
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 16 November 2017
Sec. T Cell Biology
Volume 8 - 2017 | https://doi.org/10.3389/fimmu.2017.01543
 Theresa Isabell Schindler1†
Theresa Isabell Schindler1† Johanna-Josophina Wagner1,2†
Johanna-Josophina Wagner1,2† Sybelle Goedicke-Fritz1,3*
Sybelle Goedicke-Fritz1,3* Tobias Rogosch1Verena Coccejus1Verena Laudenbach1Wilfried Nikolaizik1
Tobias Rogosch1Verena Coccejus1Verena Laudenbach1Wilfried Nikolaizik1 Christoph Härtel4Rolf Felix Maier1Sebastian Kerzel5‡
Christoph Härtel4Rolf Felix Maier1Sebastian Kerzel5‡ Michael Zemlin1,3‡
Michael Zemlin1,3‡
Background: The prevalence of obesity has dramatically increased in children in the last few decades and is associated with chronic inflammatory diseases. Fat tissue produces IL-6 and TNF-α, which are stimuli for TH17 cell differentiation. These cells are characterized by expression of the transcription factor receptor-related orphan receptor C (RORC) and by IL-17A production. In murine models, obesity has been linked with elevated TH17 cell frequencies. The aim of this study was to explore whether being overweight was associated with an elevated frequency of circulating TH17 cells or elevated messenger RNA (mRNA)-levels of IL-17A and RORC in children without chronic inflammatory diseases.
Methods: We studied peripheral blood samples from 15 overweight and 50 non-overweight children without a history of autoimmune diseases, asthma, atopic dermatitis or allergic rhinoconjunctivitis. TH17 cells were quantified in Ionomycin stimulated peripheral blood mononuclear cells by flow cytometry using intracellular IL-17A staining. RORC- and IL-17A expressions were measured by real-time PCR.
Results: We found significantly elevated TH cell frequencies in overweight children compared then on-overweight controls with 34.7 ± 1.5% of CD3+CD4+ cells versus 25.4 ± 2.4% (mean ± SEM, p = 0.0023), respectively. Moreover, TH cell frequencies correlated positively with body mass index (r = 0.42, p = 0.0005, respectively). The relative mRNA expression of RORC (p = 0.013) and IL-17A (p = 0.014) were upregulated in overweight compared to non-overweight children.
Conclusion: Childhood obesity is an independent factor that is associated with an elevated frequency of circulating TH17 cells and higher expression of RORC- and IL-17A-mRNA after in vitro stimulation with Ionomycin. This might be due to the inflammatory activity of the fat tissue. Studies on TH17 immunity should not only be adjusted for acute and chronic inflammatory diseases but also for overweight.
Being overweight not only affects adults by causing chronic diseases including type 2 diabetes, arterial hypertension, or coronary heart disease, but is also an increasingly widespread health issue in childhood (1, 2). In 2010, 43 million children worldwide were overweight and this number is likely to increase to 60 million by 2020 (3). Obesity is a risk factor for allergic diseases like bronchial asthma and for various autoimmune diseases (4, 5). It contributes to a low-grade chronic inflammation with higher numbers of macrophages, mast cells, neutrophils, B- and T-cells in adipose tissue (6, 7), but the underlying pathogenesis and complex mutual interactions between immunity and metabolism are not yet completely understood.
TH17 cells play a role in autoimmune and allergic inflammation (8–10). TH17 cells are characterized by retinoic acid receptor-related orphan receptor C (RORC) expression (11, 12) and IL-17A production which attracts and activates neutrophil granulocytes (13). Physiologically, TH17 cells seem to be primarily important for clearance of pathogens, which cannot be fought effectively by TH1 nor TH2 cells—for example, Klebsiella pneumoniae, Borrelia burgdorferi, and helminths or fungi (14, 15). On the other hand, TH17 cells contribute to various autoimmune and allergic diseases, for example, inflammatory bowel diseases or bronchial asthma (16).
Furthermore, there is growing evidence from animal models that TH17 cells might also contribute to a low-grade chronically sustained inflammation in obesity. Interestingly, TH17 cells were elevated in the spleens of mice with a diet-induced obesity compared to normal weight mice (17). Likewise, more TH17 cells were detected in adipose tissue of obese compared to normal weight mice (18). Łuczyński et al. found more TH17 cells in peripheral blood samples of children with central obesity and diabetes mellitus (19). However, to our knowledge, none of the published studies on TH17 cells in overweight was controlled for potentially coinciding chronic inflammatory disease like bronchial asthma or autoimmunity. Thus, it remains unknown whether elevated TH17 cells in overweight individuals are directly linked with overweight or are a consequence of associated chronic inflammatory diseases (20). In this study, we wanted to test the hypothesis that the TH17 cell frequency is elevated in the peripheral blood of overweight children without allergic or autoimmune disease.
The study was performed at the Department of Pediatrics, Philipps University, Marburg, and was approved by the local Ethics Committee. Written informed consent was obtained from parents and in older children additionally from themselves. Children admitted to the outpatient clinic because of adiposity or non-inflammatory conditions like headache, elective surgery, or abdominal pain were screened for eligibility. Children were excluded from our study when they were younger than 4 years or older than 17 years old, had asthma, atopic dermatitis, allergic rhinoconjunctivitis, or any autoimmune disease in their medical history. Symptoms of an acute infection during the previous 2 weeks led to exclusion as well as clinical symptoms of a current infection, current C-reactive protein elevation or leukocytosis. Being overweight was defined as a body mass index (BMI) ≥90th percentile, while children with a BMI < 90th percentile were included in our non-overweight control group. BMI percentiles were assessed with percentiles for German children published by Kromeyer-Hauschild et al. (21).
2.6 ml of venous blood were collected after informed consent during a routinely performed venipuncture or during routine placement of a peripheral venous catheter.
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque gradient (PAA, Linz, Austria) and were stimulated in a 12-well plate in X-VIVO 15 (X-VIVO 15 w/o Gentamycin and Phenol Red, Lonza, Velvier, Belgium) with Ionomycin (Sigma Aldrich Chemie, Steinheim, Germany) for 4 h (37°C, 5% CO2). Brefeldin A (eBioscience, San Diego, CA, USA) was added after 3 h. After staining these cells with anti-CD3 (FITC) and anti-CD4 (PE) antibodies (R&D Systems, Minneapolis, MN, USA), the PBMCs were fixated, permeabilized, and stained intracellularly with anti-IL-17A antibody (APC) (Flow Cytometry Fixation and Permeabilization Buffer Kit I, R&D Systems, Minneapolis, MN, USA).
Flow cytometry was performed on a 4-color FacsCalibur flow cytometer (BD Biosciences) using summit 4.3 software (Beckman Coulter, Krefeld, Germany). Gates were preset and the measurements were performed blinded for sample identity. TH17 cells were defined as CD3+CD4+IL-17Aic+ (22) and were evaluated as percentage of TH lymphocytes (CD3+CD4+) (Figure 1).
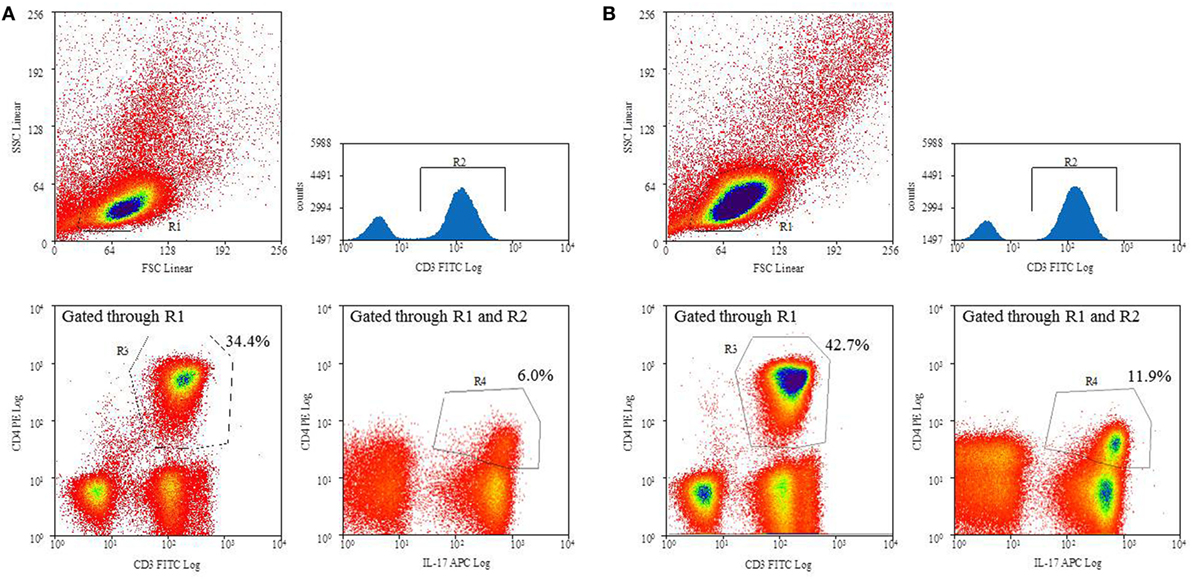
Figure 1. Definition of TH17 cells: peripheral blood mononuclear cells were stimulated with Ionomycin in vitro followed by staining with anti-CD3 (FITC), anti-CD4 (PE), and anti-IL-17A (APC). Representative flow cytometry analyses of a normal weight (A) and overweight (B) child, respectively.
An aliquot of the stimulated PBMCs was used for messenger RNA (mRNA) isolation with TriFast (peqlab, Erlangen, Germany) and subsequent DNase I digestion (Desoxyribonuclease I, Amplification Grade, invitrogen, Darmstadt, Germany), following the manufacturer’s instructions, respectively. cDNA was synthesized from 2 µl of mRNA using Omniscript R Kit (QIAGEN, Hilden) and Oligo(dT)18 primer and RiboLock RNase Inhibitor (Thermo Fisher Scientific, Nidderau, Germany). cDNA concentration was measured on a NanoDrop 2000c (Thermo Fisher Scientific, Nidderau, Germany) and was diluted by addition of RNase free water (Water, Mol Bio grade DNase-, RNase-, and Protease-free; 5 PRIME, Hamburg, Germany) into a concentration of 1,000 ng/µl.
Real-time PCR was performed on each cDNA sample as duplicate to determine relative expression of RORC- and IL-17A-mRNA and was normalized by Elongation factor 1-α (EF-1-α) as a housekeeping gene and calculated by the 2−ΔΔCt formula. EF-1-α was used because its expression in T cell culture is more stable than other housekeeping genes such as glyceraldehyde 3-phosphate dehydrogenase (23). 4.7 µl fluorochrom (SsoAdvanced SYBR Green Supermix; Biorad Laboratories, Munich, Germany), 4.2 µl RNase free water (Water, Mol Bio grade DNase-, RNase-, and Protease-free; 5 PRIME, Hamburg, Germany), 0.1 µl primer mix (all eurofins MWG Synthesis, Ebersberg, Germany) and 1 µl template were used per well of a 96-well plate. The following primers were used for real-time PCRs:
| EF-1-α-primer: | 5′-CTGAACCATCCAGGCCAAAT-3′; 5′-GCCGTGTGGCAATCCAAT-3′ |
| RORC-primer: | 5′-GCCTTTCATCATCATCTCTGC-3′; 5′-GAAGATCTGCAGCCTTTCCA-3′ |
| IL-17A-primer: | 5′-CCCCAGTTGATTGGAAGAAA-3′; 5′-TTCGTGGGATTGTGATTCCT-3′. |
An iQ5 (iQ5 Multicolor Real Time PCR Detector System; Biorad Laboratories, Munich, Germany) was used as thermocycler for real-time PCR. Real-time PCR analyses were executed with Bio-rad iQ5 Standard Edition software (Biorad Laboratories, Munich, Germany).
Statistical analyses were performed using Prism 5.0 (GraphPad Software, La Jolla, CA, USA). Group differences were tested using either the two-tailed Student’s t-test or, in case of non-normally distributed data, the Mann–Whitney U test. Violation of the normality assumption was assessed by Kolmogorov–Smirnov test. Analogously, Pearson and Spearman correlation was calculated for surface and intracellular staining for normally or non-normally distributed data, respectively. Differences with p-values of p < 0.05 were deemed significant.
65 children between 4 and 17 years of age were enrolled in our study—15 were overweight and 50 were not overweight as controls. The overweight children had a mean BMI-percentile of 97.12 ± 2.68 in comparison to 39.74 ± 27.04 in the control group. The patients’ anthropometric data are shown in Table 1.
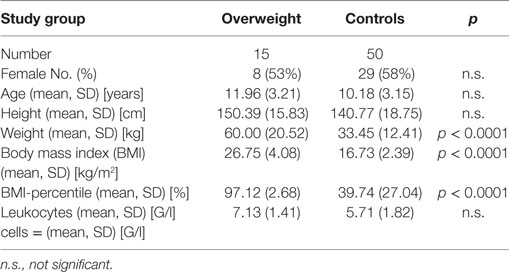
Table 1. Characteristics of the study subjects.
According to the inclusion criteria, all patients had a negative history of asthma, atopic dermatitis, allergic rhinoconjunctivitis, and autoimmune diseases. None of the children had an acute infection during the 2 weeks prior to the study enrollment as confirmed by medical history. A current infection was excluded by physical examination and measurement of C-reactive protein.
Flow cytometry analysis showed significantly higher TH17 cell frequencies after stimulation in overweight children than in non-overweight controls [34.7 ± 1.54% (mean ± SEM), and 25.4 ± 2.38%], respectively (p = 0.0023; Figure 2A).
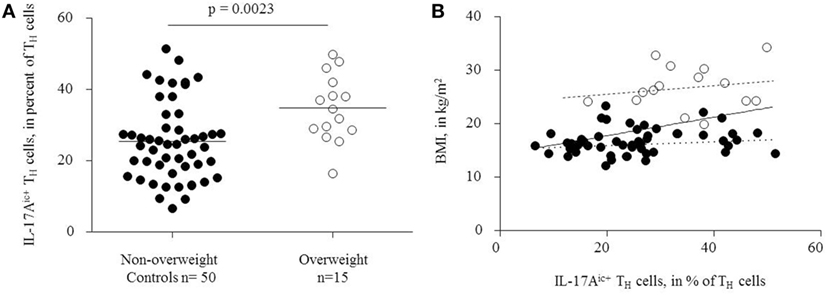
Figure 2. (A) Overweight children had significantly higher TH17 cell frequencies, defined as IL-17Aic+ TH cells, than controls. (B) Positive correlation between circulating TH17 cells and body mass index, solid regression line (r = 0.42, p = 0.0005). Filled dots: non-overweight controls, dotted regression line (r = 0.17, p = 0.27). Empty dots: overweight children, dashed regression line (r = 0.27, p = 0.33).
TH17 cell frequencies correlated positively with the absolute BMI (r = 0.42, p = 0.0005, Figure 2B). In contrast, IL-17 expression was similar among CD3+CD4− cells from non-overweight controls (45.9 ± 16.0) and overweight children (50.1 ± 17.3%) (p = 0.384, data not shown). Thus, we did not find signs of an unspecific stimulation of IL-17 production in non-TH17 cells. Separate analyses of the overweight and non-overweight groups yielded no significant correlation between BMI and TH17 cell frequencies (r = 0.27, p = 0.33; and r = 0.17, p = 0.23, respectively, Figure 2B).
Real-time PCR revealed that the relative mRNA expression of the transcription factor RORC and of IL-17A, which are both associated with the TH17 cell lineage, were 1.8- and 2.3-fold elevated in stimulated PBMCs from overweight children compared to non-overweight children, respectively (1.76 ± 0.35 versus 1.00 ± 0.13; p = 0.013; Figure 3A and 2.27 ± 0.72 versus 1.00 ± 0.16; p = 0.014; Figure 3B).
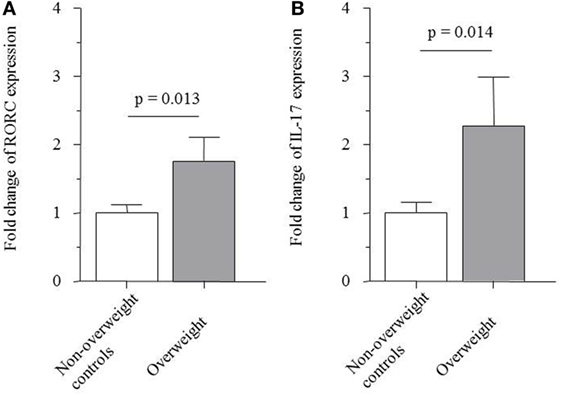
Figure 3. Messenger RNA expression of (A) transcription factor receptor-related orphan receptor C (RORC) (1.76 ± 0.35 versus 1.00 ± 0.13; p = 0.013) and (B) IL-17A (2.27 ± 0.72 versus 1.00 ± 0.16; p = 0.014) by PBMCs from non-overweight controls and overweight patients.
In this study, we observed that in the absence of acute or chronic inflammatory diseases, the frequency of circulating TH17 cells was significantly increased in overweight children compared to non-overweight controls and there was a positive correlation between TH17 cell frequency and BMI. In agreement with this, we registered a significantly higher expression of RORC- and IL-17A-mRNA transcripts after stimulation in PBMCs from overweight children.
Overweight and chronic inflammatory diseases are imprinted during the fetal and early postnatal periods, and the metabolic and inflammatory phenotypes of these diseases occur during the dynamic development of the immune system in children and juveniles (24). It is known from other studies that biomarkers associated with overweight may differ significantly between obese juveniles and adults [e.g., Ref. (25)]. This demonstrates that findings on metabolic and inflammatory diseases cannot be extrapolated from adults to children without restrictions. Thus, we chose to study the frequency of TH17 cells in children and juveniles before the metabolism and the immune system reach maturity.
Łuczyński et al. recently reported elevated TH17 cells in peripheral blood of children with central obesity and in children with diabetes mellitus I (19). Since obesity is linked with an increased risk for allergic and autoimmune diseases and TH17 cells were increased in patients with such conditions, we studied obese individuals without concomitant allergies and autoimmune diseases (5, 6, 8–10, 22).
The role of TH17 cells in the development of allergies is still under investigation. Probably, various pro-inflammatory and anti-inflammatory subpopulations of TH17 cells play unique roles in orchestrating the allergic inflammation (26). The cytokine production of TH17 cells appears to be more dynamic than previously anticipated depending on the signals provided by the microenvironment (26). Compared to healthy controls, IL-17A expression was higher in cultured T-lymphocytes from patients with mild–moderate asthma and persistent allergic rhinitis and decreased after anti-inflammatory therapy with inhalative corticosteroids (22). IL-17A influences the acute inflammatory response by upregulating IL-8 secretion in airway epithelial cells and also initiates airway remodeling by affecting the airway smooth muscle cells (27).
Since BMI standard values vary throughout childhood and between various ethnicities, we used the German percentiles that were obtained from measurements of 34,422 children (21). Due to the great variance of TH17 cell frequencies, they overlap between the overweight and non-overweight groups (Figure 2A). Intriguingly, BMI and TH17 cell frequency apparently do not show a linear correlation, since we found no significant correlation between BMI and TH17 cells when analyzing overweight and non-overweight children separately. This could indicate that the TH17 bias develops above a threshold BMI that provides a pro TH17 inflammatory environment or that the sample sizes were too small within the subgroups.
One limitation of the study is that due to the limited cell numbers in the pediatric samples, the expression of RORC and IL-17A transcripts were investigated in unsorted PBMCs. However, multiple previous studies have confirmed that CD4+ T cells are the major (although not exclusive) source of IL-17 and RORC expression in similar experimental settings as ours [e.g., Ref. (8, 22)]. In accord with this, we found that the number of IL-17ic+ cells did not increase among CD3+CD4− cells but only among CD3+CD4+ cells after in vitro stimulation (data not shown).
TH cell subsets are defined by characteristic expression patterns of surface antigens, cytokines, and master transcription factors. For instance, human TH1 cells produce interferon-γ and Tbet (28), TH2 cells produce IL-4 and GATA3 (29, 30), and TH17 cells produce IL-17A and RORC, which is orthologous to retinoic acid receptor-related orphan receptor γt (RORγt) in mice (8, 31). RORC as a master transcription factor is necessary for the differentiation of naïve TH cells into TH17 cells and is important for the production of TH17 cell effector cytokines in humans (32, 33).
In murine models, RORγt could induce IL-17A production in vitro (31). RORγt- and RORα-loss results in a total TH17 deficiency, single RORγt-loss in a relative one. In human TH17 cells, similar effects were observed: RORC-inhibition led to a reduced IL-17A-production and RORC-overexpression to an elevated differentiation into TH17 cells (34).
As effector memory TH17 cells, they secrete IL-21, IL-22, and IL-26, and express CCL20, CCR6, and CD161 (12, 35, 36). Multiple different staining protocols have been deduced from these lineage specific characteristics, but the complexity and insufficient understanding of the TH17 cell subsets precluded the development of a universally accepted standard definition. In this study, the staining protocol defines TH17 cells as CD3+CD4+IL-17Aic+ lymphocytes after stimulation—which is a common definition of flow cytometric assessed TH17 cells—with the strong, but unspecific stimulus Ionomycin (22). Possibly, a fraction of the CD3+CD4+IL-17Aic+ cells might not be classical TH17 cells but NKT cells, some of which can be CD4+ and may have the potential to express IL-17; however, according to previous work, this fraction is probably very small (37). It is unlikely that this short time culture has a polarizing effect toward the development of TH17 cells but it stimulates IL-17A expression in those cells that have already undergone polarization (38). According to previous studies with similar protocols, it is also unlikely that non-IL-17A-expressing TH cells vanish more quickly than their IL-17A-positive counterparts (38). Probably, less TH17 cells are actively producing IL-17A in vivo than in vitro following stimulation, since TH17 cells are characterized by their ability to produce IL-17A and not by actively producing it continuously.
Obesity does not only interfere with metabolic, but also with immunological pathways and is associated with low-grade chronic inflammation (39). Several groups reported an association between obesity and TH17 cells in mouse models. Winer et al. induced obesity by high fat diet and measured elevated TH17 cells and IL-17A production in murine spleens (17), and Chen et al. detected more TH17 cells in adipose tissue of obese mice compared to normal weight controls (18). Furthermore, TH17 cells were elevated in a model of obese mice that developed inflammatory arthritis. Thus, TH17 cells might contribute to the pathogenesis of inflammatory arthritis in obese mice (40). Some studies found elevated frequencies of TH17 cells or their cytokines in children with type 1 diabetes mellitus (41, 42). IL-17A secretion by human TH17 cells was enhanced in the presence of adipose-derived stem cells in vitro (43). Dendritic cells, sorted from subcutaneous adipose tissue of obese patients, can induce TH17 cell differentiation in vitro (44). Interestingly, IL-17A gene expression in CD3+ cells from subcutaneous adipose tissue was elevated overweight patients compared to normal weight patients (44). In lean state, M2 macrophages, Treg cells, TH2 cells, and NKT cells predominate, while M1 macrophages, TH1 cells, B cells, mast cells, and neutrophils predominate in obese individuals (6, 39). Adipose tissue produces adipokines which predominantly have a pro-inflammatory effect. Some of the identified adipokines include IL-6, IL-8, TNF-α, and IL-18 (39). TNF-α and IL-6 are required for TH17 cell differentiation (45). This pathway potentially leads to an increase of TH17 cell frequency in obese individuals. Leptin receptor signaling seems to be a further requirement for murine TH17 cell differentiation (46). The serum leptin concentration is elevated in obese individuals. Moreover, in humans, being overweight is associated with higher amounts of circulating neutrophils, which are attracted, among other things, by IL-17A (14, 47–49). In addition, elevated levels of myeloperoxidase were found in obese women, which coincide with higher levels of neutrophils as well (47).
Obesity can be a reason as well as a consequence of oxidative stress (50). One can hypothesize that oxidative stress might in part cause the association between obesity and allergies since obesity is associated with an overload of antioxidant compounds that promote a TH2-biased immune state as known from allergies (50). In support of this hypothesis, oxidative stress led to upregulated IL-17A serum levels in mice (51). In systemic lupus erythematosus, oxidative stress can cause mammalian target of rapamycin (mTOR) activation, and this mTOR activation in turn is suggested to stimulate TH17 cell differentiation (52, 53).
In this study, we observed an elevated TH17 cell frequency as well as elevated RORC- and IL-17A-mRNA expression in peripheral blood of overweight children without allergic asthma, allergic rhinoconjunctivitis, atopic dermatitis, or autoimmune disease. To our knowledge, this is the first study on TH17 cells in overweight pediatric patients that excludes potentially confounding chronic inflammatory diseases other than obesity. Elevated TH17 cells might contribute to the pro-inflammatory state in overweight children. Further studies are needed to specify the underlying pathomechanism of this association. In conclusion, future studies on the frequency of circulating TH17 cells should control for the BMI.
The datasets supporting the conclusions of this article are included within the article or are available from the authors upon request.
Theresa Isabell Schindler, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:dGhlcmVzYXNjaGluZGxlckBnbXguZGU=; Johanna-Josophina Wagner, HELIOS Klinikum Berlin-Buch, Department of Pediatrics, Schwanebecker Chaussee 50, D-13125 Berlin, Phone: +49 30 9401-14574, Fax: +49 309401-54509, E-Mail:am9oYW5uYWoud2FnbmVyQGdtYWlsLmNvbQ==; Sybelle Goedicke-Fritz, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:Z29lZGlja2VAbWVkLnVuaS1tYXJidXJnLmRl; Tobias Rogosch, CSL Behring, Clinical Development, Emil-von-Behring-Straße 76, D-35041 Marburg, Germany, Phone: +49 6421 39-7179, Fax: +49 6421 39-865-7179, E-Mail:VG9iaWFzLlJvZ29zY2hAY3NsYmVocmluZy5jb20=; Verena Coccejus, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:dmVyZW5hY29jY2VqdXNAYW9sLmNvbQ==; Verena Laudenbach, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:di5sYXVkZW5iYWNoQHdlYi5kZSw= Wilfried Nikolaizik, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:d2lsZnJpZWQubmlrb2xhaXppa0BtZWQudW5pLW1hcmJ1cmcuZGUs Christoph Härtel, Department of Pediatrics, University of Luebeck, RatzeburgerAllee 160, 23538 Lübeck, Germany, Phone: +49-451-500 2685, Fax: +49-451-500 6222, E-mail:Y2hyaXN0b3BoLmhhZXJ0ZWxAdWtzaC5kZQ==; Rolf Felix Maier, University Hospital Marburg, Medical Faculty, Philipps University Marburg, Children’s Hospital, Baldingerstraße, D-35043 Marburg, Phone: +49 6421-58 62650, Fax: +49 6421-58 68970, E-Mail:cm9sZi5tYWllckBtZWQudW5pLW1hcmJ1cmcuZGU=; Sebastian Kerzel, Department of Pediatric Pneumology and Allergy, University Children’s Hospital Regensburg (KUNO), Campus St. Hedwig, D-93049 Regensburg, Germany, Phone: +49 941 369 95817, Fax: +49 941 369 5424, E-Mail:c2ViYXN0aWFuLmtlcnplbEB1a3IuZGU=; Michael Zemlin, Department of Pediatrics and Neonatology, University Children’s Hospital of Saarland, Homburg Germany, Kirrberger Str. 100, D-66241 Homburg, Phone: +49 6841 16 28301, Fax: +49 6841 16 28310, E-Mail:bWljaGFlbC56ZW1saW5AdWtzLmV1.
This study was carried out in accordance with the recommendations of the Ethics Committee of the Philipps-University Marburg. All parents gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Ethics Committee of the Philipps-University Marburg with the identifier AZ 91/11.
SK, MZ, TR, WN, J-JW, and RM designed the study. TS, VC, VL, and SG-F performed the experiments and evaluated the data. All authors contributed to writing the manuscript.
Tobias Rogosch is an employee of CSL Behring. The remaining authors declare that they have no competing interests.
We wish to thank Dr. Hae-Sook Kim-Berger and the nurses of the outpatients clinics as well as all patients and their families for their support. We thank Sabine Jennemann and Regina Stoehr for excellent technical help.
The project was funded by the “von Behring-Roentgen Foundation” grants 63-LV04 and 62031685 and the “Rhoen Klinikum AG” grant FI-47.
BMI, body mass index; CCR, chemokine receptor; CD, cluster of differentiation; cDNA, complementary DNA; EF-1-α, elongation factor 1-α; IL, interleukin; mRNA, messenger RNA; PBMC, peripheral blood mononuclear cell; RORγt, retinoic acid receptor-related orphan receptor γt; RORC, retinoic acid receptor-related orphan receptor C; TH, T helper; TH1, T helper 1; TH2, T helper 2; TH17, T helper 17.
1. Guilherme A, Virbasius JV, Puri V, Czech MP. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat Rev Mol Cell Biol (2008) 9(5):367–77. doi:10.1038/nrm2391
2. Mandviwala T, Khalid U, Deswal A. Obesity and cardiovascular disease: a risk factor or a risk marker? Curr Atheroscler Rep (2016) 18(5):21. doi:10.1007/s11883-016-0575-4
3. de Onis M, Blössner M, Borghi E. Global prevalence and trends of overweight and obesity among preschool children. Am J Clin Nutr (2010) 92(5):1257–64. doi:10.3945/ajcn.2010.29786
4. Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. (2014). Available from: http://www.ginasthma.org
5. Stavropoulos-Kalinoglou A, Metsios GS, Koutedakis Y, Kitas GD. Obesity in rheumatoid arthritis. Rheumatology (Oxford) (2011) 50(3):450–62. doi:10.1093/rheumatology/keq266
6. Kanneganti TD, Dixit VD. Immunological complications of obesity. Nat Immunol (2012) 13(8):707–12. doi:10.1038/ni.2343
7. Cox AJ, West NP, Cripps AW. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol (2015) 3(3):207–15. doi:10.1016/S2213-8587(14)70134-2
8. Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM, et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol (2005) 6(11):1123–32. doi:10.1038/ni1254
9. Park H, Li Z, Yang XO, Chang SH, Nurieva R, Wang YH, et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat Immunol (2005) 6(11):1133–41. doi:10.1038/ni1261
10. Mosmann TR, Cherwinski H, Bond MW, Giedlin MA, Coffman RL. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol (1986) 136(7):2348–57.
11. Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, et al. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat Immunol (2007) 8(6):639–46. doi:10.1038/ni1467
12. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol (2009) 27:485–517. doi:10.1146/annurev.immunol.021908.132710
13. Ye P, Rodriguez FH, Kanaly S, Stocking KL, Schurr J, Schwarzenberger P, et al. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense. J Exp Med (2001) 194(4):519–27. doi:10.1084/jem.194.4.519
14. Tesmer LA, Lundy SK, Sarkar S, Fox DA. Th17 cells in human disease. Immunol Rev (2008) 223:87–113. doi:10.1111/j.1600-065X.2008.00628.x
15. Infante-Duarte C, Horton HF, Byrne MC, Kamradt T. Microbial lipopeptides induce the production of IL-17 in Th cells. J Immunol (2000) 165(11):6107–15. doi:10.4049/jimmunol.165.11.6107
16. Bedoya SK, Lam B, Lau K, Larkin J. Th17 cells in immunity and autoimmunity. Clin Dev Immunol (2013) 2013:986789. doi:10.1155/2013/986789
17. Winer S, Paltser G, Chan Y, Tsui H, Engleman E, Winer D, et al. Obesity predisposes to Th17 bias. Eur J Immunol (2009) 39(9):2629–35. doi:10.1002/eji.200838893
18. Chen Y, Tian J, Tian X, Tang X, Rui K, Tong J, et al. Adipose tissue dendritic cells enhances inflammation by prompting the generation of Th17 cells. PLoS One (2014) 9(3):e92450. doi:10.1371/journal.pone.0092450
19. Łuczyński W, Grubczak K, Moniuszko M, Głowińska-Olszewska B, Bossowski A. Elevated levels of Th17 cells in children with central obesity. Scand J Clin Lab Invest (2015) 75(7):595–601. doi:10.3109/00365513.2015.1066845
20. Moreno-Macias H, Romieu I. Effects of antioxidant supplements and nutrients on patients with asthma and allergies. J Allergy Clin Immunol (2014) 133(5):1237–44; quiz 1245. doi:10.1016/j.jaci.2014.03.020
21. Kromeyer-Hauschild K, Wabitsch M, Kunze D, Geller F, Geiß HC, Hesse V, et al. Perzentile für den Body-mass-Index für das Kindes- und Jugendalter unter Heranziehung verschiedener deutscher Stichproben. Monatsschr Kinderheilkd (2001) 149:807–18. doi:10.1007/s001120170107
22. Albano GD, Di Sano C, Bonanno A, Riccobono L, Gagliardo R, Chanez P, et al. Th17 immunity in children with allergic asthma and rhinitis: a pharmacological approach. PLoS One (2013) 8(4):e58892. doi:10.1371/journal.pone.0058892
23. Dheda K, Huggett JF, Bustin SA, Johnson MA, Rook G, Zumla A. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques (2004) 37(1):112–4. doi:10.2144/3701A0112
24. Goedicke-Fritz S, Härtel C, Krasteva-Christ G, Kopp M, Meyer S, Zemlin M. Preterm birth affects the risk of immune-mediated diseases. Front Immunol (2017) 8:1266. doi:10.3389/fimmu.2017.01266
25. Mangge H, Freytag F, Almer G, Weghuber D, Bauer-Denk C, Fuchs D. Serum neopterin is not increased in obese juveniles. J Obes (2011) 2011:946795. doi:10.1155/2011/946795
26. Sundrud MS, Trivigno C. Identity crisis of Th17 cells: many forms, many functions, many questions. Semin Immunol (2013) 25(4):263–72. doi:10.1016/j.smim.2013.10.021
27. Molet S, Hamid Q, Davoine F, Nutku E, Taha R, Pagé N, et al. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J Allergy Clin Immunol (2001) 108(3):430–8. doi:10.1067/mai.2001.117929
28. Szabo SJ, Kim ST, Costa GL, Zhang X, Fathman CG, Glimcher LH. A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell (2000) 100(6):655–69. doi:10.1016/S0092-8674(00)80702-3
29. Zheng W, Flavell RA. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell (1997) 89(4):587–96. doi:10.1016/S0092-8674(00)80240-8
30. Romagnani S. The Th1/Th2 paradigm. Immunol Today (1997) 18(6):263–6. doi:10.1016/S0167-5699(97)80019-9
31. Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell (2006) 126(6):1121–33. doi:10.1016/j.cell.2006.07.035
32. Morishima Y, Ano S, Ishii Y, Ohtsuka S, Matsuyama M, Kawaguchi M, et al. Th17-associated cytokines as a therapeutic target for steroid-insensitive asthma. Clin Dev Immunol (2013) 2013:609395. doi:10.1155/2013/609395
33. Newcomb DC, Peebles RS. Th17-mediated inflammation in asthma. Curr Opin Immunol (2013) 25(6):755–60. doi:10.1016/j.coi.2013.08.002
34. Manel N, Unutmaz D, Littman DR. The differentiation of human T(H)-17 cells requires transforming growth factor-beta and induction of the nuclear receptor RORgammat. Nat Immunol (2008) 9(6):641–9. doi:10.1038/ni.1610
35. Kleinschek MA, Boniface K, Sadekova S, Grein J, Murphy EE, Turner SP, et al. Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation. J Exp Med (2009) 206(3):525–34. doi:10.1084/jem.20081712
36. Cosmi L, De Palma R, Santarlasci V, Maggi L, Capone M, Frosali F, et al. Human interleukin 17-producing cells originate from a CD161+CD4+ T cell precursor. J Exp Med (2008) 205(8):1903–16. doi:10.1084/jem.20080397
37. Eger KA, Sundrud MS, Motsinger AA, Tseng M, Van Kaer L, Unutmaz D. Human natural killer T cells are heterogeneous in their capacity to reprogram their effector functions. PLoS One (2006) 1:e50. doi:10.1371/journal.pone.0000050
38. Olsen I, Sollid LM. Pitfalls in determining the cytokine profile of human T cells. J Immunol Methods (2013) 390(1–2):106–12. doi:10.1016/j.jim.2013.01.015
39. Ahmed M, Gaffen SL. IL-17 in obesity and adipogenesis. Cytokine Growth Factor Rev (2010) 21(6):449–53. doi:10.1016/j.cytogfr.2010.10.005
40. Jhun JY, Yoon BY, Park MK, Oh HJ, Byun JK, Lee SY, et al. Obesity aggravates the joint inflammation in a collagen-induced arthritis model through deviation to Th17 differentiation. Exp Mol Med (2012) 44(7):424–31. doi:10.3858/emm.2012.44.7.047
41. Marwaha AK, Crome SQ, Panagiotopoulos C, Berg KB, Qin H, Ouyang Q, et al. Cutting edge: Increased IL-17-secreting T cells in children with new-onset type 1 diabetes. J Immunol (2010) 185(7):3814–8. doi:10.4049/jimmunol.1001860
42. Alnek K, Kisand K, Heilman K, Peet A, Varik K, Uibo R. Increased blood levels of growth factors, proinflammatory cytokines, and Th17 cytokines in patients with newly diagnosed type 1 diabetes. PLoS One (2015) 10(12):e0142976. doi:10.1371/journal.pone.0142976
43. Eljaafari A, Robert M, Chehimi M, Chanon S, Durand C, Vial G, et al. Adipose tissue-derived stem cells from obese subjects contribute to inflammation and reduced insulin response in adipocytes through differential regulation of the Th1/Th17 balance and monocyte activation. Diabetes (2015) 64(7):2477–88. doi:10.2337/db15-0162
44. Bertola A, Ciucci T, Rousseau D, Bourlier V, Duffaut C, Bonnafous S, et al. Identification of adipose tissue dendritic cells correlated with obesity-associated insulin-resistance and inducing Th17 responses in mice and patients. Diabetes (2012) 61(9):2238–47. doi:10.2337/db11-1274
45. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity (2006) 24(2):179–89. doi:10.1016/j.immuni.2006.01.001
46. Reis BS, Lee K, Fanok MH, Mascaraque C, Amoury M, Cohn LB, et al. Leptin receptor signaling in T cells is required for Th17 differentiation. J Immunol (2015) 194(11):5253–60. doi:10.4049/jimmunol.1402996
47. Andrade VL, Petruceli E, Belo VA, Andrade-Fernandes CM, Caetano Russi CV, Bosco AA, et al. Evaluation of plasmatic MMP-8, MMP-9, TIMP-1 and MPO levels in obese and lean women. Clin Biochem (2012) 45(6):412–5. doi:10.1016/j.clinbiochem.2012.01.008
48. Dixon JB, O’Brien PE. Obesity and the white blood cell count: changes with sustained weight loss. Obes Surg (2006) 16(3):251–7. doi:10.1381/096089206776116453
49. Ilavská S, Horváthová M, Szabová M, Nemessányi T, Jahnová E, Tulinská J, et al. Association between the human immune response and body mass index. Hum Immunol (2012) 73(5):480–5. doi:10.1016/j.humimm.2012.02.023
50. Rani V, Deep G, Singh RK, Palle K, Yadav UC. Oxidative stress and metabolic disorders: pathogenesis and therapeutic strategies. Life Sci (2016) 148:183–93. doi:10.1016/j.lfs.2016.02.002
51. Al-Harbi NO, Nadeem A, Al-Harbi MM, Ansari MA, AlSharari SD, Bahashwan SA, et al. Airway oxidative stress causes vascular and hepatic inflammation via upregulation of IL-17A in a murine model of allergic asthma. Int Immunopharmacol (2016) 34:173–82. doi:10.1016/j.intimp.2016.03.003
52. Delgoffe GM, Pollizzi KN, Waickman AT, Heikamp E, Meyers DJ, Horton MR, et al. The kinase mTOR regulates the differentiation of helper T cells through the selective activation of signaling by mTORC1 and mTORC2. Nat Immunol (2011) 12(4):295–303. doi:10.1038/ni.2005
Keywords: TH17 cells, IL-17A, receptor-related orphan receptor C, inflammation, overweight, children
Citation: Schindler TI, Wagner J-J, Goedicke-Fritz S, Rogosch T, Coccejus V, Laudenbach V, Nikolaizik W, Härtel C, Maier RF, Kerzel S and Zemlin M (2017) TH17 Cell Frequency in Peripheral Blood Is Elevated in Overweight Children without Chronic Inflammatory Diseases. Front. Immunol. 8:1543. doi: 10.3389/fimmu.2017.01543
Received: 27 July 2017; Accepted: 30 October 2017;
Published: 16 November 2017
Edited by:
Rene De Waal Malefyt, Merck Research Laboratories, United StatesReviewed by:
Thomas Ciucci, National Cancer Institute (NIH), United StatesCopyright: © 2017 Schindler, Wagner, Goedicke-Fritz, Rogosch, Coccejus, Laudenbach, Nikolaizik, Härtel, Maier, Kerzel and Zemlin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sybelle Goedicke-Fritz, Z29lZGlja2VAbWVkLnVuaS1tYXJidXJnLmRl
†Co-first authors.
‡Co-last authors.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.