 Angela Castoldi
Angela Castoldi Cristiane Naffah de Souza
Cristiane Naffah de Souza Niels Olsen Saraiva Câmara
Niels Olsen Saraiva Câmara Pedro M. Moraes-Vieira
Pedro M. Moraes-Vieira
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 05 January 2016
Sec. Molecular Innate Immunity
Volume 6 - 2015 | https://doi.org/10.3389/fimmu.2015.00637
This article is part of the Research Topic Macrophages Role in Integrating Tissue Signals and Biological Processes in Chronic Inflammation and Fibrosis View all 11 articles
Immune cell infiltration in (white) adipose tissue (AT) during obesity is associated with the development of insulin resistance. In AT, the main population of leukocytes are macrophages. Macrophages can be classified into two major populations: M1, classically activated macrophages, and M2, alternatively activated macrophages, although recent studies have identified a broad range of macrophage subsets. During obesity, AT M1 macrophage numbers increase and correlate with AT inflammation and insulin resistance. Upon activation, pro-inflammatory M1 macrophages induce aerobic glycolysis. By contrast, in lean humans and mice, the number of M2 macrophages predominates. M2 macrophages secrete anti-inflammatory cytokines and utilize oxidative metabolism to maintain AT homeostasis. Here, we review the immunologic and metabolic functions of AT macrophages and their different facets in obesity and the metabolic syndrome.
Obesity is a prevalent metabolic disease characterized by excess accumulation of white adipose tissue (AT) due to increased food intake and changes in lifestyle (1, 2). Obesity leads to the development of a low-grade systemic chronic inflammatory state (3–6). According to the World Health Organization (WHO), 39% of adults over 18 years of age are overweight and 13% are clinically obese, translating to approximately 2 billion overweight adults where more than half a billion are obese (7).
A major player in systemic low-grade chronic inflammation in obesity is the increased numbers of AT pro-inflammatory macrophages and deregulated production and function of AT hormones and cytokines (2, 4). Besides its role in storing energy, AT is an important endocrine organ (8, 9), such that its dysfunction strongly contributes to the initiation and exacerbation of type 2 diabetes (T2D) (8, 10).
Insulin resistance is defined as a reduced response to insulin in liver, muscle, and AT. This impairment is due to the inhibition of the insulin-signaling pathway, leading to hyperglycemia. Insulin resistance is commonly associated with obesity and may precede the onset of T2D (11–13). One hypothesized reason for impaired insulin signaling has been thought to be due to the chronic systemic low-grade inflammation in obesity (14).
The finding that infiltration of monocytes, which differentiate into macrophages, is augmented in obesity is fundamental (15, 16). This results in pro-inflammatory macrophage and polarization leading to AT inflammation and insulin resistance (15, 17). Importantly, macrophages are crucial for regulating the immune system, specifically by restoring and maintaining AT homeostasis (18, 19).
In this review, we highlight the different functions of AT macrophages (ATMs) in the maintenance AT tissue homeostasis during lean, obese, and insulin resistant states.
The mechanisms by which inflammation increases during obesity are not fully understood. Increased pro-inflammatory cytokine secretion contributes to insulin resistance in obesity. Among these cytokines, tumor necrosis factor-α (TNF-α) was the first cytokine identified to be capable of inducing insulin resistance in adipocytes in vitro. In AT, the secretion of TNF-α is primarily derived from macrophages (20–22), and the accumulation of these immune cells in obesity contributes to the development of insulin resistance (23). This supports a key role for inflammation in the regulation of systemic metabolic homeostasis.
Macrophages make up to 40% of all AT cells in obese mice compared to 10% in lean mice (23). These cells are increased in AT during obesity due to increased amounts of several factors, including free fatty acids (FFAs), cholesterol, and lipopolysaccharide (LPS). Serum levels of LPS are elevated in obesity and, this cell wall component from Gram-negative bacteria, is linked to changes in the gut microbiota (metabolic endotoxemia) (24). LPS binds to and activates toll-like receptor 4 (TLR4) and its downstream signaling pathways in AT resident cells. These activated macrophages secrete cytokines and chemokines, such as monocyte chemoattractant protein-1 (MCP-1), and express C–C motif chemokine receptor-2 (CCR2) and CCR5, which in turn augment the recruitment of more monocytes and other leukocytes into AT (25–27). Macrophages share the same differentiation and recruitment molecules with other myeloid cells in many inflammatory conditions (28). As observed during bacterial inflammation (29), in obesity, macrophage activation is dependent on I kappa B kinase-β (IKK-β) (30). Arkan et al. showed that IKK-β activation in macrophages is sufficient for the development of insulin resistance, and mice with loss of IKK-β function only in myeloid cells are protected from obesity development and insulin resistance (30). These findings demonstrate the importance of macrophages in the context of insulin resistance development.
In addition to the activation and inflammatory profile of macrophages in the obese state, ATMs are highly adaptive to its lipid-rich environment. To maintain AT homeostasis in this lipid-rich microenvironment, macrophages increase their adiposity by activating lysosomal lipid metabolism (31). This may be a physiological response to buffer the increase in lipid concentrations released by adipocytes during obesity. This process does not classically activate ATMs, but it activates an immune cell differentiation program where high concentrations of lipids and FFAs induce a macrophage phenotype distinct from differentiated bone marrow macrophages (BMDM) (31). This phenotype is characterized by lipid accumulation in ATMs and increased expression of fatty acids transporters, such as CD36 and the lipid scavenger receptor Msr1 (31).
Several immune cells regulate AT inflammation, insulin resistance (32), and macrophage recruitment and differentiation (19, 33–35). There are two distinct macrophage populations found in AT. In healthy/lean AT, alternatively activated macrophages (M2) that express CD206 and CD301 on their surface and secrete anti-inflammatory cytokines predominates. On the other hand, obesity triggers the accumulation of classically activated macrophages (M1) characterized by CD11c surface expression, and expression of pro-inflammatory cytokines (17, 36), although this pan-classification spans a broad range of macrophage subtypes.
However, Kratz et al. recently described a different subtype of macrophage (37). They observed that treating macrophages with a mix of glucose, palmitate, and insulin (“metabolic activation”) generates a unique macrophage pro-inflammatory phenotype that is different from M1. This type of macrophage secretes pro-inflammatory cytokines, such as interleukin-1β (IL-1β) and TNF-α, whereby the secretion is dependent on peroxisome proliferator-activated receptor gamma (PPAR-γ) and p62 expression. In vivo, this phenotype is due to continuous and excessive exposure of ATMs to FFAs, such as palmitate, in a microenvironment that is saturated with glucose and insulin. In obesity, this differentiated macrophage subtype indicates the importance and the necessity to identify differentiated profiles of immune cells. Since there is a large spectrum of ATMs that have different immune profiles, we choose to focus on M1 and M2 subtypes of ATMs to better understand how metabolic alterations in ATMs impact obesity and insulin resistance.
M1 macrophages are associated with a pro-inflammatory profile. These macrophages are generally stimulated by T-helper 1 (Th1) type of cytokines, such as interferon-γ (IFN-γ), or by pathogen-associated molecular patterns (PAMPs), such as LPS (38). In turn, M1 macrophages secrete cytokines, including IL-6, TNF-α, IL-1β (39), IL-12, and IL-23 (40). M1 macrophages can also induce Th1 responses (41, 42). In general, these cells express high levels of major histocompatibility complex class II (MHC-II), CD80 and CD86 costimulatory molecules and CD68 (43). Moreover, M1 macrophages express Th1 cell-attracting chemokines, including CXCL9 and CXCL10 (44).
In addition to IFN-γ and LPS, there are several other molecules involved in M1 macrophage polarization, such as interferon regulatory factor (IRF), signal transducers and activators of transcription (STAT), and suppressor of cytokine signaling (SOCS). IRF5 is involved in M1 polarization by inducing the transcription of interleukin-12 subunit p40 (IL-12p40), IL-12p35, and IL-23p19, and by repressing the transcription of IL-10 (45). M1 macrophages express SOCS3, which promotes nitric oxide (NO) production (46). SOCS3 controls nuclear factor-κB (NF-κB) and phosphatidylinositol 3-kinase (PI3K) activity, favoring NO production in macrophages (46). The induction of inducible nitric oxide synthase (iNOS), another important molecule induced in M1 macrophages is dependent on TLR ligands, such as LPS, and activation of NF-κB, PI3K, and IFN-γ secretion (47, 48) (Table 1). Furthermore, myeloid differentiation primary response gene 88 (MyD88)-dependent pathway is also important for M1 polarization (49). The expression of TLR4/TLR2 is significantly higher in M1 when compared to M2 macrophages (50). The absence of TLR4 drives macrophages toward an M2 phenotype (51), indicating that activation and polarization of macrophages is, at least, in part dependent on TLRs.

Table 1. Differential requirement for stimuli and differential expression of transcription factors, cytokines, chemokines, and other molecules by M1 and M2 macrophages.
In contrast to M1 macrophages generated in vitro, which do not express CD11c, M1 ATMs express CD11c concomitant with F4/80 and CD11b (17, 52–54). Interestingly, the expression of CD11c in vitro by BMDM can be induced if BMDMs are differentiated in the presence of adipocytes (31, 37). This indicates the importance and requirement of adipocytes in orchestrating the functional phenotype of ATMs.
The recruitment of monocytes, which in AT gives rise to CD11c+ ATMs, is dependent on CCR2, CCR5, and MCP-1 (55, 56). Nagareddy et al. demonstrated that ATM-derived IL-1β promotes monocyte release from the bone marrow (57) and MCP-1 induces M1 ATM proliferation in AT (58). These processes are important to promote macrophage accumulation in the AT during obesity and sustain AT inflammation and insulin resistance (58).
Obesity-associated insulin resistance correlates with elevated levels of pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6 (42, 59–62). These cytokines are secreted by both adipocytes and ATMs due to increased levels of pro-inflammatory factors released during obesity development. These factors include FFA, triglycerides, resistin, leptin, retinol-binding protein 4 (RBP4), IL-6, TNF-α, and IL-1β, among others (31, 63, 64).
Secretion of these factors activates several inflammatory signal transduction pathways in macrophages and adipocytes, which are required for obesity-induced insulin resistance. The stress-responsive c-Jun NH2-terminal kinase (JNK 1 and 2) (65), inhibitor of κB kinase (IKK) (66), extracellular signal-regulated kinase 1 and 2 (ERK 1 and 2) (67), and mitogen-activated protein kinase p38 (p38 MAPK) are responsible for alterations in the insulin receptor signaling pathway (68). These alterations lead to decreased tyrosine phosphorylation of insulin receptor substrate (IRS-1 and -2), PI3K activation followed by a decreased serine phosphorylation of Akt and consequently insulin resistance (66, 68–72). There is a crosstalk between the two isoforms of JNK (JNK1 and JNK2) that contributes to obesity-induced insulin resistance development. The balance between these two molecules determines the total activity of JNK in fat tissues (73). Hematopoietic activation of JNK1 is a major player in obesity-induced inflammation and insulin resistance (74). Corroborating this, Han et al. verified that knockdown of both JNK 1 and 2 in macrophages protect mice from HFD-induced insulin resistance and AT inflammation (65). Similarly, Vallerie et al. showed that myeloid JNK1 is a regulator of cytokine expression in AT during the late, but not early states of obesity development (75).
Toll-like receptors and inflammasomes are activated in obesity by damage-associated molecular pattern molecules (DAMPs), such as high-mobility group box 1 (HMGB1) and oxidized low-density lipoprotein (Ox-LDL), RBP4 or PAMPs, such as LPS (24, 76–80). TLRs and inflammasomes modulate macrophage polarization due to activation of NF-κB, STAT1, and caspase-1 to induce IL-1β production (81, 82). Upon activation, these receptors contribute to low-grade chronic inflammation in obesity, leading to M1 polarization of ATMs. Importantly, TLR4 expression is increased in ATMs during obesity (83). Thus, many studies have investigated the role of TLR4 and nod-like receptor protein 3 (NLRP3) in knockout mouse models in HFD-induced obesity (17, 23, 51, 84).
Toll-like receptor 4 deficiency in HFD-fed mice ameliorates AT inflammation, insulin resistance, and adiposity (83, 85, 86). The reduction in inflammation is due to decreased macrophage infiltration and a switch from M1 to M2 macrophage profile (51, 83, 85, 87).
Nod-like receptor protein 3 inflammasome also plays a key role in the development of AT inflammation and insulin resistance (88, 89). Expression of NLRP3, apoptosis-associated speck-like protein containing CARD (ASC), caspase-1, and IL-1β are all upregulated in AT of obese mice, as well as the mature form of IL-1β (82, 90). The secreted IL-1β binds to IL-1R and activates NF-κB and MAPK pathways, thereby impairing insulin signaling through the activation of IRS-1 in adipocytes leading to insulin resistance (82, 91).
Functional deletion of NLRP3 and caspase-1 ameliorate HFD-induced insulin resistance and AT inflammation (82, 90, 92). Moreover, weight loss and insulin sensitivity in patients with T2D is associated with decreased AT expression of NLRP3 and IL-1β (82). Protection from insulin resistance and inflammation following loss of functional NLRP3 may be due to a shift in macrophage polarization, since NLRP3-knockout mice have decreased M1 and increased M2 gene expression profiles in AT (84).
In addition to these important signaling pathways, the mammalian target of rapamycin (TOR) has an important function in insulin resistance. It is able to sense nutrients and respond by altering the cellular metabolism in different kind of cells, including ATMs (93). Insulin, glucose, leptin, and other growth factors and cytokines activate mTOR pathway via PI3K–Akt signaling pathway (94). The protein kinase Akt phosphorylates and inhibits TSC2 and, consequently, activates mTORC1 (95, 96). Activation of these metabolic sensors, mainly PI3Kγ, is important for immune cell functions. PI3Kγ activation in hematopoietic cells contributes to the development of obesity and insulin resistance. PI3Kγ activity in the non-hematopoietic compartment is critical during obesity (97). Moreover, the catalytic subunit of PI3Kγ, p110γ, was shown to be activated during obesity. Absence of functional p110 improved insulin sensitivity with reduced infiltration of pro-inflammatory macrophages and inflammatory marker expression in AT. In addition, specific depletion of PI3Kγ in bone marrow cells as well as pharmacological blockade also inhibited macrophage infiltration during obesity and insulin resistance (98). Together, these data indicate that activation of metabolic sensors in immune cells during obesity is essential for inflammation and insulin resistance development.
Defects in mTORC1 regulation can lead to metabolic dysfunction, including T2D (93). Deletion of mTORC1 in macrophages diminishes AT inflammation and protects mice against HFD-induced insulin resistance (99, 100). mTORC1 disruption suppresses HK1-dependent glycolysis, caspase-1 activation, IL-1β, and IL-18 secretion in vitro and in vivo and induces M2 polarization (100). In accordance, Jiang et al. showed that mTORC1 depletion in macrophages protects mice against HFD-induced AT inflammation and insulin resistance through the inhibition of IRE1α/JNK/NF-κB pathways (99).
In 2013, Horng et al. demonstrated in vitro and in vivo that TSC1 deletion (Tsc1 deficiency, thereby mTORC1 is constitutively active) in macrophages leads to a marked defect in M2 polarization in response to IL-4, although LPS stimulation induced inflammatory responses in an mTOR-dependent manner (101). Moreover, in obesity, nutrient sensing by mTORC1 regulates the switch of ATMs from M2 to M1 (12).
More recently, Zhu et al. proposed that TSC1 deletion in macrophages intensifies the M1 polarization (102). TSC1 inhibits M1 polarization by suppressing the Ras GTPase/Raf1/MEK/ERK signaling pathway in an mTOR-independent manner, whereas TSC1 promotes M2 properties by mTOR-dependent CCAAT/enhancer-binding protein-β pathway (102). These findings indicate a critical role for TSC1 in orchestrating macrophage polarization via mTOR-dependent and -independent pathways (102) (Figure 1).
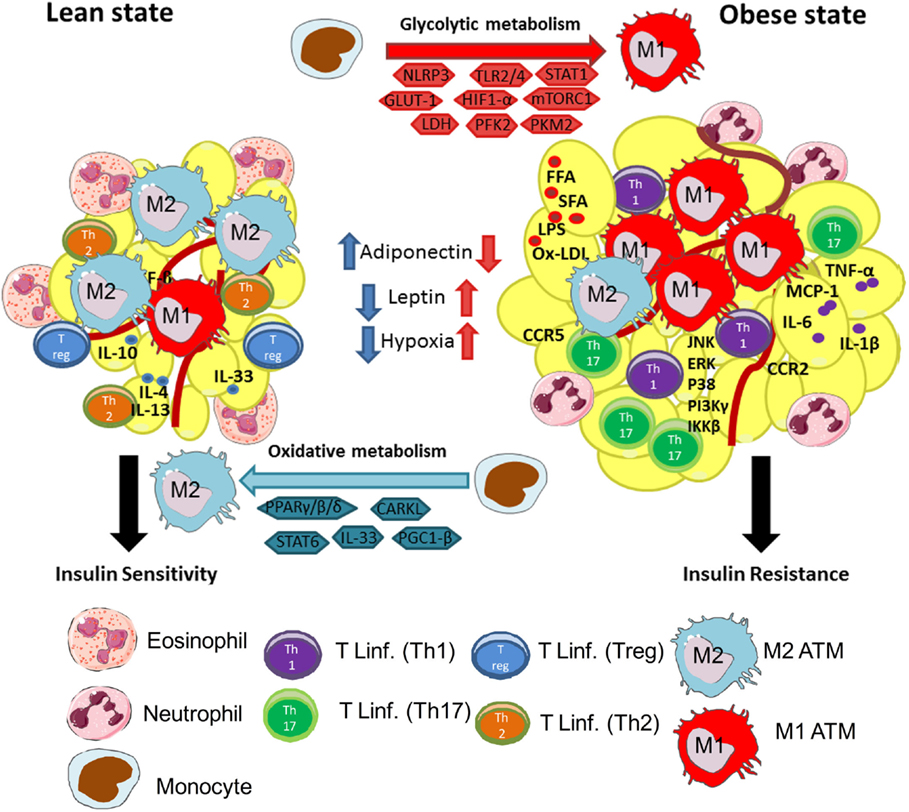
Figure 1. Macrophages are central players in lean and obese states. Lean adipose tissue is abundant in immune cells, such as eosinophils, Th2 T cells, ILC2, regulatory T cells (Treg), and M2 (anti-inflammatory) macrophages. These cells are known to secrete anti-inflammatory cytokines, such as IL-10, IL-4, IL-13, and IL-33, to maintain AT homeostasis and controlling insulin sensitivity. M2 macrophages use oxidative metabolism through PPARγ/β/δ, CARKL, STAT6, and PGC-1β. These events are central to maintain a healthy environment in adipose tissue. In the other hand, during obesity, AT is characterized by infiltration of several immune cells, such as monocytes, neutrophils, Th1 and Th17 lymphocytes, and M1 (pro-inflammatory) macrophages. The increased secretion of FFA, SFA, Ox-LDL, and LPS in obesity activates resident macrophages and adipocytes leading to secretion of pro-inflammatory cytokines, such as TNF-α, IL-6, IL-1β, and chemokines MCP-1, CCR2, and CCR5. This process will instigate the recruitment of monocytes and differentiation of M1 macrophages in AT. Besides, activation of pro-inflammatory signaling pathways downstream to TLRs, such as JNK, ERK, p38, IκB, IKKβ, and Pi3Kγ, inhibit insulin receptor signaling, leading to insulin resistance. Moreover, in obese AT, M1 macrophages use glycolytic metabolism and require activation of intracellular molecules, such as NLRP3, TLR2/4, STAT1, GLUT-1, HIF-1α, mTORC1, PFK2, and PKM2, and conversion of pyruvate to lactate by LDH. Activation of glycolysis in macrophages is central to maintain their pro-inflammatory profile.
Increased M1 activation in AT is involved in activation of the adaptive immune response through the recruitment and activation of T cells. Increased recruitment of CD4+ T cells correlates with increased M1 polarization. Also, M1 polarization appears to be dependent on AT Th1 polarization (42, 103–106). In addition, during obesity, the activation of Th1 responses in AT are mediated by mTORC1, since this molecule is necessary for polarization of T lymphocytes toward a Th1 phenotype (107). Moreover, circulating leptin, which is elevated during obesity, activates mTOR pathway, and also induces Th1 responses (108, 109). Thus, Th1 polarization is dependent on M1 polarization, and it is critical for the development of insulin resistance (104).
Together, several pathways mediate the induction/activation of ATMs to maintain AT homeostasis, which can also be affected by changes in systemic and cellular metabolism.
M2 macrophages are associated with tissue remodeling and inflammation resolution (110). M2 macrophages have immunosuppressive properties, have high phagocytic capacity, and secrete extracellular matrix components, angiogenic and chemotactic factors, anti-inflammatory cytokines, and growth factors, such as IL-10 and transforming growth factor β (TGF-β) (111, 112). M2 macrophages are characterized by upregulated expression of Dectin-1, CD206, scavenger receptor A, scavenger receptor B-1, CD163, CCR2, CXCR1, CXCR2, and MgL 1/2 (36). Moreover, the expression of arginase-1 (Arg1), PPAR-γ, and transcription factor found in inflammatory zone 1 (FIZZ1), which is specific of murine M2 macrophages, are necessary for collagen synthesis, further supporting the role of these cells in tissue remodeling (44) (Table 1).
In vitro, M2 macrophages appear to be a heterogeneous population induced by a variety of stimuli. M2a is induced by IL-4 or IL-13 express high levels of CD206 and has immunoregulatory functions (38, 113–115). M2b is induced by immune complexes and TLRs or IL-1R agonists. Both M2a and M2b have an immunoregulatory role through down-regulation of IL-12, IL-6, and TNF (116). M2c is induced by IL-10 and glucocorticoids. It has an immunosuppressive phenotype and participates in tissue remodeling. M2c secretes pro-fibrotic factors, such as TGF-β, CCL17, and CCL22 (38, 116). In addition, expansion of M2c macrophages is negatively regulated by PPAR-γ, which is expressed in M2 ATM (117). Although significant progress has been made in characterizing M2 subpopulations, it still not completely understood how these cells behave in vivo.
The microenvironment in a lean AT is composed of a 4:1 M2:M1 ratio (118). The presence of eosinophils and regulatory T cells (Tregs), which secrete the cytokines IL-4/IL-13 and IL-10, respectively, polarizes ATMs toward an anti-inflammatory phenotype (119–121). In lean AT, adipocytes secrete higher levels adiponectin compared with obese AT. Adiponectin enhances insulin sensitivity and increases M2 macrophage polarization (121). These cells and their secretome maintain the positive balance of M2 macrophages in lean AT.
Obesity inversely correlates with AT Tregs (122, 123). Moreover, Tregs can induce M2 macrophage differentiation in mice through IL-10 and TGF-β (124). In lean AT, these cells are involved in the regulation of tissue homeostasis and help to maintain the M2 macrophage population (122).
Recently, new regulatory players in AT homeostasis have been identified: innate lymphoid type 2 cells (ILC2s) and IL-33. ILC2s are a regulatory subtype of ILCs. These cells were divided into three distinct populations, ILCs 1, 2, and 3 (125–127). These subpopulations of ILCs are analogous to the largely known CD4+ T helper subsets: Th1, Th2, and Th17, respectively, with respect to cytokine profile expression (128). However, ILCs do not have T-cell receptors and respond to antigenic signals in the absence of antigen specificity (128). ILCs are activated by the cytokine IL-33 and produce large amounts of the type 2 cytokines: IL-5 and IL-13 (129).
Interleukin-33 is constitutively present in humans and mice, mainly in specialized populations of epithelial and endothelial cells (130, 131). Its receptor (ST2) is highly expressed in ILC2s and Th2 lymphocytes, and it is also found in eosinophils, mast cells, dendritic cells, basophils, myeloid-derived suppressor cells, and Tregs (132).
Interleukin-33, as well as ILC2s, has been in the spotlight due to their putative contributions in the improvement of obesity-induced insulin resistance. Upon binding to its receptor, IL-33 induces the production of large amounts of anti-inflammatory cytokines by AT ILC2s and also the polarization of ATMs toward an M2 phenotype (133). This results in AT mass reduction and improves insulin resistance (133, 134). Han and colleagues investigated ST2 expression in murine Tregs in lean and obese visceral AT. AT Tregs from lean mice express higher levels of ST2 compared to AT Tregs from obese mice. Moreover, treatment with IL-33 restored the ST2-positive Treg population, reduced AT inflammation, and improved insulin resistance (133).
In this context, Brestoff et al. demonstrated that IL-33 plays an important role in the maintenance of ILC2s in AT, promoting energy expenditure, and reducing adiposity in mice (135). This decrease in adiposity was due to caloric expenditure upon the induction of uncoupling protein 1 (UCP1) expression in AT, a process called “beiging” or “browning” (136, 137). UCP1 protein is limited to beige and brown adipocytes and regulates caloric expenditure (135). In agreement with Artis et al., Chalwa’s et al. found that IL-33 promoted the accumulation and activation of ILC2s in mouse AT, leading to the biogenesis of beige fat, which is crucial for AT metabolic homeostasis (138) (Figure 1).
Taken together, these studies demonstrate the importance of alternatively activated macrophages to maintain the tissue homeostasis, especially in AT. Moreover, the discovery of new alternative pathways for the polarization of ATMs toward an M2 phenotype is necessary to better understand the mechanisms by which insulin sensitivity in obesity.
In addition to cytokines, the availability of substrates in tissues orchestrates macrophage function. Cellular metabolism is not static but is rather a dynamic process that allows cells to adapt to the microenvironment (139). The type of nutrient substrate is critical for ATM function. Saturated fatty acids (SFAs) are pro-inflammatory and induce M1-like phenotype, while certain types of unsaturated fatty acids (UFAs), such as omega-3 and branched fatty acid esters of hydroxy fatty acids (FAHFA) (140), are anti-inflammatory and induce an M2-like phenotype (141).
M1 macrophages preferentially metabolize glucose as an energy substrate (142). During activation, macrophages alter its metabolism to support survival and cellular functions. The metabolism of M1 macrophages upon activation is characterized by induced aerobic glycolysis with increased glucose uptake and the conversion of pyruvate to lactate by lactate dehydrogenase (LDH) (143). This activation in aerobic glycolysis decreases respiratory chain activity due to increased ROS levels (144). This metabolic switch is necessary for NO production, an important effector of immune microbicidal activity and pro-inflammatory M1 macrophage responses (144).
In addition, the expression of glucose transporter-1 (GLUT-1) drives the pro-inflammatory phenotype of M1 macrophages, increases glucose uptake, and, subsequently, augments glucose metabolism (145).
One important molecule regulating glycolysis and macrophage activation is hypoxia inducible factor-1α (Hif-1α). Hif-1α induces a pro-inflammatory phenotype in macrophages (146) via TLR4 activation, which involves the PI3K/Akt signaling pathway (147). Low oxygen (O2) tension and inflammatory responses increase TLR4 expression in macrophages (148). Moreover, M1 macrophages co-localize with AT hypoxic areas in obese mice and are associated with increased inflammatory responses (147–149). Because these macrophages need to adapt to the obesity-induced hypoxic tissue environment, activating anaerobic glycolysis under these circumstances best serves these immune cells to support their rapid and demanding energy requirements (143).
Activation of macrophages with LPS also results in increased levels of succinate and malate (150). Succinate, in particular, drives IL-1β production, which is dependent on Hif-1α activation (150). In addition, pyruvate kinase M2 (PKM2), a critical determinant of macrophage activation by LPS, promotes inflammatory responses (151). Activation of PKM2 plays a key role in stabilizing Hif-1α and Hif-1α-dependent genes, such as IL-1β expression. LPS induces dimerization of PKM2 that in turn complexes with Hif-1α. This complex directly binds to the IL-1β promoter, an event that is inhibited by the activation of tetrameric PKM2, which induces M2 macrophage differentiation and attenuates LPS-induced M1 macrophages (151). Thus, PKM2 in its dimeric form is required for glycolytic reprograming in response to LPS. The dimeric form of PKM2 plays role in Hif-1α function, whereas the tetrameric form of PKM2 impairs the ability of PKM2 to promote transcriptional activity of Hif-1α and LPS-induced IL-1β expression (151).
Nonetheless, the microenvironment rich in LPS and IFN-γ also enhances M1 macrophage polarization and glycolysis activation independently of Hif-1α. This occurs upon 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase (PFK2) induction (152).
In contrast to M1 glycolytic metabolism, M2 macrophages utilize oxidative metabolism (142). The induction of oxidative metabolism in M1 macrophages shifts their phenotype toward an M2 profile (152). Moreover, the overexpression of carbohydrate kinase-like protein (CARKL), which regulates the production of sedoheptulose-7-phosphate (S7P), an intermediate of the pentose phosphate pathway (PPP) (153) results in decreased production of pro-inflammatory cytokines, which suggests a shift toward an M2 macrophages phenotype (154).
Besides CARKL, the coactivator protein PPAR-γ-coactivator-1β (PGC1-β) induces mitochondrial respiration as well as mitochondrial biogenesis. This is a key player in the metabolic switch of macrophages from M1 to M2 phenotype (142, 144). Blocking PGC1-β results in impaired M2 macrophage metabolism and function (142). Thus, identifying mechanisms that modulate the metabolism of macrophages may dampen the onset and exacerbation of inflammatory processes.
Adipose tissue-derived IL-4 and IL-13 signals through IRF/STAT to activate STAT6 in M2 macrophages (44, 113). STAT6 induces the expression of transcriptional regulators, such as PPAR-γ (44). PPAR-γ maintains the metabolic switch toward oxidative metabolism and promotes M2 gene expression (Arg1) to amplify the effector phenotype of M2 macrophages (collagen synthesis) (31, 155, 156). Other members of the PPAR family, PPARβ/δ, appears to differentially influence macrophage activation, along with IL-4 and IL-13, and promotes an alternative M2 macrophage phenotype (156). Myeloid deletion of PPARβ/δ leads to glucose intolerance and insulin resistance (27), indicating that expression of PPARs transcription factors is crucial to maintain the M2 phenotype through the secretion of Th2 cytokines (Figure 1).
Hypoxia inducible factor-2α has been shown to regulate the transcription of Arg1, which is expressed by M2 macrophages (157). However, Hif-2α also controls IL-1β production and NF-κB activity, which is associated with an M1 phenotype (150, 157). Thus, although Hif-2α appears to have a role in macrophage polarization, more studies are needed to better understand the importance of this transcription factor for macrophage phenotype, metabolism, and function.
It is still unclear how M2 macrophages metabolism is regulated during obesity and the role of M2 macrophage metabolism for the development of insulin resistance. Nevertheless, in lean state, they have an oxidative metabolism, which may shift to glycolytic metabolism, during obesity, due to a pro-inflammatory environment and further studies are needed to better understand their role in obesity.
Macrophages are central mediators of obesity-induced AT inflammation and insulin resistance. They also are key cells for maintenance of AT homeostasis. Recently, several reports described the importance of these cells as regulators of insulin sensitivity, which involves the activation of innate immune receptors, transcription factors, and intracellular metabolism to support the either pro- or anti-inflammatory AT phenotype. Thus, macrophages have a dual role, changing their status to support immune responses, obesity development, and related diseases.
AC and CN have contributed equally in the writing of this manuscript. NC and PM-V contributed in the writing and corrections of this manuscript. AC and CN have contributed equally to this review.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Dr. Jennifer Lee from Division of Endocrinology and Metabolism, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA, USA for helpful comments and edits on the manuscript.
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, grant numbers 11/15682-4, 12/02270-2, 14/10910-7, 15/18121-4) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Renal Immunopathogy Laboratory CNPq/Inserm and Complex Fluids INCT).
2. Mokdad AH, Ford ES, Bowman BA, Dietz WH, Vinicor F, Bales VS, et al. Prevalence of obesity, diabetes, and obesity-related health risk factors, 2001. JAMA (2003) 289:76–9. doi:10.1001/jama.289.1.76
3. Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science (1993) 259:87–91. doi:10.1126/science.7678183
4. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest (2003) 112:1821–30. doi:10.1172/JCI200319451
5. Qatanani M, Lazar MA. Mechanisms of obesity-associated insulin resistance: many choices on the menu. Genes Dev (2007) 21:1443–55. doi:10.1101/gad.1550907
6. Hotamisligil GS, Erbay E. Nutrient sensing and inflammation in metabolic diseases. Nat Rev Immunol (2008) 8:923–34. doi:10.1038/nri2449
7. WHO. World Health Organization. Obesity and Overweight, 2015 [Online] (2015). Available from: http://www.who.int/mediacentre/factsheets/fs311/en/
8. Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab (2004) 89(6):2548–56. doi:10.1210/jc.2004-0395
9. Andrade-Oliveira V, Camara NO, Moraes-Vieira PM. Adipokines as drug targets in diabetes and underlying disturbances. J Diabetes Res (2015) 2015:681612. doi:10.1155/2015/681612
10. Moraes-Vieira PM, Bassi EJ, Araujo RC, Camara NO. Leptin as a link between the immune system and kidney-related diseases: leading actor or just a coadjuvant? Obes Rev (2012) 13:733–43. doi:10.1111/j.1467-789X.2012.00997.x
11. Cefalu WT. Insulin resistance: cellular and clinical concepts. Exp Biol Med (Maywood) (2001) 226:13–26.
12. Chawla A, Nguyen KD, Goh YP. Macrophage-mediated inflammation in metabolic disease. Nat Rev Immunol (2011) 11:738–49. doi:10.1038/nri3071
13. Lee BC, Lee J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim Biophys Acta (2014) 1842:446–62. doi:10.1016/j.bbadis.2013.05.017
14. Laakso M, Kuusisto J. Insulin resistance and hyperglycaemia in cardiovascular disease development. Nat Rev Endocrinol (2014) 10:293–302. doi:10.1038/nrendo.2014.29
15. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest (2006) 116:1494–505. doi:10.1172/JCI26498
16. Masoodi M, Kuda O, Rossmeisl M, Flachs P, Kopecky J. Lipid signaling in adipose tissue: connecting inflammation & metabolism. Biochim Biophys Acta (2015) 1851:503–18. doi:10.1016/j.bbalip.2014.09.023
17. Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest (2007) 117:175–84. doi:10.1172/JCI29881
18. Ferrante AW Jr. Macrophages, fat, and the emergence of immunometabolism. J Clin Invest (2013) 123:4992–3. doi:10.1172/JCI73658
19. Brestoff JR, Artis D. Immune regulation of metabolic homeostasis in health and disease. Cell (2015) 161:146–60. doi:10.1016/j.cell.2015.02.022
20. Hofmann C, Lorenz K, Braithwaite SS, Colca JR, Palazuk BJ, Hotamisligil GS, et al. Altered gene expression for tumor necrosis factor-alpha and its receptors during drug and dietary modulation of insulin resistance. Endocrinology (1994) 134:264–70. doi:10.1210/en.134.1.264
21. Hotamisligil GS, Budavari A, Murray D, Spiegelman BM. Reduced tyrosine kinase activity of the insulin receptor in obesity-diabetes. Central role of tumor necrosis factor-alpha. J Clin Invest (1994) 94:1543–9. doi:10.1172/JCI117495
22. Hotamisligil GS, Spiegelman BM. Tumor necrosis factor alpha: a key component of the obesity-diabetes link. Diabetes (1994) 43:1271–8. doi:10.2337/diab.43.11.1271
23. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest (2003) 112:1796–808. doi:10.1172/JCI200319246
24. Caricilli AM, Picardi PK, De Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol (2011) 9:e1001212. doi:10.1371/journal.pbio.1001212
25. Wang P, Mariman E, Renes J, Keijer J. The secretory function of adipocytes in the physiology of white adipose tissue. J Cell Physiol (2008) 216:3–13. doi:10.1002/jcp.21386
26. Kosteli A, Sugaru E, Haemmerle G, Martin JF, Lei J, Zechner R, et al. Weight loss and lipolysis promote a dynamic immune response in murine adipose tissue. J Clin Invest (2010) 120:3466–79. doi:10.1172/JCI42845
27. Olefsky JM, Glass CK. Macrophages, inflammation, and insulin resistance. Annu Rev Physiol (2010) 72:219–46. doi:10.1146/annurev-physiol-021909-135846
28. Pollard JW. Trophic macrophages in development and disease. Nat Rev Immunol (2009) 9:259–70. doi:10.1038/nri2528
29. Yang F, Tang E, Guan K, Wang CY. IKK beta plays an essential role in the phosphorylation of RelA/p65 on serine 536 induced by lipopolysaccharide. J Immunol (2003) 170:5630–5. doi:10.4049/jimmunol.170.11.5630
30. Arkan MC, Hevener AL, Greten FR, Maeda S, Li ZW, Long JM, et al. IKK-beta links inflammation to obesity-induced insulin resistance. Nat Med (2005) 11:191–8. doi:10.1038/nm1185
31. Xu X, Grijalva A, Skowronski A, Van Eijk M, Serlie MJ, Ferrante AW Jr. Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation. Cell Metab (2013) 18:816–30. doi:10.1016/j.cmet.2013.11.001
32. Odegaard JI, Chawla A. Pleiotropic actions of insulin resistance and inflammation in metabolic homeostasis. Science (2013) 339:172–7. doi:10.1126/science.1230721
33. Molofsky AB, Nussbaum JC, Liang HE, Van Dyken SJ, Cheng LE, Mohapatra A, et al. Innate lymphoid type 2 cells sustain visceral adipose tissue eosinophils and alternatively activated macrophages. J Exp Med (2013) 210:535–49. doi:10.1084/jem.20121964
34. Mathis D. Immunological goings-on in visceral adipose tissue. Cell Metab (2013) 17:851–9. doi:10.1016/j.cmet.2013.05.008
35. Kolodin D, Van Panhuys N, Li C, Magnuson AM, Cipolletta D, Miller CM, et al. Antigen- and cytokine-driven accumulation of regulatory T cells in visceral adipose tissue of lean mice. Cell Metab (2015) 21:543–57. doi:10.1016/j.cmet.2015.03.005
36. Martinez FO, Helming L, Gordon S. Alternative activation of macrophages: an immunologic functional perspective. Annu Rev Immunol (2009) 27:451–83. doi:10.1146/annurev.immunol.021908.132532
37. Kratz M, Coats BR, Hisert KB, Hagman D, Mutskov V, Peris E, et al. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophages. Cell Metab (2014) 20:614–25. doi:10.1016/j.cmet.2014.08.010
38. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol (2004) 25:677–86. doi:10.1016/j.it.2004.09.015
39. Goerdt S, Politz O, Schledzewski K, Birk R, Gratchev A, Guillot P, et al. Alternative versus classical activation of macrophages. Pathobiology (1999) 67:222–6. doi:10.1159/000028096
40. Hao NB, Lu MH, Fan YH, Cao YL, Zhang ZR, Yang SM. Macrophages in tumor microenvironments and the progression of tumors. Clin Dev Immunol (2012) 2012:948098. doi:10.1155/2012/948098
41. Noel W, Raes G, Hassanzadeh Ghassabeh G, De Baetselier P, Beschin A. Alternatively activated macrophages during parasite infections. Trends Parasitol (2004) 20:126–33. doi:10.1016/j.pt.2004.01.004
42. Moraes-Vieira PM, Yore MM, Dwyer PM, Syed I, Aryal P, Kahn BB. RBP4 activates antigen-presenting cells, leading to adipose tissue inflammation and systemic insulin resistance. Cell Metab (2014) 19:512–26. doi:10.1016/j.cmet.2014.01.018
43. Cucak H, Grunnet LG, Rosendahl A. Accumulation of M1-like macrophages in type 2 diabetic islets is followed by a systemic shift in macrophage polarization. J Leukoc Biol (2014) 95:149–60. doi:10.1189/jlb.0213075
44. Martinez FO, Gordon S, Locati M, Mantovani A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: new molecules and patterns of gene expression. J Immunol (2006) 177:7303–11. doi:10.4049/jimmunol.177.10.7303
45. Krausgruber T, Blazek K, Smallie T, Alzabin S, Lockstone H, Sahgal N, et al. IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat Immunol (2011) 12:231–8. doi:10.1038/ni.1990
46. Arnold CE, Whyte CS, Gordon P, Barker RN, Rees AJ, Wilson HM. A critical role for suppressor of cytokine signalling 3 in promoting M1 macrophage activation and function in vitro and in vivo. Immunology (2014) 141:96–110. doi:10.1111/imm.12173
47. Bogdan C, Rollinghoff M, Diefenbach A. The role of nitric oxide in innate immunity. Immunol Rev (2000) 173:17–26. doi:10.1034/j.1600-065X.2000.917307.x
48. Niedbala W, Cai B, Liew FY. Role of nitric oxide in the regulation of T cell functions. Ann Rheum Dis (2006) 65(Suppl 3):iii37–40. doi:10.1136/ard.2006.058446
49. Braga TT, Correa-Costa M, Guise YF, Castoldi A, De Oliveira CD, Hyane MI, et al. MyD88 signaling pathway is involved in renal fibrosis by favoring a TH2 immune response and activating alternative M2 macrophages. Mol Med (2012) 18:1231–9. doi:10.2119/molmed.2012.00131
50. Wang N, Liang H, Zen K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front Immunol (2014) 5:614. doi:10.3389/fimmu.2014.00614
51. Orr JS, Puglisi MJ, Ellacott KL, Lumeng CN, Wasserman DH, Hasty AH. Toll-like receptor 4 deficiency promotes the alternative activation of adipose tissue macrophages. Diabetes (2012) 61:2718–27. doi:10.2337/db11-1595
52. Fujisaka S, Usui I, Bukhari A, Ikutani M, Oya T, Kanatani Y, et al. Regulatory mechanisms for adipose tissue M1 and M2 macrophages in diet-induced obese mice. Diabetes (2009) 58:2574–82. doi:10.2337/db08-1475
53. Shaul ME, Bennett G, Strissel KJ, Greenberg AS, Obin MS. Dynamic, M2-like remodeling phenotypes of CD11c+ adipose tissue macrophages during high-fat diet-induced obesity in mice. Diabetes (2010) 59:1171–81. doi:10.2337/db09-1402
54. Oh DY, Morinaga H, Talukdar S, Bae EJ, Olefsky JM. Increased macrophage migration into adipose tissue in obese mice. Diabetes (2012) 61:346–54. doi:10.2337/db11-0860
55. Weisberg SP, Hunter D, Huber R, Lemieux J, Slaymaker S, Vaddi K, et al. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J Clin Invest (2006) 116:115–24. doi:10.1172/JCI24335
56. Kitade H, Sawamoto K, Nagashimada M, Inoue H, Yamamoto Y, Sai Y, et al. CCR5 plays a critical role in obesity-induced adipose tissue inflammation and insulin resistance by regulating both macrophage recruitment and M1/M2 status. Diabetes (2012) 61:1680–90. doi:10.2337/db11-1506
57. Nagareddy PR, Kraakman M, Masters SL, Stirzaker RA, Gorman DJ, Grant RW, et al. Adipose tissue macrophages promote myelopoiesis and monocytosis in obesity. Cell Metab (2014) 19:821–35. doi:10.1016/j.cmet.2014.03.029
58. Amano SU, Cohen JL, Vangala P, Tencerova M, Nicoloro SM, Yawe JC, et al. Local proliferation of macrophages contributes to obesity-associated adipose tissue inflammation. Cell Metab (2014) 19:162–71. doi:10.1016/j.cmet.2013.11.017
59. Borst SE. The role of TNF-alpha in insulin resistance. Endocrine (2004) 23(2–3):177. doi:10.1385/ENDO
60. Maachi M, Pieroni L, Bruckert E, Jardel C, Fellahi S, Hainque B, et al. Systemic low-grade inflammation is related to both circulating and adipose tissue TNFalpha, leptin and IL-6 levels in obese women. Int J Obes Relat Metab Disord (2004) 28:993–7. doi:10.1038/sj.ijo.0802718
61. Monteiro R, Azevedo I. Chronic inflammation in obesity and the metabolic syndrome. Mediators Inflamm (2010) 2010:289645. doi:10.1155/2010/289645
62. Fjeldborg K, Pedersen SB, Moller HJ, Christiansen T, Bennetzen M, Richelsen B. Human adipose tissue macrophages are enhanced but changed to an anti-inflammatory profile in obesity. J Immunol Res (2014) 2014:309548. doi:10.1155/2014/309548
63. Boden G, Chen X, Ruiz J, White JV, Rossetti L. Mechanisms of fatty acid-induced inhibition of glucose uptake. J Clin Invest (1994) 93:2438–46. doi:10.1172/JCI117252
64. Boden G. Obesity and free fatty acids. Endocrinol Metab Clin North Am (2008) 37:635–646, viii–ix. doi:10.1016/j.ecl.2008.06.007
65. Han MS, Jung DY, Morel C, Lakhani SA, Kim JK, Flavell RA, et al. JNK expression by macrophages promotes obesity-induced insulin resistance and inflammation. Science (2013) 339:218–22. doi:10.1126/science.1227568
66. Gao Z, Hwang D, Bataille F, Lefevre M, York D, Quon MJ, et al. Serine phosphorylation of insulin receptor substrate 1 by inhibitor kappa B kinase complex. J Biol Chem (2002) 277:48115–21. doi:10.1074/jbc.M209459200
67. Banks AS, McAllister FE, Camporez JP, Zushin PJ, Jurczak MJ, Laznik-Bogoslavski D, et al. An ERK/Cdk5 axis controls the diabetogenic actions of PPARgamma. Nature (2015) 517:391–5. doi:10.1038/nature13887
68. Carlson CJ, Koterski S, Sciotti RJ, Poccard GB, Rondinone CM. Enhanced basal activation of mitogen-activated protein kinases in adipocytes from type 2 diabetes: potential role of p38 in the downregulation of GLUT4 expression. Diabetes (2003) 52:634–41. doi:10.2337/diabetes.52.3.634
69. Hirosumi J, Tuncman G, Chang L, Gorgun CZ, Uysal KT, Maeda K, et al. A central role for JNK in obesity and insulin resistance. Nature (2002) 420:333–6. doi:10.1038/nature01137
70. Saltiel AR, Pessin JE. Insulin signaling pathways in time and space. Trends Cell Biol (2002) 12:65–71. doi:10.1016/S0962-8924(01)02207-3
71. Schinner S, Scherbaum WA, Bornstein SR, Barthel A. Molecular mechanisms of insulin resistance. Diabet Med (2005) 22:674–82. doi:10.1111/j.1464-5491.2005.01566.x
72. Jager J, Gremeaux T, Cormont M, Le Marchand-Brustel Y, Tanti JF. Interleukin-1beta-induced insulin resistance in adipocytes through down-regulation of insulin receptor substrate-1 expression. Endocrinology (2007) 148:241–51. doi:10.1210/en.2006-0692
73. Tuncman G, Hirosumi J, Solinas G, Chang L, Karin M, Hotamisligil GS. Functional in vivo interactions between JNK1 and JNK2 isoforms in obesity and insulin resistance. Proc Natl Acad Sci U S A (2006) 103:10741–6. doi:10.1073/pnas.0603509103
74. Solinas G, Vilcu C, Neels JG, Bandyopadhyay GK, Luo JL, Naugler W, et al. JNK1 in hematopoietically derived cells contributes to diet-induced inflammation and insulin resistance without affecting obesity. Cell Metab (2007) 6:386–97. doi:10.1016/j.cmet.2007.09.011
75. Vallerie SN, Furuhashi M, Fucho R, Hotamisligil GS. A predominant role for parenchymal c-Jun amino terminal kinase (JNK) in the regulation of systemic insulin sensitivity. PLoS One (2008) 3:e3151. doi:10.1371/journal.pone.0003151
76. Cani PD, Delzenne NM. The role of the gut microbiota in energy metabolism and metabolic disease. Curr Pharm Des (2009) 15:1546–58. doi:10.2174/138161209788168164
77. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on toll-like receptors. Nat Immunol (2010) 11:373–84. doi:10.1038/ni.1863
78. Burcelin R, Serino M, Chabo C, Blasco-Baque V, Amar J. Gut microbiota and diabetes: from pathogenesis to therapeutic perspective. Acta Diabetol (2011) 48:257–73. doi:10.1007/s00592-011-0333-6
79. Jialal I, Kaur H. The role of toll-like receptors in diabetes-induced inflammation: implications for vascular complications. Curr Diab Rep (2012) 12:172–9. doi:10.1007/s11892-012-0258-7
80. Norseen J, Hosooka T, Hammarstedt A, Yore MM, Kant S, Aryal P, et al. Retinol-binding protein 4 inhibits insulin signaling in adipocytes by inducing proinflammatory cytokines in macrophages through a c-Jun N-terminal kinase- and toll-like receptor 4-dependent and retinol-independent mechanism. Mol Cell Biol (2012) 32:2010–9. doi:10.1128/MCB.06193-11
81. Toshchakov V, Jones BW, Perera PY, Thomas K, Cody MJ, Zhang S, et al. TLR4, but not TLR2, mediates IFN-beta-induced STAT1alpha/beta-dependent gene expression in macrophages. Nat Immunol (2002) 3:392–8. doi:10.1038/ni774
82. Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med (2011) 17:179–88. doi:10.1038/nm.2279
83. Shi H, Kokoeva MV, Inouye K, Tzameli I, Yin H, Flier JS. TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest (2006) 116:3015–25. doi:10.1172/JCI28898
84. Nanwang H, Liang H, Zen K. Molecular mechanisms that influence the macrophage M1-M2 polarization balance. Front Immunol (2014) 5:614. doi:10.3389/fimmu.2014.00614
85. Tsukumo DM, Carvalho-Filho MA, Carvalheira JB, Prada PO, Hirabara SM, Schenka AA, et al. Loss-of-function mutation in toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Diabetes (2007) 56:1986–98. doi:10.2337/db06-1595
86. Fresno M, Alvarez R, Cuesta N. Toll-like receptors, inflammation, metabolism and obesity. Arch Physiol Biochem (2011) 117:151–64. doi:10.3109/13813455.2011.562514
87. Davis JE, Gabler NK, Walker-Daniels J, Spurlock ME. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Obesity (Silver Spring) (2008) 16:1248–55. doi:10.1038/oby.2008.210
88. Watanabe Y, Nakamura T, Ishikawa S, Fujisaka S, Usui I, Tsuneyama K, et al. The radioprotective 105/MD-1 complex contributes to diet-induced obesity and adipose tissue inflammation. Diabetes (2012) 61:1199–209. doi:10.2337/db11-1182
89. Wen H, Ting JP, O’neill LA. A role for the NLRP3 inflammasome in metabolic diseases – did Warburg miss inflammation? Nat Immunol (2012) 13:352–7. doi:10.1038/ni.2228
90. Stienstra R, Joosten LA, Koenen T, Van Tits B, Van Diepen JA, Van Den Berg SA, et al. The inflammasome-mediated caspase-1 activation controls adipocyte differentiation and insulin sensitivity. Cell Metab (2010) 12:593–605. doi:10.1016/j.cmet.2010.11.011
91. Wen H, Gris D, Lei Y, Jha S, Zhang L, Huang MT, et al. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat Immunol (2011) 12:408–15. doi:10.1038/ni.2022
92. Zhou R, Tardivel A, Thorens B, Choi I, Tschopp J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol (2010) 11:136–40. doi:10.1038/ni.1831
93. Howell JJ, Manning BD. mTOR couples cellular nutrient sensing to organismal metabolic homeostasis. Trends Endocrinol Metab (2011) 22:94–102. doi:10.1016/j.tem.2010.12.003
94. Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell (2012) 149:274–93. doi:10.1016/j.cell.2012.03.017
95. Inoki K, Li Y, Zhu T, Wu J, Guan KL. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat Cell Biol (2002) 4:648–57. doi:10.1038/ncb839
96. Manning BD, Tee AR, Logsdon MN, Blenis J, Cantley LC. Identification of the tuberous sclerosis complex-2 tumor suppressor gene product tuberin as a target of the phosphoinositide 3-kinase/akt pathway. Mol Cell (2002) 10:151–62. doi:10.1016/S1097-2765(02)00568-3
97. Becattini B, Marone R, Zani F, Arsenijevic D, Seydoux J, Montani JP, et al. PI3Kgamma within a nonhematopoietic cell type negatively regulates diet-induced thermogenesis and promotes obesity and insulin resistance. Proc Natl Acad Sci U S A (2011) 108:E854–63. doi:10.1073/pnas.1106698108
98. Kobayashi N, Ueki K, Okazaki Y, Iwane A, Kubota N, Ohsugi M, et al. Blockade of class IB phosphoinositide-3 kinase ameliorates obesity-induced inflammation and insulin resistance. Proc Natl Acad Sci U S A (2011) 108:5753–8. doi:10.1073/pnas.1016430108
99. Jiang H, Westerterp M, Wang C, Zhu Y, Ai D. Macrophage mTORC1 disruption reduces inflammation and insulin resistance in obese mice. Diabetologia (2014) 57:2393–404. doi:10.1007/s00125-014-3350-5
100. Moon JS, Hisata S, Park MA, Denicola GM, Ryter SW, Nakahira K, et al. mTORC1-Induced HK1-dependent glycolysis regulates nlrp3 inflammasome activation. Cell Rep (2015) 12:102–15. doi:10.1016/j.celrep.2015.05.046
101. Byles V, Covarrubias AJ, Ben-Sahra I, Lamming DW, Sabatini DM, Manning BD, et al. The TSC-mTOR pathway regulates macrophage polarization. Nat Commun (2013) 4:2834. doi:10.1038/ncomms3834
102. Zhu L, Yang T, Li L, Sun L, Hou Y, Hu X, et al. TSC1 controls macrophage polarization to prevent inflammatory disease. Nat Commun (2014) 5:4696. doi:10.1038/ncomms5696
103. Fabbrini E, Cella M, McCartney SA, Fuchs A, Abumrad NA, Pietka TA, et al. Association between specific adipose tissue CD4+ T-cell populations and insulin resistance in obese individuals. Gastroenterology (2013) 145:.e361–3. doi:10.1053/j.gastro.2013.04.010
104. Stolarczyk E, Vong CT, Perucha E, Jackson I, Cawthorne MA, Wargent ET, et al. Improved insulin sensitivity despite increased visceral adiposity in mice deficient for the immune cell transcription factor T-bet. Cell Metab (2013) 17:520–33. doi:10.1016/j.cmet.2013.02.019
105. Dalmas E, Venteclef N, Caer C, Poitou C, Cremer I, Aron-Wisnewsky J, et al. T cell-derived IL-22 amplifies IL-1beta-driven inflammation in human adipose tissue: relevance to obesity and type 2 diabetes. Diabetes (2014) 63:1966–77. doi:10.2337/db13-1511
106. Khan IM, Dai Perrard XY, Perrard JL, Mansoori A, Smith CW, Wu H, et al. Attenuated adipose tissue and skeletal muscle inflammation in obese mice with combined CD4+ and CD8+ T cell deficiency. Atherosclerosis (2014) 233:419–28. doi:10.1016/j.atherosclerosis.2014.01.011
107. Delgoffe GM, Pollizzi KN, Waickman AT, Heikamp E, Meyers DJ, Horton MR, et al. The kinase mTOR regulates the differentiation of helper T cells through the selective activation of signaling by mTORC1 and mTORC2. Nat Immunol (2011) 12:295–303. doi:10.1038/ni.2005
108. Moraes-Vieira PM, Bassi EJ, Larocca RA, Castoldi A, Burghos M, Lepique AP, et al. Leptin deficiency modulates allograft survival by favoring a Th2 and a regulatory immune profile. [corrected]. Am J Transplant (2013) 13:36–44. doi:10.1111/j.1600-6143.2012.04283.x
109. Hill AA, Reid Bolus W, Hasty AH. A decade of progress in adipose tissue macrophage biology. Immunol Rev (2014) 262:134–52. doi:10.1111/imr.12216
110. Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol (2013) 229:176–85. doi:10.1002/path.4133
111. Fuentes L, Roszer T, Ricote M. Inflammatory mediators and insulin resistance in obesity: role of nuclear receptor signaling in macrophages. Mediators Inflamm (2010) 2010:219583. doi:10.1155/2010/219583
112. Bohlson SS, O’conner SD, Hulsebus HJ, Ho MM, Fraser DA. Complement, c1q, and c1q-related molecules regulate macrophage polarization. Front Immunol (2014) 5:402. doi:10.3389/fimmu.2014.00402
113. Lee CG, Homer RJ, Zhu Z, Lanone S, Wang X, Koteliansky V, et al. Interleukin-13 induces tissue fibrosis by selectively stimulating and activating transforming growth factor beta(1). J Exp Med (2001) 194:809–21. doi:10.1084/jem.194.6.809
114. Sierra-Filardi E, Vega MA, Sanchez-Mateos P, Corbi AL, Puig-Kroger A. Heme oxygenase-1 expression in M-CSF-polarized M2 macrophages contributes to LPS-induced IL-10 release. Immunobiology (2010) 215:788–95. doi:10.1016/j.imbio.2010.05.020
115. Spencer M, Yao-Borengasser A, Unal R, Rasouli N, Gurley CM, Zhu B, et al. Adipose tissue macrophages in insulin-resistant subjects are associated with collagen VI and fibrosis and demonstrate alternative activation. Am J Physiol Endocrinol Metab (2010) 299:E1016–27. doi:10.1152/ajpendo.00329.2010
116. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol (2008) 8:958–69. doi:10.1038/nri2448
117. Zizzo G, Cohen PL. The PPAR-gamma antagonist GW9662 elicits differentiation of M2c-like cells and upregulation of the MerTK/Gas6 axis: a key role for PPAR-gamma in human macrophage polarization. J Inflamm (Lond) (2015) 12:36. doi:10.1186/s12950-015-0081-4
118. Lumeng CN, Delproposto JB, Westcott DJ, Saltiel AR. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes (2008) 57:3239–46. doi:10.2337/db08-0872
119. Wu D, Molofsky AB, Liang HE, Ricardo-Gonzalez RR, Jouihan HA, Bando JK, et al. Eosinophils sustain adipose alternatively activated macrophages associated with glucose homeostasis. Science (2011) 332:243–7. doi:10.1126/science.1201475
120. Qiu Y, Nguyen KD, Odegaard JI, Cui X, Tian X, Locksley RM, et al. Eosinophils and type 2 cytokine signaling in macrophages orchestrate development of functional beige fat. Cell (2014) 157:1292–308. doi:10.1016/j.cell.2014.03.066
121. Vieira-Potter VJ. Inflammation and macrophage modulation in adipose tissues. Cell Microbiol (2014) 16:1484–92. doi:10.1111/cmi.12336
122. Feuerer M, Herrero L, Cipolletta D, Naaz A, Wong J, Nayer A, et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat Med (2009) 15:930–9. doi:10.1038/nm.2002
123. Cipolletta D, Kolodin D, Benoist C, Mathis D. Tissular T(regs): a unique population of adipose-tissue-resident Foxp3+CD4+ T cells that impacts organismal metabolism. Semin Immunol (2011) 23:431–7. doi:10.1016/j.smim.2011.06.002
124. Liu G, Ma H, Qiu L, Li L, Cao Y, Ma J, et al. Phenotypic and functional switch of macrophages induced by regulatory CD4+CD25+ T cells in mice. Immunol Cell Biol (2011) 89:130–42. doi:10.1038/icb.2010.70
125. Spits H, Cupedo T. Innate lymphoid cells: emerging insights in development, lineage relationships, and function. Annu Rev Immunol (2012) 30:647–75. doi:10.1146/annurev-immunol-020711-075053
126. Kim BS, Wojno ED, Artis D. Innate lymphoid cells and allergic inflammation. Curr Opin Immunol (2013) 25:738–44. doi:10.1016/j.coi.2013.07.013
127. Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, et al. Innate lymphoid cells – a proposal for uniform nomenclature. Nat Rev Immunol (2013) 13:145–9. doi:10.1038/nri3365
128. Kim BS, Artis D. Group 2 innate lymphoid cells in health and disease. Cold Spring Harb Perspect Biol (2015) 7:a016337. doi:10.1101/cshperspect.a016337
129. Moro K, Yamada T, Tanabe M, Takeuchi T, Ikawa T, Kawamoto H, et al. Innate production of T(H)2 cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature (2010) 463:540–4. doi:10.1038/nature08636
130. Moussion C, Ortega N, Girard JP. The IL-1-like cytokine IL-33 is constitutively expressed in the nucleus of endothelial cells and epithelial cells in vivo: a novel ‘alarmin’? PLoS One (2008) 3:e3331. doi:10.1371/journal.pone.0003331
131. Pichery M, Mirey E, Mercier P, Lefrancais E, Dujardin A, Ortega N, et al. Endogenous IL-33 is highly expressed in mouse epithelial barrier tissues, lymphoid organs, brain, embryos, and inflamed tissues: in situ analysis using a novel Il-33-LacZ gene trap reporter strain. J Immunol (2012) 188:3488–95. doi:10.4049/jimmunol.1101977
132. Cayrol C, Girard JP. IL-33: an alarmin cytokine with crucial roles in innate immunity, inflammation and allergy. Curr Opin Immunol (2014) 31:31–7. doi:10.1016/j.coi.2014.09.004
133. Han JM, Wu D, Denroche HC, Yao Y, Verchere CB, Levings MK. IL-33 reverses an obesity-induced deficit in visceral adipose tissue ST2+ T regulatory cells and ameliorates adipose tissue inflammation and insulin resistance. J Immunol (2015) 194:4777–83. doi:10.4049/jimmunol.1500020
134. Miller AM, Asquith DL, Hueber AJ, Anderson LA, Holmes WM, McKenzie AN, et al. Interleukin-33 induces protective effects in adipose tissue inflammation during obesity in mice. Circ Res (2010) 107:650–8. doi:10.1161/CIRCRESAHA.110.218867
135. Brestoff JR, Kim BS, Saenz SA, Stine RR, Monticelli LA, Sonnenberg GF, et al. Group 2 innate lymphoid cells promote beiging of white adipose tissue and limit obesity. Nature (2015) 519:242–6. doi:10.1038/nature14115
136. Harms M, Seale P. Brown and beige fat: development, function and therapeutic potential. Nat Med (2013) 19:1252–63. doi:10.1038/nm.3361
137. Cohen P, Levy DJ, Zhang Y, Frontini A, Kolodin PD, Svensson KJ, et al. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell (2014) 156:304–16. doi:10.1016/j.cell.2013.12.021
138. Lee MW, Odegaard JI, Mukundan L, Qiu Y, Molofsky AB, Nussbaum JC, et al. Activated type 2 innate lymphoid cells regulate beige fat biogenesis. Cell (2015) 160:74–87. doi:10.1016/j.cell.2014.12.011
139. Ganeshan K, Chawla A. Metabolic regulation of immune responses. Annu Rev Immunol (2014) 32:609–34. doi:10.1146/annurev-immunol-032713-120236
140. Yore MM, Syed I, Moraes-Vieira PM, Zhang T, Herman MA, Homan EA, et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell (2014) 159:318–32. doi:10.1016/j.cell.2014.09.035
141. Orr JS, Kennedy A, Anderson-Baucum EK, Webb CD, Fordahl SC, Erikson KM, et al. Obesity alters adipose tissue macrophage iron content and tissue iron distribution. Diabetes (2014) 63:421–32. doi:10.2337/db13-0213
142. Vats D, Mukundan L, Odegaard JI, Zhang L, Smith KL, Morel CR, et al. Oxidative metabolism and PGC-1beta attenuate macrophage-mediated inflammation. Cell Metab (2006) 4:13–24. doi:10.1016/j.cmet.2006.05.011
143. Nizet V, Johnson RS. Interdependence of hypoxic and innate immune responses. Nat Rev Immunol (2009) 9:609–17. doi:10.1038/nri2607
144. Galvan-Pena S, O’neill LA. Metabolic reprograming in macrophage polarization. Front Immunol (2014) 5:420. doi:10.3389/fimmu.2014.00420
145. Freemerman AJ, Johnson AR, Sacks GN, Milner JJ, Kirk EL, Troester MA, et al. Metabolic reprogramming of macrophages: glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. J Biol Chem (2014) 289:7884–96. doi:10.1074/jbc.M113.522037
146. Fujisaka S, Usui I, Ikutani M, Aminuddin A, Takikawa A, Tsuneyama K, et al. Adipose tissue hypoxia induces inflammatory M1 polarity of macrophages in an HIF-1alpha-dependent and HIF-1alpha-independent manner in obese mice. Diabetologia (2013) 56:1403–12. doi:10.1007/s00125-013-2885-1
147. Kim SY, Jeong E, Joung SM, Lee JY. PI3K/Akt contributes to increased expression of toll-like receptor 4 in macrophages exposed to hypoxic stress. Biochem Biophys Res Commun (2012) 419:466–71. doi:10.1016/j.bbrc.2012.02.015
148. Kim SY, Choi YJ, Joung SM, Lee BH, Jung YS, Lee JY. Hypoxic stress up-regulates the expression of toll-like receptor 4 in macrophages via hypoxia-inducible factor. Immunology (2010) 129:516–24. doi:10.1111/j.1365-2567.2009.03203.x
149. Rausch ME, Weisberg S, Vardhana P, Tortoriello DV. Obesity in C57BL/6J mice is characterized by adipose tissue hypoxia and cytotoxic T-cell infiltration. Int J Obes (Lond) (2008) 32:451–63. doi:10.1038/sj.ijo.0803744
150. Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, et al. Succinate is an inflammatory signal that induces IL-1beta through HIF-1alpha. Nature (2013) 496:238–42. doi:10.1038/nature11986
151. Palsson-McDermott EM, Curtis AM, Goel G, Lauterbach MA, Sheedy FJ, Gleeson LE, et al. Pyruvate kinase M2 regulates Hif-1alpha activity and IL-1beta induction and is a critical determinant of the warburg effect in LPS-activated macrophages. Cell Metab (2015) 21:65–80. doi:10.1016/j.cmet.2014.12.005
152. Rodriguez-Prados JC, Traves PG, Cuenca J, Rico D, Aragones J, Martin-Sanz P, et al. Substrate fate in activated macrophages: a comparison between innate, classic, and alternative activation. J Immunol (2010) 185:605–14. doi:10.4049/jimmunol.0901698
153. Blagih J, Jones RG. Polarizing macrophages through reprogramming of glucose metabolism. Cell Metab (2012) 15:793–5. doi:10.1016/j.cmet.2012.05.008
154. Haschemi A, Kosma P, Gille L, Evans CR, Burant CF, Starkl P, et al. The sedoheptulose kinase CARKL directs macrophage polarization through control of glucose metabolism. Cell Metab (2012) 15:813–26. doi:10.1016/j.cmet.2012.04.023
155. Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, Morel CR, Subramanian V, Mukundan L, et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature (2007) 447:1116–20. doi:10.1038/nature05894
156. Kang K, Reilly SM, Karabacak V, Gangl MR, Fitzgerald K, Hatano B, et al. Adipocyte-derived Th2 cytokines and myeloid PPARdelta regulate macrophage polarization and insulin sensitivity. Cell Metab (2008) 7:485–95. doi:10.1016/j.cmet.2008.04.002
Keywords: obesity, adipose tissue, insulin resistance, macrophage, adipokines, macrophage polarization, adipose tissue inflammation
Citation: Castoldi A, Naffah de Souza C, Câmara NO and Moraes-Vieira PM (2016) The Macrophage Switch in Obesity Development. Front. Immunol. 6:637. doi: 10.3389/fimmu.2015.00637
Received: 28 September 2015; Accepted: 04 December 2015;
Published: 05 January 2016
Edited by:
Paige Lacy, University of Alberta, CanadaReviewed by:
Giovanni Solinas, University of Gothenburg, SwedenCopyright: © 2016 Castoldi, Naffah de Souza, Câmara and Moraes-Vieira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro M. Moraes-Vieira, cHZpZWlyYUBiaWRtYy5oYXJ2YXJkLmVkdQ==
†Angela Castoldi and Cristiane Naffah de Souza contributed equally to this review.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.