 Shan Xueqing1
Shan Xueqing1 Wang Guizhi
Wang Guizhi Yang Liuxu
Yang Liuxu Chao Tianle
Chao Tianle
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 11 May 2023
Sec. RNA
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1185952
Fluvalinate is widely used in apiculture as an acaricide for removing Varroa mites, but there have been growing concerns about the negative effects of fluvalinate on honeybees in recent years. Previous research revealed changes in the miRNA and mRNA expression profiles of Apis mellifera ligustica brain tissues during fluvalinate exposure, as well as key genes and pathways. The role of circRNAs in this process, however, is unknown. The goal of this study was to discover the fluvalinate-induced changes in circular RNA (circRNA) expression profiles of brain tissue of A. mellifera ligustica workers. A total of 10,780 circRNAs were detected in A. mellifera ligustica brain tissue, of which eight were differentially expressed between at least two of the four time periods before and after fluvalinate administration, and six circRNAs were experimentally verified to be structurally correct, and their expression patterns were consistent with transcriptome sequencing results. Furthermore, ceRNA analysis revealed that five differentially expressed circRNAs (DECs) (novel_circ_012139, novel_circ_011690, novel_circ_002628, novel_circ_004765, and novel_circ_010008) were primarily involved in apoptosis-related functions by competitive binding with miRNAs. This study discovered changes in the circRNA expression profile of A. mellifera ligustica brain tissue caused by fluvalinate exposure, and it provides a useful reference for the biological function study of circRNAs in A. mellifera ligustica.
Apis mellifera ligustica is a high-value insect species that performs plant pollination in agricultural production (Aizen and Harder, 2009; Beck et al., 2015; Andonov et al., 2019) and produces high-value byproducts such as honey and royal jelly (Berry et al., 2013). The most common bee parasite is Varroa destructor, which is especially harmful to A. mellifera ligustica (Burley et al., 2008).
Fluvalinate is a pyrethroid acaricide that is commonly used to kill V. destructor (Camp et al., 2010). Fluvalinate can kill V. destructor by affecting the nervous system’s voltage-gated sodium channels, causing overexcited nerves and paralysis (Chao et al., 2022a). Fluvalinate, on the other hand, is toxic to A. mellifera ligustica (Chen et al., 2020; Chao et al., 2022b). Several studies have reported negative effects of fluvalinate exposure on A. mellifera ligustica, with the results indicating that A. mellifera ligustica’s sensory and memory systems may be impaired (Choi et al., 2006; Davies et al., 2007; Du et al., 2017; Ding et al., 2022). Our previous research has revealed changes in protein-coding gene and miRNA expression profile of A. mellifera ligustica brains caused by fluvalinate exposure, and some differentially expressed genes and miRNAs related to apoptosis, visual function, and neural response have been identified (Fischer and Raabe, 2018; El Agrebi et al., 2019). In our two previous studies, the phototransduction pathway was the primary pathway for significant enrichment, which may indicate that the toxicological mechanism of fluvalinate is associated with visual impairment. We also predicted that key genes and miRNAs including LOC412299, LOC411188, and ame-miR-3477-5p may be involved in fluvalinate resistance (Fischer and Raabe, 2018; El Agrebi et al., 2019). However, at present, the expression profile changes and functions of circRNA during fluvalinate exposure are still unclear.
Circular RNAs are naturally occurring RNAs found in eukaryotic transcriptomes (Frost et al., 2013). CircRNAs have been shown to have a “sponge” function for miRNAs (Glažar et al., 2014; Gan et al., 2017), and this mechanism can also be used to indirectly regulate mRNA expression (Gujar et al., 2019). Several studies have demonstrated the expression of circRNAs in multiple species of honeybees (Gujar et al., 2019; Han et al., 2021), with reports on the involvement of circRNAs in the regulation of A. mellifera ligustica brain functions related to foraging and nursing functions (Han et al., 2021). Currently, to the best of our knowledge, there have been no reports on the function of circRNAs in the fluvalinate exposure process.
In this study, we combined transcriptome sequencing data from previous studies to detect and analyze the changes in the circRNA expression profile of A. mellifera ligustica brain tissue before and after fluvalinate treatment, and we built a competing endogenous RNA (ceRNA) network by integrating the differentially expressed circRNA with miRNA and mRNA for a more in-depth analysis of circRNA function.
All animal protocols used in this study were approved by the Animal Protection and Ethics Committee of Shandong Agricultural University (protocol number: SDAUA-2018-055).
This study followed the same overall strategy, sample collection, total RNA extraction, and cDNA synthesis as our prior investigations (Fischer and Raabe, 2018; El Agrebi et al., 2019). Six A. mellifera ligustica colonies were obtained from the Fuxin Apiary in Tai’an City, Shandong Province, China. All colonies had the same potential and were randomly divided into two groups (three colonies as the test group and three colonies as the control group). The test colonies were administered with a standard dose of fluvalinate. The sample honeybees were obtained 1 day before fluvalinate treatment began and 10 days, 20 days, and 30 days later, with 20 adult worker bees gathered from each colony for each sampling. Afterward, honeybee brains were dissected under a microscope, and bee brains from the same colony were pooled for total RNA extraction and cDNA synthesis. Total RNA was isolated from A. mellifera ligustica brain tissue samples according to the manufacturer’s instructions using TRIzol reagent (Invitrogen, Carlsbad, CA, United States). The quantity and quality of RNA were determined using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, United States) and sequenced using the Illumina HiSeq TM 4000 system’s standard protocol.
The transcriptome sequencing data used in this study were identical to those used in our prior work, as were the quality control data (Fischer and Raabe, 2018; El Agrebi et al., 2019). Bowtie2 (version 2.3.5.1) (Hansen et al., 2013) was used to align and exclude ribosomal reads, and TopHat2 (version 2.0.3.12) (He et al., 2017) was used to align the remaining reads to the honeybee reference genome GCF-003254395.2. Then, to obtain anchor reads, we extracted unmapped reads and intercepted both ends of unmapped reads (default 20 bp). The anchor reads were matched once again with the reference genome GCF-003254395.2, and the alignments were concatenated for circRNA identification.
circRNA identification was performed using find_circ (Hung et al., 2018). The identified circRNAs were aligned to the circBase (Johnson, 2015) database using BLAST, and circRNAs with the threshold of E < e−10 were identified as highly credible annotated circRNAs, otherwise new predicted circRNAs (novel_circRNAs). DESeq2 (Kanehisa and Goto, 2000) was used to detect DECs with an FDR of 0.05 and |log2(FC)| > 1.
cDNA was synthesized using the Evo M-MLV Plus cDNA synthesis kit. All the eight DECs were selected for junction region and qRT-PCR verification with their sequence-specific primers (Table 1). The qRT-PCR was performed using the SYBR® Green Pro Taq HS Master Mix qPCR Kit II (Accurate Biology). qPCR reactions were performed in two steps on the Bio-Rad CFX96 Real-Time Detection System (Bio-Rad Laboratories, Inc., United States): 95°C for 30 s; 95°C for 5 s, and 60°C for 30 s, 40 cycles; and adding a dissolution curve of 95°C for 1 min, 62°C for 30 s, and 95°C for 30 s. The 2−ΔΔCT method was used to calculate the relative expression levels of circRNAs. A. mellifera ligustica AmAct (Kim et al., 2013) was used as a control. Three biological replicates were used for each group of samples, and each biological replicate was tested with three technical replicates.
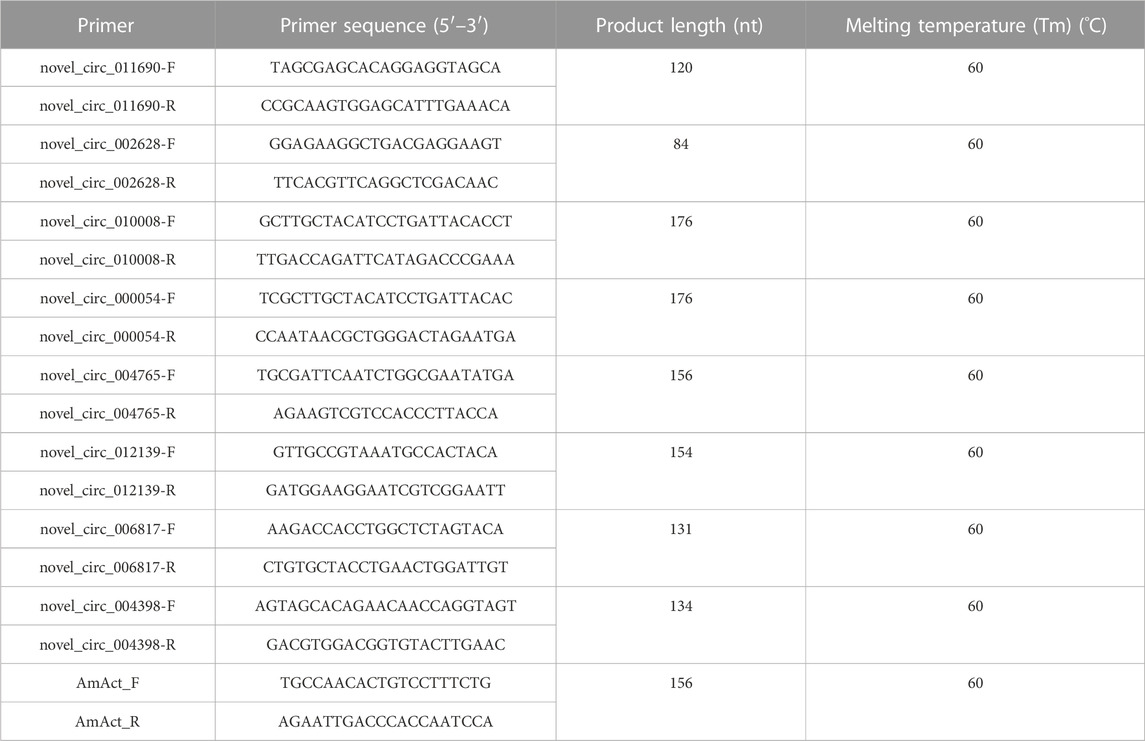
TABLE 1. Primer information for circRNA loop junction site verification and qRT-PCR testing.
Pearson’s correlation analysis was performed on differentially expressed circRNAs and differentially expressed mRNAs (with |r| > 0.7 and significant p < 0.05 as thresholds). The expression profiles of differentially expressed mRNAs were obtained from our previous research (Fischer and Raabe, 2018). A hierarchical clustering was performed with the correlation r-values using MultiExperiment Viewer (MeV) (Kozomara et al., 2019). GO and KEGG (Krüger and Rehmsmeier, 2006) enrichment analyses were performed on mRNAs associated with differentially expressed circRNAs using clusterProfiler (Langmead and Salzberg, 2012), and FDR <0.05 was used as a significant threshold.
The potential targeting relationships of miRNA–mRNA and miRNA–circRNA were predicted using RNAhybrid (version 2.1.2) (Lee et al., 2013), Miranda (version 3.3a), and TargetScan (version 7.0) (Lewis et al., 2005; Love et al., 2014), and the intersection of the three software results was taken as a trusted targeting relationship. We also performed Pearson’s correlation analysis between miRNAs and mRNAs with |r| > 0.7 and p < 0.05 as thresholds. Finally, the intersection of correlation analysis and targeting relationship results was considered potential regulatory relationships and was used for the construction of the ceRNA network using Cytoscape (version 3.8.0) (Marco and Griffiths-Jones, 2012).
Similar to our previous research, 956,250,750 clean reads were obtained from the brain tissue transcriptome sequencing of A. mellifera ligustica after treatment with fluvalinate (Fischer and Raabe, 2018).
The high-quality clean reads were aligned to the ribosome database, and 719,788,610 ribosome unaligned reads were obtained. The ribosome unaligned reads were then mapped with the A. mellifera ligustica reference genome GCF-003254395.2, and 65,829,717 unmapped reads were used for anchor read construction. The anchor reads were compared to the genome GCF-003254395.2 again, and a total of 63,050,916 mapped anchor reads were obtained for further circRNA identification.
A total of 10,780 newly discovered circRNAs were identified, of which 10,764 circRNAs were annotated on 16 known chromosomes in the A. mellifera ligustica genome and 16 circRNAs were annotated to unknown chromosomal locations. It was found that 7,192 (66.72%) circRNAs were composed entirely of annotated exons, 424 (3.93%) circRNAs were composed of introns, 1,262 (11.71%) circRNAs were composed of both exons and introns, 371,287 (2.66%) circRNAs were located in intergenic regions, 1,243 (11.53%) circRNAs were composed of single exons, and only one circRNA (novel_circ_004083) belonged to chromosome type unknown.
The expression values of circRNAs were normalized as reads per million mapped reads (RPM).
With an expression fold change >2 and FDR <0.05 as the cutoff value, eight DECs were identified. The results of cluster analysis showed that the expression patterns of DECs could be divided into three categories (Figure 1A): the first category (C1) had three DECs (novel_circ_002628, novel_circ_000054, and novel_circ_010008), the second had two DECs (novel_circ_006817 and novel_circ_012139), and the third had three DECs (novel_circ_004765, novel_circ_004398, and novel_circ_011690). C1 DEC expression decreased immediately after fluvalinate administration and then gradually increased with the passage of time (Figure 1B). C2 DEC expression showed the highest expression value at 20 days after fluvalinate treatment (Figure 1C). C3 DEC expression increased immediately after fluvalinate administration and then gradually decreased to the pre-dose level (Figure 1D).
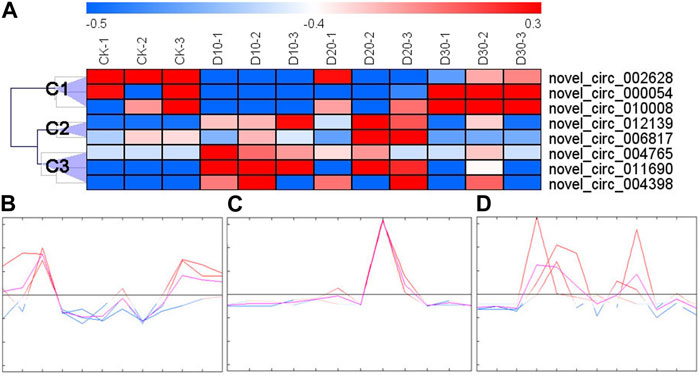
FIGURE 1. Clustering heat map and expression pattern of differentially expressed circRNAs. (A) The clustering heat map of the differentially expressed circRNAs shows that each column represents a sample, and each row represents a differentially expressed circRNA. The z-score is used to normalize the expression level of genes. Red represents higher expression, and blue represents lower expression. (B) C1 circRNA expression pattern, including novel_circ_002628, novel_circ_000054, and novel_circ_010008. (C) C2 circRNA expression pattern, including novel_circ_006817 and novel_circ_012139. (D) C3 circRNA expression pattern, including novel_circ_004765, novel_circ_004398, and novel_circ_011690.
Of the eight DECs, the circular structure and sequence of six DECs were successfully verified (Figure 2A). Furthermore, qRT-PCR results proved that the expression patterns of all six DECs were basically consistent with the transcriptome sequencing results (Figures 2B, C).
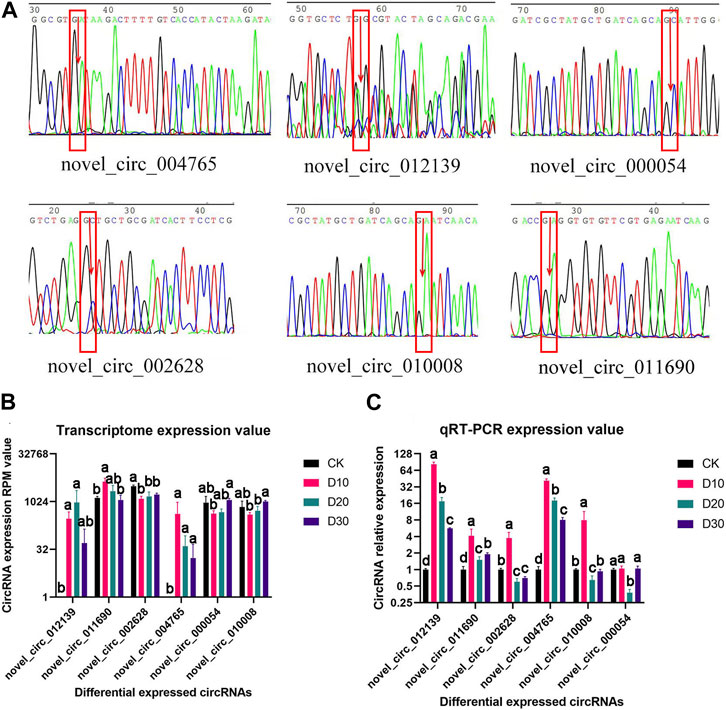
FIGURE 2. Structural verification and real-time quantitative PCR verification of DEC sequence and expression patterns. (A) Sequencing peak map of DECs at loop junction location; the position is marked by the red rectangle, and arrow is the loop junction. (B) Transcriptome expression value of six DECs. (C) qRT-PCR expression value of six DECs. CircRNAs received structural and significant differences which are indicated by different symbols with FDR <0.05 in RNA-Seq results and p < 0.05 in qRT-PCR results.
After taking the intersection of the results of the co-expressed gene–circRNA relationship, miRNA–circRNA targeting relationship, miRNA–mRNA targeting relationship, and miRNA–mRNA expression correlation relationship, a ceRNA network of five circRNAs, 41 genes, and 18 miRNAs was constructed (Figure 3).
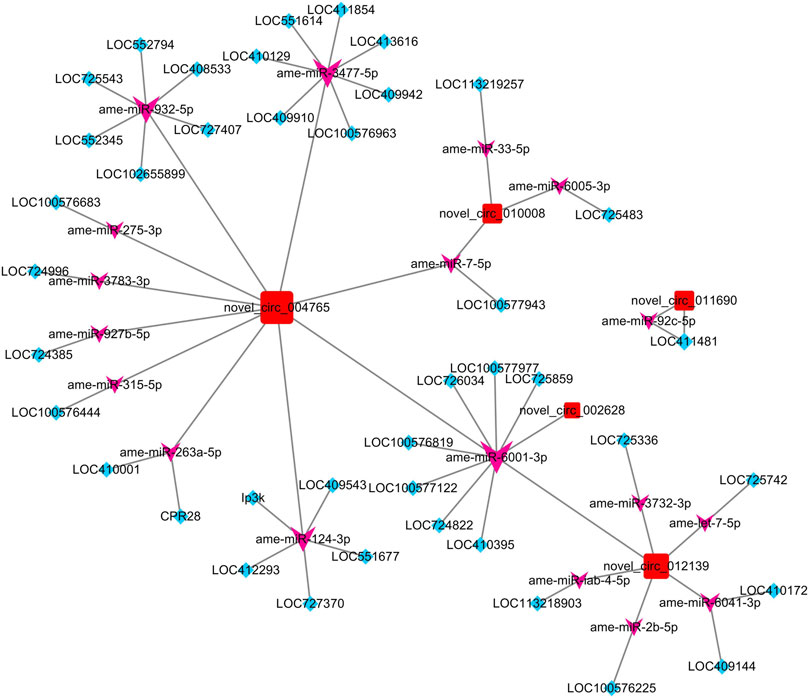
FIGURE 3. ceRNA network constructed by DECs, predicted miRNAs that can be adsorbed by DECs, and predicted targeting genes of miRNAs. The red square nodes represent circRNAs, the purple arrow nodes represent miRNAs, and the blue diamond nodes represent genes.
To confirm the function of five DECs in the network, we performed GO enrichment on genes regulated by each DEC through the ceRNA mechanism (Table 2). The GO enrichment results showed that novel_circ_002628 may be involved in apoptosis and nucleic acid binding-related regulation, and novel_circ_012139 may have the function of regulating ion transmembrane transport. Other circRNAs failed to obtain significant enrichment results.

TABLE 2. Significantly enriched GO terms and KEGG pathways of genes regulated by five differentially expressed circRNAs through the ceRNA mechanism.
To further identify the biological pathways and processes that circRNAs are involved in regulating through the ceRNA mechanism, we also performed KEGG pathway analysis. Of the five DECs, four received significantly enriched pathways, except for novel_circ_010008 (Table 2). Among them, both novel_circ_002628 and novel_circ_012139 regulate LOC100576819 by adsorbing ame-miR-6001-3p, while the gene LOC100576819 was significantly enriched in the apoptosis pathway. novel_circ_011690 regulates LOC411481 by adsorbing ame-miR-92c-5p, while LOC411481 is significantly enriched in the mTOR signaling pathway. novel_circ_004765 received the most significantly enriched pathways and genes. It regulates five protein-coding genes by adsorbing ame-miR-3477-5p, ame-miR-932-5p, ame-miR-6001-3p, and ame-miR-3477-5p.
To construct the ceRNA network, miRNA–circRNA and miRNA–mRNA targeting sites were predicted in this study. From the circRNA–miRNA interactions, we found that various circRNAs interact with different miRNAs. Among them, several circRNAs can bind to multiple miRNAs with different target sites. For example, novel_circ_004765 contains 17 potential binding sites for miRNAs, which may indicate its powerful function as a miRNA sponge. To ensure the sequence accuracy of circRNAs, we performed structural verification of link sites for all differential circRNAs. However, it should be noted that among the six differential circRNAs that were validated, the sequencing electropherogram of novel_circ_012139 always presented an excessive background that may lead to erroneous results.
To explore the functions of DECs, each circRNA in the ceRNA network was subjected to functional enrichment analysis with its related genes. The GO enrichment results showed that the predicted genes regulated by differentially expressed circRNAs are mainly enriched in cell death and ion transmembrane transport-related terms. In addition, the KEGG pathway analysis results showed that apoptosis, Hedgehog signaling pathway-fly, Wnt signaling pathway, and mTOR signaling pathway received significant enrichment.
For novel_circ_002628 and novel_circ_012139, LOC100576819 was the only gene affected by their ceRNA regulation and obtained significant enrichment in the KEGG pathway analysis. In addition, LOC100576819 was also predicted to be regulated by novel_circ_004765. Encoding a homologous protein to that of Drosophila caspase protein (DRONC), the function of LOC100576819 in A. mellifera ligustica has not been reported yet. In Drosophila, DRONC has been confirmed as a major initiator caspase for the programmed cell death of peptidergic neurons (Memczak et al., 2013; Qi et al., 2020) and demonstrates important functions in optic lobe development. Our previous research has proved that the expression level of LOC100576819 in the brain tissue of A. mellifera ligustica increased after the administration of fluvalinate and gradually decreased to the pre-dose level in the following time stages (Fischer and Raabe, 2018). During the ceRNA network, ame-miR-6001-3p was the only miRNA in the ceRNA network, and ame-miR-6001-3p targets novel_circ_002628, novel_circ_012139, and novel_circ_004765 at the same time and is also the only miRNA predicted to target LOC100576819, which may therefore play an important role in this mediating process. However, the detailed functions of miR-6001-3p are still unknown, while the only report of miR-6001 on animals is seen in a study where it was discovered as a newly identified miRNA (Rinderer et al., 1999). Furthermore, although predicted to be regulated by the ceRNA machinery of multiple circRNAs simultaneously, only LOC100576819 showed a similar expression pattern to novel_circ_004765. A possible inference is that novel_circ_004765, as the main ceRNA, regulates the expression of LOC100576819 by competitively adsorbing ame-miR-6001-3p, thereby activating the apoptotic signaling pathway and positive regulation of cell death, but there may be more complex mechanisms affecting the expression of LOC100576819. Overall, the aforementioned results and speculation are subjected to further experimental verification.
In this study, novel_circ_004765 gained the most predicted ceRNA mechanism-regulated genes, targeted miRNAs, and significantly enriched KEGG pathways. Our results showed that novel_circ_004765 may affect the apoptosis pathway by regulating LOC100576819 and LOC408533. The function of gene LOC408533 in A. mellifera ligustica remains unknown. LOC408533 encodes the homologous protein of ASK1 in Drosophila, which is orthologous to several human genes, including MAP3K15 and MAP3K5 (Saeed et al., 2003). In Drosophila, ASK1 has already been reported as a core kinase component of the insulin/insulin-like growth factor pathway, and it also acts as a highly sensitive sensor to activate the JNK and p38 signaling pathways when cells are damaged (Santabárbara-Ruiz et al., 2019). In Caenorhabditis elegans, ASK1 plays an important role in the Sarm1/TIR-1-ASK1/NSY-1-p38 MAPK pathway, which is closely related to the process of inhibition of axonal degeneration activated by CaMKII/UNC-43 (Shannon et al., 2003). In our previous research, CaMKII has been identified as a key gene in the post-transcriptional regulation after fluvalinate administration (Fischer and Raabe, 2018). Therefore, novel_circ_004765 may regulate the expression of LOC408533 (ASK1) gene by competitive adsorption of ame-miR-932-5p, realize the regulation of JNK, p38, and MAPK signaling pathways, and play an important role in the resistance to fluvalinate-induced injury and stress.
Our results also predicted that novel_circ_004765 can regulate genes in the Hedgehog signaling pathway (LOC410129 and LOC409942) and Wnt signaling pathway (LOC410129 and LOC409910) through the ceRNA mechanism. Casein kinase 1 (CK1) and Brother of Ihog (Boi) homologs are encoded by LOC410129 and LOC409942, respectively. CK1 regulates Hh signaling at multiple levels in the Hedgehog (Hh) pathway and is essential for neural function maintenance (Shi et al., 2014; Simon et al., 2021). Boi is part of the Ptc-co-receptor complex for Hh signaling (Tangredi et al., 2012). Its mutations have effects on eye development, neural differentiation, and wing patterning (Thölken et al., 2019). Gene LOC409910 encodes the ras-like GTP-binding protein Rho1 (Rho1), which participates in the axon regeneration of C. elegans and the Drosophila blood–brain barrier formation (Turner, 1985). Interestingly, novel_circ_004765 was predicted to regulate all three genes (LOC410129, LOC409942, and LOC409910) by adsorbing ame-miR-3477-5p. miR-3477 has only been found in three species, including honeybees, jewel wasps, and red flour beetles, and its function is unknown (Van Alphen and Fernhout, 2020). The results of expression correlation analysis and targeting relationship prediction showed that ame-miR-3477-5p can target LOC410129, LOC409942, and LOC409910, which can be used as a clue to reveal the function of miR-3477-5p and its sponge RNA, novel_circ_004765.
novel_circ_011690 was only predicted to adsorb one miRNA (ame-miR-92c-5p) and regulate one gene (LOC411481) through the ceRNA mechanism. The function of miR-92c is still unknown. LOC411481 encodes ribosomal protein S6 kinase alpha-2, RSK2, also known as S6kII. To the best of our knowledge, there is currently no research report on RSK2 and S6kII in honeybees. RSK2 has been shown to be important in the maintenance of normal neural function in Drosophila (Wang et al., 1996; Wieczorek et al., 2020). RSK2 is important in the development of the nervous system, and its absence can cause cognitive and emotional problems (Yu et al., 2012). Our results indicate that novel_circ_011690 may promote LOC411481 expression through competitive adsorption of ame-miR-92c-5p, which plays a role in maintaining the function of brain nerve tissue and resisting fluvalinate poisoning.
In summary, our results revealed the circRNA expression profile changes in the A. mellifera ligustica brains before and after fluvalinate treatment. A total of six circRNAs were identified as DECs and obtained structural and sequence verification, and five of them were predicted to be involved in post-transcriptional regulation of brain tissue through ceRNA regulatory mechanisms as miRNA sponges. Further functional analysis revealed that they may be involved in the regulation of apoptosis in the brain nerves of A. mellifera ligustica by indirectly affecting the genes in the apoptosis, Hedgehog signaling, Wnt signaling, and mTOR signaling pathways. This study improves our understanding of the expression and function of circRNAs in the brain tissue of A. mellifera ligustica and provides information on the role of the ceRNA mechanism in response to fluvalinate stress.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Conceptualization, CT and WG; methodology, CT; resources, SX and FY; data curation, SX; writing—original draft preparation, SX and FY; writing—review and editing, CT and WG; visualization, CT; supervision, CT and WG; project administration, CT and WG; funding acquisition, CT and WG. All authors have read and agreed to the published version of the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the Funds of Shandong Province Agricultural Variety Project (Grant Number: 2016LZGC039) and Shandong “Double Tops” Program (Grant Number SYL2017YSTD12).
The author would like to thank all the members of the Animal Protection Laboratory, School of Animal Science and Technology, Shandong Agricultural University, who collected the whole sample of our experiment. The authors are grateful to the livestock department and friends of apiculture in the sampling area for their help in sample collection on-site/for providing samples.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aizen, M. A., and Harder, L. D. (2009). The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 915–918. doi:10.1016/j.cub.2009.03.071
Andonov, S., Costa, C., Uzunov, A., Bergomi, P., Lourenco, D., and Misztal, I. (2019). Modeling honey yield, defensive and swarming behaviors of Italian honey bees (Apis mellifera ligustica) using linear-threshold approaches. BMC Genet. 20, 78. doi:10.1186/s12863-019-0776-2
Beck, K., Ehmann, N., Andlauer, T. F. M., Ljaschenko, D., Strecker, K., Fischer, M., et al. (2015). Loss of the Coffin-Lowry syndrome-associated gene RSK2 alters ERK activity, synaptic function and axonal transport in Drosophila motoneurons. DMM Dis. Model. Mech. 8, 1389–1400. doi:10.1242/dmm.021246
Berry, J. A., Hood, W. M., Pietravalle, S., and Delaplane, K. S. (2013). Field-level sublethal effects of approved bee hive chemicals on honey bees (Apis mellifera L). PLoS One 8, e76536. doi:10.1371/journal.pone.0076536
Burley, L. M., Fell, R. D., and Saacke, R. G. (2008). Survival of honey bee (Hymenoptera: Apidae) spermatozoa incubated at room temperature from drones exposed to miticides. J. Econ. Entomol. 101, 1081–1087. doi:10.1603/0022-0493(2008)101[1081:SOHBHA]2.0.CO;2
Camp, D., Currie, K., Labbé, A., van Meyel, D. J., and Charron, F. (2010). Ihog and Boi are essential for Hedgehog signaling in Drosophila. Neural Dev. 5, 28. doi:10.1186/1749-8104-5-28
Chao, T., Fan, Y., Lou, D., Xia, H., Ma, L., Shan, X., et al. (2022). Transcriptomic analysis to elucidate the response of Apis mellifera ligustica brain tissue to fluvalinate exposure. Anim. Biotechnol., 1–12. doi:10.1080/10495398.2022.2061506
Chao, T., Yang, L., Lou, D., Fan, Y., He, Y., Shan, X., et al. (2022). Fluvalinate-induced changes in MicroRNA expression profile of Apis mellifera ligustica brain tissue. Front. Genet. 13, 855987. doi:10.3389/fgene.2022.855987
Chen, D., Chen, H., Du, Y., Zhu, Z., Wang, J., Geng, S., et al. (2020). Systematic identification of circular RNAs and corresponding regulatory networks unveil their potential roles in the midguts of eastern honeybee workers. Appl. Microbiol. Biotechnol. 104, 257–276. doi:10.1007/s00253-019-10159-9
Choi, Y. J., Lee, G., and Park, J. H. (2006). Programmed cell death mechanisms of identifiable peptidergic neurons in drosophila melanogaster. Development 133, 2223–2232. doi:10.1242/dev.02376
Davies, T. G. E., Field, L. M., Usherwood, P. N. R., and Williamson, M. S. (2007). DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 59, 151–162. doi:10.1080/15216540701352042
Ding, C., Wu, Y., Dabas, H., and Hammarlund, M. (2022). Activation of the CaMKII-Sarm1-ASK1-p38 MAP kinase pathway protects against axon degeneration caused by loss of mitochondria. Elife 11, e73557. doi:10.7554/eLife.73557
Du, W. W., Zhang, C., Yang, W., Yong, T., Awan, F. M., and Yang, B. B. (2017). Identifying and characterizing circRNA-protein interaction. Theranostics 7, 4183–4191. doi:10.7150/thno.21299
El Agrebi, N., Wilmart, O., Urbain, B., Danneels, E. L., de Graaf, D. C., and Saegerman, C. (2019). Belgian case study on flumethrin residues in beeswax: Possible impact on honeybee and prediction of the maximum daily intake for consumers. Sci. Total Environ. 687, 712–719. doi:10.1016/j.scitotenv.2019.05.493
Fischer, M., and Raabe, T. (2018). Animal models for coffin-lowry syndrome: RSK2 and nervous system dysfunction. Front. Behav. Neurosci. 12, 106. doi:10.3389/fnbeh.2018.00106
Frost, E. H., Shutler, D., and Hillier, N. K. (2013). Effects of fluvalinate on honey bee learning, memory, responsiveness to sucrose, and survival. J. Exp. Biol. 216, 2931–2938. doi:10.1242/jeb.086538
Gan, H., Feng, T., Wu, Y., Liu, C., Xia, Q., and Cheng, T. (2017). Identification of circular RNA in the Bombyx mori silk gland. Insect biochem. Mol. Biol. 89, 97–106. doi:10.1016/j.ibmb.2017.09.003
Glažar, P., Papavasileiou, P., and Rajewsky, N. (2014). CircBase: A database for circular RNAs. RNA 20, 1666–1670. doi:10.1261/rna.043687.113
Gujar, M. R., Stricker, A. M., and Lundquist, E. A. (2019). RHO-1 and the rho GEF RHGF-1 interact with UNC-6/netrin signaling to regulate growth cone protrusion and microtubule organization in caenorhabditis elegans. PLoS Genet. 15, e1007960. doi:10.1371/journal.pgen.1007960
Han, B., Cao, B., Yang, Y., Wang, X., Geng, L., Diao, Q., et al. (2021). Effects of bt Cry78Ba1 toxin on larvae and adults of Apis mellifera (hymenoptera: Apidae). J. Econ. Entomol. 114, 403–408. doi:10.1093/jee/toaa261
Hansen, T. B., Jensen, T. I., Clausen, B. H., Bramsen, J. B., Finsen, B., Damgaard, C. K., et al. (2013). Natural RNA circles function as efficient microRNA sponges. Nature 495, 384–388. doi:10.1038/nature11993
He, Q., Wu, B., Price, J. L., and Zhao, Z. (2017). Circadian rhythm neuropeptides in drosophila: Signals for normal circadian function and circadian neurodegenerative disease. Int. J. Mol. Sci. 18, 886. doi:10.3390/ijms18040886
Hung, K. L. J., Kingston, J. M., Albrecht, M., Holway, D. A., and Kohn, J. R. (2018). The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. B Biol. Sci. 285, 20172140. doi:10.1098/rspb.2017.2140
Johnson, R. M. (2015). Honey bee toxicology. Annu. Rev. Entomol. 60, 415–434. doi:10.1146/annurev-ento-011613-162005
Kanehisa, M., and Goto, S. (2000). Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. doi:10.1093/nar/28.1.27
Kim, D., Pertea, G., Trapnell, C., Pimentel, H., Kelley, R., and Salzberg, S. L. (2013). TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36. doi:10.1186/gb-2013-14-4-r36
Kozomara, A., Birgaoanu, M., and Griffiths-Jones, S. (2019). MiRBase: From microRNA sequences to function. Nucleic Acids Res. 47, D155–D162. doi:10.1093/nar/gky1141
Krüger, J., and Rehmsmeier, M. (2006). RNAhybrid: MicroRNA target prediction easy, fast and flexible. Nucleic Acids Res. 34, W451–W454. doi:10.1093/nar/gkl243
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi:10.1038/nmeth.1923
Lee, G. G., Kikuno, K., Nair, S., and Park, J. H. (2013). Mechanisms of postecdysis-associated programmed cell death of peptidergic neurons in drosophila melanogaster. J. Comp. Neurol. 521, 3972–3991. doi:10.1002/cne.23387
Lewis, B. P., Burge, C. B., and Bartel, D. P. (2005). Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 120, 15–20. doi:10.1016/j.cell.2004.12.035
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi:10.1186/s13059-014-0550-8
Marco, A., and Griffiths-Jones, S. (2012). Detection of microRNAs in color space. Bioinformatics 28, 318–323. doi:10.1093/bioinformatics/btr686
Memczak, S., Jens, M., Elefsinioti, A., Torti, F., Krueger, J., Rybak, A., et al. (2013). Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 495, 333–338. doi:10.1038/nature11928
Qi, S., Zhu, L., Wang, D., Wang, C., Chen, X., Xue, X., et al. (2020). Flumethrin at honey-relevant levels induces physiological stresses to honey bee larvae (Apis mellifera L) in vitro. Ecotoxicol. Environ. Saf. 190, 110101. doi:10.1016/j.ecoenv.2019.110101
Rinderer, T. E., De Guzman, L. I., Lancaster, V. A., Delatte, G. T., and Stelzer, J. A. (1999). Varroa in the mating yard: I. The effects of Varroa jacobsoni and Apistan® on drone honey bees. Am. Bee J. 139, 134–139.
Saeed, A. I., Sharov, V., White, J., Li, J., Liang, W., Bhagabati, N., et al. (2003). TM4: A free, open-source system for microarray data management and analysis. Biotechniques 34, 374–378. doi:10.2144/03342mt01
Santabárbara-Ruiz, P., Esteban-Collado, J., Pérez, L., Viola, G., Abril, J. F., Milán, M., et al. (2019). Ask1 and Akt act synergistically to promote ROS-dependent regeneration in Drosophila. PLoS Genet. 15, e1007926. doi:10.1371/journal.pgen.1007926
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi:10.1101/gr.1239303
Shi, Q., Li, S., Li, S., Jiang, A., Chen, Y., and Jiang, J. (2014). Hedgehog-induced phosphorylation by CK1 sustains the activity of Ci/Gli activator. Proc. Natl. Acad. Sci. U. S. A. 111, E5651–E5660. doi:10.1073/pnas.1416652111
Simon, E., Jiménez-Jiménez, C., Seijo-Barandiarán, I., Aguilar, G., Sánchez-Hernández, D., Aguirre-Tamaral, A., et al. (2021). Glypicans define unique roles for the Hedgehog co-receptors Boi and Ihog in cytoneme-mediated gradient formation. Elife 10, e64581. doi:10.7554/ELIFE.64581
Tangredi, M. M., Ng, F. S., and Jackson, F. R. (2012). The C-terminal kinase and ERK-binding domains of Drosophila S6KII (RSK) are required for phosphorylation of the protein and modulation of circadian behavior. J. Biol. Chem. 287, 16748–16758. doi:10.1074/jbc.M111.315929
Thölken, C., Thamm, M., Erbacher, C., and Lechner, M. (2019). Sequence and structural properties of circular RNAs in the brain of nurse and forager honeybees (Apis mellifera). BMC Genomics 20, 88. doi:10.1186/s12864-018-5402-6
Turner, D. A. (1985). Miranda: A non-strict functional language with polymorphic types. Lect. Notes Comput. Sci. Incl. Subser. Lect. Notes Artif. Intell. Lect. Notes Bioinforma., 1–16. doi:10.1007/3-540-15975-4_26
Van Alphen, J. J. M., and Fernhout, B. J. (2020). Natural selection, selective breeding, and the evolution of resistance of honeybees (Apis mellifera) against Varroa. Zool. Lett. 6, 6. doi:10.1186/s40851-020-00158-4
Wang, X. S., Diener, K., Jannuzzi, D., Trollinger, D., Tan, T. H., Lichenstein, H., et al. (1996). Molecular cloning and characterization of a novel protein kinase with a catalytic domain homologous to mitogen-activated protein kinase kinase kinase. J. Biol. Chem. 271, 31607–31611. doi:10.1074/jbc.271.49.31607
Wieczorek, P., Frąckowiak, P., and Obrępalska-Stęplowska, A. (2020). Evaluation of the expression stability of reference genes in Apis mellifera under pyrethroid treatment. Sci. Rep. 10, 16140. doi:10.1038/s41598-020-73125-w
Keywords: Apis mellifera ligustica, circular RNAs, fluvalinate, brain tissue, ceRNA
Citation: Xueqing S, Delong L, Guizhi W, Yunhan F, Liuxu Y and Tianle C (2023) Effect of fluvalinate on the expression profile of circular RNA in brain tissue of Apis mellifera ligustica workers. Front. Genet. 14:1185952. doi: 10.3389/fgene.2023.1185952
Received: 14 March 2023; Accepted: 28 April 2023;
Published: 11 May 2023.
Edited by:
Rajnish Kumar, University of Missouri, United StatesReviewed by:
Suneel Gupta, University of Missouri, United StatesCopyright © 2023 Xueqing, Delong, Guizhi, Yunhan, Liuxu and Tianle. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chao Tianle, Y2hhb3RpYW5sZUBzZGF1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.