 Nancy H. Ruddle
Nancy H. Ruddle
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 09 November 2016
Sec. Inflammation
Volume 7 - 2016 | https://doi.org/10.3389/fimmu.2016.00491
This article is part of the Research Topic Tertiary Lymphoid Organs (TLOs): powerhouses of disease immunity View all 22 articles
High endothelial venules (HEVs) and lymphatic vessels (LVs) are essential for the function of the immune system, by providing communication between the body and lymph nodes (LNs), specialized sites of antigen presentation and recognition. HEVs bring in naïve and central memory cells and LVs transport antigen, antigen-presenting cells, and lymphocytes in and out of LNs. Tertiary lymphoid organs (TLOs) are accumulations of lymphoid and stromal cells that arise and organize at ectopic sites in response to chronic inflammation in autoimmunity, microbial infection, graft rejection, and cancer. TLOs are distinguished from primary lymphoid organs – the thymus and bone marrow, and secondary lymphoid organs (SLOs) – the LNs, spleen, and Peyer’s patches, in that they arise in response to inflammatory signals, rather than in ontogeny. TLOs usually do not have a capsule but are rather contained within the confines of another organ. Their structure, cellular composition, chemokine expression, and vascular and stromal support resemble SLOs and are the defining aspects of TLOs. T and B cells, antigen-presenting cells, fibroblast reticular cells, and other stromal cells and vascular elements including HEVs and LVs are all typical components of TLOs. A key question is whether the HEVs and LVs play comparable roles and are regulated similarly to those in LNs. Data are presented that support this concept, especially with regard to TLO HEVs. Emerging data suggest that the functions and regulation of TLO LVs are also similar to those in LNs. These observations support the concept that TLOs are not merely cellular accumulations but are functional entities that provide sites to generate effector cells, and that their HEVs and LVs are crucial elements in those activities.
Lymphoid and stromal cells accumulate and organize into tertiary lymphoid organs (TLOs) at ectopic sites in response to chronic inflammation in autoimmunity, microbial infection, graft rejection, and cancer where they assume structural and cellular characteristics of lymph nodes (LNs). High endothelial venules (HEVs) and lymphatic vessels (LVs) play key roles in LNs in transporting cells and antigens from and to the body. The questions to be addressed here are whether the HEVs and LVs in TLOs function and are regulated in a manner similar to those in LNs.
My research group became intrigued by the concept of TLOs in the course of two apparently unrelated series of investigations. The first was the characterization of mice that were transgenic for a construct of the rat insulin promoter driving expression of lymphotoxin alpha (LTα) (1) (in those days known as TNFβ, despite having been described as LT previous to the discovery of TNF). We made the rat insulin promoter lymphotoxin (RIPLT) mouse in order to develop a model of type 1 diabetes, since we knew that LT could induce inflammation. The transgene was not only expressed in the β cells in the islets of Langerhans in the pancreas as expected but also in the kidney and skin, most likely because the entire promoter with its negative regulatory elements was not included in the construct. At all sites of transgene expression, lymphoid cells accumulated, which were organized into distinct T and B cell areas (“compartmentalization”). Despite several attempts to drive the animals to β cell destruction and diabetes, the mice were healthy (2) unless a costimulator molecule such as B7-1 was also expressed in the β cells. Thus, the model resembled the early peri insulitis and non-destructive insulitis of diabetes. At the same time, we were collaborating with David Chaplin on the LTα knock out mouse that has no LNs (3). We realized that the consequence of ectopic expression of LT in the RIPLT mouse was the production of organized infiltrates that resembled LNs. We called them TLOs (4), a term that had been previously used to designate any lymphoid infiltrate (5). The process by which TLOs arise and organize was designated as lymphoid neogenesis (4).
In later years, I became especially interested in the vasculature of TLOs as I realized that understanding how cells enter into TLOs would provide insight into this accumulation and would indicate whether or not the apparent organization reflected function. That is, the presence of HEVs might indicate that naïve cells could enter the TLO, and the presence of LVs could indicate a method of entrance of antigen-presenting cells, thus providing in a single location, the elements to generate an immune response. This manuscript addresses these questions.
Tertiary lymphoid organs, which have been described in almost every organ of the body, are also known as tertiary lymphoid structures, ectopic lymphoid tissues, or tertiary lymphoid tissues. They are distinguished from primary lymphoid organs – the thymus and bone marrow, and secondary lymphoid organs (SLOs) – the LNs, spleen, and Peyer’s patches, in that they arise in response to inflammation or inflammatory cues, rather than in ontogeny and are ectopic to canonical lymphoid organs. They usually do not have a capsule but are rather contained within the confines of another organ.
Tertiary lymphoid organs are similar to LNs (6) with regard to their cellular content, stromal components, lymphoid chemokines (7), vasculature, and organization. Cells include compartmentalized T and B cells and antigen-presenting cells, including follicular dendritic cells and dendritic cells. CD8 and CD4 subsets include naïve, Treg, and T follicular helper cells (8, 9). B cells may be organized into germinal centers with plasma cells. HEVs (10), LVs (11, 12) (Figure 1), and conduits with fibroblastic reticular cells (13), all components of LNs, have also been described. In LNs, CCL19 and CCL21 direct T cells and DCs to the paracortical region, and CXCL13 directs B cells to the B cell follicles. These chemokines and cells that express their receptors are also expressed in TLOs (7). TLOs can be distinguished from acute inflammation; they generally include few granulocytes, and they are not necessarily destructive, although they may transform into tissue damaging entities. The plasticity of TLOs is seen in the case of the infiltrates in the pancreas in type 1 diabetes in the NOD mouse. Initially, the cellular infiltrates are disorganized and lack HEVs; then the infiltrates assume the characteristics of TLOs, with T and B compartmentalization and HEVs and LVs (14, 15); later, the lymphoid cells become activated, β cells are destroyed, and eventually the inflammation and thus, the TLO, is resolved as antigen is eliminated.
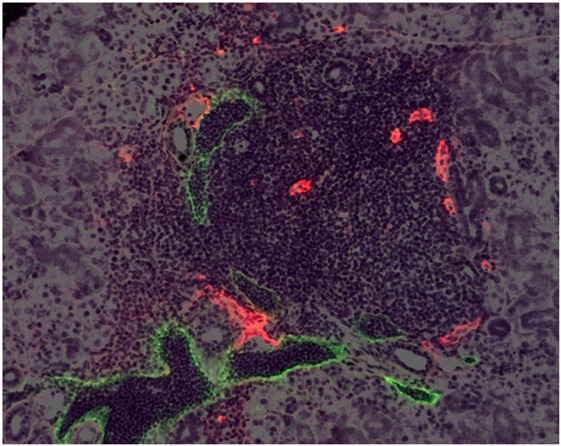
Figure 1. High endothelial venules and lymphatic vessels in a TLO. A mouse salivary gland TLO. HEVs are stained red with an antibody to MECA-79. LVs are stained green with an antibody to LYVE-1. From “Transgenic LacZ under control of Hec-6ST regulatory sequences recapitulates endogenous gene expression on high endothelial venules” by Liao et al. (11). Copyright (2007) National Academy of Sciences, USA.
Tertiary lymphoid organs differ from LNs in that they generally do not have a capsule, they are not confined to a fixed location in the body, they develop postnatally, and as noted above, they exhibit plasticity. This is not to say that LNs do not respond to their environment; they most certainly do with proliferation and changes in vasculature and cell and antigen accessibility in the course of inflammation [see, e.g., Ref. (16)].
Tertiary lymphoid organ functions vary depending on the location, stimulus and kinetics of inflammation, and cellular activation. The strongest evidence that TLOs are harmful in exacerbating autoimmune disease derives from studies in rheumatoid arthritis. In some patients, evidence that somatic mutation and affinity maturation occur in the locus of the TLO in the joint provides support for a harmful role leading to determinant spreading. On the other hand, the presence of Tregs in some TLOs (17) suggests that they can play a beneficial role by limiting inflammation. Additional evidence for a beneficial role is provided from several clinical studies of cancer, which indicate that the presence of TLOs in tumors in breast, colon, or lung predicts a favorable outcome, suggesting that the TLO site provides a locus for antigen activation and destruction of tumor, reducing dissemination of the malignant cells through the body (18). Nevertheless, Tregs in tumor TLOs can act as brakes on their defensive role (19, 20).
The presence of HEVs could be considered an essential trait distinguishing TLOs from acute inflammation. The endothelial cells in postcapillary venules in TLOs, as in LNs, tonsils, and Peyer’s patches, exhibit a typical cuboidal appearance. LN HEVs express a particular set of genes that facilitate their interactions with blood stream naïve and central memory cells that result in rolling, firm adhesion, and transmigration from the vessel into the parenchyma. HEVs in TLOs express the same molecules: CCL21 (7), ICAM-1 (4), and peripheral and/or mucosal addressins, PNAd (10) and MAdCAM-1 (4). Expression of these proteins allows the egress from the blood stream into the parenchyma of LNs of cells of the naïve and central memory phenotype that express CCR7, LFA-1, L-selectin (CD62L), and α4β7.
The evidence is quite strong that HEVs in TLOs function similarly to those in LNs, allowing naïve and central memory cells to leave the blood stream and enter into the parenchyma of the tissue where they can interact with their cognate antigen. First, as noted above, they express the molecules that allow naïve and central memory cells to interact. Second, cells expressing CCR7, LFA-1, L-selectin (CD62L), and α4β7, the ligands for the receptors on HEVs, are found in TLOs. Third, several instances of T cell activation and memory generation occurring directly in the TLO have been described. These include generation of memory cells for graft rejection in skin TLOs (21) and presentation and activation of Teffector or Treg cells (19, 22). In vivo imaging of the transit of naïve cells into TLOs and their interaction with antigen-presenting cells will solidify the conclusion that HEVs function similarly in LNs and TLOs, and that HEVs in TLOs are the sites of entrance of naïve cells to undergo activation and differentiation and generation of memory cells.
High endothelial venules are regulated similarly in TLOs and SLOs. LTα alone induces MAdCAM-1 in endothelial cells in vitro (23, 24), in vivo in mesenteric LN HEVs (16), and in HEVs in TLOs (23) through TNFR1 (25). Abluminal PNAd in LN HEVs is generated through modification of a variety of glycoproteins. These modifications include sulfation, which is essential for PNAd (also called L-selectin ligand) interaction with its receptor, L-selectin (CD62L) that is expressed on the surface of naïve and central memory lymphocytes. Sulfation is induced in peripheral LN HEVs by sulfotransferases (26, 27). LTαβ regulates the HEV sulfotransferase in both LNs (16, 28) and TLOs (10) through the alternative NFκB pathway (29).
Lymphatic vessels play key roles in the body in fluid and lipid balance. They are crucial in the immune system in providing communication of the lymphoid organs with the rest of the body. Lymphatic capillaries are thin-walled, blind-ended vessels that express CCL21, LYVE-1, PROX-1, podoplanin, VEGFR-2, and VEGFR-3 and are the initial entry point into LNs from the tissues for antigen and antigen-presenting cells. The endothelial cells on the tips of lymphatic capillaries are most frequently in a zipper-like arrangement (30). They connect to collecting vessels whose cells exhibit a button-like arrangement that are usually low or negative for LYVE-1, but do express PROX-1. The latter is especially highly expressed in valves that are characteristic of collecting vessels. A layer of smooth muscle cells surrounding collecting vessels contributes to their pumping action. Afferent collecting vessels carry substances to LNs, whereas efferent vessels allow egress of activated cells from the LN into the next LN in the chain and eventually into the blood stream via the right or left subclavian veins. In addition to serving as routes of fluid, lipid, cell, and cytokine transport, recent publications attest to the ability of LN LVs to present self or foreign antigens, either directly or by transfer to antigen-presenting cells (31–34).
Thin-walled vessels that are positive for lymphatic markers, including LYVE-1, PROX-1, podoplanin in mouse and human or D2-40 in human have been noted in many TLOs [summarized in Ref. (12)]. These include chronic kidney rejection (35, 36), cardiac allografts (37), transgenic mouse models (38), age-related Sjögren’s-like disease in the mouse (11), and a transgenic model of primary Sjögren’s in the mouse (Truman et al., in preparation). Confusingly, a reduced number of LVs in kidneys of mouse strains with a higher preponderance of spontaneous kidney TLOs have been noted (39). However, the latter report did not indicate the actual location of the LVs (i.e., in the vicinity or not of the TLO). CCL21-expressing TLO-associated vessels have been described in rheumatoid arthritis, Crohn’s disease, Sjögren’s syndrome, chronic allograft rejection (40), and pancreatic infiltrates in NOD (15) and RIPLTα mice (7). Nevertheless, much still needs to be learned. Collecting vessels with valves and smooth muscle cells neither have been specifically identified entering or leaving TLOs nor have the vessel walls been characterized with regard to their zipper or button-like morphology.
Do the LVs in TLOs carry out the same functions as those in LNs? It is likely that they contribute to fluid drainage, although this has not been carefully analyzed. Do LVs carry antigen and cells to TLOs and cells away from TLOs, as do afferent and efferent vessels in LNs? TLO LVs frequently contain cells (11, 39), supporting the concept that they act as transporters as does their expression of CCL21 indicating they interact with CCR7-expressing cells. However, the fact that LVs in some TLOs appear to be packed with cells suggests that there could be a defect in cellular drainage and that their efferent function is compromised. Sphingosine-1 phosphate (S1P) is expressed in lymph and downregulates its receptor (S1P1) on lymphocytes. Lymphocytes in LNs reexpress the receptor and migrate toward the S1P in lymph and egress from the LN. FTY720 (fingolimod) is an agent that is used in transplantation and multiple sclerosis treatment that acts as an agonist for the receptor, causing its internalization resulting in lymphocyte accumulation in LNs (41), thus acting as an immunosuppressant. When NOD mice with pancreatic TLOs are treated with this agent, they are protected from islet destruction and diabetes, consistent with the concept that their LVs carry out an efferent function (42). In our hands, this treatment inhibits disease only at the time that the mice exhibit TLOs (15), although others have determined that FTY720 treatment is partially effective even after the development of elevated blood sugar (43). The pancreatic TLOs exhibit an increased insulitis score after FTY720 treatment, indicating that cells are trapped in these structures. Within days of cessation of drug treatment, islet destruction and diabetes occurs (15, 42). These data are consistent with the concept that the S1P gradient affects lymphocyte trafficking in TLO LVs. Further supporting the concept that the FTY720 effects are at least partially due to an effect on the TLOs is the observation that FTY720 treatment inhibits cellular migration from inflamed tissues into afferent LVs (44, 45). It must be noted that FTY720 treatment is also most likely affecting trafficking from LNs in this context, complicating interpretation of the data. This needs to be evaluated in situations where the events in TLOs can be isolated from LNs, as was done in a previous transplantation model (21). A straightforward test of these conclusions would be to determine if LVs in TLOs produce S1P as they do in LNs (46). If so, systemic inhibitors of lymphocyte trafficking may function directly at the TLO site by preventing traffic to the LNs from the TLO, a potential site of self antigen presentation.
Lymphatic vessels transport soluble or cell-associated antigens into LNs. Recently, it has become apparent that plasmalemma vesicle-associated protein (PLVAP), visualized by reactivity with the MECA-32 antibody, heretofore considered limited to blood vessels, is also expressed on the lymphatic endothelial cells in the lymphatic sinus in the LN. PLVAP positive lymphatic endothelial cells contribute to sieving of lymphocytes and high molecular weight antigens entering the LN via the conduits (47). Since TLOs include conduits (13), it seems reasonable to ask whether LVs in TLOs perform antigen and cell transport and sieving functions similar to those in LNs. Antigen transport may be less important than in SLOs because the antigen is an actual component of the TLO. As long as antigen-presenting cells are in the TLO (as they usually are), the issue is moot. Proteins such as insulin in the pancreatic islet are in immediate proximity or, as constituents of β cells, even contribute to the structure of the TLO in type 1 diabetes. With regard to the sieving function, an analysis of expression of PLVAP in TLOs by co-staining with MECA-32 and LYVE-1 or PROX-1 should be fairly straightforward. Functional analysis by crossing PLVAP-deficient mice to mice with TLOs or MECA-32 inhibition of migration of cells or labeled antigen to TLOs could address the function of LVs in TLOs. As noted above, LVs in LNs present self antigens (31–34), either directly through their expression of MHC molecules or by passing antigen on to “classical” antigen-presenting cells. Such presentation of self antigen by LVs (31) could be a way to induce either tolerance or T cell activation in LNs or in TLOs. The ability of TLO LVs to present antigen to induce either of these outcomes has not been investigated. Tregs are found in tumor TLOs and can inhibit cytotoxic T cells from attacking the tumor (19), indicating that understanding the mechanisms of self and tumor presentation to both potential effector T cells and Tregs is crucial to our ability to harness TLOs for both prophylaxis and therapy of cancer and autoimmune diseases.
The most commonly accepted scenario for the development of LVs in ontogeny is that they sprout from veins (48) under the influence of SOX18, PROX-1, growth factors and their receptors (VEGF-C and VEGF-D and VEGFR-2 and VEGFR-3), and platelets (49) [reviewed in Ref. (50)]. Although the evidence is quite strong for this mechanism in the case of the LVs sprouting from the cardinal vein, it has become apparent that the situation is somewhat more complex. The first indication that additional mechanisms of lymphangiogenesis existed was the discovery of lymphangioblasts that could be distinguished from blood endothelial cells, in developing animals as distinct as tadpoles (51), chickens (52), and mice (53–55). Several recent studies have revealed that the origin of LVs is quite heterogeneous. Mahadevan et al. reported that LVs in the intestine are derived from arteries, rather than veins (56); Stanczuk et al. described hemangiogenic precursors that contribute to mesenteric LVs (57); Martinez-Corral et al. described the non-venous origin of dermal LVs in a process these authors termed lymphvasculogenesis (58); Klotz et al. also described a non-venous origin of cardiac LVs (59); and Nicenboim et al. reported that LVs derive from angioblasts in zebra fish (60).
Given the rapidly emerging data regarding the heterogeneity and the likelihood of organ-specific regulation of lymphangiogenesis in ontogeny (61), it becomes more important, and perhaps even more daunting, to understand the regulation of lymphangiogenesis in inflammation, particularly in chronic inflammation in TLOs. Do LVs in TLOs arise from veins? The presence of angiogenesis and platelets in inflammation supports such a scenario, as does the existence of vessels that express both HEV and LV markers in the inflamed LN (16). On the other hand, host-derived bone marrow precursors have been noted in association with LVs in the TLOs of chronically rejecting kidneys (36) suggesting a non-venous origin. Lymphangiogenesis in inflammation could occur by sprouting from existing LVs. But what cells orchestrate these events? DCs, macrophages, T and B cells have been implicated in the regulation of LVs in acute inflammation (16, 62–64), but different cells may be important at different times in different tissues. For example, B cells appear to be important in stimulating lymphangiogenesis that occurs in LNs during inflammation, but only at the early stages after immunization (16, 62) suggesting that they may be of lesser importance in chronic inflammation in TLOs. The participation of macrophages in lymphangiogenesis in acute inflammation has been documented, although the precise nature of their role is controversial. Various possibilities include integration into LVs, trans differentiation into lymphatic endothelial cells (65), and provision of growth factors [summarized in Ref. (66)] and cytokines. The expression of LYVE-1 by macrophages is supportive evidence for the former possibility; on the other hand, the expression of this marker on both macrophages and LECs may be serendipitous.
Several studies have evaluated the negative and positive roles of cytokines in lymphangiogenesis, although the bulk of these studies have evaluated acute inflammation rather than TLOs. There have been reports of negative regulation of lymphangiogenesis by IFNγ (67) and TH2 cytokines IL-4 and IL-13 (68) and positive regulation by IL-17 (69), LTα, and TNF (38, 70). LT is crucial for both lymphoid organ development and TLOs, and LTα3 contributes to lymphangiogenesis in development (38). LVs are apparent in RIPLT TLOs even in the absence of LTβ and before extensive cellular infiltration, suggesting a direct activity of the cytokine (38). On the other hand, LVs are inhibited by treatment with a LTβR–Ig in a CXCL13-induced model of a thyroid TLO (71). Further analysis of lymphangiogenesis in spontaneous TLOs, such as Sjögren’s syndrome, rheumatoid arthritis, and type 1 diabetes, may reveal which cytokines regulate this process. Additional studies in vivo and in vitro should reveal the mechanism of cytokines’ regulation as direct effects on lymphatic endothelial cells and/or as indirect effects through the facilitation of lymphatic growth factor producing cells.
Recent research reveals LV plasticity in gene function and regulation. It is obvious that their different environments (mesentery, skin, etc.) influence their gene expression. Inflammation in these diverse locales also contributes to changes in cytokine and growth factor expression. TNF and oxazolone treatment induce higher levels of CCL21 on dermal LVs, and presumably enhance cellular migration (72). Immunofluorescence and microarray studies revealed an increase in several additional inflammatory genes (73), although some genes, including VEGFR-3 and PROX-1, are downregulated. As yet, no such comparisons have included TLO LECs, which would be of particular interest because of the chronic nature of stimulation. Recently described methods to isolate LVs by virtue of their transgenically induced expression of a tomato red fluorescent protein should allow direct comparison of gene expression and function of LVs from different sites and acute and chronic inflammation (74, 75) and provide precise characterization of TLO LVs.
In this communication, I have provided background from a personal perspective of the development of the TLO field, and more particularly the role of the vasculature present and employed in TLOs. Although questions remain concerning the precise functions of HEVs and LVs in TLOs, the evidence is quite strong that they do behave as they do in LNs. The appropriate experimental tools (in vivo imaging, mice with fluorescent HEVs and LVs) are available to address these issues. The answers to these questions will provide insight, not only into TLOs but also into processes of antigen presentation in LNs and tissue destruction in acute inflammation. This is turn will provide understanding and methods to induce or inhibit TLOs in autoimmunity, microbial infection, organ rejection, and cancer.
The author confirms being the sole contributor of this work and approved it for publication.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author gratefully acknowledges the contributions of her many colleagues, collaborators, and trainees who have provided data and insight that gave rise to the concept of TLOs.
This work was supported by 13-SCD-YALE-27, a grant from the Connecticut Regenerative Medicine Research Fund of Connecticut Innovations.
HEV, high endothelial venule; LEC, lymphatic endothelial cell; LN, lymph node; LT, lymphotoxin; LV, lymphatic vessel; SLO, secondary lymphoid organ; TLO, tertiary lymphoid organ.
1. Picarella DE, Kratz A, Li CB, Ruddle NH, Flavell RA. Insulitis in transgenic mice expressing tumor necrosis factor beta (lymphotoxin) in the pancreas. Proc Natl Acad Sci U S A (1992) 89(21):10036–40. doi:10.1073/pnas.89.21.10036
2. Picarella DE, Kratz A, Li CB, Ruddle NH, Flavell RA. Transgenic tumor necrosis factor (TNF)-alpha production in pancreatic islets leads to insulitis, not diabetes. Distinct patterns of inflammation in TNF-alpha and TNF-beta transgenic mice. J Immunol (1993) 150(9):4136–50.
3. De Togni P, Goellner J, Ruddle NH, Streeter PR, Andrea F, Mariathasan S, et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science. 1994. 264: 703-707. J Immunol (2014) 192(5):2010–4. doi:10.1126/science.8171322
4. Kratz A, Campos-Neto A, Hanson MS, Ruddle NH. Chronic inflammation caused by lymphotoxin is lymphoid neogenesis. J Exp Med (1996) 183(4):1461–72. doi:10.1084/jem.183.4.1461
5. Picker LJ, Butcher EC. Physiological and molecular mechanisms of lymphocyte homing. Annu Rev Immunol (1992) 10:561–91. doi:10.1146/annurev.iy.10.040192.003021
6. Drayton DL, Liao S, Mounzer RH, Ruddle NH. Lymphoid organ development: from ontogeny to neogenesis. Nat Immunol (2006) 7(4):344–53. doi:10.1038/ni1330
7. Hjelmstrom P, Fjell J, Nakagawa T, Sacca R, Cuff CA, Ruddle NH. Lymphoid tissue homing chemokines are expressed in chronic inflammation. Am J Pathol (2000) 156(4):1133–8. doi:10.1016/S0002-9440(10)64981-4
8. Kielczewski JL, Horai R, Jittayasothorn Y, Chan CC, Caspi RR. Tertiary lymphoid tissue forms in retinas of mice with spontaneous autoimmune uveitis and has consequences on visual function. J Immunol (2016) 196(3):1013–25. doi:10.4049/jimmunol.1501570
9. Gu-Trantien C, Willard-Gallo K. Tumor-infiltrating follicular helper T cells: the new kids on the block. Oncoimmunology (2013) 2(10):e26066. doi:10.4161/onci.26066
10. Drayton DL, Ying X, Lee J, Lesslauer W, Ruddle NH. Ectopic LT alpha beta directs lymphoid organ neogenesis with concomitant expression of peripheral node addressin and a HEV-restricted sulfotransferase. J Exp Med (2003) 197(9):1153–63. doi:10.1084/jem.20021761
11. Liao S, Bentley K, Lebrun M, Lesslauer W, Ruddle FH, Ruddle NH. Transgenic LacZ under control of Hec-6st regulatory sequences recapitulates endogenous gene expression on high endothelial venules. Proc Natl Acad Sci U S A (2007) 104(11):4577–82. doi:10.1073/pnas.0700334104
12. Ruddle NH. Lymphatic vessels and tertiary lymphoid organs. J Clin Invest (2014) 124(3):953–9. doi:10.1172/JCI71611
13. Stranford S, Ruddle NH. Follicular dendritic cells, conduits, lymphatic vessels, and high endothelial venules in tertiary lymphoid organs: parallels with lymph node stroma. Front Immunol (2012) 3:350. doi:10.3389/fimmu.2012.00350
14. Astorri E, Bombardieri M, Gabba S, Peakman M, Pozzilli P, Pitzalis C. Evolution of ectopic lymphoid neogenesis and in situ autoantibody production in autoimmune nonobese diabetic mice: cellular and molecular characterization of tertiary lymphoid structures in pancreatic islets. J Immunol (2010) 185(6):3359–68. doi:10.4049/jimmunol.1001836
15. Penaranda C, Tang Q, Ruddle NH, Bluestone JA. Prevention of diabetes by FTY720-mediated stabilization of peri-islet tertiary lymphoid organs. Diabetes (2010) 59(6):1461–8. doi:10.2337/db09-1129
16. Liao S, Ruddle NH. Synchrony of high endothelial venules and lymphatic vessels revealed by immunization. J Immunol (2006) 177(5):3369–79. doi:10.4049/jimmunol.177.5.3369
17. Grabner R, Lotzer K, Dopping S, Hildner M, Radke D, Beer M, et al. Lymphotoxin beta receptor signaling promotes tertiary lymphoid organogenesis in the aorta adventitia of aged ApoE-/- mice. J Exp Med (2009) 206(1):233–48. doi:10.1084/jem.20080752
18. Martinet L, Garrido I, Filleron T, Le Guellec S, Bellard E, Fournie JJ, et al. Human solid tumors contain high endothelial venules: association with T- and B-lymphocyte infiltration and favorable prognosis in breast cancer. Cancer Res (2011) 71(17):5678–87. doi:10.1158/0008-5472.CAN-11-0431
19. Joshi NS, Akama-Garren EH, Lu Y, Lee DY, Chang GP, Li A, et al. Regulatory T cells in tumor-associated tertiary lymphoid structures suppress anti-tumor T cell responses. Immunity (2015) 43(3):579–90. doi:10.1016/j.immuni.2015.08.006
20. Martinet L, Filleron T, Le Guellec S, Rochaix P, Garrido I, Girard JP. High endothelial venule blood vessels for tumor-infiltrating lymphocytes are associated with lymphotoxin beta-producing dendritic cells in human breast cancer. J Immunol (2013) 191(4):2001–8. doi:10.4049/jimmunol.1300872
21. Nasr IW, Reel M, Oberbarnscheidt MH, Mounzer RH, Baddoura FK, Ruddle NH, et al. Tertiary lymphoid tissues generate effector and memory T cells that lead to allograft rejection. Am J Transplant (2007) 7(5):1071–9. doi:10.1111/j.1600-6143.2007.01756.x
22. Hu D, Mohanta SK, Yin C, Peng L, Ma Z, Srikakulapu P, et al. Artery tertiary lymphoid organs control aorta immunity and protect against atherosclerosis via vascular smooth muscle cell lymphotoxin beta receptors. Immunity (2015) 42(6):1100–15. doi:10.1016/j.immuni.2015.05.015
23. Cuff CA, Sacca R, Ruddle NH. Differential induction of adhesion molecule and chemokine expression by LTalpha3 and LTalphabeta in inflammation elucidates potential mechanisms of mesenteric and peripheral lymph node development. J Immunol (1999) 162(10):5965–72.
24. Cuff CA, Schwartz J, Bergman CM, Russell KS, Bender JR, Ruddle NH. Lymphotoxin alpha3 induces chemokines and adhesion molecules: insight into the role of LT alpha in inflammation and lymphoid organ development. J Immunol (1998) 161(12):6853–60.
25. Sacca R, Cuff CA, Lesslauer W, Ruddle NH. Differential activities of secreted lymphotoxin-alpha3 and membrane lymphotoxin-alpha1beta2 in lymphotoxin-induced inflammation: critical role of TNF receptor 1 signaling. J Immunol (1998) 160(1):485–91.
26. Bistrup A, Bhakta S, Lee JK, Belov YY, Gunn MD, Zuo FR, et al. Sulfotransferases of two specificities function in the reconstitution of high endothelial cell ligands for L-selectin. J Cell Biol (1999) 145(4):899–910. doi:10.1083/jcb.145.4.899
27. Hemmerich S, Bistrup A, Singer MS, van Zante A, Lee JK, Tsay D, et al. Sulfation of L-selectin ligands by an HEV-restricted sulfotransferase regulates lymphocyte homing to lymph nodes. Immunity (2001) 15(2):237–47. doi:10.1016/S1074-7613(01)00188-1
28. Browning JL, Allaire N, Ngam-Ek A, Notidis E, Hunt J, Perrin S, et al. Lymphotoxin-beta receptor signaling is required for the homeostatic control of HEV differentiation and function. Immunity (2005) 23(5):539–50. doi:10.1016/j.immuni.2005.10.002
29. Drayton DL, Bonizzi G, Ying X, Liao S, Karin M, Ruddle NH. Ikappa B kinase complex alpha kinase activity controls chemokine and high endothelial venule gene expression in lymph nodes and nasal-associated lymphoid tissue. J Immunol (2004) 173(10):6161–8. doi:10.4049/jimmunol.173.10.6161
30. Baluk P, Fuxe J, Hashizume H, Romano T, Lashnits E, Butz S, et al. Functionally specialized junctions between endothelial cells of lymphatic vessels. J Exp Med (2007) 204(10):2349–62. doi:10.1084/jem.20062596
31. Cohen JN, Guidi CJ, Tewalt EF, Qiao H, Rouhani SJ, Ruddell A, et al. Lymph node-resident lymphatic endothelial cells mediate peripheral tolerance via Aire-independent direct antigen presentation. J Exp Med (2010) 207(4):681–8. doi:10.1084/jem.20092465
32. Rouhani SJ, Eccles JD, Riccardi P, Peske JD, Tewalt EF, Cohen JN, et al. Roles of lymphatic endothelial cells expressing peripheral tissue antigens in CD4 T-cell tolerance induction. Nat Commun (2015) 6:6771. doi:10.1038/ncomms7771
33. Tewalt EF, Cohen JN, Rouhani SJ, Guidi CJ, Qiao H, Fahl SP, et al. Lymphatic endothelial cells induce tolerance via PD-L1 and lack of costimulation leading to high-level PD-1 expression on CD8 T cells. Blood (2012) 120(24):4772–82. doi:10.1182/blood-2012-04-427013
34. Tamburini BA, Burchill MA, Kedl RM. Antigen capture and archiving by lymphatic endothelial cells following vaccination or viral infection. Nat Commun (2014) 5:3989. doi:10.1038/ncomms4989
35. Kerjaschki D, Huttary N, Raab I, Regele H, Bojarski-Nagy K, Bartel G, et al. Lymphatic endothelial progenitor cells contribute to de novo lymphangiogenesis in human renal transplants. Nat Med (2006) 12(2):230–4. doi:10.1038/nm1340
36. Kerjaschki D, Regele HM, Moosberger I, Nagy-Bojarski K, Watschinger B, Soleiman A, et al. Lymphatic neoangiogenesis in human kidney transplants is associated with immunologically active lymphocytic infiltrates. J Am Soc Nephrol (2004) 15(3):603–12. doi:10.1097/01.ASN.0000113316.52371.2E
37. Motallebzadeh R, Rehakova S, Conlon TM, Win TS, Callaghan CJ, Goddard M, et al. Blocking lymphotoxin signaling abrogates the development of ectopic lymphoid tissue within cardiac allografts and inhibits effector antibody responses. FASEB J (2012) 26(1):51–62. doi:10.1096/fj.11-186973
38. Mounzer RH, Svendsen OS, Baluk P, Bergman CM, Padera TP, Wiig H, et al. Lymphotoxin-alpha contributes to lymphangiogenesis. Blood (2010) 116(12):2173–82. doi:10.1182/blood-2009-12-256065
39. Huang Y, Caputo CR, Noordmans GA, Yazdani S, Monteiro LH, van den Born J, et al. Identification of novel genes associated with renal tertiary lymphoid organ formation in aging mice. PLoS One (2014) 9(3):e91850. doi:10.1371/journal.pone.0091850
40. Nykanen AI, Sandelin H, Krebs R, Keranen MA, Tuuminen R, Karpanen T, et al. Targeting lymphatic vessel activation and CCL21 production by vascular endothelial growth factor receptor-3 inhibition has novel immunomodulatory and antiarteriosclerotic effects in cardiac allografts. Circulation (2010) 121(12):1413–22. doi:10.1161/CIRCULATIONAHA.109.910703
41. Mandala S, Hajdu R, Bergstrom J, Quackenbush E, Xie J, Milligan J, et al. Alteration of lymphocyte trafficking by sphingosine-1-phosphate receptor agonists. Science (2002) 296(5566):346–9. doi:10.1126/science.1070238
42. Maki T, Gottschalk R, Ogawa N, Monaco AP. Prevention and cure of autoimmune diabetes in nonobese diabetic mice by continuous administration of FTY720. Transplantation (2005) 79(9):1051–5. doi:10.1097/01.TP.0000161220.87548.EE
43. Tsuji T, Inoue M, Yoshida Y, Fujita T, Kaino Y, Kohno T. Therapeutic approach for type 1 diabetes mellitus using the novel immunomodulator FTY720 (fingolimod) in combination with once-daily injection of insulin glargine in non-obese diabetic mice. J Diabetes Investig (2012) 3(2):132–7. doi:10.1111/j.2040-1124.2011.00160.x
44. Brown MN, Fintushel SR, Lee MH, Jennrich S, Geherin SA, Hay JB, et al. Chemoattractant receptors and lymphocyte egress from extralymphoid tissue: changing requirements during the course of inflammation. J Immunol (2010) 185(8):4873–82. doi:10.4049/jimmunol.1000676
45. Ledgerwood LG, Lal G, Zhang N, Garin A, Esses SJ, Ginhoux F, et al. The sphingosine 1-phosphate receptor 1 causes tissue retention by inhibiting the entry of peripheral tissue T lymphocytes into afferent lymphatics. Nat Immunol (2008) 9(1):42–53. doi:10.1038/ni1534
46. Pham TH, Baluk P, Xu Y, Grigorova I, Bankovich AJ, Pappu R, et al. Lymphatic endothelial cell sphingosine kinase activity is required for lymphocyte egress and lymphatic patterning. J Exp Med (2010) 207(1):17–27. doi:10.1084/jem.20091619
47. Rantakari P, Auvinen K, Jappinen N, Kapraali M, Valtonen J, Karikoski M, et al. The endothelial protein PLVAP in lymphatics controls the entry of lymphocytes and antigens into lymph nodes. Nat Immunol (2015) 16(4):386–96. doi:10.1038/ni.3101
48. Srinivasan RS, Dillard ME, Lagutin OV, Lin FJ, Tsai S, Tsai MJ, et al. Lineage tracing demonstrates the venous origin of the mammalian lymphatic vasculature. Genes Dev (2007) 21(19):2422–32. doi:10.1101/gad.1588407
49. Abtahian F, Guerriero A, Sebzda E, Lu MM, Zhou R, Mocsai A, et al. Regulation of blood and lymphatic vascular separation by signaling proteins SLP-76 and Syk. Science (2003) 299(5604):247–51. doi:10.1126/science.1079477
50. Tammela T, Alitalo K. Lymphangiogenesis: molecular mechanisms and future promise. Cell (2010) 140(4):460–76. doi:10.1016/j.cell.2010.01.045
51. Ny A, Koch M, Schneider M, Neven E, Tong RT, Maity S, et al. A genetic Xenopus laevis tadpole model to study lymphangiogenesis. Nat Med (2005) 11(9):998–1004. doi:10.1038/nm1285
52. Wilting J, Aref Y, Huang R, Tomarev SI, Schweigerer L, Christ B, et al. Dual origin of avian lymphatics. Dev Biol (2006) 292(1):165–73. doi:10.1016/j.ydbio.2005.12.043
53. Buttler K, Badar M, Seiffart V, Laggies S, Gross G, Wilting J, et al. De novo hem- and lymphangiogenesis by endothelial progenitor and mesenchymal stem cells in immunocompetent mice. Cell Mol Life Sci (2013) 71(8):1513–27. doi:10.1007/s00018-013-1460-8
54. Buttler K, Ezaki T, Wilting J. Proliferating mesodermal cells in murine embryos exhibiting macrophage and lymphendothelial characteristics. BMC Dev Biol (2008) 8:43. doi:10.1186/1471-213X-8-43
55. Buttler K, Kreysing A, von Kaisenberg CS, Schweigerer L, Gale N, Papoutsi M, et al. Mesenchymal cells with leukocyte and lymphendothelial characteristics in murine embryos. Dev Dyn (2006) 235(6):1554–62. doi:10.1002/dvdy.20737
56. Mahadevan A, Welsh IC, Sivakumar A, Gludish DW, Shilvock AR, Noden DM, et al. The left-right Pitx2 pathway drives organ-specific arterial and lymphatic development in the intestine. Dev Cell (2014) 31(6):690–706. doi:10.1016/j.devcel.2014.11.002
57. Stanczuk L, Martinez-Corral I, Ulvmar MH, Zhang Y, Lavina B, Fruttiger M, et al. cKit Lineage hemogenic endothelium-derived cells contribute to mesenteric lymphatic vessels. Cell Rep (2015) 10(10)1708–21. doi:10.1016/j.celrep.2015.02.026
58. Martinez-Corral I, Ulvmar MH, Stanczuk L, Tatin F, Kizhatil K, John SW, et al. Nonvenous origin of dermal lymphatic vasculature. Circ Res (2015) 116(10):1649–54. doi:10.1161/CIRCRESAHA.116.306170
59. Klotz L, Norman S, Vieira JM, Masters M, Rohling M, Dube KN, et al. Cardiac lymphatics are heterogeneous in origin and respond to injury. Nature (2015) 522(7554):62–7. doi:10.1038/nature14483
60. Nicenboim J, Malkinson G, Lupo T, Asaf L, Sela Y, Mayseless O, et al. Lymphatic vessels arise from specialized angioblasts within a venous niche. Nature (2015) 522(7554):56–61. doi:10.1038/nature14425
61. Ulvmar MH, Makinen T. Heterogeneity in the lymphatic vascular system and its origin. Cardiovasc Res (2016) 111(4):310–21. doi:10.1093/cvr/cvw175
62. Angeli V, Ginhoux F, Llodra J, Quemeneur L, Frenette PS, Skobe M, et al. B cell-driven lymphangiogenesis in inflamed lymph nodes enhances dendritic cell mobilization. Immunity (2006) 24(2):203–15. doi:10.1016/j.immuni.2006.01.003
63. Chyou S, Benahmed F, Chen J, Kumar V, Tian S, Lipp M, et al. Coordinated regulation of lymph node vascular-stromal growth first by CD11c+ cells and then by T and B cells. J Immunol (2011) 187(11):5558–67. doi:10.4049/jimmunol.1101724
64. Muniz LR, Pacer ME, Lira SA, Furtado GC. A critical role for dendritic cells in the formation of lymphatic vessels within tertiary lymphoid structures. J Immunol (2011) 187(2):828–34. doi:10.4049/jimmunol.1004233
65. Maruyama K, Ii M, Cursiefen C, Jackson DG, Keino H, Tomita M, et al. Inflammation-induced lymphangiogenesis in the cornea arises from CD11b-positive macrophages. J Clin Invest (2005) 115(9):2363–72. doi:10.1172/JCI23874
66. Ran S, Montgomery KE. Macrophage-mediated lymphangiogenesis: the emerging role of macrophages as lymphatic endothelial progenitors. Cancers (Basel) (2012) 4(3):618–57. doi:10.3390/cancers4030618
67. Kataru RP, Kim H, Jang C, Choi DK, Koh BI, Kim M, et al. T lymphocytes negatively regulate lymph node lymphatic vessel formation. Immunity (2011) 34(1):96–107. doi:10.1016/j.immuni.2010.12.016
68. Shin K, Kataru RP, Park HJ, Kwon BI, Kim TW, Hong YK, et al. TH2 cells and their cytokines regulate formation and function of lymphatic vessels. Nat Commun (2015) 6:6196. doi:10.1038/ncomms7196
69. Chauhan SK, Jin Y, Goyal S, Lee HS, Fuchsluger TA, Lee HK, et al. A novel pro-lymphangiogenic function for Th17/IL-17. Blood (2011) 118(17):4630–4. doi:10.1182/blood-2011-01-332049
70. Seeger H, Bonani M, Segerer S. The role of lymphatics in renal inflammation. Nephrol Dial Transplant (2012) 27(7):2634–41. doi:10.1093/ndt/gfs140
71. Furtado GC, Marinkovic T, Martin AP, Garin A, Hoch B, Hubner W, et al. Lymphotoxin beta receptor signaling is required for inflammatory lymphangiogenesis in the thyroid. Proc Natl Acad Sci U S A (2007) 104(12):5026–31. doi:10.1073/pnas.0606697104
72. Johnson LA, Jackson DG. Inflammation-induced secretion of CCL21 in lymphatic endothelium is a key regulator of integrin-mediated dendritic cell transmigration. Int Immunol (2010) 22(10):839–49. doi:10.1093/intimm/dxq435
73. Vigl B, Aebischer D, Nitschke M, Iolyeva M, Rothlin T, Antsiferova O, et al. Tissue inflammation modulates gene expression of lymphatic endothelial cells and dendritic cell migration in a stimulus-dependent manner. Blood (2011) 118(1):205–15. doi:10.1182/blood-2010-12-326447
74. Truman LA, Bentley KL, Smith EC, Massaro SA, Gonzalez DG, Haberman AM, et al. ProxTom lymphatic vessel reporter mice reveal Prox1 expression in the adrenal medulla, megakaryocytes, and platelets. Am J Pathol (2012) 180(4):1715–25. doi:10.1016/j.ajpath.2011.12.026
Keywords: lymph node, lymphatic vessel, high endothelial venule, tertiary lymphoid organ, autoimmunity, inflammation, cancer, lymphotoxin
Citation: Ruddle NH (2016) High Endothelial Venules and Lymphatic Vessels in Tertiary Lymphoid Organs: Characteristics, Functions, and Regulation. Front. Immunol. 7:491. doi: 10.3389/fimmu.2016.00491
Received: 16 August 2016; Accepted: 25 October 2016;
Published: 09 November 2016
Edited by:
Andreas Habenicht, Ludwig Maximilian University of Munich, GermanyReviewed by:
Ingrid E. Dumitriu, St. George’s University of London, UKCopyright: © 2016 Ruddle. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nancy H. Ruddle, nancy.ruddle@yale.edu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.