 Pavel Dráber*
Pavel Dráber* Vadym Sulimenko and Eduarda Dráberová
Vadym Sulimenko and Eduarda Dráberová
- Department of Biology of Cytoskeleton, Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, Prague, Czech Republic
Mast cell activation mediated by the high affinity receptor for IgE (FcεRI) is a key event in allergic response and inflammation. Other receptors on mast cells, as c-Kit for stem cell factor and G protein-coupled receptors (GPCRs) synergistically enhance the FcεRI-mediated release of inflammatory mediators. Activation of various signaling pathways in mast cells results in changes in cell morphology, adhesion to substrate, exocytosis, and migration. Reorganization of cytoskeleton is pivotal in all these processes. Cytoskeletal proteins also play an important role in initial stages of FcεRI and other surface receptors induced triggering. Highly dynamic microtubules formed by αβ-tubulin dimers as well as microfilaments build up from polymerized actin are affected in activated cells by kinases/phosphatases, Rho GTPases and changes in concentration of cytosolic Ca2+. Also important are nucleation proteins; the γ-tubulin complexes in case of microtubules or Arp 2/3 complex with its nucleation promoting factors and formins in case of microfilaments. The dynamic nature of microtubules and microfilaments in activated cells depends on many associated/regulatory proteins. Changes in rigidity of activated mast cells reflect changes in intermediate filaments build up from vimentin. This review offers a critical appraisal of current knowledge on the role of cytoskeleton in mast cells signaling.
Introduction
Mast cells play an essential role in innate immunity, allergy, and inflammation. When activated they release mediators that are pivotal for initiation of inflammatory reactions associated with allergic disorders. Activation of mast cells in allergic inflammatory response occurs via the high affinity receptor for IgE (FcεRI) following receptor aggregation induced by antigen-mediated cross-linking of IgE occupied FcεRI. This response is profoundly influenced by other factors that modulate the threshold levels of mast cell triggering. Cell-surface receptors, such as the stem cell factor (SCF) receptor, c-Kit, and the specific G protein-coupled receptors (GPCRs) synergistically enhance FcεRI-mediated mast cell exocytosis (degranulation) (Kalesnikoff and Galli, 2008; Gilfillan and Rivera, 2009).
The first defined step in FcεRI signaling is phosphorylation of the immunoreceptor tyrosine-based activation motifs (ITAMs) in the cytoplasmic tails of the FcεRI β and γ subunits by Src-family protein tyrosine kinase (PTK) Lyn. Tyrosine-phosphorylated ITAMs of the FcεRI γ subunits serve as docking sites for the tandem Src homology (SH)2 domains of the non-receptor tyrosine kinase Syk (spleen tyrosine kinase) of the Syk/Zap-70 family. Syk bound to ITAMs adopts an active conformation that facilitates its phosphorylation by Lyn and further increase in enzymatic activity. Consequently, Syk phosphorylates a number of its targets critical for further propagation of the signal, including the transmembrane adaptor linker proteins (TRAPs), linker for activation of T cells (LAT), and non-T cell activation linker (NTAL) (Dráber et al., 2012). Phosphorylated TRAPs serve as plasma membrane docking sites for cytoplasmic SH2 domain containing molecules like growth factor receptor-bound protein 2 (Grb2) and phospholipase Cγ (PLCγ). After membrane anchoring and activation, PLCγ hydrolyses phosphatidylinositol 4,5-biphosphate (PIP2) and produces diacylglycerol (DAG) and inositol 1,4,5,-triphosphate (IP3) that binds to its receptors on endoplasmic reticulum (ER). This results in Ca2+ efflux from ER. Subsequently, depletion of Ca2+ from ER lumen induces Ca2+ influx across the plasma membrane through store-operated Ca2+ channels. The influx leads to enhancement in free cytoplasmic Ca2+ concentration, a step which is substantial for further signaling events (Blank and Rivera, 2004; Rivera et al., 2008). The stimulatory effect of cross-linking of IgE receptors with antigen can be mimicked in part by pervanadate, a compound that effectively inhibits protein tyrosine phosphatases (Teshima et al., 1994).
Mast cells also express on their surfaces receptors for a number of ligands known to be potent mast cell chemoattractants. The most thoroughly investigated of these is SCF, a natural ligand for c-Kit (Meininger et al., 1992). Others include agonists of GPCRs, such as prostaglandin E2 (PGE2), which positively regulates mast cell responses through the EP3 receptor (Weller et al., 2007). There is some evidence for the hypothesis that IgE sensitized mast cells migrate toward the antigen, potentially through the release of chemoattractants from the mast cells themselves (Ishizuka et al., 2001).
Degranulation can also be induced, independently of FcεRI aggregation, by thapsigargin, a compound that discharges intracellular Ca2+ stores by inhibition of the Ca2+-ATPase pumps located in ER membrane (Thastrup et al., 1990). Similarly, stimulation with basic secretagogues, such as anaphylatoxins, neuropeptides, compound 48/80, and poly-L-lysine, results in activation of distinct FcεRI-independent pathways (Lagunoff et al., 1983). These stimuli share a common characteristic of being polycations which are able to penetrate through the plasma membrane and stimulate the G-proteins (Bueb et al., 1992; Deng et al., 2009).
Activation of mast cells is accompanied by changes in cell morphology, enhanced adhesion to various substrates, migration, and exocytosis. All these processes are dependent on the activity of cytoskeletal proteins. The cytoskeleton of mast cells is formed by three types of filamentous structures: microtubules formed by tubulin dimers, microfilaments composed of actin, and intermediate filaments of vimentin-type. These cytoskeletal networks differ in their organization, protein compositon, and functions. In cells, there is equilibrium between soluble and polymeric state of principal building proteins that form corresponding cytoskeletal structures. Research has mostly focused on microfilaments and microtubules, that both play a critical role in these processes. Data on the involvement of intermediate filaments in mast cell activation are still very limited. A simplified model of early signaling events leading to cytoskeleton reorganization after FcεRI aggregation is shown in Figure 1.
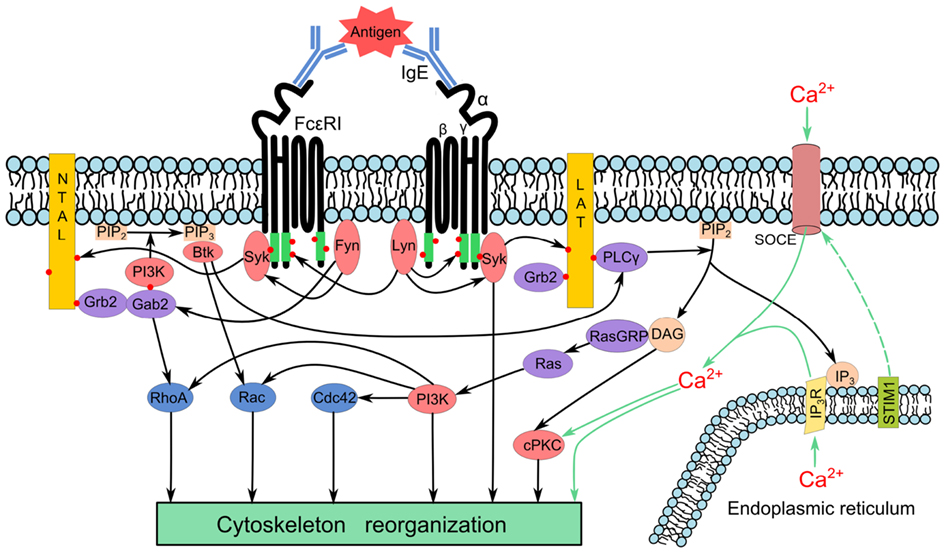
Figure 1. Simplified model of early signaling events leading to cytoskeleton reorganization after FcεRI aggregation. Aggregation of the FcεRI by multivalent antigen induces activation of the Src kinase Lyn, which phosphorylates (red dots) ITAM motifs of FcεRI β and γ subunits (green), followed by anchoring of the Syk kinase to FcεRI through interaction of Syk-SH2 domains with phosphorylated ITAMs. Aggregation also leads to activation of Fyn kinase that phosphorylates the adaptor Gab2. Transmembrane adaptor proteins NTAL and LAT are phosphorylated by Syk giving rise to formation of binding sites for various SH2-containing proteins such as Grb2. In this way it brings Gab2 and phosphoinositide-3-kinase (PI3K) to the membrane. Activated PI3K phosphorylates phosphatidylinositol- 4,5-biphosphate (PIP2) to yield phosphatidylinositol-3,4,5-triphosphate (PIP3), a binding site for PH-domain containing proteins like kinase Btk. This process further propagates the signal through activation of phospholipase Cγ (PLCγ) that hydrolyses PIP2 to inositol 1,4,5,-triphosphate (IP3) and diacylglycerol (DAG). The binding of IP3 to its receptor (IP3R) triggers release of Ca2+ from endoplasmic reticulum; STIM1 couples depletion of Ca2+ from endoplasmic reticulum with activation of Ca2+ channels and influx of extracellular Ca2+ by the so called store-operated Ca2+ entry (SOCE). DAG and Ca2+ activate conventional protein kinase C (cPKC). DAG also recruits Ras guanyl nucleotide-releasing proteins (RasGRPs) for subsequent activation of Ras family proteins that stimulate PI3K. The PIP3 also recruits nuclotide exchange factors that subsequently lead to activation of the Rho family of GTPases (RhoA, Rac, and Cdc42). Enhanced concentration of Ca2+ as well as activated kinases and GTPases modulate cytoskeleton arrangements.
Cytoskeleton of activated mast cells has been studied in various cellular models ranging from mouse bone marrow-derived mast cells (BMMCs) and isolated rat peritoneal mast cells (RPMCs) to mouse BMMC lines (e.g., MC/9), rat basophilic leukemia cell lines (e.g., RBL-2H3), or human mast cell lines (e.g., HMC-1). Typical distribution of microtubules and microfilaments in interphase RBL-2H3 cells is shown in Figure 2.
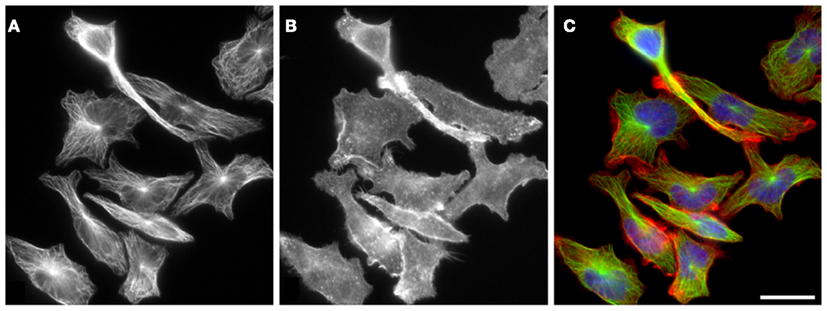
Figure 2. Distribution of microtubules and F-actin in RBL-2H3 cells. Microtubules (A) and F-actin (B) have distinct subcellular organization in resting interphase RBL-2H3 cells. While microtubule arrays radiate from perinuclear centrosomes, F-actin does not have a single organizing center within the cell and is more concentrated at cell periphery. Cells were fixed with formaldehyde and extracted with Triton X-100 before staining with rabbit antibody to α-tubulin [(A); green] and rhodamine-conjugated phalloidin [(B); red]. DNA was stained with DAPI (blue). Superposition of α-tubulin and F-actin is shown in (C). Scale bar, 20 μm. Photography E. Dráberová (Institute of Molecular Genetics AS CR, Prague).
This review presents a critical survey of current knowledge on the role of cytoskeleton in mast cells signaling. Basic characteristic properties of particular cytoskeletal systems are outlined at the beginning followed by evaluation of their participation in mast cell activation.
Microtubules
Microtubules and Tubulins
Microtubules are cytoskeletal polymers essential for many cellular activities such as maintenance of cell shape, division, migration, positioning of organelles, and ordered vesicle transport powered by motor proteins. They are organized into radial cytoplasmic networks in interphase cells, bipolar spindles in mitotic cells, and midbodies during cytokinesis. The basic building blocks of microtubules are heterodimers of globular α- and β-tubulin subunits, which are arranged in a head-to-tail fashion to form 13 protofilaments that constitute cylindrical and left-handed helical microtubule wall with outer diameter around 25 nm (Kreis and Vale, 1999). Microtubules are thus inherently polar, and contain two structurally distinct ends: a slow-growing minus end, exposing α-tubulin subunits, and a fast-growing plus end, exposing β-tubulin subunits (Nogales and Wang, 2006). Typically, microtubule minus ends are stably anchored in microtubule-organizing centers (MTOC) as centrosomes, whereas the plus ends are highly dynamic and switch between phases of growth and shrinkage. The properties of microtubule can be regulated by incorporation of alternative tubulin isotypes, post-translational modification (PTM) of tubulin subunits, and binding of microtubule associated proteins (MAPs) (Amos and Schlieper, 2005; Verhey and Gaertig, 2007).
Both tubulin subunits are encoded by multiple genes, which are in a large part phylogenetically conserved; eight α-tubulin and seven β-tubulin isotypes were identified in human (Luduena and Banerjee, 2008; Katsetos and Dráber, 2012). The secondary and tertiary structures of the α- and β-monomers are essentially identical, and reflect more than 40% identity over their entire sequence (Nogales et al., 1998). Each tubulin monomer binds one molecule of GTP, non-exchangeably in α-subunit and exchangeably in β-subunit. Most of the studied microtubules appeared to have a seam along their length. This is due to 12-nm helical pitch in combination with the 8-nm longitudinal repeat between αβ-tubulin dimers (Wade and Hyman, 1997).
Microtubule Dynamics and Nucleation
Polymerization and depolymerization of microtubules is driven by the binding, hydrolysis, and exchange of GTP on the β-tubulin monomer. GTP hydrolysis is necessary for switching between alternating phases of growth and shrinkage separated by catastrophe (transition from growth to shrinkage) and rescue (transition from shortening to growth) events. Polymerization is initiated from a pool of GTP-loaded tubulin subunits. Growing microtubule sheets maintain a “cap” of tubulin-GTP subunits to stabilize the straight tubulin conformation within the microtubule lattice (Carlier, 1991). A loss of this cap results in rapid depolymerization. The polymerization-depolymerization cycle is completed by exchanging GDP of the disassembly products with GTP (Figure 3) (Akhmanova and Steinmetz, 2008). These characteristics result in dynamic instability (Mitchison and Kirschner, 1984), an essential feature of microtubules that allows them to search through the cell for targets, such as the chromosomal kinetochores, the cell cortex, and actin cytoskeleton (Desai and Mitchison, 1997). The stability and dynamics of microtubules are actively regulated by a number of cellular factors as well as a variety of ligands, among them are well-known drugs (e.g., vinblastine, nocodazole and paclitaxel) that potently suppress the dynamic instability of microtubules (Jordan and Kamath, 2007).
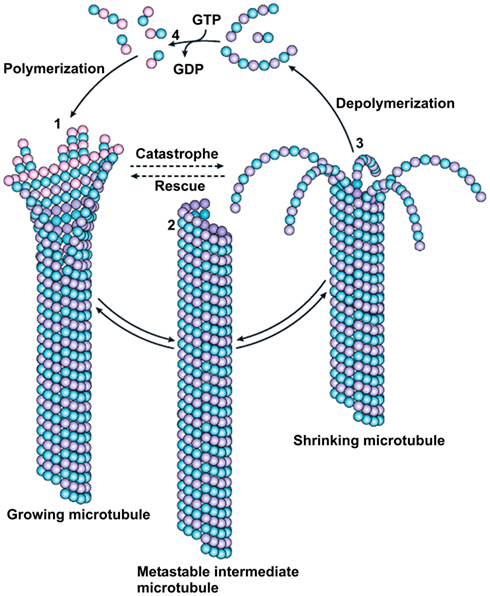
Figure 3. Dynamic instability of microtubules. Polymerization of microtubules is initiated from a pool of GTP-loaded tubulin subunits (pink beads). GTP hydrolysis changes the conformation of the protofilament from a slightly curved tubulin-GTP to a more sharply curved tubulin-GDP structure. The curved tubulin-GDP (lilac beads) is forced to remain straight when it is part of the microtubule wall. Growing microtubule sheets presumably maintain the “cap” of tubulin-GTP subunits to stabilize the straight tubulin conformation within the microtubule lattice (1). Closure of the terminal sheet structure generates a metastable, blunt-ended microtubule intermediate (2) which may pause, undergo further growth or switch to the depolymerization phase. A shrinking microtubule is characterized by fountain-like arrays of ring and spiral protofilament structures (3). The polymerization–depolymerization cycle is completed by exchanging GDP of the disassembly products with GTP (4). Transition from growth to shrinkage is termed catastrophe; transition from shortening to growth is called rescue. Reprinted by permission from Macmillan Publishers Ltd: (Nature Reviews in Molecular Biology; Akhmanova and Steinmetz, 2008).
One of the key components required for microtubule nucleation and stabilization is the γ-tubulin (Oakley and Oakley, 1989), a highly conserved, but minor, member of the tubulin family concentrated in interphase cells in MTOCs (Stearns et al., 1991). In mitotic cells γ-tubulin appears on spindle poles and along spindle fibers, during cytokinesis it is found in midbodies (Julian et al., 1993; Nováková et al., 1996). γ-Tubulin has also been found to bind to membranous components of the cell (Chabin-Brion et al., 2001; Dryková et al., 2003). Contrary to αβ-tubulin dimers only two functional genes exist in mammalian cells that code very similar γ-tubulins (Wise et al., 2000; Vinopal et al., 2012). The γ-tubulin is associated in complexes with other proteins. The human γ-tubulin small complex (γTuSC) comprises two molecules of γ-tubulin and one molecule each of GCP (γ-tubulin complex protein) 2 and 3. The large γ-tubulin-ring complex (γTuRC) derives from γ-TuSCs by condensation and association with proteins GCP4, GCP5 and GCP6 (Murphy et al., 2001). Soluble γ-tubulin can associate with αβ-tubulin dimers irrespective of the size of γ-tubulin complexes (Sulimenko et al., 2002). Several reports indicated that kinases might be involved in the regulation of γ-tubulin interactions (Kapeller et al., 1995; Vogel et al., 2001; Kukharskyy et al., 2004).
Modulation of Microtubule Functions by Posttranslational Modifications and Associated Proteins
αβ-Tubulin dimers can be separated according to their isoelectric points to more than 20 isoforms, far more than expected from the number of isotypes that are actually expressed (Wolff et al., 1982; Linhartová et al., 1992). This fact reflects extensive PTMs of both tubulin subunits. Most PTMs of tubulin subunits take place after polymerization into microtubules, and modified tubulins are non-uniformly distributed along microtubules. More than fifteen tubulin PTMs have been described; well characterized PTMs include acetylation, detyrosination, polyglutamylation, and polyglycylation. Diverse PTMs form the biochemical “tubulin code” that can be read by factors interacting with microtubules. Specific microtubule regions can be distinguished biochemically and functionally by the presence of PTMs on tubulins (Westermann and Weber, 2003; Verhey and Gaertig, 2007). It appears that modifications could participate in targeting the molecular motors and MAPs to defined subsets of microtubules inside the cell (Janke and Bulinski, 2011).
Proteins interacting with microtubules are involved in microtubule growth, stabilization, destabilization, and connection of microtubules to other cellular organelles. The mechanochemical ATPases kinesins and dyneins (microtubule motor proteins) use microtubules as pathways for intracellular transport. A wide variety of microtubule regulatory proteins promote the functional diversity of microtubules (Lyle et al., 2009a,b; Wade, 2009). Regulation can occur on many levels, including regulation of tubulin monomer folding or microtubule nucleation. Microtubule stability and dynamics is regulated by a large number of proteins that belongs to microtubule-stabilizing MAPs, microtubule severing proteins, microtubule-disassembly, or assembly promoters and microtubule plus-end tracking proteins (+TIPs) that specifically accumulate at growing microtubule plus ends. To essential +TIPs belongs the end-binding protein 1 (EB1) that directly interacts with tubulin dimers (Dráber and Dráberová, 2012). Subcellular localization of γ-tubulin and EB1 in resting RBL-2H3cells is shown in Figure 4.
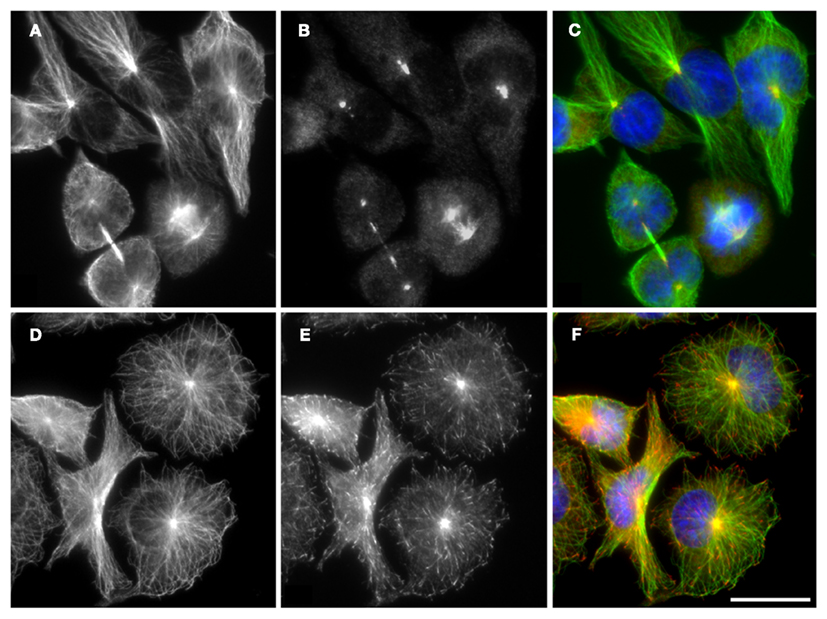
Figure 4. Distribution of microtubule proteins in RBL-2H3 cells. In resting RBL-2H3 cells, microtubules, formed by αβ-tubulin dimers (A,D) originate from centrosomes where the γ-tubulin is concentrated (B). Growing microtubules are marked by plus-end tracking protein EB1 (E). Cells were fixed with cold methanol before staining with rabbit antibody to α-tubulin dimer [(A,D); green fluorescence], mouse monoclonal antibody to γ-tubulin [(B), red fluorescence], or monoclonal antibody to EB1 [(E), red fluorescence]. DNA was stained with DAPI (blue). Superposition of α-tubulin and γ-tubulin is shown in (C). Superposition of α-tubulin and EB1 is shown in (F). Scale bar, 20 μm. Photography E. Dráberová (Institute of Molecular Genetics AS CR, Prague).
Microtubules in Activated Mast Cells
Changes in Microtubule Organization and Dynamics in Activated Cells
It is well established that agents inhibiting tubulin polymerization suppress the exocytosis (Urata and Siraganian, 1985; Tasaka et al., 1991) and that the movement of secretory granules in mast cells is dependent on intact microtubules (Martin-Verdeaux et al., 2003; Smith et al., 2003; Nishida et al., 2005).
Activation of BMMCs cells leads to changes in topography of microtubules. Compared to resting cells, FcεRI aggregation in BMMCs attached to poly-L-lysine resulted in accumulation of microtubules in cell periphery detectable by immunofluorescence microscopy. Similarly, activation of BMMCs by pervanadate, that mimics the stimulatory effect of antigen, induced enhanced formation of microtubules (Sulimenko et al., 2006). The same effect was also observed when BMMCs were activated and fixed in cell suspension before attachment to glass slides by cytospin (Nishida et al., 2005). When prior to activation the cells were attached to fibronectin, what more closely resembles the natural conditions existing in connective tissue where mast cells are congregated (Galli et al., 2008), prominent protrusions containing microtubules appeared in activated cells (Hájková et al., 2011). Attachment of BMMCs to fibronectin alone failed to generate such protrusions (Figure 5).
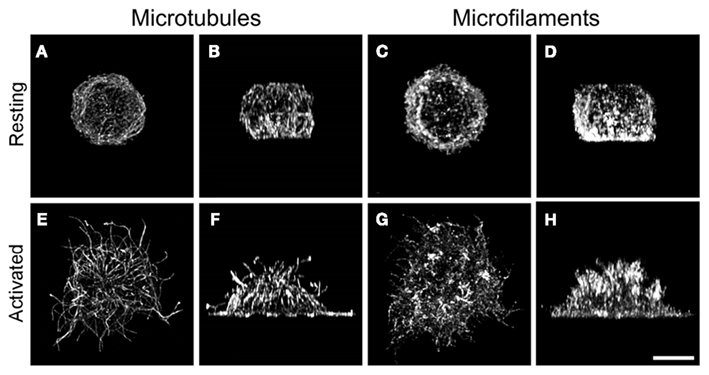
Figure 5. Reorganization of microtubules and F-actin in activated BMMCs. Resting BMMC (A–D) or thapsigargin-activated BMMC (E–H) attached to fibronectin were fixed in formaldehyde and extracted in Triton X-100. Cells were double label stained for α-tubulin (A,B,E,F) and F-actin (C,D,G,H), and examined with laser scanning confocal microscope. The stacks of confocal sections were deconvoluted and subjected to three-dimensional reconstruction using Huygens deconvolution software. The resulting 3-D images are viewed from top of the cells (A,C,E,G) and from the plane perpendicular to the plane of cell adhesion (B,D,F,H). The same cell is represented in (A–D) or (E–H). Scale bar, 5 μm. Photography Z. Hájková (Institute of Molecular Genetics AS CR, Prague).
Microtubule dynamics can be examined by live cell imaging of tagged-EB1. When microtubule plus-end dynamics in cells expressing EB1-GFP was monitored by means of time-lapse imaging using total internal reflection fluorescence microscopy (TIRFM), the number of growing microtubules in the periphery of activated cells was found substantially increased compared to non-activated cells (Figure 6). It is known that the engagement of integrins by their ligands activates some signaling pathways that modulate signals originating from other receptors (Schwartz et al., 1995). When mast cells were activated via FcεRI and integrins simultaneously, phosphorylation events were prolonged, and intensified (Lam et al., 2003). Thus, generation of protrusions in activated cells could reflect a response to such integrated signals.
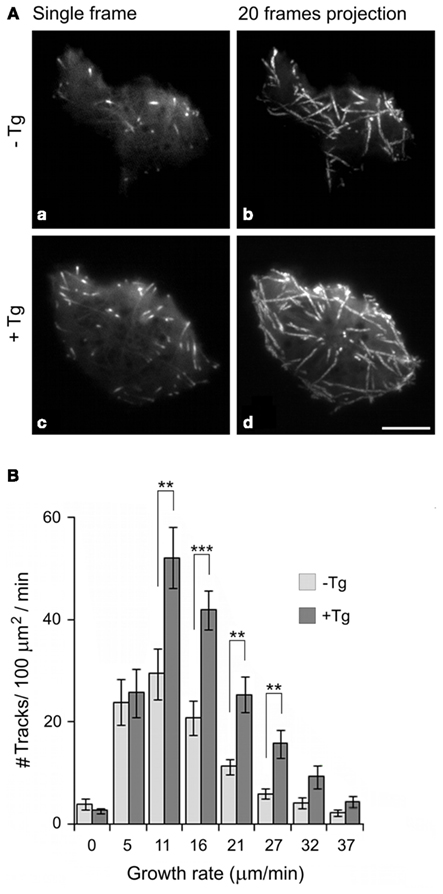
Figure 6. Changes in the number of growing microtubules in cell periphery during mast cell activation as determined by TIRFM time-lapse imaging in living cells. (A) Time-lapse imaging of resting (a,b) and thapsigargin-activated (c,d) BMMCs expressing EB1-GFP. Still images of EB1 (a,c) and tracks of EB1 comets over 20 s created by maximum intensity projection of 20 consecutive frames (b,d). Scale bar, 5 μm. (B) Histogram of microtubule growth rates in the cell periphery of resting (−Tg) and thapsigargin-activated (+Tg) cells. A total of 15 different cells were tracked in five independent experiments. Values indicate mean ±SE, n = 15 (**p < 0.01; ***p < 0.001). Reproduced by permission from reference Hájková et al. (2011). Copyright 2011; The American Association of Immunologists, Inc.
Store-Operated Ca2+ Entry and Microtubules in Activated Cells
The store-operated Ca2+ entry (SOCE) is important for the replenishment of intracellular stores by means of sarco/endoplasmatic reticulum Ca2+-ATPase (SERCA) pumps located in ER membrane (Parekh and Putney Jr, 2005; Smyth et al., 2006). A key component in SOCE is the stromal interacting protein 1 (STIM1; Roos et al., 2005), that represents the Ca2+ sensor responsible for communicating the depleted state of intracellular Ca2+ compartments to store-operated Ca2+ channels. In quiescent cells with ER filled with Ca2+, STIM1 is distributed homogenously throughout ER (Dziadek and Johnstone, 2007) but relocates upon release of Ca2+ from ER stores to distinct puncta on ER in close proximity to the plasma membrane (Liou et al., 2005). Aggregated STIM1 activates members of the Orai family of store-operated Ca2+ channels, resulting in opening of plasma membrane Ca2+ release-activated Ca2+ (CRAC) channels and Ca2+ influx into the cell (Prakriya et al., 2006). In this way STIM1 serves as a major regulator of SOCE. In addition, transient receptor potential channels (TRPC) and ion exchangers also contribute to the generation of Ca2+ signals that may be global or have dynamic (e.g., waves and oscillations) and spatial resolution for specific functional readouts (Ma and Beaven, 2009). SOCE has been described in various cell types including mast cells activated by antigen stimulation, which induces IP3 mediated Ca2+ release from ER stores. Aggregation of FcεRI receptors triggered in RBL cells significant oligomerization of STIM1 and formation of STIM1 puncta near the plasma membrane (Liou et al., 2007). Further study showed that clustering of STIM1 with Orai 1 at plasma membrane of antigen-activated RBL-2H3 cells was depended on the extent of Ca2+ store depletion (Calloway et al., 2009). The results pointed to the capability of TRPC proteins to interact with STIM1 and Orai proteins in mast cells. This could add further flexibility to the Ca2+ “tool kit” that is available to activated mast cells (Ma and Beaven, 2011).
Colocalization of ER-embedded STIM1 with microtubules has been described for several cell types including DT40 B cells (Baba et al., 2006), HEK293 cells (Mercer et al., 2006; Smyth et al., 2007), and mast RBL-2H3 cells (Calloway et al., 2009). Moreover, comet-like movement of STIM1, resembling +TIPs, was reported in living cells (Grigoriev et al., 2008), and STIM1 was found to contain a short sequence (SxIP) responsible for direct binding to EB1 (Honnappa et al., 2009). Comet-like movement of STIM1 was substantially reduced after addition of thapsigargin (Hájková et al., 2011), a compound that inhibits SERCA (Thastrup et al., 1990). This is in agreement with the impaired association of STIM1 with microtubules observed in antigen-activated RBL-2H3 cells (Calloway et al., 2009). Data on fluorescence resonance energy transfer (FRET) imaging of EB1 and STIM1 showed that STIM1 dissociated from EB1 and associated with SERCA upon store depletion of Ca2+. The process seems to be reversible, since the replenishment of intracellular Ca2+ stores also restored the STIM1-EB1 interactions (Sampieri et al., 2009). Moreover, no effect on SOCE was observed in cells with depleted EB1 (Grigoriev et al., 2008). Considering these findings together, it is likely that interaction of STIM1 with EB1 on growing microtubules is not essential for the transport of STIM1 to plasma membrane during mast cell activation.
After depletion of intracellular Ca2+ stores, STIM1 accumulates into puncta, i.e., discrete subregions of ER located in a close proximity (10–25 nm) to plasma membrane (Cahalan, 2009). STIM1 puncta are formed several seconds before the opening of Ca2+ channels (Wu et al., 2006), and it could therefore be inferred that microtubules are involved in this process. However, microtubule disruption by nocodazole had no effect on puncta formation in activated BMMCs, while the comet-like movement of STIM1 was abolished (Hájková et al., 2011). This is in line with observations that uptake of extracellular Ca2+ was only partially inhibited in nocodazole-pretreated and thapsigargin-activated BMMCs. This may suggest that STIM1 aggregation beneath the plasma membrane and subsequent opening of Ca2+ release-activated Ca2+ channels do not require intact microtubules in activated mast cells (Hájková et al., 2011). Previous studies often reported discordant effects of nocodazole treatment on SOCE or ICRAC, the current most frequently associated with SOCE, in various cell types. While no effect of nocodazole was observed in RBL-1 (Bakowski et al., 2001; Smyth et al., 2007) and DT40 cells (Baba et al., 2006), significant inhibition was demonstrated in other cell types, such as RBL-2H3, BMMCs (Oka et al., 2005), and HEK 293 (Smyth et al., 2007). It appears that other factors next to cell type, like the treatment protocol or the method of Ca2+ depletion, might modify the results of the experiments. It is also possible that microtubules play a supporting role in SOCE signaling by optimizing the location of ER containing STIM1 before cell activation (Smyth et al., 2007; Galan et al., 2011).
Protrusions containing microtubules were found in BMMCs stimulated by three types of activators that induced depletion of Ca2+ from internal stores (FcεRI aggregation, pervanadate, or thapsigargin treatment). Formation of protrusions was inhibited in cells with decreased level of STIM1 and with correspondingly reduced influx of extracellular Ca2+. Protrusions in BMMCs with depleted level of STIM1 were restored after introduction of human STIM1. These data together with the absence of protrusions in cells activated in Ca2+-free media suggest that STIM1 regulated Ca2+ influx plays a crucial role in generation of these microtubule enriched structures. Changes in the concentration of cytoplasmic Ca2+also affected a microtubule plus-end dynamics and altered chemotactic response. Altogether these results support the concept of a tight crosstalk between microtubular network and Ca2+ signaling machinery in the course of mast cell activation (Hájková et al., 2011). The presence of aggregated STIM1 in protrusions could help organize Ca2+ release-activated Ca2+ channels (Cahalan, 2009) and open locally these channels to bring about SOCE. These interactions may be subjected to modulations by Ca2+ channel regulators, such as calmodulin (Mullins et al., 2009) or CRACR2A, a cytosolic Ca2+ sensor that stabilizes CRAC channels (Srikanth et al., 2010).
γ-Tubulin in Activated Mast Cells
The formation of microtubules can be effectively regulated by microtubule nucleation, a process where γ-tubulin plays a key role. It was reported that Lyn kinase, a major Src-family kinase in RBL-2H3 cells (Eiseman and Bolen, 1992), forms complexes containing γ-tubulin and phosphotyrosine proteins in these cells activated by FcεRI aggregation or by an exposure to pervanadate (Dráberová et al., 1999). However, Lyn kinase is dispensable for the formation of functional γ-tubulin complexes, as indicated by normal topography of microtubules in Lyn−/− BMMCs. Tyrosine-phosphorylated proteins were found to be associated with immunoprecipitated γ-tubulin in resting Lyn−/− BMMCs, and the level of tyrosine phosphorylation of these proteins substantially increased after activation. Immunoprecipitation experiments and in vitro kinase assays combined with specific inhibitors revealed that Fyn and Syk kinases in complexes containing γ-tubulin are capable of phosphorylating various substrates (Sulimenko et al., 2006). Interaction of γ-tubulin complexes with Fyn was confirmed by pull-down experiments, where γ-tubulin complex bound to SH2 domain of Fyn kinase in a phosphotyrosine dependent manner. Similar binding to SH2 domains of Fyn or Src kinases has also been described in mouse embryonal carcinoma cells (Kukharskyy et al., 2004). Although γ-tubulin possesses a potential binding site for SH2 domain, experiments with several candidate synthetic peptide inhibitors failed to confirm a direct binding of γ-tubulin to this domain. Thus, the association with SH2 domain is probably mediated via adaptor-like tyrosine-phosphorylated protein(s) (Sulimenko et al., 2006). The Fyn kinase, like other Src-family kinases, is commonly involved in the formation of multi-protein complexes engaged in interaction with the SH2 and SH3 domains. It is therefore likely that association of Src-family kinases with γ-tubulin is mediated through other proteins that might be important for microtubule nucleation. Significance of Src kinase for microtubule nucleation from centrosomes was ascertained by microtubule regrowth experiments on human fibroblasts adherent to fibronectin (Colello et al., 2010). Src-family kinases could regulate the recruitment of γ-tubulin to the centrosome by different mechanisms. They can either phosphorylate γTuRC proteins to promote the assembly of the complex, or may regulate the association or activity of NEDD1/GCP-WD, a protein that is required for the centrosomal recruitment of γTuRC (Lüders et al., 2006). Alternatively, kinases may indirectly affect this process by regulating the assembly of the pericentriolar matrix (PCM) surrounding centrioles. Identification of Src-family kinase substrates that promote the recruitment of γ-tubulin to the centrosome and microtubule nucleation will be an important step forward in the elucidation of the mechanisms involved.
The molecular mechanism of the association of Syk kinase with γ-tubulin is not fully understood. In RBL-2H3 cells as well as in BMMCs, Syk is one of the preferable substrates for Lyn kinase (Jouvin et al., 1994). However, even in Lyn−/− BMMCs there is still some phosphorylation of Syk on tyrosine, which is dependent on FcεRI activation (Parravicini et al., 2002). Because the Src-family selective tyrosine kinase inhibitor PP2 inhibited phosphorylation of Syk in pervanadate-activated Lyn−/− BMMCs, and the Syk-selective tyrosine kinase inhibitor piceatannol reduced phosphorylation of proteins in γ-tubulin immunocomplexes, it is likely that it is the crosstalk between Fyn and Syk which is responsible for tyrosine phosphorylation of proteins associated with γ-tubulin immunocomplexes in Lyn−/− BMMCs (Sulimenko et al., 2006).
There are reports pointing to the localization of Fyn and Syk kinases in centrosomal region. Fyn kinase was found in centrosomes in myelocytic leukemia cell line HL-60 (Katagiri et al., 1993) and in human T lymphocytes (Ley et al., 1994). Moreover, in human Jurkat T lymphocytes tubulin phosphorylated on tyrosine interacted with SH2 domain of Fyn kinase (Marie-Cardine et al., 1995). Syk was located at the centrosomes in B lymphocytes (Navara et al., 1999). Thus, tyrosine phosphorylation of centrosomal proteins by Fyn and Syk kinases might be the process linking microtubules to early activation events in mast cells.
Tubulin has been shown to serve as a substrate for Syk kinase in vivo (Peters et al., 1996). Syk can phosphorylate both soluble tubulin (Fernandez et al., 1999) and tubulin in microtubules (Faruki et al., 2000). Syk phosphorylates α-tubulin on the conserved tyrosine residue (Tyr432) and Syk-selective inhibitor piceatannol blocks the receptor-stimulated tubulin phosphorylation in B lymphocytes (Peters et al., 1996) as well in BMMCs (Sulimenko et al., 2006). However, phosphorylation of tubulin by Syk did not have any profound effect on microtubule assembly in pervanadate-treated cells (Faruki et al., 2000). Besides, phosphorylation of tubulin by Src kinase did not cause any significant changes in microtubule polymer (Simon et al., 1998). It is therefore unlikely that phosphorylation of tubulin dimers plays a key role in the increase of microtubule formation in activated mast cells (Sulimenko et al., 2006).
It has been repeatedly reported that γ-tubulin is phosphorylated (Vogel et al., 2001; Stumpff et al., 2004; Alvarado-Kristensson et al., 2009). Phosphorylation of the γ-tubulin residue Tyr 445, which is invariably present in all γ-tubulins, was described and a mutation of this residue changed the microtubule dynamics (Vogel et al., 2001). Similarly phosphorylation of multiple serines on γ-tubulin can regulate microtubule organization (Lin et al., 2011). There are other data that point to an association of γ-tubulin with kinases. Phosphoinositide-3-kinase (PI3K) binds to γ-tubulin (Kapeller et al., 1995), and the regulatory subunits of PI3K interacts with γ-tubulin in various cell types (Inukai et al., 2000; Macurek et al., 2008), including BMMCs (V. Sulimenko, unpublished data). Collectively these data suggest that kinases take part in the regulation of γ-tubulin function(s). This could lead to changes in nucleation properties of centrosomes or alternatively to an enhancement of non-centrosomal microtubule nucleation.
Signaling Pathways for Stabilization of the Plus Ends of Microtubules
The formation of microtubules can also be regulated by stabilizing their plus ends. It has been shown that the Fyn/Gab2/RhoA signaling pathway, though not the Lyn/SLP-76, plays a critical role in microtubule-dependent degranulation of mast cells, and that RhoA kinase could be involved in stabilization of the plus ends of microtubules (Nishida et al., 2005). It is known that an important role in stabilization of growing microtubules is to be assigned to the +TIPs, whose interactions with microtubules are regulated by phosphorylation (Akhmanova and Steinmetz, 2008; Galjart, 2010). Ca2+-dependent kinases [e.g., conventional protein kinases C (cPKC); Ca2+/calmodulin-dependent kinases] or phosphatases (e.g., PP2B) might participate in the regulation of microtubule stability in activated BMMCs. It has been reported that Ca2-dependent activation of Rac (from the Rho family of small GTPases) depends on the activity of cPKC (Price et al., 2003). It was demonstrated that FcεRI stimulation of BMMCs activated RhoA (Nishida et al., 2005). Interestingly, this GTPase participates in stabilization of microtubule plus ends through its target mDia (Palazzo et al., 2001). Microtubule dynamics may also be influenced by +TIPs that interact at cell periphery with cytoskeletal scaffold IQ domain GTPase-activating protein1 (IQGAP1) which sequesters calmodulin and Rac. An increase in Ca2+ liberates calmodulin and IQGAP1 interacts with the A-kinase anchoring protein 220 (AKAP220). This leads to association of IQGAP1 with factors that modulate microtubule dynamics, e.g., CLIP-associated protein-2 (CLASP2). Recruitment of CLASP2 to AKAP220/IQGAP1 complex is regulated by glycogen synthase kinase-3β (GSK3β) and cAMP-dependent protein kinase (PKA) (Logue et al., 2011). It has been shown that FcεRI stimulation of BMMCs triggered the formation of microtubules in a manner independent of Ca2+ (Nishida et al., 2005). On the other hand, other results demonstrated Ca2+-dependent formation of protrusions containing microtubules (Hájková et al., 2011). This discrepancy could be attributed to differences in cell activation (absence or presence of integrin engagement) and various methods of preparation of samples for microscopic evaluation. However, it is also possible that the initial stages of microtubule formation as well as the transport of granules along microtubules are Ca2+-independent, but that later stages of activation and formation of microtubule protrusions depend on sustained influx of Ca2+.
Ras guanyl nucleotide-releasing proteins (RasGRPs) belong to a family of proteins that are recruited to the plasma membrane by binding to DAG for subsequent activation of Ras family proteins. GTP-bound Ras has been shown to interact with the catalytic p110 subunit of PI3K, and to induce its activation (Kodaki et al., 1994). PI3K then stimulates Rho GTPases (Kuehn et al., 2010). It was demonstrated that FcεRI activated RasGRP1-depleted BMMCs were defective in RhoA activation, microtubule formation and granule translocation. RasGRP1 could thus play an important role in the regulation of microtubule formation in activated mast cells (Liu et al., 2007).
Microfilaments
Microfilaments and Actins
Microfilaments (actin filaments) are essential for a number of cell functions, such as maintenance of cell shape, cell division, migration, junction formation, and intracellular vesicle trafficking powered by motor proteins. Microfilaments are formed by two-stranded helical polymers with a diameter of ∼7 nm. Actin filaments are assembled into two types of structures – bundles and networks. The basic component of microfilaments is a globular protein actin (called G-actin), which forms filaments (called F-actin). The molecule of actin is folded into two domains stabilized by an adenine nucleotide lying in between. The uniform orientation of asymmetrical subunits along the polymer causes polarization of actin filament with fast-growing plus-end (or barbed end) and slow-growing minus end (or pointed-end). Microfilaments are dynamic structures; subunits can be added or lost on both ends of the polymer. The different rate constants for association and dissociation depend on nucleotide bound to the monomer. Autocatalyzed hydrolysis of ATP to ADP in F-actin reduces the binding strength and modifies the behavior of microfilaments, including their affinity to regulatory proteins (Kreis and Vale, 1999).
Actin is one of the most highly conserved proteins in evolution. It is encoded by multiple genes. Mammals have six genes, and each encodes one protein isoform. Four of them, are expressed primarily in muscles. The remaining two isoforms (βcyto-actin, γcyto-actin) are expressed ubiquitously. All isoforms possess very similar amino acid sequences, with no isoform sharing less than 93% identity with any other isoform (Perrin and Ervasti, 2010). Although actin isoforms can copolymerize within the cells they are usually sorted out in different structures and perform different cellular functions due to interactions with specific subsets of actin-binding proteins (dos Remedios et al., 2003).
In contrast to microtubules, microfilaments do not have one prominent nucleation center. They can form tight bundles making up the core of microvilli or can be arranged in a less-ordered network as in the cell cortex. In dividing cells they form the contractile ring that is important for cytokinesis. In migrating cells, a branched network of actin filaments is found in the lamellipodia, broad and flat protrusions at the leading edge of moving cells. The thin cellular processes extending from lamellipodium and containing parallel bundles of actin filaments are called filopodia. Many cells have contractile filaments called stress fibers that consist of microfilaments, myosin II motor proteins and other interacting proteins. The ends of stress fibers anchor to the cell membrane in focal adhesions, cell-to-substrate adhesion structures accountable for a strong attachment to substrate (Parsons et al., 2010). There are three categories of stress fibers: ventral stress fibers that are attached to focal adhesions at both ends, dorsal stress fibers that are attached to focal adhesions typically at one end, and transverse arcs that do not directly attach to focal adhesions (Naumanen et al., 2008). Microfilaments are also important for the formation of transient membrane surface structures as podosomes and invadopodia, which attach cells to the extracellular matrix, and take part in the generation of circular dorsal ruffles (Buccione et al., 2004). The stability and dynamics of actin structures is regulated by a variety of ligands including drugs and toxins. Phalloidins bind along the sides of actin filaments, preventing their depolymerization. Phalloidin labeled with a fluorophore is often used for visualization of microfilaments. Jasplakinolides are other actin-filament stabilizers. On the other hand, cytocholasins cap the barbed end of microfilaments, and latrunculins sequester actin monomers. These processes lead to inhibition of microfilament formation (Kustermans et al., 2008).
Actin-binding Proteins Regulate Microfilament Organization and Dynamics
Actin binds a substantial number of associated proteins. They can be divided into several functional groups. The first group contains monomer-binding proteins that sequester G-actin and prevent its polymerization (e.g., profilin). The second group covers filament-depolymerizing proteins (e.g., ADF/cofilin). The third group includes filament end-binding proteins that cap the ends of actin filament and prevent the exchange of monomers at the pointed-end (e.g., tropomodulin) and at the barbed end (e.g., CapZ). The fourth group contains filament severing proteins (e.g., gelsolin). The fifth group consists of cross-linking proteins that contain at least two binding sites for F-actin, facilitating thus the formation of filament bundles, branching filaments, and three-dimensional networks (e.g., α-actinin, vilin, fimbrin, Arp2/3). In the sixth group are stabilizing proteins that bind to the sides of actin filaments and prevent depolymerization (e.g., tropomyosin). Finally, the seventh group consists of motor proteins that use F-actin as a track upon which to move (e.g., myosins) (dos Remedios et al., 2003). Actin-binding proteins are functionally not limited to one class; for example, gelsolin is capable of severing and capping the barbed end of actin filaments.
Cortical F-actin is connected to the plasma membrane through an array of closely related cytoplasmic proteins of the ERM (Ezrin, Radixin, Moesin) family. In addition to their role in binding filamentous actin, ERMs regulate the signaling pathways through their ability to bind transmembrane receptors and link them to downstream signaling components. ERMs thus serve as scaffolds to facilitate efficient signal transduction on the cytoplasmic face of the plasma membrane (Neisch and Fehon, 2011).
Inside the cells, the de novo nucleation of actin filaments from monomers require the participation of actin-nucleating proteins. These proteins fall into three main families: the Arp2/3 complex and its nucleation promoting factors (NPFs), formins, and tandem-monomer-binding nucleators. They have important roles in many essential cellular processes. The Arp2/3 complex is composed of evolutionarily conserved subunits including the actin-related proteins Arp2, Arp3, and five additional subunits ARPC1–5. The Arp2/3 complex by itself is an inefficient nucleator, and its activation requires binding to the sides of actin filaments and to NPF (Goley and Welch, 2006). Among the well characterized NPFs are Wiskott-Aldrich syndrome protein (WASP), Wiskott-Aldrich syndrome protein-family verprolin homologous protein (WAVE), WASP and Scar homolog (WASH), and the more recently identified WASP homolog associated with actin, membranes, and microtubules (WHAMM; Firat-Karalar and Welch, 2011). Once activated, the Arp2/3 complex nucleates the formation of new filaments that extend from the sides of existing filaments at a 70° to form a Y-branched network. In contrast, nucleation by formins (e.g., mammalian Diaphanous formin, mDia) and tandem-monomer-binding nucleators (e.g., adenomatous polyposis coli, APC) leads to unbranched filaments (Firat-Karalar and Welch, 2011). Arp2/3 complex activity is inhibited by coronin 1b that promotes debranching of actin filaments and recycling of the Arp 2/3 complex (Cai et al., 2007). Similarly, cofilin also dissociates Arp2/3 complex and branches from actin filaments (Chan et al., 2009).
Rho GTPases as Key Regulators of Microfilaments
Diverse cell-surface receptors trigger global structural rearrangements of actin cytoskeleton in response to external signals. These signals converge inside the cell on a group of closely related monomeric GTPases that belong to the Rho protein family. The three prevailing subclasses are Rho, Rac, and Cdc42. Rho activates the formation of stress fibers and focal adhesions (Ridley and Hall, 1992), Rac activates the formation of lamellipodia and membrane ruffles (Ridley et al., 1992) and Cdc42 activates the formation of filopodia (Nobes and Hall, 1995). In each case the active form of protein binds GTP. Down regulation of Rho GTPases involves GTPase-activating proteins (GAPs) that facilitate GTP hydrolysis. Activation of Rho proteins is promoted by guanine nucleotide exchange factors (GEFs) that control the release of GDP from the Rho protein and its replacement with GTP. GEFs are often activated through signal cascades initiated via plasma membrane receptors. Guanine nucleotide dissociation inhibitors (GDI) bind to Rho and prevent GDP/GTP exchange (Bement et al., 2006). GTPases also function as molecular switches in multiple signaling processes including regulation of phospholipase D (PLD) and PI3K (Bishop and Hall, 2000).
The downstream targets of Rho GTPases include kinases, formins, families of WASP proteins, and other scaffolding molecules. Of these major subclasses, the Rho-associated coiled-coil kinase (ROCK), the p21-activated kinase (PAK), the mDia, and proteins of the WASP and WAVE families, have direct effects on actin cytoskeleton rearrangements. In addition to activation of formins promoting actin-filament growth, Rho GTPase also promotes myosin actin interactions inducing development and contraction of stress fibers through ROCK. ROCK has been shown to directly phosphorylate a number of actin cytoskeleton regulators including myosin II light chain (MLC), myosin light chain phosphatase (MLCP), and LIM-motif containing kinase (LIMK). Direct phosphorylation of MLC or MLCP has an immediate impact on the level of phosphorylated myosin light chain, which enhances the contractility. Activation of LIMK by ROCK is linked to phosphorylation and inhibition of cofilin, thereby regulating the actin-filament turnover (Spiering and Hodgson, 2011). Feeding directly into this pathway, Rac and Cdc42 activate PAK and consecutively also LIMK. Rac activates the ARP2/3 complex through the WAVE complex, and CDC42 induces actin polymerization by binding to WASP (Heasman and Ridley, 2008). Localization of WASP can be regulated by interaction with other binding partners including Grb2, likely targeting the molecule to sites of receptor stimulation and active actin cytoskeleton remodeling (Carlier et al., 2000). Interestingly, mDia proteins through association with +TIPs has been shown to stabilize microtubules, tying the actin cytoskeleton rearrangements to the microtubule dynamics (Bartolini et al., 2008). Downstream effector targets of the Rho family of GTPases for generation of actin cytoskeletal structures are schematically shown in Figure 7.
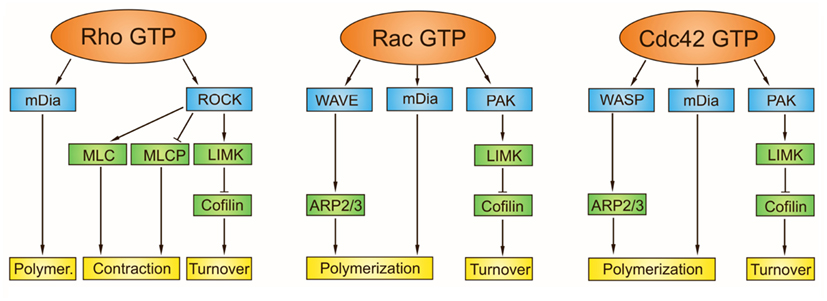
Figure 7. Downstream effector targets of Rho family GTPases for generation of actin cytoskeletal structures. The downstream targets of active Rho GTPases, with bound GTP, include kinases (p21-activated kinase, PAK; Rho-associated coiled-coil kinase; ROCK) and nucleation promoting factors (mammalian Diaphanous formin, mDia; Wiskott-Aldrich syndrome protein, WASP; Wiskott-Aldrich syndrome protein-family verprolin homologous protein, WAVE). While nucleation by mDia produces unbranched actin filaments, WASP and WAVE interact with Arp2/3 complex and generate branched microfilaments. PAK phosphorylates LIM-motif containing kinase (LIMK), that in turn phosphorylates and inhibits cofilin thereby regulating the actin-filament turnover. Besides stimulating the actin-filament growth, Rho GTPase promotes myosin actin interactions through ROCK. The ROCK phosphorylates a number of actin cytoskeleton regulators including myosin II light chain (MLC), myosin light chain phosphatase (MLCP), and LIMK. Direct phosphorylation of MLC or MLCP has an immediate impact on the level of phosphorylated myosin light chain, which contributes to contractility. Rho mainly activates the formation of stress fibers and focal adhesions, Rac activates the formation of lamellipodia and membrane ruffles, and Cdc42 activates the formation of filopodia.
Microfilaments in Activated Mast Cells
Changes in Actin-Filament Organization
Dramatic reorganization of actin cytoskeleton has been observed in mast cells activated by FcεRI aggregation. The F-actin content of the detergent-extracted cell matrices in RBL-2H3 cells decreased during the first 10–30 s after antigen binding and then increased within 1 min to almost double the control levels. The antigen-stimulated increase in F-actin coincided with the transformation of cell-surface from a finely microvillous to a highly folded topography, and with increased cell spreading (Pfeiffer et al., 1985). Enhanced formation of F-actin after cell triggering was reported in several other studies using RBL-2H3 cells (Apgar, 1994; Frigeri and Apgar, 1999; Holowka et al., 2000; Tolarová et al., 2004), RPMCs (Pendleton and Koffer, 2001), and BMMCs (Tumová et al., 2010). Sensitization with IgE alone resulted in an enhancement of cortical F-actin in RBL-2H3 cells (Oka et al., 2004) and BMMCs (Allen et al., 2009).
Inhibitors that disrupt microfilaments, such as latrunculin and cytochalasin D, did not by themselves cause any degranulation in RBL-2H3 cells, but they enhanced the FcεRI-mediated degranulation. However, latrunculin was ineffective if pervanadate was used for activation. This suggests that microfilaments are indispensible for the downstream signaling cascade (Frigeri and Apgar, 1999). Good correlation was found between inhibition of actin polymerization and increased activity of tyrosine kinase and phospholipase. Microfilaments appear to down-regulate the response by affecting the level of receptor tyrosine phosphorylation (Frigeri and Apgar, 1999; Holowka et al., 2000). Similarly, FcεRI-mediated degranulation was enhanced in BMMCs treated with latrunculin B, while jasplakinolide slightly inhibited β-hexosaminidase release (Nishida et al., 2005). It was proposed that actin controls/modulates a multitude of signaling steps leading from the FcεRI aggregation to the final degranulation (Pendleton and Koffer, 2001; Oka et al., 2002; Andrews et al., 2008).
Microfilaments are crucial not only in early stages of mast cell activation but also in later exocytosis steps. Mast cell exocytosis is accompanied with extensive reorganization of the actomyosin cortex, and FcεRI stimulation triggers F-actin ring disassembly in a Ca2+-dependent manner (Nishida et al., 2005). Both positive and negative roles have been suggested for actin in exocytosis. Actomyosin may facilitate the transport and/or extrusion of secretory vesicles, while cortical F-actin may serve as a barrier preventing the access of vesicles to the plasma membrane (Pendleton and Koffer, 2001). Alternatively, cortical F-actin may act as a barrier between the reserve and the release-ready secretory vesicle pools. F-actin disassembly may cause disruption of this barrier (Malacombe et al., 2006).
The combination of atomic force microscopy and laser scanning confocal microscopy enabling simultaneous visualization and correlation of membrane morphology with actin arrangement revealed that reorganization of microfilaments in activated BMMCs depended on the type of stimuli used for cell activation. In cells triggered by FcεRI aggregation, characteristic membrane ridges formed in accordance with the rearrangement of underlying F-actin networks. On the other hand, in BMMCs stimulated just only by poly-L-lysine coated on glass surface, lamellopodia and filopodia were observed in association with the F-actin assemblies at and near the cell periphery, whereas “craters” occurred on the central membrane lacking F-actin (Deng et al., 2009). Actin reorganization in activated cells is also strongly modulated by cell adherence to proteins of extracellular matrix (e.g., fibronectin) followed by integrin signaling (Hamawy et al., 1992, 1994). It was reported that co-stimulation of FcεRI and chemokine receptor 1 resulted in generation of long and thin tubes formed from plasma membrane (cytonemes). Cytonemes containing F-actin may participate in intercellular communication during allergic and inflammatory response (Fifadara et al., 2010). Organization of F-actin in non-activated and thapsigargin-activated BMMCs attached to fibronectin is illustrated in Figure 5.
Rho GTPases in Regulation of Actin Filaments in Activated Cells
In mast cells, small Rho-related GTPases are essential to the exocytotic function (Price et al., 1995; Brown et al., 1998). Expression of dominant negative mutant forms of the Cdc42 and Rac1 in RBL-2H3 cells substantially affected the microfilament reorganization. Inhibition of Cdc42 function decreased cell adhesion, interfered with FcεRI-induced actin plaque assembly and reduced the recruitment of vinculin at the cell-substratum interface. On the other hand, the inhibitory Rac1 mutant abolished FcεRI-mediated membrane ruffling. The expression of inhibitory forms of either Cdc42 or Rac1 significantly inhibited the antigen-induced degranulation. Cdc42 and Rac1 thus control distinct pathways downstream of FcεRI engagement leading to microfilament organization. Both pathways are decisively engaged during the degranulation response induced by FcεRI aggregation (Guillemot et al., 1997). Mast cells derived from Rac 2-deficient mice showed defects in actin-based functions, including adhesion, migration, and degranulation (Yang et al., 2000).
Rho was found to be responsible for de novo actin polymerization in RPMCs (Norman et al., 1994), and it also participated in the control of cortical actin disassembly (Sullivan et al., 1999). Stimulation of BMMCs by FcεRI aggregation resulted in an increased activity of Rac (1,2,3) but decreased activity of RhoA followed by slow recovery of the latter. It was suggested that active RhoA may be necessary to initiate the secretory process (perhaps by activating a fusogen), and its immediate subsequent inactivation may be required for cell attachment (Arthur et al., 2000) and/or F-actin disassembly (Norman et al., 1996). The later increase in RhoA activity would then be required for the formation of newly polymerized actin inhibiting exocytosis and promoting cell spreading and migration (Struckhoff et al., 2010). In the pathway utilizing RhoA and ROCK, the adaptor protein NTAL could serve as a positive regulator of actin polymerization and cell spreading (Tumová et al., 2010).
Proteins that become activated during the c-Kit signaling include PI3K and Rac; activation of this pathway triggers actin reorganization. An important PI3K-dependent GEF interacting with Rac is switch-associated protein 70 (SWAP-70), that affects ruffle formation in activated cells. SWAP-70-deficient BMMCs had impaired Rac function and defects in adhesion, migration, and degranulation. Stimulated mutant cells displayed filopodia-like protrusions possibly indicating a shift in balance between Rac and Cdc42 (Sivalenka and Jessberger, 2004). Enhanced activity of PI3K in c-Kit activated cells leads to activation of Bruton’s tyrosine kinase (Btk) and Rac. Btk-deficient BMMCs had defects in Rac activation and F-actin rearrangement resulting in impaired migration and chemotaxis. The Btk is not just an important upstream regulator of Ras GTPase but integrates in addition the signaling pathways from activated c-Kit, GPCR, and aggregated FcεRI (Kuehn et al., 2010).
Wiskott-Aldrich syndrome protein interacting protein (WIP) is another important regulator of the actin cytoskeleton that inhibits Cdc42-mediated activation of WASP. Most of WASP in lymphocytes appear to be sequestered with WIP, and binding to WIP is essential for the stability of WASP (Ramesh and Geha, 2009). More efficient generation of actin filaments was detected in WIP-deficient BMMCs activated by FcεRI aggregation (Kettner et al., 2004). Impaired actin polymerization, cell spreading, formation of ruffles and degranulation were observed in WASP-deficient BMMCs stimulated by FcεRI signaling (Pivniouk et al., 2003). All these data strongly indicate that Rho GTPases and their effectors have a major role in organization of microfilaments in activated mast cells.
Regulation of F-actin in Activated Cells by Changes in Ca2+ Concentration
Rapid dissolution of cortical F-actin following FcεRI stimulation of mast cells has been interpreted in many studies as a crucial step in facilitating the granule-plasma membrane fusion necessary for the release of granule contents to extracellular environment (Nielsen et al., 1989; Narasimhan et al., 1990; Frigeri and Apgar, 1999; Oka et al., 2004; Deng et al., 2009). Cortical F-actin disassembly is, however, also required for the motile responses. Numerous studies further also revealed that the influx of Ca2+ from extracellular space is a prerequisite for cortical F-actin depolymerization (Koffer et al., 1990; Nishida et al., 2005; Shimizu et al., 2009; Suzuki et al., 2010). In contrast, other reports concluded that cortical F-actin disassembly can be induced by both Ca2+-dependent and Ca2+-independent pathways (Sullivan et al., 1999; Guzman et al., 2007). More prominent F-actin depolymerization was observed in NTAL-depleted BMMCs simultaneously activated by FcεRI aggregation and SCF, when compared to activated BMMCs controls. However, the influx of Ca2+ was basically unchanged (Tumová et al., 2010).
One of the possible mechanisms of Ca2+-dependent cortical F-actin depolymerization is based on activation of severing protein gelsolin (Borovikov et al., 1995). Another actin-severing protein from the gelsolin family, strongly upregulated in mouse mast cells, is adseverin (D5) (Robbens et al., 1998). Calmodulin also plays an important role in Ca2+-dependent disassembly of microfilaments. This multifunctional protein transduces Ca2+ signals by binding the Ca2+ and interacting then with various target proteins. Ca2+-calmodulin causes changes in the activity of target proteins. In this connection, it should be mentioned that the disassembly of the actin cortex in RPMC is preceded by acto-myosin-II-based contraction, activated by Ca2+-calmodulin/myosin light chain kinase (MLCK) that phosphorylates the regulatory light chain of myosin II (MLC), activating the motor protein. Inhibition of myosin II significantly inhibited the Ca2+-evoked cortical actin disassembly. Thus, calmodulin plays an essential role in Ca2+-induced cortical F-actin disassembly (Sullivan et al., 2000). In activated RBL-2H3 cells calmodulin concentrated in actin-rich cell cortex (Psatha et al., 2004) where it was recruited by an interaction with the actin-binding scaffold protein IQGAP1 that sequesters Rho GTPases Rac and Cdc42 in GTP-bound state. Rho GTPases are released from IQGAP1 when IQGAP1-associated calmodulin binds Ca2+. Calcium elevation therefore not only initiates the disassembly of the existing cortical actin cytoskeleton but simultaneously releases the GTP-loaded Cdc42 and Rac1 to mediate actin-based cell spreading and ruffling (Psatha et al., 2007).
Role of Kinases and Phosphatases in Regulation of Microfilaments in Activated Cells
Activation of mast cells via both FcεRI and c-Kit receptors is mediated and regulated by numerous protein tyrosine and serine/threonine kinases which transmit the downstream signals. It has been proposed that Fyn kinase, which in BMMCs participates in signal transduction from activated FcεRI, phosphorylates the non-selective Ca2+ channel TRPC1 (transient receptor potential cation channel subfamily C). This results in modulation of Ca2+ influx and F-actin depolymerization (Suzuki et al., 2010).
PI3K regulates microfilament organization in BMMCs by activation of Btk that in turn induces Rac-dependent F-actin rearrangement and enhancement of the Ca2+ signal. This in combination leads to synergy in mast cell chemotaxis (Kuehn et al., 2010). In BMMCs stimulated by PGE2, PI3K activates Ser-Thr kinase, mammalian target of rapamycin (mTOR). mTOR binds to specific regulator, named rictor, to form mTORC2 complexes in association with other binding partners (Foster and Fingar, 2010). Interestingly, mTORC2 stimulated both polymerization of actin and chemotaxis, though by a mechanism independent of Ca2+ mobilization and Rac acctivation (Kuehn et al., 2011).
An important players are also members of p21-activated kinases (PAKs) which receive independent signals from both PI3K and Rho GTPases. PAKs induce the temporal and spatial formation of cortical actin structures similar to those regulated by Rac and Cdc42, including membrane ruffles, lamellipodia filopodia, and focal complexes (Sells et al., 2000). PAK 1-defficient BMMCs showed diminished Ca2+ mobilization, degranulation, and altered depolymerization of cortical F-actin in response to FcεRI stimulation. While cortical F-actin ring formed as expected in stimulated cells, subsequent fragmentation of F-actin occurred to a much smaller extent than in normal BMMCs (Allen et al., 2009). PAK 1 can modulate F-actin turnover by the well established LIMK/coffilin pathway, yet it can also phosphorylate Ser-Thr protein phosphatase 2A (PP2A) that is involved in vesicular fusion in mast cells, and transiently associate with cortical myosin II (Blank and Rivera, 2004).
New roles in regulation of microfilaments were recently ascribed tor cPKCs that are activated by DAG and by Ca2+. Some data showed that cPKCβ in FcεRI-stimulated RBL-2H3 cells colocalized with F-actin in membrane-ruffled regions, and inhibition of cPKCβ resulted in the blocking of actin ruffle formation and adhesion (Yanase et al., 2011). Similar effects were observed in activated BMMCs, where furthermore migration toward SCF was suppressed (Yanase et al., 2011). In antigen-stimulated RBL-2H3 cells the cPKCβII phosphorylated non-muscle myosin heavy chain IIA, and the time course of phosphorylation correlated with degranulation (Ludowyke et al., 2006).
Recent data also confirmed the significant role of protein tyrosin phosphatases (PTPs) in regulation of microfilaments in activated mast cells. When antibody specific for oxidized (inactive) phosphatases was used and isolated membranes were examined by electron microscopy, clear colocalization of oxidized PTPs with cytoskeleton-like structures was observed in non-activated mast cells. Enhanced association of oxidized phosphatases with these structures was observed in cells activated by antigen or by pervanadate. The same structures were labeled with phalloidin. This indicates that oxidized phosphatases are preferentially associated with actin cytoskeleton (Heneberg et al., 2010). These data suggest that actin cytoskeleton is involved in early signaling events by regulating the topography of phosphatases. Alternatively, actin could play a role in sequestering and/or scavenging irreversibly oxidized PTPs (Bugajev et al., 2010). In this connection it should be mentioned that actin is probably effective in sequestering Lyn kinase to the periphery of large FcεRI aggregates which are subsequently internalized (Wilson et al., 2000).
One of the phosphatases associated with F-actin and actin-binding protein filamin is the SH2 domain containing protein tyrosine phosphatase-1 (SHP-1). This complex associated with FcεRI coaggregated with FcγRIIB. Dissociation of actin and filamin-1 from the FcR complex caused activation of FcR-bound SHP-1 and inhibition of FcεRI-stimulated signal in RBL-2H3 cells (Lesourne et al., 2005).
As expected, phosphorylation of actin associated proteins affects microfilament dynamics in activated mast cells. An important family of such proteins are coronins. Their regulatory effects include the binding/bundling, disassembly, and inhibition of the Arp2/3 complex. Recent data showed that FcεRI-mediated phosphorylation of coronin 1a in peritoneal mast cells resulted in its relocation from the cell cortex to cytoplasm. This relocation was accompanied with reduced cortical actin stability and enhanced degranulation (Foger et al., 2011).
Intermediate Filaments
Intermediate filaments provide crucial structural support in the cytoplasm and nucleus; any disturbance of intermediate filaments causes cell fragility. Intermediate filaments also play a role in determining cellular architecture, cell migration, and signal modulation. More than 70 conserved genes encode intermediate filament proteins that can self-assemble into 10-nm wide filaments. Each protein has N- and C-terminal end domains, termed “head” and “tail,” respectively, surrounding the α-helical rod domain. The basic building-block for intermediate filaments is a parallel dimer formed by the winding of α-helical rods into coiled coil. Dimers then associate along their lateral surfaces with anti-parallel orientation and form apolar staggered tetramers. A tetramer represents the soluble subunit analogous to the αβ-tubulin dimer or actin monomer. However, unlike actin or tubulin, the intermediate filament subunits do not contain binding sites for nucleoside triphosphate (Coulombe and Wong, 2004). The final 10-nm filament is a helical array of tetramers. The anti-parallel orientation of tetramers means that, contrary to microtubules and microfilaments, intermediate filaments do not bear polarized unidirectional properties. Assembly and disassembly is regulated by cycles of phosphorylation and dephosphorylation. Polymerization occurs rapidly irrespective of nucleating or associated proteins. In contrast to microtubules and microfilaments, intermediate filaments do not serve as tracks for molecular motors (Kim and Coulombe, 2007).
Intermediate filament proteins are subcategorized into six types (I-VI), based on similarities in amino acid sequences and protein structure, and individual types have different cell and tissue distribution. Vimentin (type III intermediate filament protein) is the most widely distributed type that can be found in mesenchymal and connective tissue cells. Vimentin plays a significant role in supporting and anchoring the position of organelles in the cytosol. Vimentin filaments possess unique viscoelastic physical properties that render them more resistant to mechanical stress in comparison to microtubules and microfilaments (Kreis and Vale, 1999). The mechanical strength of vimentin filaments network appears to be further enhanced after interactions with microtubules (Dráberová and Dráber, 1993) and microfilaments mediated by a family of multifunctional intermediate filament-associated proteins (Green et al., 2005). Within the cells vimentin exists in multiple structural forms including mature filaments, short filaments called “squiggles,” and non-filamentous precursors called “particles.” The forms are intercovertible and their relative abundance is cell type- and cell cycle stage-dependent. These structures are often associated with microtubule motor proteins, and are therefore capable of translocating along microtubules (Chou et al., 2007).
Intermediate Filaments in Activated Mast Cells
Data on the role of intermediate filaments during mast cell activation events are limited. The key building protein of intermediate filaments in mast cells is vimentin (Horny et al., 1988). It was reported that vimentin was phosporylated on serine after activation of RPMC with compound 48/80. Phosphorylation took place within 5 s of stimulation and reached its maximum in 10 s. When cells were treated with calphostin C, a specific inhibitor of protein kinase C, phosphorylation was markedly reduced. Interestingly, cells stained with anti-vimentin antibody showed filaments surrounding granules in cytoplasm; after stimulation these filaments promptly disappeared, indicating rapid depolymerization (Izushi et al., 1992). It was assumed that the disruption of intermediate filaments took place after stimulation with compound 48/80 as a consequence of vimentin phosphorylation. The absence of filaments surrounding granules facilitated then the movement of granules toward the cell membrane and degranulation (Tasaka, 1994). Mass spectrometric analysis of proteins binding to the SH2 and SH3 domains of Fyn led to identification of vimentin as the binding partner to Fyn in MC/9 mast cells. After IgE-receptor mediated stimulation, binding of vimentin to Fyn increased; this interaction occurred via binding to the SH2, but not SH3, domain of Fyn. Mast cells from vimentin-deficient mice showed an increase in mediator release and tyrosine phosphorylation of intracellular proteins including NTAL and LAT. These results suggest that vimentin association with Fyn may have a negative regulatory role in mast cell degranulation and tyrosine phosphorylation of signaling molecules induced by FcεRI stimulation (Nahm et al., 2003). Lymphocytes from vimentin-deficient mice are subject to greater deformation than lymphocytes from wild type mice, and vimentin was therefore suggested to be the primary source of lymphocyte rigidity (Brown et al., 2001). It may be that more deformable mast cells degranulate more easily following FcεRI aggregation. The observation that vimentin binds to Fyn points to a critical role for Fyn in mast cell degranulation during interaction with both cytosolic and structural proteins (Nahm et al., 2003).
Concluding Remarks
A broad range of experimental findings shows that cytoskeletal proteins in activated mast cells play an important role in propagation of the signals from stimulated cell-surface receptors FcεRI, c-Kit, or GPCR. Cytoskeletal proteins are also accountable for observed prominent changes in cell morphology, adhesion to substrates, degranulation, and migration. Some controversial results mentioned in this review probably reflect variations in the origin of mast cells, culture conditions, cell activation, methods used for visualization of cytoskeletal structures, and possibly others. The crosstalk between microtubules, microfilaments, and intermediate filaments in the course of mast cell activation is still an open question. Various cytoskeletal scaffolding proteins with numerous protein interaction domains and integrating signals from kinases/phosphatases, Rho GTPases and changes in Ca2+concentration apparently affect the regulatory mechanisms of interactions between microtubules and microfilaments. The roles of posttranslational modifications of tubulins and MAPs in nucleation and modulation of microtubule dynamics in activated cells are other fields with many unsolved problems. Similarly, little is known about the role of motor proteins and their regulation in granule transport, and cytoskeleton reorganization during activation events. New innovative techniques of live cell imaging will undoubtedly play a growing role in studies on cytoskeleton dynamics in activated mast cells under physiological conditions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Work on mast cell cytoskeleton in the laboratory of the authors has been funded, over the years, by several grants, including P302/12/1673 (Pavel Dráber), P302/10/1701 (Eduarda Dráberová), and P302/11/P709 (Vadym Sulimenko) from the Grant Agency of the Czech Republic, by the Institutional Research Support (AVOZ 50520514); and Action BM1007 (Mast Cells and Basophils – Targets for Innovative Therapies) from European Cooperation in Science and Technology.
References
Akhmanova, A., and Steinmetz, M. O. (2008). Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 9, 309–322.
Allen, J. D., Jaffer, Z. M., Park, S. J., Burgin, S., Hofmann, C., Sells, M. A., Chen, S., Derr-Yellin, E., Michels, E. G., McDaniel, A., Bessler, W. K., Ingram, D. A., Atkinson, S. J., Travers, J. B., Chernoff, J., and Clapp, D. W. (2009). p21-Activated kinase regulates mast cell degranulation via effects on calcium mobilization and cytoskeletal dynamics. Blood 113, 2695–2705.
Alvarado-Kristensson, M., Rodriguez, M. J., Silio, V., Valpuesta, J. M., and Carrera, A. C. (2009). SADB phosphorylation of gamma-tubulin regulates centrosome duplication. Nat. Cell Biol. 11, 1081–1092.
Andrews, N. L., Lidke, K. A., Pfeiffer, J. R., Burns, A. R., Wilson, B. S., Oliver, J. M., and Lidke, D. S. (2008). Actin restricts FcεRI diffusion and facilitates antigen-induced receptor immobilization. Nat. Cell Biol. 10, 955–963.
Apgar, J. R. (1994). Polymerization of actin in RBL-2H3 cells can be triggered through either the IgE receptor or the adenosine receptor but different signaling pathways are used. Mol. Biol. Cell 5, 313–322.
Arthur, W. T., Petch, L. A., and Burridge, K. (2000). Integrin engagement suppresses RhoA activity via a c-Src-dependent mechanism. Curr. Biol. 10, 719–722.
Baba, Y., Hayashi, K., Fujii, Y., Mizushima, A., Watarai, H., Wakamori, M., Numaga, T., Mori, Y., Iino, M., Hikida, M., and Kurosaki, T. (2006). Coupling of STIM1 to store-operated Ca2+ entry through its constitutive and inducible movement in the endoplasmic reticulum. Proc. Natl. Acad. Sci. U.S.A. 103, 16704–16709.
Bakowski, D., Glitsch, M. D., and Parekh, A. B. (2001). An examination of the secretion-like coupling model for the activation of the Ca2+ release-activated Ca2+ current I(CRAC) in RBL-1 cells. J. Physiol. Paris 532, 55–71.
Bartolini, F., Moseley, J. B., Schmoranzer, J., Cassimeris, L., Goode, B. L., and Gundersen, G. G. (2008). The formin mDia2 stabilizes microtubules independently of its actin nucleation activity. J. Cell Biol. 181, 523–536.
Bement, W. M., Miller, A. L., and von Dassow, G. (2006). Rho GTPase activity zones and transient contractile arrays. Bioessays 28, 983–993.
Bishop, A. L., and Hall, A. (2000). Rho GTPases and their effector proteins. Biochem. J. 348, 241–255.
Blank, U., and Rivera, J. (2004). The ins and outs of IgE-dependent mast-cell exocytosis. Trends Immunol. 25, 266–273.
Borovikov, Y. S., Norman, J. C., Price, L. S., Weeds, A., and Koffer, A. (1995). Secretion from permeabilised mast cells is enhanced by addition of gelsolin: contrasting effects of endogenous gelsolin. J. Cell Sci. 108, 657–666.
Brown, A. M., O’Sullivan, A. J., and Gomperts, B. D. (1998). Induction of exocytosis from permeabilized mast cells by the guanosine triphosphatases Rac and Cdc42. Mol. Biol. Cell 9, 1053–1063.
Brown, M. J., Hallam, J. A., Colucci-Guyon, E., and Shaw, S. (2001). Rigidity of circulating lymphocytes is primarily conferred by vimentin intermediate filaments. J. Immunol. 166, 6640–6646.
Buccione, R., Orth, J. D., and McNiven, M. A. (2004). Foot and mouth: podosomes, invadopodia and circular dorsal ruffles. Nat. Rev. Mol. Cell Biol. 5, 647–657.
Bueb, J. L., Da, S. A., Mousli, M., and Landry, Y. (1992). Natural polyamines stimulate G-proteins. Biochem. J. 282, 545–550.
Bugajev, V., Bambousková, M., Dráberová, L., and Dráber, P. (2010). What precedes the initial tyrosine phosphorylation of the high affinity IgE receptor in antigen-activated mast cell? FEBS Lett. 584, 4949–4955.
Cai, L., Marshall, T. W., Uetrecht, A. C., Schafer, D. A., and Bear, J. E. (2007). Coronin 1B coordinates Arp2/3 complex and cofilin activities at the leading edge. Cell 128, 915–929.
Calloway, N., Vig, M., Kinet, J. P., Holowka, D., and Baird, B. (2009). Molecular clustering of STIM1 with Orai1/CRACM1 at the plasma membrane depends dynamically on depletion of Ca2+ stores and on electrostatic interactions. Mol. Biol. Cell 20, 389–399.
Carlier, M. F. (1991). Nucleotide hydrolysis in cytoskeletal assembly. Curr. Opin. Cell Biol. 3, 12–17.
Carlier, M. F., Nioche, P., Broutin-L’Hermite, I., Boujemaa, R., Le, C. C., Egile, C., Garbay, C., Ducruix, A., Sansonetti, P., and Pantaloni, D. (2000). GRB2 links signaling to actin assembly by enhancing interaction of neural Wiskott-Aldrich syndrome protein (N-WASp) with actin-related protein (ARP2/3) complex. J. Biol. Chem. 275, 21946–21952.
Chabin-Brion, K., Marceiller, J., Perez, F., Settegrana, C., Drechou, A., Durand, G., and Pous, C. (2001). The Golgi complex is a microtubule-organizing organelle. Mol. Biol. Cell 12, 2047–2060.
Chan, C., Beltzner, C. C., and Pollard, T. D. (2009). Cofilin dissociates Arp2/3 complex and branches from actin filaments. Curr. Biol. 19, 537–545.
Chou, Y. H., Flitney, F. W., Chang, L., Mendez, M., Grin, B., and Goldman, R. D. (2007). The motility and dynamic properties of intermediate filaments and their constituent proteins. Exp. Cell Res. 313, 2236–2243.
Colello, D., Reverte, C. G., Ward, R., Jones, C. W., Magidson, V., Khodjakov, A., and LaFlamme, S. E. (2010). Androgen and Src signaling regulate centrosome activity. J. Cell Sci. 123, 2094–2102.
Coulombe, P. A., and Wong, P. (2004). Cytoplasmic intermediate filaments revealed as dynamic and multipurpose scaffolds. Nat. Cell Biol. 6, 699–706.
Deng, Z., Zink, T., Chen, H. Y., Walters, D., Liu, F. T., and Liu, G. Y. (2009). Impact of actin rearrangement and degranulation on the membrane structure of primary mast cells: a combined atomic force and laser scanning confocal microscopy investigation. Biophys. J. 96, 1629–1639.
Desai, A., and Mitchison, T. J. (1997). Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 13, 83–117.
dos Remedios, C. G., Chhabra, D., Kekic, M., Dedova, I. V., Tsubakihara, M., Berry, D. A., and Nosworthy, N. J. (2003). Actin binding proteins: regulation of cytoskeletal microfilaments. Physiol. Rev. 83, 433–473.
Dráber, P., and Dráberová, E. (2012). “Microtubules,” in Cytoskeleton and Human Disease, ed. M. Kavallaris (New York, NY: Humana Press), 29–54.
Dráber, P., Hálová, I., Levi-Schaffer, F., and Dráberová, L. (2012). Transmembrane adaptor proteins in the high-affinity IgE receptor signaling. Front. Immunol. 2:95. doi:10.3389/fimmu.2011.00095
Dráberová, E., and Dráber, P. (1993). A microtubule-interacting protein involved in coalignment of vimentin intermediate filaments with microtubules. J. Cell Sci. 106, 1263–1273.
Dráberová, L., Dráberová, E., Surviladze, Z., Dráber, Pe., and Dráber, P. (1999). Protein tyrosine kinase p53/p56lyn form complexes with γ-tubulin in rat basophilic leukemia cells. Int. Immunol. 11, 1829–1839.
Dryková, D., Sulimenko, V., Cenklová, V., Volc, J., Dráber, P., and Binarová, P. (2003). Plant γ-tubulin interacts with αβ-tubulin dimers and forms membrane-associated complexes. Plant Cell 15, 465–480.
Dziadek, M. A., and Johnstone, L. S. (2007). Biochemical properties and cellular localisation of STIM proteins. Cell Calcium 42, 123–132.
Eiseman, E., and Bolen, J. B. (1992). Engagement of the high-affinity IgE receptor activates Src protein-related tyrosine kinases. Nature 355, 78–80.
Faruki, S., Geahlen, R. L., and Asai, D. J. (2000). Syk-dependent phosphorylation of microtubules in activated B-lymphocytes. J. Cell Sci. 113, 2557–2565.
Fernandez, J. A., Keshvara, L. M., Peters, J. D., Furlong, M. T., Harrison, M. L., and Geahlen, R. L. (1999). Phosphorylation- and activation-independent association of the tyrosine kinase Syk and the tyrosine kinase substrates Cbl and Vav with tubulin in B-cells. J. Biol. Chem. 274, 1401–1406.
Fifadara, N. H., Beer, F., Ono, S., and Ono, S. J. (2010). Interaction between activated chemokine receptor 1 and FcepsilonRI at membrane rafts promotes communication and F-actin-rich cytoneme extensions between mast cells. Int. Immunol. 22, 113–128.
Firat-Karalar, E. N., and Welch, M. D. (2011). New mechanisms and functions of actin nucleation. Curr. Opin. Cell Biol. 23, 4–13.
Foger, N., Jenckel, A., Orinska, Z., Lee, K. H., Chan, A. C., and Bulfone-Paus, S. (2011). Differential regulation of mast cell degranulation versus cytokine secretion by the actin regulatory proteins Coronin1a and Coronin1b. J. Exp. Med. 208, 1777–1787.
Foster, K. G., and Fingar, D. C. (2010). Mammalian target of rapamycin (mTOR): conducting the cellular signaling symphony. J. Biol. Chem. 285, 14071–14077.
Frigeri, L., and Apgar, J. R. (1999). The role of actin microfilaments in the down-regulation of the degranulation response in RBL-2H3 mast cells. J. Immunol. 162, 2243–2250.
Galan, C., Dionisio, N., Smani, T., Salido, G. M., and Rosado, J. A. (2011). The cytoskeleton plays a modulatory role in the association between STIM1 and the Ca2+ channel subunits Orai1 and TRPC1. Biochem. Pharmacol. 82, 400–410.
Galjart, N. (2010). Plus-end-tracking proteins and their interactions at microtubule ends. Curr. Biol. 20, R528–R537.
Galli, S. J., Tsai, M., and Piliponsky, A. M. (2008). The development of allergic inflammation. Nature 454, 445–454.
Gilfillan, A. M., and Rivera, J. (2009). The tyrosine kinase network regulating mast cell activation. Immunol. Rev. 228, 149–169.
Goley, E. D., and Welch, M. D. (2006). The ARP2/3 complex: an actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 7, 713–726.
Green, K. J., Bohringer, M., Gocken, T., and Jones, J. C. (2005). Intermediate filament associated proteins. Adv. Protein Chem. 70, 143–202.
Grigoriev, I., Gouveia, S. M., van der Vaart, B., Demmers, J., Smyth, J. T., Honnappa, S., Splinter, D., Steinmetz, M. O., Putney, J. W. Jr., Hoogenraad, C. C., and Akhmanova, A. (2008). STIM1 is a MT-plus-end-tracking protein involved in remodeling of the ER. Curr. Biol. 18, 177–182.
Guillemot, J. C., Montcourrier, P., Vivier, E., Davoust, J., and Chavrier, P. (1997). Selective control of membrane ruffling and actin plaque assembly by the Rho GTPases Rac1 and CDC42 in FcεRI-activated rat basophilic leukemia (RBL-2H3) cells. J. Cell Sci. 110, 2215–2225.
Guzman, R. E., Bolanos, P., Delgado, A., Rojas, H., DiPolo, R., Caputo, C., and Jaffe, E. H. (2007). Depolymerisation and rearrangement of actin filaments during exocytosis in rat peritoneal mast cells: involvement of ryanodine-sensitive calcium stores. Pflugers Arch. 454, 131–141.
Hájková, Z., Bugajev, V., Dráberová, E., Vinopal, S., Dráberová, L., Janáček, J., Dráber, Pe., and Dráber, P. (2011). STIM1-directed reorganization of microtubules in activated cells. J. Immunol. 186, 913–923.
Hamawy, M. M., Mergenhagen, S. E., and Siraganian, R. P. (1994). Adhesion molecules as regulators of mast-cell and basophil function. Immunol. Today 15, 62–66.
Hamawy, M. M., Oliver, C., Mergenhagen, S. E., and Siraganian, R. P. (1992). Adherence of rat basophilic leukemia (RBL-2H3) cells to fibronectin-coated surfaces enhances secretion. J. Immunol. 149, 615–621.
Heasman, S. J., and Ridley, A. J. (2008). Mammalian Rho GTPases: new insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 9, 690–701.
Heneberg, P., Dráberová, L., Bambousková, M., Pompach, P., and Draber, P. (2010). Down-regulation of protein-tyrosine phosphatases activates an immune receptor in the absence of its translocation into lipid rafts. J. Biol. Chem. 285, 12787–12802.
Holowka, D., Sheets, E. D., and Baird, B. (2000). Interactions between FcεRI and lipid raft components are regulated by the actin cytoskeleton. J. Cell Sci. 113, 1009–1019.
Honnappa, S., Gouveia, S. M., Weisbrich, A., Damberger, F. F., Bhavesh, N. S., Jawhari, H., Grigoriev, I., van Rijssel, F. J., Buey, R. M., Lawera, A., Jelesarov, I., Winkler, F. K., Wuthrich, K., Akhmanova, A., and Steinmetz, M. O. (2009). An EB1-binding motif acts as a microtubule tip localization signal. Cell 138, 366–376.
Horny, H. P., Reimann, O., and Kaiserling, E. (1988). Immunoreactivity of normal and neoplastic human tissue mast cells. Am. J. Clin. Pathol. 89, 335–340.
Inukai, K., Funaki, M., Nawano, M., Katagiri, H., Ogihara, T., Anai, M., Onishi, Y., Sakoda, H., Ono, H., Fukushima, Y., Kikuchi, M., Oka, Y., and Asano, T. (2000). The N-terminal 34 residues of the 55 kDa regulatory subunits of phosphoinositide 3-kinase interact with tubulin. Biochem. J. 346, 483–489.
Ishizuka, T., Okajima, F., Ishiwara, M., Iizuka, K., Ichimonji, I., Kawata, T., Tsukagoshi, H., Dobashi, K., Nakazawa, T., and Mori, M. (2001). Sensitized mast cells migrate toward the antigen: a response regulated by p38 mitogen-activated protein kinase and Rho-associated coiled-coil-forming protein kinase. J. Immunol. 167, 2298–2304.
Izushi, K., Fujiwara, Y., and Tasaka, K. (1992). Identification of vimentin in rat peritoneal mast cells and its phosphorylation in association with histamine release. Immunopharmacology 23, 153–161.
Janke, C., and Bulinski, J. C. (2011). Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 12, 773–786.
Jordan, M. A., and Kamath, K. (2007). How do microtubule-targeted drugs work? An overview. Curr. Cancer Drug Targets 7, 730–742.
Jouvin, M. H. E., Adamczewski, M., Numerof, R., Letourneur, O., Valle, A., and Kinet, J. P. (1994). Differential control of the tyrosine kinases Lyn and Syk by the 2 signaling chains of the high-affinity immunoglobulin-e receptor. J. Biol. Chem. 269, 5918–5925.
Julian, M., Tollon, Y., Lajoie-Mazenc, I., Moisand, A., Mazarguil, H., Puget, A., and Wright, M. (1993). γ-Tubulin participates in the formation of midbody during cytokinesis in mammalian cells. J. Cell Sci. 105, 145–156.
Kalesnikoff, J., and Galli, S. J. (2008). New developments in mast cell biology. Nat. Immunol. 9, 1215–1223.
Kapeller, R., Toker, A., Cantley, L. C., and Carpenter, C. L. (1995). Phosphoinositide 3-kinase binds constitutively to α/β-tubulin and binds to γ-tubulin in response to insulin. J. Biol. Chem. 270, 25985–25991.
Katagiri, K., Katagiri, T., Kajiyama, K., Yamamoto, T., and Yoshida, T. (1993). Tyrosine-phosphorylation of tubulin during monocytic differentiation of HL-60 cells. J. Immunol. 150, 585–593.
Katsetos, C. D., and Dráber, P. (2012). Tubulins as therapeutic targets in cancer: from bench to bedside. Curr. Pharm. Des. 18, 2778–2792.
Kettner, A., Kumar, L., Anton, I. M., Sasahara, Y., de la, F. M., Pivniouk, V. I., Falet, H., Hartwig, J. H., and Geha, R. S. (2004). WIP regulates signaling via the high affinity receptor for immunoglobulin E in mast cells. J. Exp. Med. 199, 357–368.
Kim, S., and Coulombe, P. A. (2007). Intermediate filament scaffolds fulfill mechanical, organizational, and signaling functions in the cytoplasm. Genes Dev. 21, 1581–1597.
Kodaki, T., Woscholski, R., Hallberg, B., Rodriguez-Viciana, P., Downward, J., and Parker, P. J. (1994). The activation of phosphatidylinositol 3-kinase by Ras. Curr. Biol. 4, 798–806.
Koffer, A., Tatham, P. E., and Gomperts, B. D. (1990). Changes in the state of actin during the exocytotic reaction of permeabilized rat mast cells. J. Cell Biol. 111, 919–927.
Kreis, T., and Vale, R. (1999). Guidebook to the Cytoskeletal and Motor Proteins. Oxford: Oxford University Press.
Kuehn, H. S., Jung, M. Y., Beaven, M. A., Metcalfe, D. D., and Gilfillan, A. M. (2011). Prostaglandin E2 activates and utilizes mTORC2 as a central signaling locus for the regulation of mast cell chemotaxis and mediator release. J. Biol. Chem. 286, 391–402.
Kuehn, H. S., Radinger, M., Brown, J. M., Ali, K., Vanhaesebroeck, B., Beaven, M. A., Metcalfe, D. D., and Gilfillan, A. M. (2010). Btk-dependent Rac activation and actin rearrangement following FcεRI aggregation promotes enhanced chemotactic responses of mast cells. J. Cell Sci. 123, 2576–2585.
Kukharskyy, V., Sulimenko, V., Macurek, L., Sulimenko, T., Dráberová, E., and Dráber, P. (2004). Complexes of γ-tubulin with non-receptor protein tyrosine kinases Src and Fyn in differentiating P19 embryonal carcinoma cells. Exp. Cell Res. 298, 218–228.
Kustermans, G., Piette, J., and Legrand-Poels, S. (2008). Actin-targeting natural compounds as tools to study the role of actin cytoskeleton in signal transduction. Biochem. Pharmacol. 76, 1310–1322.
Lagunoff, D., Martin, T. W., and Read, G. (1983). Agents that release histamine from mast cells. Annu. Rev. Pharmacol. Toxicol. 23, 331–351.
Lam, V., Kalesnikoff, J., Lee, C. W., Hernandez-Hansen, V., Wilson, B. S., Oliver, J. M., and Krystal, G. (2003). IgE alone stimulates mast cell adhesion to fibronectin via pathways similar to those used by IgE+ antigen but distinct from those used by Steel factor. Blood 102, 1405–1413.
Lesourne, R., Fridman, W. H., and Daeron, M. (2005). Dynamic interactions of Fcγ receptor IIB with filamin-bound SHIP1 amplify filamentous actin-dependent negative regulation of Fcε receptor I signaling. J. Immunol. 174, 1365–1373.
Ley, S. C., Marsh, M., Bebbington, C. R., Proudfoot, K., and Jordan, P. (1994). Distinct intracellular-localization of Lck and Fyn protein-tyrosine kinases in human T-lymphocytes. J. Cell Biol. 125, 639–649.
Lin, T. C., Gombos, L., Neuner, A., Sebastian, D., Olsen, J. V., Hrle, A., Benda, C., and Schiebel, E. (2011). Phosphorylation of the yeast γ-tubulin Tub4 regulates microtubule function. PLoS ONE 6, e19700. doi:10.1371/journal.pone.0019700
Linhartová, I., Dráber, P., Dráberová, E., and Viklický, V. (1992). Immunological discrimination of β-tubulin isoforms in developing mouse brain. Posttranslational modification of non-class III β-tubulins. Biochem. J. 288, 919–924.
Liou, J., Fivaz, M., Inoue, T., and Meyer, T. (2007). Live-cell imaging reveals sequential oligomerization and local plasma membrane targeting of stromal interaction molecule 1 after Ca2+ store depletion. Proc. Natl. Acad. Sci. U.S.A. 104, 9301–9306.
Liou, J., Kim, M. L., Heo, W. D., Jones, J. T., Myers, J. W., Ferrell, J. E. Jr., and Meyer, T. (2005). STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 15, 1235–1241.
Liu, Y., Zhu, M., Nishida, K., Hirano, T., and Zhang, W. (2007). An essential role for RasGRP1 in mast cell function and IgE-mediated allergic response. J. Exp. Med. 204, 93–103.
Logue, J. S., Whiting, J. L., Tunquist, B., Sacks, D. B., Langeberg, L. K., Wordeman, L., and Scott, J. D. (2011). AKAP220 protein organizes signaling elements that impact cell migration. J. Biol. Chem. 286, 39269–39281.
Lüders, J., Patel, U. K., and Stearns, T. (2006). GCP-WD is a γ-tubulin targeting factor required for centrosomal and chromatin mediated microtubule nucleation. Nat. Cell Biol. 8, 137–147.
Ludowyke, R. I., Elgundi, Z., Kranenburg, T., Stehn, J. R., Schmitz-Peiffer, C., Hughes, W. E., and Biden, T. J. (2006). Phosphorylation of nonmuscle myosin heavy chain IIA on Ser1917 is mediated by protein kinase C βII and coincides with the onset of stimulated degranulation of RBL-2H3 mast cells. J. Immunol. 177, 1492–1499.
Luduena, R. F., and Banerjee, A. (2008). “The isotypes of tubulin: distribution and functional significance,” in The Role of Microtubules in Cell Biology, Neurobiology and Oncology, ed. T. Fojo (Totowa, NJ: Humana Press), 123–175.
Ma, H.-T., and Beaven, A. (2011). “Regulators of Ca2+ signalling in mast cells: potential targets for treatment of mast cell-related disease,” in Mast Cell Biology: Contemporary and Emerging Topics, eds A. M. Gilfillan and D. D. Metcalfe (Heidelberg: Landes Bioscience and Springer Science+Business Media), 62–90.
Ma, H. T., and Beaven, M. A. (2009). Regulation of Ca2+ signaling with particular focus on mast cells. Crit. Rev. Immunol. 29, 155–186.
Macurek, L., Dráberová, E., Richterová, V., Sulimenko, V., Sulimenko, T., Dráberová, L., Marková, V., and Dráber, P. (2008). Regulation of microtubule nucleation in differentiating embryonal carcinoma cells by complexes of membrane-bound γ-tubulin with Fyn kinase and phosphoinositide 3-kinase. Biochem. J. 416, 421–430.
Malacombe, M., Bader, M. F., and Gasman, S. (2006). Exocytosis in neuroendocrine cells: new tasks for actin. Biochim. Biophys. Acta 1763, 1175–1183.
Marie-Cardine, A., Kirchgessner, H., Eckerskorn, C., Mener, S. C., and Schraven, B. (1995). Human T lymphocyte activation induces tyrosine phosphorylation of α-tubulin and its association with the SH2 domain of the p59fyn protein tyrosine kinase. Eur. J. Immunol. 25, 3290–3297.
Martin-Verdeaux, S., Pombo, I., Iannascoli, B., Roa, M., Varin-Blank, N., Rivera, J., and Blank, U. (2003). Evidence of a role for Munc18-2 and microtubules in mast cell granule exocytosis. J. Cell Sci. 116, 325–334.
Meininger, C. J., Yano, H., Rottapel, R., Bernstein, A., Zsebo, K. M., and Zetter, B. R. (1992). The c-kit receptor ligand functions as a mast cell chemoattractant. Blood 79, 958–963.
Mercer, J. C., Dehaven, W. I., Smyth, J. T., Wedel, B., Boyles, R. R., Bird, G. S., and Putney, J. W. Jr. (2006). Large store-operated calcium selective currents due to co-expression of Orai1 or Orai2 with the intracellular calcium sensor, Stim1. J. Biol. Chem. 281, 24979–24990.
Mitchison, T., and Kirschner, M. (1984). Dynamic instability of microtubule growth. Nature 312, 237–242.
Mullins, F. M., Park, C. Y., Dolmetsch, R. E., and Lewis, R. S. (2009). STIM1 and calmodulin interact with Orai1 to induce Ca2+-dependent inactivation of CRAC channels. Proc. Natl. Acad. Sci. U.S.A. 106, 15495–15500.
Murphy, S. M., Preble, A. M., Patel, U. K., O’Connell, K. L., Dias, D. P., Moritz, M., Agard, D., Stults, J. T., and Stearns, T. (2001). GCP5 and GCP6: two new members of the human γ-tubulin complex. Mol. Biol. Cell 12, 3340–3352.
Nahm, D. H., Tkaczyk, C., Fukuishi, N., Colucci-Guyon, E., Gilfillan, A. M., and Metcalfe, D. D. (2003). Identification of Fyn-binding proteins in MC/9 mast cells using mass spectrometry. Biochem. Biophys. Res. Commun. 310, 202–208.
Narasimhan, V., Holowka, D., and Baird, B. (1990). Microfilaments regulate the rate of exocytosis in rat basophilic leukemia cells. Biochem. Biophys. Res. Commun. 171, 222–229.
Naumanen, P., Lappalainen, P., and Hotulainen, P. (2008). Mechanisms of actin stress fibre assembly. J. Microsc. 231, 446–454.
Navara, C. S., Vassilev, A. O., Tibbles, H. E., Marks, B., and Uckun, F. M. (1999). The spleen tyrosine kinase (Syk) is present at the centrosome in cultured B-cells. Blood 94, 9A.
Neisch, A. L., and Fehon, R. G. (2011). Ezrin, Radixin and Moesin: key regulators of membrane-cortex interactions and signaling. Curr. Opin. Cell Biol. 23, 377–382.
Nielsen, E. H., Braun, K., and Johansen, T. (1989). Reorganization of the subplasmalemmal cytoskeleton in association with exocytosis in rat mast cells. Histol. Histopathol. 4, 473–477.
Nishida, K., Yamasaki, S., Ito, Y., Kabu, K., Hattori, K., Tezuka, T., Nishizumi, H., Kitamura, D., Goitsuka, R., Geha, R. S., Yamamoto, T., Yagi, T., and Hirano, T. (2005). FcεRI-mediated mast cell degranulation requires calcium-independent microtubule-dependent translocation of granules to the plasma membrane. J. Cell Biol. 170, 115–126.
Nobes, C. D., and Hall, A. (1995). Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 81, 53–62.
Nogales, E., and Wang, H. W. (2006). Structural mechanisms underlying nucleotide-dependent self-assembly of tubulin and its relatives. Curr. Opin. Struct. Biol. 16, 221–229.
Nogales, E., Wolf, S. G., and Downing, K. H. (1998). Structure of the αβ tubulin dimer by electron crystallography. Nature 391, 199–203.
Norman, J. C., Price, L. S., Ridley, A. J., Hall, A., and Koffer, A. (1994). Actin filament organization in activated mast cells is regulated by heterotrimeric and small GTP-binding proteins. J. Cell Biol. 126, 1005–1015.
Norman, J. C., Price, L. S., Ridley, A. J., and Koffer, A. (1996). The small GTP-binding proteins, Rac and Rho, regulate cytoskeletal organization and exocytosis in mast cells by parallel pathways. Mol. Biol. Cell 7, 1429–1442.
Nováková, M., Dráberová, E., Schürmann, W., Czihak, G., Viklický, V., and Dráber, P. (1996). γ-Tubulin redistribution in taxol-treated mitotic cells probed by monoclonal antibodies. Cell Motil. Cytoskeleton 33, 38–51.
Oakley, C. E., and Oakley, B. R. (1989). Identification of γ-tubulin, a new member of the tubulin superfamily encoded by mipA gene of Aspergillus nidulans. Nature 338, 662–664.
Oka, T., Hori, M., and Ozaki, H. (2005). Microtubule disruption suppresses allergic response through the inhibition of calcium influx in the mast cell degranulation pathway. J. Immunol. 174, 4584–4589.
Oka, T., Hori, M., Tanaka, A., Matsuda, H., Karaki, H., and Ozaki, H. (2004). IgE alone-induced actin assembly modifies calcium signaling and degranulation in RBL-2H3 mast cells. Am. J. Physiol. Cell Physiol. 286, C256–C263.
Oka, T., Sato, K., Hori, M., Ozaki, H., and Karaki, H. (2002). FcεRI cross-linking-induced actin assembly mediates calcium signalling in RBL-2H3 mast cells. Br. J. Pharmacol. 136, 837–846.
Palazzo, A. F., Cook, T. A., Alberts, A. S., and Gundersen, G. G. (2001). mDia mediates Rho-regulated formation and orientation of stable microtubules. Nat. Cell Biol. 3, 723–729.
Parekh, A. B., and Putney, J. W. Jr. (2005). Store-operated calcium channels. Physiol. Rev. 85, 757–810.
Parravicini, V., Gadina, M., Kovarova, M., Odom, S., Gonzalez-Espinosa, C., Furumoto, Y., Saitoh, S., Samelson, L. E., O’Shea, J. J., and Rivera, J. (2002). Fyn kinase initiates complementary signals required for IgE-dependent mast cell degranulation. Nat. Immunol. 3, 741–748.
Parsons, J. T., Horwitz, A. R., and Schwartz, M. A. (2010). Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 11, 633–643.
Pendleton, A., and Koffer, A. (2001). Effects of latrunculin reveal requirements for the actin cytoskeleton during secretion from mast cells. Cell Motil. Cytoskeleton 48, 37–51.
Perrin, B. J., and Ervasti, J. M. (2010). The actin gene family: function follows isoform. Cytoskeleton 67, 630–634.
Peters, J. D., Furlong, M. T., Asai, D. J., Harrison, M. L., and Geahlen, R. L. (1996). Syk, activated by cross-linking the B-cell antigen receptor, localizes to the cytosol where it interacts with and phosphorylates α-tubulin on tyrosine. J. Biol. Chem. 271, 4755–4762.
Pfeiffer, J. R., Seagrave, J. C., Davis, B. H., Deanin, G. G., and Oliver, J. M. (1985). Membrane and cytoskeletal changes associated with IgE-mediated serotonin release from rat basophilic leukemia cells. J. Cell Biol. 101, 2145–2155.
Pivniouk, V. I., Snapper, S. B., Kettner, A., Alenius, H., Laouini, D., Falet, H., Hartwig, J., Alt, F. W., and Geha, R. S. (2003). Impaired signaling via the high-affinity IgE receptor in Wiskott-Aldrich syndrome protein-deficient mast cells. Int. Immunol. 15, 1431–1440.
Prakriya, M., Feske, S., Gwack, Y., Srikanth, S., Rao, A., and Hogan, P. G. (2006). Orai1 is an essential pore subunit of the CRAC channel. Nature 443, 230–233.
Price, L. S., Langeslag, M., ten Klooster, J. P., Hordijk, P. L., Jalink, K., and Collard, J. G. (2003). Calcium signaling regulates translocation and activation of Rac. J. Biol. Chem. 278, 39413–39421.
Price, L. S., Norman, J. C., Ridley, A. J., and Koffer, A. (1995). The small GTPases Rac and Rho as regulators of secretion in mast cells. Curr. Biol. 5, 68–73.
Psatha, M., Koffer, A., Erent, M., Moss, S. E., and Bolsover, S. (2004). Calmodulin spatial dynamics in RBL-2H3 mast cells. Cell Calcium 36, 51–59.
Psatha, M. I., Razi, M., Koffer, A., Moss, S. E., Sacks, D. B., and Bolsover, S. R. (2007). Targeting of calcium:calmodulin signals to the cytoskeleton by IQGAP1. Cell Calcium 41, 593–605.
Ramesh, N., and Geha, R. (2009). Recent advances in the biology of WASP and WIP. Immunol. Res. 44, 99–111.
Ridley, A. J., and Hall, A. (1992). The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 70, 389–399.
Ridley, A. J., Paterson, H. F., Johnston, C. L., Diekmann, D., and Hall, A. (1992). The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell 70, 401–410.
Rivera, J., Fierro, N. A., Olivera, A., and Suzuki, R. (2008). New insights on mast cell activation via the high affinity receptor for IgE. Adv. Immunol. 98, 85–120.
Robbens, J., Louahed, J., De Pestel, K., Van Colen, I., Ampe, C., Vandekerckhove, J., and Renauld, J. C. (1998). Murine adseverin (D5), a novel member of the gelsolin family, and murine adseverin are induced by interleukin-9 in T-helper lymphocytes. Mol. Cell Biol. 18, 4589–4596.
Roos, J., DiGregorio, P. J., Yeromin, A. V., Ohlsen, K., Lioudyno, M., Zhang, S., Safrina, O., Kozak, J. A., Wagner, S. L., Cahalan, M. D., Velicelebi, G., and Stauderman, K. A. (2005). STIM1, an essential and conserved component of store-operated Ca2+ channel function. J. Cell Biol. 169, 435–445.
Sampieri, A., Zepeda, A., Asanov, A., and Vaca, L. (2009). Visualizing the store-operated channel complex assembly in real time: identification of SERCA2 as a new member. Cell Calcium 45, 439–446.
Schwartz, M. A., Schaller, M. D., and Ginsberg, M. H. (1995). Integrins: emerging paradigms of signal transduction. Annu. Rev. Cell Dev. Biol. 11, 549–599.
Sells, M. A., Pfaff, A., and Chernoff, J. (2000). Temporal and spatial distribution of activated Pak1 in fibroblasts. J. Cell Biol. 151, 1449–1458.
Shimizu, T., Owsianik, G., Freichel, M., Flockerzi, V., Nilius, B., and Vennekens, R. (2009). TRPM4 regulates migration of mast cells in mice. Cell Calcium 45, 226–232.
Simon, J. R., Graff, R. D., and Maness, P. F. (1998). Microtubule dynamics in a cytosolic extract of fetal rat brain. J. Neurocytol. 27, 119–126.
Sivalenka, R. R., and Jessberger, R. (2004). SWAP-70 regulates c-kit-induced mast cell activation, cell-cell adhesion, and migration. Mol. Cell Biol. 24, 10277–10288.
Smith, A. J., Pfeiffer, J. R., Zhang, J., Martinez, A. M., Griffiths, G. M., and Wilson, B. S. (2003). Microtubule-dependent transport of secretory vesicles in RBL-2H3 cells. Traffic 4, 302–312.
Smyth, J. T., Dehaven, W. I., Bird, G. S., and Putney, J. W. Jr. (2007). Role of the microtubule cytoskeleton in the function of the store-operated Ca2+ channel activator STIM1. J. Cell Sci. 120, 3762–3771.
Smyth, J. T., Dehaven, W. I., Jones, B. F., Mercer, J. C., Trebak, M., Vazquez, G., and Putney, J. W. Jr. (2006). Emerging perspectives in store-operated Ca2+ entry: roles of Orai, Stim and TRP. Biochim. Biophys. Acta 1763, 1147–1160.
Spiering, D., and Hodgson, L. (2011). Dynamics of the Rho-family small GTPases in actin regulation and motility. Cell Adh. Migr. 5, 170–180.
Srikanth, S., Jung, H. J., Kim, K. D., Souda, P., Whitelegge, J., and Gwack, Y. (2010). A novel EF-hand protein, CRACR2A, is a cytosolic Ca2+ sensor that stabilizes CRAC channels in T cells. Nat. Cell Biol. 12, 436–446.
Stearns, T., Evans, L., and Kirschner, M. (1991). γ-Tubulin is highly conserved component of the centrosome. Cell 65, 825–836.
Struckhoff, A. P., Vitko, J. R., Rana, M. K., Davis, C. T., Foderingham, K. E., Liu, C. H., Vanhoy-Rhodes, L., Elliot, S., Zhu, Y., Burow, M., and Worthylake, R. A. (2010). Dynamic regulation of ROCK in tumor cells controls CXCR4-driven adhesion events. J. Cell Sci. 123, 401–412.
Stumpff, J., Duncan, T., Homola, E., Campbell, S. D., and Su, T. T. (2004). Drosophila Wee1 kinase regulates Cdk1 and mitotic entry during embryogenesis. Curr. Biol. 14, 2143–2148.
Sulimenko, V., Dráberová, E., Sulimenko, T., Macurek, L., Richterová, V., Dráber, Pe., and Dráber, P. (2006). Regulation of microtubule formation in activated mast cells by complexes of γ-tubulin with Fyn and Syk kinases. J. Immunol. 176, 7243–7253.
Sulimenko, V., Sulimenko, T., Poznanovic, S., Nechiporuk-Zloy, V., Böhm, J. K., Macurek, L., Unger, E., and Dráber, P. (2002). Association of brain γ-tubulins with αβ-tubulin dimers. Biochem. J. 365, 889–895.
Sullivan, R., Burnham, M., Torok, K., and Koffer, A. (2000). Calmodulin regulates the disassembly of cortical F-actin in mast cells but is not required for secretion. Cell Calcium 28, 33–46.
Sullivan, R., Price, L. S., and Koffer, A. (1999). Rho controls cortical F-actin disassembly in addition to, but independently of, secretion in mast cells. J. Biol. Chem. 274, 38140–38146.
Suzuki, R., Liu, X., Olivera, A., Aguiniga, L., Yamashita, Y., Blank, U., Ambudkar, I., and Rivera, J. (2010). Loss of TRPC1-mediated Ca2+ influx contributes to impaired degranulation in Fyn-deficient mouse bone marrow-derived mast cells. J. Leukoc. Biol. 88, 863–875.
Tasaka, K. (1994). Molecular mechanism of histamine release: the role of intermediate filaments and membrane skeletons. J. Physiol. Pharmacol. 45, 479–492.
Tasaka, K., Mio, M., Fujisawa, K., and Aoki, I. (1991). Role of microtubules on Ca2+ release from the endoplasmic-reticulum and associated histamine-release from rat peritoneal mast-cells. Biochem. Pharmacol. 41, 1031–1037.
Teshima, R., Ikebuchi, H., Nakanishi, M., and Sawada, J. (1994). Stimulatory effect of pervanadate on calcium signals and histamine secretion of RBL-2H3 cells. Biochem. J. 302, 867–874.
Thastrup, O., Cullen, P. J., Drobak, B. K., Hanley, M. R., and Dawson, A. P. (1990). Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc. Natl. Acad. Sci. U.S.A. 87, 2466–2470.
Tolarová, H., Dráberová, L., Heneberg, P., and Dráber, P. (2004). Involvement of filamentous actin in setting the threshold for degranulation in mast cells. Eur. J. Immunol. 34, 1627–1636.
Tumová, M., Koffer, A., Šimícek, M., Dráberová, L., and Dráber, P. (2010). The transmembrane adaptor protein NTAL signals to mast cell cytoskeleton via the small GTPase Rho. Eur. J. Immunol. 40, 3235–3245.
Urata, C., and Siraganian, R. P. (1985). Pharmacologic modulation of the IgE or Ca2+ Ionophore A23187 mediated Ca2+ influx, phospholipase activation, and histamine release in rat basophilic leukemia cells. Int. Arch. Allergy Immunol. 78, 92–100.
Vinopal, S., černohorská, M., Sulimenko, V., Sulimenko, T., Vosecká, V., Flemr, M., Dráberová, E., and Dráber, P. (2012). γ-Tubulin 2 nucleates microtubules and is downregulated in mouse early embryogenesis. PLoS ONE 7, e29919. doi:10.1371/journal.pone.0029919
Vogel, J., Drapkin, B., Oomen, J., Beach, D., Bloom, K., and Snyder, M. (2001). Phosphorylation of γ-tubulin regulates microtubule organization in budding yeast. Dev. Cell 1, 621–631.
Wade, R. H., and Hyman, A. A. (1997). Microtubule structure and dynamics. Curr. Opin. Cell Biol. 9, 12–17.
Weller, C. L., Collington, S. J., Hartnell, A., Conroy, D. M., Kaise, T., Barker, J. E., Wilson, M. S., Taylor, G. W., Jose, P. J., and Williams, T. J. (2007). Chemotactic action of prostaglandin E2 on mouse mast cells acting via the PGE2 receptor 3. Proc. Natl. Acad. Sci. U.S.A. 104, 11712–11717.
Westermann, S., and Weber, K. (2003). Post-translational modifications regulate microtubule function. Nat. Rev. Mol. Cell Biol. 4, 938–945.
Wilson, B. S., Pfeiffer, J. R., and Oliver, J. M. (2000). Observing FcεRI signaling from the inside of the mast cell membrane. J. Cell Biol. 149, 1131–1142.
Wise, D. O., Krahe, R., and Oakley, B. R. (2000). The γ-tubulin gene family in humans. Genomics 67, 164–170.
Wolff, A., Denoulet, P., and Jeantet, C. (1982). High level of tubulin microheterogeneity in the mouse brain. Neurosci. Lett. 31, 323–328.
Wu, M. M., Buchanan, J., Luik, R. M., and Lewis, R. S. (2006). Ca2+ store depletion causes STIM1 to accumulate in ER regions closely associated with the plasma membrane. J. Cell Biol. 174, 803–813.
Yanase, Y., Hide, I., Mihara, S., Shirai, Y., Saito, N., Nakata, Y., Hide, M., and Sakai, N. (2011). A critical role of conventional protein kinase C in morphological changes of rodent mast cells. Immunol. Cell Biol. 89, 149–159.
Keywords: actins, intermediate filaments, mast cell activation, microfilaments, microtubules, signal transduction, tubulins, vimentin
Citation: Dráber P, Sulimenko V and Dráberová E (2012) Cytoskeleton in mast cell signaling. Front. Immun. 3:130. doi: 10.3389/fimmu.2012.00130
Received: 10 March 2012; Accepted: 05 May 2012;
Published online: 25 May 2012.
Edited by:
Ulrich Blank, Inserm U699, Université Paris-Diderot Paris 7, FranceReviewed by:
Ulrich Blank, Inserm U699, Université Paris-Diderot Paris 7, FranceMin Wu, University of North Dakota, USA
Copyright: © 2012 Dráber, Sulimenko and Dráberová. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Pavel Dráber, Laboratory of Biology of Cytoskeleton, Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, Vídeňská 1083, 142 20 Prague 4, Czech Republic. e-mail:cGF2ZWxkcmFAaW1nLmNhcy5jeg==