
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 05 July 2023
Sec. Genetics of Common and Rare Diseases
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1163162
 Mingchong Liu1Yongheng Wang2
Mingchong Liu1Yongheng Wang2 Wentao Shi2Chensong Yang1Qidong Wang1Jingyao Chen3Jun Li3
Wentao Shi2Chensong Yang1Qidong Wang1Jingyao Chen3Jun Li3 Bingdi Chen3*†
Bingdi Chen3*† Guixin Sun1*†
Guixin Sun1*†Sarcopenia and osteoporosis, two degenerative diseases in older patients, have become severe health problems in aging societies. Muscles and bones, the most important components of the motor system, are derived from mesodermal and ectodermal mesenchymal stem cells. The adjacent anatomical relationship between them provides the basic conditions for mechanical and chemical signals, which may contribute to the co-occurrence of sarcopenia and osteoporosis. Identifying the potential common crosstalk genes between them may provide new insights for preventing and treating their development. In this study, DEG analysis, WGCNA, and machine learning algorithms were used to identify the key crosstalk genes of sarcopenia and osteoporosis; this was then validated using independent datasets and clinical samples. Finally, four crosstalk genes (ARHGEF10, PCDH7, CST6, and ROBO3) were identified, and mRNA expression and protein levels of PCDH7 in clinical samples from patients with sarcopenia, with osteoporosis, and with both sarcopenia and osteoporosis were found to be significantly higher than those from patients without sarcopenia or osteoporosis. PCDH7 seems to be a key gene related to the development of both sarcopenia and osteoporosis.
In recent years, sarcopenia and osteoporosis, two degenerative diseases in the motor system of older adults, have attracted increasing attention from researchers due to increasingly aging societies (Hu et al., 2022; Khosla et al., 2022). For instance, the proportion of the aged population in South Korea was 16.6% in 2021, and this may reach 25.5% after 10 years and 40.1% after 30 years (Kwon et al., 2022). Across the world, the number of individuals aged more than 65 years outnumbered the number of children in 2018, and it is predicted that this number may double in the next 30 years (Mastnak et al., 2022).
Sarcopenia is an aging syndrome characterized by decreased skeletal muscle mass, function, and strength (Kim et al., 2022). The first practical clinical definition and consensus diagnostic criteria for it were developed in 2010 by the European Working Group on Sarcopenia in Older People (EWGSOP), and the consensus was updated in 2019 (EWGSOP2) (Cruz-Jentoft et al., 2010; Cruz-Jentoft et al., 2019). EWGSOP2 developed and emphasized low muscle strength as a key feature of sarcopenia, muscle content as a diagnostic basis, and physical function as an indicator of sarcopenia severity (Spexoto et al., 2022). An observational study conducted in Spain reported that the prevalence of sarcopenia in older people was 28.9% in men and 26.2% in women (Guillamon-Escudero et al., 2022). Sarcopenia may lack specific clinical features; patients with it may present with weakness, difficulty walking, slow gait, and thin and weak limbs (Aoki et al., 2022). Sarcopenia may increase the risk of falls and fractures (de Almeida Marques Bernabe et al., 2022), decrease the quality of daily living activities, and relate to many comorbidities such as cardiac disease (Loh et al., 2022), diabetes (Jun et al., 2022), and cognitive impairment (Noda et al., 2022)—although these symptoms can be easy to ignore (Malmstrom et al., 2016; Mijnarends et al., 2018).
Osteoporosis is a disease characterized by low bone mass and deterioration of the microstructure of bone tissue (Collins et al., 2017). Its diagnosis is mainly dependent on the evaluation of bone mineral density (BMD); patients with a BMD of more than 2.5 standard deviations lower than the average for young adults may be diagnosed with osteoporosis (Kanis et al., 2019). According to a European Union (EU) survey, there may be 22 million women and 5.5 million men in the EU with osteoporosis (Hernlund et al., 2013). Fragility fractures caused by osteoporosis have become one of the most severe and deadly health problems in the world, and the prevention of osteoporotic fractures has become the most significant aspect of managing the condition in patients (Zhang H. et al., 2022). The commonest site of fragility fractures is the spine, and the deadliest is the hip. Patients with fragility hip fractures had an annual mortality rate of more than 20%, even after surgery (Liu M. et al., 2022).
Muscle and bone are the most important components of the motor system, and patients with both muscle and bone disorders may face worse health and a higher risk of falls and fractures (Cevei et al., 2020). Osteosarcopenia was defined for the first time for patients with both sarcopenia and osteoporosis (Polito et al., 2022). A study conducted in China showed that the prevalence of sarcopenia and osteoporosis in men and women over 65 years old was 10.4% and 15.1%, respectively (Wang et al., 2015). Co-occurrence of sarcopenia and osteoporosis may affect patients additively and synergistically (Clynes et al., 2021; Teng et al., 2021) and contribute to a higher risk of falls and dysfunction as well as poorer outcomes after fractures than those with just one of these diseases (Reiss et al., 2019; Okayama et al., 2022).
Muscle and bone are derived from mesodermal and ectodermal mesenchymal stem cells, with the adjacent anatomical relationship between muscle and bone providing the basic conditions for mechanical and chemical signals (Lam and Qin, 2008). The interaction between bone and muscle is mainly achieved through mechanical stimulation and their secretion of bioactive factors (Tagliaferri et al., 2015). In addition, endocrine, disease, genetics, exercise, and other factors also jointly affect bone and muscle phenotypes (Kawao and Kaji, 2015). In the occurrence and development of osteosarcopenia, the communication of mechanical and chemical signals between muscle and bone may be the pathogenesis of osteosarcopenia, resulting in dysfunction of the skeletal muscle system (Shen et al., 2015). Osteoporosis and sarcopenia have common risk factors, and the reasons behind these factors obscure the pathogenesis of osteosarcopenia (Huang et al., 2014). Therefore, it is significant to identify the correlation and co-pathway between sarcopenia and osteoporosis, and to then explore potential treatment that may improve life quality and decrease the risk of adverse events for patients with it.
With the development of microarray technology and high throughput sequencing, more and more data for comparing differentially expressed genes between individuals with different diseases state have become available (Hephzibah Cathryn et al., 2022). Bioinformatic analysis and machine learning algorithms based on these data may provide us with more information about relationships, maps, and expressions of genes and proteins (Cheng et al., 2021). However, few studies have used bioinformatic analysis and machine learning algorithms to explore the common genes that contribute to the co-occurrence of sarcopenia and osteoporosis. In this study, differentially expressed gene (DEG) analysis, weighted gene co-expression network analysis (WGCNA), and two kinds of machine learning algorithms were used to identify the common key crosstalk genes between sarcopenia and osteoporosis. Moreover, experimental validations of expression and protein levels were performed with real clinical samples. The study’s flow chart is in Figure 1.
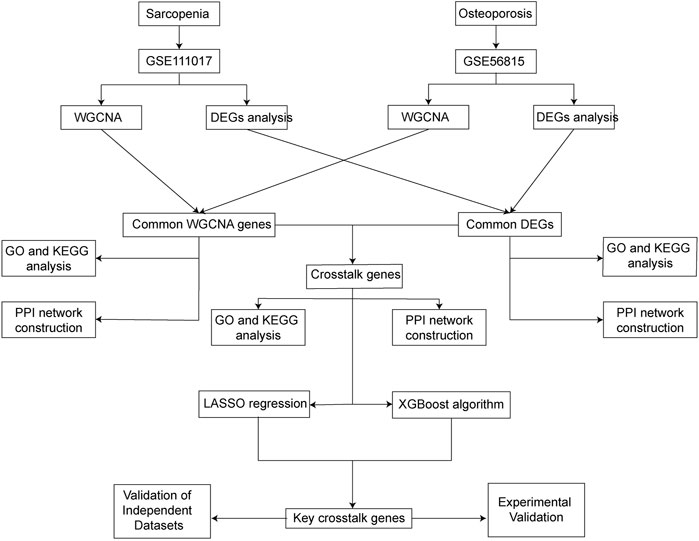
FIGURE 1. Study’s flow chart.
All datasets included in our study were downloaded and collected from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/). GEO is one of the largest public databases to include microarray data and high-throughput gene expression data submitted by research institutions around the world (Patra et al., 2020). To collect the expression data of sarcopenia and osteoporosis, the keywords “osteoporosis” and “sarcopenia” were searched, and then the datasets “GSE111017,” “GSE56815,” “GSE167186,” and “GSE35956” were included in our study. GSE167186 and GSE35956 as the independent datasets were used to validate the key crosstalk genes that we identified.
Sangerbox, an online tool based on R, was used to perform the DEG analysis of sarcopenia and osteoporosis datasets (Weitao Shen et al., 2022). GSE111017 is a profile of high-throughput gene expression, and the DEG analysis for GSE111017 was performed by using the DESeq2 package (version 1.32.0), while the limma package was used to conduct the DEG analysis for GSE56815. In the sarcopenia dataset, genes with p < 0.05 and fold change >1.2 were defined as DEGs; for osteoporosis, genes with p < 0.05 and fold change >1.1 were identified as DEGs. Visualized modules based on the ggplot2 (version 3.3.2) and pheatmap packages (version 1.0.12) in Sangerbox were used to visualize the DEGs.
WGCNA as a novel developed bioinformatic approach aims to find co-expressed gene modules and explore the association between gene networks and phenotypes of interest, as well as a network's core genes (Langfelder and Horvath, 2008). Age and gender are significant factors in the development of sarcopenia and osteoporosis, and the clinical characteristics in datasets GSE111017 and GSE56815 are quite different. Therefore, in our study, WGCNA was performed using the R packages “WGCNA” to identify the sarcopenia and osteoporosis modules and reduce the bias caused by different platforms and databases. First, the outlier sample was identified and eliminated using the Hclust function. Next, soft-thresholding power ranging from 1 to 20 was analyzed and calculated based on the criterion R2 > 0.85. Based on the soft-thresholding power calculated previously, co-expression genes were clustered and modulized and the minimum number of module genes was set at 30. The modules with a correlation of more than 80% in sarcopenia and more than 70% in osteoporosis were merged for the second time. Finally, the module eigengene and the correlation between the module eigengenes and clinical data were visualized, and all the module genes with significant correlations with sarcopenia and osteoporosis were identified and exported for further analysis.
After separately identifying DEGs and WGCNA genes of sarcopenia and osteoporosis, these were imported into R software to obtain the intersection. The intersection of the DEGs of sarcopenia and osteoporosis was identified as the common DEGs, and the common WGCNA genes were defined as the intersection of WGCNA genes of sarcopenia and osteoporosis. Finally, the intersection of common DEGs and common WGCNA were collected as the crosstalk genes of sarcopenia and osteoporosis.
Functional enrichment analysis was performed to explore the biological functions and pathways involved in the genes we identified. Biological process (BP), cellular component (CC), and molecular function (MF) from Gene Ontology (GO) were used to describe gene information, functions, and interaction, and the pathway results were obtained from the Kyoto Encyclopedia of Genes and Genomes (KEGG) databases (Kanehisa et al., 2016; Pomaznoy et al., 2018). R package “enrichplot” was used to perform the GO and KEGG analysis and the results of the enrichment analysis were imported to Cytoscape (version 3.9.1) for visualization.
The protein–protein interaction (PPI) network may reveal the protein interactive relationships. The common DEGs, common WGCNA genes, and crosstalk genes were uploaded to STRING (https://string-db.org/) to construct PPI networks with a confidence of more than 0.7 (Szklarczyk et al., 2019). CytoNSA as a Cytoscape plugin was used to calculate the core protein genes of the PPI network. The size of the nodes in the PPI network was proportional to the significance of genes calculated by CytoNSA.
To better screen the key crosstalk genes of sarcopenia and osteoporosis, the XGBoost algorithm and least absolute shrinkage and selection operator (LASSO) regression were conducted based on R packages “xgboost” and “glmnet.” In the XGBoost analysis, the importance of each gene was viewed and collected. The coefficient of punishment (λ) was identified according to the AUC of the modules with different λ, and then the predictive value of genes was exported from the modules with the most suitable λ. The intersection of genes screened by XGBoost and LASSO regression was identified as the key crosstalk genes.
The expression of key crosstalk genes from GSE167186 and GSE35956 datasets were collected to verify the key crosstalk genes. The t-test was used to compare the expression between groups, and all p-values < 0.05 were considered statistically significant.
All participants were recruited from patients awaiting hip arthroplasty due to hip fractures who were admitted to the Department of Traumatology, Shanghai East Hospital, Tongji University, School of Medicine, Shanghai, China. The study was approved by the Institutional Ethics Committee of East Hospital ([2022] Research Review (No. 278), 7 September 2022) and was performed in accordance with the Declaration of Helsinki. The mean age of patients enrolled in this study was 65 years. Finally, 12 patients who underwent hip arthroplasty were included in our study and divided into four groups according to whether they had sarcopenia and osteoporosis. The diagnosis of sarcopenia and osteoporosis followed the guidelines mentioned previously (Cruz-Jentoft et al., 2019; Zhang H. et al., 2022). Skeletal muscle tissue and bone tissue samples—5 mm × 5 mm × 5 mm each—were collected from the vastus lateralis muscle and femoral head during surgery; due to the nature of joint replacement, this had no impact on the patients. After sample collection, visible fat and blood were removed, and the samples were put into liquid nitrogen and stored at ≤−80°C until analysis.
RNA isolation was conducted following the manufacturer’s instructions for RNAeasy™ Animal RNA Isolation Kit with Spin Column (Beyotime, China). cDNA was synthesized following the manufacturer’s instructions for PrimeScript II 1st Strand cDNA Synthesis Kit (Takara, Japan). RNA quantity and integrity were measured with a NanoDrop 1000 (Thermo Scientific). The reaction system for Real-time Quantitative PCR followed the manufacturer’s instructions for SYBR® Premix Ex Taq™ (Takara, Japan). Data were normalized to GAPDH mRNA and analyzed according to the 2−ΔΔCT method. The gene primers are: forward primer TTGTGGGAGCAGGAGACAAC and reverse primer ACTCTACGAAATGGCTGTTTGC for PCDH7; forward primer GAAAGCCTGCCGGTGACTAA and reverse primer GCCCAATACGACCAAATCAGAG for GAPDH.
Proteins were extracted from tissue homogenate, and the protein concentration was quantified using a BCA kit (Epizyme, China). Total proteins of skeletal muscle and bone were subjected to 12% SDS-PAGE electrophoresis and were then wet migrated to the PVDF membrane. The membrane was sealed with 5% skimmed milk at 37°C for 2 h and then incubated with primary antibody (1:1000; HPA011866, Sigma-Aldrich) at 4°C overnight. The membrane was then washed with TBST three times, 15 min each time, and then cultivated with secondary antibody Goat anti-rabbit IgG with 1:2500 dilution at 25°C for 1 h. Finally, the signal was detected on the Azure imaging biosystem C300 using ECL Kit (Kangweishiji Biotechnology).
Immune infiltration analysis was performed to identify the roles of the immune system in the bone and muscle system. The online platform CIBERSORTx (https://cibersortx.stanford.edu/) was used for immune infiltration analysis and then the immune infiltration matrix generated from CIBERSORTx was imported into R software for visualization using the R package “ggplot2.” The differences between the two groups were compared using the Wilcoxon test and the correlation between the 22 infiltrating immune cells and key genes was then visualized.
Continuous variables were presented as mean ± standard deviation and were evaluated by independent Student’s t-test for normally distributed variables and by Wilcoxon rank-sum test for non-normally distributed data. All p-values < 0.05 were considered statistically significant, and statistical analyses were performed using GraphPad Prism version 8.0.1 (GraphPad Software San Diego, United States) and R software version 4.1.1 (R Foundation for Statistical Computing, Vienna, Austria).
In this study, GSE111017 and GSE167186 were used as the datasets of sarcopenia, and GSE56815 and GSE35956 as the datasets of osteoporosis. GSE111017 was a dataset containing three sub-datasets (GSE111006, GSE111010, and GSE111016) of data from a multi-center study in Hertfordshire, Jamaica, and Singapore. All the sub-datasets in GSE111017 were expression profiled by high-throughput sequencing using the same platforms, and the batch effect between sub-datasets was evaluated. From the evaluation results of the batch effect, there was no significant difference between the datasets (Supplementary Figure S1). GSE167186 contains the data of transcriptome profiling on muscle samples from 72 subjects with or without sarcopenia at different ages. GSE56815 and GSE35956 are the datasets with samples from patients with or without osteoporosis.
Based on the criteria that we set for p-value and fold change of DEGs, 1,426 upregulated DEGs and 470 downregulated DEGs in sarcopenia and 1,482 upregulated DEGs and 879 downregulated DEGs in osteoporosis were identified. As shown in Figures 2A–D, the expression pattern of the DEGs of sarcopenia and osteoporosis were visualized by volcano plots and heatmaps. The DEG overlap was calculated to identify the common DEGs of both diseases (Figures 2E, F). Identified for further analysis were 126 common upregulated genes and 25 common downregulated genes, as summarized in Supplementary Table S1.
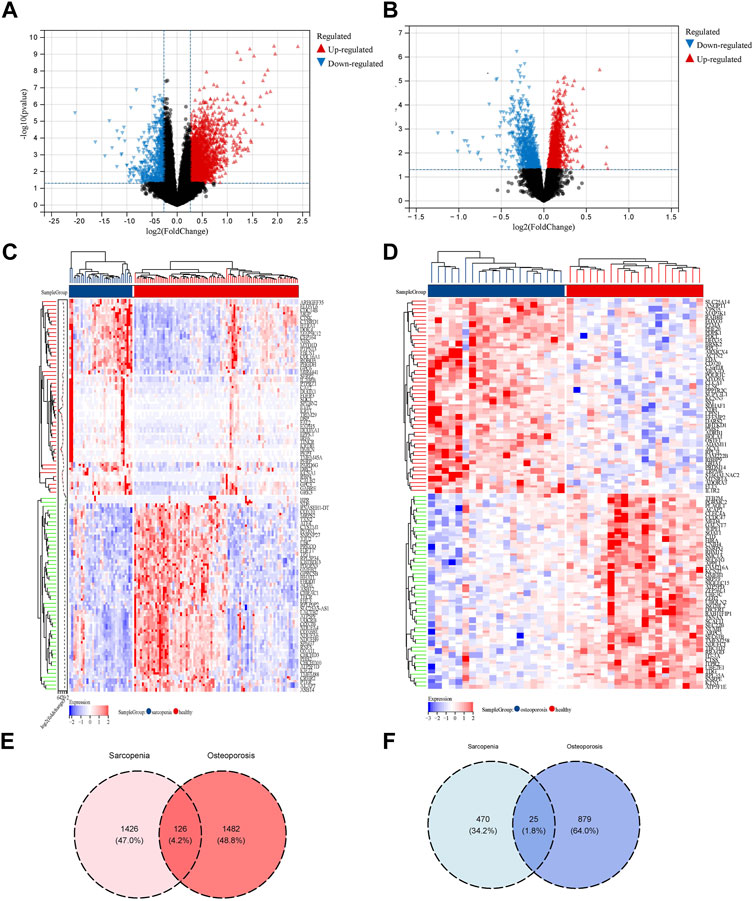
FIGURE 2. Identification of DEGs related to the co-occurrence of sarcopenia and osteoporosis. (A) Volcano plot of DEGs of sarcopenia: blue dots represent downregulated genes in sarcopenia, red dots represent upregulated genes. (B) Volcano plot of DEGs of osteoporosis: the blue dots represent downregulated genes in osteoporosis, red dots represent upregulated genes. (C) Heatmap of top 50 genes in sarcopenia: genes in blue were downregulated in sarcopenia, while those in red were upregulated. (D) Heatmap of top 50 genes in osteoporosis: genes in blue were downregulated in osteoporosis, while those in red were upregulated. (E) Venn diagram of the intersection of upregulated DEGs in sarcopenia and osteoporosis. (F) Venn diagram of the intersection of downregulated DEGs in sarcopenia and osteoporosis.
GO enrichment and KEGG enrichment analysis were conducted to explore the biological features of common DEGs. From the enrichment analysis visualized in Figures 3A–D, many energy-related pathways were enriched in the common DEGs, such as the generation of precursor metabolites and energy derivation by oxidation of organic compounds; energy metabolism is one of the most significant pathways in the research of bone and muscle degeneration (Gomes et al., 2022; Oh et al., 2022). The network of common DEGs and both GO and KEGG terms is shown in Figure 3E; all the common DEGs and the GO and KEGG terms they enriched were connected with the genes to further understand their relationships.
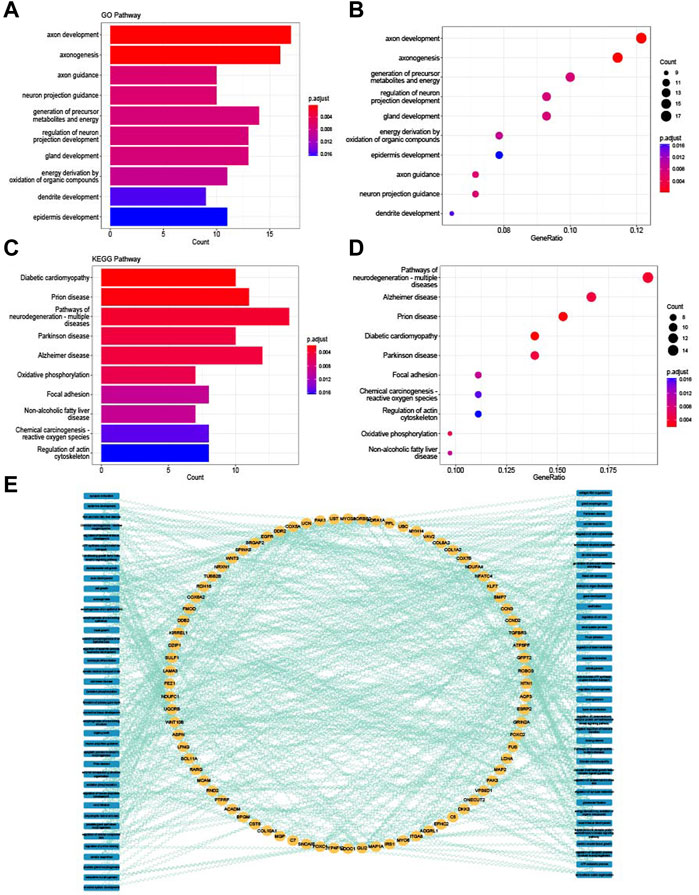
FIGURE 3. Enrichment analysis of common DEGs. (A) Top 10 GO BP terms of common DEGs visualized as bar plot. (B) Top 10 GO BP terms of common DEGs visualized as dot plot. (C) Top 10 KEGG terms of common DEGs visualized as bar plot. (D) Top 10 KEGG terms of common DEGs visualized as dot plot. (E) Network of relationships between common DEGs and GO and KEGG pathways.
WGCNA analysis was performed to identify the gene modules of sarcopenia and osteoporosis. As shown in Figure 4A, the soft threshold was calculated and selected for sarcopenia; a soft threshold of 16 was identified for further analysis due to the cutoff value R2 = 0.90. Figure 4B shows the mean connectivity in different soft thresholds. The gene modules were then established and clustered (Figure 4C), and those with a correlation of more than 80% were merged. The relationships between different modules, as well as between modules and clinical features, are visualized in Figures 4D, E. Modules with a p-value ≤0.01 in the correlation analysis for sarcopenia were selected for further analysis (Figures 4F–I). As with the WGCNA analysis of sarcopenia, the gene modules were constructed and clustered based on the soft threshold of seven (Figures 5A–C); and correlation analysis was then performed (Figures 5D, E). The black, cyan, green, yellow, and blue modules were chosen for further analysis (Figures 5F–I). For sarcopenia, 562 up-related and 8,791 down-related module genes were identified, and 6,895 up-related and 95 down-related module genes for osteoporosis were selected. The intersection of WGCNA modules of sarcopenia and osteoporosis, containing 427 up-related genes and 32 down-related genes, was identified as common WGCNA genes (Figures 6A, B). Lists of genes in different modules of sarcopenia and osteoporosis and the common WGCNA genes are provided in Supplementary Data Sheet S1.
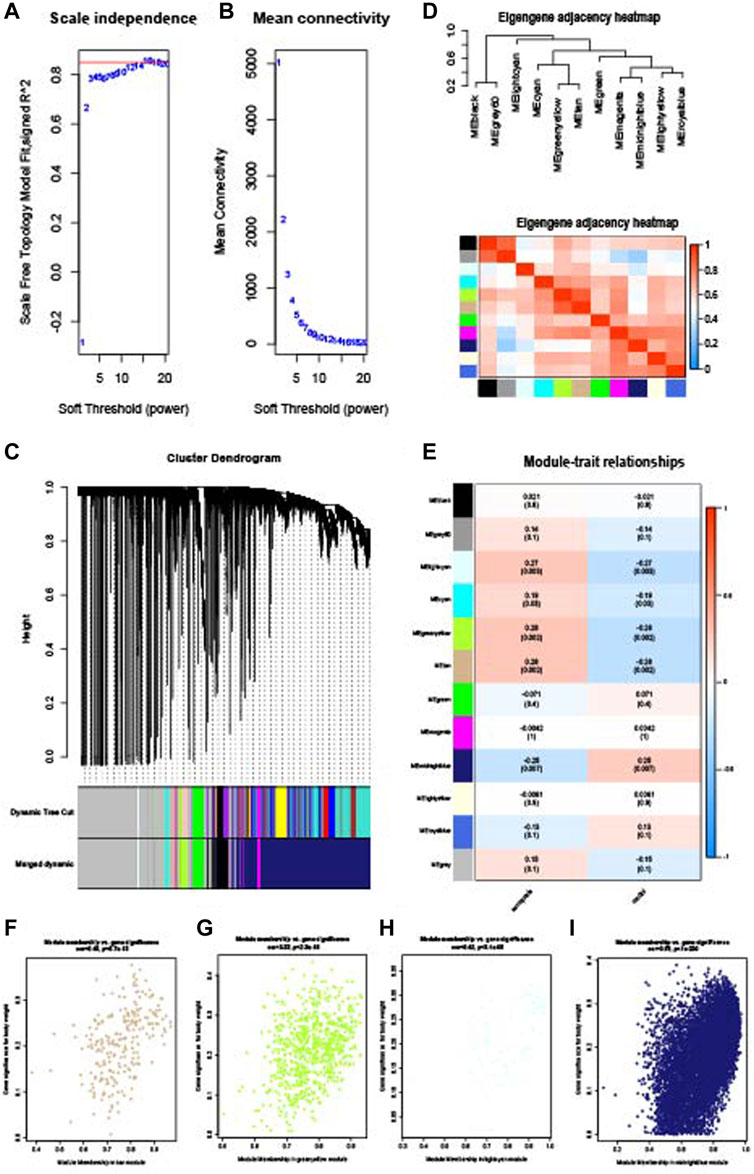
FIGURE 4. WGCNA of the sarcopenia dataset. (A) Selection of soft threshold. (B) Connectivity of different soft thresholds. (C) Cluster dendrogram before and after merge of modules. (D) Correlations of modules. (E) Correlations between modules and sarcopenia. (F) Scatter plots of gene significance in tan module. (G) Scatter plots of gene significance in the green–yellow module. (H) Scatter plots of gene significance in the light cyan module. (I) Scatter plots of gene significance in the midnight blue module.
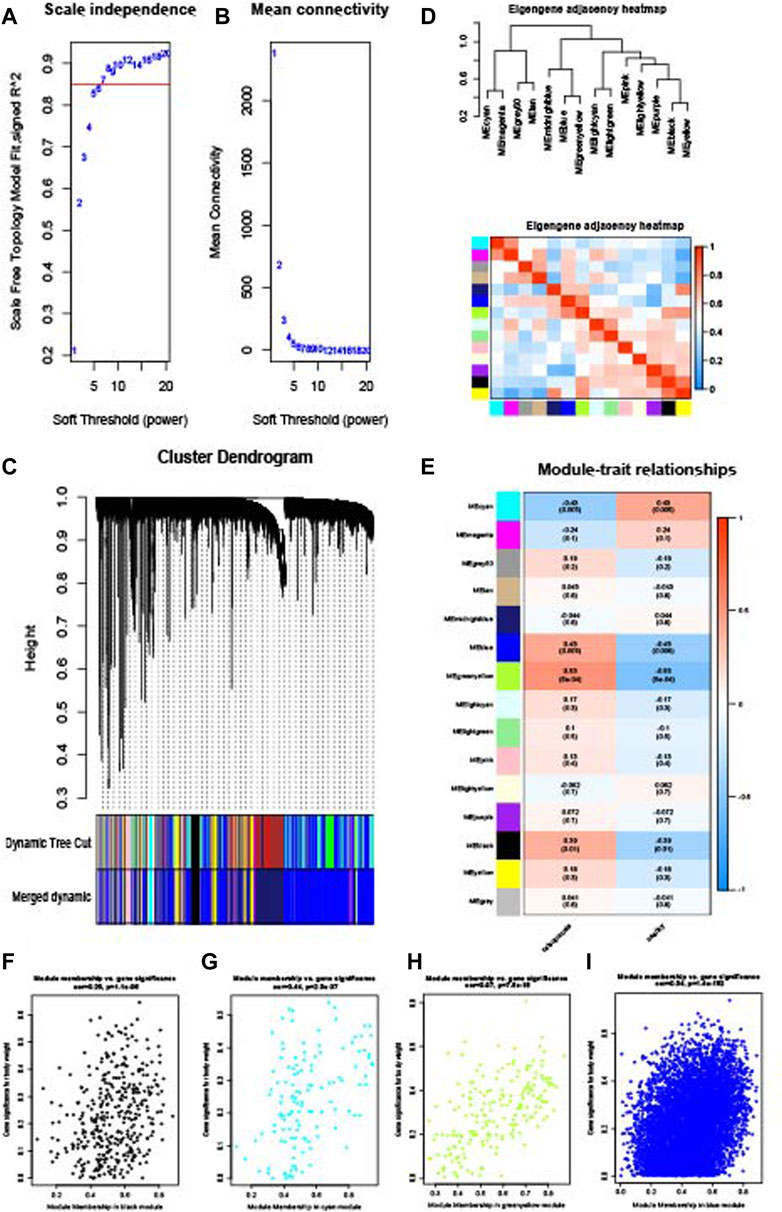
FIGURE 5. WGCNA of the osteoporosis dataset. (A) Selection of soft threshold. (B) Connectivity of different soft thresholds. (C) Cluster dendrogram before and after merge of modules. (D) Correlations of modules. (E) Correlations between modules and osteoporosis. (F) Scatter plots of gene significance in the black module. (G) Scatter plots of gene significance in the cyan module. (H) Scatter plots of gene significance in the green–yellow module. (I) Scatter plots of gene significance in the blue module.
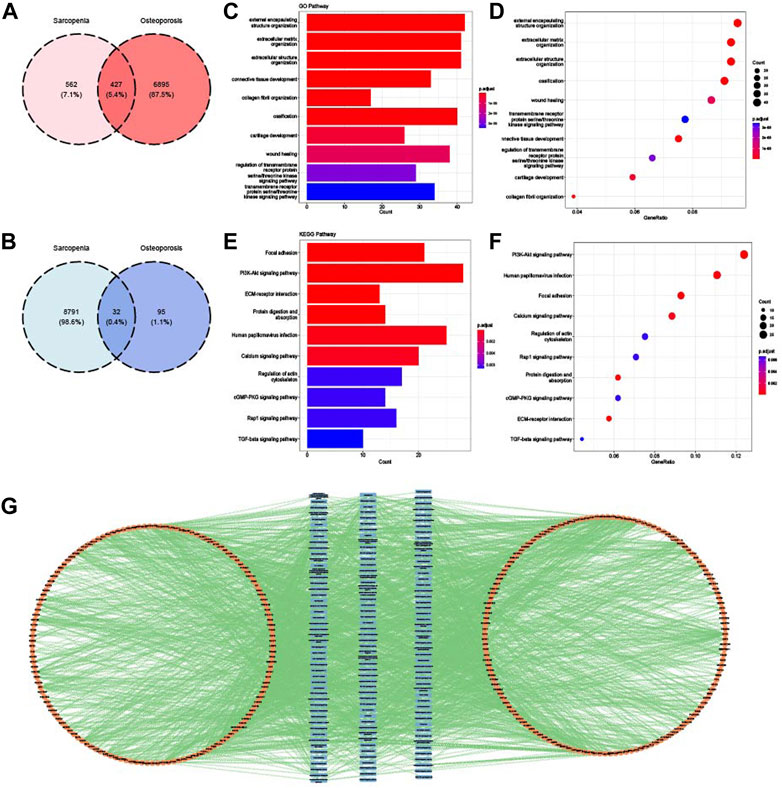
FIGURE 6. Identification and enrichment analysis of common WGCNA genes. (A) Venn diagram of the intersection of upregulated WGCNA module genes in sarcopenia and osteoporosis. (B) Venn diagram of the intersection of downregulated WGCNA module genes in sarcopenia and osteoporosis. (C) Top 10 GO BP terms of common WGCNA genes visualized as bar plot. (D) Top 10 GO BP terms of common WGCNA genes visualized as dot plot. (E) Top 10 KEGG terms of common WGCNA genes visualized as bar plot. (F) Top 10 KEGG terms of common WGCNA genes visualized as dot plot. (G) Network of relationships between common WGCNA genes and GO and KEGG pathways.
Biological features of common WGCNA genes were summarized and analyzed by GO and KEGG analysis. As shown in Figures 6C and D, the main pathways of common WGCNA genes were enriched in connective tissue development and extracellular function in GO analysis. Many muscle and bone metabolism signal pathways were involved for KEGG, such as the PI3K-Akt, cGMP-PKG, and TGF-beta pathways (Figures 6E, F). Moreover, consistent with the enrichment analysis of common DEGs, some energy metabolism pathways were also enriched, such as protein digestion and absorption. The relationships between pathways and genes are mapped in Figure 6G.
Finally, crosstalk genes were identified by intersecting the common DEGs and WGCNA genes. As shown in Figure 7A, 67 upregulated crosstalk genes were identified, and no downregulated genes overlapped. GO and KEGG analysis for 67 crosstalk genes were conducted and visualized in Figures 7B–E. Consistent with the results of enrichment analysis of common DEGs and common WGCNA genes, the crosstalk genes were mainly enriched in extracellular regulation, connective tissue development, vascular endothelial growth factor receptor pathway, and bone mineralization. The relationship map for enrichment analysis of crosstalk genes is shown in Figure 7F.
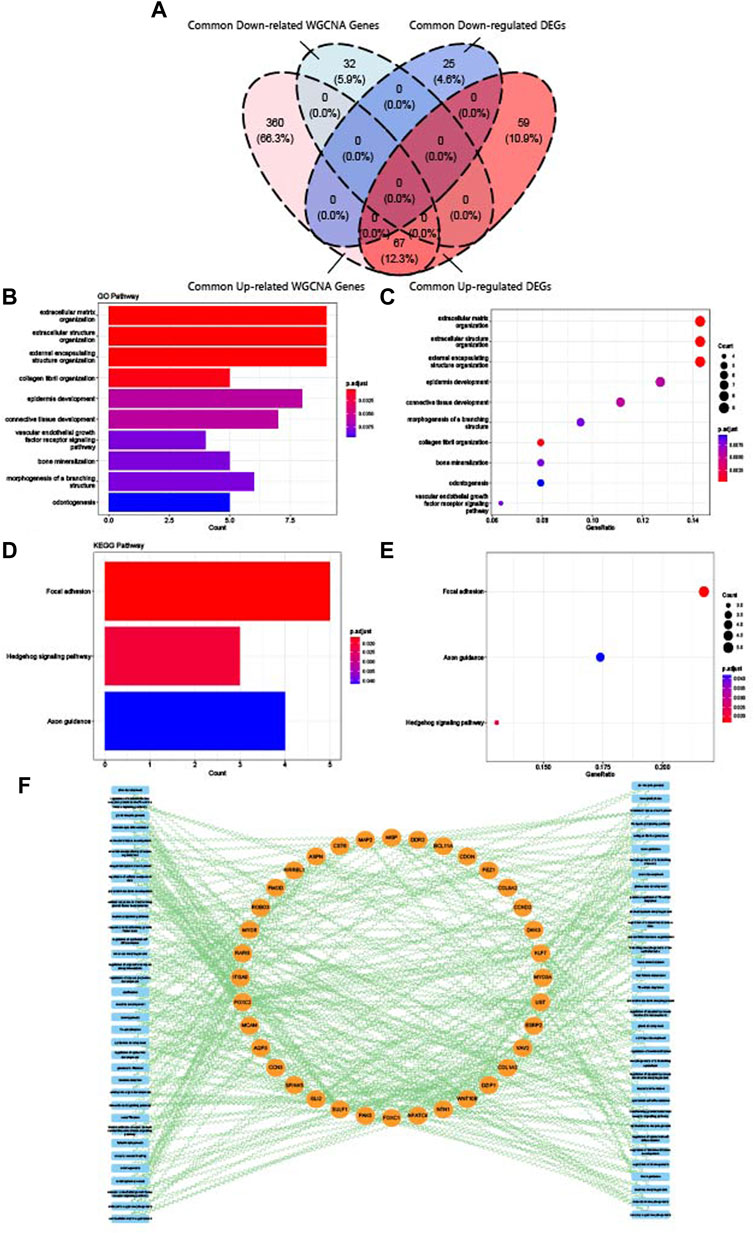
FIGURE 7. Identification and enrichment analysis of crosstalk genes. (A) Venn diagram of the intersection of upregulated common DEGs, downregulated common DEGs, upregulated common WGCNA genes, and downregulated common WGCNA genes. (B) Top 10 GO BP terms of crosstalk genes visualized as bar plot. (C) Top 10 GO BP terms of crosstalk genes visualized as dot plot. (D) Top 3 KEGG terms of crosstalk genes visualized as bar plot. (E) Top three KEGG terms of crosstalk genes visualized as dot plot. (F) Network of relationships between crosstalk genes and GO and KEGG pathways.
The common DEGs, common WGCNA genes, and crosstalk genes were uploaded to STRING (https://string-db.org/) to construct PPI networks and then imported to Cytoscape for visualization (Figures 8A–C). Genes were ranked according to the results of calculation by CytoNSA, with the size presenting the score calculated by the “degree” algorithm. The top five genes in the PPI network of common DEGs were epidermal growth factor receptor (EGFR), collagen type I alpha 1 chain (COL1A1), P21 (RAC1) activated kinase 1 (PAK1), cytochrome C oxidase subunit 5A (COX5A), and cytochrome C oxidase subunit 6A2 (COX6A2); the top five genes in PPI networks of common WGCNA were fibronectin 1 (FN1), COL1A1, collagen type I alpha 2 chain (COL1A2), matrix metallopeptidase 2 (MMP2), and collagen type V alpha 1 chain (COL5A1); in crosstalk, the genes were COL1A1, aggrecan (ACAN), fibromodulin (FMOD), netrin 1 (NTN1), and asporin (ASPN).
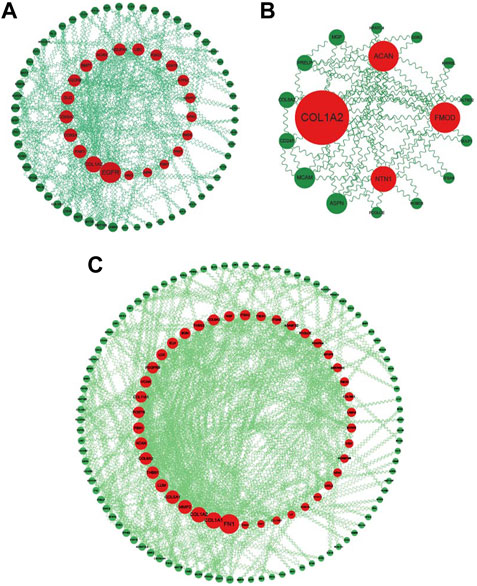
FIGURE 8. PPI networks of common DEGs, WGCNA genes, and crosstalk genes. (A) PPI network of common DEGs: genes ranked by size according to degree score in Cytoscape. (B) PPI network of common crosstalk genes: genes ranked by size according to degree score in Cytoscape. (C) PPI network of common WGCNA genes: genes ranked by size according to degree score in Cytoscape.
Two kinds of machine learning models were established to identify the key crosstalk genes of sarcopenia and osteoporosis. In LASSO analysis, the change trends of gene coefficients in different λ in sarcopenia and osteoporosis are shown in Figures 9A and C: as λ rises, efficiency falls to zero. The AUC of models with different λ is then calculated (Figures 9B, D), and the minimum of λ was selected to establish the models. Some 38 and 31 genes were included in the sarcopenia LASSO model and osteoporosis LASSO model, respectively (Supplementary Table S2). The XGBoost models based on the 67 crosstalk genes were constructed, and the genes’ importance in the models was calculated (Figures 9E, F). In the XGBoost models, 41 genes were included in sarcopenia models, and, in osteoporosis, 17 (Supplementary Table S3). After the intersection of the results of LASSO and XGBoost, four genes were identified as key crosstalk genes (Figure 9G): Rho guanine nucleotide exchange factor 10 (ARHGEF10), Protocadherin 7 (PCDH7), cystatin 6 (CST6), and roundabout guidance receptor 3 (ROBO3).
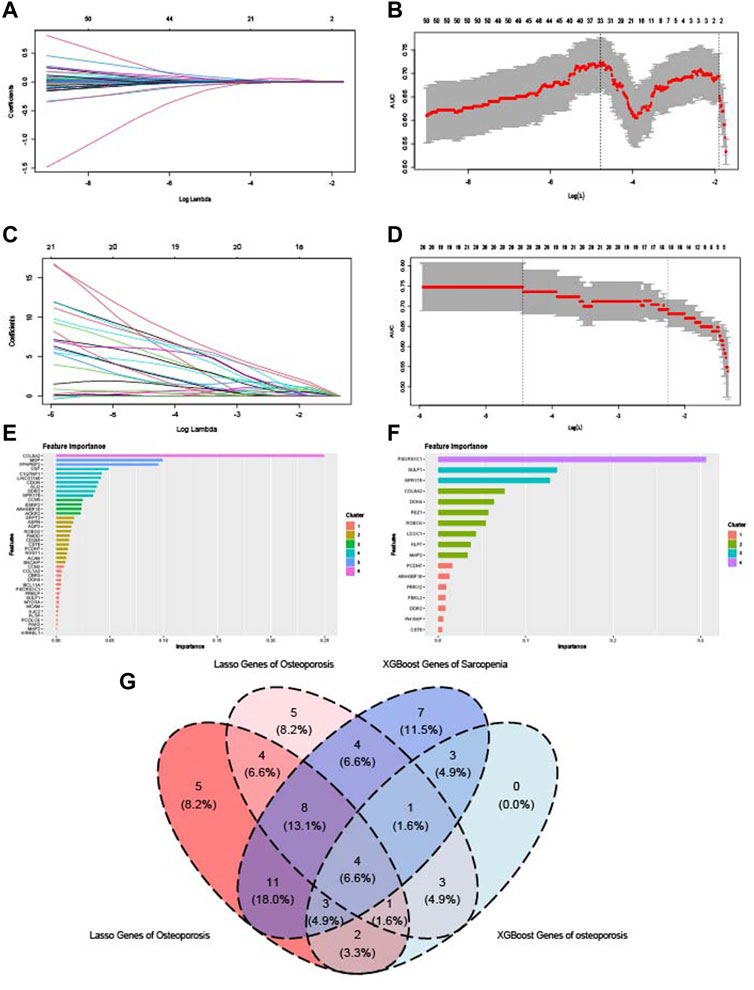
FIGURE 9. Identification of key crosstalk genes by LASSO analysis and XGBoost algorithm. (A,B) LASSO model constructed on the sarcopenia dataset. (C,D) LASSO model constructed on the osteoporosis dataset. (E) Importance of genes calculated by XGBoost algorithm in the sarcopenia dataset. (F) Importance of genes calculated by XGBoost algorithm in osteoporosis dataset. (G) Venn diagram of the intersection of genes with significant importance calculated by LASSO and XGBoost algorithm.
GSE167186 and GSE35956 were used as the datasets of sarcopenia and osteoporosis to validate the key crosstalk genes. The expression level of four key crosstalk genes was compared between patients with and without osteoporosis or sarcopenia. As shown in Figures 10A–H, only PCDH7 was validated in both sarcopenia and osteoporosis test datasets. The patients were then grouped according to whether they had sarcopenia and osteoporosis, with the baseline characteristics of patients summarized in Supplementary Table S4. The mRNA expression and protein level of PCDH7 in muscle and bone tissue between patients in different groups were compared by real-time quantitative PCR and Western blot. Those patients with sarcopenia, osteoporosis, and osteosarcopenia had a significantly higher PCDH7 expression in both mRNA and protein levels than patients without sarcopenia and osteoporosis (Figures 10I–K).
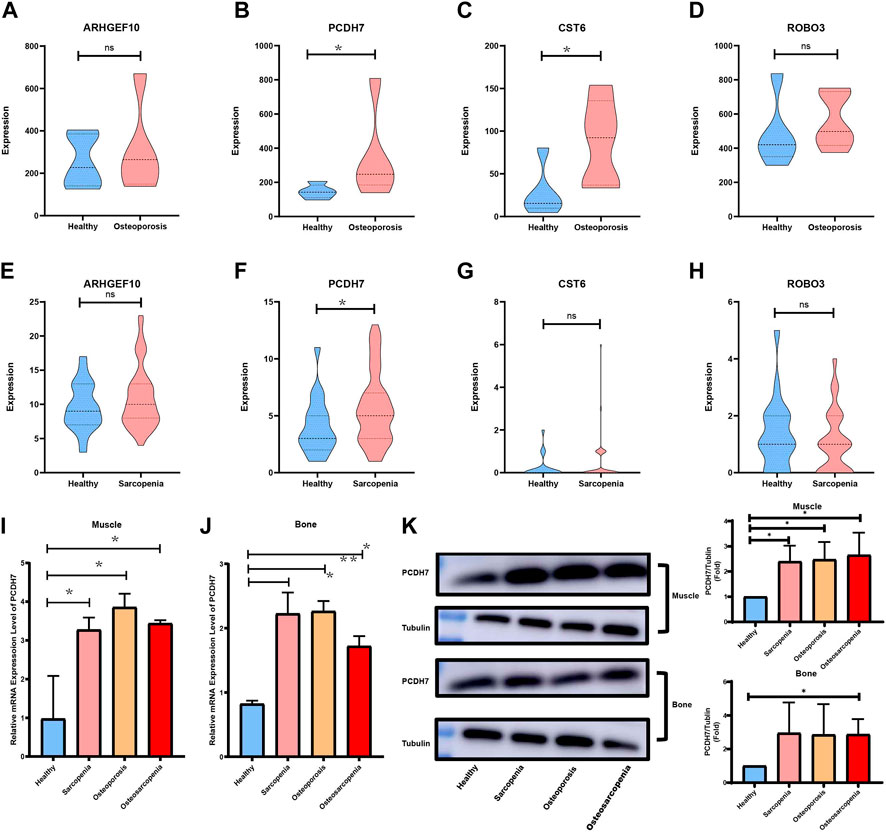
FIGURE 10. Validation of key crosstalk genes in independent datasets and experience. (A) Comparison of mRNA expression levels of ARHGEF10 in the sarcopenia dataset. (B) Comparison of mRNA expression levels of PCDH7 in the sarcopenia dataset. (C) Comparison of mRNA expression levels of CST6 in the sarcopenia dataset. (D) Comparison of mRNA expression levels of ROBO3 in the sarcopenia dataset. (E) Comparison of mRNA expression levels of ARHGEF10 in the osteoporosis dataset. (F) Comparison of mRNA expression levels of PCDH7 in the osteoporosis dataset. (G) Comparison of mRNA expression levels of CST6 in the osteoporosis dataset. (H) Comparison of mRNA expression levels of ROBO3 in the osteoporosis dataset. (I) Comparison of mRNA expression levels of PCDH7 in muscle samples. (J) Comparison of mRNA expression levels of PCDH7 in bone samples. (K) Protein levels of PCDH7 in muscle and bone samples.
To explore the relationships between the immune system and the co-occurrence of sarcopenia and osteoporosis, immune infiltration analysis was performed on the sarcopenia and osteoporosis datasets. Figure 11A shows the proportion of immune cells in different samples of the sarcopenia dataset. The relationships between key crosstalk genes and immune cells is analyzed and visualized in Figure 11B, Plasma cells and macrophages M2 were cells the most relevant to the key crosstalk genes. The relationships between sarcopenia and immune cells, as well as between immune cells, are visualized in Figures 11C and D as a heatmap and correlogram, respectively. Moreover, the levels of each type of immune cell were compared in patients with and without sarcopenia, and the levels of T cells CD4-naïve and monocytes significantly differed between groups (Figure 11E). Similarly, the results of the immune infiltration of osteoporosis are shown in Supplementary Figure S2.
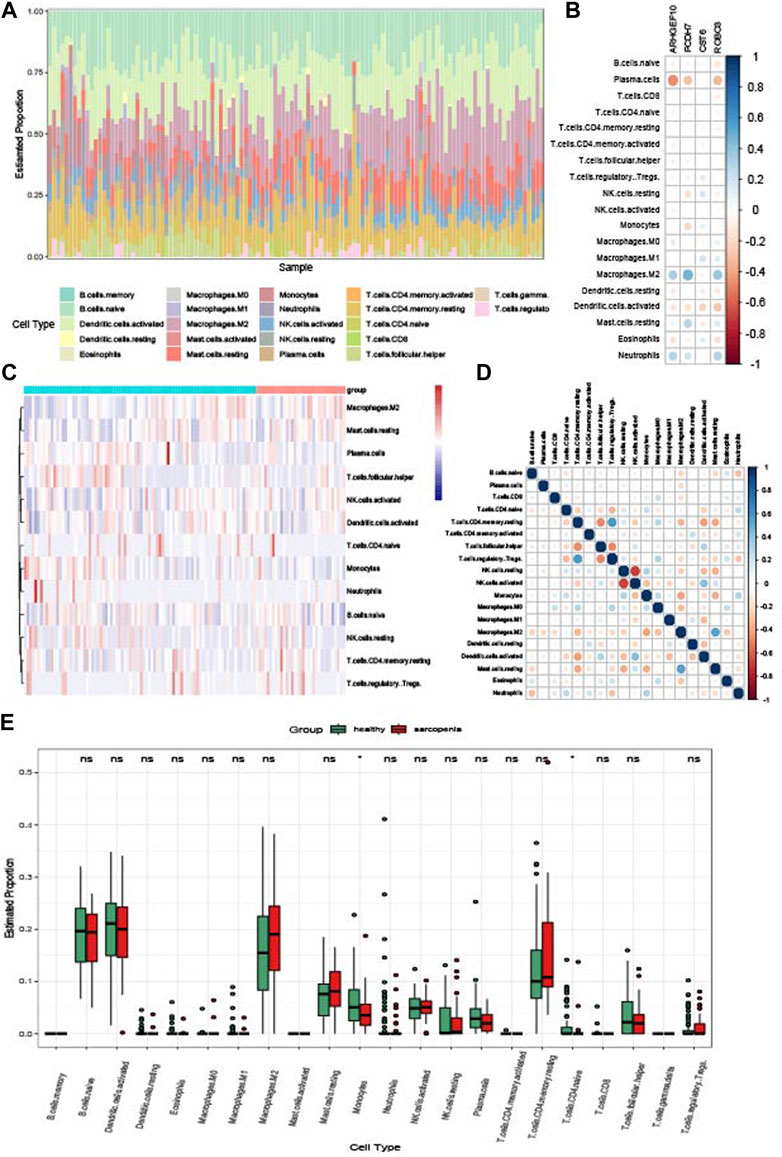
FIGURE 11. Immune infiltration analysis of sarcopenia. (A) Relative percentage of 22 immune cell subpopulations in the sarcopenia dataset. (B) Correlations between immune cell subpopulations and key crosstalk genes. (C) Heatmap of immune cell subpopulations in the sarcopenia dataset. (D) Correlations between immune cell subpopulations. (E) Box plot of immune cell subpopulations in the sarcopenia dataset.
Sarcopenia and osteoporosis as degenerative diseases in the most significant part of the motor system have become severe health problems in aging societies. In this study, datasets of sarcopenia and osteoporosis were included to identify their possible crosstalk genes. The specimens of osteoporosis were blood monocytes and of sarcopenia were muscle biopsies. Monocytes as a type of circulation blood cells have been proven in many studies to relate to sarcopenia and osteoporosis (Deng et al., 2021; Nie et al., 2022; Zeng et al., 2022). Consistent with previous studies used different kind of specimens (Wang Z. et al., 2022; Liu J. et al., 2022; Mo et al., 2022), we have concluded that the intersection between gene expression maps of muscle tissues and monocytes may provide us much wider insights into the development of osteosarcopenia (Liu et al., 2017). In our study, key crosstalk genes (ARHGEF10, PCDH7, CST6, and ROBO3) were identified by DEG analysis, WGCNA, and machine learning methods, and then validated by independent datasets and clinical samples. Finally, PCDH7 was identified and validated as the gene that may be significantly related to the co-occurrence of sarcopenia and osteoporosis.
The pathogenesis behind sarcopenia is still a controversial topic and is hotly debated. It is widely believed that the skeletal muscle mass in humans peaks at age 30 and then decreases significantly with age, while, in individuals aged over 75, skeletal muscle accounts for only a quarter of total body weight (Weitao Shen et al., 2022). Moreover, myodynamia may face a more severe decrease than muscle mass, and it is estimated that muscle strength declines at about two to five times the rate of muscle mass loss (Tournadre et al., 2019). Many studies have demonstrated that the skeletal muscle satellite cells—myogenic stem cells in skeletal muscles—play a significant role in the regeneration and recovery of muscle by regulating their proliferation and differentiation, and that the degeneration in both quantity and quality of satellite cells may relate to the development of sarcopenia (Hirano et al., 2023). Satellite cells from young mice may have a more sensitive and quicker ability to activate their cell cycle of proliferation than those from older mice when subject to external stimuli on a muscle (Leiter et al., 2011). However, the disability of satellite cells may be the result of sarcopenia but not a contributor to its development. Researchers have tried to transplant the muscle samples from cadavers of different ages to immunodeficient mice, and, interestingly, there was no significant difference in myogenic capacity (Novak et al., 2021). Therefore, the development of sarcopenia might not be caused by the degeneration of satellite cells but might be attributable to progressive and detrimental changes in the muscle microenvironment.
Many factors may influence the homeostasis of the muscle microenvironment. Neuromuscular junction as highly specialized chemical synapses formed between motor neurons and skeletal muscle fibers plays an important role in maintaining muscle tonus and preventing muscular atrophy (Cheng et al., 2022). The function of the neuromuscular junction was mainly achieved by regulating the release of acetylcholine (ACh) and the activity of ACh receptors (AChRs); in older adults, agrin, an important factor in Agrin-Lrp4-MuSK signaling pathway regulating AChR, may face a structural abnormality and then contribute to the scrambled AChR localization and dysfunctional neuromuscular junction (Butikofer et al., 2011; Gros et al., 2022). Moreover, α-synuclein, a protein localized in motor neuron axons and neuromuscular junction, may also increase myofiber apoptosis by inhibiting the release of Ach and aggregating oxidative stress (Yang et al., 2022). Many researchers believe that mitochondrial dysfunction may be another factor in the development of sarcopenia. Within aging bodies, severe mitochondrial oxidative injury may cause dysfunction of the calcium signaling pathway and then induce sarcopenia (Cheema et al., 2015). Many downstream pathways and factors relating to muscle regeneration may be regulated and activated by mitochondria, including the mammalian target of rapamycin complex 1 (mTORC1), Kelch-like ECH-associated protein 1 (KEAP1), and NF-E2-related factor 2 (NRF2) (Rai and Dey, 2020). The mammalian target of rapamycin (mTOR) has been proven to be the key regulator of muscle regeneration and to regulate bone metabolism by stimulating protein synthesis and inhibiting proteolysis (Sirago et al., 2022). Inflammaging, a chronic inflammatory phenomenon occurring in older individuals and characterized by an increase in proinflammatory cytokines and a decrease in anti-inflammatory cytokine levels, was also a cause of many geriatric syndromes, including sarcopenia (Bencivenga et al., 2022). With inflammation, the excessive release of inflammatory factors, such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and C-reaction protein (CRP), may influence the capacity of satellite cells and contribute to the disorder of the muscle microenvironment (Savary-Auzeloux et al., 2022). TNF-α may induce the overexpression of muscle atrophy-related genes, such as MuRF-1 and MAFbx, by regulating the TNF receptor-associated factor 6 (TRAF6) and activating the nuclear factor-kappa B (NF-κB) signaling pathway (Li et al., 2020). CRP may negatively affect muscle cell size directly by reducing the phosphorylation of protein kinase B (AKT) and ribosome protein subunit 6 kinase 1 (S6K1) (Oflazoglu et al., 2022).
Osteoporosis, one of the most common geriatric syndromes of older adults and postmenopausal women, has also attracted increasing attention from researchers around the world. It was widely believed that the imbalance between osteoclasts and osteoblasts contributed significantly to the development of osteoporosis, and may affect bone metabolism by regulating the activity of osteoclasts and osteoblasts (Yan et al., 2022). Many signaling pathways involved in osteoporosis have been explored, including OPG/RANKL/RANK, Wnt/β-catenin, PI3K/Akt/mTOR, MAPK, and Hedgehog (Patel and Wairkar, 2022). OPG/RANKL/RANK is one of the most significant signaling pathways in the development of osteoporosis: osteoprotegerin (OPG), a factor secreted by osteoblasts, and a receptor activator of NF-κB ligand (RANKL) competitively bind to the receptor activator of NF-κB (RANK) (Wang F. et al., 2022). Many factors may affect bone metabolism by regulating OPG/RANKL/RANK, including parathyroid hormone (PTH), vitamin D, prostaglandin, and estrogen (Chen et al., 2018). Wnt/β-catenin is a key pathway involved in the osteogenic differentiation of mesenchymal stem cells (Marini et al., 2022). It has been found that the Wnt/β⁃catenin signal directly induces BMP-2 expression and enhanced BMP-2 transcription activity in osteoblasts via the TCF/LEF reaction element, and that β-catenin can enhance the sensitivity of mesenchymal stem cells to BMP-2, inducing its differentiation into osteoblasts and promoting its formation (Huybrechts et al., 2020; Xie et al., 2021). The PI3K/Akt signaling pathway is mediated by enzyme-linked receptors (Yang et al., 2022). PI3K/Akt can activate the signaling pathway with the participation of various growth factors, cytokines, and extracellular matrix and also plays an important role in cell proliferation, apoptosis, tissue inflammation, and tumor growth and invasion (Wang Q. et al., 2022; Hashemi et al., 2022; Quan et al., 2022). Insulin-like growth factor-1(IGF-1) as well as vascular endothelial growth factor (VEGF) may promote the differentiation of bone marrow mesenchymal stem cells into osteoblasts by regulating the PI3K/Akt/mTOR signaling pathway (Karar and Maity, 2011; Bakker et al., 2016).
The interaction between bone metabolism and the immune system was also an interesting topic of osteoporosis (Terashima and Takayanagi, 2018); we therefore conducted immune infiltration to explore their relationships. In previous studies, OPG/RANKL/RANK had been reported to have an important role in osteoimmunology due to its impact on the differentiation from monocytes to osteoclasts (Okamoto and Takayanagi, 2019). Many cells involved in the immune function express RANK and RANKL, and many factors may play a role in both immune activities and bone metabolism. M1/M2 dichotomy is a commonly known immune cell due to its impact on the bone microenvironment, and, with a decreased of estrogen level and increased RANKL level, the M1/M2 ratio was influenced (Yang et al., 2019). Similarly, immune cytokines including TNF-α, IFN-γ, IL-1, IL-4, IL-6, and IL-17, may also affect the skeletal system (Zhou et al., 2021).
Both bone and muscle are derived from the mesenchymal stem cells of the mesoderm and ectoderm; the anatomical relationships between them provide more possible interactions of mechanical and chemical signals (Brotto and Bonewald, 2015). Decreased muscle mass may promote the loss of bone mass, while the muscle contractions may partially inhibit bone loss and trabecular structural degradation due to a lack of daily weight-bearing activities (Lam and Qin, 2008; Manske et al., 2011). Myostatin, a member of the TGF-β superfamily secreted by skeletal muscle cells, also inhibits the increase of muscle and bone mass (Buehring and Binkley, 2013). Similarly, Indian hedgehog (Ihh), a factor secreted by osteoblast, may regulate muscle function and mass (Bren-Mattison et al., 2011). Moreover, the gap junction protein connexin 43 also affects both bone and muscle: the connexin 43 gene ablation mouse may have lower muscle weight and strength and defective skeletal development (Shen et al., 2015).
Extracellular matrix organization and structure, connective tissue development, VEGF signaling pathway, focal adhesion, and hedgehog signaling pathway, as we showed in the enrichment analysis of crosstalk genes, are all pathways involved in the regulation of the connective tissue microenvironment. Consistent with the mechanism of bone and muscle mentioned previously, the signaling pathway between bone and muscle plays an important role in the development of musculoskeletal disorders. Therefore, there must be some genes that regulate both bone and muscle development, as well as the interaction between them, and contribute to the co-occurrence of sarcopenia and osteoporosis.
By using the bioinformatic methods, this study identified PCDH7 as the crosstalk gene that bridges sarcopenia and osteoporosis—validated by real-time quantitative PCR and Western blot on clinical samples. Cadherin and its related proteins constitute a cadherin superfamily, a group of calcium-dependent transmembrane glycoproteins that mediate cell–cell adhesion and bonding in tissues (Hatta and Takeichi, 1986). The cadherin superfamily is roughly divided into four categories: classical cadherins, pontoon cadherins, protocadherins (PCDHs), and other types of cadherin-related proteins (Shirayoshi et al., 1986). PCDH7 is a member of the PCDH family, the largest subfamily of cadherin, and belongs to the PCDHδ1 family (Yoshida et al., 1999). The extracellular domain of PCDH7 consists of seven repeats of the cadherin motif (EC1–7), and the EC2 of PCDH7 is unique due to its 55-amino-acid insertion into the middle of the motif (Yoshida et al., 1998). PCDH7 plays a role in many cancers, including lung adenocarcinoma (Ma et al., 2022) and gastric cancer (Xin et al., 2022), colon (Liu Z. et al., 2022), ovarian (Salgado-Albarran et al., 2022), and breast cancers (Zhang X. et al., 2022). Moreover, many studies have reported its impact on physiological processes, such as dendritic spine morphology, synaptic function, and the formation of homotypic cell-in-cell structures (Wang C. et al., 2020; Wang Y. et al., 2020).
The relationships between PCDH7 and sarcopenia, as well as osteoporosis, are not fully understood. Several studies have reported that PCDH7 may affect bone metabolism by regulating osteoclast differentiation: the bone marrow-derived monocytes retrovirally transduced with shRNA for PCDH7 may face significantly reduced osteoclast maturation; similarly, PCDH7-deficient mice had impaired osteoclast differentiation but increased bone mass and normal osteoblast function (Kim et al., 2020). However, few studies have reported whether PCDH7 plays a role in muscle development and atrophy, as well as whether PCDH7 contributes to the development of osteoporosis.
Three other key crosstalk genes were also identified: ARHGEF10, CST6, and ROBO3, which may contribute to bone and muscle metabolism. ARHGEF10 regulates the function of vascular endothelial cells by contributing to cell junction disruption and reorientation (Abiko et al., 2015; Khan et al., 2021), and vascular function plays significant role in bone regeneration after damage (Gautvik et al., 2022; Qin et al., 2022). CST6, an extracellular polypeptide inhibitor of cysteine proteases, also affects bone metastasis by regulating osteoclastogenesis in many cancers, such as breast cancer and multiple myeloma (Li et al., 2021; Gai et al., 2022). ROBO3 is a member of the ROBO protein family, a subfamily of the immunoglobulin transmembrane receptor superfamily (Pak et al., 2020). ROBO proteins affect bone metabolism through Slit-ROBO signaling and neural epidermal growth factor-like proteins (Niimi, 2021), as well as muscle patterning and vascular injury regulation (Long and Van Vactor, 2012; Yuen and Robinson, 2013; Ordan and Volk, 2015).
After validation by independent datasets, only PCDH7 was selected for further validation due to its significantly different expression level in test datasets. To prove the relationship between PCDH7 and osteosarcopenia, we collected clinical samples of patients with sarcopenia, osteoporosis, both sarcopenia and osteoporosis, and patients without sarcopenia or osteoporosis. Gene expression and protein levels were detected and compared; consistent with the results, patients with sarcopenia, osteoporosis, or both have a significantly higher level of PCDH7 than healthy adults. This suggests that PCDH7 must play a role in the development of both sarcopenia and osteoporosis—but how? More research is needed to explore the mechanism behind them and to provide stronger evidence.
Of four key crosstalk genes (ARHGEF10, PCDH7, CST6, and ROBO3), PCDH7 was identified and validated as relating the development of both sarcopenia and osteoporosis. This may provide new insights for treating older patients with sarcopenia and osteoporosis.
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
The studies involving human participants were reviewed and approved by the Institutional Ethics Committee of East Hospital. The patients/participants provided their written informed consent to participate in this study.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This work was supported by the National Natural Science Foundation of China (Grant/Award Number: 81971169) and Leading Talents Training Program of Pudong New Area Health Commission (Grant ID: PWR 12020-06).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2023.1163162/full#supplementary-material
Abiko, H., Fujiwara, S., Ohashi, K., Hiatari, R., Mashiko, T., Sakamoto, N., et al. (2015). Rho guanine nucleotide exchange factors involved in cyclic-stretch-induced reorientation of vascular endothelial cells. J. Cell. Sci. 128 (9), 1683–1695. doi:10.1242/jcs.157503
Aoki, Y., Furukawa, K., Suzuki, D., Takayashiki, T., Kuboki, S., Takano, S., et al. (2022). Influence of sarcopenia as defined by EWGSOP2 on complications after pancreaticoduodenectomy and on the prognosis of pancreatic head cancer: A prospective cohort study. Nutrition 99-100, 111660. doi:10.1016/j.nut.2022.111660
Bakker, A. D., Gakes, T., Hogervorst, J. M., de Wit, G. M., Klein-Nulend, J., and Jaspers, R. T. (2016). Mechanical stimulation and IGF-1 enhance mRNA translation rate in osteoblasts via activation of the AKT-mTOR pathway. J. Cell. Physiol. 231 (6), 1283–1290. doi:10.1002/jcp.25228
Bencivenga, L., Strumia, M., Rolland, Y., Martinez, L., Cestac, P., Guyonnet, S., et al. (2022). Biomarkers of mitochondrial dysfunction and inflammaging in older adults and blood pressure variability. Geroscience 45, 797–809. doi:10.1007/s11357-022-00697-y
Bren-Mattison, Y., Hausburg, M., and Olwin, B. B. (2011). Growth of limb muscle is dependent on skeletal-derived Indian hedgehog. Dev. Biol. 356 (2), 486–495. doi:10.1016/j.ydbio.2011.06.002
Brotto, M., and Bonewald, L. (2015). Bone and muscle: Interactions beyond mechanical. Bone 80, 109–114. doi:10.1016/j.bone.2015.02.010
Buehring, B., and Binkley, N. (2013). Myostatin--the holy grail for muscle, bone, and fat? Curr. Osteoporos. Rep. 11 (4), 407–414. doi:10.1007/s11914-013-0160-5
Butikofer, L., Zurlinden, A., Bolliger, M. F., Kunz, B., and Sonderegger, P. (2011). Destabilization of the neuromuscular junction by proteolytic cleavage of agrin results in precocious sarcopenia. FASEB J. 25 (12), 4378–4393. doi:10.1096/fj.11-191262
Cevei, M., Onofrei, R. R., Cioara, F., and Stoicanescu, D. (2020). Correlations between the quality of life domains and clinical variables in sarcopenic osteoporotic postmenopausal women. J. Clin. Med. 9 (2), 441. doi:10.3390/jcm9020441
Cheema, N., Herbst, A., McKenzie, D., and Aiken, J. M. (2015). Apoptosis and necrosis mediate skeletal muscle fiber loss in age-induced mitochondrial enzymatic abnormalities. Aging Cell. 14 (6), 1085–1093. doi:10.1111/acel.12399
Chen, X., Wang, Z., Duan, N., Zhu, G., Schwarz, E. M., and Xie, C. (2018). Osteoblast-osteoclast interactions. Connect. Tissue Res. 59 (2), 99–107. doi:10.1080/03008207.2017.1290085
Cheng, Q., Chen, X., Wu, H., and Du, Y. (2021). Three hematologic/immune system-specific expressed genes are considered as the potential biomarkers for the diagnosis of early rheumatoid arthritis through bioinformatics analysis. J. Transl. Med. 19 (1), 18. doi:10.1186/s12967-020-02689-y
Cheng, X., Yan, Z., Su, Z., and Liu, J. (2022). The transforming growth factor beta ligand TIG-2 modulates the function of neuromuscular junction and muscle energy metabolism in Caenorhabditis elegans. Front. Mol. Neurosci. 15, 962974. doi:10.3389/fnmol.2022.962974
Clynes, M. A., Gregson, C. L., Bruyere, O., Cooper, C., and Dennison, E. M. (2021). Osteosarcopenia: Where osteoporosis and sarcopenia collide. Rheumatol. Oxf. 60 (2), 529–537. doi:10.1093/rheumatology/keaa755
Collins, F. L., Rios-Arce, N. D., Schepper, J. D., Parameswaran, N., and McCabe, L. R. (2017). The potential of probiotics as a therapy for osteoporosis. Microbiol. Spectr. 5 (4). doi:10.1128/microbiolspec.BAD-0015-2016
Cruz-Jentoft, A. J., Baeyens, J. P., Bauer, J. M., Boirie, Y., Cederholm, T., Landi, F., et al. (2010). Sarcopenia: European consensus on definition and diagnosis: Report of the European working group on sarcopenia in older people. Age Ageing 39 (4), 412–423. doi:10.1093/ageing/afq034
Cruz-Jentoft, A. J., Bahat, G., Bauer, J., Boirie, Y., Bruyere, O., Cederholm, T., et al. (2019). Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 48 (4), 601. doi:10.1093/ageing/afz046
de Almeida Marques Bernabe, R., de Souza Vieira, M., Felicio de Souza, V., Gomes Fontana, L., Albergaria, B. H., Marques-Rocha, J. L., et al. (2022). Muscle strength is associated with fracture risk obtained by fracture risk assessment tool (FRAX) in women with breast cancer. BMC Cancer 22 (1), 1115. doi:10.1186/s12885-022-10203-4
Deng, Y. X., He, W. G., Cai, H. J., Jiang, J. H., Yang, Y. Y., Dan, Y. R., et al. (2021). Analysis and validation of hub genes in blood monocytes of postmenopausal osteoporosis patients. Front. Endocrinol. (Lausanne). 12, 815245. doi:10.3389/fendo.2021.815245
Gai, D., Chen, J. R., Stewart, J. P., Nookaew, I., Habelhah, H., Ashby, C., et al. (2022). CST6 suppresses osteolytic bone disease in multiple myeloma by blocking osteoclast differentiation. J. Clin. Invest. 132 (18), e159527. doi:10.1172/JCI159527
Gautvik, K. M., Olstad, O. K., Raue, U., Gautvik, V. T., Kvernevik, K. J., Utheim, T. P., et al. (2022). Heavy-load exercise in older adults activates vasculogenesis and has a stronger impact on muscle gene expression than in young adults. Eur. Rev. Aging Phys. Act. 19 (1), 23. doi:10.1186/s11556-022-00304-1
Gomes, M. M., da Silva, M. M. R., de Araujo, I. M., and de Paula, F. J. A. (2022). Bone, fat, and muscle interactions in health and disease. Arch. Endocrinol. Metab. 66 (5), 611–620. doi:10.20945/2359-3997000000550
Gros, K., Matkovic, U., Parato, G., Mis, K., Luin, E., Bernareggi, A., et al. (2022). Neuronal agrin promotes proliferation of primary human myoblasts in an age-dependent manner. Int. J. Mol. Sci. 23 (19), 11784. doi:10.3390/ijms231911784
Guillamon-Escudero, C., Diago-Galmes, A., Zuazua Rico, D., Maestro-Gonzalez, A., Tenias-Burillo, J. M., Soriano, J. M., et al. (2022). SarQoL questionnaire in community-dwelling older adults under EWGSOP2 sarcopenia diagnosis algorithm: A new screening method? Int. J. Environ. Res. Public Health 19 (14), 8473. doi:10.3390/ijerph19148473
Hashemi, M., Taheriazam, A., Daneii, P., Hassanpour, A., Kakavand, A., Rezaei, S., et al. (2022). Targeting PI3K/Akt signaling in prostate cancer therapy. J. Cell. Commun. Signal. doi:10.1007/s12079-022-00702-1
Hatta, K., and Takeichi, M. (1986). Expression of N-cadherin adhesion molecules associated with early morphogenetic events in chick development. Nature 320 (6061), 447–449. doi:10.1038/320447a0
Hephzibah Cathryn, R., Udhaya Kumar, S., Younes, S., Zayed, H., and George Priya Doss, C. (2022). A review of bioinformatics tools and web servers in different microarray platforms used in cancer research. Adv. Protein Chem. Struct. Biol. 131, 85–164. doi:10.1016/bs.apcsb.2022.05.002
Hernlund, E., Svedbom, A., Ivergard, M., Compston, J., Cooper, C., Stenmark, J., et al. (2013). Osteoporosis in the European union: Medical management, epidemiology and economic burden. A report prepared in collaboration with the international osteoporosis foundation (IOF) and the European federation of pharmaceutical industry associations (efpia). Arch. Osteoporos. 8, 136. doi:10.1007/s11657-013-0136-1
Hirano, K., Tsuchiya, M., Shiomi, A., Takabayashi, S., Suzuki, M., Ishikawa, Y., et al. (2023). The mechanosensitive ion channel PIEZO1 promotes satellite cell function in muscle regeneration. Life Sci. Alliance 6 (2), e202201783. doi:10.26508/lsa.202201783
Hu, Z., Yang, A., Tian, Y., and Song, X. (2022). Daytime napping, comorbidity profiles, and the risk of sarcopenia in older individuals. Front. Physiol. 13, 1000593. doi:10.3389/fphys.2022.1000593
Huang, J., Hsu, Y. H., Mo, C., Abreu, E., Kiel, D. P., Bonewald, L. F., et al. (2014). METTL21C is a potential pleiotropic gene for osteoporosis and sarcopenia acting through the modulation of the NF-κB signaling pathway. J. Bone Min. Res. 29 (7), 1531–1540. doi:10.1002/jbmr.2200
Huybrechts, Y., Mortier, G., Boudin, E., and Van Hul, W. (2020). WNT signaling and bone: Lessons from skeletal dysplasias and disorders. Front. Endocrinol. (Lausanne). 11, 165. doi:10.3389/fendo.2020.00165
Jun, J. E., Lee, S. E., Lee, Y. B., Kim, G., Jin, S. M., Jee, J. H., et al. (2022). Low skeletal muscle mass accompanied by abdominal obesity additively increases the risk of incident type 2 diabetes. J. Clin. Endocrinol. Metab., dgac662. doi:10.1210/clinem/dgac662
Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M., and Tanabe, M. (2016). KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44 (D1), D457–D462. doi:10.1093/nar/gkv1070
Kanis, J. A., Cooper, C., Rizzoli, R., and Reginster, J. Y.Scientific Advisory Board of the European Society for Clinical and Economic Aspects of Osteoporosis ESCEO and the Committees of Scientific Advisors and National Societies of the International Osteoporosis Foundation IOF (2019). European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 30 (1), 3–44. doi:10.1007/s00198-018-4704-5
Karar, J., and Maity, A. (2011). PI3K/AKT/mTOR pathway in angiogenesis. Front. Mol. Neurosci. 4, 51. doi:10.3389/fnmol.2011.00051
Kawao, N., and Kaji, H. (2015). Interactions between muscle tissues and bone metabolism. J. Cell. Biochem. 116 (5), 687–695. doi:10.1002/jcb.25040
Khan, A., Ni, W., Lopez-Giraldez, F., Kluger, M. S., Pober, J. S., and Pierce, R. W. (2021). Tumor necrosis factor-induced ArhGEF10 selectively activates RhoB contributing to human microvascular endothelial cell tight junction disruption. FASEB J. 35 (6), e21627. doi:10.1096/fj.202002783RR
Khosla, S., Wright, N. C., Elderkin, A. L., and Kiel, D. P. (2022). Osteoporosis in the USA: Prevention and unmet needs. Lancet Diabetes Endocrinol. 11, 19–20. doi:10.1016/S2213-8587(22)00322-9
Kim, H., Takegahara, N., Walsh, M. C., Ueda, J., Fujihara, Y., Ikawa, M., et al. (2020). Protocadherin-7 contributes to maintenance of bone homeostasis through regulation of osteoclast multinucleation. BMB Rep. 53 (9), 472–477. doi:10.5483/BMBRep.2020.53.9.050
Kim, T. H., Jung, Y. K., and Yim, H. J. (2022). Sarcopenia: Multiple factors to be considered in cirrhosisauthor's reply. Clin. Mol. Hepatol. 29, 173–175. doi:10.3350/cmh.2022.0372
Kwon, Y. E., Lee, J. S., Kim, J. Y., Baeg, S. I., Choi, H. M., Kim, H. B., et al. (2022). Impact of sarcopenia and phase angle on mortality of the very elderly. J. Cachexia Sarcopenia Muscle 14, 279–287. doi:10.1002/jcsm.13128
Lam, H., and Qin, Y. X. (2008). The effects of frequency-dependent dynamic muscle stimulation on inhibition of trabecular bone loss in a disuse model. Bone 43 (6), 1093–1100. doi:10.1016/j.bone.2008.07.253
Langfelder, P., and Horvath, S. (2008). Wgcna: an R package for weighted correlation network analysis. BMC Bioinforma. 9, 559. doi:10.1186/1471-2105-9-559
Leiter, J. R., Peeler, J., and Anderson, J. E. (2011). Exercise-induced muscle growth is muscle-specific and age-dependent. Muscle Nerve 43 (6), 828–838. doi:10.1002/mus.21965
Li, J., Yi, X., Yao, Z., Chakkalakal, J. V., Xing, L., and Boyce, B. F. (2020). TNF receptor-associated factor 6 mediates tnfα-induced skeletal muscle atrophy in mice during aging. J. Bone Min. Res. 35 (8), 1535–1548. doi:10.1002/jbmr.4021
Li, X., Liang, Y., Lian, C., Peng, F., Xiao, Y., He, Y., et al. (2021). CST6 protein and peptides inhibit breast cancer bone metastasis by suppressing CTSB activity and osteoclastogenesis. Theranostics 11 (20), 9821–9832. doi:10.7150/thno.62187
Liu, M., Chu, Q., Yang, C., Wang, J., Fu, M., Zhang, Z., et al. (2022a). The paradoxical relation between serum uric acid and outcomes of hip fracture in older patients after surgery: A 1-year follow-up study. Surgery 172 (5), 1576–1583. doi:10.1016/j.surg.2022.07.008
Liu, J., Zhang, D., Cao, Y., Zhang, H., Li, J., Xu, J., et al. (2022b). Screening of crosstalk and pyroptosis-related genes linking periodontitis and osteoporosis based on bioinformatics and machine learning. Front. Immunol. 13, 955441. doi:10.3389/fimmu.2022.955441
Liu, Z., Xu, Y., Liu, X., and Wang, B. (2022c). PCDH7 knockdown potentiates colon cancer cells to chemotherapy via inducing ferroptosis and changes in autophagy through restraining MEK1/2/ERK/c-Fos axis. Biochem. Cell. Biol. 100 (6), 445–457. doi:10.1139/bcb-2021-0513
Liu, Y., Reynolds, L. M., Ding, J., Hou, L., Lohman, K., Young, T., et al. (2017). Blood monocyte transcriptome and epigenome analyses reveal loci associated with human atherosclerosis. Nat. Commun. 8 (1), 393. doi:10.1038/s41467-017-00517-4
Loh, R., Tan, R. S., Lim, W. S., and Koh, A. S. (2022). Cardio-sarcopenia: A syndrome of concern in aging. Front. Med. (Lausanne). 9, 1027466. doi:10.3389/fmed.2022.1027466
Long, J. B., and Van Vactor, D. (2012). Maintaining muscle mitochondria via transsynaptic signaling. Dev. Cell. 22 (2), 238–239. doi:10.1016/j.devcel.2012.01.021
Ma, C., Li, F., He, Z., and Zhao, S. (2022). A more novel and powerful prognostic gene signature of lung adenocarcinoma determined from the immune cell infiltration landscape. Front. Surg. 9, 1015263. doi:10.3389/fsurg.2022.1015263
Malmstrom, T. K., Miller, D. K., Simonsick, E. M., Ferrucci, L., and Morley, J. E. (2016). SARC-F: A symptom score to predict persons with sarcopenia at risk for poor functional outcomes. J. Cachexia Sarcopenia Muscle 7 (1), 28–36. doi:10.1002/jcsm.12048
Manske, S. L., Boyd, S. K., and Zernicke, R. F. (2011). Vertical ground reaction forces diminish in mice after botulinum toxin injection. J. Biomech. 44 (4), 637–643. doi:10.1016/j.jbiomech.2010.11.011
Marini, F., Giusti, F., Palmini, G., and Brandi, M. L. (2022). Role of Wnt signaling and sclerostin in bone and as therapeutic targets in skeletal disorders. Osteoporos. Int. 34, 213–238. doi:10.1007/s00198-022-06523-7
Mastnak, T., Maver, U., and Finsgar, M. (2022). Addressing the needs of the rapidly aging society through the development of multifunctional bioactive coatings for orthopedic applications. Int. J. Mol. Sci. 23 (5), 2786. doi:10.3390/ijms23052786
Mijnarends, D. M., Luiking, Y. C., Halfens, R. J. G., Evers, S., Lenaerts, E. L. A., Verlaan, S., et al. (2018). Muscle, health and costs: A glance at their relationship. J. Nutr. Health Aging 22 (7), 766–773. doi:10.1007/s12603-018-1058-9
Mo, L., Ma, C., Wang, Z., Li, J., He, W., Niu, W., et al. (2022). Integrated bioinformatic analysis of the shared molecular mechanisms between osteoporosis and atherosclerosis. Front. Endocrinol. (Lausanne). 13, 950030. doi:10.3389/fendo.2022.950030
Nie, Y. Z., Yan, Z. Q., Yin, H., Shan, L. H., Wang, J. H., and Wu, Q. H. (2022). Osteosarcopenic obesity and its components-osteoporosis, sarcopenia, and obesity-are associated with blood cell count-derived inflammation indices in older Chinese People. BMC Geriatr. 22 (1), 532. doi:10.1186/s12877-022-03225-x
Niimi, T. (2021). Roles of Slit ligands and their Roundabout (robo) family of receptors in bone remodeling. Adv. Exp. Med. Biol. 21, 143–154. doi:10.1007/5584_2020_586
Noda, T., Kamiya, K., Hamazaki, N., Nozaki, K., Ichikawa, T., Yamashita, M., et al. (2022). SARC-F predicts poor motor function, quality of life, and prognosis in older patients with cardiovascular disease and cognitive impairment. Exp. Gerontol. 171, 112021. doi:10.1016/j.exger.2022.112021
Novak, J. S., Mazala, D. A. G., Nearing, M., Hindupur, R., Uapinyoying, P., Habib, N. F., et al. (2021). Human muscle stem cells are refractory to aging. Aging Cell. 20 (7), e13411. doi:10.1111/acel.13411
Oflazoglu, U., Caglar, S., Yilmaz, H. E., Onal, H. T., Varol, U., Salman, T., et al. (2022). The relationship between sarcopenia detected in newly diagnosed colorectal cancer patients and FGF21, irisin and CRP levels. Eur. Geriatr. Med. 13 (4), 795–803. doi:10.1007/s41999-022-00635-3
Oh, G. S., Lee, J. H., Byun, K., Kim, D. I., and Park, K. D. (2022). Effect of intake of leucine-rich protein supplement in parallel with resistance exercise on the body composition and function of healthy adults. Nutrients 14 (21), 4501. doi:10.3390/nu14214501
Okamoto, K., and Takayanagi, H. (2019). Osteoimmunology. Cold Spring Harb. Perspect. Med. 9 (1), a031245. doi:10.1101/cshperspect.a031245
Okayama, A., Nakayama, N., Kashiwa, K., Horinouchi, Y., Fukusaki, H., Nakamura, H., et al. (2022). Prevalence of sarcopenia and its association with quality of life, postural stability, and past incidence of falls in postmenopausal women with osteoporosis: A cross-sectional study. Healthc. (Basel). 10 (2), 192. doi:10.3390/healthcare10020192
Ordan, E., and Volk, T. (2015). A non-signaling role of Robo2 in tendons is essential for Slit processing and muscle patterning. Development 142 (20), 3512–3518. doi:10.1242/dev.128157
Pak, J. S., DeLoughery, Z. J., Wang, J., Acharya, N., Park, Y., Jaworski, A., et al. (2020). NELL2-Robo3 complex structure reveals mechanisms of receptor activation for axon guidance. Nat. Commun. 11 (1), 1489. doi:10.1038/s41467-020-15211-1
Patel, D., and Wairkar, S. (2022). Recent advances in cyclosporine drug delivery: Challenges and opportunities. Drug Deliv. Transl. Res. 9, 1067–1081. doi:10.1007/s13346-019-00650-1
Patra, B. G., Maroufy, V., Soltanalizadeh, B., Deng, N., Zheng, W. J., Roberts, K., et al. (2020). A content-based literature recommendation system for datasets to improve data reusability - a case study on Gene Expression Omnibus (GEO) datasets. J. Biomed. Inf. 104, 103399. doi:10.1016/j.jbi.2020.103399
Polito, A., Barnaba, L., Ciarapica, D., and Azzini, E. (2022). Osteosarcopenia: A narrative review on clinical studies. Int. J. Mol. Sci. 23 (10), 5591. doi:10.3390/ijms23105591
Pomaznoy, M., Ha, B., and Peters, B. (2018). GOnet: A tool for interactive gene ontology analysis. BMC Bioinforma. 19 (1), 470. doi:10.1186/s12859-018-2533-3
Qin, Q., Lee, S., Patel, N., Walden, K., Gomez-Salazar, M., Levi, B., et al. (2022). Neurovascular coupling in bone regeneration. Exp. Mol. Med. 54, 1844–1849. doi:10.1038/s12276-022-00899-6
Quan, Z., Yang, Y., Zheng, H., Zhan, Y., Luo, J., Ning, Y., et al. (2022). Clinical implications of the interaction between PD-1/PD-L1 and PI3K/AKT/mTOR pathway in progression and treatment of non-small cell lung cancer. J. Cancer 13 (13), 3434–3443. doi:10.7150/jca.77619
Rai, N., and Dey, S. (2020). Protective response of Sestrin under stressful conditions in aging. Ageing Res. Rev. 64, 101186. doi:10.1016/j.arr.2020.101186
Reiss, J., Iglseder, B., Alzner, R., Mayr-Pirker, B., Pirich, C., Kassmann, H., et al. (2019). Sarcopenia and osteoporosis are interrelated in geriatric inpatients. Z Gerontol. Geriatr. 52 (7), 688–693. doi:10.1007/s00391-019-01553-z
Salgado-Albarran, M., Spath, J., Gonzalez-Barrios, R., Baumbach, J., and Soto-Reyes, E. (2022). CTCFL regulates the PI3K-Akt pathway and it is a target for personalized ovarian cancer therapy. NPJ Syst. Biol. Appl. 8 (1), 5. doi:10.1038/s41540-022-00214-z
Savary-Auzeloux, I., Jarzaguet, M., Migne, C., Kemeny, J. L., Novais-Gameiro, L., de Azevedo, M., et al. (2022). Anti-inflammatory Streptococcus thermophilus CNRZ160 limits sarcopenia induced by low-grade inflammation in older adult rats. Front. Nutr. 9, 986542. doi:10.3389/fnut.2022.986542
Shen, H., Grimston, S., Civitelli, R., and Thomopoulos, S. (2015). Deletion of connexin43 in osteoblasts/osteocytes leads to impaired muscle formation in mice. J. Bone Min. Res. 30 (4), 596–605. doi:10.1002/jbmr.2389
Shirayoshi, Y., Hatta, K., Hosoda, M., Tsunasawa, S., Sakiyama, F., and Takeichi, M. (1986). Cadherin cell adhesion molecules with distinct binding specificities share a common structure. EMBO J. 5 (10), 2485–2488. doi:10.1002/j.1460-2075.1986.tb04525.x
Sirago, G., Picca, A., Calvani, R., Coelho-Junior, H. J., and Marzetti, E. (2022). Mammalian target of rapamycin (mTOR) signaling at the crossroad of muscle fiber fate in sarcopenia. Int. J. Mol. Sci. 23 (22), 13823. doi:10.3390/ijms232213823
Spexoto, M. C. B., Ramirez, P. C., de Oliveira Maximo, R., Steptoe, A., de Oliveira, C., and Alexandre, T. D. S. (2022). European working group on sarcopenia in older people 2010 (EWGSOP1) and 2019 (EWGSOP2) criteria or slowness: Which is the best predictor of mortality risk in older adults? Age Ageing 51 (7), afac164. doi:10.1093/ageing/afac164
Szklarczyk, D., Gable, A. L., Lyon, D., Junge, A., Wyder, S., Huerta-Cepas, J., et al. (2019). STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 47 (1), D607–D613. doi:10.1093/nar/gky1131
Tagliaferri, C., Wittrant, Y., Davicco, M. J., Walrand, S., and Coxam, V. (2015). Muscle and bone, two interconnected tissues. Ageing Res. Rev. 21, 55–70. doi:10.1016/j.arr.2015.03.002
Teng, Z., Zhu, Y., Teng, Y., Long, Q., Hao, Q., Yu, X., et al. (2021). The analysis of osteosarcopenia as a risk factor for fractures, mortality, and falls. Osteoporos. Int. 32 (11), 2173–2183. doi:10.1007/s00198-021-05963-x
Terashima, A., and Takayanagi, H. (2018). Overview of osteoimmunology. Calcif. Tissue Int. 102 (5), 503–511. doi:10.1007/s00223-018-0417-1
Tournadre, A., Vial, G., Capel, F., Soubrier, M., and Boirie, Y. (2019). Sarcopenia. Jt. Bone Spine. 86 (3), 309–314. doi:10.1016/j.jbspin.2018.08.001
Wang, C., Chen, A., Ruan, B., Niu, Z., Su, Y., Qin, H., et al. (2020a). PCDH7 inhibits the formation of homotypic cell-in-cell structure. Front. Cell. Dev. Biol. 8, 329. doi:10.3389/fcell.2020.00329
Wang, Y., Kerrisk Campbell, M., Tom, I., Foreman, O., Hanson, J. E., and Sheng, M. (2020b). PCDH7 interacts with GluN1 and regulates dendritic spine morphology and synaptic function. Sci. Rep. 10 (1), 10951. doi:10.1038/s41598-020-67831-8
Wang, Y. J., Wang, Y., Zhan, J. K., Tang, Z. Y., He, J. Y., Tan, P., et al. (2015). Sarco-Osteoporosis: Prevalence and association with frailty in Chinese community-dwelling older adults. Int. J. Endocrinol. 2015, 482940. doi:10.1155/2015/482940
Wang, Z., Xia, Q., Su, W., Zhang, M., Gu, Y., Xu, J., et al. (2022a). The commonness in immune infiltration of rheumatoid arthritis and atherosclerosis: Screening for central targets via microarray data analysis. Front. Immunol. 13, 1013531. doi:10.3389/fimmu.2022.1013531
Wang, F., Yang, G., Li, Y., Tang, Z., Du, J., Song, H., et al. (2022b). A peptide from wheat germ abolishes the senile osteoporosis by regulating OPG/RANKL/RANK/TRAF6 signaling pathway. Phytomedicine 104, 154304. doi:10.1016/j.phymed.2022.154304
Wang, Q., Shen, Z. N., Zhang, S. J., Sun, Y., Zheng, F. J., and Li, Y. H. (2022c). Protective effects and mechanism of puerarin targeting PI3K/Akt signal pathway on neurological diseases. Front. Pharmacol. 13, 1022053. doi:10.3389/fphar.2022.1022053
Weitao Shen, Z. S., Zhong, X., Huang, M., Shen, D., Gao, P., Qian, X., et al. (2022). Sangerbox: A comprehensive, interaction-friendly clinical bioinformatics analysis platform. iMeta 1 (3), e36. doi:10.1002/imt2.36
Xie, B., Chen, S., Xu, Y., Han, W., Hu, R., Chen, M., et al. (2021). The impact of glucagon-like peptide 1 receptor agonists on bone metabolism and its possible mechanisms in osteoporosis treatment. Front. Pharmacol. 12, 697442. doi:10.3389/fphar.2021.697442
Xin, D., Man, Y., Yang, Y., and Wang, F. (2022). A novel prognostic and therapeutic target biomarker based on necroptosis-related gene signature and immune microenvironment infiltration in gastric cancer. Front. Genet. 13, 953997. doi:10.3389/fgene.2022.953997
Yan, C., Zhang, J., An, F., Wang, J., Shi, Y., Yuan, L., et al. (2022). Research progress of ferroptosis regulatory network and bone remodeling in osteoporosis. Front. Public Health 10, 910675. doi:10.3389/fpubh.2022.910675
Yang, J., Park, O. J., Kim, J., Kwon, Y., Yun, C. H., and Han, S. H. (2019). Modulation of macrophage subtypes by IRF5 determines osteoclastogenic potential. J. Cell. Physiol. 234 (12), 23033–23042. doi:10.1002/jcp.28863
Yang, Q., Wang, Y., Zhao, C., Pang, S., Lu, J., and Chan, P. (2022). α-Synuclein aggregation causes muscle atrophy through neuromuscular junction degeneration. J. Cachexia Sarcopenia Muscle 14, 226–242. doi:10.1002/jcsm.13123
Yoshida, K., Watanabe, M., Kato, H., Dutta, A., and Sugano, S. (1999). BH-protocadherin-c, a member of the cadherin superfamily, interacts with protein phosphatase 1 alpha through its intracellular domain. FEBS Lett. 460 (1), 93–98. doi:10.1016/s0014-5793(99)01309-5
Yoshida, K., Yoshitomo-Nakagawa, K., Seki, N., Sasaki, M., and Sugano, S. (1998). Cloning, expression analysis, and chromosomal localization of BH-protocadherin (PCDH7), a novel member of the cadherin superfamily. Genomics 49 (3), 458–461. doi:10.1006/geno.1998.5271
Yuen, D. A., and Robinson, L. A. (2013). Slit2-Robo signaling: A novel regulator of vascular injury. Curr. Opin. Nephrol. Hypertens. 22 (4), 445–451. doi:10.1097/MNH.0b013e32836235f4
Zeng, Z., Guo, R., Wang, Z., Yan, H., Lv, X., Zhao, Q., et al. (2022). Circulating monocytes act as a common trigger for the calcification paradox of osteoporosis and carotid atherosclerosis via TGFB1-SP1 and TNFSF10-NFKB1 Axis. Front. Endocrinol. (Lausanne). 13, 944751. doi:10.3389/fendo.2022.944751
Zhang, H., Hu, Y., Chen, X., Wang, S., Cao, L., Dong, S., et al. (2022a). Expert consensus on the bone repair strategy for osteoporotic fractures in China. Front. Endocrinol. (Lausanne). 13, 989648. doi:10.3389/fendo.2022.989648
Zhang, X., Groen, K., Morten, B. C., Steffens Reinhardt, L., Campbell, H. G., Braithwaite, A. W., et al. (2022b). Effect of p53 and its N-terminally truncated isoform, Δ40p53, on breast cancer migration and invasion. Mol. Oncol. 16 (2), 447–465. doi:10.1002/1878-0261.13118
Keywords: sarcopenia, osteoporosis, crosstalk genes, machine learning, bioinformatic analysis
Citation: Liu M, Wang Y, Shi W, Yang C, Wang Q, Chen J, Li J, Chen B and Sun G (2023) PCDH7 as the key gene related to the co-occurrence of sarcopenia and osteoporosis. Front. Genet. 14:1163162. doi: 10.3389/fgene.2023.1163162
Received: 10 February 2023; Accepted: 06 April 2023;
Published: 05 July 2023.
Edited by:
Umberto Tarantino, University of Rome Tor Vergata, ItalyCopyright © 2023 Liu, Wang, Shi, Yang, Wang, Chen, Li, Chen and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bingdi Chen, aW5hbm9jaGVuQHRvbmdqaS5lZHUuY24=; Guixin Sun, c3VuZ3VpeGluQHRvbmdqaS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.