
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 30 August 2012
Sec. Inflammation
Volume 3 - 2012 | https://doi.org/10.3389/fimmu.2012.00275
This article is part of the Research Topic The Molecular Mechanisms of Chronic Inflammation Development View all 13 articles
 Masato Kubo1,2* and Yasutaka Motomura1
Masato Kubo1,2* and Yasutaka Motomura1Although the major role of the immune response is host defense from a wide range of potentially pathogenic microorganisms, excess immune responses can result in severe host damage. The host thus requires anti-inflammatory mechanisms to prevent reactivity to self. Interleukin-10 (IL-10) is a cytokine with broad anti-inflammatory properties involved in the pathogenesis of various diseases. IL-10 was originally described as a T helper (TH2) derived cytokine, but further studies indicated that IL-10 is expressed not only by many cells of the adaptive immune system, including T and B cells, but also by the innate immune cells, including dendritic cells (DCs), macrophages, mast cells, and natural killer (NK) cells. In addition, IL-10 can be induced in TH1 and TH17 cells by chronic inflammation as a system of feedback regulation. In this review, we focus on the molecular mechanisms underlying IL10 gene expression in adaptive immune cells and summarize the recent progresses in epigenetic and transcriptional regulation of the IL10 gene. Understanding the transcriptional regulatory events may help in the development of new strategies to control inflammatory diseases.
Recent genome-wide association studies (GWAS) have demonstrated tight association of polymorphisms in the genes encoding IL10 with systemic lupus erythematosis (SLE) (Gateva et al., 2009) and Bechet's disease (BD) (Mizuki et al., 2010; Remmers et al., 2010). BD is a genetically complex disease characterized by recurrent inflammation affecting urogenital mucosa, eye, and skin. Allelic imbalance of the rs158111 variant in pre-mRNA associates with expression of the IL10 gene. The disease associated haplotype results in the reduction of the pre-mRNA transcript and Interleukin-10 (IL-10) production in mononuclear cells activated with lipopolysaccharide (LPS), suggesting that a genetic predisposition for low IL-10 production is a risk factor for BD (Remmers et al., 2010). Polymorphisms in the IL10 gene region have been reported to be associated with ulcerative colitis (UC) (Franke et al., 2008), type I diabetes (Barrett et al., 2009), and severe juvenile rheumatoid arthritis (Crawley et al., 1999), and mutations in the genes encoding the subunits of the IL-10R were found in patients with inflammatory bowel disease (IBD) (Glocker et al., 2009). These observations strongly implicate IL-10 as an important regulator of the human immune system. A wide variety of cells are known to produce IL-10, but it remains unclear which cell type(s) is the major contributor to immune regulation. Therefore, it is important to better understand the source and the regulatory role of IL-10.
For identification of the cellular sources and the role of IL-10, several reporter mouse strains have been established as useful detection tools to track in vivo expression of IL-10 (Bouabe, 2012). These reporter strains often provide critical insight into the expression of IL-10 in various cell types and cell type-specific function. IL-10eYFP mice and IL10-IRES-EGFP mice are classical versions of such IL-10 reporter mice (Calado et al., 2006; Kamanaka et al., 2006; Neves et al., 2010; Bouabe et al., 2011), and reporter activity in these lines has been detected only in CD4 T cells after robust stimulation. Thus these lines have a relatively insensitive limit of detection of IL-10-driven expression of autofluorescent proteins. Improved versions of the reporter mice, IL10-IRES-eGFP-BGHpA mice, IL10Venus mice and IL10-Thy1.1-SV40pA BAC transgenic mice revealed steady expression of IL-10 in a large fraction of CD4+ T cells, including Treg cells and NKT cells, CD19+B220low B cells, CD19+ CD138+plasma cells, and in a very small subset of CD11b+ macrophages, CD11c+ dendritic cells (DCs), and NK1.1+ NK cells (Maynard et al., 2007; Madan et al., 2009; Atarashi et al., 2011). Accumulating evidence thus indicates that IL-10 is secreted by a wide variety of cells, such as helper and regulatory T cells, NKT cells, NK cells, regulatory B cells, macrophages, DCs, and monocytes, all of which may contribute to its immunoregulatory role (Figure 1).
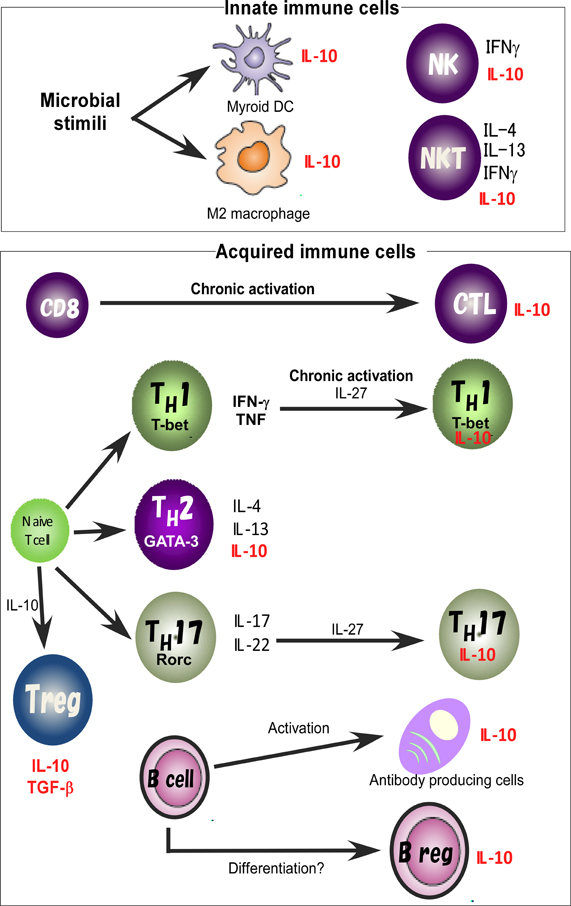
Figure 1. IL-10 expression in the immune system. IL-10 is expressed by M2 macrophages and myeloid DCs. Treg, TH1, TH2, and TH17 cell subsets share the ability to produce IL-10. IL-10 regulates the function and/or IL-10 production of Treg cells. IL-10 production by TH1 cells is induced in chronically infected mice with parasites infection and in response to high-dose antigenic stimulation. IL-27 effectively blocks IL-17 production and induces the production of IL-10. Activated B cells and Bregs are also a key B cell subset responsible for IL-10 mediated regulatory function.
IL-10 is one of the key cytokines to down-regulate a variety of inflammatory responses mediated by lymphoid and myeloid cells (Berg et al., 2001). IL-10 was originally identified as cytokine synthesis inhibitory factor (CSIF). It was secreted from type 2 T-helper cells (TH2 cells) and suppressed the differentiation and effector functions of Th1 cells (Fiorentino et al., 1989). It's inhibitory function was explained by its ability to suppress the production of pro-inflammatory cytokines such as IL-12 and TNF by DCs and macrophages, and to down regulate the expression of MHC II and the costimulatory molecules CD80 and CD86 on antigen presenting cell (APCs), thereby resulting in the subsequent inhibition of T cell activation (Fiorentino et al., 1991; Berg et al., 2001). The in vivo importance of IL-10 in controlling inflammatory responses was originally recognized based on observations made in IL-10-deficient mice, which mount exaggerated TH1 cell responses and develop spontaneous chronic enterocolitis in response to normal gut flora (Kuhn et al., 1993). IL-10-deficient mice die from spontaneous colitis (Kuhn et al., 1993), and this phenotype is partly retained even in mice lacking IL-10 only in Treg cells (Rubtsov et al., 2008). In the colitis case, natural regulatory T (nTreg) cells would be responsible for IL-10 production in response to the microflora. On the other hand, the type 1 regulatory T (Tr1) cell is a different type of Treg cell. Tr1 cells are characterized by their low proliferative capacity and their high levels of IL-10 secretion (Groux et al., 1997). Consistent with IL-10's role in suppressing inflammation, immunization of IL-10 deficient mice with myelin antigens resulted in enhanced neuroinflammation with loss of recovery from experimental autoimmune encephalomyelitis (EAE), a mouse model for human multiple sclerosis (MS) (Bettelli et al., 1998). IL-10 therefore plays an important role in regulating overactive responses that would otherwise result in autoinflammatory disease.
The differentiation of TH1, TH2, and TH17 cells is regulated by distinct signaling pathways. Despite differences in their development, these T cell subsets share the ability to make IL-10, which can be involved in the regulation of immune responses (Figure 1). The mechanism of silencing IL-10 production in TH1 cells was originally thought to be stable and immutable, but it is now clear that TH1 cells can produce IL-10 under certain conditions. IL-10-producing TH1 cells have been shown, on the one hand, to limit immunopathology during Toxplasma gondii infection (Shaw et al., 2006; Jankovic et al., 2007), but on the other hand, to attenuate protective immunity to Leishmania major (Anderson et al., 2007). IL-10 production by TH1 cells has also been reported in animals chronically infected with these parasites and in response to high-dose antigenic stimulation. Such hyperactivation of T cell receptor (TCR)-mediated signaling leads to sustained phosphorylation of ERK1 and ERK2 MAP kinases, resulting in plasticity of IL-10 production in TH1 cells (Saraiva et al., 2009). Continuous and excess antigen stimulation under TH1 skewing conditions enhances the plasticity of IL-10 production (Motomura et al., 2011), and such IL-10 production from TH1 cells may be an effective fail-safe mechanism to maintain immune homeostasis.
The presence of TH17 subsets with regulatory functions correlates with their ability to produce IL-10 (Fitzgerald et al., 2007; McGeachy et al., 2007; Stumhofer et al., 2007; Saraiva et al., 2009). IL-27 added to the cultures under TH17 skewing condition in the presence of TGF-β and IL-6 effectively blocks IL-17 production and induces the production of IL-10. The ability of IL-27 to induce IL-10 is important to suppress TH17 cell–mediated autoimmunity (Fitzgerald et al., 2007; Diveu et al., 2009). Moreover, addition of IL-27 under TH1 skewing conditions also induced the expression of IL-10. This activity was specific in that it had no suppressive effect on the ability of the cells to secrete IFN-γ (Batten et al., 2008). On the other hand, in vitro restimulation of T cells from mice with EAE in the presence of IL-23 generates cells able to efficiently transfer the disease, whereas restimulation in the presence of TGF-β and IL-6, with or without IL-23, generates nonpathogenic IL-10-producing cells (Fitzgerald et al., 2007). IL-23-induced TH17 cells are pathogenic and probably more responsive to IL-27-induced upregulation of IL-10. Therefore, IL-23 and IL-27, which may derive from different subsets of DC (Gafa et al., 2006; Kinnebrew et al., 2012), control the direction of T cell responses to pathogenesis or resistance. In the case of humans, IL-1β can modify the capacity of pathogen-induced human TH17 cells to produce either IFN-γ or IL-10 (Zielinski et al., 2012). Recently, DC and innate cell-derived IL-27 was reported to induce IL-10 production by CD8+ cytotoxic lymphocytes (CTLs) during viral infection (Sun et al., 2011).
In many cases, DCs possess unique functions to control the plasticity of T cell function, and the lectin-like receptors (LLR) on DCs contribute to alteration of the quality of the T cell response. Indeed, activation of DCs with Dectin-1 results in polarization into a TH17 response (Gringhuis et al., 2009), but Dectin-1 has also been reported to induce IL-10 production by DCs (Rogers et al., 2005). Dectin-1 in conjunction with TLR2 elicits DCs capable of inducing Treg responses (Dillon et al., 2006). Another LLR, DC-SIGN, leads to not only a TH2 response (Geurtsen et al., 2009) and but also to Treg differentiation (Zhou et al., 2010). The scavenger receptor, DC-asialoglycoprotein receptor (DC-ASGPR) leads to generation of antigen specific suppressive IL-10-producing T cells via the induction of IL-10 production by DCs (Li et al., 2012). These results and others make it abundantly clear that the regulation of T cell subset differentiation and cytokine production is complex and occurs at multiple levels.
In previous studies, involvement of various transcription factors in activation of the IL10 gene has been proposed (Figure 2). The TH2 master regulator, GATA3 controls IL-10 expression in TH2 cells through initiating changes of the chromatin structure at the IL10 locus (Shoemaker et al., 2006; Chang et al., 2007). Indeed, Gata3 deficiency cause a loss of IL-10 expression in TH2 cells (unpublished data). GATA3 binds to the IL10 promoter, but GATA3 alone does not activate the promoter activity (Shoemaker et al., 2006), supporting the idea that GATA3 may determine the transcriptional permissibility of the chromatin structure in the IL10 locus. However, in the case of other T cell types such as TH1 and TH17 that express a quite low level of GATA3, other factors would be necessary to replace its function to induce high levels of IL-10 expression (Jankovic et al., 2007; Saraiva et al., 2009).
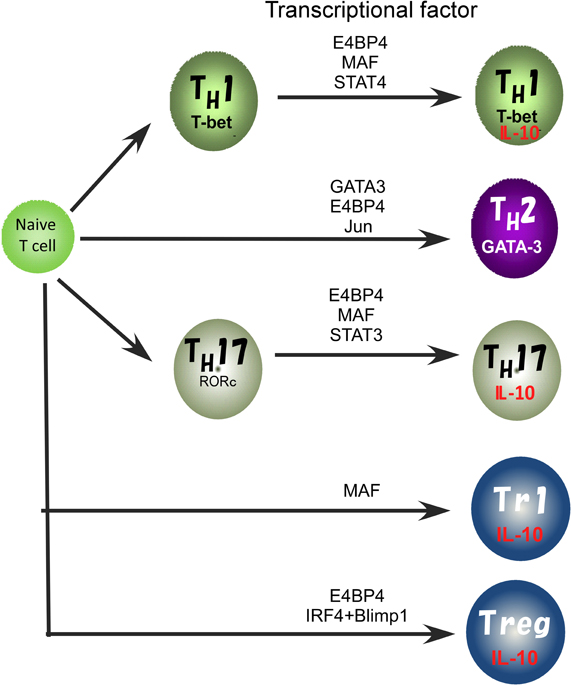
Figure 2. Transcriptional factors regulating the IL10 gene expression in Treg, TH1, TH2, and TH17 cell subsets.
We recently reported that E4BP4 has multiple functions in cytokine gene regulation and is an essential transcriptional factor to regulate IL-10 production not only in TH2, NKT, and Treg cells, but also to allow plasticity of IL-10 production in TH1 and TH17 cells (Motomura et al., 2011). E4BP4 was originally identified as a negative regulator of the mammalian circadian oscillatory system by virtue of its antagonistic binding to the same DNA regulatory sequences as a member of the PAR family of bZIP transcription factors (Cowell, 2002). The deletion of the E4bp4 gene preferentially affected NK cell development (Gascoyne et al., 2009; Kamizono et al., 2009). By contrast, the proportion of conventional helper T cell lineages that developed in the culture conditions for TH1, TH2, iTreg, and TH17 cell induction was not affected in the E4bp4 deficient mice. E4BP4 can induce IL-10 production under Gata3 deficient TH1 conditions, suggesting that GATA-3 is dispensable for plasticity of IL-10 production by TH1 cells. E4BP4 bound to intron 4 and the 3′ non-coding region of the IL10 locus and activated the histone code in the IL10 locus, as indicated by the finding that H3K9 methylation and H3K14 acetylation were completely abolished in E4bp4-deficient TH2 cells. These findings indicate that E4BP4 is an epigenetic regulator to control the permissive status of the IL10 locus (Motomura et al., 2011).
Another TH2 cell-dominant transcription factor, MAF, can bind to the IL10 promoter and plays a role in the regulation of IL-10 production in mouse macrophages stimulated with LPS and IL-4 (Cao et al., 2005). MAF is detectable in TH1, TH2 and TH17 cells, where its expression coincides with IL-10 production. IL-27 induces the expression of MAF along with IL-21 and the costimulatory receptor ICOS, and these ultimately act coordinately to promote differentiation of IL-10-producing Tr1 cells (Pot et al., 2009). Moreover, induction of MAF expression in TH1 and TH17 cells depends on ERK activation, as does IL-10 expression (Saraiva et al., 2009). These reports suggest that MAF may be a common regulator for IL-10 production in both the innate and the adaptive immune systems. However, MAF alone is not sufficient to induce IL10 expression in macrophages and T cells (Cao et al., 2005; Motomura et al., 2011), thus the role of MAF in IL-10 production is still controversial.
Distinct mechanisms seem to regulate the expression of IL10 in the innate and acquired immune systems. NF-κB activation is a major contributor to IL-10 production in macrophages, and the NF-κB p65 subunit binds to the IL10 locus 4.5 kb upstream of the transcription start site (Saraiva et al., 2005). This finding is consistent with the report that IKK2-deficient mice show a defect in IL-10 production by LPS-stimulated macrophages. On the other hand, ERK signaling is required for optimal IL-10 induction in innate immune cells as well as T cells (Agrawal et al., 2003; Kaiser et al., 2009; Saraiva et al., 2009). ERK activation leads to the binding of AP-1 to the IL10 locus through the cooperative function of Fos/Jun family proteins. Studies have suggested a role for Jun proteins in regulating IL10 expression in TH2 cells, but not in TH1 cells, and this is explained by their binding to a regulatory element located 6.45 kb downstream of the IL10 transcription start site (Jones and Flavell, 2005; Wang et al., 2005). This report addresses the idea that ERK and MAF are required for IL10 induction as common regulators in various cell types (Saraiva and O'Garra, 2010).
STAT proteins are reported to be another mechanism regulating IL-10 expression by both macrophages and T cells. In T cells, the induction of IL-10 by IL-27 seems to depend on both STAT1 and STAT3 (Stumhofer et al., 2007; Batten et al., 2008; Xu et al., 2009). STAT3 is also involved in IL-6-induced IL-10 expression (Stumhofer et al., 2007). Moreover, STAT3 is responsible for the IL-27-mediated IL-10 induction instead of the inhibition of TH17 cell differentiation. The binding of STAT3 to the IL10 promoter in human cell lines induced IL10 transcription though STAT3-dependent promoter activation in conjunction with IFN-α induced IRF1 activation (Ziegler-Heitbrock et al., 2003). STAT4, which is important for the expression of IFN-γ, also regulates IL-10 production in TH1 cells (Saraiva et al., 2009), and STAT4 was also reported to have a role in inducing its expression in NK cells (Grant et al., 2008). However, the STATs are also important in the differentiation process of TH cell subsets, thus it remains unclear whether the function of STATs in IL-10 production is direct or indirect.
Recently, the coordinated activity of IRF4 and Blimp-1 has been reported to be critical for regulation of IL-10 production by Treg cells (Cretney et al., 2011). Blimp-1 is a transcriptional repressor well known for its role in promoting the differentiation of plasma cells (Nutt et al., 2008) but is also required for the maintenance of T cell homeostasis. Mice lacking Blimp-1 specifically in T cells accumulate activated T cells and develop immune pathology, including colitis and lung inflammation (Kallies et al., 2006; Martins et al., 2006), suggesting that Blimp-1 has a critical role in Treg cell function. Indeed, Blimp-1 is required for IL-10 production and high ICOS expression in mature effector Treg cells. Blimp-1 is known to be preferentially expressed in TH2 cells that produce high levels of IL-10, and IL-10 production by CTLs is regulated by a Blimp-1 dependent mechanism (Sun et al., 2011). Therefore, Blimp-1 is an important regulator of IL10 expression.
IL-10- producing B cells play an important role in controlling autoimmunity, such as in EAE, an animal model of MS, and in a systemic lupus erythematosus (SLE)-like disease that develops in the Lyn-deficient mouse and during murine cytomegalovirus (MCMV4) infection (Fillatreau et al., 2002; Madan et al., 2009; Scapini et al., 2011). This idea has its origins in the discovery that B cell deficient μMT mice cannot recover from EAE (Wolf et al., 1996), and the identification of a regulatory role of IL-10 producing B cells in EAE lead to the realization of the importance of this type of B cell (Fillatreau et al., 2002). Furthermore, the importance of IL-10–producing B cells in controlling chronic inflammatory diseases has also been demonstrated in collagen-induced arthritis and chronic intestinal inflammation (Mizoguchi et al., 2002; Mauri et al., 2003). On the other hand, IL-10 production by B cells has been shown to prevent protective immunity to infection with Salmonella Typhimurium (Neves et al., 2010) and to decrease MCMV4-specific CD8+ T cell responses (Madan et al., 2009).
IL-10 is secreted from several B cell subsets that can be distinguished by cell surface phenotype. Regulatory B cells (Bregs) are considered as a key B cell subset responsible for IL-10 mediated regulatory function (Fillatreau et al., 2002; Mizoguchi et al., 2002; Mauri et al., 2003). However, there is no precise marker that exclusively defines the Breg (Mauri and Bosma, 2012). Transitional 2 marginal zone precursor (T2-MZP) B cells (CD19+CD21hiCD23hiCD24hiIgDhiIgMhiCD1hi) appear to be the most likely candidate for being Breg cells. T2-MZP B cells isolated from arthritic mice produced copious amount of IL-10 after stimulation with collagen type II antigen in conjunction with anti-CD40 antibody. Transfer of T2-MZP B cells suppressed the development of CIA (Evans et al., 2007). T2-MZP B cells also suppress other autoimmune diseases including antigen-induced arthritis (AIA) and lupus (Inoue et al., 2006; Carter et al., 2011), and Schistosoma mansoni infection generates IL-10 producing T2-MZP B cells with regulatory function (Amu et al., 2010). However, T2-MZP B cells do not fully satisfy of the complete phenotype of Breg cells, because T2-MZP B cells also contain the largest fraction of transitional immature B cells.
CD5+ B1 B cells are also known to be distinct source of IL-10 (O'Garra et al., 1992), and they also have an immunoregulatory function by killing of CD4 T cells by FasL/Fas-dependent mechanisms (Lundy and Fox, 2009). B1 B cells also play a role in protection from colitis, but their protective role is not due to the production of IL-10, instead IgM and IgA are responsible for the protection (Shimomura et al., 2008).
In studies to understand the mechanisms underlying control of IL10 expression in B cells, the importance of TLRs has been emphasized. In vitro stimulation with LPS, together with PMA and ionomycin, promotes the development of IL-10 producing B cells. There is a rare population expressing CD5 and CD1dhi termed IL-10 producing B cells (B10 cells) that suppresses oxazolone-induced contact hypersensitivity (Yanaba et al., 2008). Stimulation of B cells in vitro with LPS induces the expression of IL10 in plasma cells (CD19+CD138+), but only very low levels of IL-10 in the B10 population (Neves et al., 2010). On the other hand, TLR2 and TLR4 signaling predominantly controls IL-10 production in MZ B cells, but not in follicular B cells (Gray et al., 2007). The importance of IL-10 from B cells is confirmed in mice lacking TLR2, TLR4 or Myd88, a signaling molecule downstream of TLRs, which develop a chronic form of EAE (Lampropoulou et al., 2008), and expression of MyD88 is required for IL-10 production and inhibitory function of B cells. The importance of TLR9, which is a receptor for CpG, has also been proposed in regulating IL-10 production by B cells (Barr et al., 2007). However, TLR9- deficient B cells can still inhibit EAE, suggesting a redundant role of TLR9 in Breg cells (Lampropoulou et al., 2008).
Multiple studies have indicated an essential function of the CD40-CD154 interaction for the activation of Breg cells. Mice lacking CD40 on B cells develop severe EAE, with increased induction of encephalitogenic TH1 and TH17 responses, and have a profound decrease in IL-10 production (Mizoguchi et al., 2000). Furthermore, administration of an agonistic antibody against CD40 improves arthritis by the provision of IL-10 (Mauri et al., 2003; Evans et al., 2007) and IL-10 mediated TH1 inhibition (Mauri et al., 2000).
Several previous studies have suggested that Bregs require signaling through the B cell receptor (BCR) for their activity. Indeed, mice lacking the BCR co-receptor molecule CD19 develop a severe EAE, suggesting the importance of the BCR in the generation of Bregs (Sato et al., 1996; Yanaba et al., 2008). A major component downstream of BCR signaling is intracellular Ca2+ (Feske, 2007). Mice lacking the Ca2+ channel molecules, STIM1 and STIM2, develop an elaborated EAE and low numbers of IL-10 producing Bregs (Matsumoto et al., 2011). These mice showed normal B cell development and antibody responses, however, they have a defect in the activation of nuclear factor of activated T cells (NFAT). Inconsistently, mice with an NFATc1 deficiency display an increased number of IL-10 producing B cells and development of milder EAE (Bhattacharyya et al., 2011). Therefore, at present, it remains unclear which signaling pathway is the major one.
Recent GWAS have demonstrated tight association of polymorphisms in the gene encoding IL10 with several inflammatory diseases, indicating importance of understanding the source and the regulation of IL-10. IL-10 is expressed by many acquired immune cells including, TH1, TH2 and TH17 cells, Treg cells, CD8+ T cells and B cells. In this review, we have summarized the current view of IL10 regulation in these T cells and B cells. IL10 expression in different T cell subsets is regulated by a complex of multiple transcriptional factors, such as GATA-3, E4BP4, MAF, Blimp1, and so on, and these multiple levels of regulation occur at independent differentiation stages in accordance with certain rules that are just beginning to be understood. Therefore, the expression of the IL10 gene has the flexibility and plasticity. This plasticity is sometimes controlled by continuous antigen stimulation or by a particular cytokine environment, such as DC derived IL-27, and selectively occurs in inflammatory type of helper T cells, TH1 and TH17 cells. On the other hand, the importance of IL-10 from B cells is proposed in several autoimmune and inflammatory disease models. Similar to T cell, a complex of multiple signaling pathways, including BCR and CD40-CD40L pathways, is required for the generation of Bregs and the induction of IL-10 in B cells. However, it remains virtually unknown at present which signaling pathway is the major one and how IL10 is transcriptionally regulated. There may be critical transcriptional regulatory machinery that could be specific to certain cell types and this machinery may be turned on by specific signals in certain diseases. Therefore, it will be quite important to understand the meaning of the SNPs that associate with several inflammatory diseases and the molecular mechanisms underlying transcriptional regulation of the IL10 gene. Moreover, this approach may lead to the development of innovative therapeutic strategies for controlling these diseases.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Agrawal, S., Agrawal, A., Doughty, B., Gerwitz, A., Blenis, J., Van Dyke, T., and Pulendran, B. (2003). Cutting edge: different Toll-like receptor agonists instruct dendritic cells to induce distinct Th responses via differential modulation of extracellular signal-regulated kinase-mitogen-activated protein kinase and c-Fos. J. Immunol. 171, 4984–4989.
Amu, S., Saunders, S. P., Kronenberg, M., Mangan, N. E., Atzberger, A., and Fallon, P. G. (2010). Regulatory B cells prevent and reverse allergic airway inflammation via FoxP3-positive T regulatory cells in a murine model. J. Allergy Clin. Immunol. 125, 1114–1124.
Anderson, C. F., Oukka, M., Kuchroo, V. J., and Sacks, D. (2007). CD4(+)CD25(−)Foxp3(−) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J. Exp. Med. 204, 285–297.
Atarashi, K., Tanoue, T., Shima, T., Imaoka, A., Kuwahara, T., Momose, Y., Cheng, G., Yamasaki, S., Saito, T., Ohba, Y., Taniguchi, T., Takeda, K., Hori, S., Ivanov, I. I., Umesaki, Y., Itoh, K., and Honda, K. (2011). Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341.
Barr, T. A., Brown, S., Ryan, G., Zhao, J., and Gray, D. (2007). TLR-mediated stimulation of APC: distinct cytokine responses of B cells and dendritic cells. Eur. J. Immunol. 37, 3040–3053.
Barrett, J. C., Clayton, D. G., Concannon, P., Akolkar, B., Cooper, J. D., Erlich, H. A., Julier, C., Morahan, G., Nerup, J., Nierras, C., Plagnol, V., Pociot, F., Schuilenburg, H., Smyth, D. J., Stevens, H., Todd, J. A., Walker, N. M., and Rich, S. S. (2009). Genome-wide association study and meta-analysis find that over 40 loci affect risk of type 1 diabetes. Nat. Genet. 41, 703–707.
Batten, M., Kljavin, N. M., Li, J., Walter, M. J., de Sauvage, F. J., and Ghilardi, N. (2008). Cutting edge: IL-27 is a potent inducer of IL-10 but not FoxP3 in murine T cells. J. Immunol. 180, 2752–2756.
Berg, D. J., Zhang, J., Lauricella, D. M., and Moore, S. A. (2001). Il-10 is a central regulator of cyclooxygenase-2 expression and prostaglandin production. J. Immunol. 166, 2674–2680.
Bettelli, E., Das, M. P., Howard, E. D., Weiner, H. L., Sobel, R. A., and Kuchroo, V. K. (1998). IL-10 is critical in the regulation of autoimmune encephalomyelitis as demonstrated by studies of IL-10- and IL-4-deficient and transgenic mice. J. Immunol. 161, 3299–3306.
Bhattacharyya, S., Deb, J., Patra, A. K., Thuy Pham, D. A., Chen, W., Vaeth, M., Berberich-Siebelt, F., Klein-Hessling, S., Lamperti, E. D., Reifenberg, K., Jellusova, J., Schweizer, A., Nitschke, L., Leich, E., Rosenwald, A., Brunner, C., Engelmann, S., Bommhardt, U., Avots, A., Muller, M. R., Kondo, E., and Serfling, E. (2011). NFATc1 affects mouse splenic B cell function by controlling the calcineurin–NFAT signaling network. J. Exp. Med. 208, 823–839.
Bouabe, H. (2012). Cytokine reporter mice: the special case of IL-10 scand. J. Immunol. 75, 553–567.
Bouabe, H., Liu, Y., Moser, M., Bosl, M. R., and Heesemann, J. (2011). Novel highly sensitive IL-10-β-Lactamase reporter mouse reveals cells of the innate immune system as a substantial source of IL-10 in vivo. J. Immunol. 187, 3165–3176.
Calado, D. P., Paixao, T., Holmberg, D., and Haury, M. (2006). Stochastic monoallelic expression of IL-10 in T cells. J. Immunol. 177, 5358–5364.
Cao, S., Liu, J., Song, L., and Ma, X. (2005). The protooncogene c-Maf is an essential transcription factor for IL-10 gene expression in macrophages. J. Immunol. 174, 3484–3492.
Carter, N. A., Vasconcellos, R., Rosser, E. C., Tulone, C., Munoz-Suano, A., Kamanaka, M., Ehrenstein, M. R., Flavell, R. A., and Mauri, C. (2011). Mice lacking endogenous IL-10-producing regulatory B cells develop exacerbated disease and present with an increased frequency of Th1/Th17 but a decrease in regulatory T cells. J. Immunol. 186, 5569–5579.
Chang, H. D., Helbig, C., Tykocinski, L., Kreher, S., Koeck, J., Niesner, U., and Radbruch, A. (2007). Expression of IL-10 in Th memory lymphocytes is conditional on IL-12 or IL-4, unless the IL-10 gene is imprinted by GATA-3. Eur. J. Immunol. 37, 807–817.
Cowell, I. G. (2002). E4BP4/NFIL3, a PAR-related bZIP factor with many roles. Bioessays 24, 1023–1029.
Crawley, E., Kay, R., Sillibourne, J., Patel, P., Hutchinson, I., and Woo, P. (1999). Polymorphic haplotypes of the interleukin-10 5′ flanking region determine variable interleukin-10 transcription and are associated with particular phenotypes of juvenile rheumatoid arthritis. Arthritis Rheum. 42, 1101–1108.
Cretney, E., Xin, A., Shi, W., Minnich, M., Masson, F., Miasari, M., Belz, G. T., Smyth, G. K., Busslinger, M., Nutt, S. L., and Kallies, A. (2011). The transcription factors Blimp-1 and IRF4 jointly control the differentiation and function of effector regulatory T cells. Nat. Immunol. 12, 304–311.
Dillon, S., Agrawal, S., Banerjee, K., Letterio, J., Denning, T. L., Oswald-Richter, K., Kasprowicz, D. J., Kellar, K., Pare, J., van Dyke, T., Ziegler, S., Unutmaz, D., and Pulendran, B. (2006). Yeast zymosan, a stimulus for TLR2 and dectin-1, induces regulatory antigen-presenting cells and immunological tolerance. J. Clin. Invest. 116, 916–928.
Diveu, C., McGeachy, M. J., Boniface, K., Stumhofer, J. S., Sathe, M., Joyce-Shaikh, B., Chen, Y., Tato, C. M., McClanahan, T. K., de Waal Malefyt, R., Hunter, C. A., Cua, D. J., and Kastelein, R. A. (2009). IL-27 blocks RORc expression to inhibit lineage commitment of Th17 cells. J. Immunol. 182, 5748–5756.
Evans, J. G., Chavez-Rueda, K. A., Eddaoudi, A., Meyer-Bahlburg, A., Rawlings, D. J., Ehrenstein, M. R., and Mauri, C. (2007). Novel suppressive function of transitional 2 B cells in experimental arthritis. J. Immunol. 178, 7868–7878.
Feske, S. (2007). Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 7, 690–702.
Fillatreau, S., Sweenie, C. H., McGeachy, M. J., Gray, D., and Anderton, S. M. (2002). B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 3, 944–950.
Fiorentino, D. F., Bond, M. W., and Mosmann, T. R. (1989). Two types of mouse T helper cell. IV. Th2 clones secrete a factor that inhibits cytokine production by Th1 clones. J. Exp. Med. 170, 2081–2095.
Fiorentino, D. F., Zlotnik, A., Vieira, P., Mosmann, T. R., Howard, M., Moore, K. W., and O'Garra, A. (1991). IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J. Immunol. 146, 3444–3451.
Fitzgerald, D. C., Zhang, G. X., El-Behi, M., Fonseca-Kelly, Z., Li, H., Yu, S., Saris, C. J., Gran, B., Ciric, B., and Rostami, A. (2007). Suppression of autoimmune inflammation of the central nervous system by interleukin 10 secreted by interleukin 27-stimulated T cells. Nat. Immunol. 8, 1372–1379.
Franke, A., Balschun, T., Karlsen, T. H., Sventoraityte, J., Nikolaus, S., Mayr, G., Domingues, F. S., Albrecht, M., Nothnagel, M., Ellinghaus, D., Sina, C., Onnie, C. M., Weersma, R. K., Stokkers, P. C., Wijmenga, C., Gazouli, M., Strachan, D., McArdle, W. L., Vermeire, S., Rutgeerts, P., Rosenstiel, P., Krawczak, M., Vatn, M. H., Mathew, C. G., and Schreiber, S. (2008). Sequence variants in IL10, ARPC2 and multiple other loci contribute to ulcerative colitis susceptibility. Nat. Genet. 40, 1319–1323.
Gafa, V., Lande, R., Gagliardi, M. C., Severa, M., Giacomini, E., Remoli, M. E., Nisini, R., Ramoni, C., Di Francesco, P., Aldebert, D., Grillot, R., and Coccia, E. M. (2006). Human dendritic cells following Aspergillus fumigatus infection express the CCR7 receptor and a differential pattern of interleukin-12 (IL-12), IL-23, and IL-27 cytokines, which lead to a Th1 response. Infect. Immun. 74, 1480–1489.
Gascoyne, D. M., Long, E., Veiga-Fernandes, H., de Boer, J., Williams, O., Seddon, B., Coles, M., Kioussis, D., and Brady, H. J. (2009). The basic leucine zipper transcription factor E4BP4 is essential for natural killer cell development. Nat. Immunol. 10, 1118–1124.
Gateva, V., Sandling, J. K., Hom, G., Taylor, K. E., Chung, S. A., Sun, X., Ortmann, W., Kosoy, R., Ferreira, R. C., Nordmark, G., Gunnarsson, I., Svenungsson, E., Padyukov, L., Sturfelt, G., Jonsen, A., Bengtsson, A. A., Rantapaa-Dahlqvist, S., Baechler, E. C., Brown, E. E., Alarcon, G. S., Edberg, J. C., Ramsey-Goldman, R., McGwin, G. Jr., Reveille, J. D., Vila, L. M., Kimberly, R. P., Manzi, S., Petri, M. A., Lee, A., Gregersen, P. K., Seldin, M. F., Ronnblom, L., Criswell, L. A., Syvanen, A. C., Behrens, T. W., and Graham, R. R. (2009). A large-scale replication study identifies TNIP1, PRDM1, JAZF1, UHRF1BP1 and IL10 as risk loci for systemic lupus erythematosus. Nat. Genet. 41, 1228–1233.
Geurtsen, J., Chedammi, S., Mesters, J., Cot, M., Driessen, N. N., Sambou, T., Kakutani, R., Ummels, R., Maaskant, J., Takata, H., Baba, O., Terashima, T., Bovin, N., Vandenbroucke-Grauls, C. M., Nigou, J., Puzo, G., Lemassu, A., Daffe, M., and Appelmelk, B. J. (2009). Identification of mycobacterial alpha-glucan as a novel ligand for DC-SIGN: involvement of mycobacterial capsular polysaccharides in host immune modulation. J. Immunol. 183, 5221–5231.
Glocker, E. O., Kotlarz, D., Boztug, K., Gertz, E. M., Schaffer, A. A., Noyan, F., Perro, M., Diestelhorst, J., Allroth, A., Murugan, D., Hatscher, N., Pfeifer, D., Sykora, K. W., Sauer, M., Kreipe, H., Lacher, M., Nustede, R., Woellner, C., Baumann, U., Salzer, U., Koletzko, S., Shah, N., Segal, A. W., Sauerbrey, A., Buderus, S., Snapper, S. B., Grimbacher, B., and Klein, C. (2009). Inflammatory bowel disease and mutations affecting the interleukin-10 receptor. N. Engl. J. Med. 361, 2033–2045.
Grant, L. R., Yao, Z. J., Hedrich, C. M., Wang, F., Moorthy, A., Wilson, K., Ranatunga, D., and Bream, J. H. (2008). Stat4-dependent, T-bet-independent regulation of IL-10 in NK cells. Genes Immun. 9, 316–327.
Gray, M., Miles, K., Salter, D., Gray, D., and Savill, J. (2007). Apoptotic cells protect mice from autoimmune inflammation by the induction of regulatory B cells. Proc. Natl. Acad. Sci. U.S.A. 104, 14080–14085.
Gringhuis, S. I., den Dunnen, J., Litjens, M., van der Vlist, M., Wevers, B., Bruijns, S. C., and Geijtenbeek, T. B. (2009). Dectin-1 directs T helper cell differentiation by controlling noncanonical NF-kappaB activation through Raf-1 and Syk. Nat. Immunol. 10, 203–213.
Groux, H., O'Garra, A., Bigler, M., Rouleau, M., Antonenko, S., de Vries, J. E., and Roncarolo, M. G. (1997). A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 389, 737–742.
Inoue, S., Leitner, W. W., Golding, B., and Scott, D. (2006). Inhibitory effects of B cells on antitumor immunity. Cancer Res. 66, 7741–7747.
Jankovic, D., Kullberg, M. C., Feng, C. G., Goldszmid, R. S., Collazo, C. M., Wilson, M., Wynn, T. A., Kamanaka, M., Flavell, R. A., and Sher, A. (2007). Conventional T-bet(+)Foxp3(-) Th1 cells are the major source of host-protective regulatory IL-10 during intracellular protozoan infection. J. Exp. Med. 204, 273–283.
Jones, E. A., and Flavell, R. A. (2005). Distal enhancer elements transcribe intergenic RNA in the IL-10 family gene cluster. J. Immunol. 175, 7437–7446.
Kaiser, F., Cook, D., Papoutsopoulou, S., Rajsbaum, R., Wu, X., Yang, H. T., Grant, S., Ricciardi-Castagnoli, P., Tsichlis, P. N., Ley, S. C., and O'Garra, A. (2009). TPL-2 negatively regulates interferon-beta production in macrophages and myeloid dendritic cells. J. Exp. Med. 206, 1863–1871.
Kallies, A., Hawkins, E. D., Belz, G. T., Metcalf, D., Hommel, M., Corcoran, L. M., Hodgkin, P. D., and Nutt, S. L. (2006). Transcriptional repressor Blimp-1 is essential for T cell homeostasis and self-tolerance. Nat. Immunol. 7, 466–474.
Kamanaka, M., Kim, S. T., Wan, Y. Y., Sutterwala, F. S., Lara-Tejero, M., Galán, J. E., Harhaj, E., and Flavell, R. A. (2006). Expression of interleukin-10 in intestinal lymphocytes detected by an interleukin-10 reporter knockin tiger mouse. Immunity 25, 941–952.
Kamizono, S., Duncan, G. S., Seidel, M. G., Morimoto, A., Hamada, K., Grosveld, G., Akashi, K., Lind, E. F., Haight, J. P., Ohashi, P. S., Look, A. T., and Mak, T. W. (2009). Nfil3/E4bp4 is required for the development and maturation of NK cells in vivo. J. Exp. Med. 206, 2977–2986.
Kinnebrew, M. A., Buffie, C. G., Diehl, G. E., Zenewicz, L. A., Leiner, I., Hohl, T. M., Flavell, R. A., Littman, D. R., and Pamer, E. G. (2012). Interleukin 23 production by intestinal CD103(+)CD11b(+) dendritic cells in response to bacterial flagellin enhances mucosal innate immune defense. Immunity 36, 276–287.
Kuhn, R., Lohler, J., Rennick, D., Rajewsky, K., and Muller, W. (1993). Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263–274.
Lampropoulou, V., Hoehlig, K., Roch, T., Neves, P., Calderon Gomez, E., Sweenie, C. H., Hao, Y., Freitas, A. A., Steinhoff, U., Anderton, S. M., and Fillatreau, S. (2008). TLR-activated B cells suppress T cell-mediated autoimmunity. J. Immunol. 180, 4763–4773.
Li, D., Romain, G., Flamar, A. L., Duluc, D., Dullaers, M., Li, X. H., Zurawski, S., Bosquet, N., Palucka, A. K., Le Grand, R., O'Garra, A., Zurawski, G., Banchereau, J., and Oh, S. (2012). Targeting self- and foreign antigens to dendritic cells via DC-ASGPR generates IL-10-producing suppressive CD4+ T cells. J. Exp. Med. 209, 109–121.
Lundy, S. K., and Fox, D. A. (2009). Reduced Fas ligand-expressing splenic CD5+ B lymphocytes in severe collagen-induced arthritis. Arthritis Res. Ther. 11, R128.
Madan, R., Demircik, F., Surianarayanan, S., Allen, J. L., Divanovic, S., Trompette, A., Yogev, N., Gu, Y., Khodoun, M., Hildeman, D., Boespflug, N., Fogolin, M. B., Gröbe, L., Greweling, M., Finkelman, F. D., Cardin, R., Mohrs, M., Müller, W., Waisman, A., Roers, A., and Karp, C. L. (2009). Nonredundant roles for B cell-derived IL-10 in immune counter-regulation. J. Immunol. 183, 2312–2320.
Martins, G. A., Cimmino, L., Shapiro-Shelef, M., Szabolcs, M., Herron, A., Magnusdottir, E., and Calame, K. (2006). Transcriptional repressor Blimp-1 regulates T cell homeostasis and function. Nat. Immunol. 7, 457–465.
Matsumoto, M., Fujii, Y., Baba, A., Hikida, M., Kurosaki, T., and Baba, Y. (2011). The calcium sensors STIM1 and STIM2 control B cell regulatory function through interleukin-10 production. Immunity 34, 703–714.
Mauri, C., and Bosma, A. (2012). Immune regulatory function of B cells. Annu. Rev. Immunol. 30, 221–241.
Mauri, C., Gray, D., Mushtaq, N., and Londei, M. (2003). Prevention of arthritis by interleukin 10-producing B cells. J. Exp. Med. 197, 489–501.
Mauri, C., Mars, L. T., and Londei, M. (2000). Therapeutic activity of agonistic monoclonal antibodies against CD40 in a chronic autoimmune inflammatory process. Nat. Med. 6, 673–679.
Maynard, C. L., Harrington, L. E., Janowski, K. M., Oliver J. R., Zindl, C. L., Rudensky, A. Y., and Weaver, C. T. (2007). Regulatory T cells expressing interleukin 10 develop from Foxp3+ and Foxp3-precursor cells in the absence of interleukin 10. Nat Immunol. 8, 931–941.
McGeachy, M. J., Bak-Jensen, K. S., Chen, Y., Tato, C. M., Blumenschein, W., McClanahan, T., and Cua, D. J. (2007). TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat. Immunol. 8, 1390–1397.
Mizoguchi, A., Mizoguchi, E., Takedatsu, H., Blumberg, R. S., and Bhan, A. K. (2002). Chronic intestinal inflammatory condition generates IL-10-producing regulatory B cell subset characterized by CD1d upregulation. Immunity 16, 219–230.
Mizoguchi, E., Mizoguchi, A., Preffer, F. I., and Bhan, A. K. (2000). Regulatory role of mature B cells in a murine model of inflammatory bowel disease. Int. Immunol. 12, 597–605.
Mizuki, N., Meguro, A., Ota, M., Ohno, S., Shiota, T., Kawagoe, T., Ito, N., Kera, J., Okada, E., Yatsu, K., Song, Y. W., Lee, E. B., Kitaichi, N., Namba, K., Horie, Y., Takeno, M., Sugita, S., Mochizuki, M., Bahram, S., Ishigatsubo, Y., and Inoko, H. (2010). Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behcet's disease susceptibility loci. Nat. Genet. 42, 703–706.
Motomura, Y., Kitamura, H., Hijikata, A., Matsunaga, Y., Matsumoto, K., Inoue, H., Atarashi, K., Hori, S., Watarai, H., Zhu, J., Taniguchi, M., and Kubo, M. (2011). The transcription factor E4BP4 regulates the production of IL-10 and IL-13 in CD4+ T cells. Nat. Immunol. 12, 450–459.
Neves, P., Lampropoulou, V., Calderon-Gomez, E., Roch, T., Stervbo, U., Shen, P., Kuhl, A. A., Loddenkemper, C., Haury, M., Nedospasov, S. A., Kaufmann, S. H., Steinhoff, U., Calado, D. P., and Fillatreau, S. (2010). Signaling via the MyD88 adaptor protein in B cells suppresses protective immunity during Salmonella typhimurium infection. Immunity 33, 777–790.
Nutt, S. L., Kallies, A., and Belz, G. T. (2008). Blimp-1 connects the intrinsic and extrinsic regulation of T cell homeostasis. J. Clin. Immunol. 28, 97–106.
O'Garra, A., Chang, R., Go, N., Hastings, R., Haughton, G., and Howard, M. (1992). Ly-1 B (B-1) cells are the main source of B cell-derived interleukin 10. Eur. J. Immunol. 22, 711–717.
Pot, C., Jin, H., Awasthi, A., Liu, S. M., Lai, C. Y., Madan, R., Sharpe, A. H., Karp, C. L., Miaw, S. C., Ho, I. C., and Kuchroo, V. K. (2009). Cutting edge: IL-27 induces the transcription factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote differentiation of IL-10-producing Tr1 cells. J. Immunol. 183, 797–801.
Remmers, E. F., Cosan, F., Kirino, Y., Ombrello, M. J., Abaci, N., Satorius, C., Le, J. M., Yang, B., Korman, B. D., Cakiris, A., Aglar, O., Emrence, Z., Azakli, H., Ustek, D., Tugal-Tutkun, I., Akman-Demir, G., Chen, W., Amos, C. I., Dizon, M. B., Kose, A. A., Azizlerli, G., Erer, B., Brand, O. J., Kaklamani, V. G., Kaklamanis, P., Ben-Chetrit, E., Stanford, M., Fortune, F., Ghabra, M., Ollier, W. E., Cho, Y. H., Bang, D., O'Shea, J., Wallace, G. R., Gadina, M., Kastner, D. L., and Gul, A. (2010). Genome-wide association study identifies variants in the MHC class I, IL10, and IL23R-IL12RB2 regions associated with Behcet's disease. Nat. Genet. 42, 698–702.
Rogers, N. C., Slack, E. C., Edwards, A. D., Nolte, M. A., Schulz, O., Schweighoffer, E., Williams, D. L., Gordon, S., Tybulewicz, V. L., Brown, G. D., and Reis e Sousa, C. (2005). Syk-dependent cytokine induction by Dectin-1 reveals a novel pattern recognition pathway for C type lectins. Immunity 22, 507–517.
Rubtsov, Y. P., Rasmussen, J. P., Chi, E. Y., Fontenot, J., Castelli, L., Ye, X., Treuting, P., Siewe, L., Roers, A., Henderson, W. R. Jr., Muller, W., and Rudensky, A. Y. (2008). Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity 28, 546–558.
Saraiva, M., Christensen, J. R., Tsytsykova, A. V., Goldfeld, A. E., Ley, S. C., Kioussis, D., and O'Garra, A. (2005). Identification of a macrophage-specific chromatin signature in the IL-10 locus. J. Immunol. 175, 1041–1046.
Saraiva, M., Christensen, J. R., Veldhoen, M., Murphy, T. L., Murphy, K. M., and O'Garra, A. (2009). Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose. Immunity 31, 209–219.
Saraiva, M., and O'Garra, A. (2010). The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 10, 170–181.
Sato, T., McCue, P., Masuoka, K., Salwen, S., Lattime, E. C., Mastrangelo, M. J., and Berd, D. (1996). Interleukin 10 production by human melanoma. Clin. Cancer Res. 2, 1383–1390.
Scapini, P., Lamagna, C., Hu, Y., Lee, K., Tang, Q., DeFranco, A. L., and Lowell, C. A. (2011). B cell-derived IL-10 suppresses inflammatory disease in Lyn-deficient mice. Proc. Natl. Acad. Sci. U.S.A. 108, E823–E832.
Shaw, M. H., Freeman, G. J., Scott, M. F., Fox, B. A., Bzik, D. J., Belkaid, Y., and Yap, G. S. (2006). Tyk2 negatively regulates adaptive Th1 immunity by mediating IL-10 signaling and promoting IFN-gamma-dependent IL-10 reactivation. J. Immunol. 176, 7263–7271.
Shimomura, Y., Mizoguchi, E., Sugimoto, K., Kibe, R., Benno, Y., Mizoguchi, A., and Bhan, A. K. (2008). Regulatory role of B-1 B cells in chronic colitis. Int. Immunol. 20, 729–737.
Shoemaker, J., Saraiva, M., and O'Garra, A. (2006). GATA-3 directly remodels the IL-10 locus independently of IL-4 in CD4+ T cells. J. Immunol. 176, 3470–3479.
Stumhofer, J. S., Silver, J. S., Laurence, A., Porrett, P. M., Harris, T. H., Turka, L. A., Ernst, M., Saris, C. J., O'Shea, J. J., and Hunter, C. A. (2007). Interleukins 27 and 6 induce STAT3-mediated T cell production of interleukin 10. Nat. Immunol. 8, 1363–1371.
Sun, J., Dodd, H., Moser, E. K., Sharma, R., and Braciale, T. J. (2011). CD4+ T cell help and innate-derived IL-27 induce Blimp-1-dependent IL-10 production by antiviral CTLs. Nat. Immunol. 12, 327–334.
Wang, Z. Y., Sato, H., Kusam, S., Sehra, S., Toney, L. M., and Dent, A. L. (2005). Regulation of IL-10 gene expression in Th2 cells by Jun proteins. J. Immunol. 174, 2098–2105.
Wolf, S. D., Dittel, B. N., Hardardottir, F., and Janeway, C. A. Jr. (1996). Experimental autoimmune encephalomyelitis induction in genetically B cell-deficient mice. J. Exp. Med. 184, 2271–2278.
Xu, J., Yang, Y., Qiu, G., Lal, G., Wu, Z., Levy, D. E., Ochando, J. C., Bromberg, J. S., and Ding, Y. (2009). c-Maf regulates IL-10 expression during Th17 polarization. J. Immunol. 182, 6226–6236.
Yanaba, K., Bouaziz, J. D., Haas, K. M., Poe, J. C., Fujimoto, M., and Tedder, T. F. (2008). A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls T cell-dependent inflammatory responses. Immunity 28, 639–650.
Zhou, Y., Kawasaki, H., Hsu, S. C., Lee, R. T., Yao, X., Plunkett, B., Fu, J., Yang, K., Lee, Y. C., and Huang, S. K. (2010). Oral tolerance to food-induced systemic anaphylaxis mediated by the C-type lectin SIGNR1. Nat. Med. 16, 1128–1133.
Ziegler-Heitbrock, L., Lotzerich, M., Schaefer, A., Werner, T., Frankenberger, M., and Benkhart, E. (2003). IFN-alpha induces the human IL-10 gene by recruiting both IFN regulatory factor 1 and Stat3. J. Immunol. 171, 285–290.
Keywords: epigenetics, interleukin-10, plasticity, regulatory B cells, transcriptional regulation
Citation: Kubo M and Motomura Y (2012) Transcriptional regulation of the anti-inflammatory cytokine IL-10 in acquired immune cells. Front. Immun. 3:275. doi: 10.3389/fimmu.2012.00275.
Received: 23 May 2012; Paper pending published: 18 June 2012;
Accepted: 13 August 2012; Published online: 30 August 2012.
Edited by:
Masaaki Murakami, Osaka University, JapanReviewed by:
David Hildeman, Cincinnati Children's Hospital, USACopyright © 2012 Kubo and Motomura. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Masato Kubo, Division of Molecular Pathology, Research Institute for Biological Sciences, Tokyo University of Science, 2669 Yamazaki, Noda, Chiba 278-0022, Japan. e-mail:cmF5c29sZmNAcmNhaS5yaWtlbi5qcA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.