 Cecilia Da Silva1†
Cecilia Da Silva1† Natalia Mannise2,3†
Natalia Mannise2,3† Rosina Seguí4
Rosina Seguí4 Andrés Iriarte3
Andrés Iriarte3 Nadia Bou2J. Mauricio Bonifacino5,6Ary Mailhos7Lucía Anza7Santiago Chitaro7Florencia Ocampo7Rosario Gándaras7Florencia Arezo7
Nadia Bou2J. Mauricio Bonifacino5,6Ary Mailhos7Lucía Anza7Santiago Chitaro7Florencia Ocampo7Rosario Gándaras7Florencia Arezo7 Leandro Capurro7Marcelo Iturburu7Nicolás Nieto7Hernán Juan2,3Joaquín Garrido8
Leandro Capurro7Marcelo Iturburu7Nicolás Nieto7Hernán Juan2,3Joaquín Garrido8 Raúl Platero9Julián Gago10Felipe Lezama5Martín Do Carmo11Bol-Uy12
Raúl Platero9Julián Gago10Felipe Lezama5Martín Do Carmo11Bol-Uy12 Mariana Cosse2*
Mariana Cosse2*- 1Centro Universitario Regional Noreste, Universidad de La República, Tacuarembó, Uruguay
- 2Departamento de Biodiversidad y Genética, Instituto de Investigaciones Biológicas Clemente Estable, Montevideo, Uruguay
- 3Laboratorio de Biología Computacional, Departamento de Desarrollo Biotecnológico, Instituto de Higiene, Facultad de Medicina, Universidad de La República, Montevideo, Uruguay
- 4División Información Ambiental, DINACEA, Ministerio de Ambiente, Montevideo, Uruguay
- 5Facultad de Agronomía, Universidad de La República, Montevideo, Uruguay
- 6Facultad de Ciencias, Universidad de La República, Montevideo, Uruguay
- 7DNA Barcode Training Course and Grasses Barcode Pilot Project, PEDECIBA Universidad de La República, Montevideo, Uruguay
- 8Ministerio de Ganadería, Agricultura y Pesca, Montevideo, Uruguay
- 9Departamento de Bioquímica y Genómica Microbianas, Instituto de Investigaciones Biológicas Clemente Estable, Montevideo, Uruguay
- 10Museo y Jardín Botánico “Prof. Atilio Lombardo”, Intendencia de Montevideo, Montevideo, Uruguay
- 11Centro Universitario Regional Este, Universidad de La República, Rocha, Uruguay
- 12Uruguayan Barcode of Life initiative, Montevideo, Uruguay
1 Introduction
The loss of biodiversity has accelerated during the Anthropocene era, matching rates seen in previous mass extinctions (Rockström et al., 2009), resulting in unprecedented environmental changes (IPBES, 2021). In this scenario, it is crucial to make progress in effectively describing, documenting, and classifying living species. Acknowledging this urgency, the Convention on Biological Diversity (CBD) has highlighted the essential role of taxonomy in achieving these goals (CBD, 2021). However, human resources with expertise in taxonomy are increasingly scarce and often face intricate challenges like identifying cryptic species and larval stages (Thomsen and Willerslew, 2015). These challenges have sparked a need for alternative approaches in biodiversity description and monitoring. In response, Barcodes of Life (BoL) have emerged to identify organisms through genomic approaches utilizing specific DNA regions and standardized protocols (Hebert et al., 2003). The potential of this strategy has been demonstrated on a global scale as a powerful tool, with widespread applications including natural resource conservation, safeguarding endangered species, agricultural pest management, disease vector identification, water quality surveillance, verification of health and food products, forensic entomology, as well as medicinal plant identification, among others (Consortium for the Barcode of Life, 2007).
DNA techniques require the meticulous selection of suitable barcoding regions, involving the identification of standardized loci that can be systematically and reliably sequenced across extensive and diverse taxa. This enables the generation of easily comparable data that facilitates species distinction (Hollingsworth et al., 2011). In animals, cytochrome oxidase I (COI) is the standard DNA barcode, showing notable success in precisely identifying species (Hebert et al., 2003). However, identifying an equivalent DNA barcode for plants remains challenging due to the absence of a single, appropriate locus for species delimitation (Hebert et al., 2016). Plant DNA barcoding has been controversial due to the presence of hybridization, introgression processes, and polyploidy (Hebert et al., 2016). The slow evolutionary rate in plant mitochondrial genomes rules out COI as a universal plant barcode (Hollingsworth et al., 2011). Instead, several chloroplast and nuclear molecular markers like rbcL, matK, trnH-psbA, trnL (UAA), and ITS have been explored as viables alternatives (Taberlet et al., 2007; Hollingsworth et al., 2011; Marcial-Quino et al., 2015). Given the inadequate species discrimination in certain plant groups with a single molecular marker, a combination of markers has been proposed (Hollingsworth et al., 2011).
Another central aspect for taxa recognition using molecular markers is the need for a comprehensive reference database of barcode DNA sequences encompassing a wide range of species (Hollingsworth et al., 2011; Tnah et al., 2019). The quantity of sequences accessible for a specific locus impacts its discriminatory power (Hollingsworth et al., 2011). Establishing a robust public sequence repository is essential for assessing multiple potential loci for barcoding and their combination (Hollingsworth et al., 2011; Kolter and Gemeinholzer, 2021).
Based on these principles, the Consortium for the Barcode of Life (CBoL; www.barcodeoflife.org) was established and is collaboratively building the Barcode of Life Data Systems (BOLD, Ratnasingham and Hebert, 2007; www.boldsystems.org). BOLD is a global open access repository of DNA barcodes, but unlike other public genomic repositories such as GenBank (NCBI), EMBL, and DDBJ, it is specifically tailored for biodiversity and species identification through DNA barcoding (Borisenko et al., 2009), linking morphological and distributional data with DNA sequences. It also supports broader taxonomic initiatives such as the Global Taxonomic Initiative (GTI) for the Convention on Biological Diversity (CBD) and the Global Biodiversity Information Facility (GBIF) (Costa and Antunes, 2012).
Among the vast array of over 375,000 documented vascular plant species worldwide (Hong et al., 2022), only 115,768 are registered within BOLD’s database with a total of 380,813 public records (BOLD, 22/04/2024). This underscores the pressing necessity for substantial contributions from diverse researchers, institutions, and national networks to enhance the comprehensiveness of this database.
In recent years, Uruguay has experienced a decline in biodiversity, a global phenomenon impacting ecosystems worldwide. The dramatic shifts in land use since the 1990s, particularly marked by an increase in the area of tree plantations (the highest in South America) and agriculture (specifically soybean and rice), underscore the urgent need for effective advances in documenting and categorizing the country’s biodiversity (Paruelo et al., 2006; Baeza and Paruelo, 2020). Currently, vascular plants in Uruguay remains largely unexplored in terms of barcode sequence availability. The BOLD platform holds 1,131 public records for Uruguayan organisms, representing 323 species, with only 24 vascular plant species documented (searched on 22/3/2024), which accounts for less than 1% of native species reported for the country (Bonifacino et al., 2019). As a way of filling this void of DNA barcode sequences among the Uruguayan biodiversity, a major initiative is underway to establish a DNA barcode national network. During October 2018 the “Barcode of Life Training Course: example from native flora” was held, under the call for the program “Filling the capacity gap for the application of DNA technologies in taxonomy -GTI trainings/workshops driven by the trained trainers.” This project had two main objectives: i) to train undergraduate and postgraduate students, as well as technicians from public and private institutions; to conduct workshops with vascular plant specialists to list key species of the Uruguayan flora; and ii) to coordinate with key decision makers regarding national biodiversity and scientific collections, with the intention of establishing and maintaining a national DNA barcode network in the near future.
Our objective was to produce a first batch of DNA barcode sequences for the vascular plants of Uruguay, focusing on species not previously documented in BOLD that hold cultural significance in Uruguay due to their agricultural, medicinal and horticultural uses, employing four commonly used molecular markers for plants (rbcL, trnH-psbA, trnL (UAA), and ITS2).
2 Materials and methods
2.1 Species selection
In order to select the plant species to be analyzed, two meetings were organized with experts in grassland ecosystems, livestock production in natural grasslands, and native forests, as well as invited taxonomists. The aim of these meetings was to compile an initial list of key species representing Uruguayan plant biodiversity, based on their productive value, symbolic importance or conservation status.
2.2 Sample collection and storage
A total of 51 samples, representing 50 vascular plant species were collected in different regions of Uruguay: grasslands of Rivera and Río Negro departments; the greenhouse of the Instituto Nacional de Semillas (INASE) in Canelones department; the gardens of the Museo Jardín Botánico Profesor Atilio Lombardo (MJBAL-IMM), the Facultad de Agronomía of the Universidad de la República (FAgro-Udelar), and the Instituto de Investigaciones Biológicas Clemente Estable (IIBCE-MEC) in Montevideo department (Table 1).
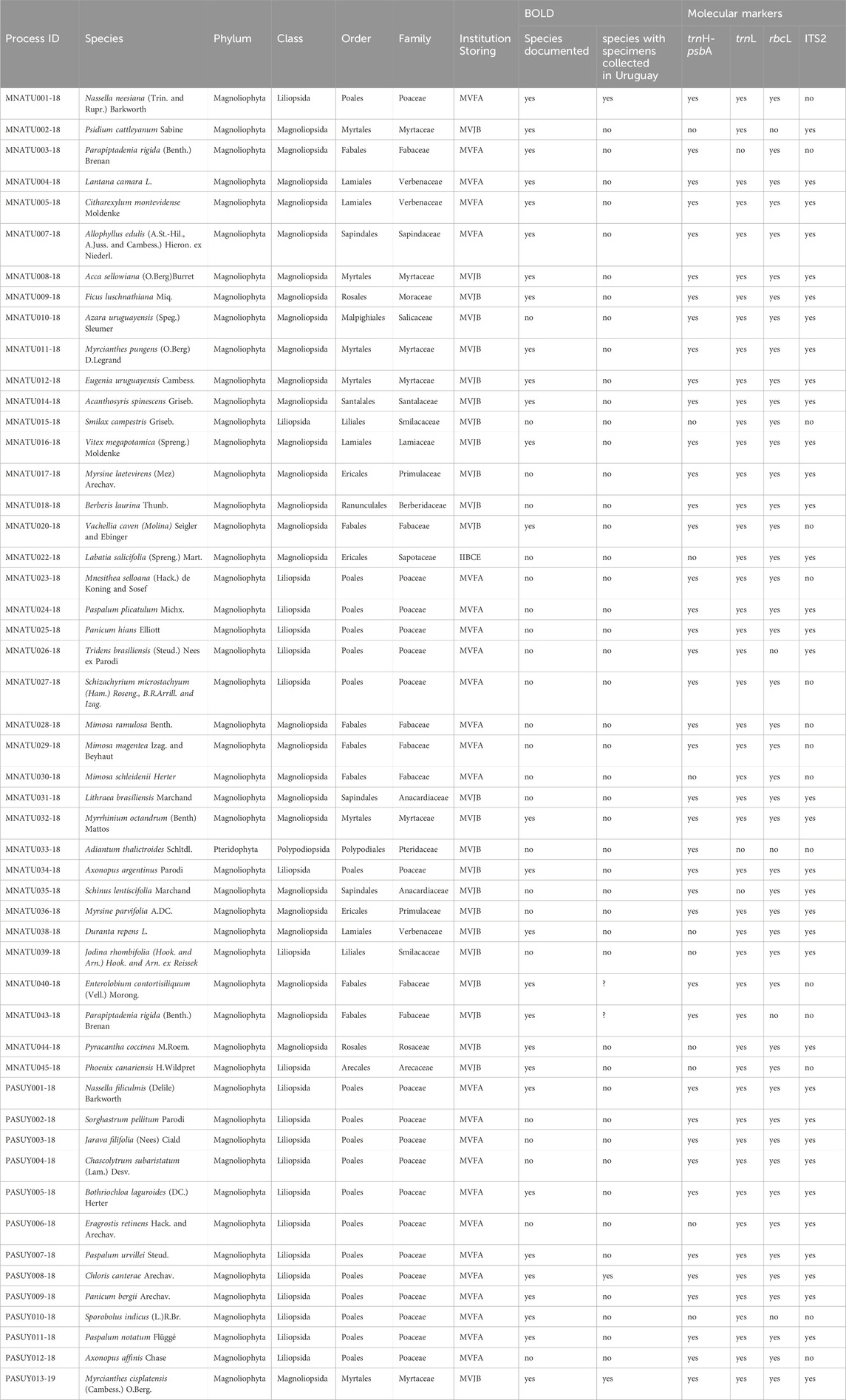
Table 1. Specimens reported in this study. For each sample, the following information is listed: process identification in BOLD, species name and taxonomy, institution where the voucher is deposited (Universidad de la República, Facultad de Agronomía, Bernardo Rosengurtt Herbarium = MVFA, Museo y Jardin Botánico Prof. Atilio Lombardo = MVJB, Instituto de Investigaciónes Biológicas Clemente Estable = IIBCE), whether the species has been previously documented in BOLD, if the documented specimens in BOLD for the species were collected in Uruguay, and whether sequencing was successful for each of the four markers used in this study. In the cases where the origin of the samples in BOLD is missing a question mark is added.
Vascular plants specimens sampled belonged to 13 orders and 16 families. The three orders best represented in the sample were Poales (37.25%), Fabales (13.73%) and Myrtales (11.76%). This includes 23 species not previously documented in BOLD and 23 species documented in BOLD, but without specimens collected in Uruguay (Table 1).
Standardized procedures were followed to obtain herbarium samples (Bridson and Forman, 1992; Seshagirirao et al., 2016) and leaf or stem tissue for DNA extraction (Wilkie et al., 2013). A high-quality image of the vouchers was obtained using an EPSON Expression 10000XL (Figure 1). Tissue samples for DNA isolation were stored in silica gel until dry. Specimens were deposited in MVJB, MVFA and IIBCE-MEC herbaria (Table 1). Herbarium acronyms according to Thiers (updated continuously).

Figure 1. Herbarium voucher. A Mimosa ramulosa Benth. specimen, submitted to BOLD with the Process ID MNATU028-18 and deposited in MVFA herbarium.
2.3 Laboratory procedures
DNA extractions were conducted from dried unexpanded young leaves, following Fazekas et al. (2012) protocol. FastPrep®- 24 (MP Biomedicals) and lysing matrix beads were used for sample lysis. Afterwards, four DNA barcode fragments were amplified, three from chloroplast: rbcL, trnH-psbA and trnL(UUA) (Fay et al., 1997; Taberlet et al., 2007; Guo et al., 2011) and one nuclear (ITS2) (Chen et al., 2010) (Supplementary Material). Each PCR reaction contained 1X Buffer (Invitrogen), 1.5 mM MgCl (Invitrogen), 0.04 U Taq polymerase (Invitrogen), 0.05 mM each dNTP, 0.2 mg/mL BSA and 0.5 µM forward and reverse primers in a final volume of 20 µL. The thermal profile consisted of an initial denaturalization step of 94°C for 3 min, followed by 35 cycles of 94°C for 30 s, annealing for 30 s, (annealing temperatures for each genetic marker are provided in the Supplementary Material), and 72°C for 1 min, a final extension step of 72°C for 15 min were inserted. PCR amplifications were conducted independently for each molecular marker and negative controls were included on each PCR and extraction reactions. Moreover, PCRs and DNA extractions were carried out in different rooms to avoid contaminations.
Amplification success was confirmed through 1% agarose electrophoresis using GoodViewTM nucleic acid stain (SBS Genetech Co., Ltd, Beijing). PCR products were purified using an enzymatic method with 0.8 U/μL of Exonuclease I and 0.2 U/μL of Thermosensitive Alkaline Phosphatase (FastAP) (ThermoFisher ScientificTM). The conditions for purifications were 37°C for 1 h and 30 min followed by 5 min at 75°C. The amount of purified PCR products was measured by NanodropTMND-1000 UV-vis Spectrophotometer (Nano-Drop Techonologies, Inc., Wilmington, DE, United States). Forward and reverse sequencing was conducted on ABI3500 (Applied Biosystems) in Institute Pasteur de Montevideo and Applied Biosystems 3730XL in Macrogen Inc.
2.4 Data treatment
Every DNA barcode sequence was analyzed by eye and alignment was built by MUSCLE using Mega5 (Tamura et al., 2011). Sequences obtained for each locus and specimen were compared with public sequences through BLAST against GenBank database (Altschul et al., 1990) to confirm sequence identity. The data generated for each sample was submitted to Barcode of Life Data System (BOLD, http://www.boldsystems.org).
3 Conclusion
The sample collection and laboratory procedures employed in this study proved to be accurate for the plant DNA barcoding protocol, resulting in 171 sequences from four loci for 50 vascular plant species. The selection of appropriate markers for taxa identification involves evaluating primer universality, sequence quality, and discriminatory power (Hollingsworth et al., 2011). Our study assessed the first two criteria, indicating that, in agreement with previous findings for land plants (Hollingsworth et al., 2011), trnL (UAA) and rbcL are the most suitable loci for the plant groups examined (Table 1). Plastid loci have been widely used in phylogenetic analyses, presenting traits similar to those seen in the mitochondrial genome of animals (Kress et al., 2005; Chase et al., 2007). Notably, the ITS2 amplification rates observed in our samples were comparatively low, diverging from previous studies (Chase et al., 2007; Chen et al., 2010). Finally, setting a comprehensive public database of sequences from several candidate loci for a wide variety of species is crucial not only for the evaluation of barcode candidates and their combinations (Hollingsworth et al., 2011), but also for an accurate species differentiation (Hollingsworth et al., 2011; Tnah et al., 2019). Our contribution aims to lay the groundwork for a robust database to further research in biodiversity and facilitate taxonomic identification.
This work marks the first step of the Uruguayan Barcode of Life Initiative (BoL-UY), providing essential insights into vascular plants with data from three chloroplast loci and one nuclear locus. Embracing the DNA barcoding protocol, voucher images of specimens remain accessible and linked to sequences for each locus in BOLD. The data generated holds promise for future research venues aimed at identifying the optimal loci and their most effective combinations for vascular plants DNA barcoding.
Data availability statement
The data presented in the study are deposited in the Barcode of Life Data System (www.boldsystems.org), accession number are listed in Table 1 as ProcessID.
Author contributions
CD: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing–original draft, Writing–review and editing. NM: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing–original draft, Writing–review and editing. RS: Data curation, Writing–review and editing, Conceptualization, Funding acquisition. AI: Data curation, Writing–review and editing, Formal Analysis. NB: Data curation, Writing–review and editing, Methodology. JB: Data curation, Methodology, Writing–review and editing, Formal Analysis, Investigation, Validation. AM: Formal Analysis, Methodology, Writing–review and editing. LA: Formal Analysis, Methodology, Writing–review and editing. SC: Formal Analysis, Methodology, Writing–review and editing. FO: Formal Analysis, Methodology, Writing–review and editing. RG: Formal Analysis, Methodology, Writing–review and editing. FA: Formal Analysis, Methodology, Writing–review and editing. LC: Formal Analysis, Methodology, Writing–review and editing. MI: Formal Analysis, Methodology, Writing–review and editing. NN: Formal Analysis, Methodology, Writing–review and editing. HJ: Formal Analysis, Methodology, Writing–reviewand editing, Data curation. JoG: Data curation,Writing–review and editing. RP: Data curation, Writing–review and editing, Methodology. JuG: Data curation, Writing–review and editing. FL: Data curation, Writing–review and editing, Methodology. MD: Data curation, Writing–review and editing, Investigation. BU: Writing–review and editing, Funding acquisition, Supervision, MC: Funding acquisition, Supervision, Writing–review and editing, Conceptualization, Formal Analysis, Investigation, Methodology, Project administration, Validation.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Agencia Nacional de Investigación e Innovación, Fondo María Viña (ANII FMV_1_2021_1_166380), Programa de Desarrollo de las Ciencias Básicas (PEDECIBA), Convention on Biological Diversity, Global Taxonomy Initiative, Japan Biodiversity Fund.
Acknowledgments
Special thanks to Susana González, Pablo Tubaro, Victor Canton, Alejandro Nario, Laura Barreiro, Viviana Mezzetta, Silvana Ravía and Eduardo Manta, for their support in obtained funds and logistic. We gratefully acknowledge the support and contributions of national organizations linked to science development and biodiversity: SNCYT, DINAMA, MVOTMA, AUCI and ANII. Thanks to Junko Shimura and Adriana Radulovici from CBD. To the editor and reviewers, which improved the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1435592/full#supplementary-material
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi:10.1016/S0022-2836(05)80360-2
Baeza, S., and Paruelo, J. M. (2020). Land use/land cover change (2000–2014) in the Rio de la Plata grasslands: an analysis based on MODIS NDVI time series. Remote Sens. 12, 381. doi:10.3390/rs12030381
Bonifacino, J. M., Marchesi, E., Valtierra, V., Perez, C., Trujillo, C., Gadea, F., et al. (2019). Next generation floristics: compositae of Uruguay. Botany 2019. Tucson, USA: Annual Meeting of the Botanical Society of America in Arizona. Available at: https://2019.botanyconference.org/engine/search/index.php?func=detail&aid=733.
Borisenko, A. V., Sones, J. E., and Hebert, P. D. (2009). The front-end logistics of DNA barcoding: challenges and prospects. Mol. Ecol. Resour. 9, 27–34. doi:10.1111/j.1755-0998.2009.02629.x
CBD (2021). The convention on biological diversity. Available at: https://www.cbd.int/convention.
Chase, M. W., Cowan, R. S., Hollingsworth, P. M., van den Berg, C., Madriñán, S., Petersen, G., et al. (2007). A proposal for a standardised protocol to barcode all land plants. Taxon 56, 295–299. doi:10.1002/tax.562004
Chen, S., Yao, H., Han, J., Liu, C., Song, J., Shi, L., et al. (2010). Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PloS one 5, e8613. doi:10.1371/journal.pone.0008613
Consortium for the Barcode of Life (2007). How DNA barcoding can help science. ScienceDaily. Available at: www.sciencedaily.com/releases/2007/09/070914120856.htm.
Costa, F. O., and Antunes, P. M. (2012). “The contribution of the barcode of life initiative to the discovery and monitoring of biodiversity,” in Natural resources, sustainability and humanity. Editors A. Mendonca, A. Cunha, and R. Chakrabarti (Dordrecht: Springer), 37–68. doi:10.1007/978-94-007-1321-5_4
Fay, M. F., Swensen, S. M., and Chase, M. W. (1997). Taxonomic affinities of Medusagyne oppositifolia (medusagynaceae). Kew Bull. 52, 111–120. doi:10.2307/4117844
Fazekas, A. J., Kuzmina, M. L., Newmaster, S. G., and Hollingsworth, P. M. (2012). DNA barcoding methods for land plants. In: Kress, W.,, and Erickson, D. (eds) DNA barcodes. Methods in molecular biology, vol 858. Humana Press, Totowa, NJ. 223–252. doi:10.1007/978-1-61779-591-6_11
Guo, X., Wang, X., Su, W., Zhang, G., and Zhou, R. (2011). DNA barcodes for discriminating the medicinal plant Scutellaria baicalensis (Lamiaceae) and its adulterants. Biol. Pharm. Bull. 34, 1198–1203. doi:10.1248/bpb.34.1198
Hebert, P. D., Cywinska, A., Ball, S. L., and DeWaard, J. R. (2003). Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 270, 313–321. doi:10.1098/rspb.2002.2218
Hebert, P. D., Hollingsworth, P. M., and Hajibabaei, M. (2016). From writing to reading the encyclopedia of life. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 371, 20150321. doi:10.1098/rstb.2015.0321
Hollingsworth, P. M., Graham, S. W., and Little, D. P. (2011). Choosing and using a plant DNA barcode. PloS6, e19254. doi:10.1371/journal.pone.0019254
Hong, Q., Jian, Z., and Jingchao, Z. (2022). How many known vascular plant species are there in the world? An integration of multiple global plant databases. Biodiv. Sci. 30, 22254. doi:10.17520/biods.2022254
IPBES (2021). Launch of IPBES-IPCC Co-sponsored workshop report on biodiversity and climate change. Available at: https://ipbes.net/events/launch-ipbes-ipcc-co-sponsored-workshop-report-biodiversity-and-climate-change.
Kolter, A., and Gemeinholzer, B. (2021). Plant DNA barcoding necessitates marker-specific efforts to establish more comprehensive reference databases. Genome 64, 265–298. doi:10.1139/gen-2019-0198
Kress, W. J., Wurdack, K. J., Zimmer, E. A., Weigt, L. A., and Janzen, D. H. (2005). Use of DNA barcodes to identify flowering plants. PNAS 102, 8369–8374. doi:10.1073/pnas.0503123102
Marcial-Quino, J., Mendoza-Espinoza, J. A., and Sierra-Palacios, E. (2015). DNA barcoding: an alternative for the identification of the medicinal plants employed in Mexico. J. Plant Sci. 10, 116–127. doi:10.3923/jps.2015.116.127
Paruelo, J. M., Guerschman, J. P., Pineiro, G., Jobbagy, E. G., Verón, S. R., Baldi, G., et al. (2006). Cambios en el uso de la tierra en Argentina y Uruguay: marcos conceptuales para su análisis. Agrocienc. Urug. 10, 47–61. doi:10.31285/AGRO.10.929
Ratnasingham, S., and Hebert, P. D. (2007). Bold: the barcode of life data system. Mol. Ecol. Notes. 7, 355–364. doi:10.1111/j.1471-8286.2007.01678.x
Rockström, J., Steffen, W., Noone, K., Persson, Å., Chapin, F. S., Lambin, E. F., et al. (2009). A safe operating space for humanity. Nature 461 (7263), 472–475. doi:10.1038/461472a
Seshagirirao, K., Harikrishnanaik, L., Venumadhav, K., Nanibabu, B., Jamir, K., Ratnamma, B. K., et al. (2016). Preparation of herbarium specimen for plant identification and voucher number. Roxburghia 6, 111–119.
Taberlet, P., Coissac, E., Pompanon, F., Gielly, L., Miquel, C., Valentini, A., et al. (2007). Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. NAR 35, e14. doi:10.1093/nar/gkl938
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi:10.1093/molbev/msr121
Thiers, B. M. (2022). Index herbariorum. Available at: https://sweetgum.nybg.org/science/ih/.
Thomsen, P. F., and Willerslev, E. (2015). Environmental DNA–An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi:10.1016/j.biocon.2014.11.019
Tnah, L. H., Lee, S. L., Tan, A. L., Lee, C. T., Ng, K. K. S., Ng, C. H., et al. (2019). DNA barcode database of common herbal plants in the tropics: a resource for herbal product authentication. Food control. 95, 318–326. doi:10.1016/j.foodcont.2018.08.022
Keywords: Bold System, taxonomy, uruguayan flora, molecular markers, rbcL, trnH-psbA, trnL (UAA), ITS2
Citation: Da Silva C, Mannise N, Seguí R, Iriarte A, Bou N, Bonifacino JM, Mailhos A, Anza L, Chitaro S, Ocampo F, Gándaras R, Arezo F, Capurro L, Iturburu M, Nieto N, Juan H, Garrido J, Platero R, Gago J, Lezama F, Do Carmo M and Cosse M (2024) Exploring biodiversity of Uruguayan vascular plants through DNA barcoding. Front. Genet. 15:1435592. doi: 10.3389/fgene.2024.1435592
Received: 20 May 2024; Accepted: 16 July 2024;
Published: 21 August 2024.
Edited by:
Viviana Solis Neffa, Instituto de Botánica del Nordeste (IBONE-CONICET), ArgentinaReviewed by:
Ercilia María Sara Moreno, Instituto de Botánica del Nordeste (IBONE-CONICET), ArgentinaCaroline Turchetto, Federal University of Rio Grande do Sul, Brazil
Copyright © 2024 Da Silva, Mannise, Seguí, Iriarte, Bou, Bonifacino, Mailhos, Anza, Chitaro, Ocampo, Gándaras, Arezo, Capurro, Iturburu, Nieto, Juan, Garrido, Platero, Gago, Lezama, Do Carmo and Cosse. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Cosse, bWNvc3NlQGlpYmNlLmVkdS51eQ==
†These authors share first authorship