
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 12 March 2018
Sec. Viral Immunology
Volume 9 - 2018 | https://doi.org/10.3389/fimmu.2018.00458
 María Ángeles Jiménez-Sousa†
María Ángeles Jiménez-Sousa† Isidoro Martínez†
Isidoro Martínez† Luz María Medrano
Luz María Medrano Amanda Fernández-Rodríguez
Amanda Fernández-Rodríguez Salvador Resino*
Salvador Resino*
People living with human immunodeficiency virus (HIV) infection typically have hypovitaminosis D, which is linked to a large number of pathologies, including immune disorders and infectious diseases. Vitamin D (VitD) is a key regulator of host defense against infections by activating genes and pathways that enhance innate and adaptive immunity. VitD mediates its biological effects by binding to the Vitamin D receptor (VDR), and activating and regulating multiple cellular pathways. Single nucleotide polymorphisms in genes from those pathways have been associated with protection from HIV-1 infection. High levels of VitD and VDR expression are also associated with natural resistance to HIV-1 infection. Conversely, VitD deficiency is linked to more inflammation and immune activation, low peripheral blood CD4+ T-cells, faster progression of HIV disease, and shorter survival time in HIV-infected patients. VitD supplementation and restoration to normal values in HIV-infected patients may improve immunologic recovery during combination antiretroviral therapy, reduce levels of inflammation and immune activation, and increase immunity against pathogens. Additionally, VitD may protect against the development of immune reconstitution inflammatory syndrome events, pulmonary tuberculosis, and mortality among HIV-infected patients. In summary, this review suggests that VitD deficiency may contribute to the pathogenesis of HIV infection. Also, VitD supplementation seems to reverse some alterations of the immune system, supporting the use of VitD supplementation as prophylaxis, especially in individuals with more severe VitD deficiency.
Vitamin D is a fat-soluble steroid synthesized from a cholesterol precursor (7-dehydrocholesterol), which has a chemical secosteroid structure (1). The major forms of VitD that are important to humans are VitD2 or ergocalciferol, synthesized from ergosterol in plants, and VitD3 or cholecalciferol synthesized naturally from cholesterol in animals (VitD3) (1, 2). They can be supplied to the body from the diet and VitD-fortified products, among other sources (1, 2). However, the main source of VitD for the human body is its synthesis in the skin. A flowchart describing VitD metabolism is represented in Figure 1.
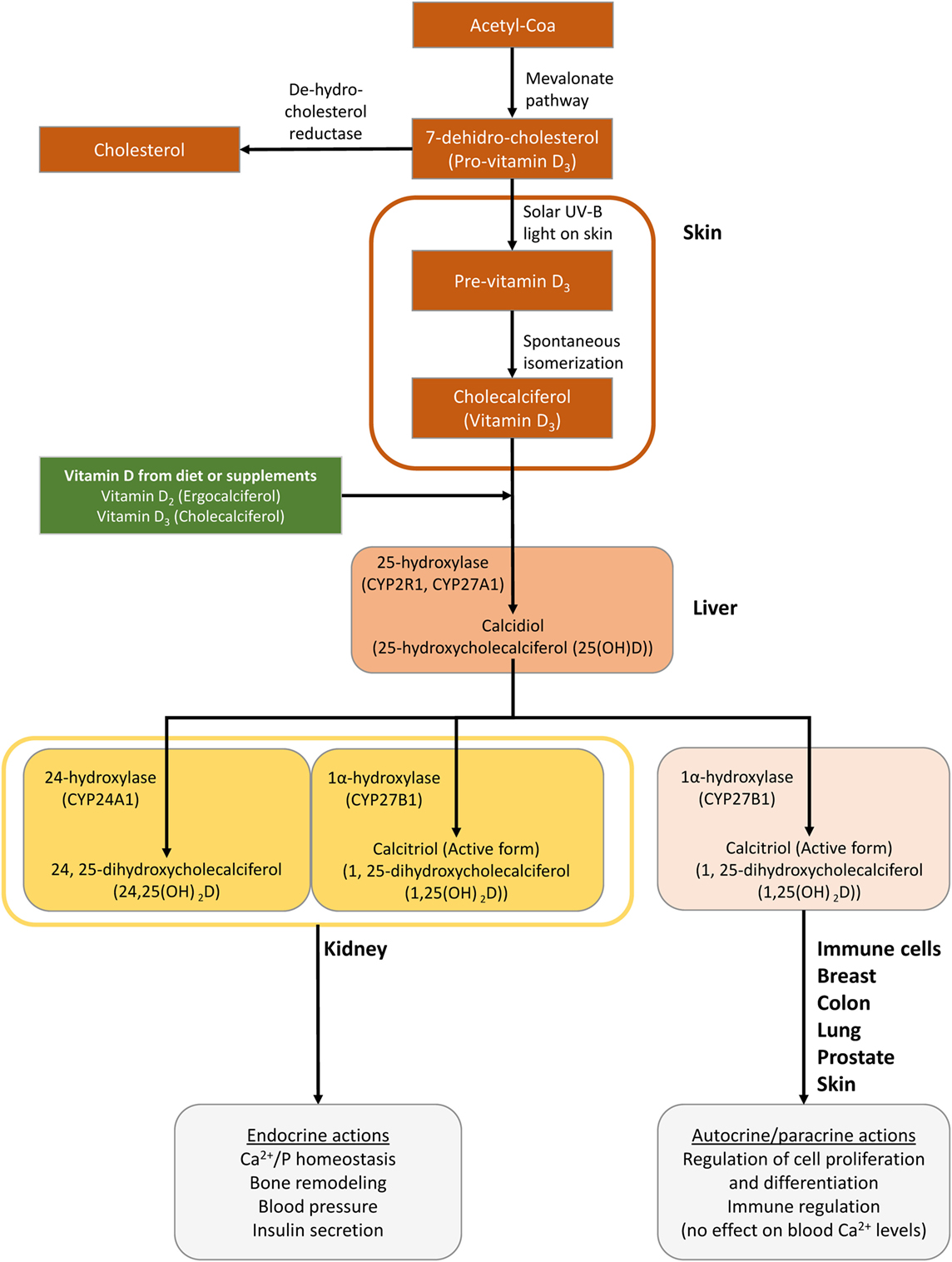
Figure 1. Schematic of the synthesis of vitamin D (VitD) in the body. Cutaneous 7-dihydrocholesterol is converted into preVitD3 following irradiation by ultraviolet light from the sun (2). Next, preVitD3 forms cholecalciferol (VitD3) by spontaneous isomerization. Subsequently, cholecalciferol is hydroxylated to 25-hydroxy-VitD (25(OH)D) or calcidiol, mainly in the liver, by the cytochrome P450 hydroxylase enzymes CYP27A1 and CYP2R1. Then, 25(OH)D is transported to the kidneys, where it is hydroxylated at the 1 alpha position by the 25-hydroxy-VitD-1 alpha hydroxylase (CYP27B1) to generate 1,25-dihydroxycholecalciferol [1,25 (OH)2D] or calcitriol, which is the metabolically active compound (1, 2). Hydroxyvitamin D-24-hydroxylase (CYP24A1) is the enzyme responsible for the multi-step catabolism of both 25(OH)D and 1,25 (OH)2D. The main product of 25(OH)D catabolism by CYP24A1 is 24,25-dihydroxycholecalciferol [24,25(OH)2D], which is less active than calcitriol and presumably represents a metabolite destined for excretion. Importantly, VitD is not only converted from 25(OH)D to 1,25 (OH)2D in the kidney but it is also activated locally by CYP27B1 in many tissues, including the brain, smooth muscle, breast, and prostate as well as cells of the immune system.
Transport and mechanism of action are shown in Figure 2A. A small fraction of VitD circulates in serum as “free” steroid and enters cells by simple diffusion. The remaining VitD in blood is transported bound to VitD-binding protein (DBP) (1, 2), which is able to bind the various types of VitD, albeit with different affinities. While DBP has a strong affinity for 25(OH)D, it has a weak affinity for 1,25(OH)2D. This low affinity together with the high affinity of the Vitamin D receptor (VDR) for 1,25(OH)2D makes 1,25(OH)2D the only ligand with direct access to the transcriptional signal transduction machinery (3). VitD binds to VDR in the nucleus, forming a complex with retinoic acid X receptor (RXR) and promotes gene transcription of several target genes by binding to VitD response elements (VDREs) (4). However, VitD can also regulate gene transcription via other mechanisms not related to VDREs. Additionally, VitD can enter the cell by binding to VDR situated on the cell membrane (VDRm), leading to non-genomics effects (5). The range of non-genomic effects is related to the cell-type and maturation status, but includes the modulation of growth factors and cytokines through cytosolic signaling pathways and effects on the activity of target transcription factors in the nucleus (5). Finally, VitD regulates its synthesis by a robust negative feedback mechanism (6).
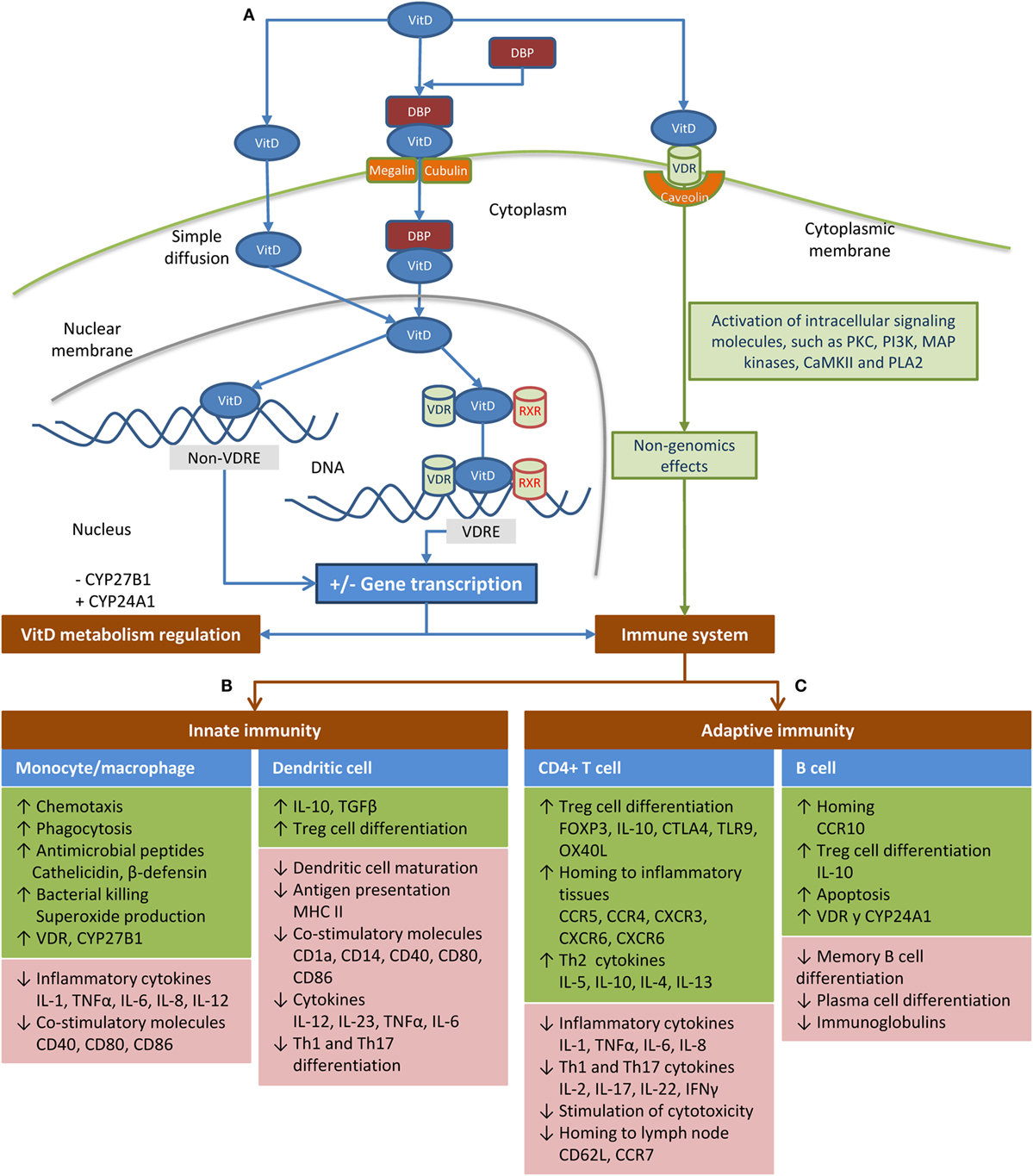
Figure 2. Schematic of transport and mechanism of action of vitamin D (VitD) in the body. (A) A small fraction of VitD circulates in the serum as a “free” steroid, having easy access to the intracellular compartment. The remaining VitD is transported in the blood while bound to the vitamin D-binding protein (DBP) (1, 2), which seems to critically regulate the bioavailability of VitD (7). This protein-bound fraction (bound to DBP) is actively transported into the cell by megalin or cubulin. Calcitriol is considered the main ligand of the vitamin D receptor (VDR) to trigger the effects of VitD, because its affinity is 1,000 times greater than calcidiol (8). When VitD binds to VDR in the nucleus of target cells, it forms a complex with the retinoic acid X receptor (RXR), which controls transcriptional activity of target genes. This heterodimer binds to VitD response elements (VDREs), a predefined promoter DNA sequence, initiating gene transcription processes, which covers around 5% of the human genome and 36 different cell types (4). However, there are genes regulated by VitD that do not contain VDREs (9). These genes may be regulated by microRNAs, phosphorylation, or other modifications of proteins, which affect their stability or the activity of proteases that target them (9). Additionally, non-genomic effects have been reported when the VDR is situated on the cell membrane (VDRm) complexed to caveolin (5), which immediately activates several intracellular pathways, such as mitogen-activated protein kinases, protein kinase C (PKC), protein kinase A, and Ca2+-calmodulin kinase II through the activation of several signaling molecules (5). VitD may reduce its synthesis by inhibiting CYP27B1 and increases its degradation by inducing CYP24A1 (6). (B) VitD modulates the function of monocytes/macrophages and dendritic cells (DCs) in response to infections. In monocytes/macrophages, 1,25(OH)2D leads to the expression of multi-target genes, among which are cathelicidin microbial peptide (10, 11), human β-defensin 4 (DEFB4) (12), and genes involved in autophagy and phagosome maturation, all of which are involved in the intracellular destruction of pathogens (7, 13). Furthermore, 1,25(OH)2D enhances the chemotactic and phagocytic capacity of macrophages (14). Moreover, VitD also promotes an anti-inflammatory response by inhibiting the maturation of DCs, resulting in a phenotype characterized by the downregulation of antigen presenting molecules (MHC-class II), costimulatory molecules (e.g., CD40, CD80, and CD86), and pro-inflammatory cytokines (e.g., IL-12 and IL-23); while an anti-inflammatory cytokine (IL-10) and T-cell inhibitory molecule (PD-1) are enhanced (15–22). Therefore, VitD induces hypo-responsiveness and allows a shift in the T-cell polarization from the pro-inflammatory Th1 and Th17 responses to a more tolerogenic Th2 response (16, 17, 20, 22–24), which leads to an altered alloreactive T cell activation (25). (C) VitD induces anti-inflammatory responses through direct effects on T-cells. Specifically, 1,25(OH)2D inhibits the proliferation of T-cells by blocking mitosis and IL-2 production (26, 27), limits the differentiation of Th1/Th17 cells, which favors Th2 differentiation (28–32), and induces the generation of IL-10 secretory Treg cells (32–34). Additionally, T-cell proliferation is significantly reduced when DCs are exposed to 1,25(OH)2D3 (16). T-cell cytokines also regulate VitD metabolism by monocytes. Thus, the Th1 cytokine IFN-γ induces CYP27B1, leading to the conversion of 25(OH)D to 1,25(OH)2D, whereas the Th2 cytokine IL-4 promotes upregulation of CYP24A1 (35). Stimulation of B-cells with 1,25(OH)2D leads to apoptosis, impaired plasma cell differentiation, decreased antibody production, inhibition of memory B-cell formation, and increased production of IL-10 (32, 36–41).
The VitD system plays a global role in many physiopathological processes since VDR is expressed in tissues and cells nearly throughout the entire organism. The tissues with the highest VDR content are the intestine, kidney, parathyroid gland, and bone, all of which are associated with maintenance of calcium homeostasis (42). Immune cells also express VDR, and they are capable of metabolizing circulating 25-hydroxy-VitD (25(OH)D) to the active form 1,25-dihydroxycholecalciferol [1,25(OH)2D], indicating a regulatory role of VitD in both the innate and adaptive immune systems (43, 44). Additionally, the effect of VitD on the immune response by binding to VDR is also present in many other cells, such as keratinocytes, bronchial/gastrointestinal epithelial cells, decidua, and trophoblastic cells (45).
The quantification of 25(OH)D in serum or plasma is the fastest and most accurate way to measure VitD levels in the body. However, this method has some drawbacks due to the hydrophobic nature of VitD, its high affinity to DBP, and the low concentration in blood (46).
The measurement of 25(OH)D levels is performed mainly via two different methodologies (46, 47): (a) competitive immunoassays, such as competitive protein-binding assays or radioimmunoassays, which do not differentiate between 25(OH)D2 and 25(OH)D3 isoforms; and (b) tests based on high-performance liquid chromatography and direct detection with liquid chromatography tandem-mass spectrometry (LC-MS/MS), which are highly sensitive and allow for the independent quantification of 25(OH)D2 and 25(OH)D3.
Vitamin D levels are expressed in nanogram per milliliter (ng/mL) or nanomol/liter (nmol/L). VitD deficiency in adults is considered to be when total 25(OH)D levels are <25 nmol/L (10 ng/mL) and inadequate/insufficient if levels are <75 nmol/L (30 ng/mL); while >75 nmol/L (30 ng/mL) is considered to be a normal healthy level (47, 48). Suboptimal VitD levels have been reported in rickets, osteomalacia, and non-skeletal diseases. There is wide variability in the prevalence of VitD deficiency across different patient groups. Regarding human immunodeficiency virus (HIV), conflicting data have been found. Some authors have described that there is no evidence of higher VitD deficiency in HIV-infected patients compared to non-HIV adults (49). However, others have described that VitD deficiency was more prevalent in HIV-positive than in HIV-negative individuals (50).
There is a lack of standardization regarding reference materials and reference methods, which makes it difficult to compare results across different laboratories (47, 51). However, the accuracy of results as well as particular aspects of 25(OH)D and 1,25(OH)2D methods (linearity, specificity, and the effect of anticoagulants) are being assessed by the large group of experts comprising the VitD External Quality Assessment Scheme (DEQAS). DEQAS has a close link to the Vitamin D Standardization Program, which promotes the standardized laboratory measurement of 25(OH)D (52).
Vitamin D deficiency is a major public health problem around the world in all age groups, even in countries closer to the equator with adequate UV radiation and in industrialized countries where VitD is typically supplemented (53). The prevalence of VitD deficiency (<25 nmol/L) is between 5 and 15%, and hypovitaminosis D (<75 nmol/L) is from 50 to 75% in high-income countries (53). This deficiency is directly involved in bone pathologies (rickets, osteoporosis, and osteomalacia). Additionally, there is growing evidence supporting its association with many other “non-classical” disorders not related to the bones, such as cardiovascular disease, cancer, multiple sclerosis, metabolic disorders, and infectious diseases (54). In fact, VitD deficiency is related to an increased incidence and severity of Mycobacterium tuberculosis (TB), HIV, and hepatitis C virus (HCV) infection (55, 56).
In a recent review article, Mansueto et al. showed that the prevalence of VitD deficiency ranges from 70 to 85% in HIV-infected patients, based on a large number of epidemiological articles that reported data of hypovitaminosis D with varying thresholds and a broad geolocalization of patients (56). VitD deficiency may be due to different reasons in these patients. On the one hand, there are many non-HIV-related risk factors for VitD deficiency, such as sex (females have higher risk), advanced age, limited sunlight exposure, skin pigmentation, black ethnicity, low levels of dietary VitD intake, gastrointestinal absorption disorders, liver and kidney diseases, higher body mass index, diabetes mellitus, and alcohol consumption (13, 56). These risk factors affect both HIV-positive and HIV-negative cohorts in a similar manner (57, 58). On the other hand, several HIV-related factors may lead to VitD deficiency. HIV infection itself leads to chronic inflammation and immune activation, and patients with VitD deficiency have been found to have increased IL6 and TNFα levels as well as activated monocyte phenotypes (59). Additionally, chronic inflammation may be responsible for impaired 1α-hydroxylase activity in the kidneys, resulting in reduced production of 1,25(OH)2D by blocking the PTH-stimulated conversion of 25(OH)D to 1,25(OH)2D (56, 60). Additionally, comorbidities, infectious complications, and hospitalizations of HIV-infected patients lead to reduced sun exposure, malnutrition, and diminished oral intake of VitD-rich foods (56, 61). In this regard, injection drug users infected with HIV suffer a disproportionate burden of VitD deficiency since they often have poor nutrition, limited and delayed access to health care, and a higher prevalence of comorbidities and infectious diseases (62). Finally, protease inhibitors (PIs) and non-nucleoside reverse transcriptase inhibitors (NNRTIs) seem to have an impact on VitD metabolic pathways. PIs seem to reduce 25(OH)D conversion to 1,25(OH)2D and NNRTIs seem to increase 25(OH)D catabolism, since low 25(OH)D levels have been seen in patients treated with these drugs (56).
Several single nucleotide polymorphisms (SNPs) in DBP gene seem to influence plasma levels of VitD (63, 64). DBP gene variants associated with reduced 25(OH)D levels were also associated with reduced DBP levels. In this setting, it has been hypothesized that altered DBP levels could affect the delivery of 1,25(OH)2D to target tissues, as well as the removal of VitD metabolites from circulation (63). A significant association has been found between rs222020 and rs2282679, which are in low linkage disequilibrium (LD), and the variation of serum 25(OH)D levels in healthy populations (64). Besides, rs222020 G allele, which has been associated with VitD deficiency, has been related to unfavorable outcome in HIV infection (65). Moreover, many SNPs located in genes related to the VitD pathway (DHCR7, CYP2R1, CYP3A4, CYP27A1, DBP, LRP2, CUB, CYP27B1, CYP24A1, VDR, and RXRA) are linked to a large number of non-skeletal health problems, especially infectious and autoimmune-related diseases (66). For example, the VDR rs1544410 G allele is related to the delayed progression of acquired immunedeficiency syndrome (AIDS) and increased resistance to HIV infection, which appears to be related to an increased response to VitD (67, 68). Additionally, VDR haplotypes conformed by rs11568820, rs4516035, rs10735810, rs1544410, and rs17878969 polymorphisms are also associated with protection against HIV infection. In this context, the protective haplotype has been related to a lower efficiency of VitD signaling, suggesting that an altered VitD pathway confers protection against HIV transmission. The exact mechanism of these polymorphisms is unclear. However, it is thought that VDR rs1544410 could reduce VDR messenger RNA production and stability, rs10735810 T allele might lead to a low transactivation capacity of the VDR protein, preventing normal VDR function, and VDR promoter rs4516035 could be biologically crucial to the immune system (69). Additionally, it is important to take into account that unknown functional genetic variants in LD with the known ones could be responsible for the regulation of serum 25(OH)D levels and the disease outcome.
Vitamin D is involved in host defense via an autocrine pathway in human monocytes and macrophages following stimulation of toll-like receptors (TLRs)2/1, TLR4, the receptor of interferon gamma (IFN-γ) or CD40 (70, 71). These receptors initiate a signaling cascade that induces upregulation of the VDR and CYP27B1, which leads to the conversion of 25(OH)D to 1,25(OH)2D. Binding of 1,25(OH)2D to the VDR leads to the expression of multitarget genes, which modulate the function of monocytes/macrophages during infection (see Figure 2B). Moreover, VitD prevents the excessive inflammatory response to infectious diseases by inhibiting the maturation of dendritic cells (DCs) (7, 13). Note that DCs also express VDR, as well as CYP27A1 and CYP27B1, thereby generating locally bioactive 1,25(OH)2D (72, 73). However, Kundu et al. described that human monocyte-derived DCs convert 25(OH)D to 1,25(OH)2D significantly less than macrophages, likely due to the fact that DCs mostly express a truncated CYP27B1 transcript, which may lead to a deficiency in VitD activation (74).
High levels of VitD and its receptor seem to be associated with a natural resistance to HIV-1 infection. This may stem from the upregulation of anti-inflammatory IL-10 and induction of anti-HIV-1 defensins in mucosa of HIV-1-exposed seronegative individuals (75). In addition, the expression of VDR is positively correlated with the expression of several anti-HIV molecules [such as elafin, TRIM5, cathelicidin microbial peptide (CAMP), HAD-4, and RNase7], which are linked to natural resistance to HIV-1 infection (76). Furthermore, it has been described that exogenous 1,25(OH)2D in monocytes decreases susceptibility to HIV infection by inhibiting viral entry, reducing surface CD4 expression and limiting monocyte proliferation (77, 78). Also, it has been demonstrated that TLR8 agonists inhibit HIV infection through a VitD- and CAMP-dependent autophagic mechanism in human macrophages (79). Furthermore, it has also been shown that VitD triggering autophagy in macrophages significantly inhibits replication of HIV-1 in a dose-dependent manner (80). However, there are other studies with contradictory data, in which VitD was found to increase HIV replication in monocytes both from patients and cell line clones (81–83).
In contrast, VitD deficiency is associated with greater inflammation (upregulation of CXCL10, IL-6, TNF-α, and D-dimer) and activated monocyte phenotypes (CX3CR1+ and CCR2+) in HIV-infected patients (55, 59, 84), which have been related to tissue dysfunction, comorbidity development, AIDS progression, and death in HIV-infected people (85, 86). In addition, chronic inflammation may also induce hypovitaminosis D (7). Thus, inflammatory processes involved in the appearance and clinical course of disease may reduce 25(OH)D levels, which would explain the low VitD status in a wide range of disorders in the general population (54) and in HIV-infected patients (7, 56). Legeai et al. found that severe VitD deficiency is associated with low CD4 counts and increased markers of inflammation in combination antiretroviral therapy (cART)-naïve HIV-infected patients (87). However, it is important to note that high 25(OH)D levels have also been recently associated with unexpectedly high levels of proinflammatory cytokines in HIV patients on cART therapy (88). Additionally, LPS and HIV gp120 upregulate the expression of CYP27B1 and CYP24A1 in monocytes and macrophages, leading to hypovitaminosis D in HIV-infected individuals and a reduction in mRNA expression of VDR and the antiviral peptides PI3 and CAMP (89, 90).
The restoration of VitD levels to normal values may minimize both ongoing inflammation and the complications of HIV and cART associated with chronic inflammation (13). For example, VitD supplementation stimulates expression of CAMP and improves antibacterial immunity in monocyte cultures and plasma from HIV-infected subjects (91, 92). VitD supplementation decreased markers of monocyte activation in HIV-infected patients (93). In vitro, VitD exposure improves the chemotactic activity of macrophages in AIDS patients (94). In addition, HIV-infected patients suffer from bacterial translocation, which is an effect of intestinal barrier damage caused by HIV itself (95). Recent studies have demonstrated the role of VitD in regulating host–bacteria interactions, intestinal innate immunity, and homeostasis (96). However, the rationale for VitD supplementation to reduce microbial translocation and systemic inflammation in the HIV population is controversial. For instance, no association between VitD levels and markers of microbial translocation was found in HIV-infected patients (97), while optimal VitD plasma levels have been associated with lower bacterial DNA translocation in HIV/HCV-coinfected patients (98). These differences between studies could be due to a high percentage of HIV/HCV-coinfected patients with advanced fibrosis/cirrhosis in the latter study, since severity of liver disease has been related to VitD deficiency (99) and bacterial translocation (100) in HIV/HCV-coinfected patients. The lack of consensus guidelines about VitD supplementation in HIV infection underlies the need for robust studies to critically evaluate the potential benefits of VitD supplementation in these patients.
Additionally, VitD has also been related to other infectious diseases in HIV-infected patients. VitD rescues impaired TB-mediated TNF release in macrophages of HIV-infected patients through an enhanced TLR signaling pathway (101). Additionally, high VitD levels have been associated with protection against the development of immune reconstitution inflammatory syndrome (IRIS) events (102) and decreased incidence of pulmonary tuberculosis and mortality among HIV-infected patients (103). However, a recent meta-analysis shows that there are other studies, particularly in HIV/TB-coinfected African patients receiving cART, which have not found any association between lower VitD levels and IRIS. Besides, they showed that VitD deficiency was not associated with an increased risk of TB in African HIV-infected patients (104). In regards to other infectious diseases, VitD levels have not been associated with better immune response to hepatitis B or pneumococcal vaccination in HIV-infected patients (105).
Vitamin D may indirectly affect T-cell responses via modulation of the DC phenotype and its stimulatory capacity toward T cells (106). Additionally, both naïve and resting memory T-cells express VDR at low levels, which suggests that VitD also acts directly on these T-cells (28, 107). T-cell activation increases the expression of VDR and CYP27B1, which allows 25(OH)D to be converted into 1,25(OH)2D to modulate effector functions of VitD (108). VitD suppresses the Th1, Th17, and Th2 profile of cytokine production, thereby altering T cell phenotype and function (106) (see Figure 2C). The effect of VitD on B-cells is thought to be the modulation of Th cells (106). However, human B-cells also express VDR and CYP27B1, which are upregulated upon activation, suggesting that B-cells may also be susceptible to 1,25(OH)2D stimulation. VitD induces hypo-responsiveness of B-cells and interferes with human plasma cell differentiation (106) (see Figure 2C).
Vitamin D induces antiviral gene expression, reduces the viral co-receptor CCR5 on CD4+ T-cells, and promotes an HIV-1-restrictive CD38+HLA-DR+ immunophenotype in in vitro assays, leading to HIV-1 infection inhibition in T cells (109). Likewise, VitD reduces the ability of TNFα to upregulate the transcription of HIV RNA from latently infected CD4+ cells (110). Thus, low levels of VitD are related to high HIV viral load in plasma (111–113), decreased CD4+ T-cells in peripheral blood (114, 115), rapid AIDS progression, and lower survival in HIV-infected patients (61, 84, 116–118).
Vitamin D deficiency seems to have no influence on T-cell and B-cell subset distribution in HIV-infected patients, but VitD supplementation is related to reduced immune activation levels (CD8+CD38+ and CD8+Ki67+) (119), and increased frequencies of antigen-specific T-cells expressing macrophage inflammatory protein-1β, an important anti-HIV blocking chemokine (91). Besides, Eckard et al. reported that VitD supplementation reduces immune activation and T-cell exhaustion, serving as an adjuvant therapy to cART in HIV-infected patients (93). However, high levels of 1,25(OH)2D have been associated with an expansion of activated CD4+ cells and Tregs in HIV-infected patients (120). In addition, high levels of 1,25(OH)2D increase the percentage of CD4+ T-cells expressing TIM-3 protein (a Th1-cell inhibitor and anti-HIV-1 molecule) in in vitro assays with peripheral blood mononuclear cells of HESN (89).
Vitamin D deficiency is also related to impaired CD4+ T-cell count recovery in HIV-positive patients on cART (121–123). Moreover, VitD levels are positively associated with CD4+ T-cell recovery after 24 weeks of VitD supplementation (115), suggesting a potential benefit of VitD supplementation on immunologic recovery during cART. However, there are other reports that showed no effect of VitD supplementation on CD4+ T-cells in HIV-infected adults (116, 124) or children (125).
Vitamin D deficiency may contribute to the pathogenesis of HIV infection by negatively modulating the innate and adaptive immune responses. Low VitD levels promote inflammation and activation of the immune system, which could increase the risk of non-AIDS-related comorbidity and mortality in the HIV-infected population. Moreover, exogenous VitD supplementation could be used as prophylaxis, as it appears to reverse some alterations of the immune system due to VitD deficiency, especially in individuals with more severe deficiency. Finally, there is a paucity of studies exploring the association between VitD levels and HIV infection.
Conceptualization, visualization, and supervision: SR. Resources and data curation: MJ-S, AF-R, LM, and IM. Writing—original draft preparation: MJ-S, IM, and SR. Writing—review and editing: AF-R and LM.
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
This work has been supported by grants given by Fondo de Investigación de Sanidad en España (FIS) (Spanish Health Founds for Research) (grant number PI14CIII/00011 and PI15CIII/00024). MJ-S, AF-R, and LM are supported by “Instituto de Salud Carlos III” (grant numbers CD13/00013, CP14CIII/00010, and CD14/00002, respectively).
1. Herrmann M, Farrell CL, Pusceddu I, Fabregat-Cabello N, Cavalier E. Assessment of vitamin D status – a changing landscape. Clin Chem Lab Med (2017) 55(1):3–26. doi:10.1515/cclm-2016-0264
2. Dusso AS, Brown AJ, Slatopolsky E. Vitamin D. Am J Physiol Renal Physiol (2005) 289(1):F8–28. doi:10.1152/ajprenal.00336.2004
3. Alshahrani F, Aljohani N. Vitamin D: deficiency, sufficiency and toxicity. Nutrients (2013) 5(9):3605–16. doi:10.3390/nu5093605
4. Hossein-nezhad A, Holick MF. Vitamin D for health: a global perspective. Mayo Clin Proc (2013) 88(7):720–55. doi:10.1016/j.mayocp.2013.05.011
5. Hii CS, Ferrante A. The non-genomic actions of vitamin D. Nutrients (2016) 8(3):135. doi:10.3390/nu8030135
6. Pike JW, Meyer MB. The vitamin D receptor: new paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Rheum Dis Clin North Am (2012) 38(1):13–27. doi:10.1016/j.rdc.2012.03.004
7. Watkins RR, Lemonovich TL, Salata RA. An update on the association of vitamin D deficiency with common infectious diseases. Can J Physiol Pharmacol (2015) 93(5):363–8. doi:10.1139/cjpp-2014-0352
8. Tsoukas CD, Provvedini DM, Manolagas SC. 1,25-dihydroxyvitamin D3: a novel immunoregulatory hormone. Science (1984) 224(4656):1438–40. doi:10.1126/science.6427926
9. Larriba MJ, Gonzalez-Sancho JM, Bonilla F, Munoz A. Interaction of vitamin D with membrane-based signaling pathways. Front Physiol (2014) 5:60. doi:10.3389/fphys.2014.00060
10. Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, et al. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science (2006) 311(5768):1770–3. doi:10.1126/science.1123933
11. Gombart AF, Borregaard N, Koeffler HP. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J (2005) 19(9):1067–77. doi:10.1096/fj.04-3284com
12. Liu PT, Schenk M, Walker VP, Dempsey PW, Kanchanapoomi M, Wheelwright M, et al. Convergence of IL-1beta and VDR activation pathways in human TLR2/1-induced antimicrobial responses. PLoS One (2009) 4(6):e5810. doi:10.1371/journal.pone.0005810
13. Gois PHF, Ferreira D, Olenski S, Seguro AC. Vitamin D and infectious diseases: simple bystander or contributing factor? Nutrients (2017) 9(7):E651. doi:10.3390/nu9070651
14. Taylor AE, Finney-Hayward TK, Quint JK, Thomas CM, Tudhope SJ, Wedzicha JA, et al. Defective macrophage phagocytosis of bacteria in COPD. Eur Respir J (2010) 35(5):1039–47. doi:10.1183/09031936.00036709
15. Lyakh LA, Sanford M, Chekol S, Young HA, Roberts AB. TGF-beta and vitamin D3 utilize distinct pathways to suppress IL-12 production and modulate rapid differentiation of human monocytes into CD83+ dendritic cells. J Immunol (2005) 174(4):2061–70. doi:10.4049/jimmunol.174.4.2061
16. Ferreira GB, van Etten E, Verstuyf A, Waer M, Overbergh L, Gysemans C, et al. 1,25-dihydroxyvitamin D3 alters murine dendritic cell behaviour in vitro and in vivo. Diabetes Metab Res Rev (2011) 27(8):933–41. doi:10.1002/dmrr.1275
17. Ferreira GB, Kleijwegt FS, Waelkens E, Lage K, Nikolic T, Hansen DA, et al. Differential protein pathways in 1,25-dihydroxyvitamin d(3) and dexamethasone modulated tolerogenic human dendritic cells. J Proteome Res (2012) 11(2):941–71. doi:10.1021/pr200724e
18. van Halteren AG, van Etten E, de Jong EC, Bouillon R, Roep BO, Mathieu C. Redirection of human autoreactive T-cells Upon interaction with dendritic cells modulated by TX527, an analog of 1,25 dihydroxyvitamin D(3). Diabetes (2002) 51(7):2119–25. doi:10.2337/diabetes.51.7.2119
19. Gauzzi MC, Purificato C, Donato K, Jin Y, Wang L, Daniel KC, et al. Suppressive effect of 1alpha,25-dihydroxyvitamin D3 on type I IFN-mediated monocyte differentiation into dendritic cells: impairment of functional activities and chemotaxis. J Immunol (2005) 174(1):270–6. doi:10.4049/jimmunol.174.1.270
20. Penna G, Adorini L. 1 alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol (2000) 164(5):2405–11. doi:10.4049/jimmunol.164.5.2405
21. Pedersen AW, Holmstrom K, Jensen SS, Fuchs D, Rasmussen S, Kvistborg P, et al. Phenotypic and functional markers for 1alpha,25-dihydroxyvitamin D(3)-modified regulatory dendritic cells. Clin Exp Immunol (2009) 157(1):48–59. doi:10.1111/j.1365-2249.2009.03961.x
22. Unger WW, Laban S, Kleijwegt FS, van der Slik AR, Roep BO. Induction of Treg by monocyte-derived DC modulated by vitamin D3 or dexamethasone: differential role for PD-L1. Eur J Immunol (2009) 39(11):3147–59. doi:10.1002/eji.200839103
23. van Etten E, Dardenne O, Gysemans C, Overbergh L, Mathieu C. 1,25-dihydroxyvitamin D3 alters the profile of bone marrow-derived dendritic cells of NOD mice. Ann N Y Acad Sci (2004) 1037:186–92. doi:10.1196/annals.1337.030
24. Ferreira GB, Gysemans CA, Demengeot J, da Cunha JP, Vanherwegen AS, Overbergh L, et al. 1,25-dihydroxyvitamin D3 promotes tolerogenic dendritic cells with functional migratory properties in NOD mice. J Immunol (2014) 192(9):4210–20. doi:10.4049/jimmunol.1302350
25. Penna G, Amuchastegui S, Giarratana N, Daniel KC, Vulcano M, Sozzani S, et al. 1,25-dihydroxyvitamin D3 selectively modulates tolerogenic properties in myeloid but not plasmacytoid dendritic cells. J Immunol (2007) 178(1):145–53. doi:10.4049/jimmunol.178.1.145
26. Alroy I, Towers TL, Freedman LP. Transcriptional repression of the interleukin-2 gene by vitamin D3: direct inhibition of NFATp/AP-1 complex formation by a nuclear hormone receptor. Mol Cell Biol (1995) 15(10):5789–99. doi:10.1128/MCB.15.10.5789
27. Rigby WF, Stacy T, Fanger MW. Inhibition of T lymphocyte mitogenesis by 1,25-dihydroxyvitamin D3 (calcitriol). J Clin Invest (1984) 74(4):1451–5. doi:10.1172/JCI111557
28. Jeffery LE, Wood AM, Qureshi OS, Hou TZ, Gardner D, Briggs Z, et al. Availability of 25-hydroxyvitamin D(3) to APCs controls the balance between regulatory and inflammatory T cell responses. J Immunol (2012) 189(11):5155–64. doi:10.4049/jimmunol.1200786
29. Ikeda U, Wakita D, Ohkuri T, Chamoto K, Kitamura H, Iwakura Y, et al. 1alpha,25-Dihydroxyvitamin D3 and all-trans retinoic acid synergistically inhibit the differentiation and expansion of Th17 cells. Immunol Lett (2010) 134(1):7–16. doi:10.1016/j.imlet.2010.07.002
30. Palmer MT, Lee YK, Maynard CL, Oliver JR, Bikle DD, Jetten AM, et al. Lineage-specific effects of 1,25-dihydroxyvitamin D(3) on the development of effector CD4 T cells. J Biol Chem (2011) 286(2):997–1004. doi:10.1074/jbc.M110.163790
31. Boonstra A, Barrat FJ, Crain C, Heath VL, Savelkoul HF, O’Garra A. 1alpha,25-Dihydroxyvitamin d3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J Immunol (2001) 167(9):4974–80. doi:10.4049/jimmunol.167.9.4974
32. Terrier B, Derian N, Schoindre Y, Chaara W, Geri G, Zahr N, et al. Restoration of regulatory and effector T cell balance and B cell homeostasis in systemic lupus erythematosus patients through vitamin D supplementation. Arthritis Res Ther (2012) 14(5):R221. doi:10.1186/ar4060
33. Jeffery LE, Burke F, Mura M, Zheng Y, Qureshi OS, Hewison M, et al. 1,25-dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J Immunol (2009) 183(9):5458–67. doi:10.4049/jimmunol.0803217
34. Baeke F, Korf H, Overbergh L, Verstuyf A, Thorrez L, Van Lommel L, et al. The vitamin D analog, TX527, promotes a human CD4+CD25highCD127low regulatory T cell profile and induces a migratory signature specific for homing to sites of inflammation. J Immunol (2011) 186(1):132–42. doi:10.4049/jimmunol.1000695
35. Edfeldt K, Liu PT, Chun R, Fabri M, Schenk M, Wheelwright M, et al. T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism. Proc Natl Acad Sci U S A (2010) 107(52):22593–8. doi:10.1073/pnas.1011624108
36. Chen S, Sims GP, Chen XX, Gu YY, Chen S, Lipsky PE. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J Immunol (2007) 179(3):1634–47. doi:10.4049/jimmunol.179.3.1634
37. Lemire JM, Adams JS, Sakai R, Jordan SC. 1 alpha,25-dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest (1984) 74(2):657–61. doi:10.1172/JCI111465
38. Provvedini DM, Tsoukas CD, Deftos LJ, Manolagas SC. 1 alpha,25-Dihydroxyvitamin D3-binding macromolecules in human B lymphocytes: effects on immunoglobulin production. J Immunol (1986) 136(8):2734–40.
39. Chen WC, Vayuvegula B, Gupta S. 1,25-Dihydroxyvitamin D3-mediated inhibition of human B cell differentiation. Clin Exp Immunol (1987) 69(3):639–46.
40. Iho S, Takahashi T, Kura F, Sugiyama H, Hoshino T. The effect of 1,25-dihydroxyvitamin D3 on in vitro immunoglobulin production in human B cells. J Immunol (1986) 136(12):4427–31.
41. Heine G, Niesner U, Chang HD, Steinmeyer A, Zugel U, Zuberbier T, et al. 1,25-dihydroxyvitamin D(3) promotes IL-10 production in human B cells. Eur J Immunol (2008) 38(8):2210–8. doi:10.1002/eji.200838216
42. Wang Y, Zhu J, DeLuca HF. Where is the vitamin D receptor? Arch Biochem Biophys (2012) 523(1):123–33. doi:10.1016/j.abb.2012.04.001
43. Hansdottir S, Monick MM, Hinde SL, Lovan N, Look DC, Hunninghake GW. Respiratory epithelial cells convert inactive vitamin D to its active form: potential effects on host defense. J Immunol (2008) 181(10):7090–9. doi:10.4049/jimmunol.181.10.7090
44. Veldman CM, Cantorna MT, DeLuca HF. Expression of 1,25-dihydroxyvitamin D(3) receptor in the immune system. Arch Biochem Biophys (2000) 374(2):334–8. doi:10.1006/abbi.1999.1605
45. Dimitrov V, White JH. Species-specific regulation of innate immunity by vitamin D signaling. J Steroid Biochem Mol Biol (2016) 164:246–53. doi:10.1016/j.jsbmb.2015.09.016
46. Hollis BW. Assessment and interpretation of circulating 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D in the clinical environment. Endocrinol Metab Clin North Am (2010) 39(2):271–86. doi:10.1016/j.ecl.2010.02.012
47. Holick MF. Vitamin D status: measurement, interpretation, and clinical application. Ann Epidemiol (2009) 19(2):73–8. doi:10.1016/j.annepidem.2007.12.001
48. SACoN (SACN). Update on Vitamin D: Position Statement by the Scientific Advisory Committee on Nutrition (2007).
49. Orkin C, Wohl DA, Williams A, Deckx H. Vitamin D deficiency in HIV: a shadow on long-term management? AIDS Rev (2014) 16(2):59–74.
50. Cervero M, Agud JL, Garcia-Lacalle C, Alcazar V, Torres R, Jusdado JJ, et al. Prevalence of vitamin D deficiency and its related risk factor in a Spanish cohort of adult HIV-infected patients: effects of antiretroviral therapy. AIDS Res Hum Retroviruses (2012) 28(9):963–71. doi:10.1089/AID.2011.0244
51. Roth HJ, Schmidt-Gayk H, Weber H, Niederau C. Accuracy and clinical implications of seven 25-hydroxyvitamin D methods compared with liquid chromatography-tandem mass spectrometry as a reference. Ann Clin Biochem (2008) 45(Pt 2):153–9. doi:10.1258/acb.2007.007091
52. Binkley N, Carter GD. Toward clarity in clinical vitamin D status assessment: 25(OH)D assay standardization. Endocrinol Metab Clin North Am (2017) 46(4):885–99. doi:10.1016/j.ecl.2017.07.012
53. Palacios C, Gonzalez L. Is vitamin D deficiency a major global public health problem? J Steroid Biochem Mol Biol (2014) 144(Pt A):138–45. doi:10.1016/j.jsbmb.2013.11.003
54. Autier P, Boniol M, Pizot C, Mullie P. Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol (2014) 2(1):76–89. doi:10.1016/S2213-8587(13)70165-7
55. Lucas RM, Gorman S, Geldenhuys S, Hart PH. Vitamin D and immunity. F1000Prime Rep (2014) 6:118. doi:10.12703/P6-118
56. Mansueto P, Seidita A, Vitale G, Gangemi S, Iaria C, Cascio A. Vitamin D deficiency in HIV infection: not only a bone disorder. Biomed Res Int (2015) 2015:735615. doi:10.1155/2015/735615
57. Havers F, Smeaton L, Gupte N, Detrick B, Bollinger RC, Hakim J, et al. 25-Hydroxyvitamin D insufficiency and deficiency is associated with HIV disease progression and virological failure post-antiretroviral therapy initiation in diverse multinational settings. J Infect Dis (2014) 210(2):244–53. doi:10.1093/infdis/jiu259
58. Dao CN, Patel P, Overton ET, Rhame F, Pals SL, Johnson C, et al. Low vitamin D among HIV-infected adults: prevalence of and risk factors for low vitamin D Levels in a cohort of HIV-infected adults and comparison to prevalence among adults in the US general population. Clin Infect Dis (2011) 52(3):396–405. doi:10.1093/cid/ciq158
59. Manion M, Hullsiek KH, Wilson EMP, Rhame F, Kojic E, Gibson D, et al. Vitamin D deficiency is associated with IL-6 levels and monocyte activation in HIV-infected persons. PLoS One (2017) 12(5):e0175517. doi:10.1371/journal.pone.0175517
60. Lake JE, Adams JS. Vitamin D in HIV-infected patients. Curr HIV/AIDS Rep (2011) 8(3):133–41. doi:10.1007/s11904-011-0082-8
61. Viard JP, Souberbielle JC, Kirk O, Reekie J, Knysz B, Losso M, et al. Vitamin D and clinical disease progression in HIV infection: results from the EuroSIDA study. AIDS (2011) 25(10):1305–15. doi:10.1097/QAD.0b013e328347f6f7
62. Lambert AA, Drummond MB, Mehta SH, Brown TT, Lucas GM, Kirk GD, et al. Risk factors for vitamin D deficiency among HIV-infected and uninfected injection drug users. PLoS One (2014) 9(4):e95802. doi:10.1371/journal.pone.0095802
63. Wang TJ, Zhang F, Richards JB, Kestenbaum B, van Meurs JB, Berry D, et al. Common genetic determinants of vitamin D insufficiency: a genome-wide association study. Lancet (2010) 376(9736):180–8. doi:10.1016/S0140-6736(10)60588-0
64. Bu FX, Armas L, Lappe J, Zhou Y, Gao G, Wang HW, et al. Comprehensive association analysis of nine candidate genes with serum 25-hydroxy vitamin D levels among healthy Caucasian subjects. Hum Genet (2010) 128(5):549–56. doi:10.1007/s00439-010-0881-9
65. Porter TR, Li X, Stephensen CB, Mulligan K, Rutledge B, Flynn PM, et al. Genetic associations with 25-hydroxyvitamin D deficiency in HIV-1-infected youth: fine-mapping for the GC/DBP gene that encodes the vitamin D-binding protein. Front Genet (2013) 4:234. doi:10.3389/fgene.2013.00234
66. Jolliffe DA, Walton RT, Griffiths CJ, Martineau AR. Single nucleotide polymorphisms in the vitamin D pathway associating with circulating concentrations of vitamin D metabolites and non-skeletal health outcomes: review of genetic association studies. J Steroid Biochem Mol Biol (2016) 164:18–29. doi:10.1016/j.jsbmb.2015.12.007
67. Moodley A, Qin M, Singh KK, Spector SA. Vitamin D-related host genetic variants alter HIV disease progression in children. Pediatr Infect Dis J (2013) 32(11):1230–6. doi:10.1097/INF.0b013e31829e4d06
68. Torres C, Sanchez de la Torre M, Garcia-Moruja C, Carrero AJ, Trujillo Mdel M, Fibla J, et al. Immunophenotype of vitamin D receptor polymorphism associated to risk of HIV-1 infection and rate of disease progression. Curr HIV Res (2010) 8(6):487–92. doi:10.2174/157016210793499330
69. de la Torre MS, Torres C, Nieto G, Vergara S, Carrero AJ, Macias J, et al. Vitamin D receptor gene haplotypes and susceptibility to HIV-1 infection in injection drug users. J Infect Dis (2008) 197(3):405–10. doi:10.1086/525043
70. Kroner Jde C, Sommer A, Fabri M. Vitamin D every day to keep the infection away? Nutrients (2015) 7(6):4170–88. doi:10.3390/nu7064170
71. Adams JS, Ren S, Liu PT, Chun RF, Lagishetty V, Gombart AF, et al. Vitamin d-directed rheostatic regulation of monocyte antibacterial responses. J Immunol (2009) 182(7):4289–95. doi:10.4049/jimmunol.0803736
72. Hata M, Takahara S, Tsuzaki H, Ishii Y, Nakata K, Akagawa KS, et al. Expression of Th2-skewed pathology mediators in monocyte-derived type 2 of dendritic cells (DC2). Immunol Lett (2009) 126(1–2):29–36. doi:10.1016/j.imlet.2009.07.008
73. Fritsche J, Mondal K, Ehrnsperger A, Andreesen R, Kreutz M. Regulation of 25-hydroxyvitamin D3-1 alpha-hydroxylase and production of 1 alpha,25-dihydroxyvitamin D3 by human dendritic cells. Blood (2003) 102(9):3314–6. doi:10.1182/blood-2002-11-3521
74. Kundu R, Chain BM, Coussens AK, Khoo B, Noursadeghi M. Regulation of CYP27B1 and CYP24A1 hydroxylases limits cell-autonomous activation of vitamin D in dendritic cells. Eur J Immunol (2014) 44(6):1781–90. doi:10.1002/eji.201344157
75. Aguilar-Jimenez W, Zapata W, Caruz A, Rugeles MT. High transcript levels of vitamin D receptor are correlated with higher mRNA expression of human beta defensins and IL-10 in mucosa of HIV-1-exposed seronegative individuals. PLoS One (2013) 8(12):e82717. doi:10.1371/journal.pone.0082717
76. Aguilar-Jimenez W, Zapata W, Rugeles MT. Antiviral molecules correlate with vitamin D pathway genes and are associated with natural resistance to HIV-1 infection. Microbes Infect (2016) 18(7–8):510–6. doi:10.1016/j.micinf.2016.03.015
77. Connor RI, Rigby WF. 1 alpha,25-dihydroxyvitamin D3 inhibits productive infection of human monocytes by HIV-1. Biochem Biophys Res Commun (1991) 176(2):852–9. doi:10.1016/S0006-291X(05)80264-5
78. Schuitemaker H, Kootstra NA, Koppelman MH, Bruisten SM, Huisman HG, Tersmette M, et al. Proliferation-dependent HIV-1 infection of monocytes occurs during differentiation into macrophages. J Clin Invest (1992) 89(4):1154–60. doi:10.1172/JCI115697
79. Campbell GR, Spector SA. Toll-like receptor 8 ligands activate a vitamin D mediated autophagic response that inhibits human immunodeficiency virus type 1. PLoS Pathog (2012) 8(11):e1003017. doi:10.1371/journal.ppat.1003017
80. Campbell GR, Spector SA. Hormonally active vitamin D3 (1alpha,25-dihydroxycholecalciferol) triggers autophagy in human macrophages that inhibits HIV-1 infection. J Biol Chem (2011) 286(21):18890–902. doi:10.1074/jbc.M110.206110
81. Kizaki M, Ikeda Y, Simon KJ, Nanjo M, Koeffler HP. Effect of 1,25-dihydroxyvitamin D3 and its analogs on human immunodeficiency virus infection in monocytes/macrophages. Leukemia (1993) 7(10):1525–30.
82. Biswas P, Mengozzi M, Mantelli B, Delfanti F, Brambilla A, Vicenzi E, et al. 1,25-Dihydroxyvitamin D3 upregulates functional CXCR4 human immunodeficiency virus type 1 coreceptors in U937 minus clones: NF-kappaB-independent enhancement of viral replication. J Virol (1998) 72(10):8380–3.
83. Skolnik PR, Jahn B, Wang MZ, Rota TR, Hirsch MS, Krane SM. Enhancement of human immunodeficiency virus 1 replication in monocytes by 1,25-dihydroxycholecalciferol. Proc Natl Acad Sci U S A (1991) 88(15):6632–6. doi:10.1073/pnas.88.15.6632
84. Shepherd L, Souberbielle JC, Bastard JP, Fellahi S, Capeau J, Reekie J, et al. Prognostic value of vitamin D level for all-cause mortality, and association with inflammatory markers, in HIV-infected persons. J Infect Dis (2014) 210(2):234–43. doi:10.1093/infdis/jiu074
85. Deeks SG, Lewin SR, Havlir DV. The end of AIDS: HIV infection as a chronic disease. Lancet (2013) 382(9903):1525–33. doi:10.1016/S0140-6736(13)61809-7
86. Deeks SG. HIV infection, inflammation, immunosenescence, and aging. Annu Rev Med (2011) 62:141–55. doi:10.1146/annurev-med-042909-093756
87. Legeai C, Vigouroux C, Souberbielle JC, Bouchaud O, Boufassa F, Bastard JP, et al. Associations between 25-hydroxyvitamin D and immunologic, metabolic, inflammatory markers in treatment-naive HIV-infected persons: the ANRS CO9 <<COPANA>> cohort study. PLoS One (2013) 8(9):e74868. doi:10.1371/journal.pone.0074868
88. Gangcuangco LM, Kohorn LB, Chow DC, Keating SM, Norris PJ, Nagamine LS, et al. High 25-hydroxyvitamin D is associated with unexpectedly high plasma inflammatory markers in HIV patients on antiretroviral therapy. Medicine (Baltimore) (2016) 95(43):e5270. doi:10.1097/MD.0000000000005270
89. Aguilar-Jimenez W, Saulle I, Trabattoni D, Vichi F, Lo Caputo S, Mazzotta F, et al. High expression of antiviral and vitamin D pathway genes are a natural characteristic of a small cohort of HIV-1-exposed seronegative individuals. Front Immunol (2017) 8:136. doi:10.3389/fimmu.2017.00136
90. Pinzone MR, Di Rosa M, Celesia BM, Condorelli F, Malaguarnera M, Madeddu G, et al. LPS and HIV gp120 modulate monocyte/macrophage CYP27B1 and CYP24A1 expression leading to vitamin D consumption and hypovitaminosis D in HIV-infected individuals. Eur Rev Med Pharmacol Sci (2013) 17(14):1938–50.
91. Lachmann R, Bevan MA, Kim S, Patel N, Hawrylowicz C, Vyakarnam A, et al. A comparative phase 1 clinical trial to identify anti-infective mechanisms of vitamin D in people with HIV infection. AIDS (2015) 29(10):1127–35. doi:10.1097/QAD.0000000000000666
92. Chun RF, Liu NQ, Lee T, Schall JI, Denburg MR, Rutstein RM, et al. Vitamin D supplementation and antibacterial immune responses in adolescents and young adults with HIV/AIDS. J Steroid Biochem Mol Biol (2015) 148:290–7. doi:10.1016/j.jsbmb.2014.07.013
93. Eckard AR, O’Riordan MA, Rosebush JC, Lee ST, Habib JG, Ruff JH, et al. Vitamin D supplementation decreases immune activation and exhaustion in HIV-1-infected youth. Antivir Ther (2017). doi:10.3851/IMP3199
94. Girasole G, Wang JM, Pedrazzoni M, Pioli G, Balotta C, Passeri M, et al. Augmentation of monocyte chemotaxis by 1 alpha,25-dihydroxyvitamin D3. Stimulation of defective migration of AIDS patients. J Immunol (1990) 145(8):2459–64.
95. Sandler NG, Douek DC. Microbial translocation in HIV infection: causes, consequences and treatment opportunities. Nat Rev Microbiol (2012) 10(9):655–66. doi:10.1038/nrmicro2848
96. Dimitrov V, White JH. Vitamin D signaling in intestinal innate immunity and homeostasis. Mol Cell Endocrinol (2017) 453:68–78. doi:10.1016/j.mce.2017.04.010
97. Missailidis C, Hoijer J, Johansson M, Ekstrom L, Bratt G, Hejdeman B, et al. Vitamin D status in well-controlled Caucasian HIV patients in relation to inflammatory and metabolic markers – a cross-sectional cohort study in Sweden. Scand J Immunol (2015) 82(1):55–62. doi:10.1111/sji.12299
98. Garcia-Alvarez M, Berenguer J, Jimenez-Sousa MA, Vazquez-Moron S, Carrero A, Gutierrez-Rivas M, et al. Optimal vitamin D plasma levels are associated with lower bacterial DNA translocation in HIV/hepatitis c virus coinfected patients. AIDS (2016) 30(7):1069–74. doi:10.1097/QAD.0000000000001007
99. Guzman-Fulgencio M, Garcia-Alvarez M, Berenguer J, Jimenez-Sousa MA, Cosin J, Pineda-Tenor D, et al. Vitamin D deficiency is associated with severity of liver disease in HIV/HCV coinfected patients. J Infect (2014) 68(2):176–84. doi:10.1016/j.jinf.2013.10.011
100. Garcia-Alvarez M, Berenguer J, Guzman-Fulgencio M, Alvarez E, Cosin J, Micheloud D, et al. Bacterial DNA translocation and liver disease severity among HIV-infected patients with chronic hepatitis C. J Acquir Immune Defic Syndr (2012) 61(5):552–6. doi:10.1097/QAI.0b013e31826ea109
101. Anandaiah A, Sinha S, Bole M, Sharma SK, Kumar N, Luthra K, et al. Vitamin D rescues impaired Mycobacterium tuberculosis-mediated tumor necrosis factor release in macrophages of HIV-seropositive individuals through an enhanced Toll-like receptor signaling pathway in vitro. Infect Immun (2013) 81(1):2–10. doi:10.1128/IAI.00666-12
102. Musselwhite LW, Andrade BB, Ellenberg SS, Tierney A, Belaunzaran-Zamudio PF, Rupert A, et al. Vitamin D, D-dimer, interferon gamma, and sCD14 levels are independently associated with immune reconstitution inflammatory syndrome: a prospective, international study. EBioMedicine (2016) 4:115–23. doi:10.1016/j.ebiom.2016.01.016
103. Sudfeld CR, Mugusi F, Aboud S, Nagu TJ, Wang M, Fawzi WW. Efficacy of vitamin D3 supplementation in reducing incidence of pulmonary tuberculosis and mortality among HIV-infected Tanzanian adults initiating antiretroviral therapy: study protocol for a randomized controlled trial. Trials (2017) 18(1):66. doi:10.1186/s13063-017-1819-5
104. Huang SJ, Wang XH, Liu ZD, Cao WL, Han Y, Ma AG, et al. Vitamin D deficiency and the risk of tuberculosis: a meta-analysis. Drug Des Devel Ther (2017) 11:91–102. doi:10.2147/DDDT.S79870
105. Viard JP, Assuied A, Levy Y, Souberbielle JC, Thiebaut R, Carrat F, et al. No positive association between vitamin D level and immune responses to hepatitis B and Streptococcus pneumoniae vaccination in HIV-infected adults. PLoS One (2016) 11(12):e0168640. doi:10.1371/journal.pone.0168640
106. Vanherwegen AS, Gysemans C, Mathieu C. Vitamin D endocrinology on the cross-road between immunity and metabolism. Mol Cell Endocrinol (2017) 453:52–67. doi:10.1016/j.mce.2017.04.018
107. Sigmundsdottir H, Pan J, Debes GF, Alt C, Habtezion A, Soler D, et al. DCs metabolize sunlight-induced vitamin D3 to ’program’ T cell attraction to the epidermal chemokine CCL27. Nat Immunol (2007) 8(3):285–93. doi:10.1038/ni1433
108. Baeke F, Korf H, Overbergh L, van Etten E, Verstuyf A, Gysemans C, et al. Human T lymphocytes are direct targets of 1,25-dihydroxyvitamin D3 in the immune system. J Steroid Biochem Mol Biol (2010) 121(1–2):221–7. doi:10.1016/j.jsbmb.2010.03.037
109. Aguilar-Jimenez W, Villegas-Ospina S, Gonzalez S, Zapata W, Saulle I, Garziano M, et al. Precursor forms of vitamin D reduce HIV-1 infection in vitro. J Acquir Immune Defic Syndr (2016) 73(5):497–506. doi:10.1097/QAI.0000000000001150
110. Nunnari G, Fagone P, Lazzara F, Longo A, Cambria D, Di Stefano G, et al. Vitamin D3 inhibits TNFalpha-induced latent HIV reactivation in J-LAT cells. Mol Cell Biochem (2016) 418(1–2):49–57. doi:10.1007/s11010-016-2732-z
111. Flauzino T, Simao ANC, de Almeida ERD, Morimoto HK, Oliveira SR, Alfieri DF, et al. Association between vitamin D status, oxidative stress biomarkers and viral load in human immunodeficiency virus type 1 infection. Curr HIV Res (2017) 15(5):336–44. doi:10.2174/1570162X15666171005170227
112. Coussens AK, Naude CE, Goliath R, Chaplin G, Wilkinson RJ, Jablonski NG. High-dose vitamin D3 reduces deficiency caused by low UVB exposure and limits HIV-1 replication in urban Southern Africans. Proc Natl Acad Sci U S A (2015) 112(26):8052–7. doi:10.1073/pnas.1500909112
113. Bearden A, Abad C, Gangnon R, Sosman JM, Binkley N, Safdar N. Cross-sectional study of vitamin D levels, immunologic and virologic outcomes in HIV-infected adults. J Clin Endocrinol Metab (2013) 98(4):1726–33. doi:10.1210/jc.2012-4031
114. Theodorou M, Serste T, Van Gossum M, Dewit S. Factors associated with vitamin D deficiency in a population of 2044 HIV-infected patients. Clin Nutr (2014) 33(2):274–9. doi:10.1016/j.clnu.2013.04.018
115. Coelho L, Cardoso SW, Luz PM, Hoffman RM, Mendonca L, Veloso VG, et al. Vitamin D3 supplementation in HIV infection: effectiveness and associations with antiretroviral therapy. Nutr J (2015) 14:81. doi:10.1186/s12937-015-0072-6
116. Sudfeld CR, Wang M, Aboud S, Giovannucci EL, Mugusi FM, Fawzi WW. Vitamin D and HIV progression among Tanzanian adults initiating antiretroviral therapy. PLoS One (2012) 7(6):e40036. doi:10.1371/journal.pone.0040036
117. Mehta S, Giovannucci E, Mugusi FM, Spiegelman D, Aboud S, Hertzmark E, et al. Vitamin D status of HIV-infected women and its association with HIV disease progression, anemia, and mortality. PLoS One (2010) 5(1):e8770. doi:10.1371/journal.pone.0008770
118. Haug C, Muller F, Aukrust P, Froland SS. Subnormal serum concentration of 1,25-vitamin D in human immunodeficiency virus infection: correlation with degree of immune deficiency and survival. J Infect Dis (1994) 169(4):889–93. doi:10.1093/infdis/169.4.889
119. Fabre-Mersseman V, Tubiana R, Papagno L, Bayard C, Briceno O, Fastenackels S, et al. Vitamin D supplementation is associated with reduced immune activation levels in HIV-1-infected patients on suppressive antiretroviral therapy. AIDS (2014) 28(18):2677–82. doi:10.1097/QAD.0000000000000472
120. Bang U, Kolte L, Hitz M, Dam Nielsen S, Schierbeck LL, Andersen O, et al. Correlation of increases in 1,25-dihydroxyvitamin D during vitamin D therapy with activation of CD4+ T lymphocytes in HIV-1-infected males. HIV Clin Trials (2012) 13(3):162–70. doi:10.1310/hct1303-162
121. Aziz M, Livak B, Burke-Miller J, French AL, Glesby MJ, Sharma A, et al. Vitamin D insufficiency may impair CD4 recovery among Women’s Interagency HIV Study participants with advanced disease on HAART. AIDS (2013) 27(4):573–8. doi:10.1097/QAD.0b013e32835b9ba1
122. Ezeamama AE, Guwatudde D, Wang M, Bagenda D, Kyeyune R, Sudfeld C, et al. Vitamin-D deficiency impairs CD4+T-cell count recovery rate in HIV-positive adults on highly active antiretroviral therapy: a longitudinal study. Clin Nutr (2016) 35(5):1110–7. doi:10.1016/j.clnu.2015.08.007
123. Ross AC, Judd S, Kumari M, Hileman C, Storer N, Labbato D, et al. Vitamin D is linked to carotid intima-media thickness and immune reconstitution in HIV-positive individuals. Antivir Ther (2011) 16(4):555–63. doi:10.3851/IMP1784
124. Giacomet V, Vigano A, Manfredini V, Cerini C, Bedogni G, Mora S, et al. Cholecalciferol supplementation in HIV-infected youth with vitamin D insufficiency: effects on vitamin D status and T-cell phenotype: a randomized controlled trial. HIV Clin Trials (2013) 14(2):51–60. doi:10.1310/hct1402-51
Keywords: vitamin D deficiency, human immunodeficiency virus, inflammation, immune activation, adaptive immunity, innate immunity
Citation: Jiménez-Sousa MÁ, Martínez I, Medrano LM, Fernández-Rodríguez A and Resino S (2018) Vitamin D in Human Immunodeficiency Virus Infection: Influence on Immunity and Disease. Front. Immunol. 9:458. doi: 10.3389/fimmu.2018.00458
Received: 01 December 2017; Accepted: 20 February 2018;
Published: 12 March 2018
Edited by:
Mario Clerici, Università degli Studi di Milano, ItalyReviewed by:
Anne L. Astier, UMR5282 Centre de Physiopathologie de Toulouse Purpan (CPTP), FranceCopyright: © 2018 Jiménez-Sousa, Martínez, Medrano, Fernández-Rodríguez and Resino. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Salvador Resino, c3Jlc2lub0Bpc2NpaWkuZXM=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.