 Léa Perra1
Léa Perra1 Viviane Balloy1Tobias Foussignière1Didier Moissenet2Hortense Petat1Imran N. Mungrue3
Viviane Balloy1Tobias Foussignière1Didier Moissenet2Hortense Petat1Imran N. Mungrue3 Lhousseine Touqui4
Lhousseine Touqui4 Harriet Corvol1,5
Harriet Corvol1,5 Michel Chignard1
Michel Chignard1 Loic Guillot1*
Loic Guillot1*- 1Sorbonne Université, Inserm, Centre de recherche Saint-Antoine (CRSA), Paris, France
- 2Department of Bacteriology, APHP, Hôpital St-Antoine, Paris, France
- 3Department of Pharmacology and Experimental Therapeutics, Louisiana State University Health Sciences Center, New Orleans, LA, United States
- 4Equipe mixte Institut Pasteur/Paris V “Mucoviscidose et Bronchopathies Chroniques” Institut Pasteur, Paris, France
- 5Pneumologie Pédiatrique, APHP, Hôpital Trousseau, Paris, France
In cystic fibrosis (CF), Pseudomonas aeruginosa (Pa) colonizes the lungs, leading to chronic inflammation of the bronchial epithelium. ChaC glutathione-specific γ-glutamylcyclotransferase 1 (CHAC1) mRNA is differentially expressed in primary human airway epithelial cells from bronchi (hAECBs) from patients with CF and healthy patients at baseline and upon infection with Pa. CHAC1 degrades glutathione and is associated with ER stress and apoptosis pathways. In this study, we examined the roles of CHAC1 in the inflammatory response and apoptosis in lung epithelial cells. First, we confirmed by reverse transcription quantitative polymerase chain reaction that CHAC1 mRNA was overexpressed in hAECBs from patients without CF compared with the expression in hAECBs from patients with CF upon Pa (PAK strain) infection. Moreover, the Pa virulence factors LPS and flagellin were shown to induce CHAC1 expression in cells from patients without CF. Using NCI-H292 lung epithelial cells, we found that LPS-induced CHAC1 mRNA expression was PERK-independent and involved ATF4. Additionally, using CHAC1 small interfering RNA, we showed that reduced CHAC1 expression in the context of LPS and flagellin stimulation was associated with modulation of inflammatory markers and alteration of NF-κB signaling. Finally, we showed that Pa was not able to induce apoptosis in NCI-H292 cells. Our results suggest that CHAC1 is involved in the regulation of inflammation in bronchial cells during Pa infection and may explain the excessive inflammation present in the respiratory tracts of patients with CF.
Introduction
Cystic fibrosis (CF), the most common severe autosomal recessive genetic disease in the Caucasian population, is caused by mutations in the CF transmembrane conductance regulator (CFTR) gene (1–3). Lung injury is predominant and is the leading cause of morbidity and mortality in CF patients. In airways, CFTR dysfunction leads to alterations in mucociliary clearance and impairment of innate immune host defenses. This results in chronic bacterial infection and inflammation, leading to degradation of the lung epithelium and progressive lung damage in CF patients (4). Pseudomonas aeruginosa (Pa) is the leading cause of chronic pulmonary infection in the airways of adults with CF (5). Pa colonization induces a strong inflammatory response in CF patients, particularly neutrophil accumulation and hypersecretion of inflammatory cytokines, such as IL-8 (6, 7).
The lung epithelium plays an essential role in the fight against infections by orchestrating innate immunity (8). Lung epithelial cells are sufficient to generate an innate protective immune response in mice infected with Pa (9). In CF, establishment of chronic pulmonary infection with Pa is linked to alterations in epithelial cell responses, which do not allow establishment of an adequate defense system (4). Previously, we showed that this response was indeed different by comparative analysis of the transcriptomic signatures of primary CF and non-CF bronchial epithelial cells during Pa infection (10). We identified several up-and downregulated mRNAs in CF cells compared with those in non-CF cells, suggesting that these genes may be involved in the exacerbated inflammatory response to Pa found in CF patients. Among these dysregulated mRNAs, ChaC glutathione-specific γ-glutamylcyclotransferase 1 (CHAC1) showed dramatically differential expression between non-CF and CF cells both at baseline and after Pa infection.
CHAC1 was identified in mammalian cells for the first time in 2009 as a new component of the unfolded protein response (UPR) pathway (11). This UPR pathway is induced in response to ER stress due to the presence of misfolded proteins in the ER lumen. The UPR consists of three pathways initiated by inositol-requiring enzyme 1α, eukaryotic translation initiation factor 2-α kinase 3 (PERK), and activation transcription factor (ATF) 6 (12). These three pathways alleviate ER stress by decreasing protein synthesis, facilitating protein folding, and increasing protein degradation. CHAC1 is induced by the PERK pathway downstream of ATF4 and the pro-apoptotic factor DNA damage-inducible transcript 3 (DDIT3 also called CHOP) and has been described as necessary and sufficient to induce markers of apoptosis, particularly the cleavage of the apoptotic factor poly(ADP-ribose) polymerase 1 (PARP) (11). Moreover, CHAC1 can degrade glutathione in the cytosol of mammalian cells through its γ-glutamylcyclotransferase activity (13). Various studies have shown a link between ER stress and inflammation (12, 14, 15). In the context of CF, atypical activation of the UPR pathway has been observed; the resulting ER stress can potentiate the inflammatory response (12). Moreover, the UPR is implicated in the regulation of cytokine production for bacterial infections (16). The role of CHAC1 in this interplay between ER stress and inflammation in the context of Pa infection has never been investigated.
Accordingly, in this study, we hypothesized that CHAC1 may play a role in regulating the inflammatory process induced by Pa infection and apoptosis.
Materials and Methods
Reagents
Pa (serotype 10) LPS (1 μg/ml), MG-132, staurosporine, tobramycine, and DMSO were from Sigma-Aldrich (Saint-Quentin Fallavier, France). Purified Pa flagellin (50 ng/ml) was from InvivoGen (San Diego, CA, USA), and Tunicamycine (TM) and GSK2656157/PERK inhibitor were from Calbiochem (Merck Millipore, Molsheim, France).
Cell Culture
NCI-H292 (ATCC-CRL-1848; Rockville, MD, USA), BEAS2-B (ATCC-CRL-9609) and A549-NF-κB luciferase cells (Panomics, Fremont, CA, USA) were grown as previously (17, 18). Primary hAECBs from CF patients or healthy donors (Table 1) were commercially obtained (Epithelix, Plan les Ouates, Switzerland) and were cultured as previously (19). The cells were seeded in different plates (TPP, Techno Plastic Products, Trasadingen, Switzerland) and after reaching confluence and were incubated overnight in DMEM containing 10% FBS and 1% antibiotics before being stimulated in the same medium free of antibiotics.
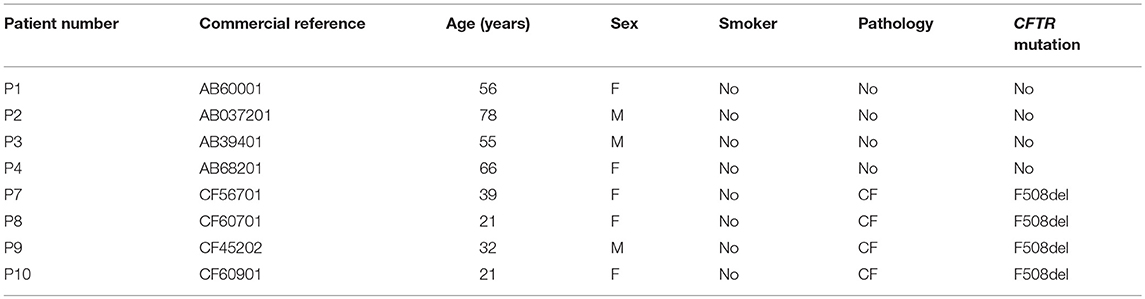
Table 1. Characteristics of patients whose hAECBs were used in this study.
Bacterial Strains, Growth Conditions, and Infection Protocols
The Pa WT PAK strain, the mutant ΔFliC with deletion of flagellin and the double mutant ΔpscFΔxcpQ with deletion of type II and III secretion systems were used and grown as previously (20, 21). The OD measured at 600 nm was adjusted to give the desired MOI. MOI were verified by serial 10-fold dilutions of the bacterial suspensions and plating on LB agar (Supplementary Figure 3). Cells seeded in 6-well (hAECBs) or 12-well plates (NCI-H292) were infected with Pa in their culture medium without antibiotics. After each stimulation, supernatants were collected and centrifuged at 3,000 × g for 15 min to remove the bacteria. Clinical strains of Pa were isolated according to the standards procedures of the microbiological department of St-Antoine Hospital, Paris, France. Sputum from patients was diluted (v/v) with prediluted Digest-Eur (Eurobio, Courtaboeuf, France), vortexed, and incubated for 15 min at room temperature. A 10-μL aliquot was plated on different selective media for 5 days at 37°C under aerobic conditions. Pa was isolated from Drigalski's agar plates, and identification was confirmed by mass spectrometry (Maldi-Biotyper, Brucker, France). Stenotrophomonas maltophilia was freshly isolated from a CF patient, and its identification was confirmed by mass spectrometry. S. maltophilia and Staphyloccocus aureus [Newman strain (22)] were cultured using the same protocol for Pa as described above.
CHAC1 and ATF4 Inhibition
To induce transient CHAC1 mRNA inhibition, we performed reverse transfection in NCI-H292 and/or A549 cells with either siRNA for CHAC1 (siRNA CHAC1, s35570) or negative control (siRNA Ctrl) from Ambion (Austin, TX, USA) using HiPerFect transfection reagent (Qiagen, Hilden, Germany). Cells were, respectively, seeded either in 96- (A549) or 12-well (NCI-H292) plates and transfected the same day with 5 nM siRNA. After 48 h of reverse transfection, cells were treated as described in the Figure Legends. To induce transient ATF4 mRNA inhibition, NCI-H292 cells seeded in 12-well plates were transfected with 5 nM siRNA for ATF4 (s1702, Sigma-Aldrich) or 5 nM negative control siRNA (siRNA Ctrl,) using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) for 48 h.
RT qPCR
RNA was isolated using a NucleoSpin RNA/Protein kit (Macherey Nagel, Duren, Germany). RT was performed using a high-capacity cDNA kit (Applied Biosystems, Foster City, CA, USA). Real-time qPCR was performed with an ABI StepOnePlus, using TaqMan Fast Universal PCR Master Mix (Applied Biosystems), TaqMan probes for CHAC1 (Hs_00225520), IL-8 (Hs_174103), IL-6 (Hs_985639), CCL2 (Hs_00234140), GAPDH (Hs_2786624) and cDNA as a template. For relative quantification, the amount of target genes was normalized to the expression of GAPDH relative to control cells used as a calibrator and was calculated using the 2−ΔΔCt method.
Western Blotting
Total proteins were extracted using a NucleoSpin RNA/Protein kit (Macherey Nagel). An equal amount of proteins was reduced, size-separated on 10 or 13.5% SDS-polyacrylamide gels, and transferred to nitrocellulose membranes (Invitrogen). The membranes were blocked in 5% milk in TBS-Tween 0.1% and incubated with specific primary antibodies against CHAC1 (AV42623; Sigma-Aldrich), PARP (9542; Cell Signaling Technology (CST), Danvers, MA, USA), ATF4 (11815; CST), cleaved caspase-3 (9661; CST), phospho- and total NF-κB-p65 (S536 and D14E12; CST), and β-actin (A2228; Sigma-Aldrich). For anti-CHAC1 antibody validation, CHAC1 overexpression was performed with a CHAC1-V5eGFP plasmid (CHAC1-GFP), as previously shown (11). The CHAC1-GFP plasmid was transfected into NCI-H292 cells using Lipofectamine 3000 transfection reagent for 24 h. Secondary antibodies were purchased from CST. Bound antibodies were detected using SuperSignal West Femto Maximum Sensitivity chemiluminescent substrate (Thermo Fisher Scientific, Rockford, IL, USA). Between successive primary antibodies, membranes were treated with Restore PLUS Western Blot Stripping Buffer (Thermo Fisher Scientific). Images were recorded with a Fujifilm LAS-3000 bioimaging system (Fujifilm, Stamford, CT, USA). Quantification was performed with ImageJ 10.2 (https://imagej.nih.gov/ij/index.html).
ELISA
Concentrations of human IL-8, IL-6, CCL2/MCP-1 (R&D, Minneapolis, MN, USA) and PGE2 (Cayman, Ann Harbor, MI, USA) were measured in cell supernatants using ELISA kits according to the manufacturer's instructions. The 3,3′,5,5′-tetramethylbenzidine substrate was from CST.
Fluorescence Microscopy
CHAC1-GFP expression was monitored in live NCI-H292 cells using an EVOS cell imaging system (Thermo Fisher Scientific).
NF-κB Luciferase Activity
NF-κB luciferase activity in A549 cells was assayed with a Luciferase kit (Promega, Madison, WI, USA) according to the manufacturer's protocol with a FLUOstar OPTIMA luminometer (BMG LABTECH, Champigny sur Marne, France).
Statistical Analysis
Differences among groups were assessed for statistical significance using Prism 7.00 software (GraphPad Software, La Jolla, CA, USA) as indicated in the Figure Legends. Differences with p < 0.05 were considered statistically significant.
Results
Defective Regulation of CHAC1 in CF Cells
In a previous transcriptomic analysis, we identified CHAC1 as one of the differentially expressed mRNAs between human airway epithelial cells from bronchi (hAECBs) of patients with and without CF during Pa infection (10). Indeed, CHAC1 mRNA was only induced in hAECBs from non-CF patients (Figure 1A, Supplementary Table 1). Because CHAC1 belongs to the UPR pathway of ER stress initiated by PERK (11), we analyzed the expression of genes shown to be involved in CHAC1-associated ER stress responses in our previous study. We observed that ATF4, PCK2, and HERPUD1 were significantly downregulated in hAECBs from CF patients vs. in hAECBs from non-CF patients (Supplementary Figures 1A–C and Supplementary Table 1). In contrast, ATF3 and TNFRSF6B were upregulated (Supplementary Figures 1D,E and Supplementary Table 1) in hAECBs from CF patients during the course of infection by Pa. These results indicated an altered expression of mRNAs involved in the CHAC1-associated ER stress response (Supplementary Figure 1F) in CF cells.
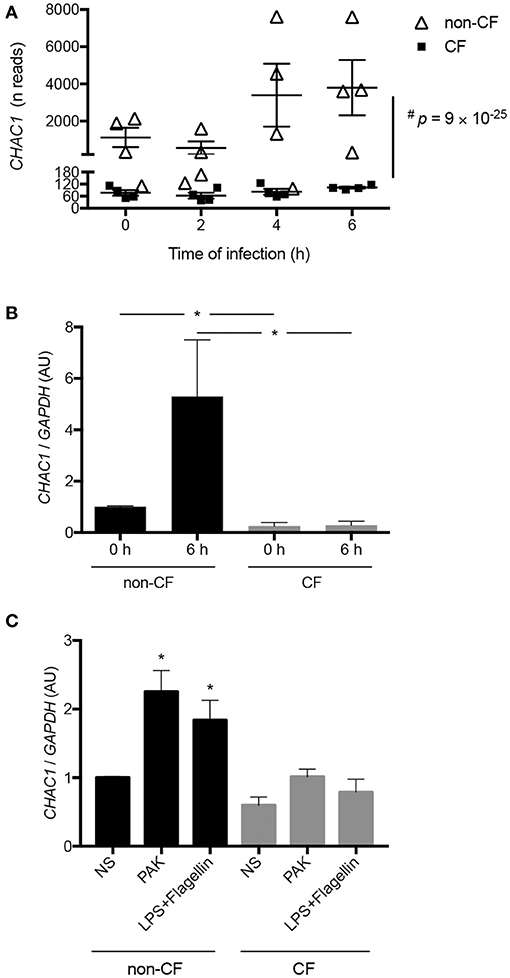
Figure 1. (A) Kinetics of CHAC1 expression measured by RNAseq in hAECBs from patients without CF (N = 4, open triangles) and with CF (N = 4, black squares) infected with Pa (PAK strain, MOI 0.25) for 0, 2, 4, or 6 h (#p : Benjamini and Hochberg corrected p-value) [data were extracted from (10)]. (B) CHAC1 mRNA expression in hAECBs from patients without CF (N = 4, black) and with CF (N = 4, gray) infected with Pa (PAK strain, MOI 0.25) for 0 or 6 h. Statistical analysis was conducted using Mann-Whitney tests (without vs. with CF groups, *p < 0.05). (C) CHAC1 mRNA expression in hAECBs from patients without CF (black) and with CF (gray) stimulated with 1 μg/mL LPS and 50 ng/mL flagellin or infected with PAK (MOI 0.25) for 6 h. Data are expressed as means ± SEMs of four independent experiments. Statistical analysis was carried out using analysis of variance followed by Dunnett's multiple comparison test (*p < 0.05, NS used as the control group).
We further validated the differential expression of CHAC1 by qPCR in the same samples used for RNA-seq. Indeed, at baseline, we observed significantly higher expression of CHAC1 in non-CF cells compared with that in CF cells. As expected from the transcriptomic data, we observed upregulation of CHAC1 mRNA expression after 6 h of Pa infection only in non-CF cells (Figure 1B).
LPS and Flagellin Induced CHAC1 Overexpression in Primary Bronchial Epithelial Cells and NCI-H292 Cells
Because Pa induced CHAC1 expression in non-CF cells, we investigated which Pa virulence factors were involved. We studied the effects of two very well-known virulence factors, LPS and flagellin from Pa, on hAECBs from CF and non-CF patients and on NCI-H292 cells, which expressed CHAC1 mRNA at sufficient levels for further knockdown experiments in comparison to BEAS-2B cells (Supplementary Figure 2A). Notably, 6 h of stimulation with LPS and flagellin induced CHAC1 expression in hAECBs from non-CF patients, similar to whole bacteria at a MOI of 0.25 for 6 h. This induction by LPS and flagellin and by living bacteria was not found in hAECBs from CF patients (Figure 1C). We observed similar effects of LPS and flagellin on CHAC1 expression in NCI-H292 cells (Figure 2A). To validate the role of flagellin in CHAC1 expression, we used a mutant of the Pa PAK strain, ΔFliC, in which flagellin was deleted. We observed a significantly lower increase in CHAC1 mRNA expression when NCI-H292 cells were stimulated with the ΔFliC strain than with the WT PAK strain (Figure 2B). We also tested a double mutant of the Pa PAK strain with both type II and III secretion systems deleted (ΔpscFΔxcpQ). This mutant induced CHAC1 mRNA expression, similar to the WT PAK strain (Figure 2B). The difference in CHAC1 expression observed with the ΔFliC strain was not due to different MOI since equal numbers of Pa colonies were found on LB agar (Supplementary Figure 3A). Subsequently, in order to determine whether the observed CHAC1 upregulation was restricted to the laboratory PAK strain, we used Pa strains freshly isolated from different patients with CF. We observed that clinical isolates of Pa were also able to induce CHAC1 mRNA expression in NCI-H292 cells (Figure 2C) with an effect similar to PAK. This induction is highly variable between the clinical strains used and may reflect difference of virulence of each strain, since the number of the bacteria used for stimulation is similar (Supplementary Figure 3B). CHAC1 induction by PAK strain is also highly variable (Figure 2C), and is likely due to the variability in the inoculum used (Supplementary Figure 3B). We also observed that the induction of CHAC1 mRNA expression by PAK was higher in NCI-H292 cells than in hAECBs (Figures 2B,C vs. Figure 1C). Finally, we tested two other bacterial species known to colonize the airways of patients with CF, i.e., S. aureus and S. maltophilia. Using a similar MOI, we observed reduced CHAC1 mRNA induction in NCI-H292 cells infected with S. aureus and S. maltophilia compared with that in the cells infected with Pa. Concomitantly, induction of IL-8 mRNA expression was reduced (Figure 2D). The differences in CHAC1 and IL-8 mRNA expression observed with S. aureus and S. maltophilia were not caused by different MOI because equal numbers of colonies were found on LB agar (Supplementary Figure 3C).
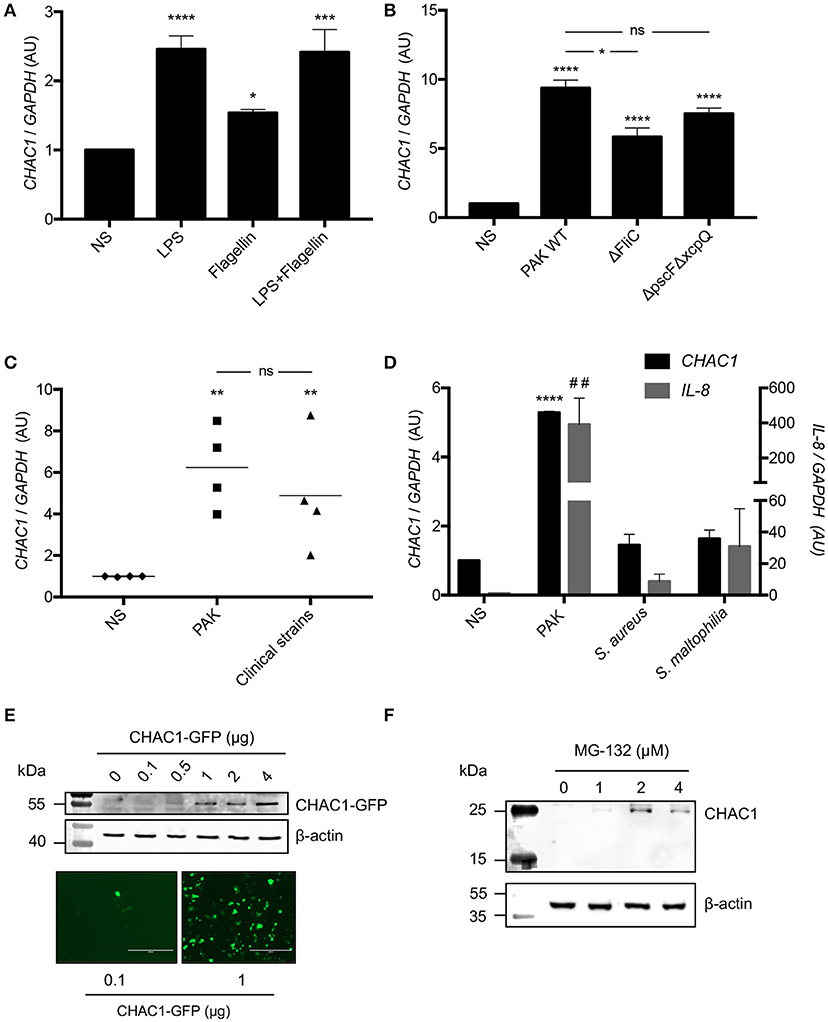
Figure 2. (A) CHAC1 mRNA expression in NCI-H292 cells stimulated with 1 μg/mL LPS and/or 50 ng/mL flagellin for 6 h. Data are expressed as the means ± SEMs of five independent experiments. Statistical analysis was performed using analysis of variance followed by Dunnett's multiple comparison test (control group: NS, *p < 0.05; ***p < 0.001; ****p < 0.0001). (B) CHAC1 mRNA expression in NCI-H292 cells infected with PAK WT, PAK ΔFliC, or PAK ΔpscFΔxcpQ (MOI 0.25) for 6 h. Data are expressed as means ± SEMs of four independent experiments. Statistical analysis was carried out using analysis of variance followed by Bonferroni's multiple comparison test (control group: NS, *p < 0.05; ****p < 0.0001). (C) CHAC1 mRNA expression in NCI-H292 cells infected with PAK strain (MOI 0.25) or different clinical strains of Pa (MOI 0.25) for 6 h. Data are expressed as means ± SEMs of four independent experiments (NS, diamonds; PAK, squares; clinical strains, triangles). Statistical analysis was carried out using analysis of variance followed by Bonferroni's multiple comparison test (control group: NS, **p < 0.01). (D) CHAC1 (black) and IL-8 (gray) mRNA expression in NCI-H292 cells infected with PAK, Staphylococcus aureus (S. aureus), or Stenotrophomonas maltophilia (S. maltophilia, MOI 0.25) for 6 h. Data are expressed as means ± SEMs of three independent experiments. Statistical analysis was carried out using analysis of variance followed by Dunnett's multiple comparison test (control group: NS, ##p < 0.01 [IL-8]; ****p < 0.0001 [CHAC1]). (E) CHAC1 and β-actin protein expression and CHAC1-GFP fluorescence in NCI-H292 cells transfected with increasing amounts of CHAC1-GFP plasmid. (F) CHAC1 and β-actin protein expression in NCI-H292 cells treated with increasing concentrations of MG-132 for 6 h.
Next, we examined CHAC1 protein expression in response to Pa stimulation. Consistent with previous reports indicating that CHAC1 protein is very unstable due to its rapid degradation by the proteasome in Neuro2a cells (23), we were unable to detect endogenous CHAC1 protein in NCI-H292 at baseline (Supplementary Figure 2B) or after Pa infection (data not shown). Additionally, we were unable to detect CHAC1 protein expression after treating the cells with tunicamycin, a known inducer of CHAC1 mRNA expression (24) (data not shown). CHAC1 was only detected at the protein level when it was either overexpressed with a plasmid in NCI-H292 cells (Figure 2E) or when the cells were treated with at least 1 μM of the proteasome inhibitor MG-132 (Figure 2F).
CHAC1 Upregulation Was ATF4 Dependent and PERK Independent
Because CHAC1 has been shown to be associated with the PERK pathway (11), we hypothesized that induction of CHAC1 by LPS from Pa may be mediated by PERK. Therefore, we used a PERK inhibitor and tunicamycin (TM) as a positive control for activation of the CHAC1 and PERK pathway (11). As expected, compared with untreated NCI-H292 cells, LPS and TM induced CHAC1 mRNA expression. When cells were treated with TM and PERK inhibitor, there was a significant 90.3% decrease in CHAC1 mRNA expression compared with that in cells treated with TM alone (Figure 3A). However, when cells were treated with LPS and PERK inhibitor, CHAC1 mRNA expression was not significantly different from that in cells treated with LPS only (Figure 3A). We then hypothesized that this induction of CHAC1 by LPS would directly pass through ATF4. Indeed, we observed that LPS was able to activate ATF4 and that siRNA against ATF4 inhibited this induction (Figure 3B). The inhibition of ATF4 expression by the siRNA ATF4 (~60%) was associated with an inhibition of CHAC1 induction by LPS (Figures 3C,D).
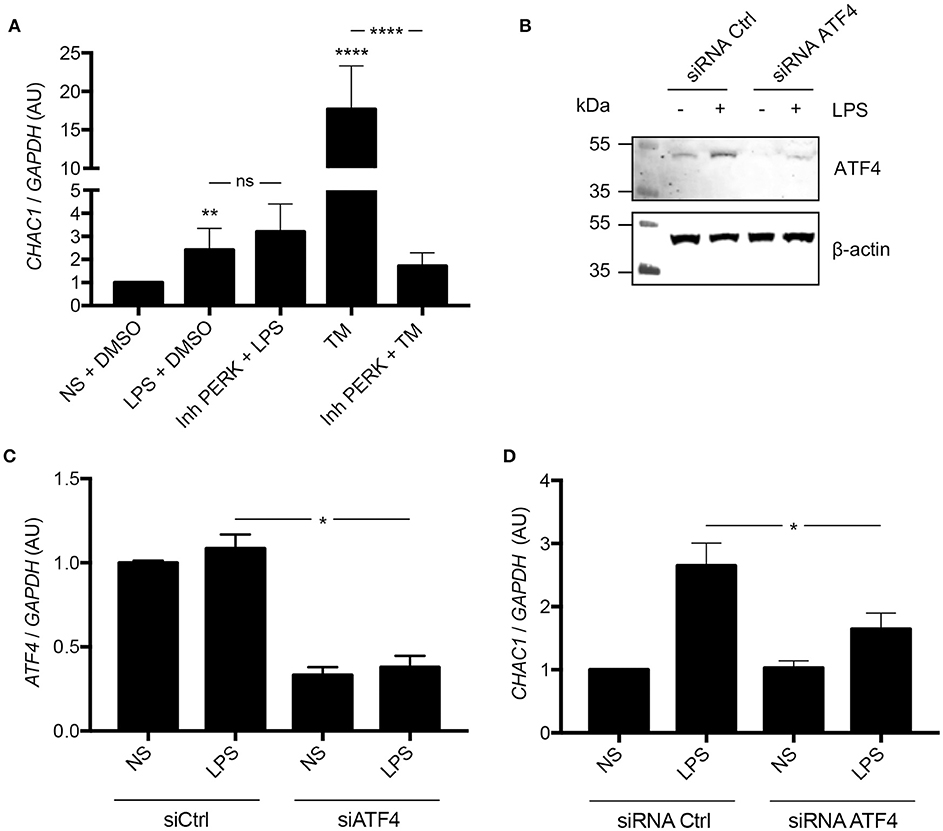
Figure 3. (A) CHAC1 mRNA expression in NCI-H292 cells stimulated with 1 μg/mL LPS in DMSO (vehicle) or 1 μM PERK inhibitor (Inh PERK) or stimulated with 0.5 μg/mL tunicamycin (TM) with or without PERK inhibitor for 6 h. Data are expressed as means ± SEMs of five independent experiments. Statistical analysis was performed with analysis of variance followed by Bonferroni's multiple comparison test (**p < 0.01, ****p < 0.0001). (B) Western blot analysis of ATF4 expression in NCI-H292 cells treated 48 h with siRNA Ctrl or siRNA ATF4 and stimulated with (+) or without (–) LPS for 1 h (images are representative of six independent experiments). (C) ATF4 mRNA expression in NCI-H292 cells treated for 48 h with siRNA Ctrl or siRNA ATF4 and stimulated with or without (NS) LPS for 1 h. Data are means ± SEMs of six independent experiments. Statistical analysis was performed using Mann-Whitney tests (*p < 0.05). (D) CHAC1 mRNA expression in NCI-H292 cells treated for 48 h with siRNA Ctrl or siRNA ATF4 and stimulated with or without (NS) LPS for 1 h. Data are means ± SEMs of six independent experiments. Statistical analysis was performed using Mann-Whitney tests (*p < 0.05).
CHAC1 Overexpression Induced by Pa Did Not Stimulate Apoptosis
CHAC1 has been shown to play a role in apoptosis (11); therefore, we investigated the effects of Pa stimulation on apoptosis. We infected NCI-H292 cells with two different MOIs (0.5 and 1) for 5 or 8 h and examined the levels of cleaved PARP and cleaved caspase-3. As expected, we observed that the positive control staurosporine induced cleavage of PARP and caspase-3. In contrast, Pa did not induce cleavage of PARP and caspase-3 (Figure 4A). PARP and caspase-3 cleavages were observed when CHAC1 was overexpressed with a plasmid in NCI-H292 cells (Figure 4B).
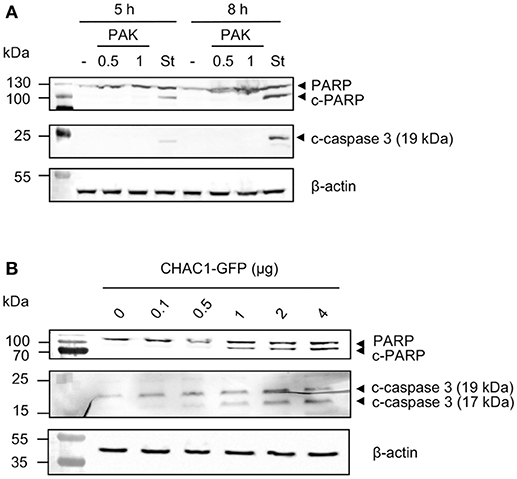
Figure 4. (A) PARP (uncleaved form), c-PARP (cleaved form), c-caspase 3 (cleaved form), and β-actin protein expression in NCI-H292 cells infected with PAK (MOI 0.5 or 1) or stimulated with 1 μM staurosporine (St) for 5 or 8 h. (B) PARP (uncleaved form), c-PARP (cleaved form), c-caspase 3 (cleaved forms) and β-actin protein expression in NCI-H292 cells transfected with p-CHAC1-GFP (same blot as shown in Figure 2E).
CHAC1 Regulated the Inflammatory Response of Bronchial Epithelial Cells Induced by LPS or Flagellin
CHAC1 is a component of the ER stress pathway, and a link between ER stress and the inflammatory response has already been described in the context of CF (12). Therefore, we investigated the role of CHAC1 in the inflammatory response induced by LPS or flagellin. Transfection with CHAC1 siRNA induced around 50% reduction in CHAC1 mRNA expression (Figure 5A). We then examined different inflammatory parameters, particularly the secretion of IL-8, IL-6, and PGE2. As expected, we observed that 1 μg/mL LPS, 50 ng/mL flagellin, and the combination of both (LPS+flagellin) induced significant increases in both IL-8 and IL-6 secretion and tended to increase PGE2 secretion (Figures 5B–D). Under these stimulation conditions, when CHAC1 mRNA expression was decreased using siRNA, greater significant increases in IL-8 secretion (Figure 5B) and PGE2 secretion (Figure 5D) were observed. In contrast, in the presence of CHAC1 siRNA, we observed significant inhibition of IL-6 secretion (Figure 5C) in cells stimulated with LPS (80% inhibition), flagellin (30% inhibition), or the combination of both virulence factors (80% inhibition). We obtained similar results for the mRNA expression of IL-8 and IL-6 (Supplementary Figures 4A,C). CCL2 mRNA expression was also modulated similar to IL-8 expression under these experimental conditions (not detectable by ELISA; Supplementary Figure 4B).
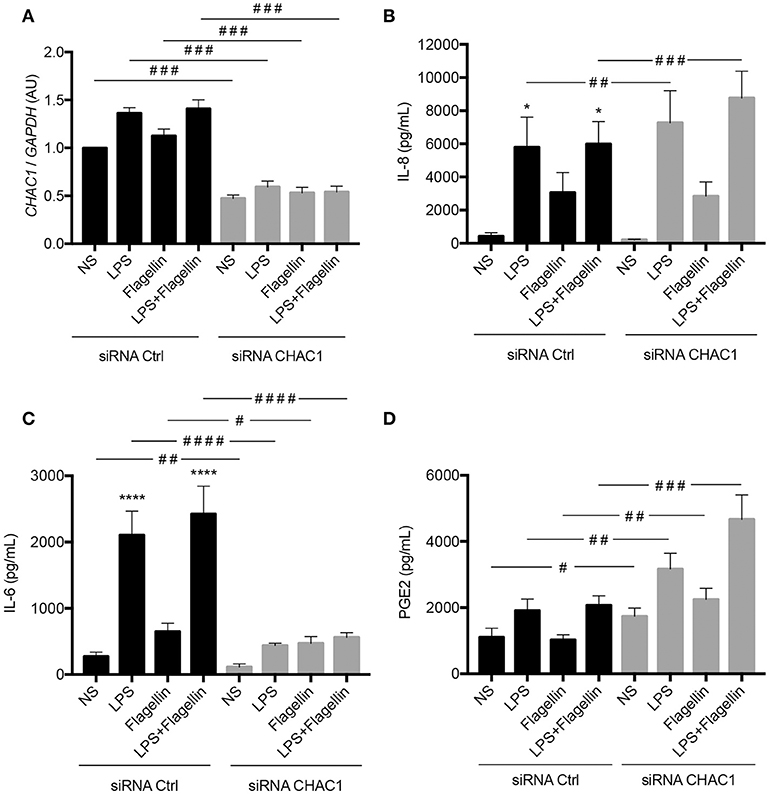
Figure 5. (A) CHAC1 mRNA expression in NCI-H292 cells transfected with siRNA control (siRNA Ctrl, black) or siRNA CHAC1 (gray) for 48 h and stimulated with LPS and/or flagellin for 6 h. Data are expressed as means ± SEMs of 10 independent experiments. (B) IL-8, (C) IL-6, and (D) PGE2 productions in supernatants from cells transfected with siRNA control (siRNA Ctrl, black) or siRNA CHAC1 (gray) for 48 h and stimulated with LPS and/or flagellin for 6 h. Data are expressed as means ± SEMs of 10 independent experiments. Statistical analysis was performed using analysis of variance followed by Dunnett's multiple comparison tests (control group: NS, *p < 0.05, ****p < 0.0001) and Wilcoxon tests (siRNA Ctrl vs. siRNA CHAC1 groups, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001).
CHAC1 Regulated NF-κB Activation
To determine whether regulation of these inflammatory parameters involved NF-κB, we studied its phosphorylation by western blot in NCI-H292 cells. We observed that LPS induced a significant increase in NF-κB p65 phosphorylation in CHAC1-knockdown cells after 1 h of stimulation (Figure 6A). To confirm these data, we used A549-NF-κB luciferase cells. As in NCI-H292 cells, 60% inhibition of CHAC1 mRNA expression was observed after siRNA transfection (data not shown). Similar to NCI-H292 cells, we observed that 1 μg/mL LPS, 50 ng/mL flagellin, and the combination of both (LPS+flagellin) induced a significant increase in IL-8 secretion when A549 cells were transfected with CHAC1 siRNA compared with that in cells transfected with siRNA control (Figure 6B). Additionally, as expected, we observed that 1 μg/mL LPS, 50 ng/mL flagellin, and the combination of both (LPS+flagellin) induced significant increases in NF-κB activity (Figure 6C). Under these experimental conditions, when we decreased CHAC1 mRNA expression, we observed an even greater increase in NF-κB activity when cells were stimulated with LPS and/or flagellin (Figure 6C). IL-6 and CCL-2 were not detectable by ELISA (data not shown).
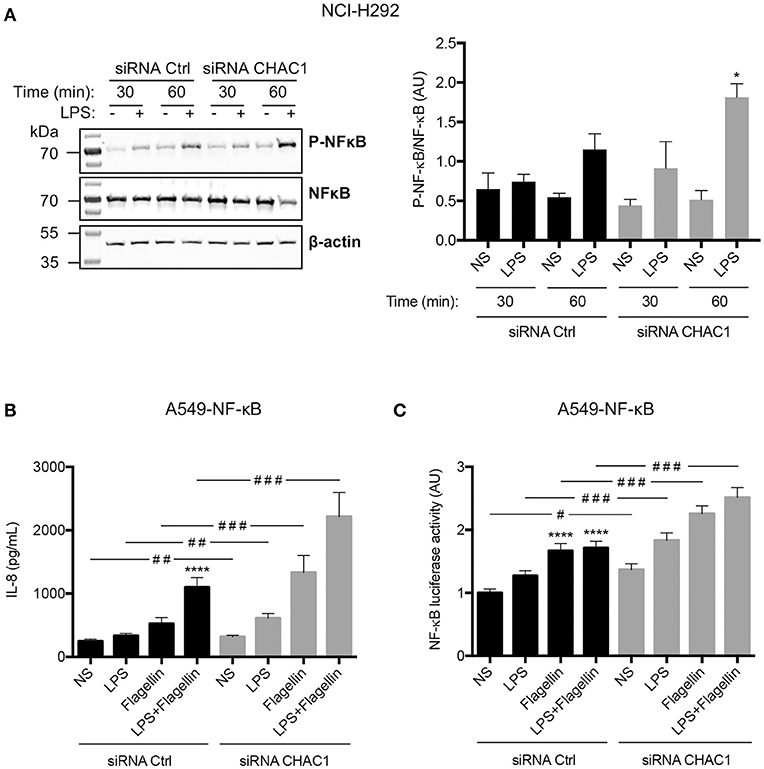
Figure 6. (A) Phospho-NF-κB (p-NF-κB), total NF-κB (NF-κB), and β-actin protein expression in NCI-H292 cells transfected with siRNA control (siRNA Ctrl) or siRNA CHAC1 for 48 h and then stimulated for 30 min or 1 h with (+) or without (–) 1 μg/mL LPS (images are representative of four independent experiments; left panel). Results are shown as p-NF-κB/NF-κB ratios (right panel). Data are expressed as means ± SEMs of four experiments. Statistical analysis was carried out using Mann-Whitney tests (*p < 0.05, siCtrl-LPS 1 h vs. siCHAC1-LPS 1 h). (B) IL-8 production and (C) NF-κB luciferase activity in A549-NF-κB luciferase cells transfected with siRNA control (siRNA Ctrl, black) or siRNA CHAC1 (gray) for 48 h and stimulated with LPS and/or flagellin for 24 h. Data are expressed as means ± SEMs of four independent experiments. Statistical analysis was performed using analysis of variance followed by Dunnett's multiple comparison tests (control group: NS, ****p < 0.0001) and Wilcoxon test (siRNA Ctrl vs. siRNA CHAC1 groups, #p < 0.05; ##p < 0.01; ###p < 0.001).
Discussion
Independent of CFTR mutations, links between ER stress and CF-related inflammation have been extensively studied, particularly with regard to inositol-requiring enzyme 1 (IRE1α), and PERK (25) pathways. In our previous study, we identified the CHAC1 mRNA as one of the most highly differentially expressed transcripts between primary bronchial cells from CF and non-CF patients, infected with Pa. We also found that CF cells exhibited an expected excessive inflammatory response, including higher IL-8 production (10). Because CHAC1 was recently described as a new component of the UPR in the PERK pathway (12), we aimed to evaluate whether CHAC1 was involved in the modulation of the inflammatory response induced by Pa in lung epithelial cells.
We confirmed our previous transcriptomic data showing that CHAC1 mRNA was overexpressed in primary non-CF epithelial cells compared with its expression in CF cells at baseline and upon Pa infection. Additionally, Pa and two of its virulence factors, LPS and flagellin, the respective ligands of Toll-Like Receptor (TLR) 4 and TLR5, were able to induce CHAC1 at the transcriptional level in bronchial epithelial NCI-H292 cells. As previously shown in Neuro2a (23) or HEK cells (26), we were able to detect CHAC1 protein expression only after MG-132 treatment, suggesting the involvement of the proteasome pathway in the stability of the protein (26). This induction of CHAC1 mRNA expression was observed not only with the laboratory strain PAK but also with freshly isolated clinical strains. Interestingly, S. maltophilia and S. aureus, both known to chronically infect CF patients, were not able to induce CHAC1 mRNA expression to similar levels. Using cellular infection models, other transcriptomic studies have consistently shown CHAC1 upregulation with microorganisms, including human coronavirus (27), human cytomegalovirus (28), tick-borne flaviviruses (29), Zika virus (30), and the bacterium Mycoplasma hominis (31). In these studies, the specific role of CHAC1 was not investigated. However, CHAC1 is essential as embryonic lethality was observed in mice with deletion of the Chac1 gene (32).
In our study, CHAC1 mRNA induction by LPS and Pa was not related to PERK activation, as is TM, but was dependent on ATF4. In contrast, LPS from E. coli has previously been shown to activate both PERK and ATF4 in A549 cells (33). Notably, ATF4 has been shown to be directly recruited by TLR4 independent of ER stress in human monocytes (34). Consistent with our results, another ER stress-associated molecule, GADD34, was recently shown to be upregulated by several virulence factors from Pa, including pyocyanin and AprA, independent of PERK but dependent on ATF4 (35). Taken together, these results were consistent with emerging evidence showing that induction of the UPR pathway by microorganisms did not reflect ER stress but was an integral part of the immune response and TLR pathways (36).
The fine tuning of immune responses by the UPR has been extensively studied (37). However, its contribution to the regulation of cytokine production in bacterial infection is not well understood (16). Here, we observed that inhibition of CHAC1 mRNA expression by siRNA promoted IL-8 and CCL2 expression and PGE2 secretion, suggesting that CHAC1 prevents excessive inflammatory responses dependent on the NF-κB p65 pathway. These results were consistent with a recent study reporting that bronchial epithelial cells undergoing ER stress showed reduced IL-8 and CXCL1 production in the presence of inflammatory stimuli (38). Indeed, pretreatment of bronchial epithelial cells with TM [shown here and by others (11) to be able to increase CHAC1 expression] can prevent IL-8 and CXCL1 production. In contrast, inhibition of CHAC1 mRNA expression by siRNA inhibits IL-6 production, suggesting that CHAC1 may promote IL-6 expression. Our results were consistent with a previous study showing that TM promotes IL-6 secretion in bronchial IB3-1 (CF deficient) and C38 cells stimulated with flagellin (12). Previous works have demonstrated that IL-6 production in A375 skin cells is dependent on CHOP (39). However, CHOP is expected to be upstream in the CHAC1 pathway and does not explain our results; future studies are needed to determine how the IL-6 pathway is affected specifically in lung epithelial cells.
In addition to its role in UPR, CHAC1, as a member of the γ-glutamylcyclotransferase family, is able to hydrolyze glutathione only in its reduced form (GSH) (40). Glutathione has been widely studied owing to the function of CFTR as a glutathione transporter (41). GSH homeostasis is defective in CF, and GSH promotes oxidative stress and related inflammation (42). The roles of CHAC1 activity in relation to cellular glutathione homeostasis and the observed cytokine regulation remain to be determined, although previous studies have demonstrated that the levels of glutathione-related enzyme can regulate cytokine production (43).
In our experiments, PAK did not induce apoptosis in NCI-H292 cells. However, these cells are not completely resistant to apoptosis since staurosporine was able to induce caspase-3 and PARP cleavage. Thus, although CHAC1 was induced by Pa, it was not sufficient to induce cell apoptosis. It is conceivable that the level of induction of CHAC1 may be too low to promote apoptosis. In contrast, high induction of CHAC1 using an overexpression plasmid led to induction of PARP cleavage, indicative of cell apoptosis. Apoptosis dysfunction is still highly controversial in CF (44), and our results are partly consistent with some previous studies. Indeed, even if Pa has been previously shown to induce apoptosis in lung epithelial cells, variable susceptibility between lung epithelial cells has been observed. Moreover, the PAO1 strain of Pa has been shown to induce apoptosis in human tracheal 9HTEo cells but not in primary nasal epithelial cells from polyps or in 16HBE14o bronchial cells, which are both resistant (45). In contrast, other bronchial cell lines are permissive to apoptosis induced by Pa, such as IB3-1 (F508del/W1282X), C38, S9 (both IB3-1 corrected cells), CFT1-LCFSN, and CFT1-LC3 (F508del) cells. More recently, using Calu-3 polarized lung epithelial cells, researchers showed that PAO1 was able to induce apoptosis markers, such as active caspase-3, within 6–8 h. Additionally, the effects of the CFTR genotype on apoptosis of lung epithelial cells at baseline (44) or after Pa infection are still debated and may be dependent on which specific marker of apoptosis is being evaluated (45, 46).
In conclusion, we showed that inhibition of CHAC1 in bronchial epithelial cells resulted in the regulation of inflammatory mediators. Moreover, CHAC1 was induced preferentially in normal hAECBs in comparison to CF hAECBs in response to Pa and its virulence factors, LPS and flagellin. Thus, these results suggest that low CHAC1 expression may contribute to the excessive inflammation observed in CF bronchial epithelial cells after Pa infection and thus may be associated with the exacerbation of inflammation characterizing the lungs from patients with CF. Inflammation is complex and thought to have a critical impact during the course of CF lung disease (47). A better understanding of the link between infection and inflammation is thus required for the development of new therapeutics.
Author Contributions
LG, MC, and VB designed the study. LG and LP wrote the manuscript. LP, VB, TF, and HP performed the experiments. LG, LP, and VB performed the data analysis. LT participated in PGE2 and Staphylococcus experiments. DM participated in bacterial sample collection. IM, DM, LT, MC, HC, and VB critically revised the manuscript.
Funding
MC received a grant from the French cystic fibrosis nonprofit organizations Vaincre La Mucoviscidose and Gregory Lemarchal. LP received a Ph.D. fellowship from the French cystic fibrosis nonprofit organization Vaincre La Mucoviscidose.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Sarah Fedele for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2018.02823/full#supplementary-material
References
1. Kerem B, Rommens JM, Buchanan JA, Markiewicz D, Cox TK, Chakravarti A, et al. Identification of the cystic fibrosis gene: genetic analysis. Science (1989) 245:1073–80. doi: 10.1126/science.2570460
2. Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science (1989) 245:1066–73. doi: 10.1126/science.2475911
3. Rommens JM, Iannuzzi MC, Kerem B, Drumm ML, Melmer G, Dean M, et al. Identification of the cystic fibrosis gene: chromosome walking and jumping. Science (1989) 245:1059–65. doi: 10.1126/science.2772657
4. Cohen TS, Prince A, Cystic fibrosis: a mucosal immunodeficiency syndrome. Nat Med. (2012) 18:509–19. doi: 10.1038/nm.2715
5. Folkesson A, Jelsbak L, Yang L, Johansen HK, Ciofu O, Hoiby N, et al. Adaptation of Pseudomonas aeruginosa to the cystic fibrosis airway: an evolutionary perspective. Nat Rev Microbiol. (2012) 10:841–51. doi: 10.1038/nrmicro2907
6. Khan TZ, Wagener JS, Bost T, Martinez J, Accurso FJ, Riches DW Early pulmonary inflammation in infants with cystic fibrosis. Am J Respir Crit Care Med. (1995) 151:1075–82. doi: 10.1164/ajrccm.151.4.7697234
7. Berube J, Roussel L, Nattagh L, Rousseau S Loss of cystic fibrosis transmembrane conductance regulator function enhances activation of p38 and ERK MAPKs increasing interleukin-6 synthesis in airway epithelial cells exposed to Pseudomonas aeruginosa. J Biol Chem. (2010) 285:22299–307. doi: 10.1074/jbc.M109.098566
8. Whitsett JA, Alenghat T. Respiratory epithelial cells orchestrate pulmonary innate immunity. Nat Immunol. (2015) 16:27–35. doi: 10.1038/ni.3045
9. Anas AA, van Lieshout MH, Claushuis TA, de Vos AF, Florquin S, de Boer OJ, et al. Lung epithelial MyD88 drives early pulmonary clearance of Pseudomonas aeruginosa by a flagellin dependent mechanism. Am J Physiol Lung Cell Mol Physiol. (2016) 311:L219–28. doi: 10.1152/ajplung.00078.2016
10. Balloy V, Varet H, Dillies MA, Proux C, Jagla B, Coppee JY, et al. Normal and cystic fibrosis human bronchial epithelial cells infected with Pseudomonas aeruginosa exhibit distinct gene activation patterns. PLoS ONE (2015) 10:e0140979. doi: 10.1371/journal.pone.0140979
11. Mungrue IN, Pagnon J, Kohannim O, Gargalovic PS, Lusis AJ CHAC1/MGC4504 is a novel proapoptotic component of the unfolded protein response downstream of the ATF4-ATF3-CHOP cascade. J Immunol. (2009) 182:466–76. doi: 10.4049/jimmunol.182.1.466
12. Blohmke CJ, Mayer ML, Tang AC, Hirschfeld AF, Fjell CD, Sze MA, et al. Atypical activation of the unfolded protein response in cystic fibrosis airway cells contributes to p38 MAPK-mediated innate immune responses. J Immunol. (2012) 189:5467–75. doi: 10.4049/jimmunol.1103661
13. Kumar A, Tikoo S, Maity S, Sengupta S, Sengupta S, Kaur A, et al. Mammalian proapoptotic factor ChaC1 and its homologues function as gamma-glutamyl cyclotransferases acting specifically on glutathione. EMBO Rep. (2012) 13:1095–101. doi: 10.1038/embor.2012.156
14. Luciani A, Villella VR, Esposito S, Brunetti-Pierri N, Medina D, Settembre C, et al. Defective CFTR induces aggresome formation and lung inflammation in cystic fibrosis through ROS-mediated autophagy inhibition. Nat Cell Biol. (2010) 12:863–75. doi: 10.1038/ncb2090
15. Park SH, Choi HJ, Yang H, Do KH, Kim J, Lee DW, et al. Endoplasmic reticulum stress-activated C/EBP homologous protein enhances nuclear factor-kappaB signals via repression of peroxisome proliferator-activated receptor gamma. J Biol Chem. (2010) 285:35330–9. doi: 10.1074/jbc.M110.136259
16. Smith JA. Regulation of cytokine production by the unfolded protein response; implications for infection and autoimmunity. Front Immunol. (2018) 9:422. doi: 10.3389/fimmu.2018.00422
17. Le Gars M, Descamps D, Roussel D, Saussereau E, Guillot L, Ruffin M, et al. Neutrophil elastase degrades cystic fibrosis transmembrane conductance regulator via calpains and disables channel function in vitro and in vivo. Am J Respir Crit Care Med. (2013) 187:170–9. doi: 10.1164/rccm.201205-0875OC
18. Balloy V, Deveaux A, Lebeaux D, Tabary O, le Rouzic P, Ghigo JM, et al. Azithromycin analogue CSY0073 attenuates lung inflammation induced by LPS challenge. Br J Pharmacol. (2014) 171:1783–94. doi: 10.1111/bph.12574
19. Corvol H, Rousselet N, Thompson KE, Berdah L, Cottin G, Foussigniere T, et al. FAM13A is a modifier gene of cystic fibrosis lung phenotype regulating rhoa activity, actin cytoskeleton dynamics and epithelial-mesenchymal transition. J Cyst Fibros. (2018) 17:190–203. doi: 10.1016/j.jcf.2017.11.003
20. Ramphal R, Balloy V, Huerre M, Si-Tahar M, Chignard M. TLRs 2 and 4 are not involved in hypersusceptibility to acute Pseudomonas aeruginosa lung infections. J Immunol. (2005) 175:3927–34. doi: 10.4049/jimmunol.175.6.3927
21. Jyot J, Balloy V, Jouvion G, Verma A, Touqui L, Huerre M, et al. Type II secretion system of Pseudomonas aeruginosa: in vivo evidence of a significant role in death due to lung infection. J Infect Dis. (2011) 203:1369–77. doi: 10.1093/infdis/jir045
22. Pernet E, Brunet J, Guillemot L, Chignard M, Touqui L, Wu Y. Staphylococcus aureus adenosine inhibits sPLA2-IIA-mediated host killing in the airways. J Immunol. (2015) 194:5312–9. doi: 10.4049/jimmunol.1402665
23. Oh-Hashi K, Nomura Y, Shimada K, Koga H, Hirata Y, Kiuchi K. Transcriptional and post-translational regulation of mouse cation transport regulator homolog 1. Mol Cell Biochem. (2013) 380:97–106. doi: 10.1007/s11010-013-1663-1
24. Crawford RR, Prescott ET, Sylvester CF, Higdon AN, Shan J, Kilberg MS, et al. Human CHAC1 protein degrades glutathione, and mRNA induction is regulated by the transcription factors ATF4 and ATF3 and a bipartite ATF/CRE regulatory element. J Biol Chem. (2015) 290:15878–91. doi: 10.1074/jbc.M114.635144
25. Ribeiro CM, Boucher RC. Role of endoplasmic reticulum stress in cystic fibrosis-related airway inflammatory responses. Proc Am Thorac Soc. (2010) 7:387–94. doi: 10.1513/pats.201001-017AW
26. Nomura Y, Hirata Y, Kiuchi K, Oh-Hashi K. Translational and post-translational regulation of mouse cation transport regulator homolog 1. Sci Rep. (2016) 6:28016. doi: 10.1038/srep28016
27. Poppe M, Wittig S, Jurida L, Bartkuhn M, Wilhelm J, Muller H, et al. The NF-kappaB-dependent and -independent transcriptome and chromatin landscapes of human coronavirus 229E-infected cells. PLoS Pathog. (2017) 13:e1006286. doi: 10.1371/journal.ppat.1006286
28. Gurczynski SJ, Das S, Pellett PE. Deletion of the human cytomegalovirus US17 gene increases the ratio of genomes per infectious unit and alters regulation of immune and endoplasmic reticulum stress response genes at early and late times after infection. J Virol. (2014) 88:2168–82. doi: 10.1128/JVI.02704-13
29. Mlera L, Lam J, Offerdahl DK, Martens C, Sturdevant D, Turner CV, et al. Transcriptome analysis reveals a signature profile for tick-borne flavivirus persistence in HEK 293T cells. mBio (2016) 7:e00314–16. doi: 10.1128/mBio.00314-16
30. Rolfe AJ, Bosco DB, Wang J, Nowakowski RS, Fan J, Ren Y. Bioinformatic analysis reveals the expression of unique transcriptomic signatures in Zika virus infected human neural stem cells. Cell Biosci. (2016) 6:42. doi: 10.1186/s13578-016-0110-x
31. Hopfe M, Deenen R, Degrandi D, Kohrer K, Henrich B. Host cell responses to persistent mycoplasmas—Different stages in infection of HeLa cells with Mycoplasma hominis. PLoS ONE (2013) 8:e54219. doi: 10.1371/journal.pone.0054219
32. Mungrue I, Prescott ET, Sylvester CF, Crawford RR. Abstract 324: knockout of the pro-apoptotic Er-stress gene Chac1 in mice results in embryonic lethality and activation of the notch pathway. Circ Res. (2015) 117:A324. doi: 10.1161/res.117.suppl_1.324
33. Li S, Guo L, Qian P, Zhao Y, Liu A, Ji F, et al. Lipopolysaccharide induces autophagic cell death through the PERK-dependent branch of the unfolded protein response in human alveolar epithelial A549 cells. Cell Physiol Biochem. (2015) 36:2403–17. doi: 10.1159/000430202
34. Zhang C, Bai N, Chang A, Zhang Z, Yin J, Shen W, et al. ATF4 is directly recruited by TLR4 signaling and positively regulates TLR4-trigged cytokine production in human monocytes. Cell Mol Immunol. (2013) 10:84–94. doi: 10.1038/cmi.2012.57
35. van't Wout EF, van Schadewijk A, van Boxtel R, Dalton LE, Clarke HJ, Tommassen J, et al. Virulence factors of Pseudomonas aeruginosa induce both the unfolded protein and integrated stress responses in airway epithelial cells. PLoS Pathog. (2015) 11:e1004946. doi: 10.1371/journal.ppat.1004946
36. Claudio N, Dalet A, Gatti E, Pierre P. Mapping the crossroads of immune activation and cellular stress response pathways. EMBO J. (2013) 32:1214–24. doi: 10.1038/emboj.2013.80
37. Grootjans J, Kaser A, Kaufman RJ, Blumberg RS. The unfolded protein response in immunity and inflammation. Nat Rev Immunol. (2016) 16:469–84. doi: 10.1038/nri.2016.62
38. Tang AC, Saferali A, He G, Sandford AJ, Strug LJ, Turvey SE. Endoplasmic reticulum stress and chemokine production in cystic fibrosis airway cells: regulation by STAT3 modulation. J Infect Dis. (2017) 215:293–302. doi: 10.1093/infdis/jiw516
39. Hattori T, Itoh S, Hayashi H, Chiba T, Takii T, Yoshizaki K, et al. CHOP, a basic leucine zipper transcriptional factor, contributes to the antiproliferative effect of IL-1 on A375 human melanoma cells through augmenting transcription of IL-6. J Interferon Cytokine Res. (2001) 21:323–32. doi: 10.1089/107999001300177510
40. Tsunoda S, Avezov E, Zyryanova A, Konno T, Mendes-Silva L, Pinho Melo E, et al. Intact protein folding in the glutathione-depleted endoplasmic reticulum implicates alternative protein thiol reductants. Elife (2014) 3:e03421. doi: 10.7554/eLife.03421
41. Linsdell P, Hanrahan JW. Glutathione permeability of CFTR. Am J Physiol. (1998) 275(1 Pt 1):C323–6. doi: 10.1152/ajpcell.1998.275.1.C323
42. Galli F, Battistoni A, Gambari R, Pompella A, Bragonzi A, Pilolli F, et al. Oxidative stress and antioxidant therapy in cystic fibrosis. Biochim Biophys Acta (2012) 1822:690–713. doi: 10.1016/j.bbadis.2011.12.012
43. Haddad JJ, Safieh-Garabedian B, Saade NE, Lauterbach R. Inhibition of glutathione-related enzymes augments LPS-mediated cytokine biosynthesis: involvement of an IkappaB/NF-kappaB-sensitive pathway in the alveolar epithelium. Int Immunopharmacol. (2002) 2;1567–83. doi: 10.1016/S1567-5769(02)00117-0
44. Soleti R, Porro C, Martinez MC. Apoptotic process in cystic fibrosis cells. Apoptosis (2013) 18:1029–38. doi: 10.1007/s10495-013-0874-y
45. Rajan S, Cacalano G, Bryan R, Ratner AJ, Sontich CU, van Heerckeren A, et al. Pseudomonas aeruginosa induction of apoptosis in respiratory epithelial cells: analysis of the effects of cystic fibrosis transmembrane conductance regulator dysfunction and bacterial virulence factors. Am J Respir Cell Mol Biol. (2000) 23:304–12. doi: 10.1165/ajrcmb.23.3.4098
46. Cannon CL, Kowalski MP, Stopak KS, Pier GB. Pseudomonas aeruginosa-induced apoptosis is defective in respiratory epithelial cells expressing mutant cystic fibrosis transmembrane conductance regulator. Am J Respir Cell Mol Biol. (2003) 29:188–97. doi: 10.1165/rcmb.4898
Keywords: cystic fibrosis, bronchial epithelium, Pseudomonas aeruginosa, inflammation, ChaC glutathione-specific γ-glutamylcyclotransferase 1, ER stress
Citation: Perra L, Balloy V, Foussignière T, Moissenet D, Petat H, Mungrue IN, Touqui L, Corvol H, Chignard M and Guillot L (2018) CHAC1 Is Differentially Expressed in Normal and Cystic Fibrosis Bronchial Epithelial Cells and Regulates the Inflammatory Response Induced by Pseudomonas aeruginosa. Front. Immunol. 9:2823. doi: 10.3389/fimmu.2018.02823
Received: 07 September 2018; Accepted: 15 November 2018;
Published: 29 November 2018.
Edited by:
Janos G. Filep, Université de Montréal, CanadaReviewed by:
Anne Hilgendorff, Ludwig Maximilian University of Munich, GermanySven Hammerschmidt, University of Greifswald, Germany
Copyright © 2018 Perra, Balloy, Foussignière, Moissenet, Petat, Mungrue, Touqui, Corvol, Chignard and Guillot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Loic Guillot, bG9pYy5ndWlsbG90QGluc2VybS5mcg==