
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 05 October 2011
Sec. Molecular Innate Immunity
volume 2 - 2011 | https://doi.org/10.3389/fimmu.2011.00044
 Aldacilene Souza Silva1
Aldacilene Souza Silva1 Mónica Marcela Castiblanco Valencia1 Aurora Marques Cianciarullo2 Sílvio Arruda Vasconcellos3
Mónica Marcela Castiblanco Valencia1 Aurora Marques Cianciarullo2 Sílvio Arruda Vasconcellos3 Angela Silva Barbosa4
Angela Silva Barbosa4 Lourdes Isaac1*
Lourdes Isaac1*Leptospirosis is a zoonosis caused by pathogenic bacteria from the genus Leptospira. The disease represents a serious public health problem in underdeveloped tropical countries. Leptospires infect hosts through small abrasions in the skin or mucous membranes and they rapidly disseminate to target organs. The capacity of some pathogenic leptospiral strains to acquire the negative complement regulators factor H (FH) and C4b binding protein correlates with their ability to survive in human serum. In this study we assessed the functional consequences of the age macular degeneration-associated polymorphism FH His402 or FH Tyr402 on FH–Leptospira interactions. In binding assays using sub-saturating amounts of FH, the FH Tyr402 variant interacted with all the strains tested more strongly than the FH His402 variant. At higher concentrations, differences tended to disappear. We then compared cofactor activities displayed by FH His402 and FH Tyr402 bound to the surface of L. interrogans. Both variants exhibit similar activity as cofactors for Factor I-mediated cleavage of C3b, thus indicating that they do not differ in their capacity to regulate the complement cascade.
Leptospirosis is a serious public health problem in tropical developing countries. The disease is transmitted by spirochetes of the genus Leptospira, directly or indirectly from animals to humans in regions which lack adequate sewage systems. Pathogenic Leptospira enter the body, reach the bloodstream and can potentially invade potentially all tissues and organs (Faine et al., 1999). Clinical manifestations are quite variable and can range from an asymptomatic, subclinical infection to a fatal hepatorenal syndrome (Weil’s disease), severe pulmonary form, including pulmonary hemorrhage and acute respiratory distress syndrome, fever and jaundice (Faine et al., 1999; Levett, 2001; Bharti et al., 2003), and uveitis (Pappachan et al., 2007).
Prompted by the necessity of developing an efficient subunit vaccine against human leptospirosis, considerable research is being conducted to understand the mechanisms and putative virulence factors that enable pathogenic Leptospira to disseminate throughout the host, culminating in the colonization of multiple organs. Of particular interest are mechanisms that allow Leptospira to evade innate immune system, notably clearance mediated by the alternative pathway (AP) of complement. Leptospira strains differ in resistance to complement-mediated lysis, and the ability to survive in human serum correlates with the capacity of some pathogenic strains to acquire the negative complement regulators factor H (FH) and C4b binding protein (C4BP) on their surfaces (Meri et al., 2005; Barbosa et al., 2009).
Complement FH, a 155-kDa plasma glycoprotein composed of 20 globular domains (termed short consensus repeats, SCRs), is the major soluble complement regulator that controls AP activation and the amplification reaction at the C3 level. It inhibits the complement AP by preventing binding of Factor B to C3b, accelerating decay of the C3-convertase C3bBb and acting as a cofactor for the cleavage of C3b by Factor I (Weiler et al., 1976; Whaley and Ruddy, 1976; Pangburn et al., 1977). FH, encoded by the HF1 gene, is as a single chain with 1231 amino acid residues. The regulatory function on complement amplification is conferred by the first four domains (Gordon et al., 1995; Kühn and Zipfel, 1996), while the remaining 16 SCRs present several binding sites for different targets, including: self surfaces, thereby preventing complement activation on host cells (Sharma and Pangburn, 1996); C3 fragments (Zipfel et al., 1999); C-Reactive Protein (Mold et al., 1999; Ormsby et al., 2006); fibromodulin (Sjöberg et al., 2005); adrenomedullin (Pio et al., 2001); cell receptors (Jarva et al., 1999; Zipfel et al., 2002); M protein and heparin (Sharma and Pangburn, 1997; Giannakis et al., 2003).
The polymorphism (Tyr402His) located in SCR7 of both FH and factor H-like protein 1 (FHL-1) has been associated with age macular degeneration (AMD) by different groups (Edwards et al., 2005; Hageman et al., 2005; Haines et al., 2005; Klein et al., 2005; Teixeira et al., 2010). The frequency of the His402 allotype of FH in the population is quite relevant. Approximately 35% of European descendants carry this allele (Clark et al., 2010). In Brazilian subjects (a highly ethnically mixed population) the frequency of this allele is 13% (Teixeira et al., 2010). Additionally, this same FH polymorphism was associated with other chronic diseases, such as Alzheimer disease (Zetterberg et al., 2008).
The hypothesis that this FH polymorphism could affect FH binding properties was approached by several groups (Laine et al., 2007; Skerka et al., 2007; Yu et al., 2007; Haapasalo et al., 2008) in studies that evaluated the effect of the Tyr402His substitution on binding of FH to heparin, C-reactive protein, and bacterial surface proteins. Using SCR 5–7 domains from FH His402 variant, Laine et al. (2007) observed a weaker binding to C-reactive protein when compared to SCR 5–7 from FH Tyr402. They obtained similar results when purified human FH variants were employed. In addition, no differences in the binding to heparin were observed (Laine et al., 2007). On the other hand, the binding of the Tyr402 and His402 allotype to C-reactive protein did not differ in a study reported by Yu et al. (2007). However, this group observed that the Tyr402 allotype binds much better to M6 protein from Streptococcus pyogenes. A study by Haapasalo et al. (2008) demonstrated that the FH Tyr402 and FH His402 allotypes display different binding affinities for several group A Streptococcus (GAS) strains: binding of the FH His402 allotype to GAS is significantly weaker compared to that exhibited by the FH Tyr402 allotype. As a consequence, GAS strains are more susceptible to phagocytosis in the presence of the His402 allele due to increased C3b deposition on the bacterial surface (Haapasalo et al., 2008).
Given the small number of investigations assessing the functional consequences of the age-related macular degeneration-associated polymorphism Tyr402His on FH–pathogen interactions and the fact that leptospires may cause eye diseases such as uveitis, the main purpose of the present study was to compare binding of Tyr402 and His402 FH variants to pathogenic Leptospira strains. We also evaluated cofactor activity displayed by both variants bound to the leptospiral surface in order to assess if they differ in their capacity to regulate the complement cascade.
The two allotypes were isolated from 100 ml EDTA plasma collected from homozygous healthy donors after previous genotyping, according to Teixeira et al. (2010). Briefly, the material was treated with 10 mM CaCl2 and 0.6 mM phenylmethylsulfonyl fluoride (final concentrations) in order to block the protease cascade, followed by dialysis in 8 mM EDTA and 10 mM benzamidine, pH 5.8, for 18 h at 4°C. The precipitate was washed twice in the same solution and solubilized in 10 mM Tris–HCl, 150 mM NaCl, 100 mM dextrose, 100 mM glycine, and 0.01% sodium azide, pH 8.5. The material was then separated by gel filtration chromatography using a Sephacryl S-200 column (GE Healthcare Life Sciences). The fractions containing FH were pooled and dialyzed against 40 mM NaH2PO4 pH 8.1 overnight and loaded in Sepharose 4B-CNBr activated resin (GE Healthcare Life Sciences) containing polyclonal anti-human FH antibodies (Calbiochem). FH was eluted with 100 mM glycine, pH 2.7. The fractions containing FH were pooled, concentrated, and re-purified in the same affinity column. The purity of the proteins was checked by SDS-PAGE and Western blot. Total protein concentration was estimated by the Bradford assay (Bio-Rad, Richmond, CA, USA). When indicated, both variants of FH were purified exclusively by affinity column.
Leptospira interrogans serovar Pomona strain Fromm, L. kirshneri serovar Grippotyphosa strain Moskva V and L. borgpetersenii serovar Javanica strain Veldrat Bataviae 46 were used in the assays. All strains were obtained from the Laboratório de Zoonoses, Faculdade de Medicina Veterinária e Zootecnia, University of São Paulo, Brazil. Moskva V and Veldrat Bataviae 46 strains were culture-attenuated by successive passages in artificial medium. Virulence of the strain Fromm was maintained by iterative passages in hamsters. Leptospires were cultured at 29°C under aerobic conditions in liquid EMJH medium (Difco® – USA) with 10% rabbit serum, enriched with 0.015% (w/v) L-asparagine, 0.001% (w/v) sodium pyruvate, 0.001% (w/v) calcium chloride, 0.001% (w/v) magnesium chloride, 0.03% (w/v) peptone, and 0.02% (w/v) meat extract.
The following protocol is modified after Barbosa et al., 2010. L. interrogans serovar Pomona strain Fromm cells were washed twice with PBS and fixed with 2% paraformaldehyde in PBS for 60 min at 22°C. After two washes with PBS, bacteria were incubated with 5 μg/ml of purified FH (His402) or FH (Tyr402) for 60 min at 22°C with gentle agitation. After two washes with PBS to remove unbound FH, bacteria were applied to electron microscopy grids and then incubated for 60 min with a polyclonal goat anti-human FH (Quidel) diluted 1:500 in PBS/1.5% BSA/0.05% Tween 20. After four washes with PBS/1% BSA, they were incubated for 60 min with rabbit anti-goat antibody–colloidal gold (10-nm particles; Sigma-Aldrich, Co., St Louis, USA) and after successive washes in PBS/1% BSA, 0.85% NaCl, and bi-distilled water, bacteria were stained with 2% uranyl acetate. Grids were examined with an electron microscope (Zeiss EM 109; Carl Zeiss, Inc., Oberkochen, Germany) at an accelerating voltage of 80 kV.
Leptospires grown to mid-log phase were harvested by centrifugation at 5400 g for 30 min and gently washed in PBS (pH 7.4) twice. Bacteria were incubated with 0–20 μg/ml of purified FH His402 or FH Tyr402 for 60 min at room temperature. After centrifugation at 10000 g for 10 min at 4°C, the pellets were washed three times with a buffer containing 1.0 mM MgCl2, 0.6 mM CaCl2, and 1% glucose, and then suspended in 0.1 M NaHCO3, pH 9.6. Bacteria were used for coating ELISA plate wells overnight at 4°C. The wells were washed twice with PBS/0.05% Tween 20 (PBST) and then blocked with 200 μl of 0.5% BSA for 2 h at 37°C. Bound FH was detected by adding 100 μl of a 1:10000 dilution of goat anti-human FH (Quidel) in PBS. Incubation proceeded for 60 min at 37°C and after three washes with PBST, 100 μl of a 1:10000 dilution of horseradish peroxidase-conjugated anti-rabbit IgG in PBS was added to each well and incubated for 60 min at 37°C. The wells were washed three times and o-phenylenediamine (0.04%) in citrate phosphate buffer (pH 5.0) plus 0.01% H2O2. The reaction was allowed to proceed for 10 min and was then interrupted by the addition of 50 μl of 8 M H2SO4. The absorbance at 492 nm was determined in a micro plate reader (Labsystems Uniscience, Multiskan EX).
Freshly harvested leptospires (1 × 109) were washed twice with PBS (pH 7.4), and incubated with 5 μg/ml of purified FH His402 or FH Tyr402 for 60 min at 22°C with gentle agitation. After five washes with PBS (the last wash fraction was collected), proteins bound to the surface of bacteria were eluted with 100 μl of 0.1 M glycine–HCl pH 2.0 and supernatants were collected after centrifugation. One fifth (20 μl) of the wash and eluate fractions was subjected to 10% SDS-PAGE under non-reducing conditions and transferred to nitrocellulose membranes. As a control, purified FH (Calbiochem) was also loaded in the same gel. Non-specific binding sites were blocked using 10% (w/v) dried milk in PBS–Tween (0.05%; pH 7.4) overnight at 4°C. Subsequently, membranes were rinsed three times in PBS–Tween (0.05%) and were incubated for 60 min at room temperature with a polyclonal goat anti-human FH (Quidel) at a 1:10000 dilution. Following three washes with PBS–Tween (0.05%), membranes were incubated with a secondary peroxidase-conjugated anti-rabbit IgG Ab (Sigma) for 60 min at room temperature at a 1:10000 dilution. The positive signals were detected by enhanced chemiluminescence (Super Signal, West Pico, Pierce).
The cofactor activity of surface-attached FH was assayed by measuring Factor I-mediated cleavage of C3b (Grosskinsky et al., 2009). One milliliter of freshly harvested leptospires (1 × 109) were washed with a binding buffer (100 mM NaCl, 50 mM Tris–HCl pH 7.4) and incubated with purified FH His402, FH Tyr402 (0.5–10 μg/ml), or 10% NHS–EDTA (from blood samples genotyped homozygous for the FH His402 allele or the FH Tyr402 allele), or binding buffer for 60 min at room temperature with gentle agitation. Bacteria were washed three times with washing buffer (100 mM NaCl, 50 mM Tris–HCl, 0.05% Tween 20, pH 7.4), and incubated with purified C3b (Calbiochem; 250 ng/assay) and Factor I (Calbiochem; 150 ng/reaction) for 60 min at 37°C. The samples were centrifuged and the supernatants were subjected to 10% SDS-PAGE under reducing conditions and transferred to nitrocellulose membranes. Blocking treatment and incubations with specific antibodies were performed as described above. Cleavage products of C3b were detected using polyclonal goat anti-human C3 (Calbiochem) at a 1:10000 dilution.
After incubation of L. interrogans Pomona strain Fromm with purified human FH His402 or FH Tyr402, immunoelectron microscopy was performed with anti-FH in order to evaluate binding of both FH variants to this pathogenic strain. Colloidal gold particles were detected on the leptospiral surface in both cases. Representative electron micrographs are shown in Figure 1. Apparently, no binding differences were observed at microscopical level thus suggesting that both variants may interact equally with L. interrogans. Organisms incubated in PBS did not present surface colloidal gold binding (data not shown).
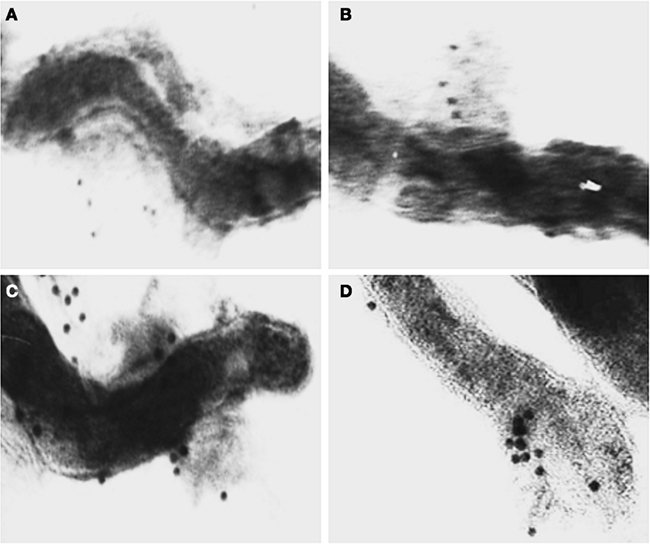
Figure 1. Immunoelectron microscopy analysis. L. interrogans serovar Pomona strain Fromm was incubated with purified human FH His402(A,B) or FH Tyr402 (C,D). After extensive washing, leptospires were incubated with goat anti-human FH polyclonal antibody followed by incubation with anti-goat IgG–colloidal gold conjugated before analysis using an electron microscope (Zeiss EM109). Both variants of FH were purified by gel filtration and affinity chromatography.
To assess the interaction of human complement FH variants with Leptospira on a quantitative basis, we first incubated L. interrogans serovar Pomona strain Fromm with increasing amounts of purified FH His420 or FH Tyr420 (over a range of 0.1–2 μg/ml) and then immobilized the bacteria on microtiter wells. Bound FH was detected with a specific antibody. FH Tyr402 binding to strain Fromm was significantly stronger when compared to FH His402 binding within the range of 0.1–2 μg/ml (Figure 2). Since FH is found in the serum at high levels, we decided to extend this range up to 20 μg/ml using three pathogenic leptospiral strains (Figure 3). At all FH concentrations tested, FH Tyr402 binding to L. interrogans serovar Pomona strain Fromm was significantly stronger when compared to FH His402 binding to this virulent strain (P < 0.05; Figure 3). L. borgpetersenii serovar Javanica strain Veldrat Bataviae 46 and L. kirshneri serovar Grippotyphosa strain Moskva V also bound FH Tyr402 with an apparently higher affinity at lower FH concentrations (Figure 3).
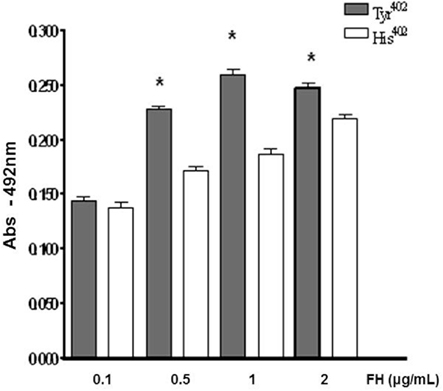
Figure 2. Dose-dependent binding of FH variants to L. interrogans serovar Pomona strain Fromm. L. interrogans were incubated with increasing amounts of purified FH His420 or FH Tyr420 (0.1–2 μg/ml) and then immobilized on microtiter wells. The binding was assessed using a polyclonal anti-human FH. Each point represents the mean absorbance value at 492 nm ± the SD of three independent experiments. *P ≤ 0.05 (Student’s two-tailed t-test). Both variants of FH were purified by gel filtration and affinity chromatography.
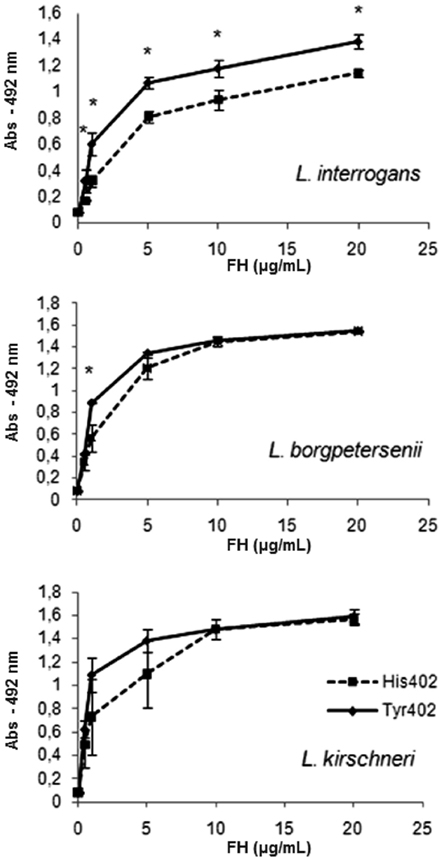
Figure 3. Dose-dependent binding of FH variants to Leptospira. L. borgpetersenii, L. kirshneri, and L. interrogans were incubated with increasing amounts of purified FH His420 or FH Tyr420 (0–20 μg/ml) and then immobilized on microtiter wells. The binding was assessed using a polyclonal anti-human FH. Each point represents the mean absorbance value at 492 nm ± the SD of three independent experiments. *P ≤ 0.05 (Student’s two-tailed t-test). Both variants of FH were purified only by affinity chromatography.
Acquisition of FH variants by pathogenic leptospires was also evaluated by Western blot. To this end, bacteria were incubated with the same amount of FH His402 or FH Tyr402 and after extensive washes the eluate fractions were separated by SDS-PAGE and subjected to Western blot with anti-FH antibody. Interestingly, FH His402 released by all three strains was more abundant when compared to FH Tyr402 probably reflecting a higher affinity of the Tyr402 variant for the Leptospira surface (Figures 4A,B). In addition to binding FH, the Leptospira strains also bound members of the FH-family, most probably FH-related protein 1 (FHR-1α and FHR-1β; Figure 4A) as has been previously reported (Meri et al., 2005).
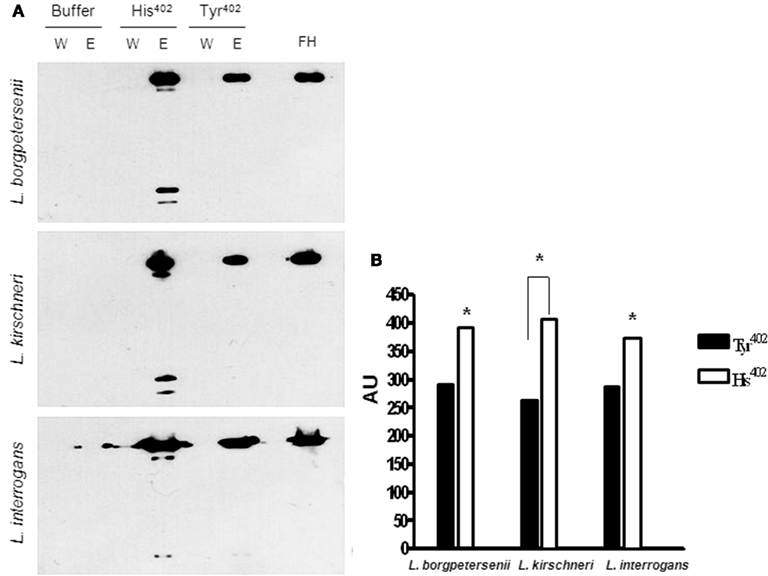
Figure 4. Acquisition of FH by L. borgpetersenii, L. kirshneri, and L. interrogans. (A) Leptospires were incubated with purified FH His402, FH Tyr402, or with buffer only. After extensive washing, surface–bound proteins were eluted and subjected to non-reducing SDS-PAGE and analyzed by Western blot using a polyclonal anti-human FH. (W) aliquots of the last wash; (E) aliquots of the eluted fractions; purified FH (100 ng) was included as a positive control. (B) Quantification of the FH bands in part (A) as described in Section “Materials and Methods.” Data are presented as arbitrary units (AU). Both variants of FH were purified only by affinity chromatography.
We next compared the regulatory role of FH His402 and FH Tyr402 attached to the surface of Leptospira, by analyzing degradation of C3b mediated by Factor I. The serum-resistant L. interrogans serovar Pomona strain Fromm was incubated with purified FH His402 and FH Tyr402 (0.5–10 μg/ml), and also with 10% NHS (His402) or 10% NHS (FH Tyr402). After washing to remove unbound FH, C3b, and Factor I were added. The cleavage fragments of C3b in the supernatant were subjected to Western blot with anti-C3 polyclonal antibody. The presence of a ∼46-kDa band indicates that acquired FH was able to promote Factor I-mediated cleavage of C3b (Figure 5). In the absence of FH, no degradation fragments were detected (Figure 5). Despite binding to Leptospira with different affinities, both variants exhibited similar cofactor activities against C3b. This indicates that they are equally efficient in regulating the complement cascade.
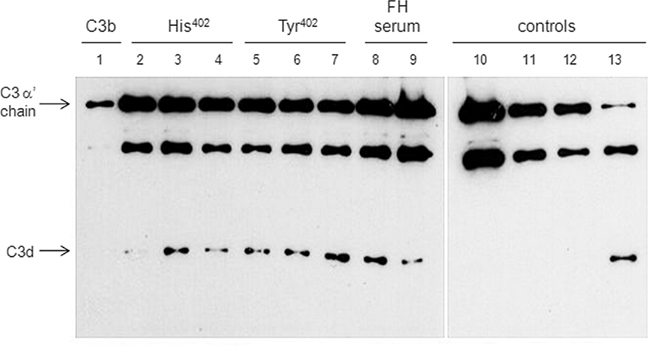
Figure 5. Cofactor activity of FH variants bound to L. interrogans serovar Pomona strain Fromm. C3b (100 ng) and Factor I (250 ng) were added to bacteria that had been pre-incubated with FH His420, FH Tyr420, or with human serum. Reaction mixtures were separated by SDS-PAGE and the cleavage fragments of C3b in the supernatant were subjected to Western blot using a polyclonal anti-human C3. Lanes 1–9: reactions with bacteria. C3b alone (1) or C3b mixed with 0.5 μg of FH (2 and 5); 2.5 μg of FH (3 and 6); 10 μg of FH (4 and 7); 100 μl of serum from FH His420 subject (8); 100 μl of serum from FH Tyr420 subject (9). Lanes 10–13: reactions without Leptospira. Only C3b (10); C3b + FH (11); C3b + FI (12); C3b + FH + Factor I (13). Both variants of FH were purified only by affinity chromatography.
The AP of the complement system plays a pivotal role in the elimination of invading microorganisms. In order to survive in the host, pathogenic Leptospira have evolved mechanisms to avoid or, at least, minimize lytic complement attack. Acquisition of the host fluid-phase complement regulators FH and C4BP is one strategy adopted by these spirochetes to inhibit complement activation (Meri et al., 2005; Barbosa et al., 2009). Binding of FH to Leptospira spp. was first assessed by Meri et al. (2005). Serum-resistant and serum-intermediate strains were able to acquire FH and also the FH-family protein member FH-related protein 1 (FHR-1). Surface-bound FH remained functionally active as a cofactor for Factor I in the cleavage of C3b (Meri et al., 2005). Leptospira’s ability to bind either purified human FH or normal equine serum FH has been examined by indirect immunofluorescence analysis: L. interrogans bound this complement regulator while the non-pathogenic L. biflexa did not (Verma et al., 2006). Our immunoelectron microscopy data presented in the present study provide ultra-structural evidence consistent with these previous findings and demonstrate that FH His402 and FH Tyr402 variants interact with pathogenic Leptospira.
Recent results by our group indicate that Leptospira interact with FH through two different binding sites, one located within SCRs 5–7 and other located at the C-terminal SCRs 18–20 (unpublished data). Considering that the His402 or Tyr402 polymorphism of FH is located in SCR7, we decided to evaluate whether these FH variants would display different binding affinities for this pathogen. According to the solid-phase binding assays, the FH Tyr402 variant bound to the virulent strain Pomona more strongly than the FH His402 did at all concentrations tested. Similar results were observed for the two other pathogenic strains when they were incubated with sub-saturating concentrations of FH. At higher concentrations, no difference in binding was detected. We speculate that intrinsic properties of each strain might affect binding to these FH variants. Since different binding affinities for FH His402 and FH Tyr402 may have functional consequences for the pathogen with regard to susceptibility to complement-mediated killing, we then compared cofactor activities of FH His402 and FH Tyr402 bound to the surface of L. interrogans. Both variants were functionally similar as cofactors for Factor I-mediated cleavage of C3b. These results indicate that the two allotypes do not differ in their capacity to regulate the complement cascade.
In a previous study, Yu et al. (2007) analyzed the interactions of both FH variants with M6 protein from S. pyogenes, pneumococcal surface protein C (PspC) from S. pneumoniae, and complement regulator-acquiring surface protein 1 from Borrelia burgdorferi (BbCRASP-1). While PspC and BbCRASP-1 bound equally well to FH His402 and FH Tyr402, M6 protein interacted with the FH Tyr402 variant with greater affinity. Furthermore, reduced binding of the FH His402 variant of both FH and FHL-1 to GAS M6 protein has been observed by Ormsby et al. (2008). A comprehensive study by Haapasalo et al. (2008) involving almost 40 GAS strains strengthened these findings, showing that the majority of strains bound more weakly FH SCRs 5–7 allotype His402. Moreover, opsonophagocytosis was more pronounced when bacteria were incubated in blood from individuals homozygous for the FH (His402) allele (Haapasalo et al., 2008).
Factor H allotype His402 has been recognized as a leading risk factor for the development of age-related macular degeneration (Edwards et al., 2005; Haines et al., 2005; Klein et al., 2005). Nevertheless, a number of studies report a high prevalence of the homozygous genotype CC (that codes for His402) in different ethnic populations (Edwards et al., 2005; Haines et al., 2005; Klein et al., 2005). A study analyzing populations from different continents found that the frequency of FH allele (His402) is between 0.15 and 0.25 (Haapasalo et al., 2008) and a recent study reported a genotype frequency of 0.13 for CC (His402) in Brazilian individuals (Teixeira et al., 2010). This allele may confer a selective advantage by, for instance, limiting evasion from the immune system by pathogenic microorganisms or increasing susceptibility of the pathogen to opsonophagocytosis (Yu et al., 2007; Haapasalo et al., 2008; Ormsby et al., 2008).
Modeling studies of SCRs 6–7 of FH/FHL-1 predict that the Tyr402His substitution will not cause a significant structural change in the molecule, but it does alter the electrostatic surface charges (Ormsby et al., 2008). This may explain the reduced binding of heparin to FHL-1 (Ormsby et al., 2008), C-reactive protein, and GAS M6 protein to both FH and FHL-1 (Laine et al., 2007; Yu et al., 2007; Haapasalo et al., 2008), and Leptospira to FH (this study). We do not believe that the lower binding affinity of the FH His402 variant for Leptospira, in particular, could have an important functional impact on bacterial survival in the host tissues in which FH is relatively abundant (i.e., plasma). First, because under physiological conditions, high FH levels are found in the serum and, according to our data, differences in binding between the two FH variants tend to disappear at higher FH concentrations. Secondly, both variants are functionally active and display similar cofactor activities. However, these binding differences may be relevant at sites such as eyes where local FH synthesis is low.
In conclusion, in this in vitro study we assessed the functional consequences of the AMD-associated polymorphism FH His402 or FH Tyr402 on FH–Leptospira interactions. While our data suggest that these variants have different binding affinities to these bacteria, both allotypes regulate the AP in a similar pattern; i.e., both act as cofactor of Factor I at similar efficiency. However, we cannot rule out the possibility that other FH polymorphisms may affect the interactions between this complement regulator and Leptospira. In this work we did not address this possibility. In addition, local tissue environment may directly influence the regulatory role of FH in vivo in the eye. For example, the variant FH His402 binds much less intensively to the Bruch’s membrane in the macula region of retina than the Tyr402 variant; which may be related to the etiopathogenesis of AMD (Clark et al., 2010). In equines, L. interrogans is responsible for causing uveitis, a major cause of blindness in these animals (Verma et al., 2005). A considerable fraction (18.4%) of infected patients from South India developed uveitis after acute leptospirosis (Pappachan et al., 2007). These considerations suggest that the role of FH polymorphism in patients with leptospiral uveitis deserves further investigations.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Prof. Shaker Chuck Farah (Instituto de Química, Universidade de São Paulo, Brazil) for the critical review of the manuscript. This work has benefited from grants provided by FAPESP (09/05979-0; 09/1778-0; 10/50043-0).
Barbosa, A. S., Abreu, P. A., Vasconcellos, A. S., Morais, Z. M., Gonçales, A. P., Silva, A. S., Daha, M. R., and Isaac, L. (2009). Immune evasion of Leptospira species by acquisition of human complement regulator C4BP. Infect. Immun. 77, 1137–1143.
Barbosa, A. S., Monaris, D., Silva, L. B., Morais, Z. M., Vasconcellos, S. A., Cianciarullo, A. M., Isaac, L., and Abreu, P. A. (2010). Functional characterization of LcpA, a surface-exposed protein of Leptospira spp. That binds the human complement regulator C4BP. Infect. Immun. 78, 3207–3216.
Bharti, A. R., Nally, J. E., Ricaldi, J. N., Matthias, M. A., Diaz, M. M., Lovett, M. A., Levett, P. N., Gilman, R. H., Willig, M. R., Gotuzzo, E., and Vinetz, J. M. (2003). Peru-united states leptospirosis consortium. Leptospirosis: a zoonotic disease of global importance. Lancet Infect. Dis. 3, 757–771.
Clark, S. J., Perveen, R., Hakobyan, S., Morgan, B. P., Sim, R. B., Bishop, P. N., and Day, A. J. (2010). Impaired binding of the age-related macular degeneration-associated complement factor H 402H allotype to Bruch’s membrane in human retina. J. Biol. Chem. 285, 30192–30202.
Edwards, A. O., Ritter, R. 3rd, Abel, K. J., Manning, A., Panhuysen, C., and Farrer, L. A. (2005). Complement factor H polymorphism and age-related macular degeneration. Science 308, 421–424.
Faine, S., Adler, B., Bolin, C., and Perolat, P. (1999). Leptospira and Leptospirosis, 2nd Edn. Armadale: MediSci.
Giannakis, E., Jokiranta, T. S., Male, D. A., Ranganathan, S., Ormsby, R. J., Fischetti, V. A., Mold, C., and Gordon, D. L. (2003). A common site within factor H SCR 7 responsible for binding heparin, C-reactive protein and streptococcal M protein. Eur. J. Immunol. 33, 962–969.
Gordon, D. L., Kaufman, R. M., Blackmore, T. K., Kwong, J., and Lublin, D. M. (1995). Identification of complement regulatory domains in human factor H. J. Immunol. 155, 348–356.
Grosskinsky, S., Schott, M., Brenner, C., Cutler, S. J., Kraiczy, P., Zipfel, P. F., Simon, M. M., and Wallich, R. (2009). Borrelia recurrentis employs a novel multifunctional surface protein with anti-complement, anti-opsonic and invasive potential to escape innate immunity. PLoS ONE 4, e4858. doi: 10.1371/journal.pone.0004858
Haapasalo, K., Jarva, H., Siljander, T., Tewodros, W., Vuopio-Varkila, J., and Jokiranta, T. S. (2008). Complement factor H allotype 402H is associated with increased C3b opsonization and phagocytosis of Streptococcus pyogenes. Mol. Microbiol. 70, 583–594.
Hageman, G. S., Anderson, D. H., Johnson, L. V., Hancox, L. S., Taiber, A. J., Hardisty, L. I., Hageman, J. L., Stockman, H. A., Borchardt, J. D., Gehrs, K. M., Smith, R. J., Silvestri, G., Russell, S. R., Klaver, C. C., Barbazetto, I., Chang, S., Yannuzzi, L. A., Barile, G. R., Merriam, J. C., Smith, R. T., Olsh, A. K., Bergeron, J., Zernant, J., Merriam, J. E., Gold, B., Dean, M., and Allikmets, R. (2005). A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. U.S.A. 102, 7227–7232.
Haines, J. L., Hauser, M. A., Schmidt, S., Scott, W. K., Olson, L. M., Gallins, P., Spencer, K. L., Kwan, S. Y., Noureddine, M., Gilbert, J. R., Schnetz-Boutaud, N., Agarwal, A., Postel, E. A., and Pericak-Vance, M. A. (2005). Complement factor H variant increases the risk of age-related macular degeneration. Science 308, 419–421.
Jarva, H., Jokiranta, T. S., Hellwage, J., Zipfel, P. F., and Meri, S. (1999). Regulation of complement activation by C-reactive protein: targeting the complement inhibitory activity of factor H by an interaction with short consensus repeat domains 7 and 8-11. J. Immunol. 163, 3957–3962.
Klein, R. J., Zeiss, C., Chew, E. Y., Tsai, J. Y., Sackler, R. S., Haynes, C., Henning, A. K., SanGiovanni, J. P., Mane, S. M., Mayne, S. T., Bracken, M. B., Ferris, F. L., Ott, J., Barnstable, C., and Hoh, J. (2005). Complement factor H polymorphism in age-related macular degeneration. Science 308, 385–389.
Kühn, S., and Zipfel, P. F. (1996). Mapping of the domains required for decay acceleration activity of the human factor H-like protein 1 and factor H. Eur. J. Immunol. 26, 2383–2387.
Laine, M., Jarva, H., Seitsonen, S., Haapasalo, K., Lehtinen, M. J., Lindeman, N., Anderson, D. H., Johnson, P. T., Järvelä, I., Jokiranta, T. S., Hageman, G. S., Immonen, I., and Meri, S. (2007). Y402H polymorphism of complement factor H affects binding affinity to C-reactive protein. J. Immunol. 178, 3831–3836.
Meri, T., Murgia, R., Stefanel, P., Meri, S., and Cinco, M. (2005). Regulation of complement activation at the C3-level by serum resistant leptospires. Microb. Pathog. 39, 139–147.
Mold, C., Gewurz, H., and Du Clos, T. W. (1999). Regulation of complement activation by C-reactive protein. Immunopharmacology 42, 23–30.
Ormsby, R. J., Jokiranta, T. S., Duthy, T. G., Griggs, K. M., Sadlon, T. A., Giannakis, E., and Gordon, D. L. (2006). Localization of the third heparin-binding site in the human complement regulator factor H1. Mol. Immunol. 43, 1624–1632.
Ormsby, R. J., Ranganathan, S., Tong, J. C., Griggs, K. M., Dimasi, D. P., Hewitt, A. W., Burdon, K. P., Craig, J. E., Hoh, J., and Gordon, D. L. (2008). Functional and structural implications of the complement factor H Y402H polymorphism associated with age-related macular degeneration. Invest. Ophthalmol. Vis. Sci. 49, 1763–1770.
Pangburn, M. K., Schreiber, R. D., and Müller-Eberhard, H. J. (1977). Human complement C3b inactivator: isolation, characterization, and demonstration of an absolute requirement for the serum protein beta1H for cleavage of C3b and C4b in solution. J. Exp. Med. 146, 257–270.
Pappachan, J. M., Mathew, S., Thomas, B., Renjini, K., Scaria, C. K., and Shukla, J. (2007). The incidence and clinical characteristics of the immune phase eye disease in treated cases of human leptospirosis. Indian J. Med. Sci. 61, 441–447.
Pio, R., Martinez, A., Unsworth, E. J., Kowalak, J. A., Bengoechea, J. A., Zipfel, P. F., Elsasser, T. H., and Cuttitta, F. (2001). Complement factor H is a serum-binding protein for adrenomedullin, and the resulting complex modulates the bioactivities of both partners. J. Biol. Chem. 276, 12292–12300.
Sharma, A. K., and Pangburn, M. K. (1996). Identification of three physically and functionally distinct binding sites for C3b in human complement factor H by deletion mutagenesis. Proc. Natl. Acad. Sci. U.S.A. 93, 10996–11001.
Sharma, A. K., and Pangburn, M. K. (1997). Localization by site-directed mutagenesis of the site in human complement factor H that binds to Streptococcus pyogenes M protein. Infect. Immun. 65, 484–487.
Sjöberg, A., Onnerfjord, P., Mörgelin, M., Heinegård, D., and Blom, A. M. (2005). The extracellular matrix and inflammation: fibromodulin activates the classical pathway of complement by directly binding C1q. J. Biol. Chem. 280, 32301–32308.
Skerka, C., Lauer, N., Weinberger, A. A., Keilhauer, C. N., Sühnel, J., Smith, R., Schlötzer-Schrehardt, U., Fritsche, L., Heinen, S., Hartmann, A., Weber, B. H., and Zipfel, P. F. (2007). Defective complement control of factor H (Y402H) and FHL-1 in age-related macular degeneration. Mol. Immunol. 44, 3398–3406.
Teixeira, A. G., Silva, A. S., Lin, F. L., Velletri, R., Bavia, L., Belfort, R. Jr., and Isaac, L. (2010). Association of complement factor H Y402H polymorphism and age-related macular degeneration in Brazilian patients. Acta Ophthalmol. 88, 165–169.
Verma, A., Artiushin, S., Matsunaga, J., Haake, D. A., and Timoney, J. F. (2005). LruA and LruB, novel lipoproteins of pathogenic Leptospira interrogans associated with equine recurrent uveitis. Infect. Immun. 73, 7259–7266.
Verma, A., Hellwage, J., Artiushin, S., Zipfel, P. F., Kraiczy, P., Timoney, J. F., and Stevenson, B. (2006). LfhA, a novel factor H-binding protein of Leptospira interrogans. Infect. Immun. 74, 2659–2666.
Weiler, J. M., Daha, M. R., Austen, K. F., and Fearon, D. T. (1976). Control of the amplification convertase of complement by the plasma protein beta1H. Proc. Natl. Acad. Sci. U.S.A. 73, 3268–3272.
Whaley, K., and Ruddy, S. (1976). Modulation of the alternative complement pathways by beta 1 H globulin. J. Exp. Med. 144, 1147–1163.
Yu, J., Wiita, P., Kawaguchi, R., Honda, J., Jorgensen, A., Zhang, K., Fischetti, V. A., and Sun, H. (2007). Biochemical analysis of a common human polymorphism associated with age-related macular degeneration. Biochemistry 46, 8451–8461.
Zetterberg, M., Landgren, S., Andersson, M. E., Palmér, M. S., Gustafson, D. R., Skoog, I., Minthon, L., Thelle, D. S., Wallin, A., Bogdanovic, N., Andreasen, N., Blennow, K., and Zetterberg, H. (2008). Association of complement factor H Y402H gene polymorphism with Alzheimer’s disease. Am. J. Med. Genet. B Neuropsychiatr. Genet. 147B, 720–726.
Zipfel, P. F., Jokiranta, T. S., Hellwage, J., Koistinen, V., and Meri, S. (1999). The factor H protein family. Immunopharmacology 42, 53–60.
Keywords: Leptospira, complement system, factor H, Y402H polymorphism
Citation: Silva AS, Valencia MM, Cianciarullo AM, Vasconcellos SA, Barbosa AS, and Isaac L (2011) Interaction of human complement factor H variants Tyr402 and His402 with Leptospira spp.. Front. Immun. 2:44. doi: 10.3389/fimmu.2011.00044
Received: 12 April 2011; Accepted: 29 August 2011
Published online: 05 October 2011.
Edited by:
Uday Kishore, Brunel University, UKCopyright: © 2011 Silva, Valencia, Cianciarullo, Vasconcellos, Barbosa and Isaac. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Lourdes Isaac, Departamento de Imunologia, Instituto de Ciências Biomédicas, Universidade de São Paulo, Av. Prof. Lineu Prestes 1730, São Paulo CEP 05508-900, Brazil. e-mail:bG91aXNhYWNAaWNiLnVzcC5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.