
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 29 June 2023
Sec. Human and Medical Genomics
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1222112
This article is part of the Research Topic CRISPR: The Game Changer in Gene and Cell Therapy View all 6 articles
 Somang Kim1,2†Jimmy B. Yuan1,2†Wendy S. Woods3Destry A. Newton1Pablo Perez-Pinera2,3,4,5,6Jun S. Song1,2,5,7,8*
Somang Kim1,2†Jimmy B. Yuan1,2†Wendy S. Woods3Destry A. Newton1Pablo Perez-Pinera2,3,4,5,6Jun S. Song1,2,5,7,8*Prime editing (PE) is a highly versatile CRISPR–Cas9 genome editing technique. The current constructs, however, have variable efficiency and may require laborious experimental optimization. This study presents statistical models for learning the salient epigenomic and sequence features of target sites modulating the editing efficiency and provides guidelines for designing optimal PEs. We found that both regional constitutive heterochromatin and local nucleosome occlusion of target sites impede editing, while position-specific G/C nucleotides in the primer-binding site (PBS) and reverse transcription (RT) template regions of PE guide RNA (pegRNA) yield high editing efficiency, especially for short PBS designs. The presence of G/C nucleotides was most critical immediately 5’ to the protospacer adjacent motif (PAM) site for all designs. The effects of different last templated nucleotides were quantified and observed to depend on the length of both PBS and RT templates. Our models found AGG to be the preferred PAM and detected a guanine nucleotide four bases downstream of the PAM to facilitate editing, suggesting a hitherto-unrecognized interaction with Cas9. A neural network interpretation method based on nonextensive statistical mechanics further revealed multi-nucleotide preferences, indicating dependency among several bases across pegRNA. Our work clarifies previous conflicting observations and uncovers context-dependent features important for optimizing PE designs.
One of the most powerful gene editing tools available today is the complex of clustered regularly interspaced short palindromic repeats (CRISPRs) with CRISPR-associated protein 9 (Cas9) (Jinek et al., 2012; Cong et al., 2013; Jinek et al., 2013; Mali et al., 2013). In nature, CRISPR–Cas9 is found in bacteria as a natural defense mechanism of excising foreign DNA in the CRISPR DNA regions. In a rapid succession of development, researchers have repurposed and further engineered this process as a laboratory tool for editing the genome of a variety of cell types across species, including diseased cells in humans (Doudna and Charpentier, 2014; Hsu et al., 2014; Wright et al., 2016). Several different techniques have been developed to date to improve the CRISPR–Cas9 system to be more programmable and suitable for in vivo editing, while reducing off-targets and unintended mutations (Komor et al., 2016; Gaudelli et al., 2017; Kim et al., 2017; Komor et al., 2017; Gapinske et al., 2018; Winter et al., 2019). In particular, prime editors (PEs) are the latest state-of-the-art tool, which are highly versatile (Anzalone et al., 2019). The biomolecular architecture of PEs consists of a partially inactivated Cas9 fused to a reverse transcriptase and a customizable prime editing guide RNA (pegRNA), which contains a scaffolding sequence bound by Cas9. A PE targets the desired edit locus via the combination of two main processes: complementary pairing of a ∼20 nucleotide (nt) guide sequence at the 5′ end of pegRNA to the non-edited DNA strand and recognition of a short protospacer adjacent motif (PAM) on the edited strand by Cas9. The modified Cas9 includes a domain fused to a reverse transcriptase (RT) and a nickase domain that nicks only the edited DNA strand, three bases upstream of the PAM sequence. The 3′ end of pegRNA hybridizes to a region in the edited strand and acts as a primer for the ensuing reverse transcription. Starting at the nick site, the RT reverse-transcribes part of the pegRNA 3′ extension that is immediately upstream of the primer-binding site (PBS) region. This region is denoted as the RT template and contains the complementary RNA template for the desired DNA edit sequence. After nicking the edited strand and reverse-transcribing the templated DNA, two single-stranded DNA flaps are formed: the 3′ flap, created via reverse transcription containing the edit of interest, and the 5′ flap, created via the Cas9 nick, which does not contain the edit. Successful edits are completed upon the action of 5′-flap-specific endonucleases, such as FEN1 (Liu et al., 2004).
There are two main advantages of PEs over other genome editors. First, compared to the CRISPR–Cas9 endonuclease construct, PEs are less likely to produce unintended nearby insertions and deletions, as they avoid DNA double-strand breaks (Chapman et al., 2012; Anzalone et al., 2019). Second, unlike base editors that can currently introduce only C > T or A > G conversions, PEs can produce all 12 base changes, as well as small insertions and deletions (Gaudelli et al., 2017; Anzalone et al., 2019). However, a notable difficulty in using PEs stems from the complication that the flexible pegRNA design has several adjustable parameters, yielding varying degrees of editing efficiency, and from the fact that there is currently a dearth of reliable computational models capable of a priori predicting these differences in efficiencies.
Anzalone et al. originally introduced three variants of PE, denoted as PE1, PE2, and PE3, where PE1 used a reverse transcriptase derived from Moloney murine leukemia virus (MMLV RT), PE2 used the MMLV RT from PE1 with an additional 5 point mutations, and PE3 used the Cas9 nickase in PE2 to perform non-concurrent nicks on both strands, and thus, perform non-concurrent edits on both strands to avoid creating double-stranded breaks (Anzalone et al., 2019). To date, numerous engineering approaches have been applied to improve the editing efficiency of PEs, such as mutating the PE components or co-expressing additional components together with the PE, resulting in multiple PE variants (Spencer and Zhang, 2017; Liu P. et al., 2021; Chen et al., 2021; Park et al., 2021; Kweon et al., 2022). There have also been additional improvements in PE design by either altering the pegRNA outside the regions directly hybridizing with the target site (Liu Y. et al., 2021; Nelson et al., 2022; Petri et al., 2022) or by impeding the mismatch repair mechanism (Kweon et al., 2022).
Despite the rapid experimental progress, accurately predicting the editing efficiency of PEs at previously untested genomic loci remains a major challenge. The prediction problem is complicated by the fact that PE efficiency may depend on numerous factors, such as sequence content and chromatin accessibility of the targeted locus, lengths of the PBS and RT regions on pegRNA, and the intended editing type. Notably, Kim et al. recently embarked on the difficult task of probing PE2 efficiency in human cells by using lentiviral plasmid libraries to screen the efficiency of ∼48,000 pegRNA designs on ∼2,000 integrated target sequences (Kim et al., 2021); analyzing the resulting data using a deep learning model, they predicted the measured PE2 efficiency based on an extensive set of variables, including the target sequence, GC counts, melting temperature, minimum self-folding energy, and the DeepSpCas9 score of Cas9 nuclease activity (Kim et al., 2019). Although the authors highlighted the most relevant features using the Tree SHAP approach, some predictive variables, such as GC content and melting temperature, were highly correlated and might have confounded the interpretation. In addition, their analysis focused on common features shared across varying lengths of PBS and RT templates, rather than features distinguishing different designs. Furthermore, the experimental method measuring the editing efficiencies mostly assayed exogenously integrated sequences rather than endogenous sequences. The specific locations of the integration sites were thus not known; as a result, data and models used by Kim et al.’s did not incorporate epigenetic information in predicting PE2 efficiency.
This study presents statistical models that systematically examine the effects of both epigenomic and sequence-dependent features on PE efficiencies by analyzing data from both existing publications and additional in-house experiments (Supplementary Method S1). We consider only PE2 editors, which have by far the most number of publicly available data. We describe preferred target features for each pair of PBS length (PBSL) and RT template length (RTTL), revealing specific nucleotide effects that depend on these lengths and resolving in the process discrepancies in the literature regarding the role of certain nucleotides. Our models, utilizing only a small number of parameters compared to those used in previous approaches (Kim et al., 2021), capture both marginal and joint effects of nucleotides across pegRNA and provide practical guidelines for choosing optimal PE2 designs.
The cell line HEK293T was obtained from the American Type Culture Collection (ATCC) and was maintained in DMEM supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin at 37°C with 5% CO2. HEK293T cells were transfected in 24-well plates with Lipofectamine 2000 (Invitrogen) following the manufacturer’s instructions. The amount of DNA used for lipofection was 1 μg per well. Transfection efficiency was always higher than 90% as determined by fluorescent microscopy or flow cytometry, following the delivery of a control GFP expression plasmid.
The plasmids encoding PE2 (Plasmid#132775), as well as the plasmid expressing the pegRNA (Plasmid #132777), were obtained from Addgene. PegRNAs were cloned into the plasmid backbone with paired oligonucleotides (IDT, Supplementary Table S1), as previously described (Anzalone et al., 2019). Briefly, the oligonucleotides used to create the guide sequences were hybridized, phosphorylated, and cloned into the sgRNA vector using BsaI-HFv2 (NEB), in a reaction that included T4-PNK (NEB) and T4 DNA Ligase (NEB).
DNA amplicons for NGS were generated by PCR using KAPA HiFi HotStart (Roche), according to the manufacturer’s instructions, using primers with overhangs compatible with Nextera XT indexing (IDT, Supplementary Table S2). Following validation of the quality of PCR products by gel electrophoresis, the PCR products were isolated using AMPure XP PCR purification beads (Beckman Coulter). Indexed amplicons were then generated using a Nextera XT DNA Library Prep Kit (Illumina), quantitated, and pooled. Libraries were sequenced with a MiSeq Nano flow cell for 251 cycles from each end of the fragment using a MiSeq Reagent Kit v2 (500 cycles). FASTQ files were created and demultiplexed using bcl2fastq v2.17.1.14 Conversion Software (Illumina). Deep sequencing was performed by the Roy J. Carver Biotechnology Center at the University of Illinois at Urbana-Champaign.
The in-house experiments of prime editing for 25 target sites (Supplementary Tables S1, S2) were conducted in three biological replicates on different days by the same person.
The editing efficiency data for PE2 at endogenously edited sites in Anzalone et al. (2019) were obtained from the Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra/) under the accession code PRJNA565979. In-house experimental sequencing data for 25 target sites can be found in the SRA under the accession code PRJNA949853. Raw sequences from Anzalone et al. and in-house experiments were aligned to the human genome (GRCh38) using the Bowtie package (version 2.4.1), with a maximum fragment length of 500 base pairs (bp) for paired-end sequences (Langmead and Salzberg, 2012). Poorly aligned sequences were filtered using the Samtools package (version 1.7) with options -h -F 4 -q 10 (Li et al., 2009). The editing efficiency for each biological replicate was calculated by dividing the number of reads that contain the edit of interest, but were otherwise perfectly aligned, by the number of perfectly aligned reads containing either the wild-type or edited sequence. For paired-end sequencing data, half of the number of reads found in strand pairs was excluded from both the mutant and wild-type sequences to prevent double-counting of reads. The final editing percentage for each target locus was determined by averaging over the editing percentages in all biological replicates. The processed editing efficiencies in Kim et al. (2021) were obtained from their Supplementary Tables 3, 4.
Genomic locations of H3K9me3 and H3K27me3 histone modifications were determined from the aggregate of chromatin immuno-precipitation sequencing (ChIP-seq) datasets in HEK293 cells from the Encyclopedia of DNA Elements (ENCODE) consortium (ENCODE Project Consortium, 2012; Luo et al., 2020), with the accession codes having the prefix “ENC,” and the Gene Expression Omnibus (GEO) (Barrett et al., 2013), with the accession codes having the prefix “GSM.” For H3K9me3, the IP sequencing data used were GSM4301086 (Broche et al., 2021), the combination of ENCFF002AAX and ENCFF002AAZ as replicates, and the combination of GSM3452796 and GSM3452797 also as replicates (Tarjan et al., 2019). The corresponding inputs were GSM4301092, ENCFF000WXY, and GSM4445881 (Lamb et al., 2019). Raw reads were aligned to GRCh38 using Bowtie (version 2.4.1). For H3K27me3, the IP sequencing data were GSM3907592 (Lamb et al., 2019), GSM4301076 (Broche et al., 2021), GSM4586041 (Fan et al., 2020), and GSM4859391 (Fan et al., 2021); the corresponding inputs were GSM3907592, GSM4301081, GSM4586041, and GSM4859385. To determine the consensus genomic locations of H3K9me3 enrichment relative to the input, we first constructed six row vectors from the
RNA-DNA hybridization energies were obtained by computing the difference in length-normalized Gibbs free energy (
Nucleosome occupancy in the lymphoblastoid cell line GM12878 was determined from MNase-sequencing bigwig tracks from the ENCODE portal under the accession code ENCFFOOOVME (Luo et al., 2020). Average nucleosome occupancy in a particular genomic range was calculated by determining the sum of MNase-seq signals at every genomic coordinate in the protospacer region, normalized by the length of the protospacer.
PE off-targets were computed by aligning the protospacer sequence to regions in the hg38 genome that are also upstream of an NGG PAM site. A genomic locus was considered to be an off-target if its aligned sequence matched the reference protospacer sequence up to three mismatches outside the PAM site and if there were no mismatches in the GG dinucleotide of the PAM. These off-targets were determined by using the Cas-OFFInder package (Bae et al., 2014).
Linear regression with combined
where
where
It was previously observed that having G as the last templated nucleotide at RTTL =
where
The data provided by Kim et al. contained editing efficiencies for PBSL = 13 at only nonconsecutive RTTLs (RTTL = 10, 12, 15, and 20); thus, the OLS was applied on the sum of stepwise differences in three ranges of RTTL: 1)
We trained a DNN model to predict the edit percentages of all pegRNA designs from Kim et al. (2021) and thereby learn the salient sequence features influencing prime editor efficiency. The inputs to the DNN were 47
To capture position-specific effects of sequence features relative to the nick site, we used independent filters for different positions in the target sequence, instead of using convolutional filters with shared weights. The input sequences were thus divided into all possible overlapping 8-mers, resulting in 40 8-mers for a sequence of length 47 nt. Each 8-mer was passed through 10 filters of kernel size
The full dataset from Kim et al. was divided into 80% training, 10% validation, and 10% test sets, grouping together pegRNA designs with the same target site but with different PBSLs or RTTLs into a common set. Since the edit percentage of approximately 60% of the pegRNA designs was less than 10%, we assigned training weights to each pegRNA design in the training set in order to balance the dataset and prevent the DNN from learning biased features from over-represented poorly edited sites. For this purpose, the training set was partitioned according to their observed edit percentages, with a bin size of 1%. A pegRNA design, indexed by
where
To extract sequence features learned by the DNN, we used a modified version of the simulated annealing (SA) algorithm to determine the optimal sequences that maximize the output of the DNN (Tsallis and Stariolo, 1996; Finnegan et al., 2020); we then used the MaxEnt algorithm to identify salient positions and nucleotide preferences in the optimal sequences (Finnegan and Song, 2017) (Supplementary Methods S3, S4).
In order for a PE to be able to edit its target site, the spacer region of pegRNA must first access and hybridize to the complementary DNA sequence at the targeted locus. We thus hypothesize that target sites residing in heterochromatin, a condensed state of highly packaged DNA, may be blocked from access by PEs and thereby have low editing efficiency. Consistent with our reasoning, it has been previously observed that chromatin structure can interfere with the CRISPR-Cas9 endonuclease activity (Wu et al., 2014; Daer et al., 2017; Yarrington et al., 2018; Liu et al., 2019). We thus investigated the effect of heterochromatin on PE efficiency using genome editing data from Anzalone et al. (2019), Kim et al. (2021), and additional in-house validation experiments. To determine heterochromatin locations in HEK293 cells, which were used in all three studies, we integrated several publicly available H3K9me3 and H3K27me3 ChIP-seq datasets as a proxy for indicating closed chromatin regions (Methods). After obtaining consensus regions of enrichment for each of these histone modifications using singular value decomposition of the joint IP and input data matrices (Methods; Supplementary Method S2; Supplementary Figure S1), we computed the genomic distances from each target site to the nearest H3K9me3- and H3K27me3-modified regions and used these distances as features for predicting editing efficiency. When considering edit percentage as a function of the distance from the protospacer to the nearest H3K9me3-modified region, we observed that the edit percentages at sites close to H3K9me3 were approximately 0%, whereas higher edit percentages were possible farther away (Supplementary Figure S2A).
This observation motivated us to search for binary classification of target sites as being either weakly or strongly editable, choosing an efficiency threshold of 1% to binarize the data. We then trained a multivariate logistic regression model to classify all 32 endogenous target sites in Kim et al., using the distances to H3K9me3- and H3K27me3-modified regions as features (Methods; Supplementary Method S2). The probability threshold separating the binary editing categories was set to maximize the finite positive likelihood ratio of the true positive rate to the false positive rate. Performing the t-test for each regression coefficient and determining the bivariate and univariate logistic regression decision boundary showed the H3K27me3 variable to be insignificant (
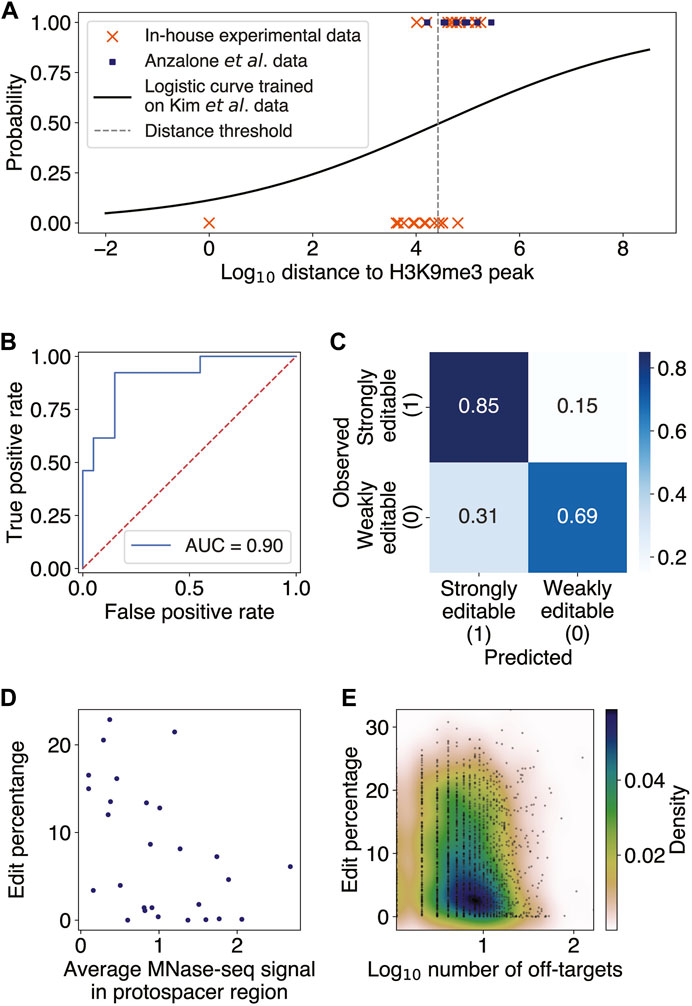
FIGURE 1. Constitutive heterochromatin near target sites impedes prime editing. (A) Scatter plot of binary classes of editability as a function of log10 distance from the target site to the nearest H3K9me3 peak (Methods; Supplementary Method S2). The distance threshold (dashed gray line) is at ∼26 kb. The logistic regression was trained on 32 endogenous targets from Kim et al. and tested on editing data from Anzalone et al. and our in-house experiments. (B) Receiver operator characteristic curve (ROC) for the logistic regression evaluated on the test set, with an area under the ROC (AUROC) of 0.90. (C) Confusion matrix for the logistic regression predictions on the test set. The overall accuracy was 75%. (D) Scatter plot of edit percentage as a function of the MNase-seq signal averaged over the protospacer region (Pearson
Having confirmed that high-order chromatin accessibility affects PE efficiency, we next investigated the effect of local chromatin structure on prime editing. It has been previously reported both in vitro and in vivo that nucleosomes inhibit the efficiency of CRISPR-Cas9 endonuclease, suggesting that histone proteins may block the target DNA access to Cas9 (Hinz et al., 2015; Yarrington et al., 2018). Given the shared components between the CRISPR-Cas9 and PE constructs, we thus sought to test whether nucleosome positioning at the target sites might also inhibit PE editing efficiencies. Since there were no publicly available MNase-seq datasets for HEK293 and most nucleosome positioning was shown to possess some degree of consistency across different cell lines (Gaffney et al., 2012) and partially exhibit intrinsic DNA sequence preferences (Ioshikhes et al., 1996; Segal et al., 2006; Gupta et al., 2008; Jin et al., 2016), we used the available nucleosome occupancy data in the lymphoblastoid cell line GM12878 (Methods). We found a significant negative correlation between PE edit percentage and nucleosome occupancy in the endogenous target sites of Kim et al. and Anzalone et al. (Pearson
Given that the PBS and spacer regions of pegRNA must hybridize to the target DNA sequence in order for editing to occur, with the spacer being the longer of the two sequences, we examined the off-target effect of having several genomic loci complementary to the spacer region on editing efficiency. We found only a weak but statistically significant negative correlation between the number of spacer off-targets and the edit rate averaged over all pegRNAs with the same spacer (Spearman
Our aforementioned logistic regression model has revealed that target sites close to heterochromatin are unlikely to be editable. Once a target site away from heterochromatin is selected, the next important step is to optimize the pegRNA design by adjusting the lengths of PBS and RT templates. It was previously reported that altering the PBS length (PBSL) and RT template length (RTTL) could have a drastic effect on PE2 efficiency, even when targeting the same site for the same edit, and that having G as the last templated nucleotide tended to decrease the PE2 efficiency (Anzalone et al., 2019). We further observed in Anzalone et al.’s data that 1) at the EMX1 locus containing no Gs in the RT region (Anzalone et al.'s Figure 2B), having A as the last templated nucleotide consistently resulted in lower edit rates, suggesting that other nucleotides apart from G might also modulate the editing efficiency; 2) at the FANCF locus (Anzalone et al.'s Figure 2B), even though the last templated nucleotide was G at RTTLs of both 10 and 18 nts, the edit rate for 18 nts was much higher than that for 10 nts, suggesting that the presence of G alone could not explain the variability of edit rates as a function of RT template sequence composition. These findings motivated us to use the sequence content in the target region as features in predicting the PE2 efficiency as PBSL and RTTL were varied. We thus developed an ordinary least squares (OLS) linear regression model to predict stepwise differences in edit percentages between two adjacent RTTLs, using the last templated nucleotides as features (Methods).
Anzalone et al. generated edit percentages (defined as the fractions of total reads with correct edits) of PE2 at eight different gene loci using pegRNAs with varying ranges of RTTL for each locus, with the largest overlap of lengths among the designs being between 10 and 20 nts. We thus chose this range [10 nt, 20 nt] to train an OLS linear regression model for predicting the stepwise differences,
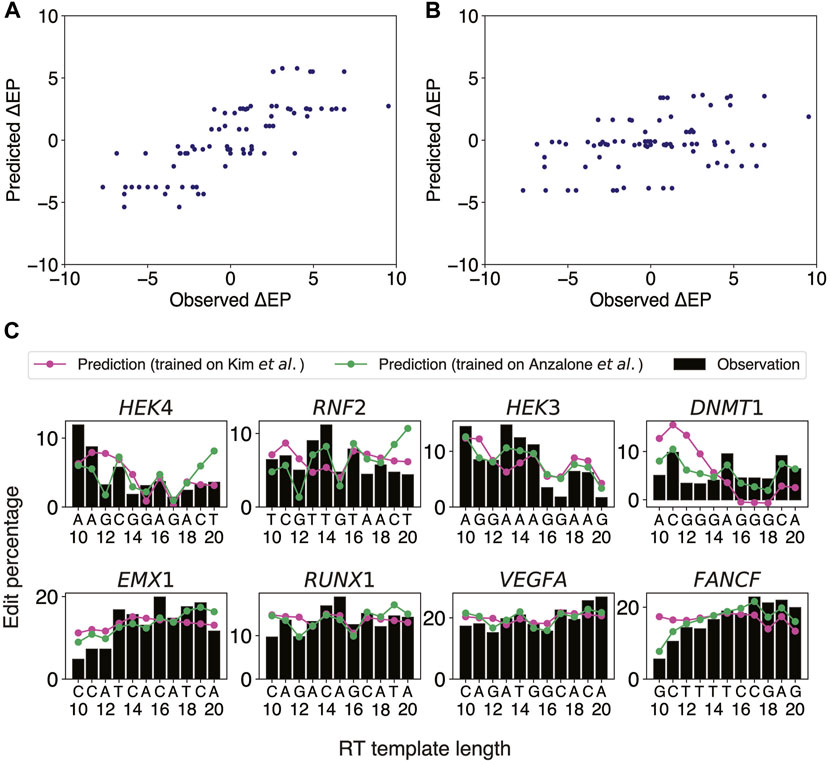
FIGURE 2. OLS linear regression using the last two templated nucleotides robustly predicts stepwise differences in edit percentage between two consecutive RTTLs. (A) Scatter plot of the predicted versus observed differences in edit percentage when the OLS linear regression model was trained and tested on Anzalone et al.’s data. (B) Scatter plot of the predicted versus observed differences in edit percentage when trained on the integrated sites from Kim et al.’s data and tested on Anzalone et al.’s data. (C) Predicted and observed edit percentages as a function of RTTL for eight different target sites from Anzalone et al. For each target, the absolute predicted edit percentage at RTTL = 10 was set such that the average of the predicted edit percentages across RTTLs matches the corresponding average of the observed edit percentages.
Having confirmed that our OLS linear regression model performed well on the eight target sites from Anzalone et al., we further investigated whether a similar approach could be generalized to Kim et al.’s data (Kim et al., 2021), which measured editing efficiencies at thousands of target sites by high-throughput sequencing. The PBSL was set to 13 nts, to be consistent with the range used in Anzalone et al. Since Kim et al. measured the edit percentages only for RTTLs = 10, 12, 15, and 20, we could not directly apply our stepwise approach on these data. We thus divided the data for different RTTLs into three ranges and trained an OLS linear regression model to predict consecutive stepwise differences in edit percentages for each range separately (Methods): 1) RTTLs from 10 to 12; 2) RTTLs from 12 to 15; and 3) RTTLs from 15 to 20 (Supplementary Figure S3). Ten-fold cross-validation verified the robustness of our model within each range (5.1% RMSE; average of Pearson correlation across folds = 0.33 for RTTLs in the range [10,12] and [15,20], and 0.25 for RTTLs of [12,15]; Supplementary Table S6). However, the effect sizes of the four nucleotides differed somewhat across the three ranges (Supplementary Tables S7–S9). When the RTTL is in the range [15,20], the effect on
Testing the regression model trained on Kim et al.’s data on Anzalone et al.’s data in the corresponding RTTL ranges (Supplementary Tables S7–S9), the predicted
There were some discrepancies between the two independent datasets regarding the regression coefficients trained on shorter RTTL designs. For example, Anzalone et al. observed a decrease in editing efficiency whenever the last templated nucleotide was G throughout all RTTLs. By contrast, Kim et al. observed that, on average, editing efficiency was highest when the last templated nucleotide was G at RTTLs = 10 and 12, while it was lowest for the last templated G only at RTTL = 20 (Kim et al.'s Figure 2F). Further investigation is needed to understand why G has an opposite effect on editing efficiency at short RTTLs and why this phenomenon is not universal across independent datasets. We shall revisit the position-specific effects of G on editing efficiency in subsequent sections.
The OLS linear regression analysis yielded insight into the preferred RTTLs of pegRNAs for a given target site, but it could not reveal the optimal target, given multiple PAM candidates in the region of interest. We therefore built an elastic net model (Zou and Hastie, 2005), a linear regression model with
The elastic net predictions significantly correlated with the observed edit percentages in each pair of PBSL and RTTL (Pearson
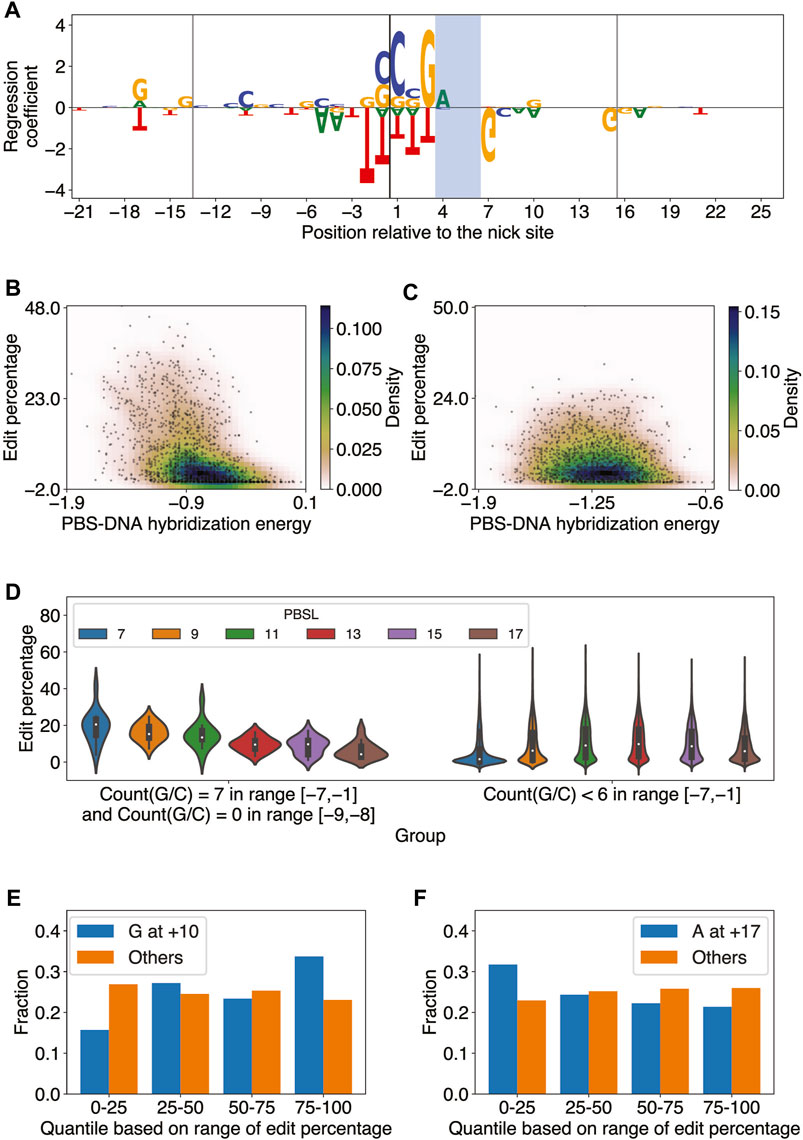
FIGURE 3. Elastic net regression model learns sequence features important for predicting PE2 efficiency. (A) Elastic net regression coefficients for all edited-strand nucleotides in the range [-21,+26] relative to the nick site for PBSL = 13 and RTTL = 15. The model was trained on the editing data for integrated target sites from Kim et al. The nucleotide heights represent the absolute value of the corresponding regression coefficients. The vertical lines denote the borders of the PBS and RT template regions, and the PAM site is shaded. (B) Scatter plot of the edit percentage versus PBS–DNA hybridization energy for PBSL = 7 and RTTL = 10, where the colors represent the density of data points. Pearson
The regression coefficients also showed that the presence of G as the last templated nucleotide tended to increase the editing efficiency when RTTL = 10, regardless of the PBSL, or when RTTL = 12 and PBSL
The large effect size of the GC content in the PBS region for short PBSL designs, together with the accompanying reduction in the negative effect of the last templated nucleotide, suggested that optimal choices of pegRNA for target loci containing high GC content in the PBS region would involve short PBSLs. Supporting this idea, we observed that when the pegRNA contained only G/C in the range [−7,−1] and A/T at −9 and −8, the edit percentage was, on average, highest for PBSL = 7 (Figure 3D; highest median edit percentage of 20.5% when PBSL = 7); here, we considered A/T at the −9 and −8 positions based on the regression coefficients learned by the elastic net for PBSL = 7 (Supplementary Figure S4). When fewer than 6 G/Cs were found in the [−7,−1] range, PBSLs between 11 and 15 had, on average, higher editing efficiency than other PBSLs (Figure 3D; highest median edit percentage of 9.8% when PBSL = 13). Given a target site, these findings thus provided a general guideline for determining the optimal PBSL based solely on the 9-bp-long sequence upstream of the nick site in the edited strand.
Some patterns of regression coefficients were shared among most combinations of PBSL and RTTL. Based on the magnitude of regression coefficients, the most salient nucleotides were those positioned around the nick site (Supplementary Figure S4): C and G nucleotides at locations from −1 to +3, immediately 5’ to the PAM site, led to high editing efficiency, whereas T in the range from −2 to +3 was to be avoided. G/A at the −17 position was positively correlated with editing efficiency, while T at the same position was negatively correlated. In general, C at the +20 position was positively correlated with editing efficiency across designs. For the PAM site, AGG was favored and CGG disfavored for optimizing editing efficiency. Immediately after the PAM site, G at the +7 position was strongly anti-correlated with editing efficiency. Surprisingly, the presence of G at the +10 position was significant and tended to increase editing efficiency, not only for RTTL = 10, in which case G would be the last templated nucleotide as described previously, but also for RTTL = 15 or 20 when PBSL
In summary, the elastic net regression models trained on each combination of PBSL and RTTL learned distinct and common sequence features modulating editing efficiency. The effect of the last templated G nucleotide on editing efficiency depended on both PBSL and RTTL of pegRNA designs. Moreover, pegRNAs with short PBSLs highly depended on the GC content of the PBS region, perhaps to help stabilize the hybridization to the complementary DNA. The PBS RNA-DNA hybridization energy was significantly anti-correlated with edit percentage in all combinations of PBSL and RTTL, but pegRNAs with shorter PBSLs showed more pronounced dependence. Finally, the presence of G/C nucleotides was most critical immediately 5’ to the PAM site for all combinations of PBSL and RTTL.
It is challenging to model nonlinear effects of coupled nucleotides at multiple positions using elastic net regression. To extend our linear regression approach, we thus trained a DNN model on Kim et al.’s data (Kim et al., 2021) to predict the edit percentage of a pegRNA design given its length and the sequence of the target site and flanking regions (Figure 4A; Methods). The Pearson correlation coefficient between the observed and predicted edit percentages in the test set was 0.73 (Figure 4B; Methods).
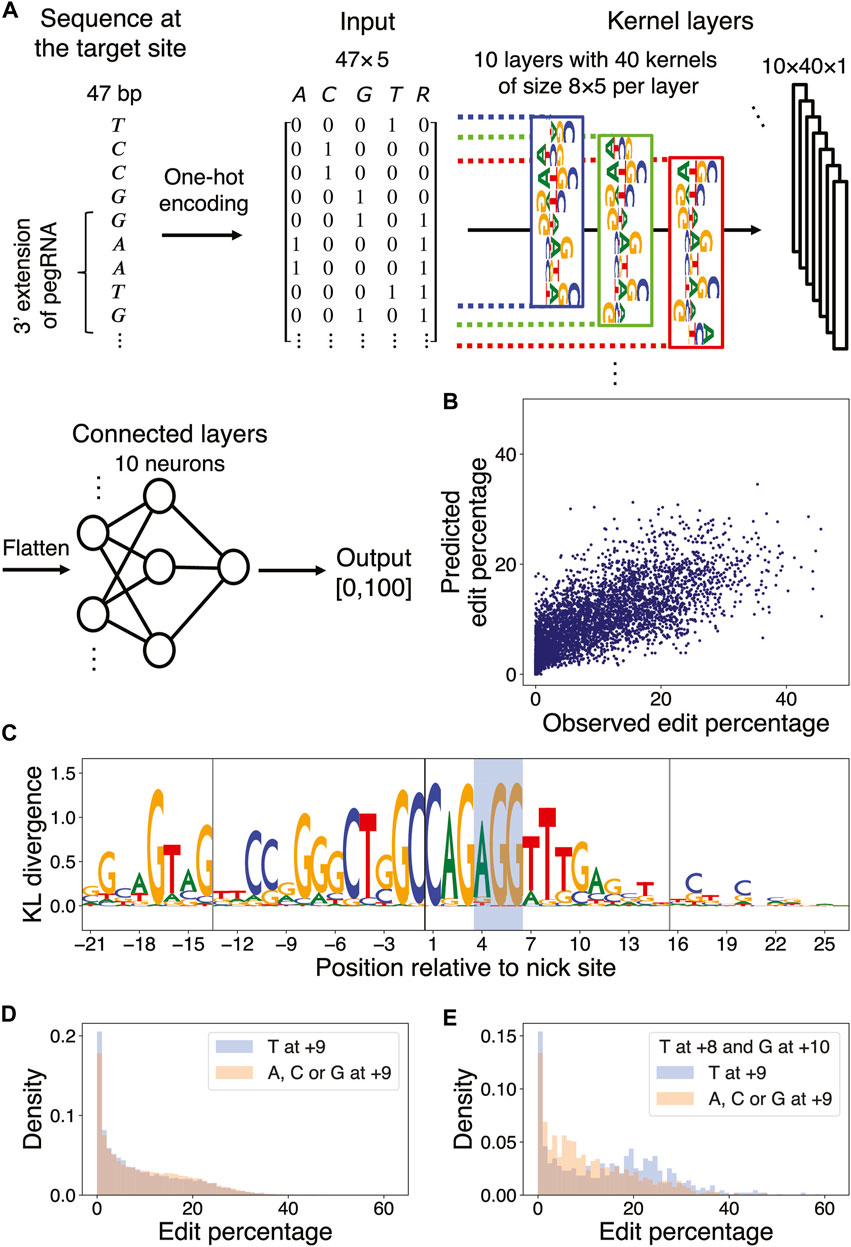
FIGURE 4. DNN learns marginal and multi-nucleotide sequence features. (A) DNN architecture. The first four columns of the input matrix indicate the presence of a particular nucleotide in the range [−21,+26] relative to the nick site via one-hot encoding; the 5th column indicates whether a particular position resides in the union of the PBS and RT template regions of the pegRNA being considered. The input is passed through a layer of kernels and two dense layers to yield an output value between 0 and 100. (B) Scatter plot of observed versus predicted edit percentages of the target sites in the test set (Pearson
To understand optimal sequences associated with high edit percentages, we applied a simulated annealing (SA) method for maximizing the prediction of a trained DNN over its input space via Markov Chain Monte Carlo (MCMC) sampling (Finnegan et al., 2020). We accelerated the simulation by using a broadened sampling distribution stemming from nonextensive statistical mechanics (Tsallis and Stariolo, 1996) and obtained the target sequences maximizing the predicted edit percentages for each combination of PBSL and RTTL (Supplementary Method S3; Supplementary Tables S11–S14). To identify the DNN-learned salient features in these optimal sequences, we next applied MaxEnt, another MCMC sampling method based on the maximum entropy principle, for generating new input sequences that produce similar DNN predictions as those of the initial input sequence (Finnegan and Song, 2017). Upon initializing the MaxEnt chains at the optimal target sequences, nucleotide preferences important for maximizing PE efficiency were extracted by calculating the position-wise Kullback–Leibler (KL) divergence between the nucleotide distribution in the sampled sequences and the uniform null distribution; in this formalism, the larger the KL divergence, the more relevant the nucleotide (Figure 4C; Supplementary Figure S7; Supplementary Method S4).
The DNN confirmed the sequence features previously detected by the elastic net model. For example, it learned that C at the +1 position, G at the +3 position, and G at the −17 position relative to the nick site were associated with high edit percentage; other similarities with the elastic net results included the preferences for A as the first nucleotide in the PAM sequence, for C or G in the PBS region, and for C at the +20 position. In addition, the DNN also discovered new features. For example, although the elastic net model detected no strong preference for any particular nucleotide at position +9, except for perhaps a weak preference for G (Supplementary Figure S4), the DNN found T at this position to be associated with optimal editing. This discrepancy in the feature importance arose from the difference in model architectures, that is, the elastic net model learned only the independent effects of individual nucleotides, whereas the DNN accounted for aggregate effects of nucleotides captured by the filters. When considering only the marginal effect of single nucleotides at the +9 position, target sequences with T at that position actually had a lower edit percentage on average than those with other nucleotides at that same position (9.00% for T vs. 9.72% for A, C, and G at +9; Figure 4D). However, imposing the +8 position to be T and the +10 position to be G showed that target sequences with T at +9 had a higher edit percentage on average than those with other nucleotides at +9 (14.33% for T vs. 10.72% for A, C, and G at +9; Figure 4E; Supplementary Figures S8A, B). Similarly, the marginal effect of T at the −4 position, residing in the PBS region, was negligible; however, the DNN uncovered a role of T in the context of a GC-rich background: for sequences with 100% GC content in the range [−7,−1], except at the −4 position, the presence of T at −4 substantially increased the edit percentage by 6.90% on average (Supplementary Figures S8C, D). Finally, the DNN found a preference for CAG at [+1,+3] in optimal target sequences, whereas the elastic net model found only a marginal effect of G/C at +2 and a less pronounced trinucleotide effect of C (G/C) G in the same region (Supplementary Figures S8E, F). In summary, our DNN was able to predict the observed edit percentage with high accuracy and yielded interpretable results regarding both marginal and context-dependent sequence features of optimal target sites.
We have demonstrated that both regional heterochromatin and local nucleosome occlusion of target sites may decrease PE2 editing efficiency, with a more pronounced effect observed for the H3K9me3 modification, perhaps because unwinding DNA may be particularly difficult in a heterochromatin environment where multiple nucleosomes are condensed together. These results are consistent with those of a recent report showing that inducing an open chromatin state improves PE and base editor efficiencies (Liu et al., 2022). Our study thus provides evidence for the hypothesis that chromatin structure modulates genome editing efficiency by interfering with the accessibility of target sites to the Cas9 protein and guide RNA. Unlike the significant predictive power of H3K9me3 in classifying strongly and weakly editable target sites, however, the feature of H3K27me3 was insignificant (Supplementary Figures S2A, B; Supplementary Table S3). While both H3K9me3 and H3K27me3 are associated with heterochromatin formation, H3K9me3 is associated with constitutive heterochromatin, whereas H3K27me3 is associated with facultative heterochromatin (Saksouk et al., 2014), which depends on the presence of certain stimuli, and thus, remains conducive to dynamic changes (Oberdoerffer and Sinclair, 2007). Further investigation is needed to decipher the precise differences in folding pattern, making constitutive heterochromatin more resistant to editing than facultative heterochromatin.
Our work shows that, in addition to the chromatin environment, the local sequence content of target sites can also modulate editing efficiency. For example, strategic positioning of G and C nucleotides in the PBS and RT regions increases the editing efficiency, indicating that G: C base pairing between DNA and pegRNA helps anchor the pegRNA to the target site prior to editing. A similar observation regarding the GC content in the PBS region was previously reported (Kim et al., 2021); however, our study further clarifies that 1) the presence of G or C is most important in the RTT region, immediately downstream of the PBS region, at positions +1 and +3; 2) within the PBS region itself, the positive effect of G or C is most pronounced near the nick site and rapidly decays away from the nick site for PBSL
When choosing a target site, our results recommend selecting target sequences that 1) have AGG in the PAM site; 2) have high GC content in the PBS region, especially when the PBS is short; 3) avoid G as the last templated nucleotide if the RT template is longer than 12; and 4) have G at the −17 position relative to the nick site on the edited strand (Figure 5). Once the target site has been chosen, a PBSL in the [11,13] nt range is generally recommended, unless the target site has high GC content that provides stable hybridization between the pegRNA and target DNA, in which case a shorter PBSL is recommended. As for the effect of RTTL, pegRNAs with shorter RTTL have, on average, higher editing efficiency than those with longer RTTL; when the available PAM site is far away from the desired site, such that having RTTL >15 nt is unavoidable, we recommend the following designs: prioritize the last templated nucleotide in the order T
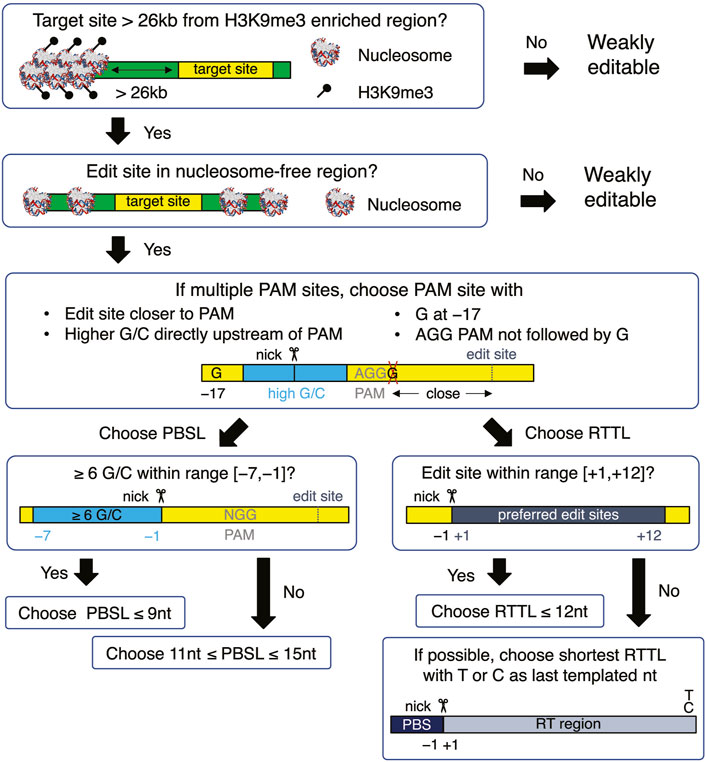
FIGURE 5. Flowchart for designing an optimal pegRNA for the selected target site. The yellow boxes represent the target site, which is defined to be the union of edited-strand nucleotides spanning the protospacer region, PBS region, and RT region. The green boxes represent the DNA flanking the target site. All nucleotide numbers are relative to the nick site.
Our computational analysis has identified several epigenetic and sequence features that need to be considered when designing pegRNAs to optimize PE efficiency. Future availability of additional high-throughput genome editing data and biophysical studies investigating how PEs search and bind target sequences will help further improve our understanding and make this technology feasible for effective biomedical applications. In particular, our current models do not take into account biochemical and biophysical processes involving reverse transcriptase activity, priming, flap equilibration, and DNA repair. Unfortunately, relevant parameters required for investigating these processes are currently limited. Incorporating such features obtained from detailed biophysical experiments and molecular dynamics simulations in the future could further improve the optimization of prime editors for practical applications.
Part of this project was completed while JS was on a sabbatical leave in the Center for Theoretical Physics at the Massachusetts Institute of Technology and the Department of Statistics at Harvard University.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, PRJNA949853.
PP-P and JS conceived and supervised the project. SK and JY carried out the computational analyses, aided by DN. WW performed the validation experiments. SK, JY, and JS wrote the manuscript with contributions from other authors. All authors contributed to the article and approved the submitted version.
The National Institutes of Health (R01CA163336 to JS, R01GM141296 to PP-P and JS); Grainger Engineering Breakthroughs Initiative to JS.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2023.1222112/full#supplementary-material
bp: base pair; CRISPR: clustered regularly interspaced short palindromic repeats; Cas9: CRISPR-associated protein 9; ChIP-seq: chromatin immune-precipitation sequencing; DNN: deep neural network; EP: edit percentage; MCMC: Markov Chain Monte Carlo; OLS: ordinary least squares; PAM: protospacer adjacent motif; PBS: primer-binding site; PBSL: primer-binding site length; PE: prime editor; pegRNA: prime editor guide RNA; RT: reverse transcription; RMSE: root mean square error; RTTL: reverse transcription template length; SA: simulated annealing.
Anders, C., Niewoehner, O., Duerst, A., and Jinek, M. (2014). Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 513, 569–573. doi:10.1038/nature13579
Anzalone, A. V., Randolph, P. B., Davis, J. R., Sousa, A. A., Koblan, L. W., Levy, J. M., et al. (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 576, 149–157. doi:10.1038/s41586-019-1711-4
Bae, S., Park, J., and Kim, J. S. (2014). Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 30, 1473–1475. doi:10.1093/bioinformatics/btu048
Barrett, T., Wilhite, S. E., Ledoux, P., Evangelista, C., Kim, I. F., Tomashevsky, M., et al. (2013). NCBI GEO: Archive for functional genomics data sets--update. Nucleic Acids Res. 41, D991–D995. doi:10.1093/nar/gks1193
Barski, A., Cuddapah, S., Cui, K., Roh, T. Y., Schones, D. E., Wang, Z., et al. (2007). High-resolution profiling of histone methylations in the human genome. Cell 129, 823–837. doi:10.1016/j.cell.2007.05.009
Broche, J., Kungulovski, G., Bashtrykov, P., Rathert, P., and Jeltsch, A. (2021). Genome-wide investigation of the dynamic changes of epigenome modifications after global DNA methylation editing. Nucleic Acids Res. 49, 158–176. doi:10.1093/nar/gkaa1169
Burrows, C. J., and Muller, J. G. (1998). Oxidative nucleobase modifications leading to strand scission. Chem. Rev. 98, 1109–1152. doi:10.1021/cr960421s
Chapman, J. R., Taylor, M. R., and Boulton, S. J. (2012). Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell 47, 497–510. doi:10.1016/j.molcel.2012.07.029
Chen, P. J., Hussmann, J. A., Yan, J., Knipping, F., Ravisankar, P., Chen, P. F., et al. (2021). Enhanced prime editing systems by manipulating cellular determinants of editing outcomes. Cell 184, 5635–5652.e29. doi:10.1016/j.cell.2021.09.018
Cong, L., Ran, F. A., Cox, D., Lin, S., Barretto, R., Habib, N., et al. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823. doi:10.1126/science.1231143
Daer, R. M., Cutts, J. P., Brafman, D. A., and Haynes, K. A. (2017). The impact of chromatin dynamics on cas9-mediated genome editing in human cells. ACS Synth. Biol. 6, 428–438. doi:10.1021/acssynbio.5b00299
Doudna, J. A., and Charpentier, E. (2014). Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096. doi:10.1126/science.1258096
Encode Project Consortium (2012). An integrated encyclopedia of DNA elements in the human genome. Nature 489, 57–74. doi:10.1038/nature11247
Fan, H., Guo, Y., Tsai, Y. H., Storey, A. J., Kim, A., Gong, W., et al. (2021). A conserved BAH module within mammalian BAHD1 connects H3K27me3 to Polycomb gene silencing. Nucleic Acids Res. 49, 4441–4455. doi:10.1093/nar/gkab210
Fan, H., Lu, J., Guo, Y., Li, D., Zhang, Z. M., Tsai, Y. H., et al. (2020). BAHCC1 binds H3K27me3 via a conserved BAH module to mediate gene silencing and oncogenesis. Nat. Genet. 52, 1384–1396. doi:10.1038/s41588-020-00729-3
Finnegan, A. I., Kim, S., Jin, H., Gapinske, M., Woods, W. S., Perez-Pinera, P., et al. (2020). Epigenetic engineering of yeast reveals dynamic molecular adaptation to methylation stress and genetic modulators of specific DNMT3 family members. Nucleic Acids Res. 48, 4081–4099. doi:10.1093/nar/gkaa161
Finnegan, A., and Song, J. S. (2017). Maximum entropy methods for extracting the learned features of deep neural networks. PLoS Comput. Biol. 13, e1005836. doi:10.1371/journal.pcbi.1005836
Gaffney, D. J., Mcvicker, G., Pai, A. A., Fondufe-Mittendorf, Y. N., Lewellen, N., Michelini, K., et al. (2012). Controls of nucleosome positioning in the human genome. PLoS Genet. 8, e1003036. doi:10.1371/journal.pgen.1003036
Gapinske, M., Luu, A., Winter, J., Woods, W. S., Kostan, K. A., Shiva, N., et al. (2018). CRISPR-SKIP: Programmable gene splicing with single base editors. Genome Biol. 19, 107. doi:10.1186/s13059-018-1482-5
Gaudelli, N. M., Komor, A. C., Rees, H. A., Packer, M. S., Badran, A. H., Bryson, D. I., et al. (2017). Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage. Nature 551, 464–471. doi:10.1038/nature24644
Gupta, S., Dennis, J., Thurman, R. E., Kingston, R., Stamatoyannopoulos, J. A., and Noble, W. S. (2008). Predicting human nucleosome occupancy from primary sequence. PLoS Comput. Biol. 4, e1000134. doi:10.1371/journal.pcbi.1000134
Hinz, J. M., Laughery, M. F., and Wyrick, J. J. (2015). Nucleosomes inhibit Cas9 endonuclease activity in vitro. Biochemistry 54, 7063–7066. doi:10.1021/acs.biochem.5b01108
Hsu, P. D., Lander, E. S., and Zhang, F. (2014). Development and applications of CRISPR-Cas9 for genome engineering. Cell 157, 1262–1278. doi:10.1016/j.cell.2014.05.010
Ioshikhes, I., Bolshoy, A., Derenshteyn, K., Borodovsky, M., and Trifonov, E. N. (1996). Nucleosome DNA sequence pattern revealed by multiple alignment of experimentally mapped sequences. J. Mol. Biol. 262, 129–139. doi:10.1006/jmbi.1996.0503
Jin, H., Rube, H. T., and Song, J. S. (2016). Categorical spectral analysis of periodicity in nucleosomal DNA. Nucleic Acids Res. 44, 2047–2057. doi:10.1093/nar/gkw101
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., and Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821. doi:10.1126/science.1225829
Jinek, M., East, A., Cheng, A., Lin, S., Ma, E., and Doudna, J. (2013). RNA-programmed genome editing in human cells. Elife 2, e00471. doi:10.7554/eLife.00471
Kim, H. K., Kim, Y., Lee, S., Min, S., Bae, J. Y., Choi, J. W., et al. (2019). SpCas9 activity prediction by DeepSpCas9, a deep learning-based model with high generalization performance. Sci. Adv. 5, eaax9249. doi:10.1126/sciadv.aax9249
Kim, H. K., Yu, G., Park, J., Min, S., Lee, S., Yoon, S., et al. (2021). Predicting the efficiency of prime editing guide RNAs in human cells. Nat. Biotechnol. 39, 198–206. doi:10.1038/s41587-020-0677-y
Kim, Y. B., Komor, A. C., Levy, J. M., Packer, M. S., Zhao, K. T., and Liu, D. R. (2017). Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 35, 371–376. doi:10.1038/nbt.3803
Komor, A. C., Kim, Y. B., Packer, M. S., Zuris, J. A., and Liu, D. R. (2016). Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 533, 420–424. doi:10.1038/nature17946
Komor, A. C., Zhao, K. T., Packer, M. S., Gaudelli, N. M., Waterbury, A. L., Koblan, L. W., et al. (2017). Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:G-to-T:A base editors with higher efficiency and product purity. Sci. Adv. 3, eaao4774. doi:10.1126/sciadv.aao4774
Kweon, J., Hwang, H. Y., Ryu, H., Jang, A. H., Kim, D., and Kim, Y. (2022). Targeted genomic translocations and inversions generated using a paired prime editing strategy. Mol. Ther. 31, 249–259. doi:10.1016/j.ymthe.2022.09.008
Lamb, K. N., Bsteh, D., Dishman, S. N., Moussa, H. F., Fan, H., Stuckey, J. I., et al. (2019). Discovery and characterization of a cellular potent positive allosteric modulator of the polycomb repressive complex 1 chromodomain, CBX7. Cell Chem. Biol. 26, 1365–1379.e22. doi:10.1016/j.chembiol.2019.07.013
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi:10.1038/nmeth.1923
Leistico, J. R., Saini, P., Futtner, C. R., Hejna, M., Omura, Y., Soni, P. N., et al. (2021). Epigenomic tensor predicts disease subtypes and reveals constrained tumor evolution. Cell Rep. 34, 108927. doi:10.1016/j.celrep.2021.108927
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi:10.1093/bioinformatics/btp352
Liu, G., Yin, K., Zhang, Q., Gao, C., and Qiu, J. L. (2019). Modulating chromatin accessibility by transactivation and targeting proximal dsgRNAs enhances Cas9 editing efficiency in vivo. Genome Biol. 20, 145. doi:10.1186/s13059-019-1762-8
Liu, N., Zhou, L., Lin, G., Hu, Y., Jiao, Y., Wang, Y., et al. (2022). HDAC inhibitors improve CRISPR-Cas9 mediated prime editing and base editing. Mol. Ther. Nucleic Acids 29, 36–46. doi:10.1016/j.omtn.2022.05.036
Liu, P., Liang, S. Q., Zheng, C., Mintzer, E., Zhao, Y. G., Ponnienselvan, K., et al. (2021a). Improved prime editors enable pathogenic allele correction and cancer modelling in adult mice. Nat. Commun. 12, 2121. doi:10.1038/s41467-021-22295-w
Liu, Y., Kao, H. I., and Bambara, R. A. (2004). Flap endonuclease 1: A central component of DNA metabolism. Annu. Rev. Biochem. 73, 589–615. doi:10.1146/annurev.biochem.73.012803.092453
Liu, Y., Yang, G., Huang, S., Li, X., Wang, X., Li, G., et al. (2021b). Enhancing prime editing by Csy4-mediated processing of pegRNA. Cell Res. 31, 1134–1136. doi:10.1038/s41422-021-00520-x
Luo, Y., Hitz, B. C., Gabdank, I., Hilton, J. A., Kagda, M. S., Lam, B., et al. (2020). New developments on the Encyclopedia of DNA Elements (ENCODE) data portal. Nucleic Acids Res. 48, D882–D889. doi:10.1093/nar/gkz1062
Mali, P., Yang, L., Esvelt, K. M., Aach, J., Guell, M., Dicarlo, J. E., et al. (2013). RNA-guided human genome engineering via Cas9. Science 339, 823–826. doi:10.1126/science.1232033
Moreno-Mateos, M. A., Vejnar, C. E., Beaudoin, J. D., Fernandez, J. P., Mis, E. K., Khokha, M. K., et al. (2015). CRISPRscan: Designing highly efficient sgRNAs for CRISPR-cas9 targeting in vivo. Nat. Methods 12, 982–988. doi:10.1038/nmeth.3543
Nelson, J. W., Randolph, P. B., Shen, S. P., Everette, K. A., Chen, P. J., Anzalone, A. V., et al. (2022). Engineered pegRNAs improve prime editing efficiency. Nat. Biotechnol. 40, 402–410. doi:10.1038/s41587-021-01039-7
Oberdoerffer, P., and Sinclair, D. A. (2007). The role of nuclear architecture in genomic instability and ageing. Nat. Rev. Mol. Cell Biol. 8, 692–702. doi:10.1038/nrm2238
Park, S. J., Jeong, T. Y., Shin, S. K., Yoon, D. E., Lim, S. Y., Kim, S. P., et al. (2021). Targeted mutagenesis in mouse cells and embryos using an enhanced prime editor. Genome Biol. 22, 170. doi:10.1186/s13059-021-02389-w
Petri, K., Zhang, W., Ma, J., Schmidts, A., Lee, H., Horng, J. E., et al. (2022). CRISPR prime editing with ribonucleoprotein complexes in zebrafish and primary human cells. Nat. Biotechnol. 40, 189–193. doi:10.1038/s41587-021-00901-y
Saksouk, N., Barth, T. K., Ziegler-Birling, C., Olova, N., Nowak, A., Rey, E., et al. (2014). Redundant mechanisms to form silent chromatin at pericentromeric regions rely on BEND3 and DNA methylation. Mol. Cell 56, 580–594. doi:10.1016/j.molcel.2014.10.001
Segal, E., Fondufe-Mittendorf, Y., Chen, L., Thastrom, A., Field, Y., Moore, I. K., et al. (2006). A genomic code for nucleosome positioning. Nature 442, 772–778. doi:10.1038/nature04979
Spencer, J. M., and Zhang, X. (2017). Deep mutational scanning of S. pyogenes Cas9 reveals important functional domains. Sci. Rep. 7, 16836. doi:10.1038/s41598-017-17081-y
Sugimoto, N., Nakano, S., Katoh, M., Matsumura, A., Nakamuta, H., Ohmichi, T., et al. (1995). Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry 34, 11211–11216. doi:10.1021/bi00035a029
Tarjan, D. R., Flavahan, W. A., and Bernstein, B. E. (2019). Epigenome editing strategies for the functional annotation of CTCF insulators. Nat. Commun. 10, 4258. doi:10.1038/s41467-019-12166-w
Tsallis, C., and Stariolo, D. A. (1996). Generalized simulated annealing. Phys. A 233, 395–406. doi:10.1016/s0378-4371(96)00271-3
Winter, J., Luu, A., Gapinske, M., Manandhar, S., Shirguppe, S., Woods, W. S., et al. (2019). Targeted exon skipping with AAV-mediated split adenine base editors. Cell Discov. 5, 41. doi:10.1038/s41421-019-0109-7
Wright, A. V., Nunez, J. K., and Doudna, J. A. (2016). Biology and applications of CRISPR systems: Harnessing nature's toolbox for genome engineering. Cell 164, 29–44. doi:10.1016/j.cell.2015.12.035
Wu, X., Scott, D. A., Kriz, A. J., Chiu, A. C., Hsu, P. D., Dadon, D. B., et al. (2014). Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells. Nat. Biotechnol. 32, 670–676. doi:10.1038/nbt.2889
Yarrington, R. M., Verma, S., Schwartz, S., Trautman, J. K., and Carroll, D. (2018). Nucleosomes inhibit target cleavage by CRISPR-Cas9 in vivo. Proc. Natl. Acad. Sci. U. S. A. 115, 9351–9358. doi:10.1073/pnas.1810062115
Keywords: prime editing, CRISPR–Cas9, heterochromatin, nucleosome positioning, DNA-RNA hybridization, nucleotide preference, machine learning, neural network interpretation
Citation: Kim S, Yuan JB, Woods WS, Newton DA, Perez-Pinera P and Song JS (2023) Chromatin structure and context-dependent sequence features control prime editing efficiency. Front. Genet. 14:1222112. doi: 10.3389/fgene.2023.1222112
Received: 13 May 2023; Accepted: 16 June 2023;
Published: 29 June 2023.
Edited by:
Meizhu Bai, Yale University, United StatesReviewed by:
Simon Sretenovic, University of Maryland, United StatesCopyright © 2023 Kim, Yuan, Woods, Newton, Perez-Pinera and Song. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun S. Song, songj@illinois.edu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.