
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 22 September 2015
Sec. Molecular Innate Immunity
Volume 6 - 2015 | https://doi.org/10.3389/fimmu.2015.00480
This article is part of the Research Topic Macrophages Role in Integrating Tissue Signals and Biological Processes in Chronic Inflammation and Fibrosis View all 11 articles
 Henrique Borges da Silva*†‡
Henrique Borges da Silva*†‡ Raíssa Fonseca‡
Raíssa Fonseca‡ Rosana Moreira Pereira
Rosana Moreira Pereira Alexandra dos Anjos Cassado
Alexandra dos Anjos Cassado José Maria Álvarez
José Maria Álvarez Maria Regina D’Império Lima*
Maria Regina D’Império Lima*
The spleen is one of the major immunological sites for maintaining blood homeostasis. Previous studies showed that heterogeneous splenic macrophage populations contribute in complimentary ways to control blood-borne infections and induce effective immune responses. Marginal metallophilic macrophages (MMMΦs) and marginal zone macrophages (MZMΦs) are cells with great ability to internalize blood-borne pathogens such as virus or bacteria. Their localization adjacent to T- and B-cell-rich splenic areas favors the rapid contact between these macrophages and cells from adaptive immunity. Indeed, MMMΦs and MZMΦs are considered important bridges between innate and adaptive immunity. Although red pulp macrophages (RpMΦs) are mainly considered scavengers for senescent erythrocytes, several data indicate a role for RpMΦs in control of infections such as blood-stage malaria as well as in the induction of innate and adaptive immunity. Here, we review current data on how different macrophage subsets recognize and help eliminate blood-borne pathogens, and, in turn, how the inflammatory microenvironment in different phases of infection (acute, chronic, and after pathogen clearance) influences macrophage function and survival.
Effective control of infections through the immune system is ensured by the well-organized structure of secondary lymphoid organs, which allow capture, processing, and presentation of antigens, ultimately leading to successful elimination of pathogens and induction of adaptive immunity. Among lymphoid organs, the spleen is particularly shaped for clearance of blood-borne pathogens. Microanatomically, the spleen is divided into the white pulp and the red pulp (Rp), separated by the marginal zone (MZ) [reviewed in Ref. (1)]. Rp and MZ have a complex macrophage (MΦ) network with distinct origins and functions in the immune response to infections. RpMΦs form a vast network inside the Rp and are characterized in mice by expression of F4/80highCD68+CD11blow/− and intense autofluorescence (2). In turn, inside the MZ, two populations of MΦs can be discerned. The MZMΦs typically express in their surface the C-type lectin SIGN-related 1 (SIGNR1) and a type I scavenger receptor called Macrophage Receptor with Collagenous structure (MARCO), which recognize non-opsonized molecules (3), mainly blood-borne antigens (4). Furthermore, marginal metallophilic MΦs (MMMΦs) are defined, among other molecules, by the expression of Sialic acid-binding Ig-like Lectin-1 (Siglec-1, Sialoadhesin, CD169) and MOMA-1 (5). A general scheme of the spleen structure is depicted in Figure 1.
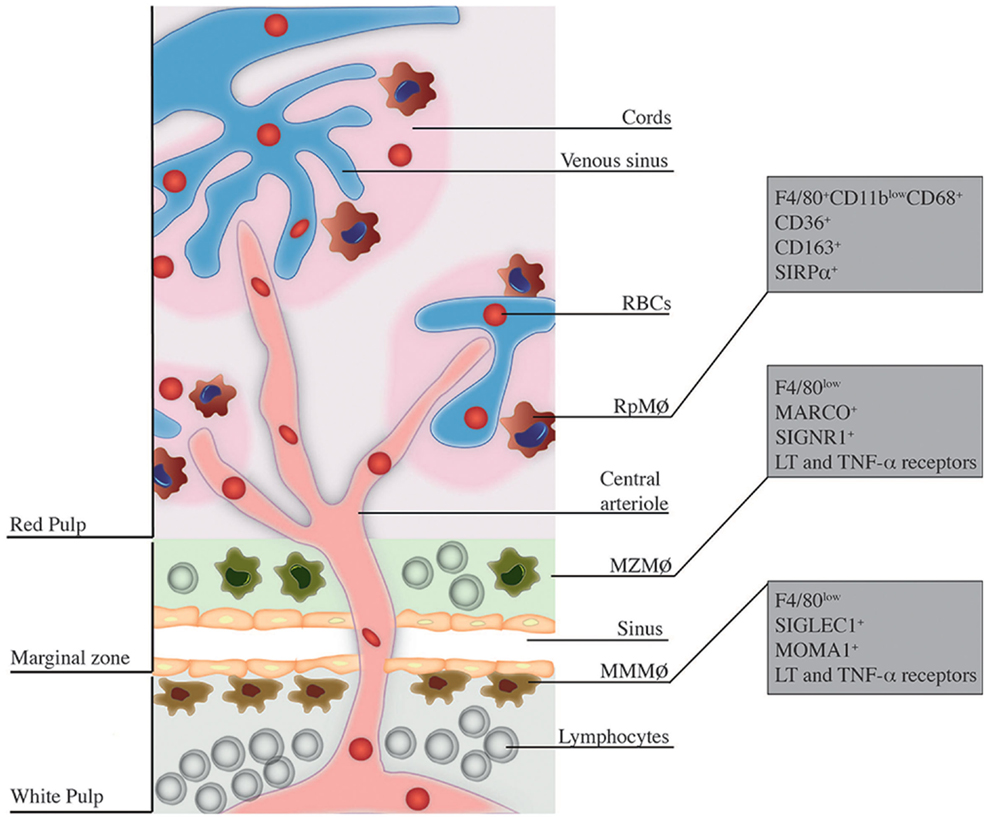
Figure 1. Localization and phenotype of splenic MΦ subsets. This figure is a broad scheme of the positioning of RpMΦs, MZMΦs, and MMMΦs inside spleen and their respective phenotypic markers. RpMΦs (in red) are typically found within cords on the red pulp, allowing direct contact with RBCs and other blood cells/particles passing through venous sinuses. They are better defined by the concomitant expression of F4/80, CD11b (at low levels), and CD68 as well as other receptors that aid in their function. MZMΦs (in green) are found in the marginal zone (MZ) outer layer – they are also in direct contact with blood-borne particles. These cells express in their surface the molecules MARCO and SIGNR1 and other receptors that help in the uptake of blood-borne pathogens. Finally, the MMMΦs (in brown) reside within the inner layer of the MZ, in the contact with the white pulp. They are also specialized in blood-borne particle uptake and express surface markers such as SIGLEC-1 and MOMA-1.
Recent studies led to a growing understanding of the precise roles different splenic MΦs play to maintain blood homeostasis, particularly in infectious diseases, in which pathogen elimination depends on the development of appropriate adaptive immune response. In this review, we addressed the roles of each one of these MΦ subsets, with special focus on blood-borne infections. We described the current knowledge on the effects of splenic microarchitecture and microenvironment on these MΦs and reciprocal influence of these cells on spleen structure and functionality.
Splenic MΦs have two main protective activities during blood-borne infections. The first and most well characterized is phagocytosis and elimination of pathogens from circulation. However, beyond the task of eliminating blood-borne pathogens, splenic MΦs can play a prominent role in immune system activation. To properly execute these functions, they are provided with a large variety of pattern-recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). Engagement of Toll-like receptor (TLR) 4 by pathogen molecules, such as lipopolysaccharides (LPS) from Gram-negative bacteria is fundamental for the induction of a proinflammatory gene and protein expression signature in MΦs, which ultimately leads to innate immune activation (6). This also holds true for several other interactions such as TLR2 and/or TLR4 with glycosylphosphatidylinositol (GPI) anchors from Trypanosoma and Plasmodium parasites (7, 8) and TLR9 engagement by CpG motifs found in bacterial (9) and plasmodial DNA (10).
On the other hand, TLRs recognize DAMPs in situations of tissue injury. For example, heat shock proteins (HSPs) are endogenous damage signals (molecules released by cells under stress or necrotic cell death) and bind to TLR2 and TLR4 in MΦs, inducing these cells to produce proinflammatory cytokines and to express costimulatory molecules (11). Release of HSPs to circulation has been reported during sepsis (12) as well as production of HSP homologues by pathogens such as Plasmodium parasites (13). Also, TLRs – especially TLR2 and TLR4 – can recognize extracellular matrix components such as fibronectin (14). TLR4 engagement by fibronectin leads to MΦ activation in a similar fashion to what happens after LPS stimulation. Fibronectin is presumably secreted by fibroblasts inside the spleen. Thus, this molecule may be produced during blood-borne infections such as malaria, where profound changes in splenic microarchitecture following acute infection occur, leading to the accumulation of fibroblasts inside the Rp (15). Expression of fibronectin-binding proteins (FnBPs) by Staphylococcus aureus is important to bacterial uptake by MΦs in inflammatory situations through binding of very late antigen 5 (VLA-5) (16). Therefore, it is reasonable to question whether TLR2 and/or TLR4 expressed in MΦs are engaged by fibronectin in those situations. Importantly, S. aureus FnBPs are crucial for the development of sepsis (16).
Another DAMP that can induce MΦ activation is the high mobility group box protein 1 (HMGB1), an intracellular DNA-binding protein involved in chromatin remodeling and transcription regulation (17). Extracellular HMGB1 binds to different endogenous ligands that are recognized by receptors such as TLR4, as well as the receptor for advanced glycation end products (RAGE) (18), and triggers inflammatory responses by the innate immune system. Release of HMGB1 by splenic MΦs occurs upon extensive splenic cell apoptosis, a feature commonly observed during sepsis. Indeed, HMGB1 is released into the extracellular milieu during sepsis and neutralization of this protein by monoclonal antibody treatment blocks sepsis development (19). Abundant splenic cell apoptosis is also typical in rodent malaria, at the peak of acute infection (20). In human malaria, endogenous HMGB1 serum levels are significantly higher in patients with severe disease compared to uncomplicated cases (21), suggesting that HMGB1 might also be involved in the development of immunopathology. Thus, it would not be surprising if acute immune response to Plasmodium and consequent immunopathology could be suppressed in great extent with neutralization of HMGB1.
Splenic MΦ receptors also include C-type lectin receptors (CLRs), such as dectin-1, mannose receptor, and dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN). CLRs have multiple functions in the immune system, including pathogen recognition and neutralization (22). Additionally, the liver synthesizes mannose-binding protein (MBP) during infectious diseases. This protein activates the complement system in order to form the membrane attack complex (MAC), and, more importantly in the spleen, to opsonize microorganisms such as virus (23) or protozoan parasites such as Trypanosoma cruzi (24). Scavenger receptors (SRs), such as SR-A1 and MARCO, are also expressed in splenic MΦs and likewise bind both self and pathogen molecules – more specificities of these receptors will be discussed later in this review. Among class B SRs, CD36 is known to mediate the uptake of oxidized low-density lipoprotein (oxLDL) and apoptotic cells, but also promotes phagocytosis of S. aureus bacteria by peritoneal MΦs (25). However, CD36 mediates cytoadherence of Plasmodium-infected red blood cells (iRBCs) to microvascular endothelium (26), a process supposed to avoid parasite clearance inside the spleen. The role of CD36 in recognizing this parasite by splenic MΦs still needs to be fully elucidated. Of note, RpMΦs express constitutively this molecule, which implies a possible role for this receptor in antiplasmodial immunity. This is a clear example of a receptor capable of mediating the recognition of both self and non-self molecules, implicating RpMΦs with both blood homeostasis and control of blood-borne infections.
Among cytoplasmic PRRs, splenic MΦs express molecules from the NOD-like receptor (NLR) family (27). For example, disturbance of cellular ionic gradient activates the pyrin subfamily member NLRP3, leading to inflammasome complex formation and in consequence to the release of inflammatory cytokines IL-1β and IL-18. Hemozoin, a disposal product formed from hemoglobin digestion by Plasmodium parasites, triggers the NLRP3 inflammasome in bone marrow-derived macrophages (BMDMs) (28), mediating the production of proinflammatory cytokines by these cells. Furthermore, the NLRP3 inflammasome is activated in mouse RpMΦs and human peripheral monocytes during acute malaria – although large amounts of IL-1β are only produced after stimulation with LPS (29). Interestingly, in mice, this process is mediated by the purinergic P2 × 7 receptor which recognizes extracellular ATP. ATP accumulates in Plasmodium-iRBCs and is released into the extracellular milieu through ion channels in the erythrocyte membrane or upon iRBC rupture (30).
As stated previously, RpMΦs form a vast network inside the Rp, and although there is no consensus about the origin of RpMΦs, recent data indicate that these MΦs are maintained by local proliferation during physiological conditions (31). Conversely, in some pathological conditions, circulating monocytes are able to differentiate into RpMΦs (32). RpMΦ population comprises macrophage colony-stimulating factor (M-CSF)-dependent and M-CSF-independent cells (33). M-CSF-dependent RpMΦs are efficient phagocytes and produce proinflammatory cytokines such as TNF-α and type I IFNs and are highly responsive to prostaglandin E2 (PGE2). In contrast, M-CSF-independent BMDMs are less efficient phagocytes that produce high amounts of PGE2 (34). If this is a general feature of M-CSF-independent MΦ populations, M-CSF-independent RpMΦs might influence the activity of M-CSF-dependent RpMΦs.
Venous cords and sinuses render the splenic Rp bloodstream in a slow pace. This characteristic allows for the filtering function of the spleen and favors elimination of aberrant red blood cells (RBCs) or Plasmodium-iRBCs (35). Of note, development of RpMΦs relies on the expression of the transcription factor Spi-C, which is induced by free heme from RBC degradation (32). Thus, iron homeostasis – which conversely is controlled by RpMΦs – might play a role in RpMΦ development. Splenic structure also facilitates the control of numerous blood-borne infections by RpMΦs. For example, RpMΦs can recognize the capsular polysaccharide glucuronoxylomannan (GXM) from Cryptococcus neoformans and subsequently phagocytize the bacteria (36). RpMΦs can also eliminate Streptococcus pneumoniae under conditions of splenomegaly (37). However, these MΦs are permissive to intracellular growth of Salmonella typhimurium (38).
Red pulp macrophages have also been implicated in the control of blood-stage malaria (35). Nevertheless, in experimental Plasmodium yoelii infection, spleen remodeling facilitates iRBC adherence to the vascular endothelium and, in consequence, allows parasites to escape from phagocytes (15). Interestingly, a proportion of Rp phagocytes exhibit strong labeling for F4/80 and CD11c, a phenotype shared by RpMΦs and DCs (39). This population participates in the early clearance of Plasmodium chabaudi parasites, but it sharply declines at the parasitemia peak. RpMΦs have a slow turnover rate and possibly undergo cell death after ingesting Plasmodium-iRBCs due to the toxic effects of hemozoin. RpMΦs, which are primarily required to maintain tissue homeostasis, might be substituted by inflammatory phagocytes as well as by MΦs derived from inflammatory monocytes. An alternative explanation is downregulation of the F4/80 molecule upon MΦ activation, as reported during mycobacterial infection (40).
Several mechanisms mediate RBC recognition and clearance by RpMΦs. One of the most studied mechanisms is the antibody binding to altered self components such as Band 3 clusters (41) or phosphatidylserine residues exposed in the outer leaflet of RBC membrane (42). In these cases, natural antibodies and complement system proteins opsonize RBCs though recognition of Band 3 clusters or phosphatidylserine residues. Another important interaction involved in RBC phagocytosis by RpMΦs is the ligation of CD47 to Signal Regulatory Protein alpha (SIRPα) (43). CD47 is a self-molecule important to avoid clearance by phagocytes, which is ubiquitously expressed on many cell types, including RBCs. CD47 expression on RBCs is an inhibitory signal for phagocytosis (44), but RBCs expressing a modified isoform of this molecule are phagocytized by RpMΦs through SIRPα binding (43). Interestingly, the conformation-dependent anti-CD47 antibody 2D3 binds sickle RBCs preferentially (45), which might explain the enhanced phagocytosis of sickle RBCs inside spleen. A recent study showed that P. yoelii parasites preferentially infect young RBCs expressing high levels of CD47 and, in consequence, escape from splenic clearance (46). Furthermore, enhanced resistance to P. yoelii observed in CD47-deficient mice is associated with a larger population of RpMΦs that ingest more iRBCs than wild-type counterparts. These findings explain why individuals with mild genetic RBC disorders (e.g., sickle cell trait and glucose-6-phosphate dehydrogenase deficiency) are protected from lethal malaria due to enhanced RBC phagocytosis.
Apart from being phagocytized by splenic MΦs, Plasmodium-iRBCs are also destroyed intravascularly as a consequence of plasma membrane damage upon release of free merozoites. Hemozoin, a disposal product formed from hemoglobin digestion by parasites, is released from lysed iRBCs. Furthermore, a massive destruction of non-infected RBCs occurs during blood-stage malaria, leading to increased hemoglobin levels in circulation [reviewed in Ref. (47)]. Another example of hemolysis induced by infections is observed in septicemia caused by Escherichia coli, which produces exotoxin α-hemolysin (Hlyα) (48). Evidencing RpMΦs crucial role in neutralizing toxic effects of hemoglobin, these MΦs have high levels of intracellular heme due to RBC phagocytosis (2) and of free hemoglobin through the scavenger receptor CD163 (49). The enzyme heme-oxygenase 1 (HO-1) plays an important role in degrading free heme, which in excess causes toxicity to MΦs (50). Importantly, RpMΦs are able to control pathogen burden through control of iron availability. For example, RpMΦs express the natural resistance associated macrophage protein-1 (NRAMP1) that is associated with protection against intraphagosomal pathogens, such as Mycobacterium bovis BCG, Leishmania donovani, or S. typhimurium. This molecule is a pH-dependent metal transporter localized in phagosomal compartments, which reduces intraphagosomal iron levels derived from RBC phagocytosis (51). NRAMP1 synthesis is upregulated in IFN-γ-activated MΦs (52), a condition likely to occur during acute blood-borne infections. RpMΦs also limit pathogen iron uptake through TLR-mediated release of lipocalin-2, which can form complexes with pathogen-secreted siderophores – molecules that help the collection of iron available for pathogens (53). RpMΦs involvement in controlling excessive immune responses is suggested by studies on autoimmune syndromes, while a similar participation in infectious diseases remains to be established. For instance, RpMΦs constitutively express peroxisome proliferator-activated receptor-γ (PPAR-γ), which might be important to curb excessive immune responses to pathogens, in a similar manner to PPAR-γ expressed on lung MΦs upon S. pneumoniae infection (54). RPMΦs can also prevent autoimmunity by producing anti-inflammatory cytokines such as TGF-β and IL-10 and by inducing generation of regulatory T (Treg) cells (55). Of note, there are many T cells scattered in Rp (55), and this population participates in acute immune responses to infections, such as blood-stage malaria (39). We present an illustrated scheme of the different roles of RpMΦs in homeostasis and disease in Figure 2.
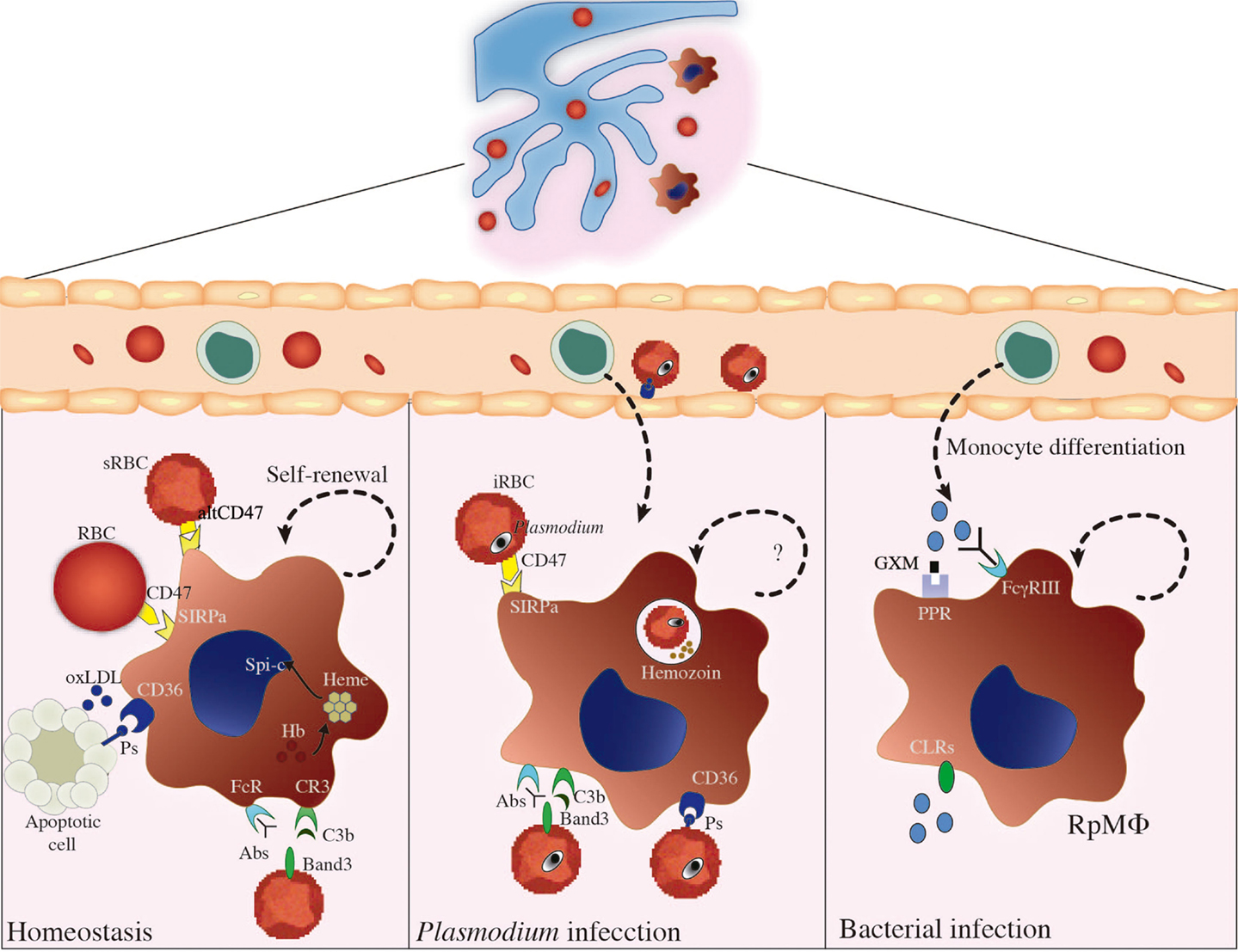
Figure 2. RpMΦ biology during homeostasis and infection. This figure summarizes the different roles of RpMΦs in maintenance of host homeostasis and in the control of different infections. In the absence of infection (left), RpMΦs play important roles in the uptake of apoptotic cells, oxidized LDL (oxLDL), or senescent RBCs (sRBCs) from the circulations, through interaction with receptors such as SIRPα, CD36, CR3, or FcRs. CD47 expression on RBCs is an inhibitory signal for phagocytosis mediated by SIRPα, but sRBCs expressing a modified isoform of this molecule (altCD47) are phagocytized by RpMΦs. CD36 binds to phosphatidylserine (PS) and, alternatively, to oxLDL. RpMΦs are also important for iron homeostasis, and conversely, iron homeostasis seems to control RpMΦ development, through the action of free heme on Spi-C transcriptional factor. In these situations, RpMΦs have the ability of self-renewal by proliferation. Beyond the task of maintaining blood homeostasis, RpMΦs contribute to control blood-borne infections such as malaria (center) or bacterial infections (right) lead to changes in RpMΦ function. Plasmodium-infected RBCs (iRBCs) are recognized through the same receptors that recognize sRBCs, such as SIRPα, CR3, FcRs, or CD36, inferring a role for RpMΦs in parasite clearance. However, the adherence of iRBCs to microvascular endothelium through CD36 prevents iRBC clearance inside the spleen. Interestingly, P. yoelii parasites preferentially infect young RBCs expressing high levels of CD47 and, in consequence, escape from splenic clearance. RpMΦs also present with other receptors such as CLRs and PPRs, which in conjunct with FcγRIII contribute to recognition and elimination of bacteria from circulation. RpMΦs can recognize the capsular polysaccharide glucuronoxylomannan (GXM) from Cryptococcus neoformans and subsequently phagocytize the bacteria. The ability of RpMΦ renewal during infections, however, is poorly understood, and substitution of dead RpMΦs for monocyte-derived RpMΦs is presumable.
Marginal zone macrophages and MMMΦs have unique characteristics that contribute to rapid phagocytosis of pathogens and other particles. Thus, these MΦs act like scavenger cells, developing pro- or anti-inflammatory responses depending on the nature of the interaction. MZMΦs express SIGNR1 that binds to yeasts and the yeast-derived particle zymosan (4), to bacteria such as Mycobacterium tuberculosis (56), S. pneumoniae (57), E. coli, and S. typhimurium (58), and to virus such as human immunodeficiency virus (HIV) (4). This receptor recognizes carbohydrate antigens from blood-borne pathogens and mediates their subsequent internalization into phagosomes (4). Although SIGNR1 in peritoneal MΦs cooperate with dectin-1 in zymosan uptake (59), these innate receptors colocalize poorly in MZMΦs (60). Similar to classical complement pathway activation, but independently of antibodies, SIGNR1 also binds C1q and assembles the complex C4bC2a or C3 convertase that catalyzes C3b opsonin formation (61). This mechanism was shown to provide resistance to intravenous S. pneumoniae infection.
Expression of the scavenger receptor MARCO is upregulated in different MΦ populations, especially in MZMΦs and MΦs in the medullary cord of lymph nodes (3). MARCO was firstly reported to bind and mediate uptake of Gram-negative bacteria and also to recognize oxLDL [reviewed in Ref. (62)]. The structure of MARCO is similar to that of the Scavenger Receptor A1 (SR-A1, CD204), which plays a role in bacteria and virus recognition (3). TLR-mediated activation of BMDMs stimulates MARCO-mediated phagocytic activity (63). Furthermore, MARCO in MZMΦs directly binds and mediates phagocytosis of E. coli and S. aureus bacteria (3). TLR engagement leads to activation of transcriptional mechanisms that increase phagocytosis and cell activation, and MARCO seems to work in conjunct with TLRs in order to mediate pathogen control (64).
Marginal zone macrophages and MMMΦs are fundamental in the early control of Listeria monocytogenes bacteremia, as evaluated by depletion of these MΦs using a low dose of clodronate liposomes (65). T-cell responses are not affected in this experimental model, ruling out the participation of MZMΦs and MMMΦs as antigen-presenting cells. Similar findings were reported during infection with Neisseria meningitidis (64), thus it is likely that these MΦs have a direct role in the elimination of bacteria from circulation. Conversely, adenoviruses colocalize with MZMΦs as soon as a few minutes after intravenous injection in mice (66). MZMΦs and MMMΦs play a similar role in lymphocytic choriomeningitis virus (LCMV) infection, corroborating the importance of these MΦs in first-line antiviral defense (67). On the other hand, localization of MZMΦs and MMMΦs in the interface between the bloodstream and lymphocyte-rich zones makes them suitable to bridge innate and adaptive immunity in several situations. For instance, mice lacking SRs MARCO and SR-A1 show a defective microarchitecture of the splenic MZ and an impaired T-independent type 2 response when challenged with pneumococcal polysaccharide (68). MMMΦs also collaborate in cytotoxic T-cell activation by transferring antigen directly to CD8α+ DCs, which are specialized in cross-presentation to CD8+ T cells (69). This observation supports the use of the MMMΦs antigen-concentrating capacity in therapeutic strategies for the development of antitumor immunity. The different roles of MZMΦs and MMMΦs in blood-borne infections are shown in Figure 3.
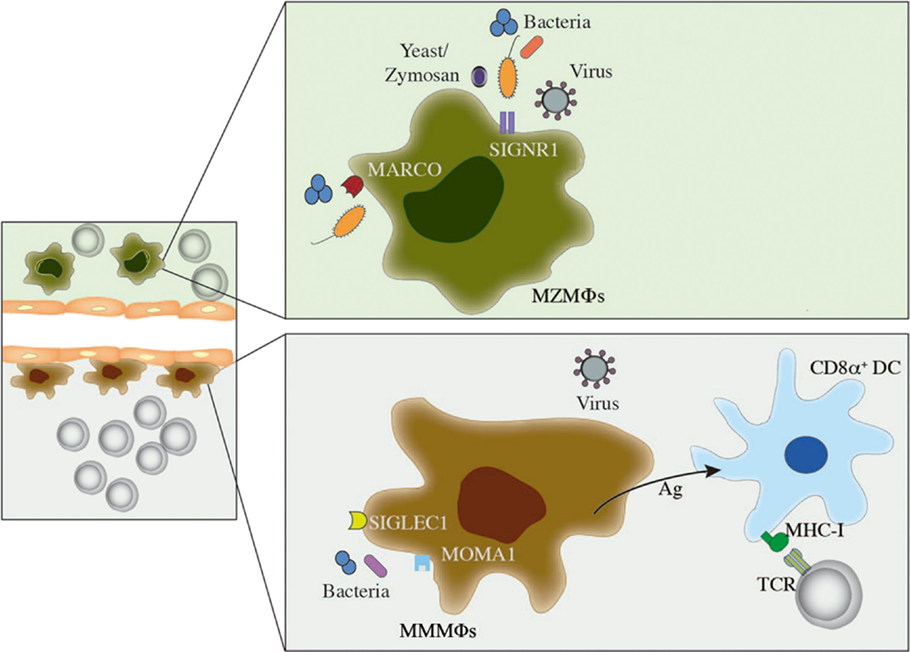
Figure 3. Role of MZMΦs and MMMΦs during infection. In this figure, a brief description on how MZMΦs and MMMΦs are able to recognize and mediate protection against blood-borne pathogens is shown. MZMΦs (above) can recognize bacterial and viral infections by receptors such as MARCO or SIGNR1, which usually induce internalization and further pathogen degradation. A similar feature can be depicted for MMMΦs, where MOMA-1 or SIGLEC can mediate pathogen recognition and elimination from circulation. MMMΦs can also interact with CD8α+ dendritic cells (DCs), which ultimately lead to CD8+ T-cell activation.
In several aspects, splenic MΦs shape splenic structure and/or microenvironment. The development of splenomegaly is typical in blood-borne infections, and it is characterized by profound changes in splenic microarchitecture, including remodeling of Rp (1). Given this, splenic MΦs are expected to play a prominent role in the recruitment of different cell types during acute immune responses. For example, RpMΦs recruit neutrophils into the spleen during early Candida infection by releasing CXCL1 and CXCL2, through autophagy-mediated depletion of the NF-κB inhibitor molecule A20 (70). Another example is the arresting of T cells inside the Rp during acute Plasmodium infection (39). RpMΦs may produce CXCR3- and/or CCR5-binding chemokines by a mechanism similar to that observed during early Candida infection – CXCR3 and CCR5 are the main upregulated chemokine receptors in splenic CD4+ T cells during acute blood-stage malaria (71). However, splenic MΦs might also act on splenic microenvironment after an acute infection. For example, apoptotic cell uptake induces CCL22 production by MMMΦs, which in turn induces Foxp3+ Tregs and DCs recruitment and accumulation, leading to a state of tolerance (72). Since the accumulation of apoptotic cells is a normal feature after acute blood-borne infections (20), a similar mechanism possibly takes place. RPMΦs can also prevent autoimmunity by producing anti-inflammatory cytokines such as TGF-β and IL-10 and by inducing the generation of Treg cells (55). These cytokines may be important – besides limiting autoimmunity – to curb an excessive immune response that could be dangerous to the host after pathogen clearance.
Conversely, the splenic structure and its microenvironment seem to play pivotal roles in MΦ homing and function. For instance, arrangement of sinusoidal endothelial cells inside Rp hampers the circulation of aging and/or iRBCs (1), facilitating their trapping inside Rp and posterior phagocytosis by RpMΦs. Importantly, the cytokine milieu in the microenvironment – which varies throughout an acute infection – may also dictate RpMΦ function. Classic M1 MΦs have an enhanced capacity to accumulate iron, which positively influences the maintenance of these cells in a proinflammatory state. On the other hand, alternative M2 MΦs have an increased ability to release iron, and increased iron availability in the microenvironment seemingly favors tissue remodeling [reviewed in Ref. (73)]. These effects can easily be associated with RpMΦs especially considering their role in iron uptake (1), thus it is possible that RpMΦs play distinct roles as M1- or M2-skewing microenvironments may occur during the beginning of an acute blood-borne infection or after pathogen clearance, respectively. Furthermore, the MZ contains a large number of resident cells that apparently depend on each other for their localization, thereby establishing and maintaining MZ integrity (1). For example, studies in which B cells were absent from the time of birth or in which they are depleted led to disappearance of MZMΦs and MMMΦs (74). Thus, the continuous interaction between resident and transmigrating cells inside the spleen MZ is crucial for efficient homing of MZMΦs and MMMΦs as well as for efficient removal and destruction of blood-borne pathogens by these cells. Lymphotoxin (LT) and TNF also influence the dynamics of MZMΦs and MMMΦs. L. donovani-infected mice have profound changes in the splenic MZ including loss of MZMΦs, which depend on TNF signaling that may increase MZMΦs susceptibility to parasite-induced cell death (75). These changes block lymphocyte traffic in the white pulp, impairing the development of an appropriate adaptive immune response. In another case, MZ B cells secrete LT-α1β2, and this leads to induction of a range of chemokines that could, in turn, influence lodging and retention of MZMΦs (76).
As discussed above, splenic MΦs (RpMΦs, MZMΦs, and MMMΦs) play important roles in the control of blood-borne infections and shape several aspects of innate and adaptive immune responses (Table 1). Thus, a clear concept on the nature of splenic MΦ populations can be drawn, in which their interplay with the splenic microenvironment guarantees efficient control of blood-borne pathogens and maintenance of homeostasis following these infections. At the same time, the splenic structure is likely fundamental for proper localization and function of these MΦs. However, several questions on the nature and function of these cells are still unanswered, especially on (a) the development of splenic MΦs during embryogenesis, (b) the exact signals required for the homeostatic maintenance of these cells, and (c) the extent of how important each of these subsets are for the development of immunity against blood-borne infections. The development of mouse models to accurately study the distinct roles of RpMΦs, MZMΦs, and MMMΦs as well as the development of more detailed studies on signaling pathways and epigenetic modifications on genes involved in the function of these cells will be of great utility to solve these questions.

Table 1. Overview of splenic MΦ subsets.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This study was supported by grants from the São Paulo Research Foundation (FAPESP) 2013/07140-2 (ML) and 2014/00810-5 (HS) and from National Council for Scientific and Technological Development (CNPq) 303676/2014-0 (ML) and 448765/2014-4 (ML). The funders had no role in the preparation of the manuscript.
1. Mebius RE, Kraal G. Structure and function of the spleen. Nat Rev Immunol (2005) 5:606–16. doi: 10.1038/nri1669
2. Kohyama M, Ise W, Edelson BT, Wilker PR, Hildner K, Mejia C, et al. Role for Spi-C in the development of red pulp macrophages and splenic iron homeostasis. Nature (2009) 457:318–21. doi:10.1038/nature07472
3. Elomaa O, Kangas M, Sahlberg C, Tuukkanen J, Sormunen R, Liakka I, et al. Cloning of a novel bacteria-binding receptor structurally related to scavenger receptors and expressed in a subset of macrophages. Cell (1995) 80:603–9. doi:10.1016/0092-8674(95)90514-6
4. Geijtenbeek TB, Groot PC, Nolte MA, van Vliet SJ, Gangaram-Panday ST, van Duijnhoven GC, et al. Marginal zone macrophages express a murine homologue of DC-SIGN that captures blood-borne antigens in vivo. Blood (2002) 100:2908–16. doi:10.1182/blood-2002-04-1044
5. Kraal G, Janse M. Marginal metallophilic cells of the mouse spleen identified by a monoclonal antibody. Immunology (1986) 58:665–9.
6. Liu G, Xia XP, Gong SL, Zhao Y. The macrophage heterogeneity: difference between mouse peritoneal exudate and splenic F4/80+ macrophages. J Cell Physiol (2006) 209:341–52. doi:10.1002/jcp.20732
7. Campos MA, Almeida IC, Takeuchi O, Akira S, Valente EP, Procópio DO, et al. Activation of toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J Immunol (2001) 167:416–23. doi:10.4049/jimmunol.167.1.416
8. Krishnegowda G, Hajjar AM, Zhu J, Douglass EJ, Uematsu S, Akira S, et al. Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: cell signaling receptors, glycosylphosphatidylinositol (GPI) structural requirement, and regulation of GPI activity. J Biol Chem (2005) 280:8606–16. doi:10.1074/jbc.M413541200
9. Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, et al. A toll-like receptor recognizes bacterial DNA. Nature (2000) 408:740–5. doi:10.1038/35047123
10. Parroche P, Lauw FN, Goutagny N, Latz E, Monks BG, Visintin A, et al. Malaria hemozoin is immunologically inert but radically enhances innate responses by presenting malaria DNA to toll-like receptor 9. Proc Natl Acad Sci U S A (2007) 104:1919–24. doi:10.1073/pnas.0608745104
11. Ohashi K, Burkart V, Flohe S, Kolb H. Cutting edge: heat shock protein 60 is a putative endogenous ligand of the toll-like receptor-4 complex. J Immunol (2000) 164:558–61. doi:10.4049/jimmunol.164.2.558
12. Zhang R, Wan XJ, Zhang X, Kang QX, Bian JJ, Yu GF, et al. Plasma HSPA12B is a potential predictor for poor outcome in severe sepsis. PLoS One (2014) 9:e101215. doi:10.1371/journal.pone.0101215
13. Bianco AE, Favaloro JM, Burkot TR, Culvenor JG, Crewther PE, Brown GV, et al. A repetitive antigen of Plasmodium falciparum that is homologous to heat shock protein 70 of Drosophila melanogaster. Proc Natl Acad Sci U S A (1986) 83:8713–7. doi:10.1073/pnas.83.22.8713
14. Okamura Y, Watari M, Jerud ES, Young DW, Ishizaka ST, Rose J, et al. The extra domain A of fibronectin activates toll-like receptor 4. J Biol Chem (2001) 276:10229–33. doi:10.1074/jbc.M100099200
15. Martin-Jaular L, Ferrer M, Calvo M, Rosanas-Urgell A, Kalko S, Graewe S, et al. Strain-specific spleen remodelling in Plasmodium yoelii infections in Balb/c mice facilitates adherence and spleen macrophage-clearance escape. Cell Microbiol (2011) 13:109–22. doi:10.1111/j.1462-5822.2010.01523.x
16. Shinji H, Seki K, Tajima A, Uchida A, Masuda S. Fibronectin bound to the surface of Staphylococcus aureus induces association of very late antigen 5 and intracellular signaling factors with macrophages. Infect Immun (2003) 71:140–6. doi:10.1128/IAI.71.1.140-146.2003
17. Stros M. HMGB proteins: interactions with DNA and chromatin. Biochim Biophys Acta (2010) 1799:101–13. doi:10.1016/j.bbagrm.2009.09.008
18. Hori O, Brett J, Slattery T, Cao R, Zhang J, Chen JX, et al. The receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin. Mediation of neurite outgrowth and coexpression of RAGE and amphoterin in the developing nervous system. J Biol Chem (1995) 270:25752–61. doi:10.1074/jbc.270.43.25752
19. Qin S, Wang H, Yuan R, Li H, Ochani M, Ochani K, et al. Role of HMGB1 in apoptosis-mediated sepsis lethality. J Exp Med (2006) 203:1637–42. doi:10.1084/jem.20052203
20. Elias RM, Sardinha LR, Bastos KR, Zago CA, da Silva AP, Alvarez JM, et al. Role of CD28 in polyclonal and specific T and B cell responses required for protection against blood stage malaria. J Immunol (2005) 174:790–9. doi:10.4049/jimmunol.174.2.790
21. Higgins SJ, Xing K, Kim H, Kain DC, Wang F, Dhabangi A, et al. Systemic release of high mobility group box 1 (HMGB1) protein is associated with severe and fatal Plasmodium falciparum malaria. Malar J (2013) 12:105. doi:10.1186/1475-2875-12-105
22. Marodi L, Korchak HM, Johnston RB Jr. Mechanisms of host defence against Candida species I. Phagocytosis by monocytes and monocyte-derived macrophages. J Immunol (1991) 146:2783–9.
23. Nguyen DG, Hildreth JE. Involvement of macrophage mannose receptor in the binding and transmission of HIV by macrophages. Eur J Immunol (2003) 33:483–93. doi:10.1002/immu.200310024
24. Kahn SJ, Wleklinski M, Ezekowitz RA, Coder D, Aruffo A, Farr A. The major surface glycoprotein of Trypanosoma cruzi amastigotes are ligands of the human serum mannose-binding protein. Infect Immun (1996) 64:2649–56.
25. Stuart LM, Deng J, Silver JM, Takahashi K, Tseng AA, Hennessy EJ, et al. Response to Staphylococcus aureus requires CD36-mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. J Cell Biol (2005) 170:477–85. doi:10.1083/jcb.200501113
26. Mota MM, Jarra W, Hirst E, Patnaik PK, Holder AA. Plasmodium chabaudi-infected erythrocytes adhere to CD36 and bind to microvascular endothelial cells in an organ-specific way. Infect Immun (2000) 68:4135–44. doi:10.1128/IAI.68.7.4135-4144.2000
27. Guarda G, Zenger M, Yazdi AS, Schroder K, Ferrero I, Menu P, et al. Differential expression of NLRP3 among hematopoietic cells. J Immunol (2011) 186:2529–34. doi:10.4049/jimmunol.1002720
28. Kalantari P, DeOliveira RB, Chan J, Corbett Y, Rathinam V, Stutz A, et al. Dual engagement of the NLRP3 and AIM2 inflammasomes by Plasmodium-derived hemozoin and DNA during malaria. Cell Rep (2014) 6:196–210. doi:10.1016/j.celrep.2013.12.014
29. Ataide MA, Andrade WA, Zamboni DS, Wang D, Souza Mdo C, Franklin BS, et al. Malaria-induced NLRP12/NLRP3-dependent caspase-1 activation mediates inflammation and hypersensitivity to bacterial superinfection. PLoS Pathog (2014) 10:e1003885. doi:10.1371/journal.ppat.1003885
30. Levano-Garcia J, Dluzewski AR, Markus RP, Garcia CR. Purinergic signalling is involved in the malaria parasite Plasmodium falciparum invasion to red blood cells. Purinergic Signal (2010) 6:365–72. doi:10.1007/s11302-010-9202-y
31. Hashimoto D, Chow A, Noizat C, Teo P, Beasley MB, Leboeuf M, et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity (2013) 38:792–804. doi:10.1016/j.immuni.2013.04.004
32. Haldar M, Kohyama M, So AY, Kc W, Wu X, Briseño CG, et al. Heme-mediated SPI-C induction promotes monocyte differentiation into iron-recycling macrophages. Cell (2014) 156:1223–34. doi:10.1016/j.cell.2014.01.069
33. Naito M, Hayashi SI, Yoshida H, Takahashi K. Abnormal differentiation of tissue macrophage populations in “osteopetrosis” (op) mice defective in the production of macrophage colony-stimulating factor (M-CSF) or CSF-1. Am J Pathol (1991) 139:657–67.
34. Rutherford MS, Schook LB. Differential immunocompetence of macrophages derived using macrophage or granulocyte-macrophage colony-stimulating factor. J Leukoc Biol (1992) 51:69–76.
35. Schnitzer B, Sodeman T, Mead ML, Contacos PG. Pitting function of the spleen in malaria: ultrastructural observations. Science (1972) 177:175–7. doi:10.1126/science.177.4044.175
36. De Jesus M, Park CG, Su Y, Goldman DL, Steinman RM, Casadevall A. Spleen deposition of Cryptococcus neoformans capsular glucuronoxylomannan in rodents occurs in red pulp macrophages and not marginal zone macrophages expressing the C-type lectin SIGN-R1. Med Mycol (2008) 46:153–62. doi:10.1080/13693780701747182
37. Kirby AC, Beattie L, Maroof A, van Rooijen N, Kaye PM. SIGNR1-negative red pulp macrophages protect against acute streptococcal sepsis after Leishmania donovani-induced loss of marginal zone macrophages. Am J Pathol (2009) 175:1107–15. doi:10.2353/ajpath.2009.090258
38. Salcedo SP, Noursadeghi M, Cohen J, Holden DW. Intracellular replication of Salmonella typhimurium strains in specific subsets of splenic macrophages in vivo. Cell Microbiol (2001) 3:587–97. doi:10.1046/j.1462-5822.2001.00137.x
39. Borges da Silva H, Fonseca R, Cassado Ados A, Machado de Salles É, de Menezes MN, Langhorne J, et al. In vivo approaches reveal a key role for DCs in CD4+ T cell activation and parasite clearance during the acute phase of experimental blood-stage malaria. PLoS Pathog (2015) 11:e1004598. doi:10.1371/journal.ppat.1004598
40. Ezekowitz RA, Gordon S. Down-regulation of mannosyl receptor-mediated endocytosis and antigen F4/80 in bacillus Calmette-Guérin-activated mouse macrophages. Role of T lymphocytes and lymphokines. J Exp Med (1982) 155:1623–37. doi:10.1084/jem.155.6.1623
41. Kay MM, Goodman SR, Sorensen K, Whitfield CF, Wong P, Zaki L, et al. Senescent cell antigen is immunologically related to band 3. Proc Natl Acad Sci U S A (1983) 80:1631–5. doi:10.1073/pnas.80.6.1631
42. Boas FE, Forman L, Beutler E. Phosphatidylserine exposure and red cell viability in red cell aging and in hemolytic anemia. Proc Natl Acad Sci U S A (1998) 95:3077–81. doi:10.1073/pnas.95.6.3077
43. Burger P, Hilarius-Stokman P, de Korte D, van den Berg TK, van Bruggen R. CD47 functions as a molecular switch for erythrocyte phagocytosis. Blood (2012) 119:5512–21. doi:10.1182/blood-2011-10-386805
44. Olsson M, Oldenborg PA. CD47 on experimentally senescent murine RBCs inhibits phagocytosis following Fc gamma receptor-mediated but not scavenger receptor-mediated recognition by macrophages. Blood (2008) 112:4259–67. doi:10.1182/blood-2008-03-143008
45. Hillery CA, Scott JP, Du MC. The carboxy-terminal cell-binding domain of thrombospondin is essential for sickle red blood cell adhesion. Blood (1999) 94:302–9.
46. Banerjee R, Khandelwal S, Kozakai Y, Sahu B, Kumar S. CD47 regulates the phagocytic clearance and replication of the Plasmodium yoelii malaria parasite. Proc Natl Acad Sci U S A (2015) 112:3062–7. doi:10.1073/pnas.1418144112
47. Haldar K, Mohandas N. Malaria, erythrocytic infection, and anemia. Hematology Am Soc Hematol Educ Program (2009) 2009:87–93. doi:10.1182/asheducation-2009.1.87
48. Grimminger F, Rose F, Sibelius U, Meinhardt M, Pötzsch B, Spriestersbach R, et al. Human endothelial cell activation and mediator release in response to the bacterial exotoxins Escherichia coli hemolysin and staphylococcal alpha-toxin. J Immunol (1997) 159:1909–16.
49. Kristiansen M, Graversen JH, Jacobsen C, Sonne O, Hoffman HJ, Law SK, et al. Identification of the haemoglobin scavenger receptor. Nature (2001) 409:198–201. doi:10.1038/35051594
50. Kovtunovych G, Eckhaus MA, Ghosh MC, Ollivierre-Wilson H, Rouault TA. Dysfunction of the heme recycling system in heme oxygenase 1-deficient mice: effects on macrophage viability and tissue iron distribution. Blood (2010) 116:6054–62. doi:10.1182/blood-2010-03-272138
51. Vidal S, Tremblay ML, Govoni G, Gauthier S, Sebastiani G, Malo D, et al. The Ity/Lsh/Bcg locus: natural resistance to infection with intracellular parasites is abrogated by disruption of the Nramp1 gene. J Exp Med (1995) 182:655–66. doi:10.1084/jem.182.3.655
52. Fritsche G, Dlaska M, Barton H, Theurl I, Garimorth K, Weiss G. Nramp1 functionality increases inducible nitric oxide synthase transcription via stimulation of IFN regulatory factor 1 expression. J Immunol (2003) 171:1994–8. doi:10.4049/jimmunol.171.4.1994
53. Flo TH, Smith KD, Sato S, Rodriguez DJ, Holmes MA, Strong RK, et al. Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron. Nature (2004) 432:917–21. doi:10.1038/nature03104
54. Gautier EL, Chow A, Spanbroek R, Marcelin G, Greter M, Jakubzick C, et al. Systemic analysis of PPARγ in mouse macrophage populations reveals marked diversity in expression with critical roles in resolution of inflammation and airway immunity. J Immunol (2012) 189:2614–24. doi:10.4049/jimmunol.1200495
55. Kurotaki D, Kon S, Bae K, Ito K, Matsui Y, Nakayama Y, et al. CSF-1-dependent red pulp macrophages regulate CD4 T cell responses. J Immunol (2011) 186:2229–37. doi:10.4049/jimmunol.1001345
56. Koppel EA, Ludwig IS, Hernandez MS, Lowary TL, Gadikota RR, Tuzikov AB, et al. Identification of the mycobacterial carbohydrate structure that binds the C-type lectins DC-SIGN, L-SIGN and SIGNR1. Immunobiology (2004) 209:117–27. doi:10.1016/j.imbio.2004.03.003
57. Lanoue A, Clatworthy MR, Smith P, Green S, Townsend MJ, Jolin HE, et al. SIGN-R1 contributes to protection against lethal pneumococcal infection in mice. J Exp Med (2004) 200:1383–93. doi:10.1084/jem.20040795
58. Nagaoka K, Takahara K, Tanaka K, Yoshida H, Steinman RM, Saitoh S, et al. Association of SIGNR1 with TLR4-MD-2 enhances signal transduction by recognition of LPS in gram-negative bacteria. Int Immunol (2005) 17:827–36. doi:10.1093/intimm/dxh264
59. Taylor PR, Brown GD, Herre J, Williams DL, Willment JA, Gordon S. The role of SIGNR1 and the beta-glucan receptor (dectin-1) in the nonopsonic recognition of yeast by specific macrophages. J Immunol (2004) 172:157–62. doi:10.4049/jimmunol.172.2.1157
60. Reid DM, Montoya M, Taylor PR, Borrow P, Gordon S, Brown GD, et al. Expression of the beta-glucan receptor, Dectin-1, on murine leukocytes in situ correlates with its function in pathogen recognition and reveals potential roles in leukocyte interactions. J Leukoc Biol (2004) 76:86–94. doi:10.1189/jlb.0104031
61. Kang YS, Do Y, Lee HK, Park SH, Cheong C, Lynch RM, et al. A dominant complement fixation pathway for pneumococcal polysaccharides initiated by SIGN-R1 interacting with C1q. Cell (2006) 125:47–58. doi:10.1016/j.cell.2006.01.046
62. Gough PJ, Gordon S. The role of scavenger receptors in the innate immune system. Microbes Infect (2000) 2:305–11. doi:10.1016/S1286-4579(00)00297-5
63. Doyle SE, O’Connell RM, Miranda GA, Vaidya SA, Chow EK, Liu PT, et al. Toll-like receptors induce a phagocytic gene program through p38. J Exp Med (2004) 199:81–90. doi:10.1084/jem.20031237
64. Chen Y, Wermeling F, Sundqvist J, Jonsson AB, Tryggvason K, Pikkarainen T, et al. A regulatory role for macrophage class A scavenger receptors in TLR4-mediated LPS responses. Eur J Immunol (2010) 40:1451–60. doi:10.1002/eji.200939891
65. Aichele P, Zinke J, Grode L, Schwendener RA, Kaufmann SH, Seiler P. Macrophages of the splenic marginal zone are essential for trapping of blood-borne particulate antigen but dispensable for induction of specific T cell responses. J Immunol (2003) 171:1148–55. doi:10.4049/jimmunol.171.3.1148
66. Di Paolo NC, Miao EA, Iwakura Y, Murali-Krishna K, Aderem A, Flavell RA, et al. Virus binding to a plasma membrane receptor triggers interleukin-1 alpha-mediated proinflammatory macrophage response in vivo. Immunity (2009) 31(1):110–21. doi:10.1016/j.immuni.2009.04.015
67. Seiler P, Aichele P, Odermatt B, Hengartner H, Zinkernagel RM, Schwendener RA. Crucial role of marginal zone macrophages and marginal zone metallophils in the clearance of lymphocytic choriomeningitis virus infection. Eur J Immunol (1997) 27:2626–33. doi:10.1002/eji.1830271023
68. Chen Y, Pikkarainen T, Elomaa O, Soininen R, Kodama T, Kraal G, et al. Defective microarchitecture of the spleen marginal zone and impaired response to a thymus-independent type 2 antigen in mice lacking scavenger receptors MARCO and SR-A. J Immunol (2005) 175:8173–80. doi:10.4049/jimmunol.175.12.8173
69. Backer R, Schwandt T, Greuter M, Oosting M, Jüngerkes F, Tüting T, et al. Effective collaboration between marginal metallophilic macrophages and CD8+ dendritic cells in the generation of cytotoxic T cells. Proc Natl Acad Sci U S A (2010) 107:216–21. doi:10.1073/pnas.0909541107
70. Kanayama M, Inoue M, Danzaki K, Hammer G, He YW, Shinohara ML. Autophagy enhances NFκB activity in specific tissue macrophages by sequestering A20 to boost antifungal immunity. Nat Commun (2015) 6:5779. doi:10.1038/ncomms6779
71. Gwyer Findlay E, Villegas-Mendez A, de Souza JB, Inkson CA, Shaw TN, Saris CJ, et al. IL-27 receptor signaling regulates CD4+ T cell chemotactic responses during infection. J Immunol (2013) 190:4553–61. doi:10.4049/jimmunol.1202916
72. Ravishankar B, Shinde R, Liu H, Chaudhary K, Bradley J, Lemos HP, et al. Marginal zone CD169+ macrophages coordinate apoptotic cell-driven cellular recruitment and tolerance. Proc Natl Acad Sci U S A (2014) 111:4215–20. doi:10.1073/pnas.1320924111
73. Recalcati S, Locati M, Gammella E, Invernizzi P, Cairo G. Iron levels in polarized macrophages: regulation of immunity and autoimmunity. Autoimmun Rev (2012) 11:883–9. doi:10.1016/j.autrev.2012.03.003
74. Nolte MA, Arens R, Kraus M, van Oers MH, Kraal G, van Lier RA, et al. B cells are crucial for both development and maintenance of the splenic marginal zone. J Immunol (2004) 172:3620–7. doi:10.4049/jimmunol.172.6.3620
75. Engwerda CR, Ato M, Cotterell SE, Mynott TL, Tschannerl A, Gorak-Stolinska PM, et al. A role for tumor necrosis factor-alpha in remodeling the splenic marginal zone during Leishmania donovani infection. Am J Pathol (2002) 161:429–37. doi:10.1016/S0002-9440(10)64199-5
Keywords: spleen, macrophages, phagocytosis, pattern-recognition receptors, tissue remodeling
Citation: Borges da Silva H, Fonseca R, Pereira RM, Cassado AA, Álvarez JM and D’Império Lima MR (2015) Splenic macrophage subsets and their function during blood-borne infections. Front. Immunol. 6:480. doi: 10.3389/fimmu.2015.00480
Received: 30 June 2015; Accepted: 03 September 2015;
Published: 22 September 2015
Edited by:
Ivan C. Moura, INSERM, FranceReviewed by:
Martin Herrmann, Universitätsklinikum Erlangen, GermanyCopyright: © 2015 Borges da Silva, Fonseca, Pereira, Cassado, Álvarez and D’Império Lima. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Henrique Borges da Silva and Maria Regina D’Império Lima, Department of Immunology, Instituto de Ciências Biomédicas, Universidade de São Paulo, 1730 – Room 226, 2nd Floor, São Paulo 05508-000, Brazil,aGVucmlxdWUuYm9yZ2VzLnNpbHZhQHVzcC5icg==;
relima@usp.br
†Present address: Henrique Borges da Silva, Department of Laboratory Medicine and Pathology, University of Minnesota, Minneapolis, MN, USA
‡Henrique Borges da Silva and Raíssa Fonseca have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.