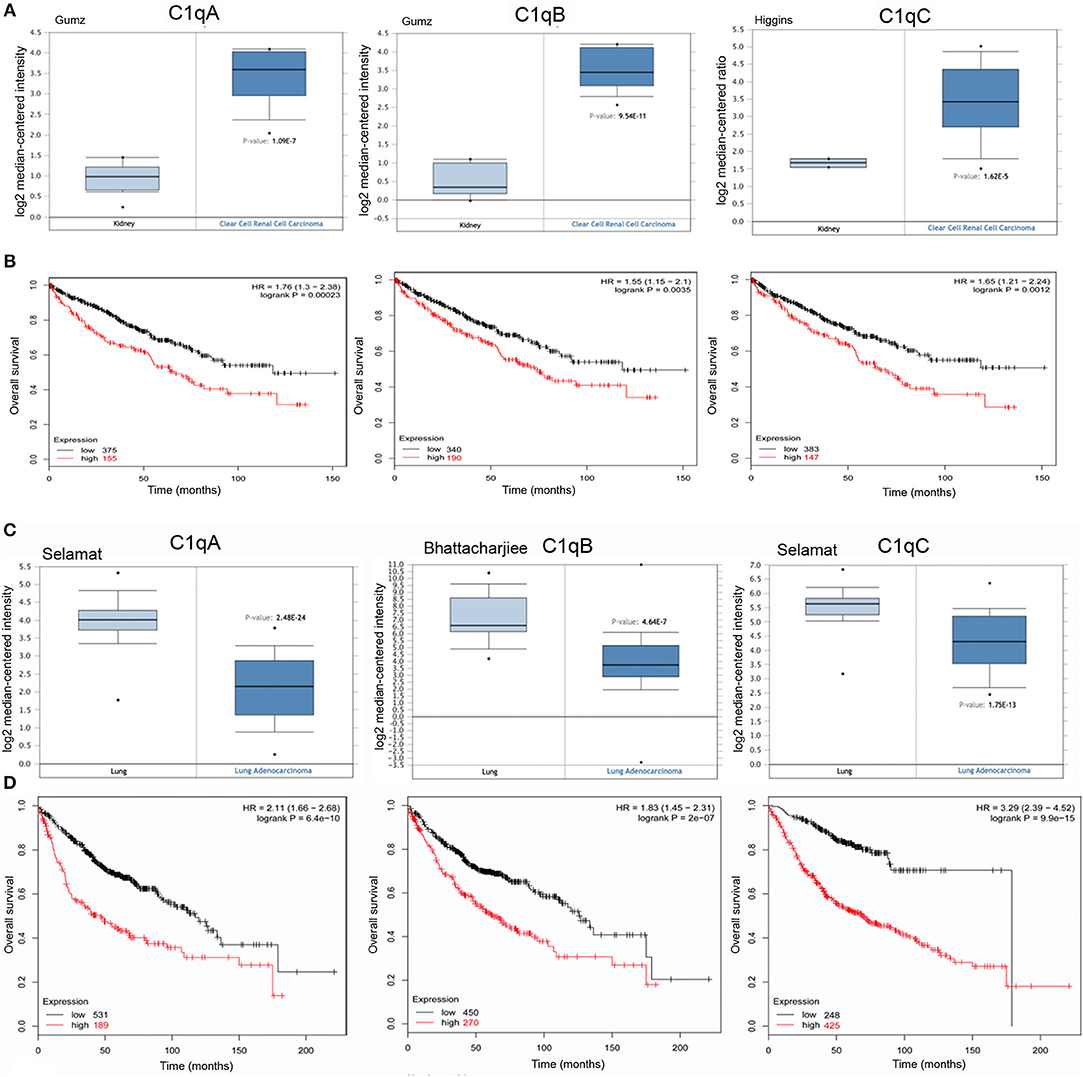
Figure 3. Pathological significance of C1q expression kidney and in lung cancer. Gumz's dataset have explored C1QA and C1QB mRNA expression in kidney, whereas Higgins' dataset were used for C1QC mRNA expression. A higher C1q expression was detectable in CCRCC cancer than that in normal tissue (A), p < 0.05. According to the data from Kaplan-Meier plotter, C1q mRNA expressions were negatively related to an overall survival rate of the patients with CCRCC (B). HR, hazard ratio. Selamat's dataset have revealed a lower C1QA and C1QC mRNA expression in lung adenocarcinoma that in normal lung tissues (C), whereas Bhattacharjiee's dataset was used for C1QB, but the results were in accordance with Selamat's one. There was a negative association between C1q mRNA expression and a favorable prognosis in patients with lung cancer, for Kaplan-Meir plotter (D). HR, hazard ratio.
Within the CCRCC microenvironment, C1q was found to be mainly expressed in the tumor stroma and in the small vessels, and it was associated with the cell membrane of tumor cells (Figure 2B).
While examing C1QA and C1QB mRNA expression in lung cancer, using Selamat's, Wachi's and Bhattacharjee's datasets, we found a lower expression level in adenocarcinoma (Figure 3C, p <0.05) and in squamous cell carcinoma (data not shown, p <0.05) than in normal lung tissue; C1QC mRNA expression was significant only in lung adenocarcinoma. As shown in Figure 3D, C1QA, C1QB, and C1QC mRNA expression levels negatively correlated with an OS rate of the patients with lung adenocarcinoma (p <0.05); no correlation with OS was observed in squamous cell carcinoma (Table 1).
IHC in lung adenocarcinoma revealed C1q staining in the stroma and some macrophage-like positive cells into the tumor mass (Figure 2C).
In this paper, we performed bioinformatics analysis to explore if C1q level could act as a possible prognostic marker in various carcinomas, in view of its reported dichotomous effects on cancer cells (pro- and anti-tumorigenic). C1q is present in colon, lung, breast, pancreatic carcinoma, and melanoma. C1q can promote adhesion, proliferation and migration of melanoma cells (17). We found C1q in abundance in all histological variants (epithelioid, sarcomatoid, and biphasic) of asbestos-induced malignant pleural mesothelioma. C1q bound high and low molecular weight HA and acted as a tumor-promoting factor (18). In addition, C1q exerted a protective effect against apoptosis, suggesting an overall pro-tumorigenic activity (17). However, Hong et al. recently observed that C1q, expressed in normal prostate, was downregulated in benign prostatic hyperplasia and prostate cancer (15). C1q was able to induce apoptosis and growth suppression of human prostate DU145 cells, through direct activation of the tumor suppressor WW-domain containing oxidoreductase (WOX1). C1q also have a pro-apoptotic effect on an ovarian cell line, SKOV3, acting via a TNF-α induced apoptosis pathway that involves upregulation of Bax and Fas (16).
In a syngeneic murine model of melanoma in C57BL/6 strain, C1q-deficient mice showed prolonged survival and slower tumor growth, as compared to wild-type mice (17). However, Bandini et al. found that neuT mice, a genetically engineered mouse model for mammary carcinoma that was made deficient for the C1qA chain (neuT-C1KO mice), manifested an accelerated tumor growth associated with an increased number of intra-tumoral vessels, compared to wild-type neuT mice. These differences in tumor progression were attributed to a reduced activation of WW domain containing oxidoreductase (WWOX) in C1q–deficient mice (25).
In view of these rather contradicting roles of C1q in tumor progression, we performed a systematic bioinformatics analysis of the expression of C1q, and its correlation with the survival rate in different carcinoma histotypes, using Oncomine and Kaplan-Meier plotter tools. We selected the carcinomas that showed all the three chains of human C1q statistically significant for the prognosis; in several cases, the prognosis was differentially linked to the C1q chains, or limited to one or two C1q chains. We often noticed the mRNA encoding for only one or two C1q chains, something that would impede synthesis of a functional C1q molecule. Indeed, we have provided evidence in the past that the expression of C1qC chain is essential for the production of functional C1q by the endothelial cells of the decidua (26). Moreover, mesothelioma cells are impaired in C1q A chain synthesis (18).
Our bioinformatics analysis highlighted that high levels of C1q have a favorable prognostic index in BLBCs for DFS and HER2+ breast cancer for OS, (Graphical Abstract) consistent with the in vivo studies by Bandini et al. using C1q-deficient mice (25). Inflammation is a major characteristic of these types of tumors. One possible explanation for the observed positive association between C1q expression and favorable prognostic index could be due to the correlation between the presence of C1q and dendritic cells (CD11c positive cells) in TME. High CD11c expression in BLBCs is associated with a significantly higher OS (p = 0.047) as compared to low CD11c expression (27). Dendritic cells themselves can be a potential source of C1q within the TME (28, 29). C1q, although present, is not able to bind BLBC cells (MDA-MB-231), and hence, not able to promote tumor progression (unpublished data), probably due to downregulation of putative C1q receptor(s). It is thus crucial to understand the differences in good prognosis survival between BLBCs and HER2+ breast cancer, the role of inflammation, and that of C1q in determining such differences.
Wilson et al. (30) found that C1q chain genes were enriched in the stroma compartment of triple-negative breast cancers. The analysis of publicly available data sets revealed that the genes encoding for the C1q chains were associated with a poor prognosis in BLBC using the TCGA dataset (504 patients). In our analysis, using the GEO dataset that include 5,143 patients, we observed a positive prognostic effect for BLBCs in DFS and HER2-positive breast cancers in OS. The opposite results were obtained for CCRCCs and lung adenocarcinomas in OS.
A negative prognostic effect arose from the analysis of kidney and lung carcinomas (Graphical Abstract). The most frequent histological subtypes include CCRCC and PRCC (CCRCC ~75%; PRCC ~10%) (31). The expression of C1q in kidney cancer is increased as compared to normal kidney tissue (Figure 3A) and C1q has a negative prognostic effect in the case of CCRCC (Figure 3B); no association was evident for PRCC. CCRCC tumor is characterized by an increased response to HIF that promotes blood vessel growth. Targeted therapies directed against VEGF, VEGF receptor, and mTOR play a crucial role in the management of metastatic CCRCC (32). We can hypothesize that C1q can also participate in promoting angiogenic processes in this particular tumor (14).
C1q has a negative prognostic value in lung tumors limited to adenocarcinomas, the most common form of lung cancer (Figure 3D). According to the WHO classification of lung tumors, there are four major histological types: adenocarcinomas, squamous cell carcinomas, large cell carcinomas, and small cell carcinomas (33). It is worth noting that C1q expression is reduced in lung cancer compared to the normal lung as we observed for surfactant protein D (SP-D) (34). Although C1q expression in lung cancer is lower than in normal tissue, lung cancer cells bind C1q present in the tumor microenvironment and activate the classical complement pathway (35). Tumor transformation is also concomitant with the loss of key defense molecules entrusted with early recognition and removal of the altered self (36).
A number of factors can modulate the role of C1q in the TME. C1q interaction with the ECM components can adversely interrupt its putative functions, as is the case with HA. It is also possible that certain tumors downregulate the putative receptor for C1q in order to escape possible apoptosis induction. Proliferative and apoptotic responses to C1q can be dictated by distinct receptors that are yet to be discovered. Last but not the least, the orientation of the C1q molecule, while engaging with the tumor cells, can also define the C1q-mediated implications. Our study encompasses all the above-mentioned possibilities, including tumor heterogeneity.
This study was carried out as per the recommendations of governmental guidelines, and approved by the CEUR (Comitato Etico Unico Regionale, FVG, Italy; number 34/2016). All subjects gave written informed consent in accordance with the Declaration of Helsinki.
AM and RB: conception and design. CA, DB, and AR: development of methodology. BB, DB, CA, and GZ: acquisition of data. AM, DB, BB, FZ and CA: analysis and interpretation of data (e.g., statistical analysis, biostatistics, and computational analysis). RB, UK, CA, PZ, and GR: writing, review, and/or revision of the manuscript. RB: study supervision.
This work was supported by grants from the Institute for Maternal and Child Health, IRCCS Burlo Garofolo, Trieste, Italy (RC 20/16, RC 23/18), AIRC to Claudio Tripodo. Fondazione Cassa di Risparmio Trieste to RB.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Andrea Balduit, Fleur Bossi (IRCCS, Burlo Garofolo, Trieste, Italy) and Alessandro Gulino (Department of Human Pathology, University of Palermo) for the immunohistochemical analysis. The contribution of Nicolò Morosini for the C1 visualization in Graphical Abstract is acknowledged.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.00865/full#supplementary-material
TME, Tumor microenvironment; ECM, extracellular matrix; BLBC, basal-like breast cancer; CCRCC, clear cell renal cell carcinoma; PRCC, papillary renal cell carcinoma; OS, overall survival; DFS, disease-free survival; WOX1, WW-domain containing oxidoreductase.
1. Lu J, Kishore U. C1 complex: an adaptable proteolytic module for complement and non-complement functions. Front Immunol. (2017) 8:592. doi: 10.3389/fimmu.2017.00592
2. Kishore U, Gaboriaud C, Waters P, Shrive AK, Greenhough TJ, Reid KB, et al. C1q and tumor necrosis factor superfamily: modularity and versatility. Trends Immunol. (2004) 25:551–61. doi: 10.1016/j.it.2004.08.006
3. Kishore U, Ghai R, Greenhough TJ, Shrive AK, Bonifati DM, Gadjeva MG, et al. Structural and functional anatomy of the globular domain of complement protein C1q. Immunol Lett. (2004) 95:113–28. doi: 10.1016/j.imlet.2004.06.015
4. Thielens NM, Tedesco F, Bohlson SS, Gaboriaud C, Tenner AJ. C1q: A fresh look upon an old molecule. Mol Immunol. (2017) 89:73–83. doi: 10.1016/j.molimm.2017.05.025
5. Petry F, Botto M, Holtappels R, Walport MJ, Loos M. Reconstitution of the complement function in C1q-deficient (C1qa−/−) mice with wild-type bone marrow cells. J Immunol. (2001) 167:4033–7. doi: 10.4049/jimmunol.167.7.4033
6. Nayak A, Pednekar L, Reid KB, Kishore U. Complement and non-complement activating functions of C1q: a prototypical innate immune molecule. Innate Immun. (2012) 18:350–63. doi: 10.1177/1753425910396252
7. Sellar GC, Cockburn D, Reid KB. Localization of the gene cluster encoding the A, B, and C chains of human C1q to 1p34.1-1p36.3. Immunogenetics. (1992) 35:214–6.
8. Ghebrehiwet B, Kandov E, Kishore U, Peerschke EIB. Is the A-chain the engine that drives the diversity of C1q functions? revisiting its unique structure. Front Immunol. (2018) 9:162. doi: 10.3389/fimmu.2018.00162
9. Reid KBM. Complement component C1q: historical perspective of a functionally versatile, and structurally unusual, serum protein. Front Immunol. (2018) 9:764. doi: 10.3389/fimmu.2018.00764
10. Agostinis C, Bulla R, Tripodo C, Gismondi A, Stabile H, Bossi F, et al. An alternative role of C1q in cell migration and tissue remodeling: contribution to trophoblast invasion and placental development. J Immunol. (2010) 185:4420–9. doi: 10.4049/jimmunol.0903215
11. Agostinis C, Tedesco F, Bulla R. Alternative functions of the complement protein C1q at embryo implantation site. J Reprod Immunol. (2017) 119:74–80. doi: 10.1016/j.jri.2016.09.001
12. Singh J, Ahmed A, Girardi G. Role of complement component C1q in the onset of preeclampsia in mice. Hypertension. (2011) 58:716–24. doi: 10.1161/HYPERTENSIONAHA.111.175919
13. Agostinis C, Stampalija T, Tannetta D, Loganes C, Vecchi Brumatti L, De Seta F, et al. Complement component C1q as potential diagnostic but not predictive marker of preeclampsia. Am J Reprod Immunol. (2016) 76:475–81. doi: 10.1111/aji.12586
14. Bossi F, Tripodo C, Rizzi L, Bulla R, Agostinis C, Guarnotta C, et al. C1q as a unique player in angiogenesis with therapeutic implication in wound healing. Proc Natl Acad Sci USA. (2014) 111:4209–14. doi: 10.1073/pnas.1311968111
15. Hong Q, Sze CI, Lin SR, Lee MH, He RY, Schultz L, et al. Complement C1q activates tumor suppressor WWOX to induce apoptosis in prostate cancer cells. PLoS ONE. (2009) 4:e5755. doi: 10.1371/journal.pone.0005755
16. Kaur A, Sultan SH, Murugaiah V, Pathan AA, Alhamlan FS, Karteris E, et al. Human C1q induces apoptosis in an ovarian cancer cell line via tumor necrosis factor pathway. Front Immunol. (2016) 7:599. doi: 10.3389/fimmu.2016.00599
17. Bulla R, Tripodo C, Rami D, Ling GS, Agostinis C, Guarnotta C, et al. C1q acts in the tumour microenvironment as a cancer-promoting factor independently of complement activation. Nat Commun. (2016) 7:10346. doi: 10.1038/ncomms10346
18. Agostinis C, Vidergar R, Belmonte B, Mangogna A, Amadio L, Geri P, et al. Complement protein C1q binds to hyaluronic acid in the malignant pleural mesothelioma microenvironment and promotes tumor growth. Front Immunol. (2017) 8:1559. doi: 10.3389/fimmu.2017.01559
19. Markiewski MM, DeAngelis RA, Benencia F, Ricklin-Lichtsteiner SK, Koutoulaki A, Gerard C, et al. Modulation of the antitumor immune response by complement. Nat Immunol. (2008) 9:1225–35. doi: 10.1038/ni.1655
20. Corrales L, Ajona D, Rafail S, Lasarte JJ, Riezu-Boj JI, Lambris JD, et al. Anaphylatoxin C5a creates a favorable microenvironment for lung cancer progression. J Immunol. (2012) 189:4674–83. doi: 10.4049/jimmunol.1201654
21. Riihila P, Nissinen L, Farshchian M, Kallajoki M, Kivisaari A, Meri S, et al. Complement component C3 and complement factor B promote growth of cutaneous squamous cell carcinoma. Am J Pathol. (2017) 187:1186–97. doi: 10.1016/j.ajpath.2017.01.006
22. Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia. (2004) 6:1–6. doi: 10.1016/S1476-5586(04)80047-2
23. Rhodes DR, Kalyana-Sundaram S, Mahavisno V, Varambally R, Yu J, Briggs BB, et al. Oncomine 3.0: genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia. (2007) 9:166–80. doi: 10.1593/neo.07112
24. Lanczky A, Nagy A, Bottai G, Munkacsy G, Szabo A, Santarpia L, et al. miRpower: a web-tool to validate survival-associated miRNAs utilizing expression data from 2178 breast cancer patients. Breast Cancer Res Treat. (2016) 160:439–46. doi: 10.1007/s10549-016-4013-7
25. Bandini S, Macagno M, Hysi A, Lanzardo S, Conti L, Bello A, et al. The non-inflammatory role of C1q during Her2/neu-driven mammary carcinogenesis. Oncoimmunology. (2016) 5:e1253653. doi: 10.1080/2162402X.2016.1253653
26. Bulla R, Agostinis C, Bossi F, Rizzi L, Debeus A, Tripodo C, et al. Decidual endothelial cells express surface-bound C1q as a molecular bridge between endovascular trophoblast and decidual endothelium. Mol Immunol. (2008) 45:2629–40. doi: 10.1016/j.molimm.2007.12.025
27. Lee H, Lee HJ, Song IH, Bang WS, Heo SH, Gong G, et al. CD11c-positive dendritic cells in triple-negative breast cancer. in vivo. (2018) 32:1561–9. doi: 10.21873/invivo.11415
28. Castellano G, Trouw LA, Fiore N, Daha MR, Schena FP, van Kooten C. Infiltrating dendritic cells contribute to local synthesis of C1q in murine and human lupus nephritis. Mol Immunol. (2010) 47:2129–37. doi: 10.1016/j.molimm.2010.02.006
29. Castellano G, Woltman AM, Nauta AJ, Roos A, Trouw LA, Seelen MA, et al. Maturation of dendritic cells abrogates C1q production in vivo and in vitro. Blood. (2004) 103:3813–20. doi: 10.1182/blood-2003-09-3046
30. Winslow S, Leandersson K, Edsjo A, Larsson C. Prognostic stromal gene signatures in breast cancer. Breast Cancer Res. (2015) 17:23. doi: 10.1186/s13058-015-0530-2
31. Lopez-Beltran A, Carrasco JC, Cheng L, Scarpelli M, Kirkali Z, Montironi R. 2009 update on the classification of renal epithelial tumors in adults. Int J Urol. (2009) 16:432–43. doi: 10.1111/j.1442-2042.2009.02302.x
32. Su D, Singer EA, Srinivasan R. Molecular pathways in renal cell carcinoma: recent advances in genetics and molecular biology. Curr Opin Oncol. (2015) 27:217–23. doi: 10.1097/CCO.0000000000000186
33. Travis WD, Brambilla E, Burke AP, Marx A, Nicholson AG. Introduction to The 2015 World Health Organization Classification of Tumors of the Lung, Pleura, Thymus, and Heart. J Thorac Oncol. (2015) 10:1240–2. doi: 10.1097/JTO.0000000000000663
34. Mangogna A, Belmonte B, Agostinis C, Ricci G, Gulino A, Ferrara I, et al. Pathological significance and prognostic value of surfactant protein D in cancer. Front Immunol. (2018) 9:1748. doi: 10.3389/fimmu.2018.01748
35. Ajona D, Pajares MJ, Corrales L, Perez-Gracia JL, Agorreta J, Lozano MD, et al. Investigation of complement activation product c4d as a diagnostic and prognostic biomarker for lung cancer. J Natl Cancer Inst. (2013) 105:1385–93. doi: 10.1093/jnci/djt205
36. Vandivier RW, Ogden CA, Fadok VA, Hoffmann PR, Brown KK, Botto M, et al. Role of surfactant proteins A, D, and C1q in the clearance of apoptotic cells in vivo and in vitro: calreticulin and CD91 as a common collectin receptor complex. J Immunol. (2002) 169:3978–86. doi: 10.4049/jimmunol.169.7.3978
Keywords: complement, classical pathway, C1q, tumor, microenvironment, prognosis
Citation: Mangogna A, Agostinis C, Bonazza D, Belmonte B, Zacchi P, Zito G, Romano A, Zanconati F, Ricci G, Kishore U and Bulla R (2019) Is the Complement Protein C1q a Pro- or Anti-tumorigenic Factor? Bioinformatics Analysis Involving Human Carcinomas. Front. Immunol. 10:865. doi: 10.3389/fimmu.2019.00865
Received: 10 December 2018; Accepted: 04 April 2019;
Published: 03 May 2019.
Edited by:
Ruben Pio, University of Navarra, SpainCopyright © 2019 Mangogna, Agostinis, Bonazza, Belmonte, Zacchi, Zito, Romano, Zanconati, Ricci, Kishore and Bulla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chiara Agostinis, Y2Fnb3N0aW5pc0B1bml0cy5pdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.