 Moustafa Hamze1
Moustafa Hamze1 Sylvain Meunier1
Sylvain Meunier1 Anette Karle2
Anette Karle2 Abdelaziz Gdoura1Amélie Goudet1Natacha Szely3
Abdelaziz Gdoura1Amélie Goudet1Natacha Szely3 Marc Pallardy3Franck Carbonnel4Sebastian Spindeldreher2
Marc Pallardy3Franck Carbonnel4Sebastian Spindeldreher2 Xavier Mariette5Corinne Miceli-Richard5
Xavier Mariette5Corinne Miceli-Richard5 Bernard Maillère1*
Bernard Maillère1*
- 1CEA-Saclay, Institut de Biologie et Technologies, Université Paris-Saclay, Gif sur Yvette, France
- 2Novartis Pharma AG, Basel, Switzerland
- 3INSERM UMR 996, Faculté de Pharmacie, Université Paris-Sud, Chatenay Malabry, France
- 4Service de gastro-entérologie, Hôpitaux Universitaires Paris-Sud, Le Kremlin-Bicêtre, France
- 5INSERM UMR 1184, Assistance Publique-Hôpitaux de Paris, Service de Rhumatologie, Hôpitaux Universitaires Paris-Sud, Université Paris-Sud, Le Kremlin-Bicêtre, France
The chimeric antibodies anti-CD20 rituximab (Rtx) and anti-TNFα infliximab (Ifx) induce antidrug antibodies (ADAs) in many patients with inflammatory diseases. Because of the key role of CD4 T lymphocytes in the initiation of antibody responses, we localized the CD4 T cell epitopes of Rtx and Ifx. With the perspective to anticipate immunogenicity of therapeutic antibodies, identification of the CD4 T cell epitopes was performed using cells collected in healthy donors. Nine T cell epitopes were identified in the variable chains of both antibodies by deriving CD4 T cell lines raised against either Rtx or Ifx. The T cell epitopes often exhibited a good affinity for human leukocyte antigen (HLA)-DR molecules and were part of the peptides identified by MHC-associated peptide proteomics assay from HLA-DR molecules of dendritic cells (DCs) loaded with the antibodies. Two-third of the T cell epitopes identified from the healthy donors stimulated peripheral blood mononuclear cells from patients having developed ADAs against Rtx or Ifx and promoted the secretion of a diversity of cytokines. These data emphasize the predictive value of evaluating the T cell repertoire of healthy donors and the composition of peptides bound to HLA-DR of DCs to anticipate and prevent immunogenicity of therapeutic antibodies.
Introduction
Therapeutic antibodies have become a major strategy in oncology and clinical immunology. However, a large subset of patients develops neutralizing antidrug antibodies (ADAs) that reduce their therapeutic efficacy (1) and induce allergic responses (2). As immunological tolerance is expected to regulate the response to self-sequences (3), almost all therapeutic antibodies are humanized, with various degrees of antibody humanization (4). Chimeric antibodies are humanized on their constant parts only, their variable regions remaining of murine origin, while humanized antibodies are also modified in the framework (FR) regions of the variable parts. Fully human antibodies are mostly produced by in vitro selection of antibodies encoded by human immunoglobulin genes or by immunization of Ig-humanized mice. Nevertheless, none of these approaches fully guarantee the absence of immune responses. Chimeric antibodies such as rituximab (Rtx) (5–7) and infliximab (Ifx) (8–10) are known to elicit specific ADAs in multiple patients, generally associated with reduced clinical efficacy. Similarly, the humanized antibodies alemtuzumab (11) and vedolizumab (12) and the fully human adalimumab (10, 13, 14) are known to generate ADAs in many patients. While humanization of the constant parts of therapeutic antibodies clearly reduces ADA responses, the benefits of humanization of the variable parts remain controversial (15), reflecting the lack of knowledge about the molecular determinants contributing to immunogenicity of therapeutic antibodies (16).
Immunogenicity of antibodies mainly relies on the presentation of antibody-derived peptides displayed on APCs and their capacity to stimulate specific CD4 T lymphocytes. CD4 T lymphocytes participate to the immune response to therapeutic proteins (16–20). However, T cell epitopes contained in marketed therapeutic proteins are largely unknown, although localization of T cell epitopes could help to mitigate immunogenicity by removing them from the initial sequence (19, 21). As T cell epitopes bind to human leukocyte antigen (HLA) class II molecules, locating HLA class II binding peptides could serve as a first step in the evaluation of the immunogenic potential of therapeutic proteins (22–24). However many good peptide binders to HLA molecules do not necessarily elicit a T cell response, especially those derived from self-proteins as many self-reactive T cells are eliminated by central tolerance. Therefore, T cell assays have been established using cells collected from healthy donors (25–29). As healthy donors have never been exposed to therapeutic proteins, T cell assays aim to detect low-frequency specific naïve T cells, assuming that the assays reproduce the memory T cell response that occurs after injection of the therapeutic proteins in the body. However, the relevance of T cell epitopes identified from healthy donors accounting for the T cell response in patients has never been formally demonstrated for therapeutic antibodies.
We therefore investigated the T cell response to the immunogenic chimeric antibodies Rtx and Ifx. Rtx is specific for CD20, a surface marker of B cell lymphocytes and is a B cell-depleting therapeutic antibody approved for the treatment of many lymphomas, leukemias, and autoimmune disorders (5–7, 30). Ifx targets TNF-α and has been proven highly effective in the treatment of inflammatory diseases (8–10). Both antibodies are immunogenic in many patients suffering from inflammatory diseases with an immunogenicity incidence ranging from 10 to 60% (5–10). In this study, we identified the T cell epitopes in the variable parts of Rtx and Ifx using cells collected from healthy donors and evaluated their capacity to stimulate T cells collected from patients with ADA. We characterized naturally presented HLA class II peptides from human dendritic cells (DCs) loaded with the antibodies and evaluated the binding affinities of overlapping peptides to HLA class II molecules. We therefore established a detailed map of the T cell epitopes of Rtx and Ifx, which helped to understand the origin of their immunogenicity.
Materials and Methods
Proteins and Peptides
Keyhole limpet hemocyanin (KLH) was purchased from Thermo Fisher Scientific (Brebières, France). Rtx (Mabthera®) was purchased from Roche (Neuilly, France) and Ifx (Remicade®) from Centocor (Horsham, PA, USA). Peptides were purchased from Pepscan (Lelystad, The Netherlands).
Characterization of Antibody-Specific CD4 T Cell Lines
Peripheral blood mononuclear cells (PBMCs) were obtained from blood cells collected at the Etablissement Français du Sang (EFS, Rungis, France), as buffy coat preparations from anonymous healthy donors who gave informed consent, in accordance with EFS guidelines. Antibody-specific CD4 T cell lines were generated as described previously (28). DCs were produced from plastic-adherent cells of PBMCs, while CD4 T cells were isolated from PBMCs by using magnetic microbeads (Miltenyi Biotech, Paris, France). DCs were loaded overnight at 37°C with the therapeutic antibody or with KLH used as a positive control (1 µM) and matured with lipopolysaccharide (1 µg/mL). CD4 T cells (200,000/w) were stimulated by protein-loaded DCs (20,000/w) and cultured during 21 days (28). Their peptide specificity was tested by interferon-γ (IFN-γ) ELISPOT (28). Spot number was determined by the AID ELISPOT Reader System (AID). CD4 T cell lines were considered as specific when a spot count was twofold higher in the presence of the protein or the peptide than in their absence, with a minimal difference of 25 spots.
HLA-DR-Specific Binding Assays
Human leukocyte antigen-DR molecules were immunopurified from homozygous EBV B lymphoblastoid cell as previously reported (31, 32). Binding of the Rtx and Ifx peptides to HLA-DR molecules was assessed by competitive ELISA, as previously described (33). A strong binder to each HLA class II molecule was introduced in each assay as reference (33). Data were reported as relative affinity corresponding to the ratio of the IC50 of the tested peptide to the IC50 of the reference peptide. Means were calculated from at least two independent experiments.
MHC-Associated Peptide Proteomics (MAPPs) Assay
Human leukocyte antigen-DR-associated peptides from DCs loaded with Ifx or Rtx were identified via the MAPPs assay as described previously (34–37). Briefly, CD14 positive mononuclear cells were purified from PBMCs isolated from human buffy coats sampled from consented healthy donors (Blood Donation Center Bern, Bern, Switzerland) and differentiated into immature DCs (37). Immature DCs were matured by adding lipopolysaccharide (1 µg/mL, Sigma) and loaded separately with one of the chimeric antibodies. After incubation for 24 h at 37°C and 5% CO2, DCs were harvested, washed in PBS, and lysed in hypotonic buffer containing 1% Triton X-100. After immunoprecipitation of HLA-DR molecules with Mab L243-conjugated beads, peptides were eluted from HLA-DR molecules by adding 0.1% trifluoroacetic acid (Fluka, Buchs, Switzerland) at 37°C and lyophilized using an Eppendorf Concentrator 5301 (Eppendorf AG, Hamburg, Germany). Lyophilized peptides were resuspended in hydrophilic buffer containing 5% acetonitrile and 1.1% formic acid. Peptide composition was analyzed by liquid chromatography (nano capillary system, Dionex Corporation, Sunnyvale, CA, USA) on a self-packed fused-silica C18 reversed-phase nano-high-performance liquid chromatography column connected to a mass spectrometer (Q-Exactive, Thermo, CA, USA) via electrospray ionization (LC–ESI–MS/MS). Peptides were identified via a database search approach using the SEQUEST algorithm as detailed (37).
Evaluation of the T Cell Response to Ifx and Rtx Peptides in Immunized Patients
Blood samples from patients immunized against Rtx or Ifx were collected at Bicêtre Hospital (Le Kremlin-Bicêtre, France) in accordance with French law and after approval by the patients. Levels of circulating Ifx or Rtx and of ADAs specific for each antibody were quantified using bridging ELISA kits (Theradiag, Croissy Beaubourg, France). PBMCs were seeded at 5 M/mL with peptides (10 µg/mL) in RPMI 1640 medium supplemented with 10% SAB, 50 IU/mL IL-2 (R&D), 1,000 U/mL IL-4 (R&D), and 1 µg/mL anti-CD28 (Miltenyi). The culture medium was changed every 2–3 days. Cells were harvested at day 10 and incubated with individual peptides (10 µg/mL) in AIM-V supplemented with 0.5 ng/mL IL-7 at 37°C in Multiscreen 96-well plates (Merck Millipore) previously coated overnight at 4°C with 4 µg/mL anti-human IL-5 MAb (TRFK5, Mabtech). After 48 h incubation, supernatants were collected, and IL-5 secretion was detected by successive addition of mouse biotinylated anti-human IL-5 Mab (TRFK4, Mabtech), Extravidin conjugate, and BCIP substrate (Sigma) as described above for IFNγ ELISPOT. Spots were counted with a computer-assisted video image analyzer (AID, Strassberg, Germany). A positive value was assigned to culture wells with a spot count that was twofold higher in the presence of the peptide than in its absence, with a minimal difference of 25 spots and positivity in Student’s t-test (P < 0.05). Supernatants of IL-5 ELISPOT plates were submitted to Luminex cytokine assays (R&D) for six different cytokines (IL-2, IL-4, IL-10, IL-13, IL-17, and IFN-γ) according to the manufacturer’s instructions (R&D). Sensitivity of detection ranged from 0.4 to 2 pg/mL, except for IL-4 (9.3 pg/mL) and IL-13 (36.6 pg/mL). Cytokine concentrations lower than the lower limits of detection were reported as undetectable. A positive value was assigned to culture wells with a concentration that was twofold higher in the presence of the peptide than in its absence and positivity in Student’s t-test (P < 0.05).
Results
Ifx- and Rtx-Specific T Cell Epitopes Were Identified in the Variable Domains Using Cells Collected in Healthy Donors
To identify CD4 T cell epitopes of Ifx and Rtx, we constituted 2 panels of 15 healthy donors on the basis of the frequency of HLA-DRB1 alleles in Europe (Table S1 in Supplementary Material). From the PBMCs of these donors we derived CD4 T cell lines specific for Rtx and Ifx by 4 weekly rounds of stimulation of the CD4 T cells by DCs loaded with either Rtx or Ifx. CD4 T cell lines were also raised against KLH to assess the ability of the donors to generate specific T cell responses (Figure S2 in Supplementary Material). Each independent T cell line (CD4 T cells present in a single well) was evaluated for its specificity using pools of 45 overlapping peptides encompassing the entire variable regions (VH and VL) of the therapeutic antibodies (Tables S3 and S4 in Supplementary Material), individual peptides, and DCs loaded with the therapeutic antibodies (Figure 1). As an example, T cell line 113.6 from donor #113 was specific for Rtx but not for Ifx and reacted with the pool 2 of Rtx peptides in the first ELISPOT and with the peptide RH56–70 in the second one (Figure 1A). In contrast, T cell line 247.26 from donor #247 was specific for Ifx but not for Rtx and reacted with the pool 3 of Ifx peptides and the peptide IH91–105 (Figure 1B). Each healthy donor gave rise to a variable number of antibody-specific T cell lines as shown for multiple donors (Figure 2A). HLA restriction was evaluated for T cell lines of donors #225 and #251 by inhibition experiments with anti-HLA class II antibodies. These T cell lines appeared to be restricted to HLA-DR molecules and not to HLA-DP or DQ (Figure 2B). Together, 11 out of 14 donors generated a CD4 T cell response to Rtx peptides, and 12 out of 15 donors generated a CD4 T cell response to Ifx peptides (Figure 3). A CD4 T cell response specific for each antibody was sustained by nine different T cell epitopes found in the variable regions of both light and heavy chains. The Rtx peptide RH56–70 was common to three donors, while RH31–45, RH41–55, and RL41–55 were shared by two donors (Figure 3A). Four donors shared the same peptide Ifx IH46–60, while peptides IH91–105, IL31–45, and IL6–20 were common to two donors (Figure 3B).
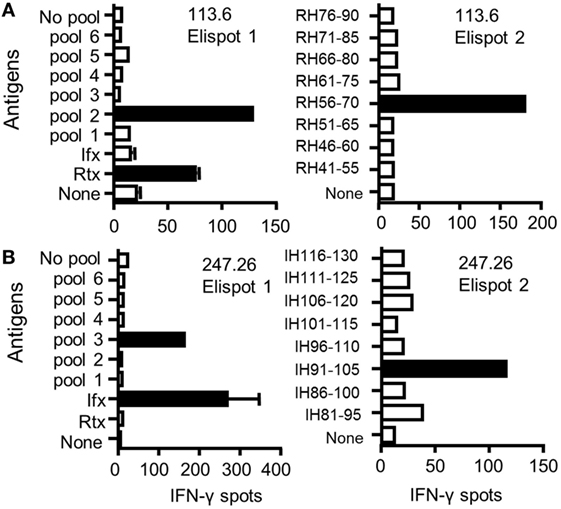
Figure 1. Identification of CD4 T cell epitopes of infliximab (Ifx) and rituximab (Rtx) from healthy donors. CD4 T cell lines were generated in vitro by four weekly rounds of stimulation with autologous dendritic cells (DCs) loaded with either Rtx or Ifx. Specificity of CD4 T cell lines #113.6 raised against Rtx (A) and #247.26 raised against Ifx (B) was analyzed by interferon-γ ELISPOT. T cells were incubated with autologous DCs alone (none) or with DCs previously loaded with each antibody (3 µM) or with autologous unloaded peripheral blood mononuclear cells (PBMCs) (no pool) or PBMCs loaded with a pool of peptides (10 µg/mL) (left panels) and with individual peptides (right panels).
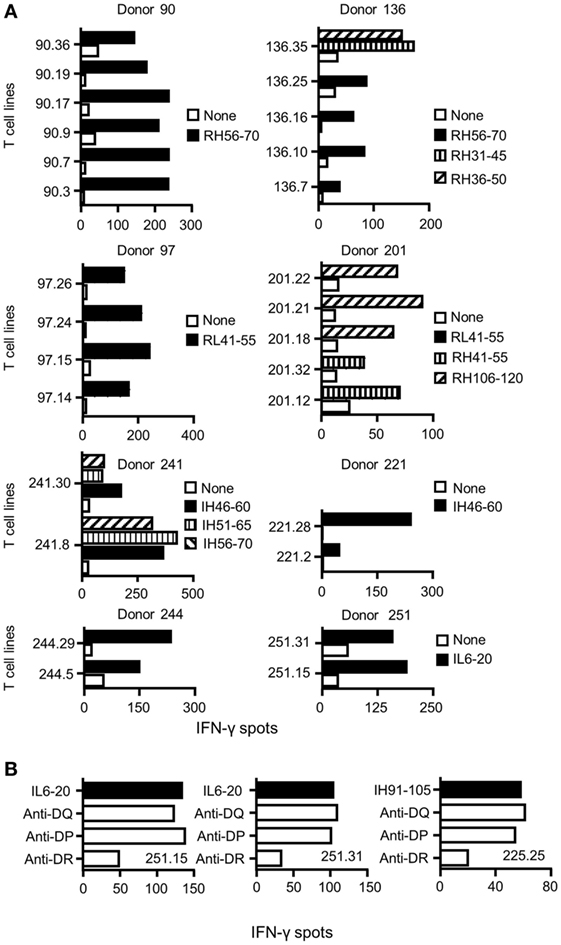
Figure 2. Specificity and restriction of T cell lines generated from healthy donors. (A) CD4 T cell lines were generated in vitro by four weekly rounds of stimulation with autologous dendritic cells loaded with either rituximab (Rtx) or infliximab (Ifx). T cells were incubated with autologous unloaded peripheral blood mononuclear cells (PBMCs) (none) and with PBMCs loaded with individual peptides. Activation of the T cells was evaluated by interferon-γ (IFN-γ) ELISPOT, and peptide specificity was confirmed in two independent experiments. Each panel reports the peptide-specific T cell lines found from one donor. Cells from donors 90, 97, 136, and 201 were used to evaluate the T cell response to Rtx, while cells from donors 221, 241, 244, and 251 were used for Ifx. (B) For inhibition assays, anti-HLA-DR (L243), -DQ (SPVL3), and -DP (B7/21) antibodies were added at 10 µg/mL to the IFN-γ ELISPOT.
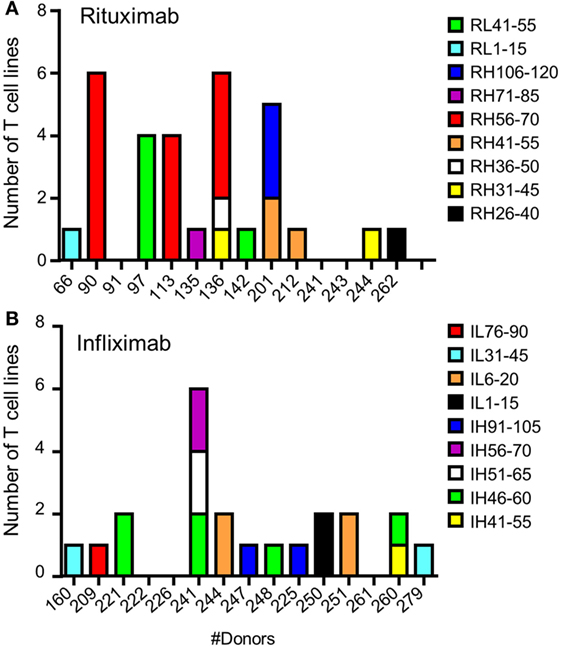
Figure 3. Specificity for rituximab (Rtx) and infliximab (Ifx) peptides of the CD4 T cells preexisting in healthy donors. T cell lines were generated, and their peptide specificity was assessed as described in the legend of Figure 1 using cells collected from 14 and 15 donors for Rtx and Ifx, respectively. Donors are identified by a number. The number of T cell lines specific for Rtx (A) and Ifx (B) is reported for each donor.
Binding of Ifx and Rtx Peptides to Purified HLA Class II Molecules
The two sets of overlapping peptides were submitted to 11 different binding assays specific for common HLA-DR alleles (Figure 4). These alleles comprise eight molecules encoded by the HLA-DRB1 (01:01, 03:01, 04:01, 07:01, 09:01, 11:01, 13:01, and 15:01) gene and three second HLA-DR molecules (DRB3*01:01, DRB4*01:01, and DRB5*01:01). Binding activities were found all along the variable sequences of both antibodies. Multiple peptides bound with strong or moderate affinity to multiple HLA-DR molecules and comprised several CD4 T cell epitopes (RH31–45, RH106–120, RL1–15, RL41–55, IH91–105, and IL6–20). As shown Table 1, a majority of CD4 T cell epitopes exhibited a strong or moderate binding to at least one of the HLA-DR molecules of the donors responding to them. It is of note that the peptide RH56–70 was highly specific for the HLA-DR11 molecule and generated a T cell response in three HLA-DR11 donors. In contrast, the peptides RL41–55 and IH46–60 bound to 6 and 3 HLA-DR molecules, respectively, and concomitantly stimulated T cells in 2 and 4 donors, respectively. In summary, most of the T cell epitopes are associated with strong or moderate peptide binding to HLA-DR molecules but not all of the strong binders lead to T cell stimulation.
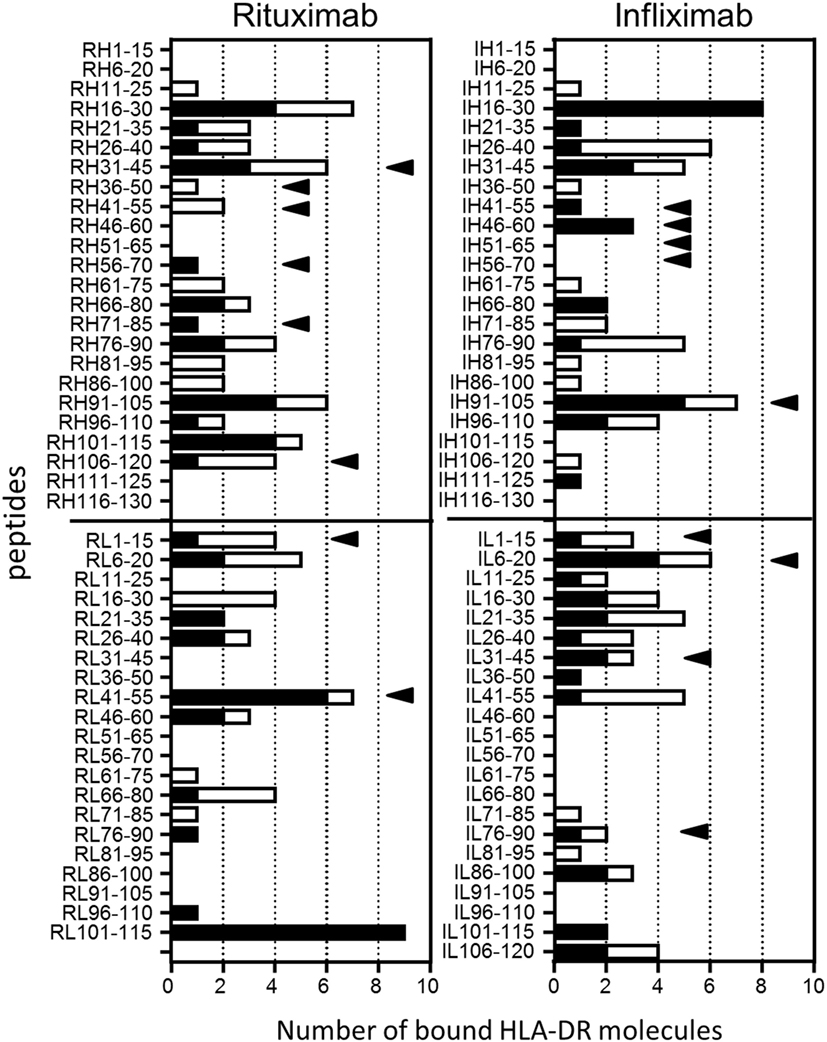
Figure 4. Binding of rituximab (Rtx) and infliximab (Ifx) overlapping peptides to common human leukocyte antigen (HLA)-DR molecules. Overlapping 15-mer peptides covering the whole sequence of the variable parts of Rtx and Ifx were submitted to competitive ELISA specific for the following HLA-DR molecules: DRB1*01:01, *04:01, *11:01, *07:01, *01:01, *03:01, *09:01, *1301, *1501, DRB3*01:01, DRB4*01:01, and DRB5*01:01. Data are reported as number of bound HLA-DR molecules for each peptide. Relative affinity below 20 corresponds to strong binding (black), while moderate binding is in the range of 20–100 (blank). Black arrows: CD4 T cell epitopes.
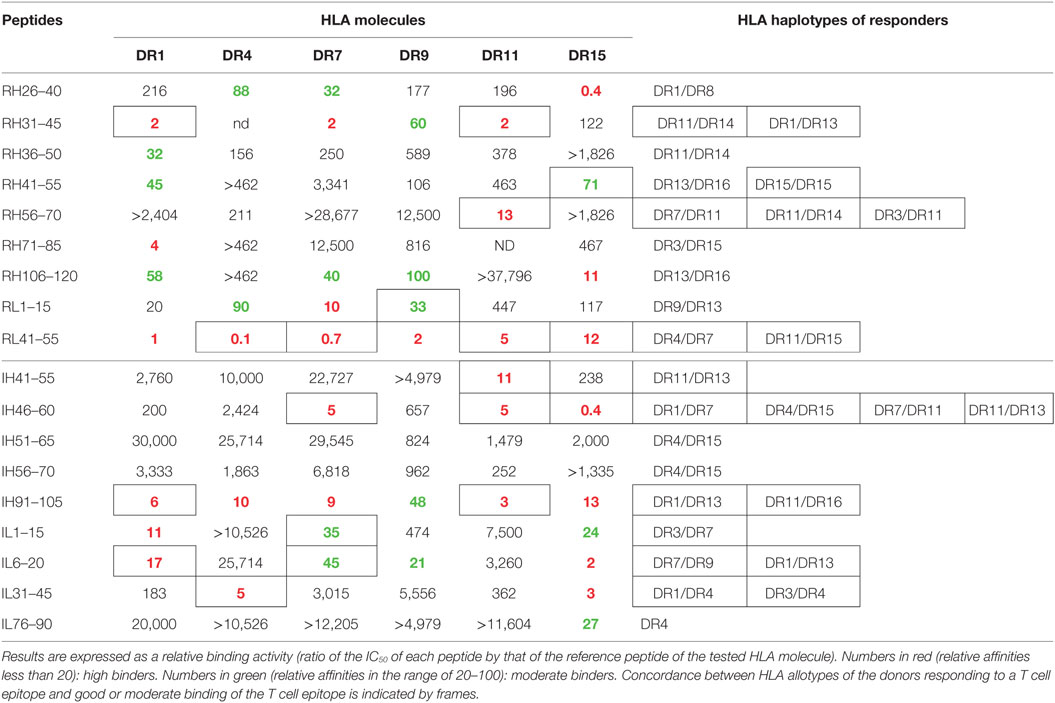
Table 1. Binding of infliximab and rituximab CD4 T cell epitopes to human leukocyte antigen (HLA)-DR molecules found in responding healthy donors.
CD4 T Cell Epitopes Are Retrieved from Elution Experiments of Naturally Presented Peptides of Rtx and Ifx
Naturally presented HLA-DR-associated peptides were identified using the MAPPs assay (34–37) from monocyte-derived DCs of 34 healthy donors exposed to Rtx or Ifx. Peptides typically occur as multiple length variants, which share the same HLA-DR binding core and form a “cluster.” Five and four clusters were found in the VH and VL domains of Rtx, respectively (Figure 5). Each VH and VL domain of Ifx hosted four clusters. Four of the nine Rtx clusters and four of the eight Ifx clusters overlapped CDR regions, while the FR regions hosted about the half of the clusters. Many clusters of Ifx and Rtx encompassed the identified T cell epitopes.

Figure 5. Rituximab (Rtx) and infliximab (Ifx) peptides identified by MHC-associated peptide proteomics assay. Human leukocyte antigen-DR molecules were immunopurified from dendritic cell loaded with Rtx or Ifx, and eluted peptides were identified by nano-high-performance liquid chromatography and electrospray ionization mass spectrometry (LC–ESI–MS/MS). Peptide sequences (black) are representative of the clusters found by each donor. Only donors with identified peptides were reported. Blue: CDRs.
CD4 T Cell Epitopes Identified in the Healthy Donors Contribute to the T Cell Response to Ifx and Rtx in Patients Having Developed ADA
We then investigated the CD4 T cell response in seven patients treated with Ifx (n = 6) or Rtx (n = 1) (one with granulomatous uveitis; five with Crohn’s disease, and one with rheumatoid arthritis) having developed ADAs against Ifx or Rtx (Table S5 in Supplementary Material). To detect memory CD4 T cells, PBMCs were submitted to a short-term T cell assay with only one step of in vitro stimulation instead of multiple rounds of antigenic stimulation as we did with healthy donors to detect naïve T cells. Accordingly, in this short-term T cell assay, PBMCs collected in three healthy donors did not generate a T cell response to the Rtx and Ifx T cell epitopes. PBMCs from five patients were cultured with a pool of the nine Ifx CD4 T cell epitopes for 10 days and then submitted to IL-5 ELISPOT using individual peptides (Figure 6A). All the patients responded to at least one peptide. Peptides IL1–15, IH41–55, and IH46–60 were common to three patients, while peptide IH91–105 was shared by two patients. Peptides IH51–65 and IL76–90 stimulated T cells in one patient, only. For two patients (L and C), sufficient amount of PBMCs was available to screen the peptide specificity of the T cells with the complete set of peptides and not with the T cell epitopes, only (Figure 6B). Four peptides (RL41–55, RH31–46, RH36–50, and RH46–60) among the 45 Rtx peptides gave rise to a T cell response for patient L. Two peptides (IH56–70 and IH41–55) from a total of the 46 Ifx peptides were active for patient C. All the peptides active in patients C and L are part of the Ifx and Rtx T cell epitopes we identified from the healthy donors.
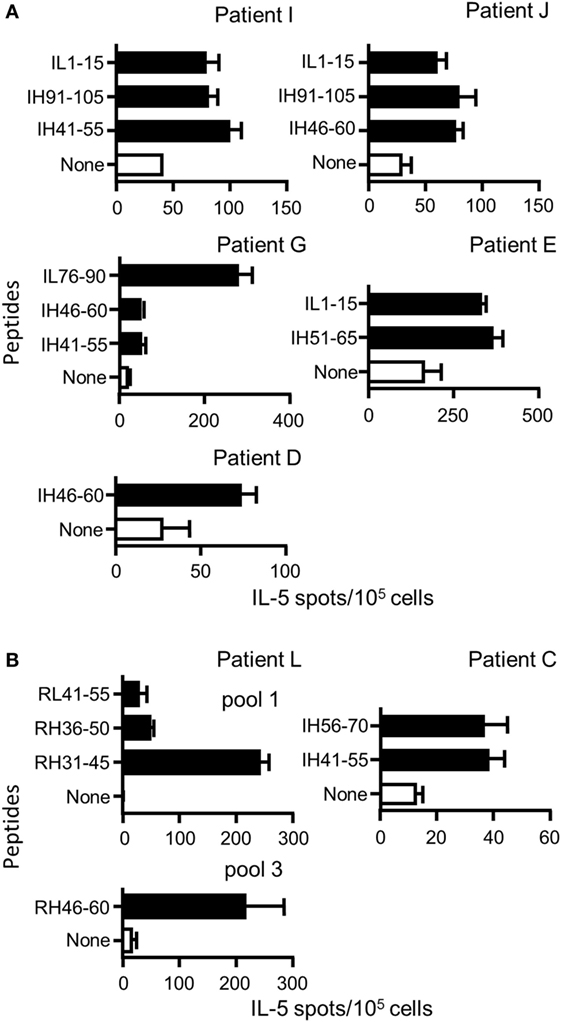
Figure 6. Peptide specificity of T cells collected in patients with antidrug antibodies (ADAs) against rituximab (Rtx) or infliximab (Ifx). Peripheral blood mononuclear cells of patients with ADA against Rtx or Ifx were cultured with pools of Rtx and Ifx peptides for 10 days, and their peptide specificity was assessed by IL-5 ELISPOT using individual peptides (A) Ifx T cell epitopes already identified in the healthy donors composed the pool of peptides (B) All the peptides covering the entire sequences of the variable parts of Rtx or Ifx were introduced in the assay to evaluate the T cell response of the donors C and L. Only positive responses are presented [spot count that twofold higher in the presence of the peptide than in its absence, with a minimal difference of 25 spots and positivity in Student’s t-test (P < 0.05)].
Finally, supernatants of the ELISPOT plates were submitted to multiplex cytokine assay (Figure 7). Under the assay conditions, IL-2, IL-4, and IL-13 were not detected in any of the patients. Besides IL-5, PBMCs of patients L and J secreted IL-10, while patients E and G secreted IL-10 and IFN-γ. Patient D developed a T cell response characterized by the secretion of IFN-γ and IL-17, while a high level of IFN-γ secretion supported by multiple peptides was found for patient E. Altogether, we demonstrated that four Rtx and eight Ifx T cell epitopes identified in the healthy donors participated to the T cell responses elicited in patients having developed ADA against Rtx or Ifx. The T cell responses to Rtx and Ifx exhibited different patterns of secretion of cytokines, including IL-5, IFN-γ, IL-10, and IL-17.
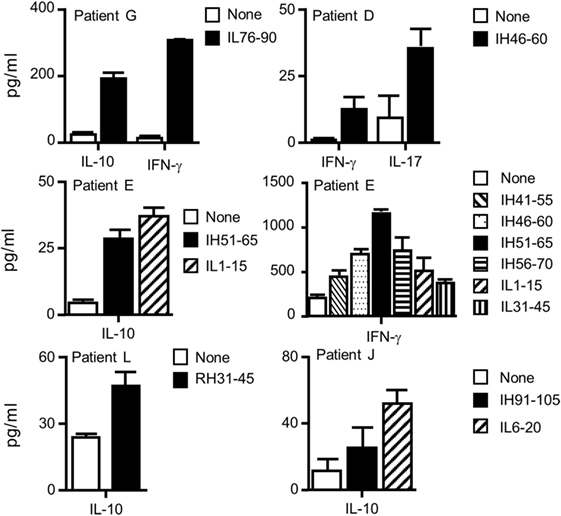
Figure 7. Cytokine profiles of the T cells collected from patients with antidrug antibodies against rituximab or infliximab. Supernatants of IL-5 ELISPOT plates were submitted to Luminex cytokine assays (R&D) for six different cytokines (IL-2, IL-4, IL-10, IL-13, IL-17, and interferon-γ). Only positive responses as defined in the Section “Materials and Methods” are presented.
Discussion
With the perspective to anticipate immunogenicity of therapeutic antibodies, we identified CD4 T cell epitopes of the immunogenic chimeric antibodies Ifx (8–10) and Rtx (5–7) from healthy donors. We evaluated the capacity of the CD4 T cell epitopes identified in the healthy donors to stimulate T cells collected from patients with ADA. We evaluated the binding affinity of overlapping VH and VL peptides to preponderant HLA-DR molecules and identified naturally presented HLA-DR-associated peptides displayed by human DCs. The main results of this comprehensive analysis of the CD4 T cell epitopes of Rtx and Ifx are summarized in Figure 8.

Figure 8. Maps of the T cell epitopes of rituximab and infliximab. T cell epitope sequences identified using cells collected in healthy donors (red) or in patients with antidrug antibodies (green) were reported, each bar corresponding to an individual response. Black: cluster identified by MHC-associated peptide proteomics assay. Occurrence of each cluster among the donors tested is indicated inside each bar.
Although the immunogenicity of therapeutic antibodies such as Ifx and Rtx is a limitation for their clinical use, their CD4 T cell epitopes have not yet been identified. We therefore isolated specific T cell lines from CD4 T lymphocytes collected from healthy donors by multiple rounds of in vitro antigenic stimulation (28), while memory CD4 T cell response was investigated in ADA patients using a short-term T cell assay. For each antibody, the number of T cell lines derived from each healthy donor ranged from 0 to 6, in agreement with the very low frequency of Ifx- and Rtx-specific T cells preexisting in the blood of healthy donors (28). Nine T cell epitopes were found in both antibodies, spreading over 25–58% of the VH and VL sequences (Figure 8). They mainly overlap CDR regions, but RH71–85, RL1–15, IL1–15, and IL6–20 are entirely embedded in the FR regions. The wide spread of the T cell epitopes along the variable parts of Rtx and Ifx and not only the CDR is consistent with their murine origin as their FR regions could be recognized as foreign by the human immune system (3). The location of Ifx and Rtx T cell epitopes also seems to rely on HLA binding affinity, as most of the T cell epitopes appeared to be associated with strong or moderate peptide binding to HLA molecules (Table 1). However, as previously observed for other antigens (33, 38), including therapeutic proteins (25), not all of HLA binders stimulated T cells (Figure 4). Naturally presented peptide sequences retrieved from the MAPPs assay (34–37) span a more restricted portion of the VH and VL sequences as compared to HLA-DR binding regions. In many cases, sequences of peptides eluted from HLA-DR molecules from human DCs nicely match with the T cell epitopes (Figure 8). Only three T cell epitopes (RH106–120, IL 31–45, and IL76–90) are not included in the peptides identified by MAPPs. The assays were performed on different donor sets, which might be a reason for the mismatches. Of particular interest are immunoprevalent T cell epitopes (39) that are common to multiple donors. Immunoprevalence of Ifx IH46–60 and Rtx RL41–55 seemed to result from their capacity to bind multiple HLA class II molecules corresponding to promiscuous T cell epitopes. Rtx RL41–55 was also naturally presented by more than half of the donors tested via MAPPs. In contrast, the immunoprevalent Rtx peptide RH56–70 bound strongly only to the HLA-DRB1*11:01 molecule, and in the MAPPs assay the corresponding sequence region was displayed by only one donor who carried the same allele. Accordingly, the three donors responding to this peptide were genotyped HLA-DRB1*11:01. Together, our data from HLA-DR binding experiments, natural peptide presentation experiments, and T cell epitope mapping experiments show that the presented peptide repertoire is smaller than the peptide binding repertoire, and that the T cell epitope repertoire appears to be smaller than the presented peptide repertoire.
We also provide new insights into the specificity of the T cell response to Rtx and Ifx in patients having developed ADAs. All patients generated a T cell response to either Rtx or Ifx peptides, confirming that the ADA response is a T-cell-dependent process (16–20). Eight out of nine Ifx T cell epitopes and four out of nine Rtx T cell epitopes, which we identified in healthy blood donors contributed to the T cell response in patients having developed ADAs. As the healthy donors have never been treated with these antibodies, the T cell epitopes we identified from the cells collected in these donors should emerge from naïve T cell cells (28). Our data are therefore consistent with previous observations in animal models using HLA tetramers (40) and in vaccinated donors (41, 42) showing that memory CD4 T cell response is shaped by the naïve repertoire. Accordingly, immunogenic therapeutic antibodies including Ifx and Rtx exhibit a larger repertoire of preexisting naïve CD4 T lymphocytes in healthy donors than most of the antibodies considered as non-immunogenic (28). T cell epitopes in therapeutic antibodies could be at least partly predicted by using cells collected from healthy donors (25–29). Such T cell assays allow therapeutic antibodies to be ranked for immunogenicity risk on the basis of their T cell epitope content. T cell epitopes could be removed from the antibody sequence to minimize their immunogenicity, as already proposed (19, 21).
Our findings also contribute to describe the diversity of the T cell response developed in patients with ADAs. Besides IL-5, we also detected secretion of IFN-γ, IL-10, and IL-17, but with different patterns of secretion across the patients. These data illustrate the heterogeneity of the T cell response to chimeric antibodies across inflammatory cytokines and the immunosuppressive cytokine IL-10 (43). Using the identified T cell epitopes, further studies would help to understand the diversity of immune responses, including the allergic response (2), to classify patients with and without an ADA response and to identify early markers of ADA response.
In conclusion, we identified the CD4 T cell epitopes of Ifx and Rtx from healthy donors and demonstrated their relevance to participate to the T cell response developed in patients with ADAs. We also compared them to HLA binding data as well as to the naturally presented peptide repertoire. These data contribute to the understanding of the root cause of immunogenicity of chimeric antibodies. They could help to improve the diagnostic of ADA responses (2) and the selection of therapeutic antibodies with low immunogenicity by removing the T cell epitopes from the initial sequence.
Ethics Statement
This study was carried out in accordance with the recommendations of the guidelines of the etablissement farnçais du sang (EFS), Comité d’ethique et deontologie de EFS with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by le Comité d’ethique et deontologie de EFS.
Author Contributions
MH, SM, AK, SS, XM, and BM designed the study and interpreted the data; BM, AK, MH, and SM wrote the manuscript; MH, SM, AGd, AGo, and NS performed the experiments; BM, MP, and AK supervised the experiments; and XM, FC, and CM-R selected the patients and supervised the collection of blood samples. All authors revised the work and gave final approval of the version to be published.
Conflict of Interest Statement
AK and SS are full-time employees of Novartis. The other authors declare no conflict of interest.
The reviewer, ER, and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
Funding
The research leading to these results was supported by the Innovative Medicines Initiative Joint Undertaking ABIRISK (Anti-Biopharmaceutical Immunization Risk) project under grant agreement #115303, the resources of which comprise financial contribution from the European Union’s Seventh Framework Program (FP7/2007–2013) and in-kind contributions from EFPIA companies. This work was also supported by the Labex in Research on Medication and Therapeutic Innovation (LERMIT) (to BM, MP, and XM) and the CEA (to BM).
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2017.00500/full#supplementary-material.
Abbreviations
DCs, dendritic cells; F, female; HLA, human leukocyte antigen; HSA, human serum albumin; IFN-γ, interferon-γ; KLH, keyhole limpet hemocyanin; Ifx, infliximab; MAPPs, MHC-associated peptide proteomics; MS, mass spectrometry; PBMCs, peripheral blood mononuclear cells; Rtx, rituximab.
References
1. Bartelds GM, Krieckaert CL, Nurmohamed MT, van Schouwenburg PA, Lems WF, Twisk JW, et al. Development of antidrug antibodies against adalimumab and association with disease activity and treatment failure during long-term follow-up. JAMA (2011) 305(14):1460–8. doi: 10.1001/jama.2011.406
2. Matucci A, Pratesi S, Petroni G, Nencini F, Virgili G, Milla M, et al. Allergological in vitro and in vivo evaluation of patients with hypersensitivity reactions to infliximab. Clin Exp Allergy (2013) 43(6):659–64. doi:10.1111/cea.12098
3. Cibotti R, Kanellopoulos JM, Cabaniols JP, Halle-Panenko O, Kosmatopoulos K, Sercarz E, et al. Tolerance to a self-protein involves its immunodominant but does not involve its subdominant determinants. Proc Natl Acad Sci U S A (1992) 89(1):416–20. doi:10.1073/pnas.89.1.416
4. Hansel TT, Kropshofer H, Singer T, Mitchell JA, George AJ. The safety and side effects of monoclonal antibodies. Nat Rev Drug Discov (2010) 9(4):325–38. doi:10.1038/nrd3003
5. Pijpe J, van Imhoff GW, Spijkervet FK, Roodenburg JL, Wolbink GJ, Mansour K, et al. Rituximab treatment in patients with primary Sjogren’s syndrome: an open-label phase II study. Arthritis Rheum (2005) 52(9):2740–50. doi:10.1002/art.21260
6. Smith KG, Jones RB, Burns SM, Jayne DR. Long-term comparison of rituximab treatment for refractory systemic lupus erythematosus and vasculitis: remission, relapse, and re-treatment. Arthritis Rheum (2006) 54(9):2970–82. doi:10.1002/art.22046
7. Thurlings RM, Teng O, Vos K, Gerlag DM, Aarden L, Stapel SO, et al. Clinical response, pharmacokinetics, development of human anti-chimeric antibodies, and synovial tissue response to rituximab treatment in patients with rheumatoid arthritis. Ann Rheum Dis (2009) 69(2):409–12. doi:10.1136/ard.2009.109041
8. Baert F, Noman M, Vermeire S, Van Assche G, D’ Haens G, Carbonez A, et al. Influence of immunogenicity on the long-term efficacy of infliximab in Crohn’s disease. N Engl J Med (2003) 348(7):601–8. doi:10.1056/NEJMoa020888
9. Bendtzen K, Geborek P, Svenson M, Larsson L, Kapetanovic MC, Saxne T. Individualized monitoring of drug bioavailability and immunogenicity in rheumatoid arthritis patients treated with the tumor necrosis factor alpha inhibitor infliximab. Arthritis Rheum (2006) 54(12):3782–9. doi:10.1002/art.22214
10. Radstake TR, Svenson M, Eijsbouts AM, van den Hoogen FH, Enevold C, van Riel PL, et al. Formation of antibodies against infliximab and adalimumab strongly correlates with functional drug levels and clinical responses in rheumatoid arthritis. Ann Rheum Dis (2009) 68(11):1739–45. doi:10.1136/ard.2008.092833
11. Weinblatt ME, Maddison PJ, Bulpitt KJ, Hazleman BL, Urowitz MB, Sturrock RD, et al. CAMPATH-1H, a humanized monoclonal antibody, in refractory rheumatoid arthritis. An intravenous dose-escalation study. Arthritis Rheum (1995) 38(11):1589–94. doi:10.1002/art.1780381110
12. Feagan BG, Greenberg GR, Wild G, Fedorak RN, Pare P, McDonald JW, et al. Treatment of active Crohn’s disease with MLN0002, a humanized antibody to the alpha4beta7 integrin. Clin Gastroenterol Hepatol (2008) 6(12):1370–7. doi:10.1016/j.cgh.2008.06.007
13. Bartelds GM, Wijbrandts CA, Nurmohamed MT, Stapel S, Lems WF, Aarden L, et al. Clinical response to adalimumab: relationship to anti-adalimumab antibodies and serum adalimumab concentrations in rheumatoid arthritis. Ann Rheum Dis (2007) 66(7):921–6. doi:10.1136/ard.2006.065615
14. Bender NK, Heilig CE, Droll B, Wohlgemuth J, Armbruster FP, Heilig B. Immunogenicity, efficacy and adverse events of adalimumab in RA patients. Rheumatol Int (2007) 27(3):269–74. doi:10.1007/s00296-006-0183-7
15. Getts DR, Getts MT, McCarthy DP, Chastain EM, Miller SD. Have we overestimated the benefit of human(ized) antibodies? MAbs (2010) 2(6):682–94. doi:10.4161/mabs.2.6.13601
16. van Schouwenburg PA, Kruithof S, Votsmeier C, van Schie K, Hart MH, de Jong RN, et al. Functional analysis of the anti-adalimumab response using patient-derived monoclonal antibodies. J Biol Chem (2014) 289(50):34482–8. doi:10.1074/jbc.M114.615500
17. Jacquemin M, Vantomme V, Buhot C, Lavend’homme R, Burny W, Demotte N, et al. CD4+ T-cell clones specific for wild-type factor VIII: a molecular mechanism responsible for a higher incidence of inhibitor formation in mild/moderate hemophilia A. Blood (2003) 101(4):1351–8. doi:10.1182/blood-2002-05-1369
18. Barbosa MD, Vielmetter J, Chu S, Smith DD, Jacinto J. Clinical link between MHC class II haplotype and interferon-beta (IFN-beta) immunogenicity. Clin Immunol (2006) 118(1):42–50. doi:10.1016/j.clim.2005.08.017
19. Mazor R, Eberle JA, Hu X, Vassall AN, Onda M, Beers R, et al. Recombinant immunotoxin for cancer treatment with low immunogenicity by identification and silencing of human T-cell epitopes. Proc Natl Acad Sci U S A (2014) 111(23):8571–6. doi:10.1073/pnas.1405153111
20. Stickler MM, Estell DA, Harding FA. CD4+ T-cell epitope determination using unexposed human donor peripheral blood mononuclear cells. J Immunother (2000) 23(6):654–60. doi:10.1097/00002371-200011000-00006
21. Yeung VP, Chang J, Miller J, Barnett C, Stickler M, Harding FA. Elimination of an immunodominant CD4+ T cell epitope in human IFN-beta does not result in an in vivo response directed at the subdominant epitope. J Immunol (2004) 172(11):6658–65. doi:10.4049/jimmunol.172.11.6658
22. Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, et al. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices [see comments]. Nat Biotechnol (1999) 17(6):555–61. doi:10.1038/9858
23. Wang P, Sidney J, Dow C, Mothe B, Sette A, Peters B. A systematic assessment of MHC class II peptide binding predictions and evaluation of a consensus approach. PLoS Comput Biol (2008) 4(4):e1000048. doi:10.1371/journal.pcbi.1000048
24. Paul S, Kolla RV, Sidney J, Weiskopf D, Fleri W, Kim Y, et al. Evaluating the immunogenicity of protein drugs by applying in vitro MHC binding data and the immune epitope database and analysis resource. Clin Dev Immunol (2013) 2013:467852. doi:10.1155/2013/467852
25. Tangri S, Mothe BR, Eisenbraun J, Sidney J, Southwood S, Briggs K, et al. Rationally engineered therapeutic proteins with reduced immunogenicity. J Immunol (2005) 174(6):3187–96. doi:10.4049/jimmunol.174.6.3187
26. Harding FA, Stickler MM, Razo J, DuBridge RB. The immunogenicity of humanized and fully human antibodies: residual immunogenicity resides in the CDR regions. MAbs (2010) 2(3):256–65. doi:10.4161/mabs.2.3.11641
27. Delluc S, Ravot G, Maillere B. Quantification of the preexisting CD4 T-cell repertoire specific for human erythropoietin reveals its immunogenicity potential. Blood (2010) 116(22):4542–5. doi:10.1182/blood-2010-04-280875
28. Delluc S, Ravot G, Maillere B. Quantitative analysis of the CD4 T-cell repertoire specific to therapeutic antibodies in healthy donors. FASEB J (2011) 25(6):2040–8. doi:10.1096/fj.10-173872
29. Jawa V, Cousens LP, Awwad M, Wakshull E, Kropshofer H, De Groot AS. T-cell dependent immunogenicity of protein therapeutics: preclinical assessment and mitigation. Clin Immunol (2013) 149(3):534–55. doi:10.1016/j.clim.2013.09.006
30. Maloney DG, Grillo-Lopez AJ, White CA, Bodkin D, Schilder RJ, Neidhart JA, et al. IDEC-C2B8 (rituximab) anti-CD20 monoclonal antibody therapy in patients with relapsed low-grade non-Hodgkin’s lymphoma. Blood (1997) 90(6):2188–95.
31. Texier C, Pouvelle S, Busson M, Herve M, Charron D, Menez A, et al. HLA-DR restricted peptide candidates for bee venom immunotherapy. J Immunol (2000) 164(6):3177–84. doi:10.4049/jimmunol.164.6.3177
32. Texier C, Pouvelle-Moratille S, Buhot C, Castelli FA, Pecquet C, Menez A, et al. Emerging principles for the design of promiscuous HLA-DR-restricted peptides: an example from the major bee venom allergen. Eur J Immunol (2002) 32(12):3699–707. doi:10.1002/1521-4141(200212)32:12<3699:AID-IMMU3699>3.0.CO;2-V
33. Chevaleyre C, Benhamouda N, Favry E, Fabre E, Mhoumadi A, Nozach H, et al. The tumor antigen cyclin B1 hosts multiple CD4 T cell epitopes differently recognized by pre-existing naive and memory cells in both healthy and cancer donors. J Immunol (2015) 195(4):1891–901. doi:10.4049/jimmunol.1402548
34. Kropshofer H, Spindeldreher S, Rohn TA, Platania N, Grygar C, Daniel N, et al. Tetraspan microdomains distinct from lipid rafts enrich select peptide-MHC class II complexes. Nat Immunol (2002) 3(1):61–8. doi:10.1038/ni750
35. Rohn TA, Reitz A, Paschen A, Nguyen XD, Schadendorf D, Vogt AB, et al. A novel strategy for the discovery of MHC class II-restricted tumor antigens: identification of a melanotransferrin helper T-cell epitope. Cancer Res (2005) 65(21):10068–78. doi:10.1158/0008-5472.CAN-05-1973
36. Rombach-Riegraf V, Karle AC, Wolf B, Sorde L, Koepke S, Gottlieb S, et al. Aggregation of human recombinant monoclonal antibodies influences the capacity of dendritic cells to stimulate adaptive T-cell responses in vitro. PLoS One (2014) 9(1):e86322. doi:10.1371/journal.pone.0086322
37. Karle A, Spindeldreher S, Kolbinger F. Secukinumab, a novel anti-IL-17A antibody, shows low immunogenicity potential in human in vitro assays comparable to other marketed biotherapeutics with low clinical immunogenicity. MAbs (2016) 8(3):536–50. doi:10.1080/19420862.2015.1136761
38. Castelli FA, Leleu M, Pouvelle-Moratille S, Farci S, Zarour HM, Andrieu M, et al. Differential capacity of T cell priming in naive donors of promiscuous CD4+ T cell epitopes of HCV NS3 and core proteins. Eur J Immunol (2007) 37(6):1513–23. doi:10.1002/eji.200636783
39. Castelli FA, Houitte D, Munier G, Szely N, Lecoq A, Briand JP, et al. Immunoprevalence of the CD4+ T-cell response to HIV Tat and Vpr proteins is provided by clustered and disperse epitopes, respectively. Eur J Immunol (2008) 38(10):2821–31. doi:10.1002/eji.200738072
40. Moon JJ, Chu HH, Pepper M, McSorley SJ, Jameson SC, Kedl RM, et al. Naive CD4(+) T cell frequency varies for different epitopes and predicts repertoire diversity and response magnitude. Immunity (2007) 27(2):203–13. doi:10.1016/j.immuni.2007.07.007
41. Kwok WW, Tan V, Gillette L, Littell CT, Soltis MA, LaFond RB, et al. Frequency of epitope-specific naive CD4(+) T cells correlates with immunodominance in the human memory repertoire. J Immunol (2012) 188(6):2537–44. doi:10.4049/jimmunol.1102190
42. Castelli FA, Szely N, Olivain A, Casartelli N, Grygar C, Schneider A, et al. Hierarchy of CD4 T cell epitopes of the ANRS Lipo5 synthetic vaccine relies on the frequencies of pre-existing peptide-specific T cells in healthy donors. J Immunol (2013) 190(11):5757–63. doi:10.4049/jimmunol.1300145
Keywords: therapeutic antibodies, immunogenicity, antidrug antibody, CD4 T cell epitopes, healthy donors, infliximab, rituximab, MHC-associated peptide proteomics
Citation: Hamze M, Meunier S, Karle A, Gdoura A, Goudet A, Szely N, Pallardy M, Carbonnel F, Spindeldreher S, Mariette X, Miceli-Richard C and Maillère B (2017) Characterization of CD4 T Cell Epitopes of Infliximab and Rituximab Identified from Healthy Donors. Front. Immunol. 8:500. doi: 10.3389/fimmu.2017.00500
Received: 12 February 2017; Accepted: 11 April 2017;
Published: 05 May 2017
Edited by:
Laurent Rénia, Agency for Science, Technology and Research (A*STAR), SingaporeReviewed by:
Alejandro Balsa, Hospital Universitario La Paz, SpainEe Chee Ren, Agency for Science, Technology and Research (A*STAR), Singapore
Copyright: © 2017 Hamze, Meunier, Karle, Gdoura, Goudet, Szely, Pallardy, Carbonnel, Spindeldreher, Mariette, Miceli-Richard and Maillère. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bernard Maillère, YmVybmFyZC5tYWlsbGVyZUBjZWEuZnI=