 Stéphanie Graff-Dubois
Stéphanie Graff-Dubois Angeline Rouers
Angeline Rouers Arnaud Moris
Arnaud Moris
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 11 November 2016
Sec. HIV and AIDS
Volume 7 - 2016 | https://doi.org/10.3389/fimmu.2016.00501
This article is part of the Research Topic CD4 T follicular helper cells in HIV View all 10 articles
The discovery of broad and potent HIV-1 neutralizing antibodies (bNAbs) has renewed optimism for developing an effective vaccine against HIV-1. The generation of most bNAbs requires multiple rounds of B cell receptor affinity maturation, suggesting a crucial role of follicular helper T (Tfh) cells in their production. However, less than 1% of HIV-infected patients develop bNAbs that arise late in the course of infection, indicating probable Tfh and B cell dysfunctions in this context. Since the last few years, many studies have characterized Tfh cells from lymph nodes and spleen of HIV-infected individuals and SIV-infected macaques. Various lymphoid Tfh cell subsets have been identified, including precursor Tfh (pTfh), germinal center Tfh (GC Tfh), and the regulatory counterpart of Tfh cells, the follicular regulatory T cells. The latter have been reported to play a crucial role in the control of T and B cell crosstalk and GC reactions. More recently, circulating Tfh-like cells (cTfh) have been identified. Meanwhile, advances in single-cell technologies have made possible to analyze the transcriptional profiles of low abundant cells, such as Tfh populations. Using transcriptional signatures, we review here the impact of chronic SIV/HIV infection on Tfh, GC Tfh, pTfh, and cTfh differentiation and helper T cell functions with regard to their capacity to induce efficient B cell maturation. We will explore some hypothesis to explain the increased proportion of Tfh cells reported in chronically infected individuals and the impact on HIV pathogenesis.
In germinal centers (GC), T follicular helper (Tfh) cells deliver helper signals and cytokines required for B cell affinity maturation and B cell differentiation into long-lived plasma cells. Optimal Tfh and B cell crosstalk is a prerequisite for the induction of efficient humoral immunity to pathogens. By providing survival and differentiation signals, Tfh cells control multiple steps of B cell maturation and antibody (Ab) production.
In addition to the cognate antigen interaction with B cells, Tfh cells express costimulatory molecules, such as CD40L, ICOS, and OX40. Tfh cells secrete high levels of interleukin-21 (IL-21) and IL-4, which are necessary for GC formation and B cell differentiation into long-lived plasma cells, respectively (1–3).
Among tissue-resident Tfh cell subsets, early committed precursor Tfh (pTfh) and germinal center Tfh (GC Tfh) represent two different stages of the Tfh cell maturation. Follicular regulatory T (Tfr) cells are identified as the regulatory counterpart of Tfh cells. Tfr cells control T and B cell crosstalk and GC reactions. Blood circulating Tfh cells (cTfhs) have been recently identified as a memory compartment of tissue-resident Tfh cells. Like tissue-resident Tfh cells, cTfhs are endowed with the capacity to produce IL-21 and to provide B cell help (4).
Since the last 5 years, Tfh cells have been extensively studied in the lymph nodes (LNs) and spleens of individuals with chronic HIV/SIV infection. HIV infection is associated with an altered B cell differentiation (5) and Tfh isolated from LNs of HIV-infected (HIV+) individuals provide inadequate B cell help in vitro (6). As lymphoid tissue-resident Tfh cells are targeted by HIV/SIV early after infection, they constitute a major compartment for HIV infection, replication, and production of viral particles in LNs of viremic individuals (7–9), even though in vivo production of viral particles by Tfh cells remains to be demonstrated. Likewise, in blood, within central memory CD4T cells, cTfh cells serve as HIV reservoir in chronic HIV-infected individuals under antiretroviral therapy (10). Very recently, in natural HIV controllers, study of HIV infection in various CD4 T cell subsets demonstrates various mechanism of HIV persistence according to the CD4 T cell compartment (11). LN-resident helper T cells (Tfh and non-Tfh) showed replicative virus, while clonally expanded blood CD4 T cells harbor inducible provirus (11). However, despite their high susceptibility to HIV/SIV infection, many studies reported an accumulation of tissue-resident or cTfh populations during the chronic phases of infection (7, 8, 12, 13). In addition, Hong et al. demonstrate that after a rapid expansion of GCs during the acute phase, slowly proliferative Tfh cells accumulate during the chronic phase of SIV infection (14).
Various hypotheses can support the higher proportions of Tfh cell subsets in the context of chronic HIV infection: (i) Tfh cells might present high proliferative or survival capacities; (ii) antigen persistence could drive CD4 T cells toward Tfh differentiation; and (iii) regulatory cells that control the Tfh/B cell crosstalk might be defective.
Here, we propose to review recent studies based on transcriptional analysis of Tfh cell subsets and to discuss the potential consequences on GC deregulations reported in chronic HIV/SIV infection.
The signals involved in Tfh cell differentiation include TCR activation, costimulation, cytokines, and migration-associated molecules. However, the origin of Tfh cells is not well defined in humans: it is not clear whether Tfh fate is established at the time of DC priming or later. Here, we review the impact of HIV infection on Tfh cell differentiation, from the priming of CD4 T cells by DCs cells until their ultimate stage of differentiation corresponding to GC Tfh and circulating memory Tfh. Two distinct differentiation pathways have been described (Figure 1).
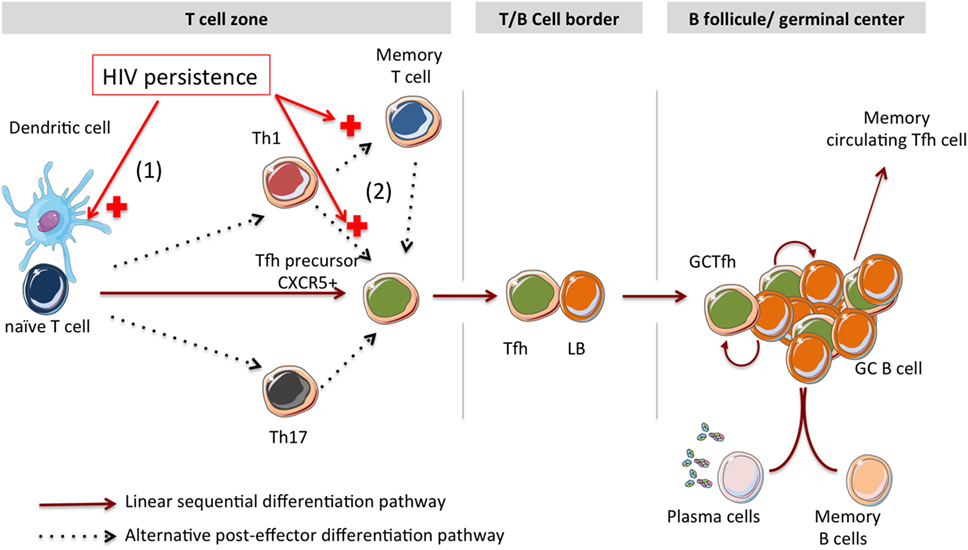
Figure 1. Impact of HIV antigen persistence on Tfh cell differentiation in lymphoid tissues. Linear and alternative post-effector Tfh cell differentiation pathways are described. (1) Interactions between HIV particles and DC-SIGN expressing DCs could support the T helper cell differentiation toward a Tfh polarization. (2) According to the alternative post-effector differentiation pathway, HIV persistence might support Th1 and memory T cell differentiation into Tfh cells.
The linear multistage Tfh differentiation pathway implicates multiple antigen-specific interactions in secondary lymphoid organs: (i) DC priming of naïve T cells leads to the rise of pTfh cells expressing CXCR5 molecule; pTfh cells migrate toward the T/B cell border zone where they experience (ii) a second antigen-specific interaction with B cells. This interaction leads to the progression of pTfh cells within the B cell follicle and differentiation into Tfh cells. (iii) In the B cell follicle, Tfh cells experience multiple interactions with B cells, leading to B cell maturation and the complete differentiation of Tfh cells into GC Tfh. Thus in this model, B cells appear central in the terminal differentiation of Tfh cell into GC Tfh and reciprocally, Tfh are also required for B cell maturation. As with other helper T cell subsets, the stimulatory cytokines produced by DCs during the priming of naïve T cells are critical parameters of Tfh cell differentiation. Using monocyte-derived DCs (MoDCs), Schmitt et al. demonstrated the key role of IL-12-producing MoDCs in the induction of IL-21-producing Tfh-like cells (15). More recently, the same group found that TGF-β acts together with IL-12 and IL-23 to induce the expression of various molecules associated with Tfh functions by human naïve helper T cells, including CXCR5, ICOS, IL-21, Bcl-6, and the transcription factors BATF and c-Maf (16). However, little is known about the type of DCs responsible for inducing Tfh cell priming. A recent study reported that engagement of DC-SIGN by fucose-based PAMPs licenses DCs for inducing Tfh polarization (17). Such activated DCs produce IL-27, which is essential for Tfh polarization. This finding highlights the importance of adjuvants in the induction of Tfh cells. Interestingly, HIV particles bind DC-SIGN through Gp120, the viral envelope (18). Therefore, one can hypothesize that, under chronic HIV infection, interactions between HIV particles and DC-SIGN expressing DCs could support the T helper cell differentiation toward a Tfh polarization. A recent study also showed that treatment with CpG (TLR-9 ligand) induces IL-6 production by MoDC, orientating helper T cells differentiation toward the Tfh-cell lineage (19). Indeed, by inducing Bcl6 early during the T cell activation, IL-6 has been shown to be critic for Tfh polarization (20). In the context of HIV/SIV infection, several groups reported higher plasma levels of IL-6 (13, 21). However, we and others (22) did not find any difference in the amount of secreted IL-6 between HIV-infected and -uninfected spleens upon activation (8). Once engaged into Tfh differentiation, the sequential differentiation model proposes that pTfh/B cell interactions dictate the fate of Tfh cells. Several groups highlighted the requirement of antigen presentation by B cells to induce Tfh cell and in turn GC reactions. In the absence of B cells, DC restricted antigen presentation initiates Tfh cell differentiation (into pTfh) but fails to complete ultimate effector Tfh cell differentiation (23). In a model of bone marrow chimera, B cells deficient for the expression of MHC-II-molecules exhibit a reduced capacity to initiate T cell expansion and differentiation (24). In fact, sequential antigen-specific interactions of Tfh cell with DC and B cells are required to initiate Tfh cell and GC differentiation (25), and antigen persistence sustains Tfh responses and GC reactions (26). Using live multiphoton imaging, Schwickert et al. suggested that the amount of peptide–MHC (pMHC) complexes presented by antigen-specific B cells to cognate T cells, at the B-cell–T-cell border, was a limiting factor regulating the entry of B cell clones into GC (27). Furthermore, highlighting the critical role of MHC-II molecules expressed by B cells in the generation of Abs of diverse functions and of memory B-cell responses, B cells lacking MHC-II expression are unable to differentiate into memory cells and are defective in producing antigen-specific IgG (28). These results demonstrate that MHC-II-restricted antigen presentation by B cells is strictly required for B cells to receive help by antigen-specific Tfh cells, and thus to establish a potent humoral immune response. Therefore, Tfh cell differentiation and GC development require the combination of DC and B cell antigen presentation.
As DC during T cell priming, B cells also provide additional signals to Tfh cells, contributing to their helper functions and maintenance. These signals include CD40L/CD40, OX40/OX40L, signaling lymphocyte activation molecule (SLAM) family members, and adhesion molecules that strengthen GC Tfh/GC B cell interaction. Interaction between ICOS and ICOS ligand (ICOSL) as well as IL-21 production has been implicated in GC formation (29, 30). PD-1/PD-1 ligand interactions also control Tfh and GC B cell differentiation (1, 31). Murine Tfh cells also express the nutrient transporter folate R 4 (FR4) and CD73 (32) although their functional relevance for Tfh cell differentiation and B cell help has not yet been uncovered. In sum, the sequential differentiation proposes that combined interactions with DC and B cell dictate the fate of Tfh cells.
The alternative “post effector” developmental pathway proposes that Tfh-like cells may develop either from the memory CD4 T cell lineage (33, 34) or from effector T helper cell subsets (35, 36), rather than arising from pTfh cells. It has been shown that Tfh and central memory T cells (TCM) are similar in their developmental pathway, including the requirement of Bcl6 and low levels of IL-2 signaling (37). In line with this, Tfh and TCM gene programs can co-initiate from effector Th1 cells upon increased Bcl-6 expression in response to a decrease of IL-2, resulting in a “Tfh/TCM-like” population. IL-7 signaling also acts as a negative feedback that downregulates the differentiation of Th1 into Tfh-like cells (38). Interestingly, in spleens from HIV-infected individuals with a high proportion of Tfh cells, we reported a markedly reduced expression of the IL-7r encoding gene in all CD4 T cell populations (8). Taken together these observations support the hypothesis that, in addition to Tfh cells, other T helper populations may contribute to B cell maturation into long-lived plasma cells. Hence, in humans, the precursors of Tfh cells might be composed of heterogeneous cell populations, which have the ability to differentiate into distinct types of Tfh cells. The latter keep some functional imprint from the parental T cell subtype. Of note, in this alternative pathway, interactions with antigen-presenting B cells are still a key event of the Tfh cell orientation thus raising the question of the impact of Tfh infection by HIV on the B cell compartments.
Germinal center Tfh cells have long been considered as the terminal stage of tissue-resident Tfh cell differentiation. Newly identified, memory Tfh cells are preferentially located in secondary lymphoid organs and bone marrow although they can recirculate in the blood. These cTfh involve several subsets that differentially support Ab secretion (4) and are related to lymphoid-tissue-resident Tfh cells by their gene expression profile, cytokine production, and functional properties (39). Recently, adding to the CXCR5 and PD-1 canonical markers, Schultz et al. proposed that cTfh can be identified by their ability to produce IL-21, the cardinal Tfh cytokine (40). Interestingly, activated memory B cells induce rapid re-expression of Bcl6 by memory Tfh cells (41), reinforcing the concept that many features of Tfh cells are highly linked with those of the B cells. Thanks to their accessibility and relative high frequencies, cTfh cell dynamics and features are the focus of growing interest in the context of infection and vaccination.
Follicular helper T cell homeostasis is critical to the induction of high affinity Ab responses that are devoid of self-reactivity. Indeed, optimal Tfh cell frequency imposes competition between B cells, thus favoring survival of high affinity B cell clones. Several cell populations maintain Tfh cell homeostasis, including regulatory T (Treg) cells, Tfr cells, CD8 regulatory cells, and plasma cells (42). Tfr cells are identified as the main T cell subset implicated in the control of Tfh cells. They migrate into follicles and directly control GC reaction (43, 44). Hence, many studies have demonstrated increased GC and T cell responses in the absence of Tfr (43, 45, 46). Tfr cells co-express Bcl6 and Blimp-1 (43) that is known to negatively regulate Tfh cell differentiation pathway (47). Indeed, Blimp-1 represses Bcl-6 and reciprocally, which might explain the lower expression of Bcl-6 in Tfr cells as compared to Tfh cells (43). Tfr cells express CTLA-4 and produce high amounts of IL-10. They have been shown to arise from Foxp3+ precursors that highjack the Tfh differentiation pathway. However, a recent study showed that, using an appropriate vaccine adjuvant, Tfr cells can derive directly from naïve CD4 T cells (48). Alteration of Tfr cells functionality might contribute to higher proportions of Tfh cells during HIV-infection. Our results indicated that HIV infection did not impact splenic Tfh/Tfr ratio suggesting that Tfr and Tfh cell subsets expended equally during HIV infection (8). However, Chowdhury et al. have shown a limited expansion of Tfr cells as compared to the one of Tfh cells during SIV infection (49). They explored the transcriptional profile of CXCR5+ PD1hiCD127-CD25+ Tfr cells after SIV infection. Overall, genes linked with Tfh differentiation and functions, such as PD-1, IL-6R, SLAMF6, and CD84, were more expressed in Tfr cells, while expression of IL-2RA linked with Treg functions was reduced after SIV infection suggesting that SIV infection might impair expression of genes associated with Treg and thus Tfr regulatory functions (49).
According to their transcription profile, Tfr cells are situated between Tfh and Treg cell subsets. However, foxp-3 expression is not taken into account in most Tfh cell studies that de facto include Tfr subset among Tfh cells. Recently, adding to Tfr cells, Treg cells expressing CTLA-4 have been reported as major inhibitors of B cell expression of CD80 and CD86, which are essential to the induction of Tfh cells (50–52).
Follicular helper T and cTfh cells are targeted by HIV/SIV very early after infection and constitute a major compartment for HIV replication and production of viral particles in LNs and periphery of viremic individuals (7–9, 11). Despite their high susceptibility to HIV/SIV infection, most studies reported an accumulation of tissue-resident or cTfh cell populations (7, 8, 12, 13). Interestingly, the Tfh cell frequency positively correlates with plasma viremia levels (7, 12), and Tfh cell accumulation is reduced in individuals that control SIV infection (53), suggesting that the persistence of viral antigens might drive Tfh cell expansion. Accordingly, cTfh cell expansion has been recently reported in untreated individuals while the frequency of cTfh cells is restored to normal levels under cART suggesting that HIV replication also drives cTfh cell dynamics (10). Most studies report an increase of Tfh cells among memory CD4 T cells during HIV/SIV infection, whereas others conclude with the opposite statement (9, 54). In SIV-infected rhesus macaques, Moukambi et al. recently showed that Tfh dynamics differs from one compartment to another (peripheral blood vs. LNs or spleen) (9). Moreover, the Tfh cell frequency varies according to (i) the stage of HIV/SIV infection (53), (ii) the severity of the disease, and (iii) the ability to develop broadly neutralizing antibodies (bNAbs) (39, 55). In Table 1, we summarized Tfh cell dynamics from various studies taking into account: the type of infection (HIV/SIV), the phase (acute vs. chronic), the disease progression (slow vs. fast), the immune compartment (peripheral blood vs. secondary lymphoid organs), the phenotype, and the antigen specificity of the Tfh cells. Irrespective of the immune compartment (LNs, spleen, or blood), Tfh cells are preserved in HIV/SIV controllers, displaying no Tfh cell accumulation or loss. On the contrary, Tfh cell loss is reported in fast progressors as well as in the late stages of disease.
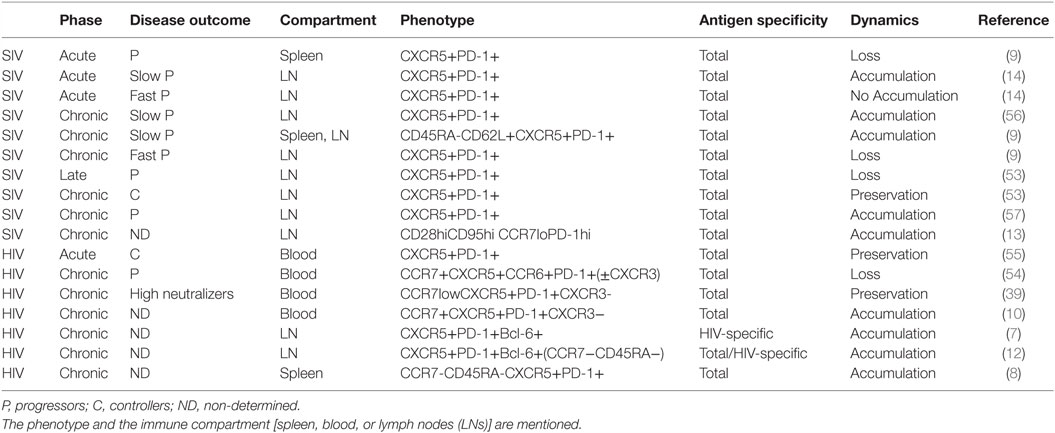
Table 1. Tfh cell dynamics in HIV/SIV infection according to the stage (acute, chronic, or late) and the outcome of the disease.
Follicular helper T cell accumulation is reported during slow progression (SP) or chronic stage of the disease. Indeed, evidences support the pivotal role of persistent viral antigen within the GC in driving Tfh cell expansion. HIV particles are associated with FDC in tonsils and LNs from infected patients (58–60) and Cheynier et al. reported the persistence of high levels of HIV particles in GC of HIV+ spleens from untreated subjects (61). In addition to FDC-bound virions, opsonized HIV particles interact with B cells trough CD21 membrane receptor (62, 63). Remarkably, the accessibility of CTL to GC is reduced, thus limiting the elimination of HIV-infected cells (64). Therefore, B cell follicles locally concentrate cell subsets implicated in HIV replication and viral production, which maintain antigen persistence and GC reactions. It appears that antigen persistence sustains ongoing GC reactions in which Tfh and GC B cell frequencies are highly correlated (8, 12, 65). However, a limited number of fully functional Tfh cells is required for the induction of bNAbs (42).
Another clue supporting the pivotal role of GC in driving Tfh cell expansion is that the disruption of GC organization coincides with the loss of Tfh cells and the onset of AIDS in terminal stages of SIV infection (53). PD1/PD-L2 axis contributes to the survival of Tfh and B cells. Interestingly, the expression of PD-L2 on B cells is severely impacted in the late stages of SIV infection potentially contributing to a decreased survival of T and B cells and the termination of GC reaction (53).
In sum, increased proportions of Tfh cells do not necessarily results in a better immune control of HIV infection, and Tfh cell proportions must be tightly regulated to allow efficient maturation and selection of B cells displaying high B cell receptor (BCR) affinity. Tfh cell functions need to be preserved to allow the production of potent bNAbs.
The key role of Tfh cells is to provide B cell helper signals and to promote their differentiation into memory B cell displaying high affinity for pathogens. These signals consist of production of cytokines, such as IL-4 and IL-21, and the expression of cell surface molecules, such as OX40, ICOS, and CD40L, by Tfh cells (1). As other groups, we analyzed the transcriptome profiles of Tfh as a mean to assess potential Tfh dysfunctions (see references in Table 2). Using single-cell sorting and high-throughput PCR (Fluidigm BioMark HD), we showed that expression of genes implicated in splenic Tfh and GC Tfh cell functions are deeply impacted by chronic HIV infection (8). In this section, we intend to review the impact of HIV/SIV infection on the main signals implicated in Tfh cell functions.
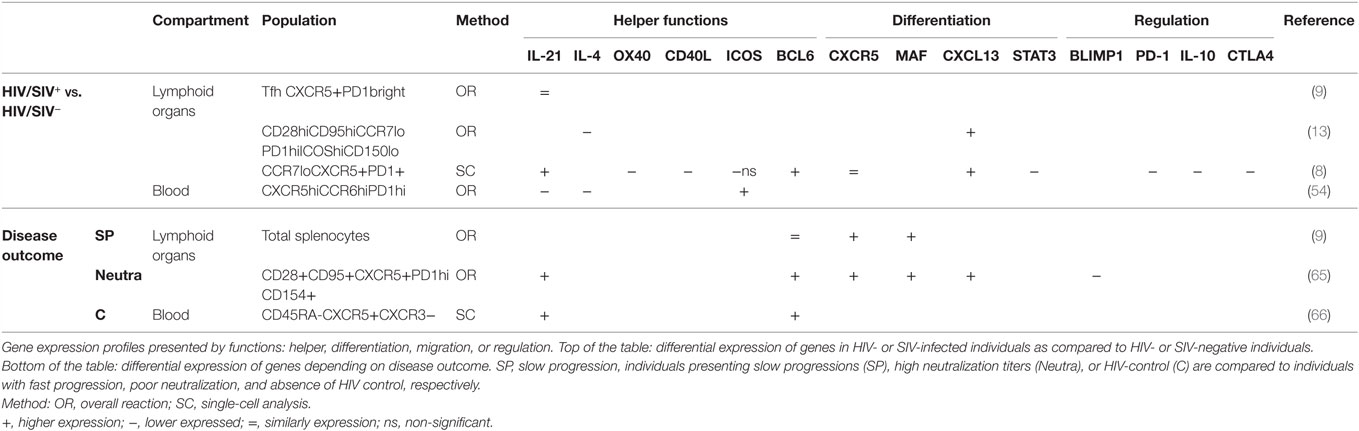
Table 2. Transcriptional profiles of Tfh cells in HIV/SIV-infection.
CD40L–CD40 (expressed by Tfh and B cell, respectively) interactions are required for the induction and maintenance of GC reaction. Blocking this molecular axis leads to GC disruption (67). In line with this, mutation in the CD40L gene is responsible for the X-linked hyper-IgM syndrome in humans characterized by a markedly decreased serum concentrations of IgA, IgE, and IgG (68). Our transcriptional data showed that the expression of CD40L gene is severely impacted in Tfh and GC Tfh from HIV-infected spleens (8). Of note, CD40L down modulation has been reported in global CD4+ T cell population during the late stages of HIV infection (69) and our unpublished data.
OX40–OX40L interaction is required for B cell differentiation into plasma cells (70). In humans, mutations in OX40 gene lead to decreased proportion of circulating memory B cells but do not impact the Ab responses (71). HIV-infected spleens exhibit defective expression of gene encoding OX40 in Tfh and GC Tfh cells (8). Intriguingly, we reported a reduction of memory B cell compartment in chronically HIV-infected individuals. Whether OX40 defective expression by Tfh is involved in this decrease in memory B cell should be further investigated.
During GC reactions, the expression ICOS by Tfh cells plays a major role in the process of selection of high affinity B cells. ICOS ligation leads to the overexpression of CD40L by Tfh cells that, in turn, promote the expression of ICOSL by GC B cells (72). In mice, recent findings emphasize the crucial role of T and B cell interactions through ICOS–ICOSL and CD40L–CD40 molecular axis in the maintenance of GC reactions and the production of high affinity bone-marrow plasma cells. ICOSL has been identified as a key regulator of positive selection of high affinity B cells during T–B cell interaction. Noteworthy, in comparison with uninfected donors, ICOS expression is enhanced in cTfh of ART-treated HIV-infected individuals (54) suggesting an overall immune activation of cTfh in those patients.
IL-21 is considered as the cardinal cytokine of the Tfh cell population. Tfh-secreted IL-21 induces B cell affinity maturation (73). A defective production of IL-21 by Tfh cells as well as a defective expression of its receptor by B cells severely impacts B cell proliferation and their differentiation into plasma cells (74). In the context of HIV infection, circulating CD4+ T cells secreting IL-21 are defined as the closest relative of tissue-resident (from secondary lymphoid organs) Tfh cells, both phenotypically and transcriptionally (40). cTfh cells from chronically HIV-infected individuals present altered expression of IL-21 gene (54) suggesting defective helper function. Conversely, we showed higher level of IL-21 transcripts in Tfh cells from chronically HIV-infected spleens (8) while Moukambi et al. reported a similar level of expression between Tfh from uninfected macaques and SIV-infected macaques during the early and the chronic phases of SIV infection (9). Discrepancies concerning the stage of the disease as well as the immune compartment (blood, spleen, and LNs) might explain these conflicting observations. Higher levels of IL-21 transcripts in cTfh cells are associated with HIV-controller status (66) or with cross-reactive neutralizing responses in rhesus macaques during the chronic phase of SIV infection (34–50 weeks post infection) (65).
IL-4 plays a major role in Tfh function (1) and confers anti-apoptotic properties to GC B cells that are particularly susceptible to cell death by apoptosis (75). IL-4 also participates to GC B cells selection (76). Tfh cells from SIV-infected rhesus macaques showed a marked reduction of IL-4 gene expression, and a global alteration of genes implicated in the network associated with IL-4 pathway (13). A defective expression of IL-4 gene has also been reported in cTfh from ART-treated HIV-infected individuals (54). We have shown a decreased secretion of IL-4 and IL-10 in splenocytes from chronically HIV-infected individuals (8). IL-10 contributes to B cell differentiation (77). Altered IL-10 secretion was also confirmed at the transcriptional level (8). However, our experimental settings did not allow the discrimination of Tfh cells from the Tfr subset and IL-10 deficiencies might also be related to defective Tfr functions.
SLAMF1 is specifically required for IL-4 and IL-21 production by GC Tfh cells (78), and SAP (SLAM-associated protein) is essential for GC reaction by stabilizing interaction between Tfh and B cells (79). In spleens from HIV-infected donors, we also observed a defective SLAMF1 gene expression in Tfh cell subsets that might explain the low level of IL-4 production by splenocytes from HIV-infected patients (8).
Altogether, these results indicate that HIV infection severely impacts Tfh cell functions in their capacity to provide adequate B cell help. These alterations result from both intrinsic Tfh cell defects and higher proportions of Tfh cells. Whether HIV infection alters Tfr cell functions is not clear. This issue will require further studies, especially addressing the capacities of Tfr cells to suppress Tfh cell functions or differentiation as well as B cell maturation. However, it will require identifying Tfr specific extracellular markers.
HIV/SIV infection results in antigen persistence that drives CD4 T cell differentiation toward the Tfh phenotype, thus maintaining high level of Tfh cells in GC. By controlling viremia, introduction of cART allows a normalization of cTfh cell proportions while Tfh cell functions are not fully restored. In particular, in vitro production of immunoglobulin is reduced in both untreated patients and HIV-infected individuals under cART (54). Hence, despite cART, HIV patients showing a detectable viral load (163 ± 178 HIV RNA copies/mL) did not respond to the 2009 H1N1/09 vaccine, whereas vaccine responders displayed undetectable viral loads (<40 HIV RNA copies/mL). This suggests that the persistence of viremia compromises Tfh cell functions (80). Altogether, early control of viremia appears crucial to preserve Tfh cells functions. Additionally, while B cell compartment is not recovered in chronically infected individuals under cART, early cART initiation restores the memory B cell compartment during acute HIV infection (81).
Early cART initiation during primo infection allows a dramatic decrease of cell-associated HIV DNA and thus limits establishment of the reservoir (82, 83). However, the poor penetration of cART into lymphoid tissues (84) could limit the efficiency of drugs even if cART was initiated early. To address these limitations, several approaches, mainly based on the molecular formulation of drugs, are actually envisaged. Injectable cART could overcome the limited access to lymphoid tissues. In the LATTE clinical trial, injection of combined cabotegravir and rilpivirine was well tolerated and efficient (85). However, the relevance of this approach on the persistence of the HIV reservoir has not been reported yet. Very recently, the efficient elimination of latently HIV-infected cells using HIV protease-sensitive toxin nanocapsules has been reported (86). This strategy presents the advantage to specifically eliminate HIV-infected cells without impacting healthy cells, allowing a less invasive approach. Combining new ART formulations with innovative route of administration could contribute to eradicate HIV reservoir from lymphoid organs.
In conclusion, HIV/SIV infections target Tfh cell subsets and severely affect Tfh cell frequency and functions, with dramatic impact on GC homeostasis. While limited number of fully functional Tfh cells is required for the induction of bNAbs, it is now well established that viral antigen persistence drives increased Tfh cell differentiation. To this regard, blocking HIV replication in lymphoid tissues might be a prerequisite to the induction of potent bNAbs. Preventing virus from entering lymphoid tissues should give another benefit to early ART initiation as well as the development of new strategies to optimize the access of ART to lymphoid tissues.
SG-D, AR, and AM wrote and approved the version to be published.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors apologize to authors and colleagues whose work was not cited. We thank lab members, Dr. B. Combadière and Pr. B. Autran for discussions. We also thank ANRS, Sidaction, UPMC, INSERM, and CNRS for continuous support.
The study was supported by Agence Nationale de Recherches sur le SIDA et les hépatites virales (ANRS).
1. Crotty S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol (2011) 29:621–63. doi:10.1146/annurev-immunol-031210-101400
2. Vogelzang A, McGuire HM, Yu D, Sprent J, Mackay CR, King C. A fundamental role for interleukin-21 in the generation of T follicular helper cells. Immunity (2008) 29:127–37. doi:10.1016/j.immuni.2008.06.001
3. Ohara J, Paul WE. Up-regulation of interleukin 4/B-cell stimulatory factor 1 receptor expression. Proc Natl Acad Sci U S A (1988) 85:8221–5. doi:10.1073/pnas.85.21.8221
4. Morita R, Schmitt N, Bentebibel S, Ranganathan R, Bourdery L, Zurawski G, et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity (2011) 34:108–21. doi:10.1016/j.immuni.2010.12.012
5. Moir S, Fauci AS. B cells in HIV infection and disease. Nat Rev Immunol (2009) 9:235–45. doi:10.1038/nri2524
6. Cubas RA, Mudd JC, Savoye A-L, Perreau M, van Grevenynghe J, Metcalf T, et al. Inadequate T follicular cell help impairs B cell immunity during HIV infection. Nat Med (2013) 19:494–9. doi:10.1038/nm.3109
7. Perreau M, Savoye A-L, De Crignis E, Corpataux J-M, Cubas R, Haddad EK, et al. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J Exp Med (2013) 210:143–56. doi:10.1084/jem.20121932
8. Colineau L, Rouers A, Yamamoto T, Xu Y, Urrutia A, Pham H-P, et al. HIV-infected spleens present altered follicular helper T cell (Tfh) subsets and skewed B cell maturation. PLoS One (2015) 10:e0140978. doi:10.1371/journal.pone.0140978
9. Moukambi F, Rabezanahary H, Rodrigues V, Racine G, Robitaille L, Krust B, et al. Early loss of splenic Tfh cells in SIV-infected rhesus macaques. PLoS Pathog (2015) 11:e1005287. doi:10.1371/journal.ppat.1005287
10. Pallikkuth S, Sharkey M, Babic DZ, Gupta S, Stone GW, Fischl MA, et al. Peripheral T follicular helper cells are the major HIV reservoir within central memory CD4 T cells in peripheral blood from chronic HIV infected individuals on cART. J Virol (2015) 90(6):2718–28. doi:10.1128/JVI.02883-15
11. Boritz EA, Darko S, Swaszek L, Wolf G, Wells D, Wu X, et al. Multiple origins of virus persistence during natural control of HIV infection. Cell (2016) 166:1004–15. doi:10.1016/j.cell.2016.06.039
12. Lindqvist M, Van Lunzen J, Soghoian DZ, Kuhl BD, Ranasinghe S, Kranias G, et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection. J Clin Invest (2012) 122:3271–80. doi:10.1172/JCI64314.action
13. Petrovas C, Yamamoto T, Gerner MY, Boswell KL, Wloka K, Smith EC, et al. CD4 T follicular helper cell dynamics during SIV infection. J Clin Invest (2012) 122:3281–94. doi:10.1172/JCI63039.factors
14. Hong JJ, Amancha PK, Rogers KA, Courtney CL, Havenar-Daughton C, Crotty S, et al. Early lymphoid responses and germinal center formation correlate with lower viral load set points and better prognosis of simian immunodeficiency virus infection. J Immunol (2014) 193:797–806. doi:10.4049/jimmunol.1400749
15. Schmitt N, Morita R, Bourdery L, Bentebibel SE, Zurawski SM, Banchereau J, et al. Human dendritic cells induce the differentiation of interleukin-21-producing T follicular helper-like cells through interleukin-12. Immunity (2009) 31:158–69. doi:10.1016/j.immuni.2009.04.016
16. Schmitt N, Liu Y, Bentebibel S-E, Munagala I, Bourdery L, Venuprasad K, et al. The cytokine TGF-β co-opts signaling via STAT3-STAT4 to promote the differentiation of human TFH cells. Nat Immunol (2014) 15:856–65. doi:10.1038/ni.2947
17. Gringhuis SI, Kaptein TM, Wevers BA, van der Vlist M, Klaver EJ, van Die I, et al. Fucose-based PAMPs prime dendritic cells for follicular T helper cell polarization via DC-SIGN-dependent IL-27 production. Nat Commun (2014) 5:5074. doi:10.1038/ncomms6074
18. Geijtenbeek TB, Kwon DS, Torensma R, van Vliet SJ, van Duijnhoven GC, Middel J, et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell (2000) 100:587–97. doi:10.1016/S0092-8674(00)80694-7
19. Chakarov S, Fazilleau N. Monocyte-derived dendritic cells promote T follicular helper cell differentiation. EMBO Mol Med (2014) 6:590–603. doi:10.1002/emmm.201403841
20. Choi YS, Eto D, Yang JA, Lao C, Crotty S. Cutting edge: STAT1 is required for IL-6-mediated Bcl6 induction for early follicular helper cell differentiation. J Immunol (2013) 190:3049–53. doi:10.4049/jimmunol.1203032
21. Kuller LH, Tracy R, Belloso W, De Wit S, Drummond F, Lane HC, et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med (2008) 5:e203. doi:10.1371/journal.pmed.0050203
22. Shive CL, Biancotto A, Funderburg NT, Pilch-Cooper HA, Valdez H, Margolis L, et al. HIV-1 is not a major driver of increased plasma IL-6 levels in chronic HIV-1 disease. J Acquir Immune Defic Syndr (2012) 61:145–52. doi:10.1097/QAI.0b013e31825ddbbf
23. Goenka R, Barnett LG, Silver JS, O’Neill PJ, Hunter CA, Cancro MP, et al. Cutting edge: dendritic cell-restricted antigen presentation initiates the follicular helper T cell program but cannot complete ultimate effector differentiation. J Immunol (2011) 187:1091–5. doi:10.4049/jimmunol.1100853
24. Crawford A, Macleod M, Schumacher T, Corlett L, Gray D. Primary T cell expansion and differentiation in vivo requires antigen presentation by B cells. J Immunol (2006) 176:3498–506. doi:10.4049/jimmunol.176.6.3498
25. Barnett LG, Simkins HMA, Barnett BE, Korn LL, Johnson AL, Wherry EJ, et al. B cell antigen presentation in the initiation of follicular helper T cell and germinal center differentiation. J Immunol (2014) 192:3607–17. doi:10.4049/jimmunol.1301284
26. Baumjohann D, Preite S, Reboldi A, Ronchi F, Ansel KM, Lanzavecchia A, et al. Persistent antigen and germinal center B cells sustain T follicular helper cell responses and phenotype. Immunity (2013) 38:596–605. doi:10.1016/j.immuni.2012.11.020
27. Schwickert TA, Victora GD, Fooksman DR, Kamphorst AO, Mugnier MR, Gitlin AD, et al. A dynamic T cell-limited checkpoint regulates affinity-dependent B cell entry into the germinal center. J Exp Med (2011) 208:1243–52. doi:10.1084/jem.20102477
28. Shimoda M, Li T, Pihkala JPS, Koni PA. Role of MHC class II on memory B cells in post-germinal center B cell homeostasis and memory response. J Immunol (2006) 176:2122–33. doi:10.4049/jimmunol.176.4.2122
29. Choi YS, Kageyama R, Eto D, Escobar TC, Robert J, Monticelli L, et al. ICOS receptor instructs T follicular helper cell versus effector cell differentiation via induction of the transcriptional repressor Bcl6. Immunity (2011) 34:932–46. doi:10.1016/j.immuni.2011.03.023.Bcl6
30. McAdam A, Greenwald RJ, Levin M, Chernova T, Malenkovich N, Ling V, et al. ICOS is critical for CD40-mediated antibody class switching. Nature (2001) 409:102–5. doi:10.1038/35051107
31. Good-Jacobson KL, Szumilas CG, Chen L, Sharpe AH, Tomayko MM, Shlomchik MJ. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nat Immunol (2010) 11:535–42. doi:10.1038/ni.1877
32. Iyer SS, Latner DR, Zilliox MJ, McCausland M, Akondy RS, Penaloza-Macmaster P, et al. Identification of novel markers for mouse CD4(+) T follicular helper cells. Eur J Immunol (2013) 43:3219–32. doi:10.1002/eji.201343469
33. Fairfax KC, Everts B, Amiel E, Smith AM, Schramm G, Haas H, et al. IL-4-secreting secondary T follicular helper (Tfh) cells arise from memory T cells, not persisting Tfh cells, through a B cell-dependent mechanism. J Immunol (2015) 194:2999–3010. doi:10.4049/jimmunol.1401225
34. Qi H, Liu D, Ma W, Wang Y, Yan H. Bcl-6 controlled TFH polarization and memory: the known unknowns. Curr Opin Immunol (2014) 28:34–41. doi:10.1016/j.coi.2014.01.016
35. Lu KT, Kanno Y, Cannons JL, Handon R, Bible P, Elkahloun AG, et al. Functional and epigenetic studies reveal multistep differentiation and plasticity of in vitro-generated and in vivo-derived follicular T helper cells. Immunity (2011) 35:622–32. doi:10.1016/j.immuni.2011.07.015
36. Oestreich KJ, Mohn SE, Weinmann AS. Molecular mechanisms that control the expression and activity of Bcl-6 in TH1 cells to regulate flexibility with a TFH-like gene profile. Nat Immunol (2012) 13:405–11. doi:10.1038/ni.2242
37. Pepper M, Pagán AJ, Igyártó BZ, Taylor JJ, Jenkins MK. Opposing signals from the Bcl6 transcription factor and the interleukin-2 receptor generate T helper 1 central and effector memory cells. Immunity (2011) 35:583–95. doi:10.1016/j.immuni.2011.09.009
38. McDonald PW, Read KA, Baker CE, Anderson AE, Powell MD, Ballesteros-Tato A, et al. IL-7 signalling represses Bcl-6 and the TFH gene program. Nat Commun (2016) 7:10285. doi:10.1038/ncomms10285
39. Locci M, Havenar-Daughton C, Landais E, Wu J, Kroenke MA, Arlehamn CL, et al. Human circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity (2013) 39:758–69. doi:10.1016/j.immuni.2013.08.031
40. Schultz BT, Teigler JE, Pissani F, Oster AF, Kranias G, Alter G, et al. Circulating HIV-specific interleukin-21(+)CD4(+) T cells represent peripheral Tfh cells with antigen-dependent helper functions. Immunity (2016) 44:167–78. doi:10.1016/j.immuni.2015.12.011
41. Ise W, Kohyama M, Schraml BU, Zhang T, Schwer B, Basu U, et al. The transcription factor BATF controls the global regulators of class-switch recombination in both B cells and T cells. Nat Immunol (2011) 12:536–43. doi:10.1038/ni.2037
42. Pratama A, Vinuesa CG. Control of TFH cell numbers: why and how? Immunol Cell Biol (2014) 92:40–8. doi:10.1038/icb.2013.69
43. Linterman MA, Pierson W, Lee SK, Kallies A, Kawamoto S, Rayner TF, et al. Foxp3(+) follicular regulatory T cells control the germinal center response. Nat Med (2011) 17:975–82. doi:10.1038/nm.2425
44. Sage PT, Francisco LM, Carman CV, Sharpe AH. The receptor PD-1 controls follicular regulatory T cells in the lymph nodes and blood. Nat Immunol (2013) 14:152–61. doi:10.1038/ni.2496
45. Wollenberg I, Agua-Doce A, Hernández A, Almeida C, Oliveira VG, Faro J, et al. Regulation of the germinal center reaction by Foxp3+ follicular regulatory T cells. J Immunol (2011) 187:4553–60. doi:10.4049/jimmunol.1101328
46. Chung Y, Tanaka S, Chu F, Nurieva RI, Martinez GJ, Rawal S, et al. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions. Nat Med (2011) 17:983–8. doi:10.1038/nm.2426
47. Johnston RJ, Poholek AC, DiToro D, Yusuf I, Eto D, Barnett B, et al. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science (2009) 325:1006–10. doi:10.1126/science.1175870
48. Aloulou M, Carr EJ, Gador M, Bignon A, Liblau RS, Fazilleau N, et al. Follicular regulatory T cells can be specific for the immunizing antigen and derive from naive T cells. Nat Commun (2016) 7:10579. doi:10.1038/ncomms10579
49. Chowdhury A, Del Rio PME, Tharp GK, Trible RP, Amara RR, Chahroudi A, et al. Decreased T follicular regulatory Cell/T follicular helper cell (TFH) in Simian immunodeficiency virus-infected rhesus Macaques may contribute to accumulation of TFH in chronic infection. J Immunol (2015) 195:3237–47. doi:10.4049/jimmunol.1402701
50. Sage PT, Paterson AM, Lovitch SB, Sharpe AH. The coinhibitory receptor ctla-4 controls B cell responses by modulating T follicular helper, T follicular regulatory, and T regulatory cells. Immunity (2014) 41:1026–39. doi:10.1016/j.immuni.2014.12.005
51. Wing JB, Ise W, Kurosaki T, Sakaguchi S. Regulatory T cells control antigen-specific expansion of Tfh cell number and humoral immune responses via the coreceptor CTLA-4. Immunity (2014) 41:1013–25. doi:10.1016/j.immuni.2014.12.006
52. Linterman MA, Denton AE. Treg cells and CTLA-4: the ball and chain of the germinal center response. Immunity (2014) 41:876–8. doi:10.1016/j.immuni.2014.12.009
53. Xu H, Wang X, Malam N, Lackner AA, Veazey RS. Persistent simian immunodeficiency virus infection causes ultimate depletion of follicular Th cells in AIDS. J Immunol (2015) 195:4351–7. doi:10.4049/jimmunol.1501273
54. Boswell KL, Paris R, Boritz E, Ambrozak D, Yamamoto T, Darko S, et al. Loss of circulating CD4 T cells with B cell helper function during chronic HIV infection. PLoS Pathog (2014) 10:e1003853. doi:10.1371/journal.ppat.1003853
55. Cohen K, Altfeld M, Alter G, Stamatatos L. Early preservation of CXCR5+ PD-1+ helper T cells and B cell activation predict the breadth of neutralizing antibody responses in chronic HIV-1 infection. J Virol (2014) 88:13310–21. doi:10.1128/JVI.02186-14
56. Hong JJ, Amancha PK, Rogers K, Ansari AA, Villinger F. Spatial alterations between CD4(+) T follicular helper, B, and CD8(+) T cells during simian immunodeficiency virus infection: T/B cell homeostasis, activation, and potential mechanism for viral escape. J Immunol (2012) 188:3247–56. doi:10.4049/jimmunol.1103138
57. Xu H, Wang X, Malam N, Aye PP, Alvarez X, Lackner AA, et al. Persistent SIV infection drives differentiation, aberrant accumulation, and latent infection of germinal center follicular T helper cells. J Virol (2015) 90:1578–87. doi:10.1128/JVI.02471-15
58. Embretson J, Zupancic M, Ribas J, Burke A, Racz P, Tenner-Racz K, et al. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS. Nature (1993) 362:359–62. doi:10.1038/362359a0
59. Pantaleo G, Cohen OJ, Schacker T, Vaccarezza M, Graziosi C, Rizzardi GP, et al. Evolutionary pattern of human immunodeficiency virus (HIV) replication and distribution in lymph nodes following primary infection: implications for antiviral therapy. Nat Med (1998) 4:341–5. doi:10.1038/nm0398-341
60. Biberfeld P, Chayt KJ, Marselle LM, Biberfeld G, Gallo RC, Harper ME. HTLV-III expression in infected lymph nodes and relevance to pathogenesis of lymphadenopathy. Am J Pathol (1986) 125:436–42.
61. Cheynier R, Henrichwark S, Hadida F, Pelletier E, Oksenhendler E, Autran B, et al. HIV and T cell expansion in splenic white pulps is accompanied by infiltration of HIV-specific cytotoxic T lymphocytes. Cell (1994) 78:373–87. doi:10.1016/0092-8674(94)90417-0
62. Moir S, Malaspina A, Li Y, Chun TW, Lowe T, Adelsberger J, et al. B cells of HIV-1-infected patients bind virions through CD21-complement interactions and transmit infectious virus to activated T cells. J Exp Med (2000) 192:637–46. doi:10.1084/jem.192.5.637
63. de Bree GJ, Lynch RM. B cells in HIV pathogenesis. Curr Opin Infect Dis (2016) 29:23–30. doi:10.1097/QCO.0000000000000225
64. Fukazawa Y, Lum R, Okoye AA, Park H, Matsuda K, Bae JY, et al. B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers. Nat Med (2015) 21:132–9. doi:10.1038/nm.3781
65. Yamamoto T, Lynch RM, Gautam R, Matus-nicodemos R, Schmidt SD, Boswell KL, et al. Quality and quantity of T FH cells are critical for broad antibody development in SHIV AD8 infection. Sci Transl Med (2015) 7:1–12. doi:10.1126/scitranslmed.aab3964
66. Cubas R, van Grevenynghe J, Wills S, Kardava L, Santich BH, Buckner CM, et al. Reversible reprogramming of circulating memory T follicular helper cell function during chronic HIV infection. J Immunol (2015) 195(12):5625–36. doi:10.4049/jimmunol.1501524
67. Casamayor-Palleja M, Feuillard J, Ball J, Drew M, MacLennan IC. Centrocytes rapidly adopt a memory B cell phenotype on co-culture with autologous germinal centre T cell-enriched preparations. Int Immunol (1996) 8:737–44. doi:10.1093/intimm/8.5.737
68. Allen RC, Armitage RJ, Conley ME, Rosenblatt H, Jenkins NA, Copeland NG, et al. CD40 ligand gene defects responsible for X-linked hyper-IgM syndrome. Science (1993) 259:990–3. doi:10.1126/science.7679801
69. Vanham G, Penne L, Devalck J, Kestens L, Colebunders R, Bosmans E, et al. Decreased CD40 ligand induction in CD4 T cells and dysregulated IL-12 production during HIV infection. Clin Exp Immunol (1999) 117:335–42. doi:10.1046/j.1365-2249.1999.00987.x
70. Stüber E, Strober W. The T cell-B cell interaction via OX40-OX40L is necessary for the T cell-dependent humoral immune response. J Exp Med (1996) 183:979–89. doi:10.1084/jem.183.3.979
71. Byun M, Ma CS, Akçay A, Pedergnana V, Palendira U, Myoung J, et al. Inherited human OX40 deficiency underlying classic Kaposi sarcoma of childhood. J Exp Med (2013) 210:1743–59. doi:10.1084/jem.20130592
72. Liu D, Xu H, Shih C, Wan Z, Ma X, Ma W, et al. T-B-cell entanglement and ICOSL-driven feed-forward regulation of germinal centre reaction. Nature (2015) 517:214–8. doi:10.1038/nature13803
73. Linterman MA, Beaton L, Yu D, Ramiscal RR, Srivastava M, Hogan JJ, et al. IL-21 acts directly on B cells to regulate Bcl-6 expression and germinal center responses. J Exp Med (2010) 207:353–63. doi:10.1084/jem.20091738
74. Zotos D, Coquet JM, Zhang Y, Light A, D’Costa K, Kallies A, et al. IL-21 regulates germinal center B cell differentiation and proliferation through a B cell-intrinsic mechanism. J Exp Med (2010) 207:365–78. doi:10.1084/jem.20091777
75. Wurster AL, Rodgers VL, White MF, Rothstein TL, Grusby MJ. Interleukin-4-mediated protection of primary B cells from apoptosis through Stat6-dependent up-regulation of Bcl-xL. J Biol Chem (2002) 277:27169–75. doi:10.1074/jbc.M201207200
76. Cunningham AF, Serre K, Toellner KM, Khan M, Alexander J, Brombacher F, et al. Pinpointing IL-4-independent acquisition and IL-4-influenced maintenance of Th2 activity of CD4 T cells. Eur J Immunol (2004) 34:686–94. doi:10.1002/eji.200324510
77. Arpin BC, Banchereau J, Liu Y. Memory B cells are biased towards terminal differentiation: a strategy that may prevent repertoire freezing. J Exp Med (1997) 186:931–40. doi:10.1084/jem.186.6.931
78. Yusuf I, Kageyama R, Monticelli L, Johnston RJ, Hansen K, Barnett B, et al. Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150). J Immunol (2010) 185:190–202. doi:10.4049/jimmunol.0903505.Germinal
79. Crotty S, Kersh EN, Cannons J, Schwartzberg PL, Ahmed R. SAP is required for generating long-term humoral immunity. Nature (2003) 421:2–7. doi:10.1038/nature01338.Published
80. Pallikkuth S, Parmigiani A, Silva SY, George VK, Fischl M, Pahwa R, et al. Impaired peripheral blood T-follicular helper cell function in HIV-infected nonresponders to the 2009 H1N1/09 vaccine. Blood (2012) 120:985–93. doi:10.1182/blood-2011-12-396648
81. Muir R, Metcalf T, Tardif V, Takata H, Phanuphak N, Kroon E, et al. Altered memory circulating T follicular helper-B cell interaction in early acute HIV infection. PLoS Pathog (2016) 12:e1005777. doi:10.1371/journal.ppat.1005777
82. Hocqueloux L, Avettand-Fènoël V, Jacquot S, Prazuck T, Legac E, Mélard A, et al. Long-term antiretroviral therapy initiated during primary HIV-1 infection is key to achieving both low HIV reservoirs and normal T cell counts. J Antimicrob Chemother (2013) 68:1169–78. doi:10.1093/jac/dks533
83. Laanani M, Ghosn J, Essat A, Melard A, Seng R, Gousset M, et al. Impact of the timing of initiation of antiretroviral therapy during primary HIV-1 infection on the decay of cell-associated HIV-DNA. Clin Infect Dis (2015) 60:1715–21. doi:10.1093/cid/civ171
84. Fletcher CV, Staskus K, Wietgrefe SW, Rothenberger M, Reilly C, Chipman JG, et al. Persistent HIV-1 replication is associated with lower antiretroviral drug concentrations in lymphatic tissues. Proc Natl Acad Sci U S A (2014) 111:2307–12. doi:10.1073/pnas.1318249111
85. Margolis DA, Brinson CC, Smith GHR, de Vente J, Hagins DP, Eron JJ, et al. Cabotegravir plus rilpivirine, once a day, after induction with cabotegravir plus nucleoside reverse transcriptase inhibitors in antiretroviral-naive adults with HIV-1 infection (LATTE): a randomised, phase 2b, dose-ranging trial. Lancet Infect Dis (2015) 15:1145–55. doi:10.1016/S1473-3099(15)00152-8
Keywords: HIV, SIV, Tfh cell differentiation, Tfh cell dynamics, germinal center reaction
Citation: Graff-Dubois S, Rouers A and Moris A (2016) Impact of Chronic HIV/SIV Infection on T Follicular Helper Cell Subsets and Germinal Center Homeostasis. Front. Immunol. 7:501. doi: 10.3389/fimmu.2016.00501
Received: 31 August 2016; Accepted: 26 October 2016;
Published: 11 November 2016
Edited by:
Smita S. Iyer, Emory University, USAReviewed by:
Christel Vérollet, Institute of Pharmacology and Structural Biology (CNRS), FranceCopyright: © 2016 Graff-Dubois, Rouers and Moris. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphanie Graff-Dubois, c3RlcGhhbmllLmdyYWZmLWR1Ym9pc0B1cG1jLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.