 Martin Kongsbak
Martin Kongsbak

94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 18 June 2013
Sec. T Cell Biology
Volume 4 - 2013 | https://doi.org/10.3389/fimmu.2013.00148
This article is part of the Research TopicLipid Signaling in T Cell Development and FunctionView all 12 articles
The vitamin D receptor (VDR) is a nuclear, ligand-dependent transcription factor that in complex with hormonally active vitamin D, 1,25(OH)2D3, regulates the expression of more than 900 genes involved in a wide array of physiological functions. The impact of 1,25(OH)2D3-VDR signaling on immune function has been the focus of many recent studies as a link between 1,25(OH)2D3 and susceptibility to various infections and to development of a variety of inflammatory diseases has been suggested. It is also becoming increasingly clear that microbes slow down immune reactivity by dysregulating the VDR ultimately to increase their chance of survival. Immune modulatory therapies that enhance VDR expression and activity are therefore considered in the clinic today to a greater extent. As T cells are of great importance for both protective immunity and development of inflammatory diseases a variety of studies have been engaged investigating the impact of VDR expression in T cells and found that VDR expression and activity plays an important role in both T cell development, differentiation and effector function. In this review we will analyze current knowledge of VDR regulation and function in T cells and discuss its importance for immune activity.
The purpose of the immune system is to recognize and clear pathogens from the body. However, occasionally unwanted immune reactions against self-tissue that lead to autoimmune diseases occur. The frequency of autoimmune diseases such as type 1 diabetes mellitus (Staples et al., 2003; Sloka et al., 2010), rheumatoid arthritis (Vieira et al., 2010), multiple sclerosis (MS) (Hogancamp et al., 1997), and inflammatory bowel disease (Khalili et al., 2012) has been linked to geographic location with a higher incidence of these diseases at higher degrees of latitude. One explanation of this geographical distribution is low exposure to sunlight and hence lower levels of vitamin D (25(OH)D3) at higher degrees of latitude, as confirmed by studies showing an association between low serum levels of 25(OH)D3 and development of autoimmune diseases (Pierrot-Deseilligny and Souberbielle, 2010; Rossini et al., 2010; Greer et al., 2012). Low serum levels of 25(OH)D3 have also been linked to higher susceptibility to infections such as tuberculosis (Nnoaham and Clarke, 2008), influenza (Cannell et al., 2006; Grant, 2008), HIV (Rodriguez et al., 2009), respiratory syncytial virus (Grant, 2008), and viral infections of the upper respiratory tract (Ginde et al., 2009). It is therefore apparent that vitamin D plays a role in immune modulation. A recent acknowledgment that the majority of immune cells expresses the vitamin D receptor (VDR) (Kreutz et al., 1993; Hewison et al., 2003; Baeke et al., 2010; von Essen et al., 2010; Geldmeyer-Hilt et al., 2011; Joseph et al., 2012) and also the enzyme CYP27B1 used for internal conversion of circulating 25(OH)D3 to the VDR-ligand 1,25(OH)2D3 (Hewison et al., 2003; Baeke et al., 2010) has further strengthen this perception.
The cellular actions of 1,25(OH)2D3 are mediated by the VDR, a ligand-dependent transcription regulator molecule belonging to the superfamily of nuclear receptors. In the absence of 1,25(OH)2D3-VDR is mainly distributed to the cytoplasm (Nagpal et al., 2005). Interaction of VDR with its ligand 1,25(OH)2D3 induces formation of two independent protein interaction surfaces on the VDR, one that facilitates association with the retinoid X receptor (RXR) necessary for DNA binding, and one that is required for recruitment of co-regulators necessary for gene modulation (Pike et al., 2012). Following interaction with 1,25(OH)2D3-VDR dimerizes with RXR and translocates to the nucleus where it binds to vitamin D response elements (VDRE) in vitamin D responsive genes. Depending on the target gene either co-activators or co-repressors are attracted to the VDR/RXR complexes to induce or repress gene transcription (Nagpal et al., 2005; Pike et al., 2012; Haussler et al., 2013). Even though details of how these co-regulatory complexes work are only slowly beginning to emerge, it is now evident that they include ATPase-containing nucleosomal remodeling capabilities, enzymes with chromatin histone modifying abilities (e.g., acetyl- or methyl-transferases) and proteins involved in recruitment of RNA polymerase II (Pike et al., 2012; Haussler et al., 2013). Besides regulation through VDRE, VDR can inhibit genes by antagonizing certain transcription factors (Alroy et al., 1995; Takeuchi et al., 1998; Towers and Freedman, 1998). One such example is VDR-dependent inhibition of the T cell cytokine IL-2. Here, VDR first competes with the transcription factor NFAT1 for binding to the enhancer motif of AP1 and subsequently VDR binds to c-Jun. This co-occupancy of VDR-c-Jun to AP1 leads to inhibition of IL-2 expression. The VDR inhibition of the IL-2 gene requires that VDR dimerizes with RXR, illustrating a need for 1,25(OH)2D3 (Alroy et al., 1995; Takeuchi et al., 1998; Towers and Freedman, 1998). Overall, the cellular action of vitamin D therefore depends on sufficient production and delivery of 1,25(OH)2D3 and adequate expression of VDR and its associated proteins. Since the VDR in 1983 was reported to be expressed in immune cells (Bhalla et al., 1983; Provvedini et al., 1983) an increasing effort to elucidate the importance of vitamin D on immune function has been undertaken. It has become increasingly clear that a major mechanism to control the immune regulatory effect of vitamin D is adjustment of the expression level and activity of the VDR.
Due to the importance of T cells in protective immunity and in development of inflammatory and autoimmune disorders, several studies have examined the impact of VDR expression on T cell development, differentiation, and function. One approach to determine the role of VDR expression in development of T cells has been to study mice lacking the VDR (VDR-KO). These mice show normal numbers of CD4+ and CD8+ T cells including naturally occurring CD4+ FoxP3+ regulatory T cells (nTreg) (Yu et al., 2008), suggesting that VDR is not required for development of either of these cell types. A study performed by Hayes and coworkers using a mouse model with defective VDR in only the T cells confirmed that VDR is not essential for development of either conventional CD4+ T cells, CD8+ T cells, or CD4+ FoxP3+ nTreg cells (Mayne et al., 2011). Even so, VDR-KO mice appear to have a more vigorous immune response as seen by their increased risk of development of autoimmune diseases (Froicu et al., 2003; Froicu and Cantorna, 2007), and the enhanced response of VDR-KO T cells in mixed lymphocyte reactions (Froicu et al., 2003). In a series of studies Cantorna and coworkers have establish that the increased immune reactivity observed in VDR-KO mice in part is caused by a failure to develop the two regulatory T cell subsets, invariant NKT (iNKT) cells and CD8αα/TCRαβ T cells (Yu and Cantorna, 2008; Yu et al., 2008; Bruce and Cantorna, 2011). iNKT cells are a subset of T cells with a regulatory role in autoimmunity and infection (Godfrey et al., 2000; Bendelac et al., 2001; Singh et al., 2001). CD8αα T cells are mainly present in the gut, where they help maintain tolerance and suppress inflammation by dampening the response to a large number of gut antigens (Poussier et al., 2002; Cheroutre, 2004). The VDR-KO mice have significant fewer iNKT cells, due to a block in development as VDR is implicated in Tbet expression and conversion to the mature NK1.1 expressing mature iNKT cell. The few iNKT cells present in the periphery are furthermore functionally defective (Yu and Cantorna, 2008; Ooi et al., 2012). Like the iNKT cells, there are also fewer CD8αα/TCRαβ precursors in the thymus of VDR-KO animals. Moreover, to complete development CD8αα/TCRαβ cells must travel from the thymus to the gastrointestinal tract where IL-15 induces proliferation and upregulation of CD8αα. Due to decreased levels of IL-15 receptor expression VDR-KO CD8αα/TCRαβ cells proliferate poorly, resulting in a diminished mature CD8αα/TCRαβ population in the VDR-KO gut (Yu et al., 2008; Bruce and Cantorna, 2011; Ooi et al., 2012). These data illustrate that in contrast to conventional T cells, VDR expression is mandatory for development of both iNKT cells and CD8αα/TCRαβ T cells.
Adaptive immune responses require priming and proliferation of naïve T cells followed by migration of the resulting effector T cells to the site of infection. Antigen-specific triggering of TCRs expressed on the surface of antigen-naïve T cells together with co-stimulation induces intracellular signaling events that promote upregulation of the VDR (Provvedini et al., 1983; von Essen et al., 2010; Joseph et al., 2012). This activation-induced upregulation of VDR in naïve human T cells encourages 1,25(OH)2D3-VDR signaling. 1,25(OH)2D3-VDR signaling induces upregulation of the VDRE containing enzyme PLC-γ1, which is a central molecule in the classical TCR signaling pathway. Following VDR-induced PLC-γ1 upregulation classical TCR signaling is established and full T cell activation accomplished (von Essen et al., 2010). VDR expression therefore contributes to priming of naïve human T cells. Interestingly, this VDR-induced PLC-γ1 upregulation is not a mechanism involved in T cell priming of mouse T cells, as naïve mouse T cells already expresses substantial amounts of PLC-γ1 (Ericsson et al., 1996). In order for T cells to proliferate they need the cytokine IL-2. IL-2 is produced and secreted by T cells in response to antigen-induced T cell stimulation. In an autocrine and paracrine fashion IL-2 binds to high affinity IL-2 receptors on the same or adjacent T cells, inducing cell proliferation and hence a clonally expanded population of antigen-specific effector T cells (Cantrell and Smith, 1984; Smith, 1988). As VDR expression has been shown to inhibit transcription of the IL-2 gene (Alroy et al., 1995; Takeuchi et al., 1998), it is likely that upregulation of VDR serves as a negative feedback mechanism to control potential overreactions of the immune system. Besides inducing the early priming phase of naïve human T cells and possibly ensuring immune integrity, Mathieu and coworkers showed that a 1,25(OH)2D3 agonist drastically changed the surface expression of homing receptors on both CD4 and CD8 T cells, resulting in a profile corresponding to an increased migration ability to sites of infection (Baeke et al., 2011); and hence implying a role for VDR in all phases of T cell differentiation.
In agreement with a suggested role of VDR in preventing immune overreaction, a changed distribution of naïve and antigen-experienced T cells was observed in a VDR-KO study performed by Bruce et al. (2011). The CD4+ T cells had a more activated phenotype and readily developed into the proinflammatory Th17 effector cells that produced twice as much IL-17 as their WT counterparts in vitro (Bruce et al., 2011). Furthermore, vitamin D has been shown to modify the phenotype of antigen presenting dendritic cells (DC) to a more tolerogenic phenotype that favors differentiation of inducible Treg (iTreg) cells instead of the inflammatory Th1 and Th17 cells (Griffin et al., 2001; Adorini et al., 2003; Adorini and Penna, 2009). In VDR-KO mice, the frequency of total DC populations were not affected, but a significant reduction in tolerogenic DCs was observed (Bruce et al., 2011). In accordance with the reduced population of tolerogenic DCs and increased population of activated inflammatory T cells, a decrease in the population of iTregs that differentiated from naïve T cells was observed (Bruce et al., 2011). This lead to an increased pathogenic potential of the T cell population, which manifested in development of more severe experimental inflammatory bowel disease (Bruce et al., 2011). These observations emphasize the importance of VDR expression in controlling the balance between effector and tolerogenic cells.
Only few studies have investigated whether there is coherence between VDR expression and T cell effector function. In the iNKT cell study performed by Cantorna and coworkers, a reduction of at least fifty percent in iNKT cells that produced the effector cytokine IL-4 and IFN-γ was observed in multiple organs (Yu and Cantorna, 2008). However, as iNKT cells most likely acquire the ability to transcribe IL-4 and IFN-γ during thymic development at the stage where they diverge from conventional T cells (Bezbradica et al., 2006), it is possible that the reduced cytokine production observed is due to defects in iNKT cell development. In a study of conventional T cells from VDR-KO mice, Bruce et al. (2011) showed that VDR-KO Th17 cells induced in in vitro cultures overproduced IL-17 as compared to WT cells. In contrast to the study performed by Cantorna using iNKT cells from VDR-KO mice, Bruce et al. found no change in IFN-γ production in the cultured conventional VDR-KO T cells. Taking this into consideration and the fact that Th17 cells are more readily induced in the VDR-KO mice, it is likely that the increased IL-17 production observed by Bruce et al. (2011) is also a developmental defect. Conversely, an in vitro study in human T cells performed by Youssef and coworkers favors a direct effect of VDR on IL-17 production. Here they showed that VDR blocks binding of the transcription factor NFAT1 to the promoter of the human IL-17 gene leading to a decrease in IL-17 production (Joshi et al., 2011). This inhibitory mechanism somehow resembles VDR’s control of both IL-2 and GM-CSF transcription in which VDR also inhibits NFAT1 binding to the DNA of the respective cytokine genes (Figure 1) (Alroy et al., 1995; Takeuchi et al., 1998; Towers and Freedman, 1998). As NFAT1 is a transcription factor involved in regulation of a wide range of genes and as VDR’s inhibition of NFAT1 appears not to include a canonical VDRE sequence in the promoter regions (Towers and Freedman, 1998), the transcriptional control of VDR’s target genes is likely far more widespread than first anticipated. Today, a direct effect of 1,25(OH)2D3-VDR signaling on the expression of effector T cell molecules includes not only cytokines but also chemokines and chemokine homing receptors as reviewed by Peelen et al. (2011).
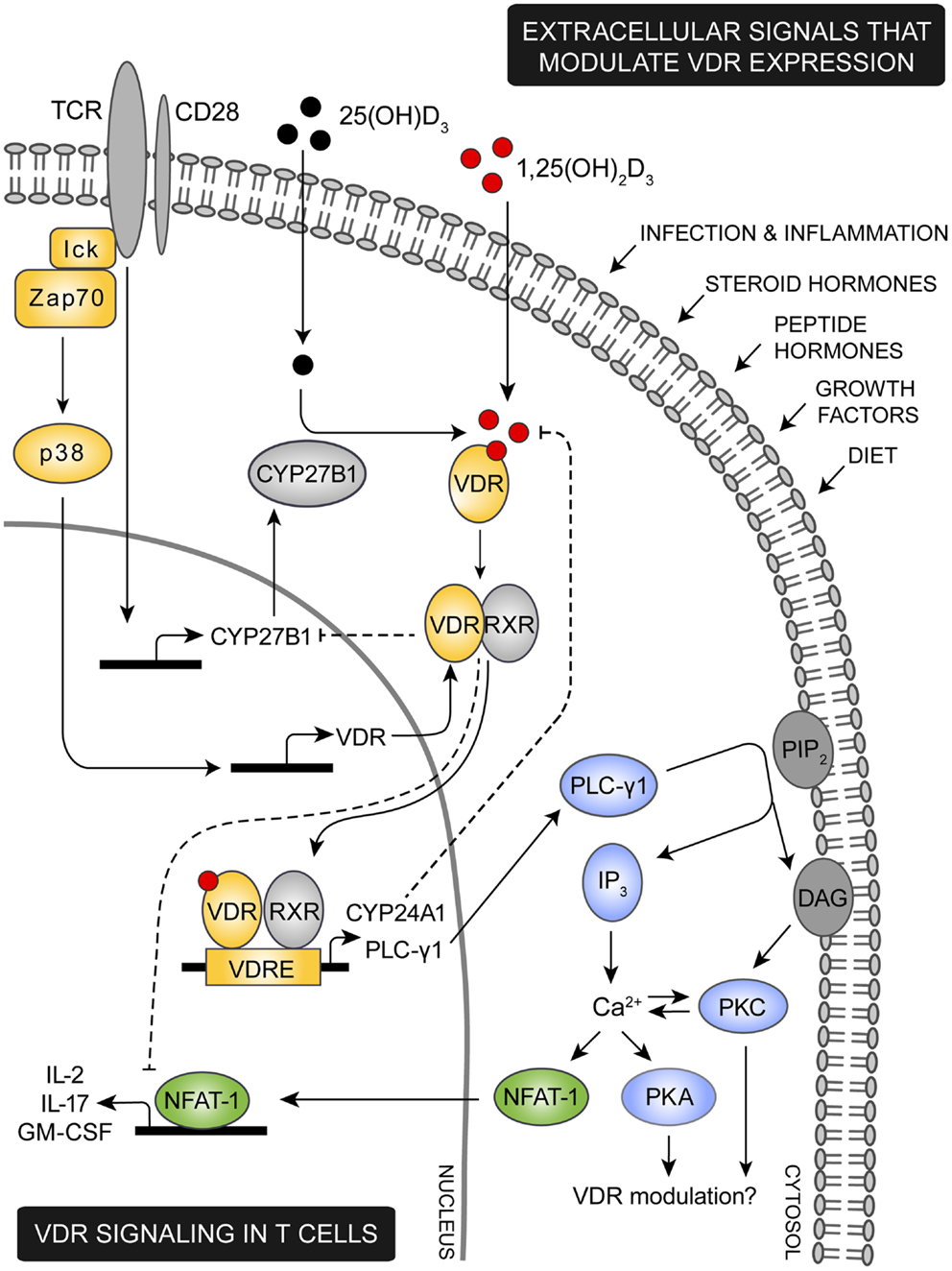
Figure 1. Proposed model for VDR signaling in T cells. Various extracellular signals including infection, inflammation, steroid and peptide hormones, and diet are involved in regulation of the intracellular VDR level. During an immune response the TCR is triggered by specific antigens, inducing a cascade of intracellular signaling events. Among these, lck and ZAP-70 are activated leading to activation of the p38 kinase which in naïve human T cells induce expression of VDR. TCR triggering also promotes expression of the 1,25(OH)2D3 synthesis enzyme CYP27B1. Through intrinsic synthesis of 1,25(OH)2D3 and uptake of 1,25(OH)2D3 from the extracellular environment, VDR is activated and translocated into the nucleus where it either induce or suppress transcription of a variety of genes. As an example, VDR induce upregulation of PLC-γ1 in naïve human T cells. Once PLC-γ1 is expressed, TCR induced activation of PLC-γ1 leads to activation of PKA and PKC and an increase in the intracellular calcium level. In other cell types PKA and PKC has been shown to modulate expression of VDR, depending on the particular cell type and cellular differentiation state investigated. An increase in intracellular calcium concentration activates NFAT1 a necessary transcription factor for expression of IL-2 and other cytokines. IL-2 is a cytokine required for proliferation of T cells and one mechanism by which VDR adjust T cell activity is to outcompete NFAT1’s binding to the IL-2 promoter and furthermore to down-regulate the actual expression of NFAT1. To control VDR activity a series of negative feedback loops exists; activated VDR both induce expression of the 1,25(OH)2D3 degrading enzyme CYP24A1 and down-regulates expression of the 1,25(OH)2D3 synthesizing enzyme CYP27B1.
Studies in which either T cell conditional VDR knock-out animals or animal models of adoptive transferred T cells from a VDR knock-out animal would substantially increase our understanding of VDR’s direct influence on T cell effector function. Along this line Hayes and coworkers developed a mouse model in which only the T cells included an inactive VDR gene in order to investigate the implication of T cells in development of autoimmunity. In this model T cells developed normally in thymus but peripheral T cells expressing an inactive VDR were resistant to the inhibitory effect of vitamin D on autoimmune disease development (Mayne et al., 2011). Future studies will likely elaborate on specific T cell effector functions in similar animal models.
Allelic variants of the VDR gene occur naturally in the human population. Even though interpretation of these polymorphic variants in relation to different diseases is difficult due to the small numbers of subjects included in the performed studies, an association with disease risk has been suggested. This includes a greater susceptibility to infections and a higher incidence of autoimmune diseases and cancer. The impact of VDR polymorphism on VDR function may in part be due to reduced VDR-mRNA stability and hence of reduced VDR expression (Feldman et al., 2005). In addition to allelic variations of the VDR gene, a rare genetic disorder has been described in which the VDR gene contain mutations that renders the gene product non-functional. This includes mutations in the DNA-binding domain and in the ligand binding domain, rendering binding of VDR to DNA, RXR, or co-regulators impossible (Malloy and Feldman, 2010). Individuals with a non-functional VDR suffer from the absence of VDR signaling giving rise to the disease hereditary vitamin D resistant rickets (HVDRR). In HVDRR patients the serum level of 1,25(OH)2D3 is exceedingly high and most patients are completely resistant to vitamin D therapy. As there are only very few cases of HVDRR, long-term effects of defective VDR signaling on immune function such as development of autoimmune diseases and control of cancer have not yet been documented (Malloy and Feldman, 2010). A promising model system regarding this issue is the VDR-KO mouse in which the VDR gene has been deleted. These mice show increased sensitivity to autoimmune diseases, and are more prone to oncogene- and chemocarcinogen-induced tumors (Bouillon et al., 2008) illustrating a possible in vivo relation between VDR expression and immune function.
The studies described above have led to an understanding of the importance of VDR expression in T cell development, differentiation, and function. Even though the abundance of VDR in T cells reflects the cells responsiveness to 1,25(OH)2D3, this concept likely is far more complex. Besides transcriptional regulation of the VDR, additional factors with an impact on VDR activity should be considered. This includes ligand availability, induction of intracellular signaling pathways, posttranslational modifications of VDR, nuclear translocation, and DNA binding as well as recruitment of activated co-regulators.
The role of the VDR-ligand 1,25(OH)2D3 is to convert VDR into a functionally active protein that can bind to RXR and to specific gene sequences and co-regulators necessary for modulation of gene expression (Pike, 2011; Pike et al., 2012; Haussler et al., 2013). Availability of 1,25(OH)2D3 is therefore a prerequisite for VDR activity. The circulating concentration of 1,25(OH)2D3 is very low (≤100 pM) compared to its metabolic inactive precursor 25(OH)D3 (≤100 nM) (Feldman et al., 2005). During an immune reaction it is therefore most likely that the source of 1,25(OH)2D3 predominantly is endogenous production from the precursor molecule 25(OH)D3. In support of this, several studies of immune cells have revealed that 25(OH)D3 can be taken up and subsequently converted into 1,25(OH)2D3 through the action of the enzyme CYP27B1 (Figure 1) (Jeffery et al., 2012). CYP27B1 has been identified in most cells of the immune system (Fritsche et al., 2003; Liu et al., 2006; Sigmundsdottir et al., 2007; Krutzik et al., 2008; Correale et al., 2009; Baeke et al., 2010), however, it is not clear whether all cells can take up the precursor 25(OH)D3 and convert it. When 1,25(OH)2D3 is synthesized a great part is secreted to adjacent cells, minimizing the need for endogenous production in all immune cells (Jeffery et al., 2012). In addition, 1,25(OH)2D3 holds the capacity to restrict its own synthesis by exerting a negative feedback on the vitamin D signaling system. 1,25(OH)2D3 induces displacement of a key transcriptional factor responsible for CYP27B1 expression leading to a decrease in CYP27B1 (Murayama et al., 1999) and also induces a rapid binding of VDR-RXR to the promoter sequence of the 1,25(OH)2D3 degrading enzyme CYP24A1 leading to an increase in CYP24A1 (Figure 1) (Ohyama et al., 1996; Kim et al., 2005). The net result is a reduction in endogenous 1,25(OH)2D3. As illustrated by Vidal et al. this negative feedback mechanism can be partly prevented by inflammatory-induced proteins. In this study, they showed that IFN-γ induced activation of STAT1 promoted binding of STAT1 to the DNA-binding domain of VDR, preventing VDR from inducing expression of CYP24A1 (Vidal et al., 2002).
A major determinant of 25(OH)D3 availability is the carrier protein DBP that binds most circulating vitamin D in the serum. In immune reactions DBP restricts the availability of 25(OH)D3 to the immune cells (Chun et al., 2010; Jeffery et al., 2012). More than 100 genotypes of DBP have been documented but most people express the three most common variants GC1S, GC1F, and GC2 (Arnaud and Constans, 1993). These DBP variants have different properties including a difference in their affinity for 25(OH)D3 (Arnaud and Constans, 1993; Wood et al., 2011). In vitro studies performed with immune cells using different DBP genotypes in addition to 25(OH)D3 have shown that the particular genotype used influences the magnitude of the immune response (Chun et al., 2010; Jeffery et al., 2012). Along this line, an association between DBP genotype and development of inflammatory diseases has been described (Papiha and Pal, 1985; Speeckaert et al., 2006; Martineau et al., 2010). 1,25(OH)2D3 availability therefore is the sum of the circulating 25(OH)D3 level, DBP genotype, CYP27B1 function, near proximity to other cells that produces and secretes 1,25(OH)2D3 and 1,25(OH)2D3 self-restriction.
Vitamin D receptor expression can be modulated by numerous physical stimuli such as dietary composition (e.g., calcium and phosphorus), steroid hormones, growth factors, peptide hormones (Feldman et al., 2011), and inflammatory agents (Provvedini et al., 1983; Liu et al., 2006, 2009; von Essen et al., 2010; Joseph et al., 2012). For example, VDR expression is significantly regulated by the steroid hormones estrogen, glucocorticoid, and retinoids which appears to be rather cell specific (Feldman et al., 2011). The effect of glucocorticoid on VDR expression in the immune system has not been evaluated, but glucocorticoids are known to have a profound anti-inflammatory and immune suppressive effect (Miller and Ranatunga, 2012). Glucocorticoid therapy is used to suppress inflammation implicated in the pathogenesis of various inflammatory diseases (Hanaoka et al., 2012; Miller and Ranatunga, 2012), and it could be speculated that one mechanism used by glucocorticoids to suppress immune responses is by increasing the expression levels of VDR. Estrogen (Chighizola and Meroni, 2012) and retinoids (Cassani et al., 2012) also appear to have strong immunomodulatory effects, but like glucocorticoid the implication of VDR regulation as a possible mechanism to modulate immune function has not been investigated. Receptors for the peptide hormone parathyroid hormone (PTH) was recently identified on T cells (Geara et al., 2010). This renders PTH-induced modulation of VDR expression in T cells a possibility as observed for other cell types (Feldman et al., 2011). Again, this is unexplored territory even though PTH possesses an immune regulatory ability (Geara et al., 2010). The most well described hormonal effect on VDR activity and expression is that of 1,25(OH)2D3 itself, as 1,25(OH)2D3 directly influences the expression levels of VDR by homologous regulation. Although varying between different cell types, 1,25(OH)2D3 in general increases VDR-mRNA production (McDonnell et al., 1987), stabilizes VDR-mRNA, and protects the VDR against degradation (Feldman et al., 2005), altogether increasing the total amount of the VDR.
Various inflammatory signals have also been shown to induce upregulation of VDR in immune cells. During an innate immune response, pathogen-induce activation of toll-like-receptors on human monocytes and macrophages results in upregulation of the VDR (Liu et al., 2006). Likewise, antigen-induced activation of TCR on human naïve T cells induce upregulation of the VDR (Provvedini et al., 1983; von Essen et al., 2010; Joseph et al., 2012). Furthermore, T cell cytokines induced during inflammation can modulate VDR expression (Edfeldt et al., 2010; Spanier et al., 2012), illustrating that regulation of the VDR level is a common mechanism used in the defense against pathogens.
Modulation of VDR expression as a result of physical stimuli is mediated by various intracellular signaling pathways. Although only a sparse numbers of publications concern this issue, a few studies agree that activation of the cAMP-dependent protein kinase A (PKA) pathway leads to an increase in VDR abundance (Pols et al., 1988; Krishnan and Feldman, 1992; Song, 1996). Both cellular responses to PTH (Pols et al., 1988) and to prostaglandin (Smith et al., 1999) activate PKA causing an increase in the VDR level. In contrast, Feldman and coworkers showed that stimuli that induce protein kinase C (PKC) activity down-regulate both VDR-mRNA and VDR protein levels in fibroblastic cells (Krishnan and Feldman, 1991). Moreover, Reinhardt and Horst (1994) has shown that the impact of PKC activation on the VDR-mRNA level highly depends on the particular cellular differentiation state investigated. This suggests that other signaling pathways may cooperate to determine the final effect on VDR expression. In support of this idea, a study by Krishnan and Feldman (1992) indicated a mutual antagonism between the PKA and PKC pathways in regulation of the VDR level, an observation confirmed by others (van Leeuwen et al., 1992). Furthermore, it has been suggested that the intracellular calcium level that is known to influence and be influenced by PKC activity is implicated in PKA induced VDR upregulation (Figure 1) (van Leeuwen et al., 1990). A new signaling pathway which leads to VDR expression has recently been described in human naïve T cells. Here, TCR stimulation induces VDR expression through activation of the p38 mitogen activated protein kinase by ZAP-70 (Figure 1) (von Essen et al., 2010). In contrast, Gocek et al. (2007) showed that VDR expression was controlled by Erk and PI3K signaling in a myeloid leukemia cell line where p38 activity appeared irrelevant. This implies that not only might different intracellular signaling pathways cooperate to regulate the expression of VDR, but also that the implicated signaling events differs between different cell types and different differentiation states of the cells.
Until recently the regulatory responses to hormones at the VDR-gene promoter were unknown. To clarify this, Zella et al. (2010) used ChIP–chip analysis to investigate the VDR gene transcription. These investigations revealed the presence of several enhancers, including the transcription factor C/EBPβ involved in basal expression of VDR as well as the transcription factor glucocorticoid receptor (GR) which mediates the action of glucocorticoids, the transcription factor retinoid acid receptor (RAR) mediating the action of retinoic acid, and the transcription factor CREB mediating the action of PTH (Zella et al., 2010). In case of VDR enhancement by 1,25(OH)2D3, Zella et al. (2006, 2010) found accumulation of VDR-RXR and RNA pol II at the VDR gene together with an increase in C/EBPβ binding. They also detected a substantial increase in histon H4 acetylation associated with enhancer regions across the VDR locus (Zella et al., 2010). An induction of transcription from promoters is often associated with an increase in H4 acetylation, and the observations therefore indicated the existence of multiple enhancers in the VDR-gene locus that may contribute to 1,25(OH)2D3-induced VDR expression. Transcriptional regulation of the VDR gene therefore includes the presence and activity of a wide range of enhancers induced by extracellular signals as well as induction of various epigenetic changes. In case of inflammatory-induced VDR upregulation, the regulatory responses at transcriptional level have not been investigated. As new techniques such as ChIP–chip and ChIP–seq have emerged, this topic will likely be explored in nearby future.
In addition to transcriptional regulation of VDR, several in vitro studies have suggested that VDR can be post-translationally modified. Studies by Haussler and coworkers revealed that 1,25(OH)2D3 binding to VDR led to serine phosphorylation at multiple sites of the receptor. PKC was implicated in phosphorylation at serine 51, an event that partly inhibited VDR transcriptional activity (Hsieh et al., 1991). Although not required for VDR transcriptional activity, casein kinase II (CK II)-induced phosphorylation at serine 208 led to an enhancement of VDR transcriptional activity (Jurutka et al., 1996). As both PKC and CKII activity is induced in cells in response to various stimuli, it can be proposed that these posttranslational modifications although probably not obligatory for VDR function represents a mode to adjust the activity of VDR according to the specific signals received by the cell. Disease-induced posttranslational modifications leading to a dysfunctional VDR has also been documented. In a study by Patel et al. (1995) plasma toxins from uremic patients was shown to bind to the patients VDR, thereby disrupting binding of VDR-RXR to DNA resulting in a diminished VDR response. It so appears that posttranslational modifications of VDR adjust VDR activity in both health and disease.
The genomic actions of 1,25(OH)2D3 also highly depends on the abundance and activity of proteins that interact with VDR. Binding of VDR to its ligand 1,25(OH)2D3 facilitates association with RXR and in the absence of RXR, VDR is unable to bind to most VDRE in vitamin D target genes (Kliewer et al., 1992; Forman et al., 1995; Chambon, 1996). In addition to RXR binding, VDR interacts with various co-activators or co-repressors once bound to the DNA (Nagpal et al., 2005; Pike et al., 2012; Haussler et al., 2013). These co-regulatory complexes are necessary for the VDR-RXR heterodimer to either induce or suppress gene transcription and include ATPase-containing nucleosomal remodeling capabilities, enzymes with chromatin histone modifying abilities (e.g., acetyl- or methyl-transferases), and proteins involved in recruitment of RNA polymerase II (Pike et al., 2012). GRIP1 (Issa et al., 2001), RAC3 (Issa et al., 2001), SRC-1 (Masuyama et al., 1997a), TIF-1 (vom et al., 1996), ACTR (Chen et al., 1997), pCIP (Torchia et al., 1997), and Mediator (Oda et al., 2010) are some of the described co-activator proteins and co-activator complexes to date. These co-activators all regulate VDR function through co-assembling with VDR but they modulate VDR activity via distinct mechanisms. GRIP1 and RAC3 for example regulate VDR activity by modulating crosstalk between VDR and RXR (Issa et al., 2001), ACTR encompass histone acetyltransferase capacity and can recruit other nuclear factors (Chen et al., 1997), and Mediator which is a large complex composed of several MED-proteins activates transcription by direct recruitment of the RNA polymerase II transcriptional machinery (Oda et al., 2010). Although most co-activators facilitate VDR-induced transcriptional activation by binding to VDR, others are shown to be released from VDR to enable transcription, e.g., TFIIB (Masuyama et al., 1997b); illustrating the functional complexity of these co-activator complexes. Only a few co-repressor proteins involved in VDR silencing of genes have been described. As an example NcoR-1, NcoR-2, and Hairless can recruit histone deacetylase activity to VDR-target genes, leading to chromatin compaction and hence gene silencing (Nagpal et al., 2005). A recent study by Singh et al. (2012) furthermore showed that recruitment of co-repressors inappropriately can change during disease, causing a deregulation of VDR-target genes. In addition to transcriptional control of VDR, co-regulator proteins can modulate VDR abundance by enhancing degradation of VDR. Certain cellular signaling events have been shown to motivate the physical interaction of VDR-1,25(OH)2D3 with SUG1 of the proteasome complex, targeting VDR for ubiquitination and subsequent proteolysis (Masuyama and MacDonald, 1998). Therefore, it is evident that regulation of the expression level of RXR and other co-regulators are important to modulate the activity of VDR, and it could be speculated that expression of particular co-regulators are dictated by the inflammatory environment.
A recent study by Edfeldt et al. revealed that VDR expression is not only modulated on a single cell level. Their study showed that VDR expression of innate immune cells could be regulated by nearby T cells (Edfeldt et al., 2010). In innate immunity, pathogen-induced signaling through Toll-like-receptors on human monocytes and macrophages up-regulate the expression of VDR. This in turn, leads to VDR-induced expression of the antimicrobial peptide cathelicidin resulting in killing of microbes (Liu et al., 2006). VDR-induced cathelicidin expression by human monocytes was shown to be adjusted by cytokines produced by T cells. By modulating the level of VDR and the amount of VDR-ligand available by adjusting the CYP27B1 level, the T cell cytokine IFN-γ increases cathelicidin expression and IL-4 attenuates cathelicidin expression (Edfeldt et al., 2010). This example illustrates how interplay between innate and adaptive immunity cooperates to mount an appropriate response to infection through regulation of the VDR-system.
This review indicates that VDR expression and activity are important for all stages of a T cells life, ranging from development to differentiation and elicitation of effector functions. In concordance, VDR expression and activity are associated with immunity against certain infections and with the prevalence of some autoimmune diseases. In animal models 1,25(OH)2D3 has been shown to prevent development of autoimmune diseases. This includes experimental autoimmune encephalomyelitis (EAE), the animal model for MS (Mayne et al., 2011). EAE studies performed in VDR-KO animals (Bouillon et al., 2008) or in animals with a dysfunctional VDR (Mayne et al., 2011) illustrates the requirement of a functional VDR in 1,25(OH)2D3 mediated EAE-inhibition. Furthermore, a study by Hayes and coworkers showed that VDR-gene inactivation selectively in the T cells completely eradicated the ability of 1,25(OH)2D3 to inhibit EAE (Mayne et al., 2011). The biological relevance of low levels of VDR in development of MS was confirmed in a microarray analysis performed by Achiron et al. Here they compared blood mononuclear cells from healthy subjects that later developed MS with healthy subjects that remained MS-free. One of the early disease markers identified turned out to be suppressed VDR expression (Achiron et al., 2010). These observations may not only reflect a change in conventional T cells (e.g., development of more memory T cells that are predisposed to develop into Th1 and Th17 cells as observed in VDR-KO mice (Bruce et al., 2011) but also a reduced development of iNKT cells (as observed in VDR-KO mice, Yu and Cantorna, 2008; Ooi et al., 2012). iNKT cells are negative regulators of EAE (Matsuda et al., 2008) and furthermore, fewer iNKT cells can be found in the blood of MS patients (Araki et al., 2003). Along this line Araki et al. (2003) showed that an increase in iNKT cell number is associated with remission from symptoms in MS patients. Altogether, these observations emphasize a role for VDR expression in development and progression of autoimmunity.
Most experiments investigating susceptibility to a given autoimmune disease is, however, based on animal models. The question therefore remains whether these animal models which are executed in a pathogen free environment reflect the real life situation where humans continuously are bombarded with a variety of pathogens. It is slowly becoming apparent that the microbial environment has a greater influence on development of autoimmune diseases than previously anticipated. For example, certain microbes have been shown to slow innate immune defenses by dysregulating the VDR. One mechanism used by the innate immune system to clear a pathogen is VDR-induced production of the antimicrobial peptide cathelicidin which possesses antiviral, antibacterial, and antifungal activity. Therefore, any microbe capable of dysregulating expression of the VDR would enhance its chance for survival (Waterhouse et al., 2009; Proal et al., 2013). Klein and coworkers illustrated in vitro that Epstein-Barr virus (EBV) were able to effectively down-regulate expression of VDR in B cells (Yenamandra et al., 2009), Modlin and coworkers that Mycobacterium leprae inhibits VDR activity through down-regulation of CYP27B1 in monocytes (Liu et al., 2012), Wang and coworkers that Mycobacterium tuberculosis down-regulate expression of VDR in macrophages (Xu et al., 2003), and McElvaney and coworkers that the fungus Aspergillus fumigates secretes a toxin capable of down-regulating VDR in macrophages (Coughlan et al., 2012). This allows pathogens to accumulate in tissue and blood and the weakened innate defense further causes susceptibility to additional infections. As more pathogens are incorporated into this microbiome, people start to show symptoms characteristic of inflammatory and autoimmune diseases. Accumulating evidence now supports the observation that a number of autoimmune diseases can be reversed by restoring VDR function (using the VDR agonist olmesartan) along with antibiotics. This includes rheumatoid arthritis, systemic lupus erythematosis, sarcoidosis, scleroderma, psoriasis, Sjogren’s syndrome, autoimmune thyroid disease, and type I and II diabetes mellitus (Waterhouse et al., 2009; Proal et al., 2013). Knowledge of the regulation of VDR abundance and activity in immune cells potentially is of great therapeutic importance, and therapeutic enhancement of VDR should therefore be considered in the clinic today.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Achiron, A., Grotto, I., Balicer, R., Magalashvili, D., Feldman, A., and Gurevich, M. (2010). Microarray analysis identifies altered regulation of nuclear receptor family members in the pre-disease state of multiple sclerosis. Neurobiol. Dis. 38, 201–209. doi:10.1016/j.nbd.2009.12.029
Adorini, L., and Penna, G. (2009). Dendritic cell tolerogenicity: a key mechanism in immunomodulation by vitamin D receptor agonists. Hum. Immunol. 70, 345–352. doi:10.1016/j.humimm.2009.01.016
Adorini, L., Penna, G., Giarratana, N., and Uskokovic, M. (2003). Tolerogenic dendritic cells induced by vitamin D receptor ligands enhance regulatory T cells inhibiting allograft rejection and autoimmune diseases. J. Cell. Biochem. 88, 227–233. doi:10.1002/jcb.10340
Alroy, I., Towers, T. L., and Freedman, L. P. (1995). Transcriptional repression of the interleukin-2 gene by vitamin D3: direct inhibition of NFATp/AP-1 complex formation by a nuclear hormone receptor. Mol. Cell. Biol. 15, 5789–5799.
Araki, M., Kondo, T., Gumperz, J. E., Brenner, M. B., Miyake, S., and Yamamura, T. (2003). Th2 bias of CD4+ NKT cells derived from multiple sclerosis in remission. Int. Immunol. 15, 279–288. doi:10.1093/intimm/dxg029
Arnaud, J., and Constans, J. (1993). Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum. Genet. 92, 183–188. doi:10.1007/BF00219689
Baeke, F., Korf, H., Overbergh, L., van, E. E., Verstuyf, A., Gysemans, C., et al. (2010). Human T lymphocytes are direct targets of 1,25-dihydroxyvitamin D3 in the immune system. J. Steroid Biochem. Mol. Biol. 121, 221–227. doi:10.1016/j.jsbmb.2010.03.037
Baeke, F., Korf, H., Overbergh, L., Verstuyf, A., Thorrez, L., Van, L. L., et al. (2011). The vitamin D analog, TX527, promotes a human CD4+CD25highCD127low regulatory T cell profile and induces a migratory signature specific for homing to sites of inflammation. J. Immunol. 186, 132–142. doi:10.4049/jimmunol.1000695
Bendelac, A., Bonneville, M., and Kearney, J. F. (2001). Autoreactivity by design: innate B and T lymphocytes. Nat. Rev. Immunol. 1, 177–186. doi:10.1038/35105052
Bezbradica, J. S., Gordy, L. E., Stanic, A. K., Dragovic, S., Hill, T., Hawiger, J., et al. (2006). Granulocyte-macrophage colony-stimulating factor regulates effector differentiation of invariant natural killer T cells during thymic ontogeny. Immunity 25, 487–497. doi:10.1016/j.immuni.2006.06.017
Bhalla, A. K., Amento, E. P., Clemens, T. L., Holick, M. F., and Krane, S. M. (1983). Specific high-affinity receptors for 1,25-dihydroxyvitamin D3 in human peripheral blood mononuclear cells: presence in monocytes and induction in T lymphocytes following activation. J. Clin. Endocrinol. Metab. 57, 1308–1310. doi:10.1210/jcem-57-6-1308
Bouillon, R., Carmeliet, G., Verlinden, L., van, E. E., Verstuyf, A., Luderer, H. F., et al. (2008). Vitamin D and human health: lessons from vitamin D receptor null mice. Endocr. Rev. 29, 726–776. doi:10.1210/er.2008-0004
Bruce, D., and Cantorna, M. T. (2011). Intrinsic requirement for the vitamin D receptor in the development of CD8alphaalpha-expressing T cells. J. Immunol. 186, 2819–2825. doi:10.4049/jimmunol.1003444
Bruce, D., Yu, S., Ooi, J. H., and Cantorna, M. T. (2011). Converging pathways lead to overproduction of IL-17 in the absence of vitamin D signaling. Int. Immunol. 23, 519–528. doi:10.1093/intimm/dxr045
Cannell, J. J., Vieth, R., Umhau, J. C., Holick, M. F., Grant, W. B., Madronich, S., et al. (2006). Epidemic influenza and vitamin D. Epidemiol. Infect. 134, 1129–1140. doi:10.1017/S0950268806007175
Cantrell, D. A., and Smith, K. A. (1984). The interleukin-2 T-cell system: a new cell growth model. Science 224, 1312–1316. doi:10.1126/science.6427923
Cassani, B., Villablanca, E. J., De, C. J., Wang, S., and Mora, J. R. (2012). Vitamin A and immune regulation: role of retinoic acid in gut-associated dendritic cell education, immune protection and tolerance. Mol. Aspects Med. 33, 63–76. doi:10.1016/j.mam.2011.11.001
Chen, H., Lin, R. J., Schiltz, R. L., Chakravarti, D., Nash, A., Nagy, L., et al. (1997). Nuclear receptor coactivator ACTR is a novel histone acetyltransferase and forms a multimeric activation complex with P/CAF and CBP/p300. Cell 90, 569–580. doi:10.1016/S0092-8674(00)80516-4
Cheroutre, H. (2004). Starting at the beginning: new perspectives on the biology of mucosal T cells. Annu. Rev. Immunol. 22, 217–246. doi:10.1146/annurev.immunol.22.012703.104522
Chighizola, C., and Meroni, P. L. (2012). The role of environmental estrogens and autoimmunity. Autoimmun. Rev. 11, A493–A501. doi:10.1016/j.autrev.2011.11.027
Chun, R. F., Lauridsen, A. L., Suon, L., Zella, L. A., Pike, J. W., Modlin, R. L., et al. (2010). Vitamin D-binding protein directs monocyte responses to 25-hydroxy- and 1,25-dihydroxyvitamin D. J. Clin. Endocrinol. Metab. 95, 3368–3376. doi:10.1210/jc.2010-0195
Correale, J., Ysrraelit, M. C., and Gaitan, M. I. (2009). Immunomodulatory effects of vitamin D in multiple sclerosis. Brain 132, 1146–1160. doi:10.1093/brain/awp033
Coughlan, C. A., Chotirmall, S. H., Renwick, J., Hassan, T., Low, T. B., Bergsson, G., et al. (2012). The effect of Aspergillus fumigatus infection on vitamin D receptor expression in cystic fibrosis. Am. J. Respir. Crit. Care Med. 186, 999–1007. doi:10.1164/rccm.201203-0478OC
Edfeldt, K., Liu, P. T., Chun, R., Fabri, M., Schenk, M., Wheelwright, M., et al. (2010). T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism. Proc. Natl. Acad. Sci. U.S.A. 107, 22593–22598. doi:10.1073/pnas.1011624108
Ericsson, P. O., Orchansky, P. L., Carlow, D. A., and Teh, H. S. (1996). Differential activation of phospholipase C-gamma 1 and mitogen-activated protein kinase in naive and antigen-primed CD4 T cells by the peptide/MHC ligand. J. Immunol. 156, 2045–2053.
Forman, B. M., Umesono, K., Chen, J., and Evans, R. M. (1995). Unique response pathways are established by allosteric interactions among nuclear hormone receptors. Cell 19, 541–550. doi:10.1016/0092-8674(95)90075-6
Fritsche, J., Mondal, K., Ehrnsperger, A., Andreesen, R., and Kreutz, M. (2003). Regulation of 25-hydroxyvitamin D3-1 alpha-hydroxylase and production of 1 alpha,25-dihydroxyvitamin D3 by human dendritic cells. Blood 102, 3314–3316. doi:10.1182/blood-2002-11-3521
Froicu, M., and Cantorna, M. T. (2007). Vitamin D and the vitamin D receptor are critical for control of the innate immune response to colonic injury. BMC Immunol. 8:5. doi:10.1186/1471-2172-8-5
Froicu, M., Weaver, V., Wynn, T. A., McDowell, M. A., Welsh, J. E., and Cantorna, M. T. (2003). A crucial role for the vitamin D receptor in experimental inflammatory bowel diseases. Mol. Endocrinol. 17, 2386–2392. doi:10.1210/me.2003-0281
Geara, A. S., Castellanos, M. R., Bassil, C., Schuller-Levis, G., Park, E., Smith, M., et al. (2010). Effects of parathyroid hormone on immune function. Clin. Dev. Immunol. 2010:418695. doi:10.1155/2010/418695
Geldmeyer-Hilt, K., Heine, G., Hartmann, B., Baumgrass, R., Radbruch, A., and Worm, M. (2011). 1,25-Dihydroxyvitamin D3 impairs NF-kappaB activation in human naive B cells. Biochem. Biophys. Res. Commun. 407, 699–702. doi:10.1016/j.bbrc.2011.03.078
Ginde, A. A., Mansbach, J. M., and Camargo, C. A. Jr. (2009). Association between serum 25-hydroxyvitamin D level and upper respiratory tract infection in the Third National Health and Nutrition Examination Survey. Arch. Intern. Med. 169, 384–390. doi:10.1001/archinternmed.2008.560
Gocek, E., Kielbinski, M., and Marcinkowska, E. (2007). Activation of intracellular signaling pathways is necessary for an increase in VDR expression and its nuclear translocation. FEBS Lett. 581, 1751–1757. doi:10.1016/j.febslet.2007.03.055
Godfrey, D. I., Hammond, K. J., Poulton, L. D., Smyth, M. J., and Baxter, A. G. (2000). NKT cells: facts, functions and fallacies. Immunol. Today 21, 573–583. doi:10.1016/S0167-5699(00)01735-7
Grant, W. B. (2008). Variations in vitamin D production could possibly explain the seasonality of childhood respiratory infections in Hawaii. Pediatr. Infect. Dis. J. 27, 853. doi:10.1097/INF.0b013e3181817bc1
Greer, R. M., Portelli, S. L., Hung, B. S., Cleghorn, G. J., McMahon, S. K., Batch, J. A., et al. (2012). Serum vitamin D levels are lower in Australian children and adolescents with type 1 diabetes than in children without diabetes. Pediatr. Diabetes 14, 31–41. doi:10.1111/j.1399-5448.2012.00890.x
Griffin, M. D., Lutz, W., Phan, V. A., Bachman, L. A., McKean, D. J., and Kumar, R. (2001). Dendritic cell modulation by 1alpha,25 dihydroxyvitamin D3 and its analogs: a vitamin D receptor-dependent pathway that promotes a persistent state of immaturity in vitro and in vivo. Proc. Natl. Acad. Sci. U.S.A. 98, 6800–6805. doi:10.1073/pnas.121172198
Hanaoka, B. Y., Peterson, C. A., Horbinski, C., and Crofford, L. J. (2012). Implications of glucocorticoid therapy in idiopathic inflammatory myopathies. Nat. Rev. Rheumatol. 8, 448–457. doi:10.1038/nrrheum.2012.85
Haussler, M. R., Whitfield, G. K., Kaneko, I., Haussler, C. A., Hsieh, D., Hsieh, J. C., et al. (2013). Molecular mechanisms of vitamin D action. Calcif. Tissue Int. 92, 77–98. doi:10.1007/s00223-012-9619-0
Hewison, M., Freeman, L., Hughes, S. V., Evans, K. N., Bland, R., Eliopoulos, A. G., et al. (2003). Differential regulation of vitamin D receptor and its ligand in human monocyte-derived dendritic cells. J. Immunol. 170, 5382–5390.
Hogancamp, W. E., Rodriguez, M., and Weinshenker, B. G. (1997). The epidemiology of multiple sclerosis. Mayo Clin. Proc. 72, 871–878. doi:10.4065/72.9.871
Hsieh, J. C., Jurutka, P. W., Galligan, M. A., Terpening, C. M., Haussler, C. A., Samuels, D. S., et al. (1991). Human vitamin D receptor is selectively phosphorylated by protein kinase C on serine 51, a residue crucial to its trans-activation function. Proc. Natl. Acad. Sci. U.S.A. 88, 9315–9319. doi:10.1073/pnas.88.20.9315
Issa, L. L., Leong, G. M., Barry, J. B., Sutherland, R. L., and Eisman, J. A. (2001). Glucocorticoid receptor-interacting protein-1 and receptor-associated coactivator-3 differentially interact with the vitamin D receptor (VDR) and regulate VDR-retinoid X receptor transcriptional cross-talk. Endocrinology 142, 1606–1615. doi:10.1210/en.142.4.1606
Jeffery, L. E., Wood, A. M., Qureshi, O. S., Hou, T. Z., Gardner, D., Briggs, Z., et al. (2012). Availability of 25-hydroxyvitamin D3 to APCs controls the balance between regulatory and inflammatory T cell responses. J. Immunol. 189, 5155–5164. doi:10.4049/jimmunol.1200786
Joseph, R. W., Bayraktar, U. D., Kim, T. K., St John, L. S., Popat, U., Khalili, J., et al. (2012). Vitamin D receptor upregulation in alloreactive human T cells. Hum. Immunol. 73, 693–698. doi:10.1016/j.humimm.2012.04.019
Joshi, S., Pantalena, L. C., Liu, X. K., Gaffen, S. L., Liu, H., Rohowsky-Kochan, C., et al. (2011). 1,25-Dihydroxyvitamin D(3) ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol. Cell. Biol. 31, 3653–3669. doi:10.1128/MCB.05020-11
Jurutka, P. W., Hsieh, J. C., Nakajima, S., Haussler, C. A., Whitfield, G. K., and Haussler, M. R. (1996). Human vitamin D receptor phosphorylation by casein kinase II at Ser-208 potentiates transcriptional activation. Proc. Natl. Acad. Sci. U.S.A. 93, 3519–3524. doi:10.1073/pnas.93.8.3519
Khalili, H., Huang, E. S., Ananthakrishnan, A. N., Higuchi, L., Richter, J. M., Fuchs, C. S., et al. (2012). Geographical variation and incidence of inflammatory bowel disease among US women. Gut 61, 1686–1692. doi:10.1136/gutjnl-2011-301574
Kim, S., Shevde, N. K., and Pike, J. W. (2005). 1,25-Dihydroxyvitamin D3 stimulates cyclic vitamin D receptor/retinoid X receptor DNA-binding, co-activator recruitment, and histone acetylation in intact osteoblasts. J. Bone Miner. Res. 20, 305–317. doi:10.1359/JBMR.041112
Kliewer, S. A., Umesono, K., Mangelsdorf, D. J., and Evans, R. M. (1992). Retinoid X receptor interacts with nuclear receptors in retinoic acid, thyroid hormone and vitamin D3 signalling. Nature 355, 446–449. doi:10.1038/355446a0
Kreutz, M., Andreesen, R., Krause, S. W., Szabo, A., Ritz, E., and Reichel, H. (1993). 1,25-Dihydroxyvitamin D3 production and vitamin D3 receptor expression are developmentally regulated during differentiation of human monocytes into macrophages. Blood 82, 1300–1307.
Krishnan, A. V., and Feldman, D. (1991). Activation of protein kinase-C inhibits vitamin D receptor gene expression. Mol. Endocrinol. 5, 605–612. doi:10.1210/mend-5-4-605
Krishnan, A. V., and Feldman, D. (1992). Cyclic adenosine 3’,5’-monophosphate up-regulates 1,25-dihydroxyvitamin D3 receptor gene expression and enhances hormone action. Mol. Endocrinol. 6, 198–206. doi:10.1210/me.6.2.198
Krutzik, S. R., Hewison, M., Liu, P. T., Robles, J. A., Stenger, S., Adams, J. S., et al. (2008). IL-15 links TLR2/1-induced macrophage differentiation to the vitamin D-dependent antimicrobial pathway. J. Immunol. 181, 7115–7120.
Liu, P. T., Schenk, M., Walker, V. P., Dempsey, P. W., Kanchanapoomi, M., Wheelwright, M., et al. (2009). Convergence of IL-1beta and VDR activation pathways in human TLR2/1-induced antimicrobial responses. PLoS ONE 4:e5810. doi:10.1371/journal.pone.0005810
Liu, P. T., Stenger, S., Li, H., Wenzel, L., Tan, B. H., Krutzik, S. R., et al. (2006). Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 311, 1770–1773. doi:10.1126/science.1123933
Liu, P. T., Wheelwright, M., Teles, R., Komisopoulou, E., Edfeldt, K., Ferguson, B., et al. (2012). MicroRNA-21 targets the vitamin D-dependent antimicrobial pathway in leprosy. Nat. Med. 18, 267–273. doi:10.1038/nm.2584
Malloy, P. J., and Feldman, D. (2010). Genetic disorders and defects in vitamin D action. Endocrinol. Metab. Clin. North Am. 39, 333–346. doi:10.1016/j.ecl.2010.02.004
Martineau, A. R., Leandro, A. C., Anderson, S. T., Newton, S. M., Wilkinson, K. A., Nicol, M. P., et al. (2010). Association between Gc genotype and susceptibility to TB is dependent on vitamin D status. Eur. Respir. J. 35, 1106–1112. doi:10.1183/09031936.00087009
Masuyama, H., Brownfield, C. M., St-Arnaud, R., and MacDonald, P. N. (1997a). Evidence for ligand-dependent intramolecular folding of the AF-2 domain in vitamin D receptor-activated transcription and coactivator interaction. Mol. Endocrinol. 11, 1507–1517. doi:10.1210/me.11.10.1507
Masuyama, H., Jefcoat, S. C. Jr., and MacDonald, P. N. (1997b). The N-terminal domain of transcription factor IIB is required for direct interaction with the vitamin D receptor and participates in vitamin D-mediated transcription. Mol. Endocrinol. 11, 218–228. doi:10.1210/me.11.2.218
Masuyama, H., and MacDonald, P. N. (1998). Proteasome-mediated degradation of the vitamin D receptor (VDR) and a putative role for SUG1 interaction with the AF-2 domain of VDR. J. Cell. Biochem. 71, 429–440. doi:10.1002/(SICI)1097-4644(19981201)71:3<429::AID-JCB11>3.3.CO;2-G
Matsuda, J. L., Mallevaey, T., Scott-Browne, J., and Gapin, L. (2008). CD1d-restricted iNKT cells, the ‘Swiss-Army knife’ of the immune system. Curr. Opin. Immunol. 20, 358–368. doi:10.1016/j.coi.2008.03.018
Mayne, C. G., Spanier, J. A., Relland, L. M., Williams, C. B., and Hayes, C. E. (2011). 1,25-Dihydroxyvitamin D3 acts directly on the T lymphocyte vitamin D receptor to inhibit experimental autoimmune encephalomyelitis. Eur. J. Immunol. 41, 822–832. doi:10.1002/eji.201040632
McDonnell, D. P., Mangelsdorf, D. J., Pike, J. W., Haussler, M. R., and O’Malley, B. W. (1987). Molecular cloning of complementary DNA encoding the avian receptor for vitamin D. Science 235, 1214–1217. doi:10.1126/science.3029866
Miller, A. V., and Ranatunga, S. K. (2012). Immunotherapies in rheumatologic disorders. Med. Clin. North Am. 96, 475–496. doi:10.1016/j.mcna.2012.04.003
Murayama, A., Takeyama, K., Kitanaka, S., Kodera, Y., Kawaguchi, Y., Hosoya, T., et al. (1999). Positive and negative regulations of the renal 25-hydroxyvitamin D3 1alpha-hydroxylase gene by parathyroid hormone, calcitonin, and 1alpha,25(OH)2D3 in intact animals. Endocrinology 140, 2224–2231. doi:10.1210/en.140.5.2224
Nagpal, S., Na, S., and Rathnachalam, R. (2005). Noncalcemic actions of vitamin D receptor ligands. Endocr. Rev. 26, 662–687. doi:10.1210/er.2004-0002
Nnoaham, K. E., and Clarke, A. (2008). Low serum vitamin D levels and tuberculosis: a systematic review and meta-analysis. Int. J. Epidemiol. 37, 113–119. doi:10.1093/ije/dym247
Oda, Y., Chalkley, R. J., Burlingame, A. L., and Bikle, D. D. (2010). The transcriptional coactivator DRIP/mediator complex is involved in vitamin D receptor function and regulates keratinocyte proliferation and differentiation. J. Invest. Dermatol. 130, 2377–2388. doi:10.1038/jid.2010.148
Ohyama, Y., Ozono, K., Uchida, M., Yoshimura, M., Shinki, T., Suda, T., et al. (1996). Functional assessment of two vitamin D-responsive elements in the rat 25-hydroxyvitamin D3 24-hydroxylase gene. J. Biol. Chem. 271, 30381–30385. doi:10.1074/jbc.271.48.30381
Ooi, J. H., Chen, J., and Cantorna, M. T. (2012). Vitamin D regulation of immune function in the gut: why do T cells have vitamin D receptors? Mol. Aspects Med. 33, 77–82. doi:10.1016/j.mam.2011.10.014
Papiha, S. S., and Pal, B. (1985). Gc (vitamin D binding protein) subtypes in rheumatoid arthritis. Hum. Genet. 70, 278–280. doi:10.1007/BF00273457
Patel, S. R., Ke, H. Q., Vanholder, R., Koenig, R. J., and Hsu, C. H. (1995). Inhibition of calcitriol receptor binding to vitamin D response elements by uremic toxins. J. Clin. Invest. 96, 50–59. doi:10.1172/JCI118061
Peelen, E., Knippenberg, S., Muris, A. H., Thewissen, M., Smolders, J., Tervaert, J. W., et al. (2011). Effects of vitamin D on the peripheral adaptive immune system: a review. Autoimmun. Rev. 10, 733–743. doi:10.1016/j.autrev.2011.05.002
Pierrot-Deseilligny, C., and Souberbielle, J. C. (2010). Is hypovitaminosis D one of the environmental risk factors for multiple sclerosis? Brain 133, 1869–1888. doi:10.1093/brain/awq147
Pike, J. W. (2011). Genome-wide principles of gene regulation by the vitamin D receptor and its activating ligand. Mol. Cell. Endocrinol. 347, 3–10. doi:10.1016/j.mce.2011.05.012
Pike, J. W., Meyer, M. B., and Bishop, K. A. (2012). Regulation of target gene expression by the vitamin D receptor – an update on mechanisms. Rev. Endocr. Metab. Disord. 13, 45–55. doi:10.1007/s11154-011-9198-9
Pols, H. A., van Leeuwen, J. P., Schilte, J. P., Visser, T. J., and Birkenhager, J. C. (1988). Heterologous up-regulation of the 1,25-dihydroxyvitamin D3 receptor by parathyroid hormone (PTH) and PTH-like peptide in osteoblast-like cells. Biochem. Biophys. Res. Commun. 156, 588–594. doi:10.1016/S0006-291X(88)80883-0
Poussier, P., Ning, T., Banerjee, D., and Julius, M. (2002). A unique subset of self-specific intraintestinal T cells maintains gut integrity. J. Exp. Med. 195, 1491–1497. doi:10.1084/jem.20011793
Proal, A. D., Albert, P. J., and Marshall, T. G. (2013). The human microbiome and autoimmunity. Curr. Opin. Rheumatol. 25, 234–240. doi:10.1097/BOR.0b013e32835cedbf
Provvedini, D. M., Tsoukas, C. D., Deftos, L. J., and Manolagas, S. C. (1983). 1,25-Dihydroxyvitamin D3 receptors in human leukocytes. Science 221, 1181–1183. doi:10.1126/science.6310748
Reinhardt, T. A., and Horst, R. L. (1994). Phorbol 12-myristate 13-acetate and 1,25-dihydroxyvitamin D3 regulate 1,25-dihydroxyvitamin D3 receptors synergistically in rat osteosarcoma cells. Mol. Cell. Endocrinol. 101, 159–165. doi:10.1016/0303-7207(94)90230-5
Rodriguez, M., Daniels, B., Gunawardene, S., and Robbins, G. K. (2009). High frequency of vitamin D deficiency in ambulatory HIV-positive patients. AIDS Res. Hum. Retroviruses 25, 9–14. doi:10.1089/aid.2008.0183
Rossini, M., Maddali, B. S., La, M. G., Minisola, G., Malavolta, N., Bernini, L., et al. (2010). Vitamin D deficiency in rheumatoid arthritis: prevalence, determinants and associations with disease activity and disability. Arthritis Res. Ther. 12, R216. doi:10.1186/ar3195
Sigmundsdottir, H., Pan, J., Debes, G. F., Alt, C., Habtezion, A., Soler, D., et al. (2007). DCs metabolize sunlight-induced vitamin D3 to ‘program’ T cell attraction to the epidermal chemokine CCL27. Nat. Immunol. 8, 285–293. doi:10.1038/ni1433
Singh, A. K., Wilson, M. T., Hong, S., Olivares-Villagomez, D., Du, C., Stanic, A. K., et al. (2001). Natural killer T cell activation protects mice against experimental autoimmune encephalomyelitis. J. Exp. Med. 194, 1801–1811. doi:10.1084/jem.194.12.1801
Singh, P. K., Doig, C. L., Dhiman, V. K., Turner, B. M., Smiraglia, D. J., and Campbell, M. J. (2012). Epigenetic distortion to VDR transcriptional regulation in prostate cancer cells. J. Steroid Biochem. Mol. Biol. doi:10.1016/j.jsbmb.2012.10.002
Sloka, S., Grant, M., and Newhook, L. A. (2010). The geospatial relation between UV solar radiation and type 1 diabetes in Newfoundland. Acta Diabetol. 47, 73–78. doi:10.1007/s00592-009-0100-0
Smith, K. A. (1988). Interleukin-2: inception, impact, and implications. Science 240, 1169–1176. doi:10.1126/science.3131876
Smith, S. J., Green, L. M., Hayes, M. E., and Mawer, E. B. (1999). Prostaglandin E2 regulates vitamin D receptor expression, vitamin D-24-hydroxylase activity and cell proliferation in an adherent human myeloid leukemia cell line (Ad-HL60). Prostaglandins Other Lipid Mediat. 57, 73–85. doi:10.1016/S0090-6980(98)00073-2
Song, L. N. (1996). Demonstration of vitamin D receptor expression in a human megakaryoblastic leukemia cell line: regulation of vitamin D receptor mRNA expression and responsiveness by forskolin. J. Steroid Biochem. Mol. Biol. 57, 265–274. doi:10.1016/0960-0760(95)00266-9
Spanier, J. A., Nashold, F. E., Olson, J. K., and Hayes, C. E. (2012). The Ifng gene is essential for Vdr gene expression and vitamin D(3)-mediated reduction of the pathogenic T cell burden in the central nervous system in experimental autoimmune encephalomyelitis, a multiple sclerosis model. J. Immunol. 189, 3188–3197. doi:10.4049/jimmunol.1102925
Speeckaert, M., Huang, G., Delanghe, J. R., and Taes, Y. E. (2006). Biological and clinical aspects of the vitamin D binding protein (Gc-globulin) and its polymorphism. Clin. Chim. Acta 372, 33–42. doi:10.1016/j.cca.2006.03.011
Staples, J. A., Ponsonby, A. L., Lim, L. L., and McMichael, A. J. (2003). Ecologic analysis of some immune-related disorders, including type 1 diabetes, in Australia: latitude, regional ultraviolet radiation, and disease prevalence. Environ. Health Perspect. 111, 518–523. doi:10.1289/ehp.5941
Takeuchi, A., Reddy, G. S., Kobayashi, T., Okano, T., Park, J., and Sharma, S. (1998). Nuclear factor of activated T cells (NFAT) as a molecular target for 1alpha,25-dihydroxyvitamin D3-mediated effects. J. Immunol. 160, 209–218.
Torchia, J., Rose, D. W., Inostroza, J., Kamei, Y., Westin, S., Glass, C. K., et al. (1997). The transcriptional co-activator p/CIP binds CBP and mediates nuclear-receptor function. Nature 387, 677–684. doi:10.1038/42652
Towers, T. L., and Freedman, L. P. (1998). Granulocyte-macrophage colony-stimulating factor gene transcription is directly repressed by the vitamin D3 receptor. Implications for allosteric influences on nuclear receptor structure and function by a DNA element. J. Biol. Chem. 273, 10338–10348. doi:10.1074/jbc.273.17.10338
van Leeuwen, J. P., Birkenhager, J. C., Buurman, C. J., van den Bemd, G. J., Bos, M. P., and Pols, H. A. (1992). Bidirectional regulation of the 1,25-dihydroxyvitamin D3 receptor by phorbol ester-activated protein kinase-C in osteoblast-like cells: interaction with adenosine 3’,5’-monophosphate-induced up-regulation of the 1,25-dihydroxyvitamin D3 receptor. Endocrinology 130, 2259–2266. doi:10.1210/en.130.4.2259
van Leeuwen, J. P., Birkenhager, J. C., Schilte, J. P., Buurman, C. J., and Pols, H. A. (1990). Role of calcium and cAMP in heterologous up-regulation of the 1,25-dihydroxyvitamin D3 receptor in an osteoblast cell line. Cell Calcium 11, 281–289. doi:10.1016/0143-4160(90)90005-F
Vidal, M., Ramana, C. V., and Dusso, A. S. (2002). Stat1-vitamin D receptor interactions antagonize 1,25-dihydroxyvitamin D transcriptional activity and enhance stat1-mediated transcription. Mol. Cell. Biol. 22, 2777–2787. doi:10.1128/MCB.22.8.2777-2787.2002
Vieira, V. M., Hart, J. E., Webster, T. F., Weinberg, J., Puett, R., Laden, F., et al. (2010). Association between residences in U.S. northern latitudes and rheumatoid arthritis: a spatial analysis of the Nurses’ Health Study. Environ. Health Perspect. 118, 957–961. doi:10.1289/ehp.0901861
vom, B. E., Zechel, C., Heery, D., Heine, M. J., Garnier, J. M., Vivat, V., et al. (1996). Differential ligand-dependent interactions between the AF-2 activating domain of nuclear receptors and the putative transcriptional intermediary factors mSUG1 and TIF1. EMBO J. 15, 110–124.
von Essen, M. R., Kongsbak, M., Schjerling, P., Olgaard, K., Odum, N., and Geisler, C. (2010). Vitamin D controls T cell antigen receptor signaling and activation of human T cells. Nat. Immunol. 11, 344–349. doi:10.1038/ni.1851
Waterhouse, J. C., Perez, T. H., and Albert, P. J. (2009). Reversing bacteria-induced vitamin D receptor dysfunction is key to autoimmune disease. Ann. N. Y. Acad. Sci. 1173, 757–765. doi:10.1111/j.1749-6632.2009.04637.x
Wood, A. M., Bassford, C., Webster, D., Newby, P., Rajesh, P., Stockley, R. A., et al. (2011). Vitamin D-binding protein contributes to COPD by activation of alveolar macrophages. Thorax 66, 205–210. doi:10.1136/thx.2010.140921
Xu, Y., Xie, J., Li, Y., Yue, J., Chen, J., Chunyu, L., et al. (2003). Using a cDNA microarray to study cellular gene expression altered by Mycobacterium tuberculosis. Chin. Med. J. 116, 1070–1073.
Yenamandra, S. P., Lundin, A., Arulampalam, V., Yurchenko, M., Pettersson, S., Klein, G., et al. (2009). Expression profile of nuclear receptors upon Epstein-Barr virus induced B cell transformation. Exp. Oncol. 31, 92–96.
Yu, S., Bruce, D., Froicu, M., Weaver, V., and Cantorna, M. T. (2008). Failure of T cell homing, reduced CD4/CD8alphaalpha intraepithelial lymphocytes, and inflammation in the gut of vitamin D receptor KO mice. Proc. Natl. Acad. Sci. U.S.A. 105, 20834–20839. doi:10.1073/pnas.0808700106
Yu, S., and Cantorna, M. T. (2008). The vitamin D receptor is required for iNKT cell development. Proc. Natl. Acad. Sci. U.S.A. 105, 5207–5212. doi:10.1073/pnas.0711558105
Zella, L. A., Kim, S., Shevde, N. K., and Pike, J. W. (2006). Enhancers located within two introns of the vitamin D receptor gene mediate transcriptional autoregulation by 1,25-dihydroxyvitamin D3. Mol. Endocrinol. 20, 1231–1247. doi:10.1210/me.2006-0015
Keywords: vitamin D receptor, T cell function, vitamin D, signaling, expression, activity
Citation: Kongsbak M, Levring TB, Geisler C and von Essen MR (2013) The vitamin D receptor and T cell function. Front. Immunol. 4:148. doi: 10.3389/fimmu.2013.00148
Received: 25 March 2013; Accepted: 29 May 2013;
Published online: 18 June 2013.
Edited by:
Karsten Sauer, The Scripps Research Institute, USAReviewed by:
Christopher E. Rudd, University of Cambridge, UKCopyright: © 2013 Kongsbak, Levring, Geisler and von Essen. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Marina Rode von Essen, Department of International Health, Immunology and Microbiology, Faculty of Health and Medical Sciences, University of Copenhagen, Blegdamsvej 3C, DK-2200 Copenhagen, Denmark e-mail:bWVzc2VuQHN1bmQua3UuZGs=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.