
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 25 September 2024
Sec. Epigenomics and Epigenetics
Volume 15 - 2024 | https://doi.org/10.3389/fgene.2024.1455872
 Dušanka Mitrović1†
Dušanka Mitrović1† Snežana Smederevac1†
Snežana Smederevac1† Lissette Delgado-Cruzata2†
Lissette Delgado-Cruzata2† Selka Sadiković1*†
Selka Sadiković1*† Dejan Pajić1†
Dejan Pajić1† Mechthild Prinz2†
Mechthild Prinz2† Zoran Budimlija3†
Zoran Budimlija3† Milan Oljača1†
Milan Oljača1† Jelena Kušić-Tišma4†
Jelena Kušić-Tišma4† Nataša Vučinić5†
Nataša Vučinić5† Aleksandra Milutinović5†
Aleksandra Milutinović5†Background: This study investigates the relationship between MB-COMT DNA methylation (DNAm) and the personality traits outlined in the NEO-PI-R model through an epigenetic study of monozygotic twins. DNAm, a critical epigenetic mechanism, regulates gene expression and has been linked to various biological processes and disorders. By leveraging the genetic similarities of monozygotic twins, this research explores how epigenetic variations influenced by environmental factors correlate with personality differences.
Methods: The study utilized the Five-Factor Model (FFM) to categorize personality traits into five domains: Neuroticism, Extraversion, Conscientiousness, Agreeableness, and Openness to Experience. Each domain comprises six facets, providing a granular view of personality. The research centered on the catechol-O-methyltransferase (COMT) gene, focusing on its role in dopamine metabolism, which is hypothesized to influence personality traits through the dopaminergic system. DNAm status in the MB-COMT promoter region was examined to determine its association with personality facets.
Results: Preliminary findings suggest a complex interaction between MB-COMT DNAm patterns and personality traits. Specific methylation patterns at different CpG sites were linked to varying expressions of traits such as impulsivity and aggression, highlighting the nuanced impact of epigenetics on personality.
Conclusion: This study underscores the potential of integrating genetic, epigenetic, and environmental data to enhance our understanding of personality formation. The results contribute to a broader understanding of how genetic predispositions shaped by environmental factors manifest in complex trait differences, paving the way for future research in genetic psychiatry and personalized medicine.
Epigenetic modifications, notably DNA methylation (DNAm), are pivotal in regulating gene expression. The prominence of DNAm within regulatory domains such as gene promoters is widely recognized, primarily due to its association with the suppression of gene expression, as elucidated by (Jones, 2012). This epigenetic mechanism is instrumental in orchestrating a myriad of biological processes and plays a significant role in the emergence and manifestation of various disorders. Contemporary research has primarily focused on exploring the impact of DNA methylation in the context of personality and mental disorders. This research trajectory is driven by the hypothesis that these disorders often display more conspicuous and distinguishable phenotypic characteristics, as noted by Gescher et al. (2018) and Thomas et al. (2019). Comprehending the epigenetic underpinnings of such disorders is imperative for deciphering their complex etiology and potentially developing targeted therapeutic strategies.
However, investigating the influence of environmental factors on epigenetic variations presents a distinct challenge, particularly when examining personality traits. In contrast to disorders, personality traits signify dimensions of individual differences that typically demonstrate consistency in their expression and intensity over time. To elucidate the epigenetic foundations of these traits, a multifaceted approach is required, encompassing various levels of the trait hierarchy.
In this context, twin studies represent an invaluable resource. These studies leverage the genetic congruence of monozygotic twins (MZ), who share nearly 100% of their genetic makeup, to scrutinize disparities in their epigenome and phenotype. By contrasting MZ twins with divergent epigenetic profiles, researchers can discern the influence of environmental factors on epigenetic alterations and their correlations with personality traits.
The FFM is widely acknowledged in the psychological literature as a relevant framework for personality analysis. The model comprises five dimensions - Neuroticism, Extraversion, Conscientiousness, Agreeableness, and Openness to Experience, called personality domains, with each domain encompassing six lower-order facets. The best-known operationalization of this model is the NEO-PI-R questionnaire, an extensively validated and reliable tool for personality assessment (Costa and McCrae, 1992; Moore et al., 2010; Soto, 2019), widely recognized for its universality. The stability of the FFM has been attributed in part to the genetic basis underlying traits and shaping their domains (McCrae et al., 2005), a proposition further supported by studies highlighting significant genetic influences on personality traits (Jang et al., 2006; Yamagata et al., 2006). However, some research (Jang et al., 2002; Sadiković, 2023; Smederevac et al., 2020) suggests that the genetic and environmental architecture of personality traits may deviate from their phenotypic manifestations, with individual variances being more influenced by specific genetic and environmental factors than common ones.
The hierarchical structure of the FFM, especially its lower levels of hierarchy, such as facets and nuances, has attracted research attention (Terracciano et al., 2010). These facets, representing more subtle and specific phenotypic expressions within each domain, have shown incremental validity and are influenced by unique genetic and environmental factors (Paunonen and Ashton, 2001; Kandler et al., 2010; Soto and John, 2017). Notably, the reliance only on aggregate scores for each dimension of the FFM is acknowledged as a potential limitation, as it may reduce the statistical power of research in this area (Jang et al., 2002).
Twin studies play a key role in examining the genetic contribution to individual differences in personality traits, indicating that heritability accounts for approximately 50% of the variance (Sadiković, 2023; Smederevac et al., 2023a; Balestri et al., 2014; Bratko et al., 2017; Munafò and Flint, 2011). However, variability in heritability estimates across studies suggests the presence of non-additive gene effects and gene interactions. High genetic correlations among personality traits are hypothesized to result from pleiotropy (Smederevac et al., 2020), linking trait similarity between individuals to their genetic resemblance [e.g., (Turkheimer, 2016)].
Examining the complex interaction between genetic effects and environmental factors, it becomes apparent that the same gene variant can produce different phenotypic outcomes, and different combinations of genes can result in identical phenotypes. This complexity underlines the importance of identifying small, consistent gene variant effects on personality traits (Wagner et al., 2020). Since genes indirectly influence psychological phenomena through biochemical and physiological processes, consideration of brain structure and function appears as a suitable endophenotype for tracing the path from genes to behavior (Moore et al., 2018; Haas et al., 2018).
The dopaminergic system, predominantly expressed in the various brain regions, plays a vital role in dopamine biosynthesis and synaptic regulation (Chen et al., 2011; Peciña et al., 2013; DeYoung et al., 2011; Smillie et al., 2010; Tsuchimine et al., 2012). Existing research, despite presenting mixed findings, generally associates dopaminergic genes with various personality traits, including approach behaviors, extraversion, particularly positive emotionality (Tsuchimine et al., 2012; Demetrovics et al., 2010; Hoth et al., 2006), impulsivity (White et al., 2009), openness to experience (Peciña et al., 2013), and neuroticism, primarily through anxiety, depression, and harm avoidance (Chen et al., 2011; Aluja et al., 2019; Valeeva et al., 2020), as well as the behavioral activation system (Smederevac et al., 2023a). The dopaminergic hypothesis of extraversion suggests a link between the COMT enzyme, critical for dopamine metabolism, and extraversion, either independently (Reuter et al., 2007) or in interaction with other dopamine-related genes (Reuter et al., 2006).
The catechol-O-methyltransferase (COMT) gene, located on chromosome 22q11, is responsible for encoding the COMT enzyme, a pivotal agent in the primary inactivation pathway of dopamine within the brain, particularly manifesting substantial expression in the prefrontal cortex (Chen et al., 2011). This enzyme plays a crucial role in the metabolic decomposition of catecholamines, including adrenaline, noradrenaline, and dopamine (Chen et al., 2011; Demetrovics et al., 2010; Hoth et al., 2006), thereby exerting a significant influence on a spectrum of cognitive functions and personality attributes (Chen et al., 2011; DeYoung et al., 2011). The activity of the COMT enzyme is influenced by various polymorphisms in the COMT gene, which can have potential implications for the pathophysiology of several neurological and psychiatric conditions. One notable single nucleotide polymorphism (SNP) at codon 158 involves a transition from guanine (G) to adenine (A), leading to a change in the amino acid that involves substitution of Valine (Val) with Methionine (Met), which consequently alters the enzyme’s thermal stability and its enzymatic activity (DeYoung et al., 2011; Hoth et al., 2006; Tunbridge et al., 2019). This modification results in a reduction of enzyme activity by approximately 30%–35% in the prefrontal cortex of individuals carrying the Met allele (Chen et al., 2011), leading to an amplification of dopamine signaling due to the predominant role of COMT in dopamine clearance from synaptic spaces (DeYoung et al., 2011; Demetrovics et al., 2010). The three genotypes of COMT—Val/Val, Val/Met, and Met/Met—correspond to high, intermediate, and low enzyme activity, respectively.
Previous research has demonstrated a correlation between neuroticism and diminished activity of the COMT enzyme (Chen et al., 2011), with a notable gender-specific manifestation. The connection with anxiety-related traits, for instance, was predominantly observed in women (Valeeva et al., 2020). Conversely, in men, the Val allele has been linked to increased negative emotionality (Chen et al., 2011), neuroticism, and harm avoidance tendencies (Lee and Prescott, 2014). Furthermore, significant phenotypic variances between carriers of the Val and Met alleles of the COMT V158M polymorphism have been documented in relation to extraversion (Hoth et al., 2006), positive emotionality (Chen et al., 2011), and novelty-seeking behaviors (Demetrovics et al., 2010). Notably, Met/Met homozygotes have demonstrated augmented prefrontal cortical activity during phases of reward anticipation (Dreher et al., 2009), suggesting that the directional association, akin to that with neuroticism, may exhibit gender-specific divergences: women with the Met allele showing reduced extraversion (Hoth et al., 2006), and men with the Val allele demonstrating the opposite (Demetrovics et al., 2010). Moreover, the COMT gene has been implicated in the realm of creative potential (Ren et al., 2019), where a decrease in dopamine removal correlates with elevated synaptic dopamine levels, potentially contributing to enhanced openness traits (DeYoung et al., 2011; Smillie et al., 2010).
The COMT gene occupies a central role in the metabolic processing of catecholamines within the brain, including neurotransmitters such as dopamine. Of particular significance is the membrane-bound catechol-O-methyltransferase (MB-COMT), a notable isoform of this gene. Elucidation of the epigenetic regulation of the MB-COMT promises to shed light on the molecular determinants underpinning individual variances in behavioral, cognitive, and emotional regulatory mechanisms. Attributes such as impulsivity, anxiety, and cognitive capabilities, notably working memory and executive control, are postulated as potential endophenotypes susceptible to modulation by MB-COMT DNAm.
Recent scholarly endeavors have delved into the role of DNAm of the MB-COMT in the modulation of personality traits, with a particular focus on impulsivity. It has been observed that individuals exhibiting specific DNAm patterns in the COMT gene may display varied capacities in modulating impulsive behavior. Dopamine, crucially modulated by COMT, is integral to decision-making and impulse control processes. Variations in COMT DNAm are hypothesized to influence the efficacy of dopamine degradation, thereby contributing to interindividual differences in impulsivity (Smederevac et al., 2023b). Furthermore, investigations into the correlation between MB-COMT DNAm and personality traits, guided by the Revised Reinforcement Sensitivity Theory, have unveiled insights into how MB-COMT gene DNAm may affect behavioral patterns and personality traits. This is particularly evident in traits associated with responses to environmental stimuli, such as impulsivity and aggression (Smederevac et al., 2023b).
This study seeks to contribute to a comprehensive understanding of the interactions between genes, environment and personality traits by integrating genetic, epigenetic and phenotypic data. By such an approach, we aim to advance the insight into the underlying mechanisms shaping personality.
Specifically, our research will explore the relationship between variations in the COMT gene and personality traits within the FFM. The COMT gene, by modulating adrenaline, noradrenaline, and dopamine levels, may affect a variety of cognitive, emotional, and behavioral outcomes, such as cognitive flexibility (Smederevac et al., 2023b), emotional regulation (Sanz et al., 2010), or reward sensitivity (Smederevac et al., 2023a). Therefore, it is important to investigate its effects across the different levels of the personality trait hierarchy outlined by the FFM.
Furthermore, we will investigate how differences in COMT promoter DNAm levels correlate with personality traits. Our approach involves utilizing twin data, relying on the unique genetic similarity inherent in monozygotic twins that allows for discerning the epigenetic changes resulting from environmental influences. Thus, our study aims to assess both genetic and environmental contributions to individual variation in personality traits, with a particular focus on the hierarchical structure of the FFM, including its facets.
The comprehensive recruitment, testing, and data collection process within the Serbian Advanced Twin Registry (STAR) is detailed in Smederevac et al. (2019). From the STAR, which includes data on 1,654 participants (827 twin pairs), we selected all monozygotic twin pairs that had data on all relevant phenotypic measures and were genotyped for COMT V158M polymorphism (rs4680). The study obtained the required ethical clearances from the Institutional Ethical Committees, with the codes #02-374/15, #01-39/229/1, and #O-EO-024/2020. Participation in the study was entirely voluntary, with all participants providing informed written consent before their involvement.
After excluding certain cases due to failed genotyping, the sample comprised 430 twins. The age of the participants ranged from 18 to 60 years (M = 24.66; SD = 7.72; 74.4% women). The sample included more monozygotic (75.8%) than dizygotic twins. Monozygotic twins were aged between 18 and 60 years (M = 25.18; SD = 8.08; 75.5% women). Dizygotic twins ranged from 18 to 48 years of age (M = 23.05; SD = 6.21; 71.2% women).
To investigate the association between DNAm differences and behavioral phenotypic traits, we analyzed a subset of 35 pairs of monozygotic twins (MZ) from the SNP sample, who had high-quality buccal swab DNA suitable for methylation studies. This subset included 16 male and 54 female twins, aged between 18 and 44 years, with an average age of 23.30 years (SD = 6.68).
Revised NEO-PI-R Personality Inventory (Costa and McCrae, 1992; Costa and McCrae, 2019). The NEO-PI-R personality inventory contains 240 items with a five-point Likert response format and is designed to operationalize the five major personality dimensions of the FFM: Neuroticism (N), Extraversion (E), Agreeableness (A), Conscientiousness (C), and Openness to experience (O). Each dimension includes 6 lower-order facets (30 in total), operationalized through 8 items for each personality aspect. Neuroticism encompasses Anxiety, Hostility, Depression, Self-consciousness, Impulsiveness and Vulnerability; Extraversion contains Warmth, Gregariousness, Assertiveness, Activity, Excitement seeking and Positive emotions; Agreeableness includes Trust, Straightforwardness, Altruism, Compliance, Modesty and Tender Mindedness; Conscientiousness covers Competence, Order, Dutifulness, Achievement striving, Self-discipline and Deliberation, while Openness to experience incorporates Fantasy, Aesthetics, Feelings, Actions, Ideas and Values. Reliabilities (α) range from 0.30 for Openness to 0.78 for Neuroticism.
The zygosity of the subjects was determined by DNA isolated from a buccal swab sample by analyzing microsatellite loci. The Investigator24plex GO! kits were used for microsatellite analysis. (Qiagen®, Valencia, CA, United States) or (Applied Biosystems®, Thermofisher Scientific, Waltham, MA, United States) according to the manufacturer’s instructions. Buccal swabs are tested using STR (short tandem repeat) megaplex sets according to the manufacturer’s instructions, and they are of two types: Investigator 24plex GO! (Qiagen®, Valencia, CA, United States) and GlobalFiler (Applied Biosystems®, Thermofisher Scientific, Waltham, MA, United States).
Both kits detect 21 autosomal microsatellite loci. Samples with partial profiles were interpreted if a result was present at least 10 gene loci. Fully concordant (identical) pairs are categorized as monozygotic twins based on microsatellite profiles, and all others as dizygotic twins. One part of the DNA samples was analyzed at the Institute of Forensic Medicine in Novi Sad, and the other at the John Jay College of Criminal Law in New York.
The COMT gene V158M polymorphism (rs4680) genotyping was performed using the TaqMan assay (TaqMan SNP, Applied Biosystems®, Warrington, United Kingdom) at the Faculty of Medicine, University of Novi Sad. TaqMan single nucleotide polymorphism genotyping assays are based on 5′-nuclease activity to detect and amplify specific polymorphisms in purified DNA samples and use probes that bind to the minor groove for better discrimination and accurate determination of allelic type. Samples for polymerase chain reaction (PCR) were prepared from 10 ng of genomic DNA with 1 µL of TaqMan genotyping assay and 12.5 µL of genotyping master mix in a final volume of 25 µL. For the polymerase chain reaction, a 96-well microplate, and an ABI Prism 7500 Fast PCR device (Applied Biosystems®, Foster City, California, United States) are used.
The COMT rs4680 polymorphism (V158M) was defined by three genotypes: 129 low active homozygotes (Met/Met), then 224 heterozygotes (Met/Val), and 77 highly active homozygotes (Val/Val). Individuals with at least one copy of the Val allele were grouped into the Val+ group (301 subjects), while Met homozygotes formed the Met + group (129 subjects). The studied population was in Hardy-Weinberg equilibrium (HWE) for the COMT V158M polymorphism (rs4680; χ2 = 0.85; df = 2; p > 0.05).
The COMT genotypes in the epigenetic sample were also categorized into the Met/Met group (23 subjects), Met/Val group (37 subjects), or Val/Val group (10 subjects). All individuals who had at least one copy of the Val allele were grouped into the Val+ group (47 subjects), while Met homozygotes formed the Met+ group (23 subjects). The studied population for epigenetics was also in HWE for COMT V158M polymorphism (rs4680; χ2 = 5.06; df = 2; p > 0.05).
Genomic DNA was isolated from buccal swabs utilizing the QIAamp DNA Mini Kit (Qiagen®, Valencia, CA, United States), adhering to the protocol provided by the manufacturer. The DNA underwent bisulfite conversion through the EZ DNA Methylation-Gold kit (Zymo Research), following its specific instructions. The bisulfite-treated DNA was then eluted in 15 μL of water, and an aliquot of 2.5–5 μL was employed for Pyromark PCR amplification (Qiagen). This assay was designed to target a 228-bp region in the promoter of MB-COMT, one of the isoforms of the COMT gene. We have selected this region for investigation because membrane-bound COMT (MB-COMT) is mainly expressed in the brain and ultimately impacts behavior. The part of the promoter under study here has been investigated in association to schizophrenia and bipolar disorder in two previous studies (Abdolmaleky et al., 2006; Nohesara et al., 2011). The assay used here to determine DNA methylation levels of MB-COMT has been previously described (Smederevac et al., 2023b). We used two primers, the forward primer sequence was 5′-TGGGGTAGATTAGGGTTGT-3′, and the biotinylated reverse primer at the 5′ end was 5′-CCACACCCTATCCCAATATTC-3′. The amplification process followed the guidelines of the Pyromark PCR Kit (Qiagen). Pyrosequencing was conducted on a PyroMark Q24 system (Qiagen), adhering to the manufacturer’s protocol. The sequencing primer used was 5′-GGATAGGGGAGGGTTTAGTT-3′, and the sequence analyzed was 5′-TYGGGYGGGTYGTYGYGGGAGAGGTGAGAG-3′. This method assessed the DNAm levels of five CpG sites. Pyrosequencing measured DNA methylation levels of five CpG sites are presented in Figure 1. DNAm was analyzed using PyroMark Q24 Advanced 3.0.0 software. The results were expressed as a percent DNAm at each of the five CpG sites. Each amplification and pyrosequencing batch included fully methylated and unmethylated DNA [using the Human Methylated and Non-Methylated (WGA) DNA Set (product number D5013) from Zymo Research] as controls, alongside no-template controls in all runs.
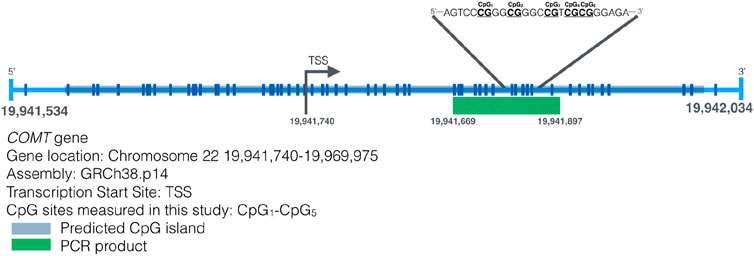
Figure 1. Map of the transcription start site of the MB-COMT gene and the CpG sites on that region (Smederevac et al., 2023b).
The statistical analyses applied to the SNP dataset encompassed descriptive statistics, correlations, t-tests, and non-parametric alternatives to the t-test for independent samples, post hoc tests, and effect size estimation. All analyses were conducted using the JASP statistical program (JASP Team, 2022). The level of statistical significance for each of the applied t-tests was adjusted using the Bonferroni correction for multiple comparisons. In cases where certain prerequisites for using the t-test were not met (conditions of homogeneity of variances or normality of distribution), alternatives to the t-test for independent samples were employed, namely, Welch’s test (for heterogeneous variances) or the non-parametric Mann-Whitney U test (for distributions significantly deviating from normal). For Welch’s test, Cohen’s d was presented as the effect size, and for the Mann-Whitney U test, rank biserial correlation coefficients were used. The common interpretation of effect size for Cohen’s d is small (d = 0.20), medium (d = 0.50), or large (d = 0.80), based on the guidelines provided by Cohen (Cohen, 1988). Along with the effect size, confidence intervals for the effect size (95% CI) were provided.
The primary goal of this analysis was to detect association strength between DNAm levels at various CpG sites and the levels of personality traits. Due to a relatively small sample size, high skewness, and large variations in DNAm value ranges (see the Results section), twin pairs were compared dichotomously, using more robust non-parametrical tests. They were divided into four categories based on two criteria: higher/lower level of methylation and higher/lower level of trait expression. For instance, the “lower-lower” group included twin pairs where the first twin exhibited both a lower methylation level at a specific CpG site and a lower score on a personality trait or facet.
Two-by-two contingency tables for each pair of CpG site and trait/facet were analyzed using Barnard’s unconditional exact test. This test was selected over the more commonly used Fisher’s exact test because the goal was to determine whether higher levels of methylation are associated with higher or lower levels of trait expression. Consequently, Barnard’s test was expected to be more appropriate since it does not condition on both margins being fixed, allowing for potentially greater test power in detecting associations. This gain in power is particularly noticeable when sample sizes are small and research scenarios imply multiple comparisons (Cheng and He, 2023).
All statistical analyses in this section were performed using the Python SciPy library (Virtanen et al., 2020). Contingency tables were visualized as bubble charts using the Python Plotly library (Plotly Technologies Inc, 2015).
In Supplementary Table SA in Supplementary Material the descriptive statistics for the FFM domains and facets of neuroticism, extraversion, openness to experience and conscientiousness on groups of COMT gene polymorphism V158M (rs4680) carriers were given. The results indicate significant and robust effects of the COMT polymorphism V158M rs4680 genotype on self-consciousness from the domain of neuroticism (U = 22,341.5; p < 0.05; pbonf < 0.05; d = 0.151; d (CI) = 0.033–0.265). Those in the Met+ group show significantly higher scores on self-consciousness than those in the Val+ group (Supplementary Table SA). Correlations between NEO-PI-R domains and facets (Supplementary Table SB) and intra-class correlations between twins on domains and facets of the NEO-PI-R (Supplementary Table SC) are given in Supplementary Material.
The COMT V158M (rs4680) genotype shows robust main effects on extraversion (U = 15,777.5; p < 0.01; pbonf < 0.01) and facets of extraversion: warmth (U = 15,371.5; p < .01; pbonf < 0.01), gregariousness (U = 15,564; p < 0.01; pbonf < 0.01) and positive emotions (U = 16,275.5; p < 0.01; pbonf < 0.05) (Table 1). The main effects of the COMT V158M (rs4680) genotype on the facets of extraversion remain significant after correction for multiple comparisons, and effect sizes are largest for warmth (mean effect size; d = −0.208; d (CI) = −0.319–−0.092), gregariousness (d = −0.198; d (CI) = −0.310–−0.082), and extraversion domain (d = −0.187; d (CI) = −0.299–−0.070), and Val+ group show statistically significantly higher scores on extraversion, warmth, gregariousness, and positive emotions (Table 1).
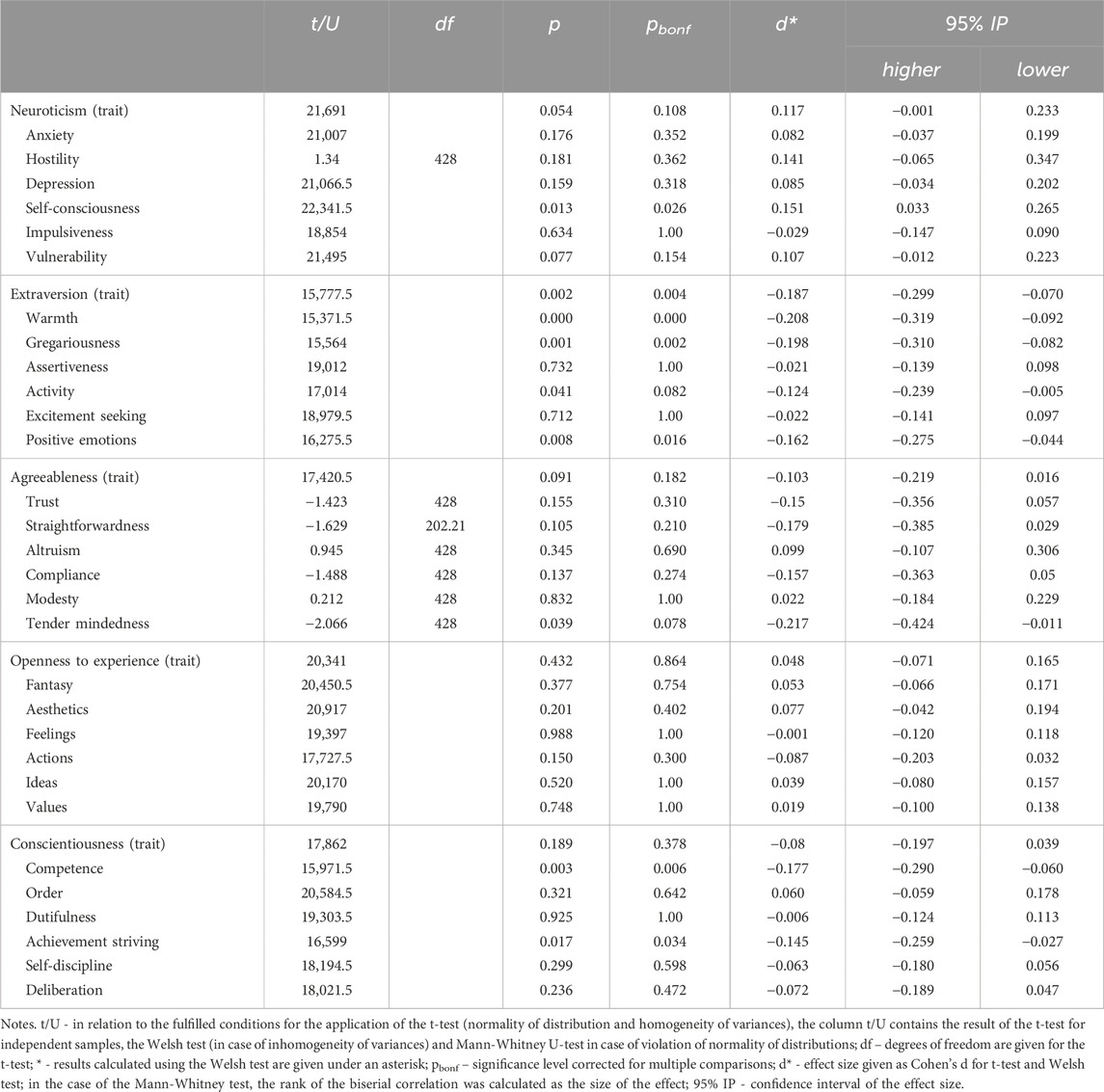
Table 1. Main effects of the COMT V158M (rs4680) genotype on domains and facets of every domain of the FFM.
Moreover, the COMT V158M (rs4680) genotype shows significant main effects on competence (U = 15,971.5; p < 0.01; pbonf < 0.01; d = −0.177; d (CI) = −0.290–−.060) and achievement striving (U = 16,599; p < 0.05; pbonf < 0.05; d = −0.145; d (CI) = −0.259–−.027). As with the extraversion domain, the carriers of the Val + allele show statistically significantly higher scores on both the competence and the achievement striving. The results indicated that the COMT V158M (rs4680) genotype showed no significant main effects on other aspects of neuroticism, extraversion, openness to experience and conscientiousness.
In Supplementary Table SD of the Supplementary Material, the main effects of COMT V158M (rs4680) genotype status on COMT promoter CpG levels are presented. There were no statistically significant effects of the V158M (rs4680) genotype on the DNAm levels of any CpGs.
Table 2 presents basic descriptive statistics for DNAm levels at five analyzed CpG sites and scores across five NEO-PI-R domains. Due to a violated assumption of normality for DNAm levels, we opted for Barnard’s test as a more robust non-parametric method for assessing the association between categorical variables. Another rationale for this choice was the intention to assess association strength in terms of contingencies, specifically to compare twins within each pair and evaluate the likelihood of a twin having higher level of methylation to also obtain higher or lower score on a specific personality domain or facet. In line with that, we divided twin pairs into four groups based on two dichotomous criteria: higher/lower score on DNAm level for the first twin and higher/lower score on NEO-PI-R domain or facet for the same twin in a twin pair.
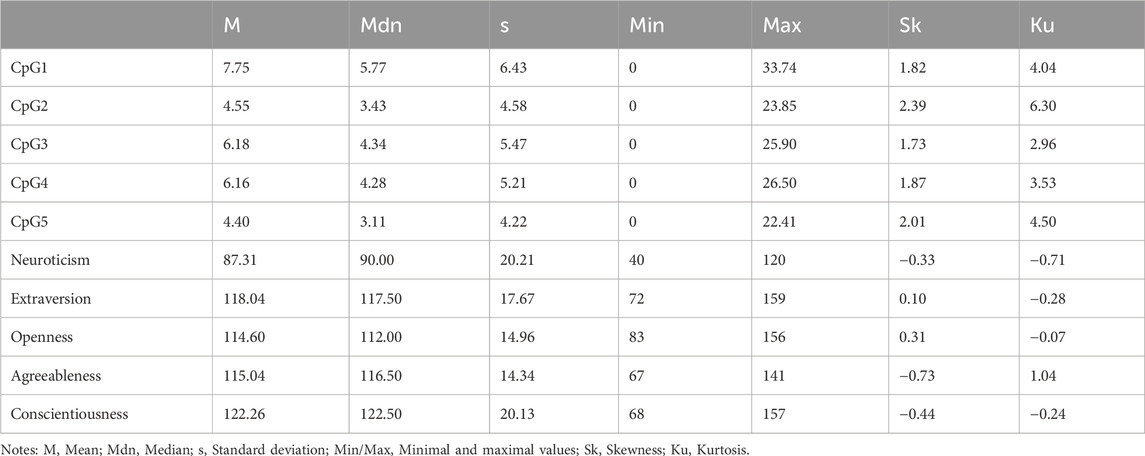
Table 2. Descriptive statistics for DNAm levels on five CpG sites and scores on five main NEO-PI-R domains.
Hypothesis testing was conducted using a one-sided approach. Let p1 represent the theoretical binomial probability that a twin has a higher DNAm level and a lower score on a personality trait compared to his or her sibling. Let p2 denote the theoretical binomial probability that a twin has both a higher DNAm level and a higher score on a personality trait than his or her sibling. Setting the alternative (tail) to “greater” would then test the assumption: H0: p1 ≤ p2 vs H1: p1 > p2. Conversely, setting the alternative (tail) to “less” would test the assumption: H0: p1 ≥ p2 vs H1: p1 < p2. In line with these hypotheses, lower score categories were treated as “not higher” under the “greater” alternative, and hence included pairs of twins with identical scores on the personality trait/facet. Similarly, higher score categories were considered “not lower” under the “less” alternative, thus including pairs of twins who had the same score on the personality trait/facet. Only one twin pair had identical DNAm levels (zero) at only one site (CpG2), and this pair was excluded from all the analyses that included this CpG site. Supplementary Table SE, F of the Supplementary Material show Barnard’s test values for association between each DNAm level and each NEO-PI-R dimension and facet.
Figure 2 shows contingency tables visualized as bubble charts and corresponding Barnard’s test values for statistically significant associations between methylation levels on five CpG sites and NEO-PI-R facets under the “greater” alternative hypothesis. In general, higher DNAm levels are associated with higher scores on facets of Extraversion (Warmth and Gregariousness) and Agreeableness (Trust and Modesty). Additionally, Values facet has a bordering significant association with the DNAm level on CpG3 site.
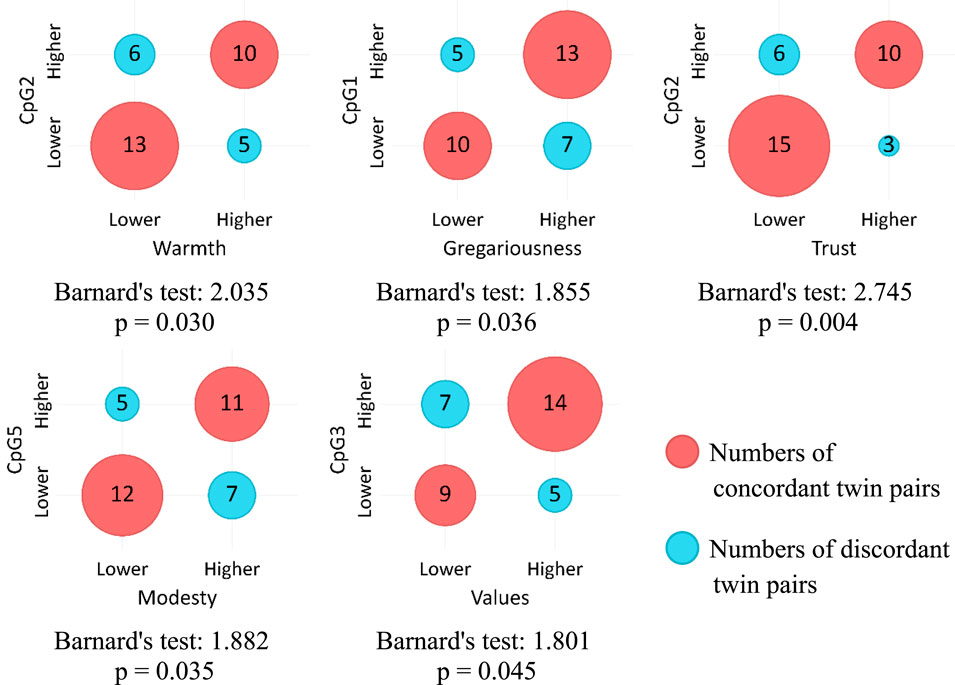
Figure 2. Association between COMT DNAm levels and NEO-PI-R facets (“greater” alternative).
Figure 3 shows contingency tables and corresponding Barnard’s test values for statistically significant associations between DNAm levels on five CpG sites and NEO-PI-R traits and facets under the “less” alternative hypothesis. Lower DNAm levels on multiple CpG sites are primarily associated with higher scores on facets of Neuroticism (Hostility, Depression, Impulsiveness). Extraversion and its facets (Excitement Seeking, Positive Emotion), as well as Agreeableness and its facets (Straightforwardness, Tender-Mindedness) again show significant associations, but this time in opposite directions and at different CpG sites.
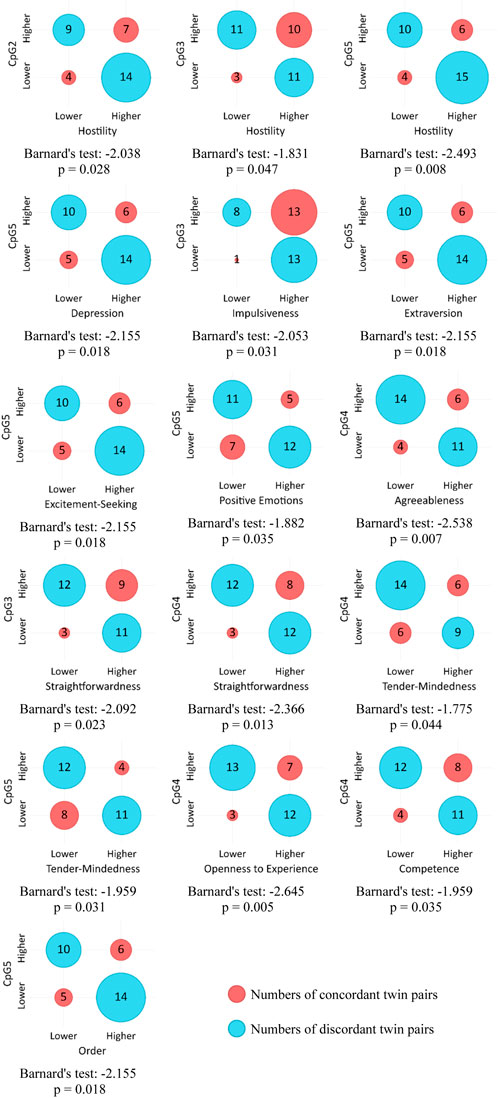
Figure 3. Association between COMT DNAm levels and NEO-PI-R facets (“less” alternative).
The main goal of this research was to examine the role of the COMT V158M (rs4680) genotype in the genetic basis of personality traits according to the FFM. In addition to the relationships between the polymorphism of this gene and the domains and facets of the FFM, we were also interested in whether the level of DNAm of the promoter region of this gene, assumed to be influenced by environmental factors, shows a connection with different levels of FFM domains and facets.
Results of the association study indicate a connection between the COMT V158M (rs4680) polymorphism and traits within the domains of Neuroticism, Extraversion, and Conscientiousness. Specifically, individuals carrying the Met allele tend to exhibit heightened levels of Self-Consciousness (N), whereas those with the Val allele display higher Extraversion, as well as its facets Gregariousness, Warmth, and Positive Emotions. They also score higher on competence and Achievement Striving within the Conscientiousness domain.
Thus, reduced COMT enzyme activity in Met allele carriers, resulting in higher synaptic dopamine levels (Demetrovics et al., 2010), is linked to social anxiety and heightened sensitivity to social reinforcement. Due to increased dopaminergic signaling, individuals carrying the Met allele may experience greater self-consciousness and display more inhibited behavior in social interactions, as dopamine is associated with behavioral control (Chmielowiec et al., 2018). Heightened reactivity and focus on internal experiences could potentially result in a tendency to interpret social cues as threatening.
Conversely, heightened COMT enzyme activity in Val allele carriers, resulting in reduced dopaminergic activity, is associated with sociability, friendly behavior, and a positive mood, aligning with the dopamine hypothesis of extraversion (Depue and Collins, 1999). Engaging in social interactions and adopting approach-oriented behavior might serve as a means to boost baseline dopamine signaling in the cortex. Moreover, the pleasure derived from social encounters could stimulate the reward dopamine system, potentially compensating for lower dopamine levels in synapses.
Furthermore, carriers of the Val allele also exhibit more pronounced self-confidence and ambition. Although Conscientiousness strongly implies volitional control of behavior, leading to the anticipation that carriers of the Met allele might exhibit higher scores, its facets Competence and Achievement Striving are more aligned with approach behavior and the pursuit of rewards rather than with impulse control or delaying reinforcement.
In general, the connections between the COMT V158M (rs4680) polymorphism and personality traits suggest that increased dopamine signaling in carriers of the Met allele is associated with a specific form of sensitivity to negative reinforcement, as well as tendencies toward avoidance behavior and behavioral inhibition. Some previous findings pointed to the increased Neuroticism in Met carriers (e.g., Chen et al., 2011; Stein et al., 2005), which is partially in line with our results. However, our study didn't show the link to the other aspects of Neuroticism, nor the Neuroticism domain as a whole, suggesting only a specific association with social inhibition. Conversely, lower dopamine levels in Val allele carriers are associated with reward sensitivity, active attitude, and behavioral approach. The expectation of a connection between higher COMT enzyme activity and impulsive behavior, drawn from previous findings (e.g., Smederevac et al., 2023a; White et al., 2009), is not supported by the results, indicating that lower dopamine levels are associated with approach-oriented behavior rather than uncontrolled impulsivity.
Given the significant role of environmental factors in the emergence of epigenetic modifications, the study also focused on exploring how these changes correlate with personality traits. The objective was to investigate the link between DNAm levels in CpG sites of the COMT promoter and personality trait expression, aiming to contribute to the understanding of the mechanisms through which environmental factors affect personality phenotypes. Findings indicate that DNAm of certain CpG sites of the COMT promoter has additional effects on dopamine function and consequently behavior, revealing a specific pattern of associations. Notably, traits that are more pronounced in individuals with higher methylated CpG sites of the COMT gene promoter are quite distinct from traits that are less pronounced in the case of elevated DNAm. Furthermore, a pattern emerges wherein methylated CpG sites associated with heightened levels of certain personality traits differ from those associated with lower levels of some traits. Hence, it appears that DNAm at different sites on the COMT gene promoter may not uniformly affect behavior.
Traits more pronounced in individuals with higher DNAm levels are primarily linked to CpG1, CpG2, and CpG5 sites. These include facets of Extraversion (Gregariousness, Warmth) and Agreeableness (Trust, Modesty), which foster quality relationships and intimacy, suggesting that DNAm at certain sites, mostly CpG2, of the MB-COMT gene promoter uniquely impact traits associated with affiliation potential through its influence on dopaminergic function. Moreover, higher DNAm of CpG1, CpG3, and CpG5 is associated with greater sociability, increased openness to values and tolerance for diversity, as well as heightened modesty. Generally, traits more pronounced in individuals with higher methylated CpGs are those relevant to smooth social interactions and harmonious relationships: friendly demeanor, trust in others, acceptance of diversity, and modesty. Since increased DNAm is predominantly associated with gene silencing, it appears that the enhanced expression of these traits results from the lower amounts of COMT enzyme, leading to heightened dopamine signaling in some parts of the brain.
On the other hand, a much larger number of traits, across all five personality domains, seems to be less pronounced in individuals with higher levels of DNAm. These include some of the domains mentioned before, but now associated with DNAm levels at different sites: CpG2, CpG3, and CpG5. For example, some facets of Extraversion were found to be associated with CpG1 and CpG2 sites under the “greater” alternative, while the lower expression of other facets of the same domain are associated with higher levels of DNAm at the CpG5 site including Excitement Seeking, Positive Emotion, and Extraversion itself. Similarly, some of the Agreeableness facets (Trust, Modesty) were associated with CpG2 and CpG5 sites under the “greater” alternative, while others (Straightforwardness, Tender-Mindedness) are associated with CpG3 and CpG4 sites under the “less” alternative.
Several facets of Neuroticism exhibit lower expression in individuals with higher DNAm levels. Most notably, lower Hostility is associated with higher DNAm at CpG2, CpG3, and CpG5. This finding indicates the association between an angry and hostile attitude and decreased dopamine signaling, potentially leading to a sense of reward deprivation. Namely, tendency to frequently experience anger may partly originate from the perception of being unfairly treated (Sanz et al., 2010) and from the belief that others receive more reinforcement than oneself. It may imply somewhat decreased control as well.
Previous research has linked impulsivity to the activity of the COMT gene (Smederevac et al., 2023a; White et al., 2009; Smederevac et al., 2023b). Our findings show that Impulsiveness is negatively related to the DNAm level of CpG5, if the COMT gene is silenced and dopamine degradation function declines, Impulsiveness (N) is lower. This finding aligns with expectations, as Impulsiveness entails a weakened control over basic urges and an immediate quest for reinforcement, which is possibly related to reduced dopamine levels in certain cortical regions. A somewhat similar explanation applies to Excitement Seeking and Positive Emotions facets within the Extraversion domain, which also exhibit decreased expression in individuals with higher DNAm levels at CpG5, and to the association of Openness and CpG4 DNAm as well. Here, lower dopamine signaling seems to drive the quest for reward and positive reinforcement. While Impulsiveness, as the Neuroticism facet, involves low self-control, heightened tension (Costa and McCrae, 1992) and the pursuit of means to reduce it, Excitement-Seeking and Positive Emotions simply involve continual searching for positive stimulation. Nevertheless, these dimensions all seem to imply some form of compensating for the deficiency of dopamine in the system through the pursuit of reward. Moreover, the association between Straightforwardness (A) and DNAm levels of CpG3 and CpG4 further supports the link between dopamine and behavior control, as honesty and simplicity may imply some form of unrestrained behavior, albeit differing in nature from Impulsiveness. Although not associated with the lack of control, the Agreeableness domain and its Tender-Mindedness facet may be linked to decreased dopamine levels through heightened sensitivity to reward signals, as these traits facilitate positive social reinforcement and acceptance.
Findings regarding the association between depression and COMT V158M (rs4680) polymorphism are inconsistent (Antypa et al., 2013), suggesting that the relationship might be complex. Furthermore, epigenetic studies suggest different effects of DNAm in the promoter of COMT on prefrontal white matter connectivity in healthy individuals compared to those with major depressive disorder (MDD). While higher COMT DNAm is associated with weaker connectivity in healthy subjects, it's linked with improved connectivity in MDD patients (Na et al., 2018). Our results indicate a negative correlation between Depressiveness and increased CpG5 DNAm, suggesting that decreased dopamine levels might lead to a propensity for frequent experiences of sadness and diminished motivation. This association was somewhat surprising, particularly given its alignment with the connection to Positive Emotion. One potential explanation involves a sense of being deprived of rewards due to decreased dopamine signaling, which could trigger feelings of anger, as demonstrated in the case of Angry Hostility, but also feelings of sadness and decline in motivation (Lyon et al., 2020). Additionally, depression is associated with reduced cognitive control, which may result from motivational deficits (Grahek et al., 2019). Furthermore, the findings that individuals with high DNAm levels in CpG5 and CpG4 score lower on the Conscientiousness facets of Order and Competence suggest that traits associated with organization and preparedness could also stem from attempts to activate the reward system in response to reduced dopamine levels, but possibly to overcompensate the sense of insufficient behavioral control as well.
The findings obviously indicate a variety of behavioral responses to decreased dopamine levels in the reward system and suggest the likely role of numerous mechanisms, many of them beyond DNAm of COMT gene, that may contribute to shaping these responses. For example, some of these behavioral traits imply a lack of control, and the others possibly an effort to maintain control. It is crucial to investigate environmental factors that contribute to decreasing or increasing DNAm, consequently leading to behavioral changes. Some of these factors, for example, could involve negative life experiences and stress (McGowan et al., 2009), as is possibly the case with Depression and Angry Hostility.
Although the small sample in the epigenetic aspect of the study made it impossible to examine the relationships of specific polymorphisms with DNAm levels, indirectly one can observe tendencies that are important for tracing future research questions. Namely, the findings regarding the relationship between DNAm status at specific CpG sites of the COMT promoter and the expression of personality traits suggest a more complex role of this gene in behavior, which is influenced by environmental factors. Notably, the observed correlation between increased DNAm at CpG1 and heightened Gregariousness, and CpG2 DNAm with Warmth, appears somewhat at odds with findings indicating increased expression of these traits in Val allele carriers, given that higher DNAm typically suppresses gene expression. Furthermore, higher CpG2 DNAm was found to have close to significant correlation with higher Competence, whereas CpG4 DNAm is associated with lower Competence. This implies that DNAm at particular CpG sites of the COMT gene exerts additional effects on dopamine function and behavior, possibly interacting with COMT V158M (rs4680) polymorphism to influence behavior (van der Knaap et al., 2014). Since epigenetic changes are dynamic and influenced by environmental factors such as effort and stress (Wiegand et al., 2021), it’s also possible that some transient factors contributed to these results.
The key strengths of this study lie in using the methodological advantages of twin samples to explore the interplay between genetic factors and environmental influences on personality traits. However, certain limitations warrant consideration, potentially hindering straightforward generalization of the findings. Firstly, the use of a SNP approach in the GWAS era may oversimplify the interpretation of results. Namely, psychological phenotypic features, which are the focus of this study, are determined by a multitude of genes and their possible interactions. Therefore, our findings should be regarded as a contribution to the growing body of evidence linking the COMT gene to behaviors associated with impulse control and reward seeking. Secondly, the sample of twins in the epigenetic part of the study is relatively modest. Although the statistical methodologies employed were tailored to address the specific research inquiries, replication on larger cohorts is warranted for robust validation. Thirdly, an important limitation of our study is the use of buccal swabs to measure DNA methylation, which may not fully capture the epigenetic landscape of brain tissues (Davies et al., 2012; Ladd-Acosta et al., 2007). However, it may be possible to identify consistencies between buccal and brain tissue through approaches such as comparing methylation quantitative trait loci (meQTLs) (Smith et al., 2014; Lowe et al., 2013). A fourth important limitation is that in this study a mechanistic link between low methylation and high COMT transcription was not investigated with a model system. However, a previous study from another group showed low levels of DNA methylation at highly expressed COMT in iPSC-derived dopaminergic neurons from patients with Parkinson’s disease, compared to healthy controls (Kuzumaki et al., 2019). Finally, the hierarchical nature of the psychological phenotypes examined in this study, such as personality traits, underscores the importance of instrument reliability in their assessment. Despite potential limitations in the reliability of individual facets, the NEO-PI-R is widely used measure of the FFM, demonstrating validity in prior studies with Serbian twin samples (Mitrović et al., 2023; Nikolašević et al., 2021) and facilitating cross-cultural and cross-linguistic comparisons (Smederevac et al., 2020).
The study reveals significant links between the V158M polymorphism of the COMT gene and personality traits like Neuroticism, Extraversion, and Conscientiousness. Met allele carriers tend to have higher self-consciousness and social anxiety, while Val allele carriers exhibit more extraversion, sociability, and ambition. This suggests that COMT has pleiotropic effects, influencing dopamine signaling and behavior, impacting approach and avoidance tendencies.
Exploring DNAm levels of CpG sites in the COMT gene’s promoter region shows a complex relationship with personality traits. Higher DNAm levels at certain CpG sites are linked to traits like warmth, trust, and sociability, while lower DNAm levels relate to reduced expression of traits like hostility and impulsivity. These findings underscore how epigenetic modifications affect dopamine function and behavior, necessitating further study on their interaction with genetic and environmental factors.
Although the study didn’t directly examine the link between specific polymorphisms and DNAm levels in COMT, indirect observations suggest a complex interplay between genetic variation and epigenetic modifications in shaping behavior. This emphasizes the importance of considering both genetic and epigenetic factors to understand individual differences in personality and behavior, paving the way for future research on gene-environment interactions.
The data presented in the study are deposited in OSF repository, accession link https://osf.io/bu3xp/.
The study obtained the required ethical clearances from the Institutional Ethical Committees of Faculty of Philosophy (University of Novi Sad, Department of Psychology), Faculty of Medicine (University of Novi Sad) and Institute of Molecular Genetics and Genetic Engineering (University of Belgrade), with the codes #02-374/15, #01-39/229/1, and #O-EO-024/2020. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
DM: Conceptualization, Investigation, Writing–original draft, Writing–review and editing. SnS: Conceptualization, Funding acquisition, Investigation, Project administration, Writing–original draft. LD-C: Formal Analysis, Resources, Writing–review and editing. SeS: Conceptualization, Data curation, Investigation, Methodology, Writing–original draft. DP: Formal Analysis, Methodology, Software, Visualization, Writing–original draft. MP: Investigation, Resources, Supervision, Writing–review and editing. ZB: Data curation, Formal Analysis, Validation, Writing–review and editing. MO: Data curation, Formal Analysis, Writing–review and editing. JK-T: Formal Analysis, Supervision, Writing–review and editing. NV: Investigation, Project administration, Validation, Writing–review and editing. AM: Data curation, Formal Analysis, Writing–original draft.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was fully supported by the Science Fund of the Republic of Serbia (#7744418, Genetic and environmental influences on psychological adaptation of children and adults–GENIUS).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1455872/full#supplementary-material
Abdolmaleky, H. M., Cheng, K. hung, Faraone, S. V., Wilcox, M., Glatt, S. J., Gao, F., et al. (2006). Hypomethylation of MB-COMT promoter is a major risk factor for schizophrenia and bipolar disorder. Hum. Mol. Genet. 15 (21), 3132–3145. doi:10.1093/hmg/ddl253
Aluja, A., Balada, F., Blanco, E., Fibla, J., and Blanch, A. (2019). Twenty candidate genes predicting neuroticism and sensation seeking personality traits: a multivariate analysis association approach. Personality Individ. Differ. 140, 90–102. doi:10.1016/j.paid.2018.03.041
Antypa, N., Drago, A., and Serretti, A. (2013). The role of COMT gene variants in depression: bridging neuropsychological, behavioral and clinical phenotypes. Neurosci. and Biobehav. Rev. 37 (8), 1597–1610. doi:10.1016/j.neubiorev.2013.06.006
Balestri, M., Calati, R., Serretti, A., and De Ronchi, D. (2014). Genetic modulation of personality traits: a systematic review of the literature. Int. Clin. Psychopharmacol. 29 (1), 1–15. doi:10.1097/YIC.0b013e328364590b
Bratko, D., Butković, A., and Vukasović Hlupić, T. (2017). Heritability of personality. Psihol. teme 26 (1), 1–24. doi:10.31820/pt.26.1.1
Chen, C., Chen, C., Moyzis, R., Dong, Q., He, Q., Zhu, B., et al. (2011). Sex modulates the associations between the COMT gene and personality traits. Neuropsychopharmacol 36 (8), 1593–1598. doi:10.1038/npp.2011.39
Cheng, H., and He, J. (2023). Comparison of tests for association of 2 × 2 tables under multiple testing setting. Commun. Statistics - Simul. Comput. 52 (6), 2336–2348. doi:10.1080/03610918.2021.1905843
Chmielowiec, J., Chmielowiec, K., Suchanecka, A., Trybek, G., Mroczek, B., Małecka, I., et al. (2018). Associations between the dopamine D4 receptor and DAT1 dopamine transporter genes polymorphisms and personality traits in addicted patients. Int. J. Environ. Res. Public Health 15 (10), 2076. doi:10.3390/ijerph15102076
Cohen, J. (1988). Statistical power analysis for the behavioral sciences. New York: Lawrence Erlbaum Associates.
Costa, P. T., and McCrae, R. R. (1992). Revised NEO personality inventory (NEO-PI-R) and NEO five-factor inventory (NEO-FFI) professional manual. Odessa, FL: Psychological Assessment Resources.
Costa, P. T., and McCrae, R. R. (2019). Srpska standardizacija NEO petofaktorskog inventara NEO-FFI: forma S [The Serbian standardization of the NEO five-factor inventory NEO-FFI: form S]. Beograd: Sinapsa edicije.
Davies, M. N., Volta, M., Pidsley, R., Lunnon, K., Dixit, A., Lovestone, S., et al. (2012). Functional annotation of the human brain methylol identifies tissue-specific epigenetic variation across brain and blood. Genome Biol. 13 (6), R43. doi:10.1186/gb-2012-13-6-r43
Demetrovics, Z., Varga, G., Szekely, A., Vereczkei, A., Csorba, J., Balazs, H., et al. (2010). Association between Novelty Seeking of opiate-dependent patients and the catechol-O-methyltransferase Val158Met polymorphism. Compr. Psychiatry 51 (5), 510–515. doi:10.1016/j.comppsych.2009.11.008
Depue, R. A., and Collins, P. F. (1999). Neurobiology of the structure of personality: dopamine, facilitation of incentive motivation, and extraversion. Behav. Brain Sci. 22 (3), 491–517. doi:10.1017/s0140525x99002046
DeYoung, C. G., Cicchetti, D., Rogosch, F. A., Gray, J. R., Eastman, M., and Grigorenko, E. L. (2011). Sources of cognitive exploration: genetic variation in the prefrontal dopamine system predicts Openness/Intellect. J. Res. Personality 45 (4), 364–371. doi:10.1016/j.jrp.2011.04.002
Dreher, J. C., Kohn, P., Kolachana, B., Weinberger, D. R., and Berman, K. F. (2009). Variation in dopamine genes influences responsivity of the human reward system. Proc. Natl. Acad. Sci. 106 (2), 617–622. doi:10.1073/pnas.0805517106
Gescher, D. M., Kahl, K. G., Hillemacher, T., Frieling, H., Kuhn, J., and Frodl, T. (2018). Epigenetics in personality disorders: today’s insights. Front. Psychiatry 9, 579. doi:10.3389/fpsyt.2018.00579
Grahek, I., Shenhav, A., Musslick, S., Krebs, R. M., and Koster, E. H. W. (2019). Motivation and cognitive control in depression. Neurosci. and Biobehav. Rev. 102, 371–381. doi:10.1016/j.neubiorev.2019.04.011
Haas, B. W., Smith, A. K., and Nishitani, S. (2018). Epigenetic modification of OXTR is associated with openness to experience. Personal. Neurosci. 1, e7. doi:10.1017/pen.2018.7
Hoth, K. F., Paul, R. H., Williams, L. M., Dobson-Stone, C., Todd, E., Schofield, P. R., et al. (2006). Associations between the COMT Val/Met polymorphism, early life stress, and personality among healthy adults. Neuropsychiatric Dis. Treat. 2 (2), 219–225. doi:10.2147/nedt.2006.2.2.219
Jang, K. L., Livesley, W. J., Ando, J., Yamagata, S., Suzuki, A., Angleitner, A., et al. (2006). Behavioral genetics of the higher-order factors of the Big Five. Personality Individ. Differ. 41 (2), 261–272. doi:10.1016/j.paid.2005.11.033
Jang, K. L., Livesley, W. J., Angleitner, A., Riemann, R., and Vernon, P. A. (2002). Genetic and environmental influences on the covariance of facets defining the domains of the five-factor model of personality. Personality Individ. Differ. 33 (1), 83–101. doi:10.1016/s0191-8869(01)00137-4
JASP Team (2022). Jasp. Available at: https://jasp-stats.org/.
Jones, P. A. (2012). Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 13 (7), 484–492. doi:10.1038/nrg3230
Kandler, C., Riemann, R., Spinath, F. M., and Angleitner, A. (2010). Sources of variance in personality facets: a multiple-rater twin study of self-peer, peer-peer, and self-self (Dis)Agreement. J. Personality 78 (5), 1565–1594. doi:10.1111/j.1467-6494.2010.00661.x
Kuzumaki, N., Suda, Y., Iwasawa, C., Narita, M., Sone, T., Watanabe, M., et al. (2019). Cell-specific overexpression of COMT in dopaminergic neurons of Parkinson’s disease. Brain 142 (6), 1675–1689. doi:10.1093/brain/awz084
Ladd-Acosta, C., Pevsner, J., Sabunciyan, S., Yolken, R. H., Webster, M. J., Dinkins, T., et al. (2007). DNA methylation signatures within the human brain. Am. J. Hum. Genet. 81 (6), 1304–1315. doi:10.1086/524110
Lee, L. O., and Prescott, C. A. (2014). Association of the catechol-O-methyltransferase val158met polymorphism and anxiety-related traits: a meta-analysis. Psychiatr. Genet. 24 (2), 52–69. doi:10.1097/YPG.0000000000000018
Lowe, R., Gemma, C., Beyan, H., Hawa, M. I., Bazeos, A., Leslie, R. D., et al. (2013). Buccal are likely to be a more informative surrogate tissue than blood for epigenome-wide association studies. Epigenetics 8 (4), 445–454. doi:10.4161/epi.24362
Lyon, K. A., Juhasz, G., Brown, L. J. E., and Elliott, R. (2020). Big Five personality facets explaining variance in anxiety and depressive symptoms in a community sample. J. Affect. Disord. 274, 515–521. doi:10.1016/j.jad.2020.05.047
McCrae, R. R., and Terracciano, A.Personality Profiles of Cultures Project (2005). Universal features of personality traits from the observer's perspective: data from 50 cultures. J. Personality Soc. Psychol. 88 (3), 547–561. doi:10.1037/0022-3514.88.3.547
McGowan, P. O., Sasaki, A., D’Alessio, A. C., Dymov, S., Labonté, B., Szyf, M., et al. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 12 (3), 342–348. doi:10.1038/nn.2270
Mitrović, D., Mihić, L., Sadiković, S., and Smederevac, S. (2023). Common genetic and environmental bases of the mental disorders and personality traits: special focus on the hierarchical model of psychopathology and NEO-PI-R facets. J. Personality 92, 1143–1157. doi:10.1111/jopy.12878
Moore, A. A., Sawyers, C., Adkins, D. E., and Docherty, A. R. (2018). Opportunities for an enhanced integration of neuroscience and genomics. Brain Imaging Behav. 12 (4), 1211–1219. doi:10.1007/s11682-017-9780-1
Moore, M., Schermer, J. A., Paunonen, S. V., and Vernon, P. A. (2010). Genetic and environmental influences on verbal and nonverbal measures of the Big Five. Personality Individ. Differ. 48 (8), 884–888. doi:10.1016/j.paid.2010.02.015
Munafò, M. R., and Flint, J. (2011). Dissecting the genetic architecture of human personality. Trends Cognitive Sci. 15 (9), 395–400. doi:10.1016/j.tics.2011.07.007
Na, K. S., Won, E., Kang, J., Kim, A., Choi, S., Tae, W. S., et al. (2018). Differential effect of COMT gene methylation on the prefrontal connectivity in subjects with depression versus healthy subjects. Neuropharmacology 137, 59–70. doi:10.1016/j.neuropharm.2018.04.030
Nikolašević, Ž., Dinić, B. M., Smederevac, S., Sadiković, S., Milovanović, I., Ignjatović, V. B., et al. (2021). Common genetic basis of the five factor model facets and intelligence: a twin study. Personality Individ. Differ. 175, 110682. doi:10.1016/j.paid.2021.110682
Nohesara, S., Ghadirivasfi, M., Mostafavi, S., Eskandari, M. R., Ahmadkhaniha, H., Thiagalingam, S., et al. (2011). DNA hypomethylation of MB-COMT promoter in the DNA derived from saliva in schizophrenia and bipolar disorder. J. Psychiatric Res. 45 (11), 1432–1438. doi:10.1016/j.jpsychires.2011.06.013
Paunonen, S. V., and Ashton, M. C. (2001). Big Five factors and facets and the prediction of behavior. J. Personality Soc. Psychol. 81 (3), 524–539. doi:10.1037//0022-3514.81.3.524
Peciña, M., Mickey, B. J., Love, T., Wang, H., Langenecker, S. A., Hodgkinson, C., et al. (2013). DRD2 polymorphisms modulate reward and emotion processing, dopamine neurotransmission and openness to experience. Cortex 49 (3), 877–890. doi:10.1016/j.cortex.2012.01.010
Plotly Technologies Inc (2015). Collaborative data science. Montréal, QC: Plotly Technologies Inc. Available at: https://plot.ly.
Ren, Z., Yang, W., and Qiu, J. (2019). Neural and genetic mechanisms of creative potential. Curr. Opin. Behav. Sci. 27, 40–46. doi:10.1016/j.cobeha.2018.09.003
Reuter, M., Kuepper, Y., and Hennig, J. (2007). Association between a polymorphism in the promoter region of the TPH2 gene and the personality trait of harm avoidance. Int. J. Neuropsychopharmacol. 10 (3), 401–404. doi:10.1017/S1461145706007073
Reuter, M., Schmitz, A., Corr, P. J., and Hennig, J. (2006). Molecular genetics support Gray's personality theory: the interaction of COMT and DRD2 polymorphisms predicts the behavioural approach system. Int. J. Neuropsychopharmacol. 9, 155–166. doi:10.1017/S1461145705005419
Sadiković, S. (2023). Bihevioralnogenetičke osnove petofaktorskog modela ličnosti. [Behavioral genetic basis of the Five-factor Model of Personality]. Univ. Novi Sad, Fac. Philosophy. Available at: https://nardus.mpn.gov.rs/bitstream/handle/123456789/21533/Disertacija_13886.pdf?sequence=1.
Sanz, J., García-Vera, M. P., and Magán, I. (2010). Anger and hostility from the perspective of the Big Five personality model. Scand. J. Psychol. 51 (3), 262–270. doi:10.1111/j.1467-9450.2009.00771.x
Smederevac, S., Delgado-Cruzata, L., Mitrović, D., Dinić, B. M., Bravo, T. A. T., Delgado, M., et al. (2023b). Differences in MB-COMT DNA methylation in monozygotic twins on phenotypic indicators of impulsivity. Front. Genet. 13, 1067276. doi:10.3389/fgene.2022.1067276
Smederevac, S., Mitrović, D., Sadiković, S., Milovanović, I., Branovački, B., Dinić, B. M., et al. (2019). Serbian twin Registry. Twin Res. Hum. Genet. 22 (6), 660–666. doi:10.1017/thg.2019.114
Smederevac, S., Mitrović, D., Sadiković, S., Riemann, R., Bratko, D., Prinz, M., et al. (2020). Hereditary and environmental factors of the Five-Factor Model traits: a cross-cultural study. Personality Individ. Differ. 162, 109995. doi:10.1016/j.paid.2020.109995
Smederevac, S., Sadiković, S., Čolović, P., Vučinić, N., Milutinović, A., Riemann, R., et al. (2023a). Quantitative behavioral genetic and molecular genetic foundations of the approach and avoidance strategies. Curr. Psychol. 42 (17), 14268–14282. doi:10.1007/s12144-022-02724-9
Smillie, L. D., Cooper, A. J., Proitsi, P., Powell, J. F., and Pickering, A. D. (2010). Variation in DRD2 dopamine gene predicts Extraverted personality. Neurosci. Lett. 468 (3), 234–237. doi:10.1016/j.neulet.2009.10.095
Smith, A. K., Kilaru, V., Kocak, M., Almli, L. M., Mercer, K. B., Ressler, K. J., et al. (2014). Methylation quantitative trait loci (meQTLs) are consistently detected across ancestry, developmental stage, and tissue type. BMC Genomics 15 (1), 145. doi:10.1186/1471-2164-15-145
Soto, C. J. (2019). How replicable are links between personality traits and consequential life outcomes? The life outcomes of personality replication Project. Psychol. Sci. 30 (5), 711–727. doi:10.1177/0956797619831612
Soto, C. J., and John, O. P. (2017). The next Big Five Inventory (BFI-2): developing and assessing a hierarchical model with 15 facets to enhance bandwidth, fidelity, and predictive power. J. Personality Soc. Psychol. 113 (1), 117–143. doi:10.1037/pspp0000096
Stein, M. B., Fallin, M. D., Schork, N. J., and Gelernter, J. (2005). COMT polymorphisms and anxiety-related personality traits. Neuropsychopharmacol 30 (11), 2092–2102. doi:10.1038/sj.npp.1300787
Terracciano, A., Tanaka, T., Sutin, A. R., Deiana, B., Balaci, L., Sanna, S., et al. (2010). BDNF Val66Met is associated with introversion and interacts with 5-HTTLPR to influence neuroticism. Neuropsychopharmacol 35 (5), 1083–1089. doi:10.1038/npp.2009.213
Thomas, N., Gurvich, C., and Kulkarni, J. (2019). Borderline personality disorder, trauma, and the hypothalamus-pituitary-adrenal axis. NDT 15, 2601–2612. doi:10.2147/NDT.S198804
Tsuchimine, S., Yasui-Furukori, N., Sasaki, K., Kaneda, A., Sugawara, N., Yoshida, S., et al. (2012). Association between the dopamine D2 receptor (DRD2) polymorphism and the personality traits of healthy Japanese participants. Prog. Neuro-Psychopharmacology Biol. Psychiatry 38 (2), 190–193. doi:10.1016/j.pnpbp.2012.03.008
Tunbridge, E. M., Narajos, M., Harrison, C. H., Beresford, C., Cipriani, A., and Harrison, P. J. (2019). Which dopamine polymorphisms are functional? Systematic review and meta-analysis of COMT, DAT, DBH, DDC, DRD1–5, MAOA, MAOB, TH, VMAT1, and VMAT2. Biol. Psychiatry 86 (8), 608–620. doi:10.1016/j.biopsych.2019.05.014
Turkheimer, E. (2016). Weak Genetic Explanation 20 Years Later: Reply to Plomin et al. Perspect. Psychol. Sci. 11 (1), 24–28. doi:10.1177/1745691615617442
Valeeva, E. V., Kashevarov, G. S., Kasimova, R. R., Ahmetov, I. I., and Kravtsova, O. A. (2020). Association of the Val158Met polymorphism of the COMT gene with measures of psychophysiological status in athletes. Neurosci. Behav. Physi 50 (4), 485–492. doi:10.1007/s11055-020-00924-z
van der Knaap, L. J., Schaefer, J. M., Franken, I. H. A., Verhulst, F. C., van Oort, F. V. A., and Riese, H. (2014). Catechol-O-methyltransferase gene methylation and substance use in adolescents: the TRAILS study. Genes., Brain Behav. 13 (7), 618–625. doi:10.1111/gbb.12147
Virtanen, P., Gommers, R., Oliphant, T. E., Haberland, M., Reddy, T., Cournapeau, D., et al. (2020). SciPy 1.0: fundamental algorithms for scientific computing in Python. Nat. Methods 17 (3), 261–272. doi:10.1038/s41592-019-0686-2
Wagner, J., Orth, U., Bleidorn, W., Hopwood, C. J., and Kandler, C. (2020). Toward an integrative model of sources of personality stability and change. Curr. Dir. Psychol. Sci. 29 (5), 438–444. doi:10.1177/0963721420924751
White, M. J., Lawford, B. R., Morris, C. P., and Young, R. M. D. (2009). Interaction between DRD2 C957T polymorphism and an acute psychosocial stressor on reward-related behavioral impulsivity. Behav. Genet. 39 (3), 285–295. doi:10.1007/s10519-008-9255-7
Wiegand, A., Blickle, A., Brückmann, C., Weller, S., Nieratschker, V., and Plewnia, C. (2021). Dynamic DNA methylation changes in the COMT gene promoter region in response to mental stress and its modulation by transcranial direct current stimulation. Biomolecules 11 (11), 1726. doi:10.3390/biom11111726
Keywords: COMT, DNA methylation (DNAm), NEO-PI-R, monozygotic twins, epigenetic modifications
Citation: Mitrović D, Smederevac S, Delgado-Cruzata L, Sadiković S, Pajić D, Prinz M, Budimlija Z, Oljača M, Kušić-Tišma J, Vučinić N and Milutinović A (2024) Personality and COMT gene: molecular-genetic and epigenetic associations with NEO-PI-R personality domains and facets in monozygotic twins. Front. Genet. 15:1455872. doi: 10.3389/fgene.2024.1455872
Received: 27 June 2024; Accepted: 11 September 2024;
Published: 25 September 2024.
Edited by:
Joel Carter Eissenberg, Saint Louis University, United StatesReviewed by:
Gabe Haller, Washington University in St. Louis, United StatesCopyright © 2024 Mitrović, Smederevac, Delgado-Cruzata, Sadiković, Pajić, Prinz, Budimlija, Oljača, Kušić-Tišma, Vučinić and Milutinović. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Selka Sadiković, c2Vsa2Euc2FkaWtvdmljQGZmLnVucy5hYy5ycw==
†ORCID: Dušanka Mitrović, orcid.org/0000-0001-8532-6795; Snežana Smederevac, orcid.org/0000-0002-3780-0576; Lissette Delgado-Cruzata, orcid.org/0000-0002-2884-0234; Selka Sadiković, orcid.org/0000-0003-0874-2511; Dejan Pajić, orcid.org/0000-0002-6417-5382; Mechthild Prinz, orcid.org/0000-0002-8169-6622; Zoran Budimlija, orcid.org/0000-0003-4086-8536; Milan Oljača, orcid.org/0000-0003-4628-4558; Jelena Kušić-Tišma, orcid.org/0000-0002-4167-404X; Nataša Vučinić, orcid.org/0000-0001-6847-2862; Aleksandra Milutinović, orcid.org/0000-0002-7001-3937
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.