 Siham A. Rahmatalla
Siham A. Rahmatalla Danny Arends
Danny Arends Gudrun A. Brockmann
Gudrun A. Brockmann
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet., 07 December 2022
Sec. Livestock Genomics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.995349
The milk casein genes in goats, are highly polymorphic genes with numerous synonymous and non-synonymous mutations. So far, 20 protein variants have been reported in goats for alpha-S1-casein, eight for beta-casein, 14 for alpha-S2-casein, and 24 for kappa-casein. This review provides a comprehensive overview on identified milk casein protein variants in goat and non-coding DNA sequence variants with some affecting the expression of the casein genes. The high frequency of some casein protein variants in different goat breeds and geographical regions might reflect specific breeding goals with respect to milk processing characteristics, properties for human nutrition and health, or adaptation to the environment. Because protein names, alongside the discovery of protein variants, go through a historical process, we linked old protein names with new ones that reveal more genetic variability. The haplotypes across the cluster of the four genetically linked casein genes are recommended as a valuable genetic tool for discrimination between breeds, managing genetic diversity within and between goat populations, and breeding strategies. The enormous variation in the casein proteins and genes is crucial for producing milk and dairy products with different properties for human health and nutrition, and for genetic improvement depending on local breeding goals.
Casein is the insoluble major protein component of milk comprising about 80% of the total milk protein fraction (Bhat et al., 2016). Milk caseins have a fundamental nutritional value since they provide calcium and phosphorus for bone formation and amino acid synthesis in offspring (Barth and Schlimme, 2012; Ghosh et al., 2016). Milk composition is a multifactorial complex trait. Besides genetic factors, the total protein and casein content is affected by various factors such as number of milking per day, stage of lactation, age of the animal, parity, rearing practices, seasonal components, and nutrition (Assan, 2014).
In goats, like in other mammals, four evolutionary conserved casein genes occur. The genes encoding alpha-S1-, beta-, alpha-S2-, and kappa-caseins are CSN1S1, CSN2, CSN1S2, and CSN3, respectively (Martin et al., 2002). All four genes are located in a 250 kb region between 85.978 and 86.211 Mb on Capra hircus chromosome 6 (Ferretti et al., 1990; Threadgill and Womack, 1990).
Caseins occur in milk in micelles which are large spherical colloidal particles. These micelles are buildup of several submicelles which are bound together by colloidal calcium phosphate (Fox and McSweeney, 1998). Alpha-S1-, α-S2-, and β-caseins, which are hydrophobic phosphate and calcium-sensitive caseins, are located inside the inner part of the micelle (Hui and Sherkat, 2005), which precipitate from raw milk upon acidification at pH 4.6 at 20°C⁰ (Park et al., 2017). Kappa-casein, the insensitive calcium protein, resides on the surface of the micelles where it stabilizes the micelles with a protruding hydrophobic chain (Guinee and O'Brien, 2010).
The most important economic effects of the milk casein proteins come from the relationship between protein quality, amount, coagulation, and techno-functional properties to produce yoghurts and cheese. Casein polymorphisms also affect the fat content, micellar size, mineralization, and sensory characteristic in milk. Therefore, casein polymorphisms play a significant role both in milk coagulation performance and cheese quality. Since the identification of the impact of milk protein polymorphisms on milk quality and its technological properties (Aschaffenburg and Drewry, 1955), milk protein variants have been intensively characterized in different species including goats (Marletta et al., 2007; Caroli et al., 2009; Giambra and Erhardt, 2012; Selvaggi et al., 2014; Ozdemir et al., 2018). As a side effect of the intensive work, simultaneous publications led to inconsistency in the naming and positioning of some sequence variants which were anchored in part to mature and in part to immature proteins and sometimes include mutations in non-coding regions. In addition, many studies have referred to the genetic and protein variants of caseins in goats with different nomenclature. This review gives a comprehensive overview on identified protein and DNA variants of the four casein genes in goats. One goal is to clarify names and nomenclatures, the positions of nucleotide and amino acid polymorphisms and provide a translation from DNA to protein variant. We focus on protein variants, but also describe linked nucleotide variants in non-coding regions of the gene, like the promoter, 5′ and 3’ untranslated regions, and introns, which could affect the transcription and translation and thereby production traits. Furthermore, we have derived haplotypes across all four linked genes and summarize the frequency of alleles in different breeds and the impact of milk protein variants on milk quality for cheese making, human health and human nutrition.
At the functional level, alpha-S1-casein is required for the efficient export of other caseins from the endoplasmic reticulum to the Golgi compartment, the casein transport in the secretory pathway, and for the biogenesis of casein micelles in milk (Chanat et al., 1999; Le Parc et al., 2010). Alpha-S1-casein has an important role in the capacity of milk to provide calcium phosphate for human nutrition (Kolb et al., 2011).
The percentage of α-S1-casein in goat milk varies from 0 to 25%. Alpha-S1-casein concentrations has been shown to be dependent on the existence of polymorphisms in the coding and regulatory sequences of the gene (Boulanger et al., 1984). In contrast, the α-S1-casein fraction in bovine milk constitutes up to 40% (Farrell et al., 2004). Strong, intermediate, low, and null expression levels of α-S1-casein in goat milk are distinguished (Moatsou et al., 2004; Caroli et al., 2007; Kupper et al., 2010). In the first group comprising CSN1S1*A, CSN1S1*A3, CSN1S1*B1, CSN1S1*B2, CSN1S1*B3, CSN1S1*B4, CSN1S1*C, CSN1S1*H, CSN1S1*L, and CSN1S1*M are “strong” variants, which are associated with a high content of α-S1-casein in milk. CSN1S1*E and CSN1S1*I are “intermediate” variants associated with a medium content. CSN1S1*D, CSN1S1*F, and CSN1S1*G are “weak” variants associated with a low content, and CSN1S1*N, CSN1S1*01, and CSN1S1*02 are “null” variants leading to the absence of α-S1-casein in the milk of animals homozygote for these alleles (Grosclaude et al., 1987; Brignon et al., 1990; Chianese et al., 1997; Grosclaude and Martin, 1997; Martin et al., 1999; Bevilacqua et al., 2002; Ramunno et al., 2005; Mestawet et al., 2013). The α-S1-casein strong variants produced around 3.6 g/L per allele, medium variants produced 1.1–1.6 g/L per allele, weak variants produced 0.45–0.6 g/L per allele, and null variants resulted in absence of the α-S1-casein in milk (Grosclaude et al., 1987; Grosclaude and Martin, 1997; Martin et al., 1999). Strong variants with high content of α-S1-casein are most desired in dairy production since they are associated with lower pH values, better coagulation properties, faster curdling rate and greater gel firmness, which contribute largely to high yield and quality of cheese, even if the rennet coagulation time was longer (Ambrosoli et al., 1988; Pirisi et al., 1994; Clark and Sherbon, 2000; Schmidely et al., 2002; Zullo et al., 2005; Caravaca et al., 2011; Devold et al., 2011; Cebo et al., 2012; Vacca et al., 2014; Johansson et al., 2015; Marković et al., 2018). Since the casein content in milk also affects the size of casein micelles and the amount and composition of milk fatty acids, cheese made from milk with strong α-S1-variants has a better structure and taste with less typical goat flavor than cheese produced from intermediate or weak variants (Delacroix Buchet et al., 1996). Generally, the lower α-S1-casein content in goat milk compared to bovine milk is associated with smaller casein micelles, with less hydrated pores (Park et al., 2007; Ingham et al., 2018; Nguyen et al., 2018). Hence, yoghurt (Miocinovic et al., 2016; Nguyen et al., 2018) and cheese (Park et al., 2007; Mestawet et al., 2014) made from goat milk have a lower dense gel structure and poor coagulation features compared to their counterparts made with cow milk.
The CSN1S1 gene is located on chromosome 6 between 85.978 and 85.995 Mb and spans 16,808 bp. The DNA and protein reference sequences NC_030813 (goat genome reference version LWLT01 (Bickhart et al., 2017)) and XP_017904616 (NCBI database), respectively, represent the CSN1S1*A protein variant (Boulanger et al., 1984; Brignon et al., 1989; Ramunno et al., 2004). The gene encodes 214 amino acids of α-S1-casein, the first 15 amino acids form the signal peptide sequence, followed by 199 amino acids long mature protein (Brignon et al., 1990). It consists of 19 exons ranging in size from 24 (exons 5, 6, 7, 8, 10, 13, 16) to 385 bp (exon 19) and 18 introns from 90 bp (intron 10) to 1,685 bp (intron 2) (Ramunno et al., 2004).
In the following, we briefly describe the 20 protein variants that have been identified so far (Supplementary Table S1).
The α-S1-casein CSNS1*A is the reference protein. The sub variants CSN1S1*A′ and CSN1S1*A2 represent the same reference protein, namely CSN1S1*A, though CSN1S1*A′ carries a T/C transition in intron 9 (Kupper et al., 2010) and CSN1S1*A2 a G/A transition in the 5′ flanking region (Ramunno et al., 2005). The same G/A transition is present also in variants CSN1S1*N and CSN1S1*01, which are null variants (Ramunno et al., 2005), which means absence of α-S1-casein in the milk of homozygous carriers. Hence, the G/A variation in the 5′ flanking region is not responsible for the absence of α-S1-casein in milk. Different from the other CSN1S1*A proteins, CSN1S1*A3 carries a transversion from A to C at position CHR6:85987198 (exon 10) leading to the amino acid exchange from Gln77 to Pro77 (Mestawet et al., 2013). CSN1S1*A is the predominant protein variant in most Indian breeds (frequency >0.7) (Kumar et al., 2007; Rout et al., 2010) and in a few breeds from Spain, Italy and France (frequency >0.5, Supplementary Table S2) (Ramunno et al., 1991).
The CSN1S1*B protein is divided into four variants (CSN1S1*B1, CSN1S1*B2, CSN1S1*B3, CSN1S1*B4) (Grosclaude and Martin, 1997). All CSN1S1*B variants differ from the reference protein through the substitution of Gln77 to Glu77 (Brignon et al., 1989), a substitution that is associated with the presence of a phosphate group on Serine at position 75 (Brignon et al., 1989). Among all α-S1-casein variants, CSN1S1*B1 is considered the ancestor allele showing the closest homology to its bovine and ovine counterparts (Grosclaude and Martin, 1997). CSN1S1*B1 and CSN1S1*B′ are identical proteins; the only difference is a synonymous SNP in the codon for Ser66 in exon 9 in CSN1S1*B′ (Caroli et al., 2007). The caseins CSN1S1*B2, CSN1S1*B3 and CSN1S1*B4 carry the additional T to C transition at position CHR6:85982615 (exon 4), leading to the substitution of Leu16 to Pro16 (Grosclaude and Martin, 1997; Cosenza et al., 2008). CSN1S1*B3 and CSN1S1*B4 carry the G to A transition at position CHR6:85988705 (exon 12) changing Arg100 to Lys100 (Grosclaude and Martin, 1997; Cosenza et al., 2008). CSN1S1*B4 differs from CSN1S1*B3 by the nucleotide substitution A to G at position CHR6:85993465 (exon 17) altering Thr195 to Ala195 (Grosclaude and Martin, 1997; Cosenza et al., 2008). With respect to names, it is necessary to mention that firstly, CSN1S1*B2 was named simply CSN1S1*B in old papers (Boulanger et al., 1984; Brignon et al., 1989), and secondly, the variant CSN1S1*E had also been named CSN1S1*B− (Grosclaude et al., 1987). CSN1S1*B protein variants can be found at high frequency (>0.5) in diverse breeds across the world (Supplementary Table S2).
CSN1S1*C and CSN1S1*C1 differ from the reference by five and four amino acid substitutions, respectively. The two variants carry the amino acid substitutions His8 to Ile8, Leu16 to Pro16, Gln77 to Glu77, and Arg100 to Lys100. The additional substitution Thr195 to Ala195 occurs only in CSN1S1*C (Leroux et al., 1990; Rahmatalla et al., 2021).
CSN1S1*D is characterized by a deletion of 11 amino acids at positions 59 to 69 which comprises the major phosphorylation site of the protein (Brignon et al., 1990). The deletion of the single nucleotide C at position CHR6:85986426 in exon 9 is a splice-site mutation which causes the exclusion of exon nine in the mature RNA assembly which also reduces the synthesis rate of the protein (Mahé and Grosclaude, 1989; Brignon et al., 1990). Besides the significant deletion, CSN1S1*D carries the substitutions Leu16 to Pro16 and Gln77 to Glu77, which also occur in most CSN1S1*B and CSN1S1*C protein variants (Mahé and Grosclaude, 1989; Brignon et al., 1990).
CSN1S1*E (formerly CSN1S1*B′) has the same amino acid variant as CSN1S1*B4. Besides that, CSN1S1*E has an insertion of 457–458 bp in exon 19, which is expressed but untranslated (Jansàpérez et al., 1994). This insert is a truncated long interspersed repeated element (LINE), containing part of the ORF-2, the 3′ UTR, and the original retrotransposon’s poly (A) tail. The mutant 3′ UTR likely controls the mRNA stability of transcripts and/or the translation rate which was seen as low casein content in goat milk of CSN1S1*E carriers (Jansàpérez et al., 1994). The CSN1S1*E allele is highly frequent (>0.5) in Saanen goats worldwide (Tadlaoui Ouafi et al., 2002; Maga et al., 2009), and in other breeds from Spain, France, and the United States (Supplementary Table S2) (Grosclaude et al., 1994; Jordana et al., 1996; Tadlaoui Ouafi et al., 2002; Caravaca et al., 2008; Maga et al., 2009).
CSN1S1*F contains the amino acid variants Pro16 and Glu77 as in CSN1S1*D (Grosclaude et al., 1987; Brignon et al., 1990). Most significant in CSN1S1*F is the lacks of 37 amino acids from position 59 to 95 as the consequence of a frameshift mutation due to a single nucleotide deletion in exon 9 (Grosclaude et al., 1987; Brignon et al., 1990). The deletion starts exactly at the same position and with the same mutation as in CSN1S1*D. Two additional insertions of 11 and 3 base pairs in intron nine are likely responsible for the skipping of exons 9, 10 and 11 in CSN1S1*F (Leroux et al., 1992). The content of this shortened α-S1-casein protein variant in milk is low, likely as a result of low efficiency and accuracy of the splicing machinery (Martin et al., 1999), which might result from either a reduced translation rate or intracellular degradation of the incomplete protein. Several breeds that originate from the Alpine region carry the CSN1S1*F allele at a frequency above 0.40 (Grosclaude et al., 1987; Ramunno et al., 1991). A higher expression of CSN1S1*F compared to CSN1S1*N might be attributed to a mutation in the promotor region creating an extra putative activator protein 1 (AP-1) binding motif (Ramunno et al., 2005). However, experimental validation does not exist. Particular high frequency >0.90 of CSN1S1*F was found in Toggenburg goats from America and Orobica goats from Italy (Supplementary Table S2) (Caroli et al., 2006; Maga et al., 2009).
Protein variant CSN1S1*G is missing 13 amino acids from position 14 to 26. The G/A transition at position CHR6:85982647 in the 5′ splice site at the beginning of intron 4 leads to the skipping of exon 4 (Martin and Leroux, 1994). The occurrence of the CSN1S1*G allele is associated with low α-S1-casein content in milk (Martin and Leroux, 1994).
CSN1S1*H differs from the reference variant CSN1S1*A by a single amino acid substitution at position 1, Arg1 to Lys1. This protein was classified as a strong variant (Chianese et al., 1997). For the Italian Garganica breed the frequency of 0.07 was reported (Sacchi et al., 2005).
For the protein variants CSN1S1*G, CSN1S1*I, CSN1S1*J, CSN1S1*K, CSN1S1*L, and CSN1S1*M there is information available for sequence variants only, but not for allele frequency in any breed. CSN1S1*I had the same amino acids sequence as variant CSN1S1*A. However, CSN1S1*I had only about half of the amount of casein in the milk than CSN1S1*A (Chianese et al., 1997), which led to the differentiation between CSN1S1*I and CSN1S1*A. The causal mutation is unidentified.
CSN1S1*J carries the combination of the amino acids substitutions Gln77 to Glu77 and Arg100 to Lys100, which are also found in other protein variants (Rahmatalla et al., 2021). CSN1S1*J detected in Bezoar ibex. This variant is similar to CSN1S1*B1 which is consider the ancestor allele, but it contains the Arg100 to Lys100 substitution (Rahmatalla et al., 2021).
The CSN1S1*K protein variant differs from the reference by the unique amino acid substitution from Ile44 to Val44 as a results of a to G transition at position CHR6:85984154 (exon 7) (Rahmatalla et al., 2021). The CSN1S1*K was found in wild Alpine ibex (Rahmatalla et al., 2021).
CSN1S1*L harbors the two amino acids Pro16 and Glu77 instead of Leu16 and Gln77 in the reference sequence and additionally the amino acid substitution from Arg90 to His90 (Chianese et al., 1997). The transition of G to A at CHR6:85987327 in exon 11 leads to the substitution of Arg90 to His90.
CSN1S1*M carries the Leu16 to Pro16 substitution, which occurs also in other protein variants. CSN1S1*M also contains the amino acid substitution of Ser66 to Leu66 which results from a C/T transition at CHR6:85986426 in exon 9 (Chianese et al., 1998; Bevilacqua et al., 2002).
CSN1S1*N, CSN1S1*01 and CSN1S1*02 are null variants resulting in truncated gene products and subsequently no casein in the milk. CSN1S1*N descends from protein variant CSN1S1*M, but contains also the nucleotide deletion of a single C at CHR6:85986426 in exon 9 that also occurs in variants CSN1S1*D and CSN1S1*F, where it causes exon skipping. However, in CSN1S1*N the one-nucleotide deletion leads to a premature stop codon. The amount of the truncated transcript is reduced to one-third and the amount of α-S1-casein in milk in homozygous carriers is null (Ramunno et al., 2002; Ramunno et al., 2005). CSN1S1*01 is a truncated protein where the last seven exons are missing. The truncation results from a deletion of about 8.5 kb starting from nucleotide 181 of intron 12. In homozygous carriers, α-S1-casein is absent in milk (Grosclaude et al., 1987; Leroux et al., 1990; Cosenza et al., 2003). CSN1S1*02 is another null variant, leading to the absence of the respective protein in milk. Martin et al. (1999) proposed a large insertion that has not been identified so far. Albeit the goats carrying the null alleles homozygous do not have contain α-S1-casein in their milk, the Indian breeds Ganjam and Local MP have an allele frequencies of 0.40 and 0.45, respectively (Rout et al., 2010).
Goat milk that lacks α-S1-casein has a higher pH value and a low content of chemical components like total solids, protein, casein, Nitrogen, fat, and Phosphorus compared to milk with strong α-S1-casein variants (Ambrosoli et al., 1988; Pirisi et al., 1994; Clark and Sherbon, 2000; Devold et al., 2011; Johansson et al., 2015; Marković et al., 2018). In addition, milk containing α-S1-casein null variants is characterized by large casein micelles with weak gel firmness and long coagulation time (Devold et al., 2011; Johansson et al., 2015). The gross yield of cheese and the content of fat and non-fat solids as well as the fat to dry matter ratio and the total solids recovery rate from milk that lacks α-S1-casein is low (Pirisi et al., 1994). Furthermore, textural parameters of cheese such as hardness and plasticity were low in cheese that is produced from goat milk lacking α-S1-casein (Pirisi et al., 1994). In contrast, milk containing strong α-S1-casein variants produce firmer curd that is associated with high gross yield of cheese with a better composition (Devold et al., 2011; Johansson et al., 2015).
Beta-casein is the most hydrophobic casein as a result of a highly charged N-terminal domain and an anionic phosphoserine cluster (Swaisgood, 2003). Beta-casein is the major protein in goat milk (Clark and Mora Garcia, 2017) comprising up to 50% of the total casein content. In comparison, human milk casein contains 60%–70% and bovine milk casein 36% β-casein (Guo, 2009; Elagamy, 2016). Beta-casein is a primary source of essential amino acids and facilitates the mineral transport for infants (Sadler and Smith, 2013). The peptides derived from β-casein during digestion have antihypertensive and immune-stimulating properties (Silva et al., 2006; Lamothe et al., 2007).
The CSN2 gene is located on chromosome 6 between 86.006 and 86.015 Mb (NC_030813, goat genome reference version LWLT01) (Bickhart et al., 2017). The 9,072 bp long gene consists of nine exons ranging in size from 24 (exon 5) to 492 bp (exon 7) (Roberts et al., 1992; Hayes and Petit, 1993). The protein contains 222 amino acids, which comprise 207 functional amino acids and 15 amino acids of a signal peptide (XP_005681778 (NCBI database).
So far, eight protein variants have been identified (Table 1). Variants CSN2*A, CSN2*B, CSN2*C, CSN2*D, and CSN2*E are “strong” variants associated with high β-casein content in milk (5 g/L per allele) (Roberts et al., 1992; Mahé and Grosclaude, 1993; Neveu et al., 2002; Galliano et al., 2004; Cosenza et al., 2005; Caroli et al., 2006). The variants CSN2*C2 and CSN2*F1 were associated with intermediate β-casein content (3.3 and 2.7 g/L per allele, respectively) (Nicolai et al., 2021). The two null alleles CSN2*0 and CSN2*0′ are associated with a lack of β-casein in milk (Persuy et al., 1999). In the CSN2 gene, the existence of the strong variants were associated with the highest milk protein content and yield in Sarda goats from Italy (Vacca et al., 2014).
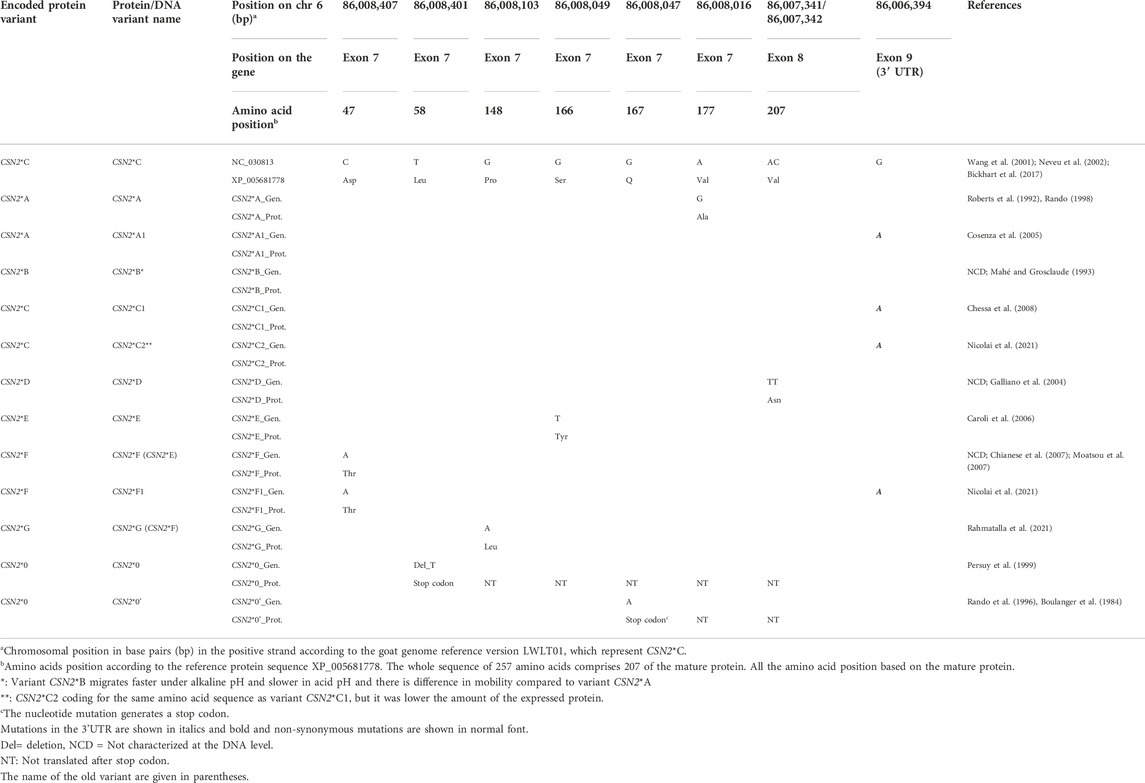
TABLE 1. Nucleotide and amino acid variants within the gene and mature protein of the CSN2.
The reference protein is variant CSN2*C (Wang et al., 2001; Neveu et al., 2002). CSN2*A differs from the reference by the amino acid substitution of Val177 to Ala177 (Neveu et al., 2002) as a result of an A/G transition at position CHR6:86008016 in exon 7 (Roberts et al., 1992). CSN2*A is considered the ancestral allele in goats because the amino acid alanine at position 177 is also found in sheep, cattle and buffalo (Chessa et al., 2005). The variant CSN2*A1 differs from CSN2*A by an additional G/A SNP in exon 9, which belongs to the 3′ untranslated region (Cosenza et al., 2005). CSN2*A was predominant with a frequency above 0.90 in Girgentana and Argentata dell’Etna from Italy, and in most Indian goats (Supplementary Table S3) (Marletta et al., 2005; Rout et al., 2010). A high frequency above 0.70 was evident in French Creole goats, West African breeds, and Buren goat from Germany (Supplementary Table S3) (Mahé and Grosclaude, 1993; Caroli et al., 2007; Kupper et al., 2010).
The CSN2*B variant was characterized via isoelectric protein electrophoresis, where CSN2*B migrates faster than CSN2*A (Mahé and Grosclaude, 1993). The mobility differences between CSN2*A and CSN2*B might be due to an extra phosphate group in CSN2*B (Mahé and Grosclaude, 1993).
CSN2*C1 and CSN2*C2 encode the same protein CSN2*C, the reference protein sequence. The protein variant CSN2*C is predominant with a frequency of ≥0.70 in most Italian goats, Angora and Hair goats from Turkey, Rajasthan and South goats from India as well as Banat White goats from Romania (Supplementary Table S3) (Chessa et al., 2005; Caroli et al., 2006; Chessa et al., 2007; Kusza et al., 2016). The two subtypes differ in a synonymous transition of G to A in the 3′ untranslated region of exon 9 in CSN2*C (Chessa et al., 2008; Nicolai et al., 2021). This single nucleotide polymorphism likely leads to lower amount of ß-casein if the animals is homozygouse for the CSN2*C2 subtype compared to CSN2*C1. An impact on the mRNA stability and or translation rate might be causal for the low amount of the protein (Nicolai et al., 2021). The variant CSN2*C1 was found with a frequency of more than 0.10 in Bunte Deutsche Edelziege and Weiße Deutsche Edelziege goats from Germany and Italian Girgentana goats (Kupper et al., 2010; Tortorici et al., 2014).
CSN2*D differs from the reference protein in the amino acid substitution from Val207 to Asn207 (Galliano et al., 2004). DNA sequence comparisons show a AC/TT substitution at positions CHR6:86007341 and CHR6:86007342 in exon 8 that is responsible for the amino acid change.
CSN2*E is characterized by the amino acid exchange from Ser166 to Thr166 which results from the G/T transversion at position CHR6:86008049 in exon 7 (Caroli et al., 2006). CSN2*E was found at a low frequency (0.08) only in Italian Frisa goats (Caroli et al., 2006).
The protein variant CSN2*F differs from the reference by the substitution of Asp47 to Thr47 (Chianese et al., 2007; Moatsou et al., 2007). The underlying SNP is a C/A transversion at position CHR6:86008407 in exon 7. By mistake, this protein variant had been named CSN2*E by Caroli et al. (2006) although CSN2*E already existed. It was correctly renamed as CSN2*F by Martin et al. (2013). CSN2*F1 and CSN2*F represent the same β-casein protein. CSN2*F1 differs from CSN2*F by a synonymous transition G/A in the 3′ untranslated region in exon 9 (Nicolai et al., 2021).
CSN2*G (previously CSN2*F) is characterized by the amino acid substitution from Pro148 to Leu148 and found in Wild Alpine ibex (Rahmatalla et al., 2021). The nucleotide mutation is a G/A substitution at position CHR6:86008103 in exon 7 (Rahmatalla et al., 2021).
Observations of milk lacking β-casein led to the conclusion that at least two null alleles exist (Dall’Olio et al., 1989; Mahé and Grosclaude, 1989). Variant CSN2*0 has one nucleotide deletion (T/-) at position CHR6:86008401 in exon 7 which introduces a premature stop codon at position 58 in the mature protein (Persuy et al., 1999). The CSN2*0 null variant was detected in a frequency below 0.10 in Creole goats from France and some Italian and Indian goats (Supplementary Table S3) (Mahé and Grosclaude, 1993; Marletta et al., 2005; Sacchi et al., 2005; Rout et al., 2010; Vacca et al., 2014). However, the frequency was 0.19 in Italian Garganica goats (Albenzio et al., 2009). CSN2*0′ carries the single nucleotide substitution C to T at position CHR6:86008047 in exon 7, which introduces a premature stop codon at position 167 of the mature protein (Rando et al., 1996). The CSN2*0′ null variant was found at a frequency below 0.10 in some Italian breeds and in Arbi goats from Tunis (Supplementary Table S3) (Chessa et al., 2005; Gigli et al., 2008; Vacca et al., 2009a; Tortorici et al., 2014). In CSN2*0′, another SNP in the promoter region was detected which negatively affects the transcriptional activity of the gene (Cosenza et al., 2007; Cosenza et al., 2016).
No information was available for the allele frequency of CSN2*C2, CSN2*D, CSN2*F, CSN2*F1, and CSN2*G.
The β-caseins in milk are primarily classified into the two types A1 and A2. A1 and A2 differ in the generation of β-casomorphin-7 (BCM-7) peptides during digestion (Jianqin et al., 2016). The peptide β-casomorphin-7 gained attention because it was associated with gastrointestinal disturbances, ischemic heart disease, arteriosclerosis, type 1 diabetes, and sudden infant death syndrome in humans (Birgisdottir et al., 2002; Tailford et al., 2003; Ho et al., 2014). The peptide β-casomorphin-7 is uniquely derived from the digestion of β-casein type A1, but not from A2. Different from cow milk, goat milk mainly contains the β-casein type A2 (Cieslinska et al., 2012). Therefore, β-casein of the type A2 contributes to the healthy properties of goat milk because it does not produce the peptide β-casomorphin-7 during digestion (De Noni and Cattaneo, 2010).
It is worth noting that with the increasing awareness of consumers’ behavior and their interest in healthy eating and functional food, different companies launched β-casein type A2 milk from cattle on the market. Costs are associated with the separate handling of this type of milk during processing. Since goat milk naturally contains β-casein type A2, this milk plays a crucial role in providing functional milk and milk products to the food market. A positive benefit for human health is realized here without additional marketing costs.
The alpha-S2-casein protein plays an important role in the transport of calcium phosphate. The alpha-S2-casein content in goat milk is relatively high comprising about 19%, while it constitutes up to 10% in bovine milk (McMahon and Brown, 1984). The antimicrobial peptide cathelicidin-1 which derives from the digestion of alpha-S2-casein has the activity to inhibit the growth of Gram-positive and Gram-negative bacteria (Zucht et al., 1995; López-Expósito et al., 2008; Gigli, 2016).
The CSN1S2 gene resides on chromosome 6 between 86.077 and 86.094 Mb, which is about 16,695 bp long and consists of 18 exons (NCBI, accession no. NC_030813). The gene encodes 223 amino acids comprising 208 functional amino acids and 15 amino acids of a signal peptide (https://www.uniprot.org/uniprot/P33049).
The DNA [NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)] and protein reference sequence (XP_013820127.2, NCBI database) represent the CSN1S2*A variant (Boulanger et al., 1984; Bouniol et al., 1993; Bickhart et al., 2017). CSN1S2*A is considered the ancestor variant for goat according to the evolutionary path from sheep and cattle (Sacchi et al., 2005). This variant is common with a frequency of ≥0.70 in distinct Saanen goats, most Italian, German and Indian goats, Dwarf goats from Nigeria, and Nubian goats from Sudan (Supplementary Table S4) (Boulanger et al., 1984; Bouniol et al., 1994; Ramunno et al., 2001b; Erhardt et al., 2002; Marletta et al., 2004; Chiatti et al., 2005; Marletta et al., 2005; Caroli et al., 2007; Chessa et al., 2007; Kupper et al., 2010; Rout et al., 2010; Yue et al., 2013).
So far, 14 alpha-S2-casein protein variants have been identified (CSN1S2*A, CSN1S2*B, CSN1S2*C, CSN1S2*D, CSN1S2*E, CSN1S2*F, CSN1S2*G, CSN1S2*H, CSN1S2*I, CSN1S2*J, CSN1S2*K, CSN1S2*0, truncated sub-variant A, and truncated sub-variant E) (Table 2) (Boulanger et al., 1984; Bouniol, 1993; Bouniol et al., 1994; Veltri et al., 2000; Ramunno et al., 2001a; Ramunno et al., 2001b; Lagonigro et al., 2001; Erhardt et al., 2002; Cunsolo et al., 2006; Rahmatalla et al., 2021). While the protein variants CSN1S2*A, CSN1S2*B, CSN1S2*C, CSN1S2*E, CSN1S2*F, and CSN1S2*G are associated with normal amounts of α-S2-casein of about 2.5 g/L per allele (Boulanger et al., 1984; Grosclaude et al., 1987; Bouniol et al., 1994; Ramunno et al., 2001a), CSN1S2*D is associated with a reduced amount of about 1.5 g/L per allele (Ramunno et al., 2001a). In CSN1S2*0 homozygous goats, alpha-S2-casein was not detectable (Ramunno et al., 2001b).
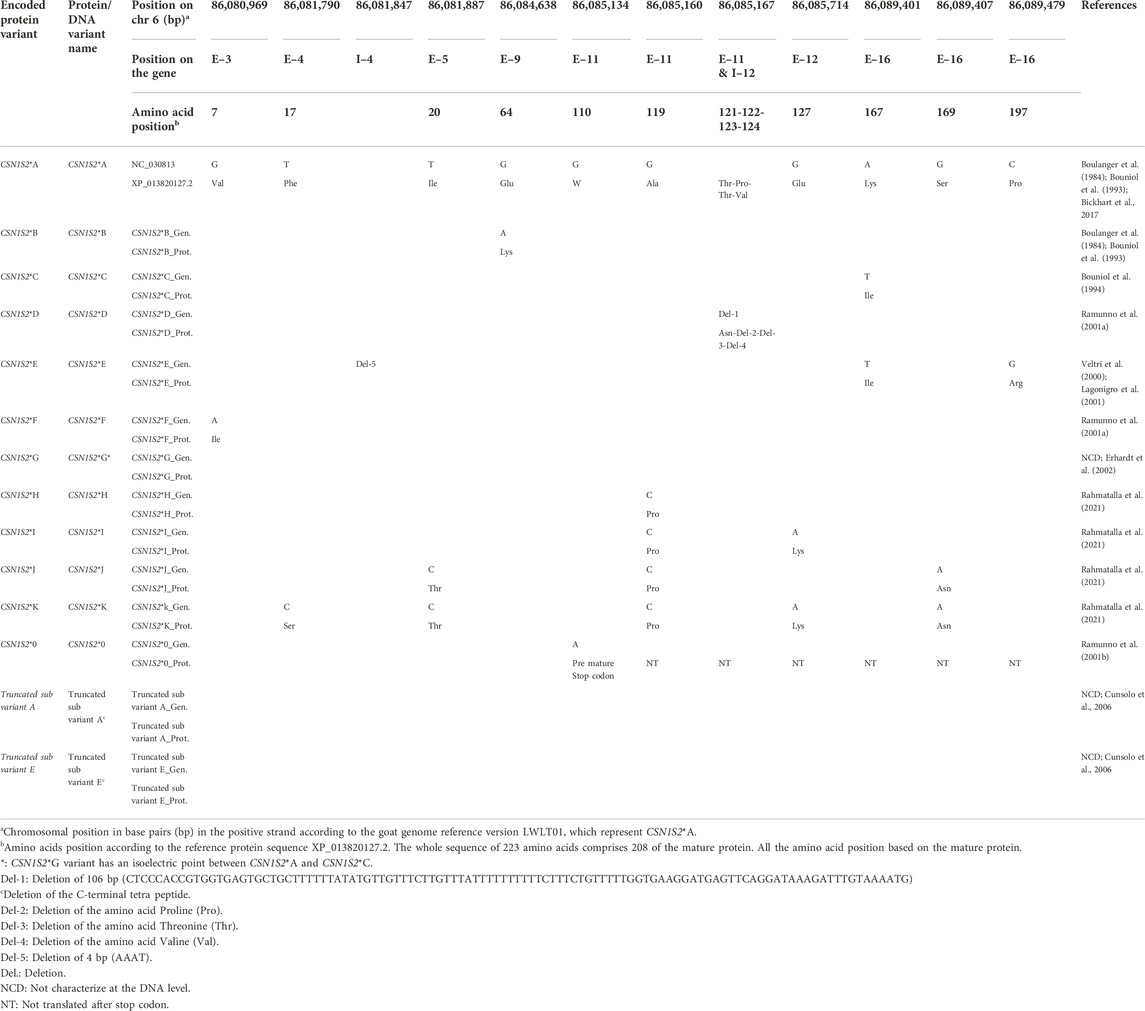
TABLE 2. Nucleotide and amino acid variants within the gene and mature protein of the CSN1S2.
CSN1S2*B differs from the reference (CSN1S2*A) by a G/A substitution at CHR6:86084638 in exon 9, which is responsible for the amino acid substitution Glu64 to Lys64 that affects a phosphorylation site (Boulanger et al., 1984; Bouniol et al., 1994). CSN1S2*B occurs at a frequency above 0.40 in Egyptian and Lithuanian Native goats (Othman and Ahmed, 2006; Baltrėnaitė et al., 2009).
CSN1S2*C distinguishes from the reference by the transversion A/T at CHR6:86089401 in exon 16 which changing Lys167 to Ile167 (Bouniol et al., 1994). This variant occurs at a frequency ≥0.40 in Vallesana and Roccaverano Italian goats, Dwarf Cameroon goats, Borno goats from Nigeria, Hair goats from Turkey, as well as Maharashtra and Rajasthan from India (Supplementary Table S4) (Sacchi et al., 2005; Caroli et al., 2007; Chessa et al., 2007).
CSN1S2*D contains a 106 bp deletion comprising the last 11 bp of exon 11 and the first 95 bp of the following intron (Ramunno et al., 2001a). This deletion is responsible for the lack of the three codons Pro122, Thr123, and Val124. Furthermore, the last undeleted nucleotide (A) of exon 11 together with the two nucleotides (AT) of the following intron generate a codon for the amino acid Asn121 instead of Thr121.which is followed by a new GT dinucleotide splicing donor site. The rare CSN1S2*D variant was found in Italian Argentata dell’Etna and Girgentana goats and Hungarian goats with a frequency of 0.01 (Kusza et al., 2007; Gigli et al., 2008).
CSN1S2*E is characterized by a C/G mutation at CHR6: 86089479 in exon 16, leading to the amino acid exchange Pro197 to Arg197 (Chianese et al., 1998; Veltri et al., 2000; Lagonigro et al., 2001). Additionally, the of A/T transversion of CSN1S2*C changing Lys167 to Ile167 is present also in CSN1S2*E. Furthermore, a deletion of 4 bp (AAAT) was identified in intron 4 (Lagonigro et al., 2001). CSN1S2*E was found at low frequency (below 0.11) in some Italian goats, Hair goats from Turkey, and South goats from India (Sacchi et al., 2005; Caroli et al., 2006; Chessa et al., 2007; Gigli et al., 2008; Albenzio et al., 2009; Vacca et al., 2009b; Palmeri et al., 2014; Vacca et al., 2014). In addition to the full-length protein variants CSN1S2*A and CSN1S2*E, two truncated sub-variants were identified, which lack the C-terminal tetrapeptide (Gln199, Pro200, Leu201, Gln202, Pro203, Leu204). These truncated variants likely originate from differential splicing of pre-messenger RNA during the translation process of CSN1S2*A and CSN1S2*E variants (Cunsolo et al., 2006).
CSN1S2*F differs from the reference by a G/A transition at CHR6:86080969 in exon 3 substituting Val7 to Ile7 (Ramunno et al., 2001a). CSN1S2*F is common (frequency ≥0.40) in Italian Garganica, Orobica, Frisa, Sarda goats, and Weiße Deutsche Edelziege (Sacchi et al., 2005; Caroli et al., 2006; Vacca et al., 2009b; Kupper et al., 2010).
There is not much known about the nucleotide and protein sequence of the variant CSN1S2*G. This protein has an isoelectric point between CSN1S2*A and CSN1S2*C (Erhardt et al., 2002).
CSN1S2*H differs from CSN1S2*A by a G/C substitution at CHR6:86085160 in exon 11 leading to the substitution of Ala119 by Pro119 (Rahmatalla et al., 2021). This variant is predominant in Sudanese goats (Supplementary Table S4) (Rahmatalla et al., 2021).
CSN1S2*I contains the same mutations that distinguish CSN1S2*G from the reference and additionally a G/A substitution at CHR6:86085714 in exon 12 that is responsible for the change of Glu127 to Lys127 (Rahmatalla et al., 2021). So far, CSN1S2*I has been detected in Nubian and Desert goats only at a frequency of 0.21 and 0.10, respectively (Rahmatalla et al., 2021).
CSN1S2*J diverges from the reference by a T/C substitution at CHR6:86081887 in exon 5 and G/A substitution at CHR6:86089407 in exon 16 that account for the substitutions Ile20 to Thr20 and Ser169 to Asn169, respectively (Rahmatalla et al., 2021). In addition, the mutation which distinguishes CSN1S2*H from CSN1S2*A is present also in CSN1S2*J (Rahmatalla et al., 2021).
CSN1S2*K carries the same three amino acid substitution that occur in CSN1S2*J (Rahmatalla et al., 2021) and additionally the amino acid substitutions Phe17 to Ser17 and Glu127 to Lys127 as a result of the nucleotide substitutions of T to C at CHR6:86081790 in exon 4 and G to A at CHR6:86085714 in exon 12, respectively (Rahmatalla et al., 2021) The CSN1S2*J and CSN1S2*K were identified in wild Nubian ibex (Rahmatalla et al., 2021).
CSN1S2*0 carries a stop codon at the amino acid position 110 as a result of a G to A transition at CHR6:86085134 in exon 11, which disrupts the codon for Try110 (Ramunno et al., 2001b). As a consequence, a truncated non-functional protein of 109 instead of 208 amino acids is generated. Homozygous CSN1S2*0 carriers do not have α-S2-casein in milk. Nevertheless, the allele frequency in the Indian breeds Ganjam and Local MP is 0.55 and 0.48, respectively (Rout et al., 2010).
No information was available for the allele frequency of CSN1S2*J, CSN1S2*K, and the truncated sub-variants of CSN1S2*A and CSN1S2*E.
CSN1S2 variants affected the milk protein content and milk yield in Sarda goats from Italy (Vacca et al., 2014). In Sarda goats, CSN1S2*AC genotyped animals produce the highest fat and protein yield, which affect the technological properties for cheese processing, while animals carrying the CSN1S2*CF genotype have the highest daily milk yield (Vacca et al., 2014). Interestingly, the variants CSN1S2*A, CSN1S2*C, and CSN1S2*F were associated with a high amount of α-S2-casein in milk and produced 2.5 g/l per allele (Boulanger et al., 1984; Bouniol et al., 1994; Ramunno et al., 2001a). In Chinese dairy goats, animals carrying the CSN1S2*FF genotype have higher milk fat and total solids content than CSN1S2*AA and CSN1S2*AF genotypes, but do not have lower milk yield (Yue et al., 2013). However, Lan et al. (2005) found significant lower milk yield of Xinong Saanen goats carrying the CSN1S2*FF genotypes compared to other genotypes. The contradictory effects of CSN1S2*FF genotyped animals might be attributed to the small sample size in the study of Lan et al. (2005). The variation in the α-S2-casein level in milk is associated with the unique physicochemical characteristic of milk. Thereby, α-S2-casein levels affect the cheese-making properties (Selvaggi and Tufarelli, 2011).
Kappa-casein (k-casein) is a calcium insensitive protein that forms a protective layer around the calcium-sensitive caseins α-S1, α-S2, and β, resulting in stable casein micelles (Quaglia and Gennaro, 2003). The formation, stabilization, and aggregation of casein micelles affect the physical characteristics and technological properties of milk (Alexander et al., 1988; Glantz et al., 2010). Kappa-casein is the only casein that is highly glycosylated (Haratifar and Guri, 2017). The mature kappa-casein protein has a labile peptide bond whose cleavage by chymosin or rennin produces a soluble hydrophilic glycopeptide called caseino-macropeptide (amino acids 106–171) and an insoluble peptide or para-k-casein (amino acids 1–105) (Yahyaoui et al., 2003). The caseino-macropeptide is important for milk coagulation.
Kappa-casein comprises about 20% in goat and sheep milk, in comparison, only 12–14% in cow milk (Fox and Brodkorb, 2008; Molik et al., 2012). Kappa-casein is essential for lactation. For example, mice lacking this gene (CSN3−/−) fail to lactate because of destabilization of the micelles in the lumen of the mammary gland (Shekar et al., 2006). Kappa-casein contributes to the nutritional properties of milk. The protein has beneficial effects on oral health (Vacca-Smith and Bowen, 1995; Johansson and Lif Holgerson, 2011) and the digestion of k-casein generates an antimicrobial peptide (Malkoski et al., 2001).
The CSN3 gene is located on chromosome 6 between 86.197 and 86.211 Mb [NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)]. The gene spans 14,114 bp and comprises five exons (Martin et al., 2002). The protein contains 192 amino acids which include 21 amino acids of the signal peptide and 171 amino acids of the mature protein (Mercier et al., 1976; Coll et al., 1993). The mature protein is encoded in only two exons, exons 3 (9 amino acids) and exon 4 (162 amino acids) (Yahyaoui et al., 2003).
The CSN3 gene in goats is highly polymorphic. To date, 35 variants are known that yield 24 protein variants and 11 synonymous mutations which are detectable only at the DNA level (Supplementary Table S5). In this review we follow the new nomenclatures of the k-casein variants recently described by Gautam et al. (2019). The DNA (NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)) and protein reference sequences of CSN3 (NP_001272516, NCBI database) represent the protein variant CSN3*B as the reference (Yahyaoui et al., 2001; Jann et al., 2004). We supplement the information with nucleotide positions in the coding sequence that is given in the reference X60763 (NCBI database) (Supplementary Table S5) (Coll et al., 1993) to facilitate the comparison between the position in the new goat reference genome of k-casein variants with previous studies that used the position in the coding sequence to name variants.
The reference protein CSN3*B is the predominant k-casein variant that occurs at a frequency >0.70 in most Italian and East Africa goats, Toggenberg and Buren goats from Germany, and Polish Saanen, fawn and white improved goats (Supplementary Table S6) (Chessa et al., 2003; Yahyaoui et al., 2003; Sacchi et al., 2005; Caroli et al., 2006; Kiplagat et al., 2010; Kupper et al., 2010; Strzelec and Niżnikowski, 2011; Rahmatalla et al., 2021).
The difference between CSN3*A and CSN3*B is the amino acid substitution Ile119 to Val119 which is the result of the transition of A to G at CHR6:86209184 in exon 4 (Coll et al., 1993; Yahyaoui et al., 2001). CSN3*A occurs also at a frequency ≥0.7 in Ionica and Montefalcone from Italy, Bunte and Weise Deutsche Edelziege, and Thueringer Waldziege from Germany as well as Ardi and Syrian goats from Saudi Arabia (Supplementary Table S6) (Caroli et al., 2001; Angiolillo et al., 2002; Kumar et al., 2009; El-Shazly et al., 2017). CSN3*A′ and CN3*A″ encode the same protein variant CSN3*A, but, carry the synonymous mutations T to C at CHR6:86209207 and C to T at CHR6:86209258 in exon 4, respectively (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*A″ was found at low frequency in goats from Kenya and Maasai from Ethiopia. In that study, CSN3*A″ was named CSN3*O (Kiplagat et al., 2010).
CSN3*B′, CSN3*B″, CSN3*B‴ have the equivalent CSN3*B protein variant. In addition to the transition of G to A at CHR6:86209184 that alters the amino acid Val119 to Ile119, CSN3*B′, CSN3*B″, CSN3*B″ carry the synonymous mutations C to T at CHR6:86208883, C to T at CHR6:86209003, and A to G at CHR6:86209297 in exon 4, respectively (Supplementary Table S5) (Chessa and Bolla, 2003; Yahyaoui et al., 2003; Jann et al., 2004; Prinzenberg et al., 2005; Kiplagat et al., 2010). Previously, CSN3*B, CSN3*B′, CSN3*B″, and CSN3*B‴ had the names CSN3*D, CSN3*D′, CSN3*D″, and CSN3*P, respectively (Jann et al., 2004; Kiplagat et al., 2010). The homozygous CSN3*B genotype was associated with a higher milk protein content in Saanen goats (Catota-Gómez et al., 2017) and longer milk coagulation time in Spanish Murciano-Granadina goats compared to CSN3*A carriers (Caravaca et al., 2011).
CSN3*C differs from the reference variant CSN3*B in the three amino acid substitutions Val65 to Ile65, Ala156 to Val156, and Ser159 to Pro159 (Yahyaoui et al., 2001). The amino acid substitutions occurred as a result of a G to A, C to T, and T to C transitions at position CHR6:86209022, CHR6:86209296, and CHR6:86209304 in exon 4 (Yahyaoui et al., 2001), respectively. CSN3*C occurs at a frequency below 0.20 in some breeds from Italy, Canaria Spanish goats, goats from Germany and Turkey, Saanen goats worldwide, and Lithuania goats (Supplementary Table S6) (Chessa et al., 2003; Yahyaoui et al., 2003; Prinzenberg et al., 2005; Sacchi et al., 2005; Caroli et al., 2006; Gigli et al., 2008; Baltrėnaitė et al., 2009; Vacca et al., 2014; Catota-Gómez et al., 2017). CSN3*C′ differs from CSN3*C through a synonymous mutation in codon 131 (Prinzenberg et al., 2005).
In the CSN3*D (previously named CSN3*B), the amino acid substitutions Val65 to Ile65 and Ser159 to Pro159 that exist in CSN3*C were also found. In addition, CSN3*D carries the Gln44 to Arg44 substitution because of an A to G transition at CHR6:86208960 in exon 4 (Caroli et al., 2001; Yahyaoui et al., 2001). This variant occurs at a frequency of less than 0.40 in Italian and German goats and Ardi goats from Saudi Arabia (Supplementary Table S6).
CSN3*D′ differs from CSN3*D by a synonymous mutation in codon 56 and occurs at a frequency of 0.01 in Girgentana goats from Italy (Gerlando et al., 2015).
The CSN3*E protein variant differs from the reference by the amino acid substitution of Asp90 by Gly90 as the results of an A to G transition at position CHR6:86209098 in exon 4 (Angiolillo et al., 2002). CSN3*E occurs in Montefalcone goats from Italy at a frequency up to 0.45 (Angiolillo et al., 2002; Yahyaoui et al., 2003).
CSN3*F differs from the reference protein in the amino acid substitution of Ser159 by Pro159 which occurred due to the T to C transition at CHR6:86209304 in exon 4 (Yahyaoui et al., 2003). This variant was detected in breeds from Italy at a frequency below 0.15 and in Wild Spanish at a frequency of 0.98 (Yahyaoui et al., 2003). The variants CSN3*F′ and CSN3*F encode the same CSN3*F protein. CSN3*F differs from CSN3*F′ by a synonymous mutation in codon 77. CSN3*F′ occurs at a low frequency (0.01) in Raigarh goats from India (Gautam et al., 2019).
CSN3*G is characterized by G to A and T to C mutations at CHR6:86209022, and CHR6:86209304 in exon 4, respectively, in comparison to the reference variant CSN3*B. The mutations cause the amino acid exchange Val65 to Ile65 and Ser159 to Pro159, respectively (Yahyaoui et al., 2003). An addition synonymous mutation is present in codon 43 (Yahyaoui et al., 2003). CSN3*G was found at a frequency of less than 0.15 in a few breeds from Italy and Hair and Angora goats from Turkey (Yahyaoui et al., 2003; Prinzenberg et al., 2005; Gigli et al., 2008; Gerlando et al., 2015). Variant CSN3*G′ has the same amino acid substitutions as variant CSN3*G, but does not contain the synonymous mutation in codon 43 (Supplementary Table S5). The name CSN3*G′ was introduced by Gautam et al. (2019); before that this variant had been named CSN3*F or CSN3*L (Jann et al., 2004; Kiplagat et al., 2010). CSN3*G″ contains the same amino acid substitution as variant CSN3*G and in addition two synonymous mutations in codon 43 and 56 (Vacca et al., 2014; Gerlando et al., 2015; Gautam et al., 2019). The name CSN3*G″ was also introduced by Gautam et al. (2019) before this variant had been named CSN3*N and CSN3*S (Vacca et al., 2014; Gerlando et al., 2015).
CSN3*H differs from CSN3*B in the amino acid substitution from Asn53 to Ser53 due to the transition of A to G in exon 4 at CHR6:86208987 (Jann et al., 2004).
CSN3*I varies from the reference in the amino acid substitution Val65 to Ile65 as a result of a G to A transition at CHR6:86209022 in exon 4 (Jann et al., 2004).
CSN3*J varies from the reference CSN3*B in the substitution of the amino acid Tyr61 to Cys61 because of the transition of A to G in exon 4 at CHR6:86209011. (Jann et al., 2004).
CSN3*K distinguishes from the reference variant by the amino acid substitution Gln44 to Arg44 that is caused by an A to G transition at CHR6:86208960 in exon 4 (Jann et al., 2004). CSN3*K and CSN3*I were found at a frequency ≤0.10 in Bunte and Weise Deutsche Edelziege, and Buren goats from Germany (Kupper et al., 2010). In addition, CSN3*K was detected in Taggar and Saanen goats from Sudan (Rahmatalla et al., 2021).
CSN3*L contains the same amino acid substitution Gln44 to Arg44 that occurs in CSN3*K. The additional substitution of Ser159 by Pro159 differentiate CSN3*L from the reference. This substitution to Pro159 has been detected before in the other protein variants CSN3*C, CSN3*D, CSN3*F, and CSN3*G (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*L had been named before as CSN3*D (Kiplagat et al., 2010). This protein variant was found in Afar goats from Ethiopia only at a frequency of 0.05 (Kiplagat et al., 2010).
CSN3*M (provisionally named CSN3*Y) differs from the reference protein by three non-synonymous mutations. CSN3*M is characterized by a G to A transition at CHR6:86209097 resulting in the amino acid exchange of Asp90 to Asn90, a T to C transition at CHR6:86209263 resulting in a Val145 to Ala145 substitution, and a T to C transition at CHR6:86209304 resulting in the amino acid change from Ser159 to Pro159 (Chessa et al., 2003; Prinzenberg et al., 2005). CSN3*M occurs at a frequency of ≤0.05 in few Italian goats and Red Sokoto goats from Cameroon and Nigeria (Supplementary Table S6) (Chessa et al., 2003; Prinzenberg et al., 2005; Sacchi et al., 2005; Caroli et al., 2007; Gigli et al., 2008; Vacca et al., 2014). CSN3*M′ (previously named CSN3*N) differs from the variant CSN3*M by a synonymous mutation in codon 156 (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*M′ occurs at a frequency of 0.01 in Short Eared goats from Somalia (Kiplagat et al., 2010).
CSN3*N, named before as CSN3*M, carries the two amino acid substitutions Asp90 to Asn90 and Val145 to Ala145 (Kiplagat et al., 2010; Gautam et al., 2019).
The CSN3*O protein variant (previously CSN3*Q) carries the substitutions of Val65 to Ile65, Ile119 to Val119 and Ser159 to Pro159 compared to the reference (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*O and CSN3*N found in a few goats from Africa at a frequency of less than 0.05 and 0.15, respectively (Kiplagat et al., 2010; Rahmatalla et al., 2021).
The replacements of Gln44 to Arg44 and Val65 to Ile65 lead to the variant CSN3*P (Gautam et al., 2019).
CSN3*Q carries the substitutions Gln44 to Arg44, Ile119 to Val119 and Ser159 to Pro159, that had been detected and described before in other k-casein variants. Furthermore, CSN3*Q has the amino acid substitution Thr82 to Ala82 which is the result of an A to G transition at CHR6:86209073 in exon 4 (Gautam et al., 2019). CSN3*Q and CSN3*P were found at a frequency of 0.01 in Blank Bengal Indian goats (Gautam et al., 2019).
CSN3*R distinguishes from the reference in the amino acids Pro80 to Leu80 and Ile119 to Val119. The exchanges from Pro80 to Leu80 and Ile119 to Val119 result from a transition of C to T and A to G in exon 4 at CHR6:86,209,068 and CHR6:86209184, respectively (Gautam et al., 2019).
CSN3*S is distinguished from the reference by the amino acid substitutions Val67 to Ala67 and Ile119 to Val119. The substitution of Val67 to Ala67 is a result of a T to C transition at CHR6:86209029 in exon 4 (Gautam et al., 2019). The frequency of CSN3*S and CSN3*R is about 0.01 in Siroh goats from India (Gautam et al., 2019).
CSN3*T carries also the Ile119 to Val119 substitution. In addition, the substitution Ser37 to Arg37 is present which results from an A to C transversion at CHR6:86208938 in exon 4 (Gautam et al., 2019).
CSN3*U varies from the reference by two amino acid substitutions, Val78 to Ala78 and Asn143 to His143. The substitution of Val78 to Ala78 results from the mutation TT to CA at CHR6:86209062 and CHR6:86209063; the substitution of Asn143 to His143 occurs due to an A to C transversion at CHR6:86209256 in exon 4 (Gautam et al., 2019).
CSN3*V variant carries the Asn53 to Ser53 substitution that occurs also in variant CSN3*H, but variant CSN3*V contains also the amino acid change from Ser159 to Pro159 that can be found also in other k-casein variants (Gautam et al., 2019).
In CSN3*W, the amino acid substitution Asn53 to Ser53 is found as in CSN3*H and CSN3*V. In addition, the amino acid substitution Thr138 to Ala138 as a result of an A to G transition at CHR6:86209241 occurs in CSN3*W (Gautam et al., 2019). The protein variants CSN3*H, CSN3*T, CSN3*U, CSN3*V, and CSN3*W are rare. They were found at a frequency of 0.01 in Rajgarh Indian breed (Gautam et al., 2019).
CSN3*X differs from the reference by the amino acid substitutions Ser33 to Asn33 and Ser37 to Thr37 which are the result of G to A transitions at CHR6:86208927 and CHR6:86208939, respectively. CSN3*X was identified in Alpine ibex (Rahmatalla et al., 2021).
No information is available for the allele frequency of CSN3*A′, CSN3*B″, CSN3*J, and CSN3*X.
The CSN3 protein variants are classified into two groups based on the isoelectric focusing point (IEP). CSN3*A, CSN3*B, CSN3*B′, CSN3*B″, CSN3*C, CSN3*C′, CSN3*F, CSN3*G, CSN3*H, CSN3*I, CSN3*J, CSN3*L have an isoelectric focusing point of 5.29 and belong to group AIEF. CSN3*D, CSN3*E, CSN3*K, CSN3*M have an isoelectric focusing point of 5.66 and belong to group BIEF (Prinzenberg et al., 2005). The IEF pattern of CSN3*D′ and CSN3*N were not experimentally tested, but estimated using the ExPASy tool (Gasteiger et al., 2003). The D′ variant was classified to group BIEF and the N variant (new name G”) was classified to group AIEF (Gerlando et al., 2015). The k-casein variants in group BIEF were significantly associated with higher milk casein and protein content, which was shown in different goat breeds (Chiatti et al., 2005; Chiatti et al., 2007; Caravaca et al., 2009).
Due to the strong genetic linkage of the four casein genes within a cluster of 250 kb, the haplotype approach has been a suitable tool instead of evaluating a single gene for accessing the variability within and among populations in diversity studies (Sacchi et al., 2005; Finocchiaro et al., 2008; Gigli et al., 2008; Kupper et al., 2010; Criscione et al., 2019), for breeding strategies (Haplotype Assisted Selection) (Caroli et al., 2006; Hayes et al., 2006), and for prediction of individual milk composition and suitability for milk processing. However, the full definition of the casein haplotypes would require genomic sequence data and is also complicated because of the large number of genetic variations found at the four casein loci. One of the major problems in comparing the different research when considering haplotypes is the different mutations used for deriving casein haplotypes. Furthermore, the number of genotyped animals per population or breeds, and the availability of pedigree and parent information affect the reconstructing and thereby the accuracy of the exact haplotypes carried by each animal.
The evaluation of the haplotype variability in five Italian goat breeds from Northern and Southern Italy identified 18 common haplotypes (Sacchi et al., 2005). However, the study failed to genotype CSN2*C which, therefore, could not be considered it in the haplotype construction. The haplotype CSN1S1*F-CSN1S2*F-CSN3*D occurred in all breeds and was the most common haplotype in the Southern Italian breeds. In the Northern Italian breed, the haplotypes CSN1S1*E-CSN1S2*C-CSN3*D and CSN1S1*F-CSN1S2*C-CSN3*D were most prevalent. A high frequency of the CSN1S1*0-CSN1S2*C-CSN3*A haplotype, containing the null variant CSN1S1*0, was detected in Vallesana Italian goats. Interestingly, the haplotype CSN1S1*F-CSN2*0, which was found in Maltese goats, was associated with a very low casein level of around 2.5 g/L, even if it was linked to a strong variant of CSN1S2. Eventually, the “Maltese” haplotype is the result of selection for milk with specific nutritional properties rather than for cheese-making (Sacchi et al., 2005).
In a study of Caroli et al. (2006) in four goat breeds from Northern Italy, 13 haplotypes were identified that occurred at a frequency >0.05 in at least one breed. Based on the evolution model, CSN1S1*B-CSN2*A-CSN1S2*A-CSN3*B was suggested to be the ancestral haplotype (Caroli et al., 2006).
Fifty four haplotypes were detected by using 30 SNPs within the casein genes in the analysis of six Southern Italian breeds and Norwegian dairy goat which highlighted the significant diversity among the breed (Criscione et al., 2019).
The haplotype structure within each casein gene was constructed in Norwegian goats based on 39 SNPs (Hayes et al., 2006). The unique number of haplotypes were 10, 6, 4, and 8 for CSN1S1, CSN2, CSN1S2, and CSN3, respectively. Since CSN1S1 haplotypes significantly affected the protein percentage and fat yield, and CSN3 haplotypes the protein and fat percentage, the authors suggest the use of a haplotype-assisted selection approach in Norwegian dairy goats, which are used specifically for cheese production (Hayes et al., 2006).
The haplotype structure was based on 22 SNPs and one deletion within the four casein genes in two geographically distant populations, the Sicilian Girgentana breed that likely originated from Afghanistan and the Himalaya area and the Norwegian goat breed (Finocchiaro et al., 2008). The number of haplotypes were 10, 8, 3, and 11 for CSN1S1, CSN2, CSN1S2, and CSN3, respectively. Although considerably geographically distant and phenotypically divergent, casein haplotypes were identical between both breeds.
In five Italian Sicilian goat breeds, 27 haplotypes were observed at a frequency >0.03. The most frequent haplotype in Derivata di Siria and Maltese was CSN1S1*F-CSN2*C-CSN1S2*F-CSN3*B. This haplotype is associated with low milk casein content, and therefore superior for the production of hypoallergenic goat milk (Gigli et al., 2008). In Argentata dell’Etna, Girgentana and Messinese the most frequent haplotype was CSN1S1*A-CSN2*C-CSN1S2*A-CSN3*B which is also most favorable for cheese production (Gigli et al., 2008).
In Bunte Deutsche Edelziege and Weiße Deutsche Edelziege dairy goats from Germany, 30 haplotypes with a frequency higher than 0.01 were detected, only four haplotypes were found in Buren meat goats (Kupper et al., 2010). The ancestral haplotype CSN1S1*B-CSN2*A-CSN1S2*A-CSN3*B was evident in all three breeds with the highest frequency in Buren goats (Kupper et al., 2010). The CSN1S1*E-CSN2*A-CSN1S2*A-CSN3*B haplotype was most frequent in both dairy breeds followed by CSN1S1*F-CSN2*C-CSN1S2*F-CSN3*B in Bunte Deutsche Edelziege and CSN1S1*F-CSN2*C1-CSN1S2*F-CSN3*B in Weiße Deutsche Edelziege, which might be attributed to the selection of milk quantity. The deep divergence in the casein gene haplotypes in dairy goat breeds was attributed to different selection aims among breeds.
The selection for favorable alleles of major genes within a breed is cost-effective if it can be implemented in an existing selection system. Major genes affecting milk protein content, particularly caseins, are well known. The prevalence of specific casein variants in different goat breeds is probably the result of breeding goals but could be also due to random fixation. Currently, there are several kits on the market for testing milk protein variants in goats (Simões and Gutiérrez, 2018). Genotyping CSN1S1 is of particular interest for breeders and the milk industry since genetic variation in the CSN1S1 gene is one of the primary factors affecting milk protein content and technological properties, such as cheese yield and cheese curd formation. Selection schemes in Alpine and Saanen goats currently use CSN1S1 variants for the pre-selection of young males undergoing the progeny test (Manfredi et al., 2000).
It is also conceivable to select goats carrying casein gene variants with anti-allergenic properties to produce healthy milk as functional food for human nutrition (Robinson, 2001).
Goat milk proteins possess unique biologically active components that affect the milk properties and have an impact on numerous aspects of human nutrition. Goat milk contains less lactose and higher alkalinity compared to cow milk (Robinson, 2011). Goat milk is highly digestible and has been approved repeatedly as an alternative milk source for people, who suffer from lactose intolerance and cow milk protein allergies (Park and Haenlein, 2017). Therefore, goat milk has a therapeutic value in human medicine and nutrition (Park, 2009).
Many people who are allergic and have gastrointestinal disorders to cow milk can tolerate goat milk because of considerable variation in goat milk protein sequences (Park and Haenlein, 2017). Numerous cow milk proteins have been identified as allergens (Wal, 2002). Alpha-S1-casein, α-S2-casein, and beta-lactoglobulin are among the most crucial milk protein allergens (Wal, 1998; Crittenden and Bennett, 2005). Beta-lactoglobulin does not occur in human milk (O'Mahony et al., 2014). For example, α-S1-casein from cow milk forms hard curds in the stomach which can cause digestion problems. Therefore, the advantage of goat milk is the low concentration or even null content of α-S1-casein and other allergic casein protein variants (Bernacka, 2011; Yangilar, 2013).
For instance, the association of the casein protein variants with the null or low content of those proteins might affect the production of goat milk with hypoallergenic characteristics and selecting milk with null or reduced content of the particular protein. In this concern, some variants in goat α-S1-casein, β-casein, and α-S2-casein in different goat breeds are associated with null or reduced expression of the encoded protein (Persuy et al., 1999; Rando et al., 2000; Ramunno et al., 2001a; Ramunno et al., 2001b; Ramunno et al., 2005; Caroli et al., 2006).
Through the digestion of goat milk proteins and milk products, several interesting peptides have been identified with bioactive properties. The beneficial health effects of these peptides include antihypertensive, anti-diabetic, antithrombotic, immunomodulatory, antibacterial, antifungal, antiviral, antioxidant, binding and transporting metals, preventing amnesia, and causing smooth muscle contractions properties (Ebringer et al., 2008; Atanasova and Ivanova, 2010; Zhang et al., 2015; Gong et al., 2020).
In this review, we provide a comprehensive overview on the genetic variation of the four casein proteins in different goat breeds. We provide a translation of genetic DNA variants to the corresponding amino acid changes, as well as the protein variants. In addition, we also updated the names of different protein variants according to the information we found in a literature search on reported protein and genetic variants in casein to summarize more than 30 years of research. The effect of different casein gene variants on the casein content, milk composition, and cheese-making properties is discussed. The wide variation occurring in the milk caseins has impact on the nutritional value for humans and on technological properties in milk processing. The different characteristics of goat milk likely affected breeding goals and adaptation, which led to high frequency of some casein variants in various goat breeds and country. The casein genomic and proteomic variations as well as the casein haplotypes approach might be useful in selection of the animals according to the specific breeding goals.
SAR, DA, and GAB substantially contributed to the conception and design of the article and interpreting the relevant literature. SAR drafted the manuscript. DA and GAB critical revision of the manuscript. All authors read and approved the final manuscript.
This study was supported by a German Research Foundation (DFG), Germany, to SAR (funding number RA 3482/2-1). We acknowledge the support of the article processing charge by the Deutsche Forschungsgemeinschaft DFG, German Research Foundation)—491192747 and the Open Access Publication Fund of Humboldt-Universität zu Berlin.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.995349/full#supplementary-material
Supplementary Table S1 | Nucleotide and amino acid variants within the gene and mature protein of the CSN1S1.
Supplementary Table S2 | Frequency of CSN1S1 variants in different goat breeds and countries.
Supplementary Table S3 | Frequency of CSN2 variants in different goat breeds and countries.
Supplementary Table S4 | Frequency of CSN1S2 variants in different goat breeds and countries.
Supplementary Table S5 | Nucleotide and amino acid variants within the gene and mature protein of the CSN3.
Supplementary Table S6 | Frequency of CSN3 variants in different goat breeds and countries.
Albenzio, M., Santillo, A., D'angelo, F., and Sevi, A. (2009). Focusing on casein gene cluster and protein profile in Garganica goat milk. J. Dairy Res. 76, 83–89. doi:10.1017/S0022029908003853
Alexander, L. J., Stewart, A. F., Mackinlay, A. G., Kapelinskaya, T. V., Tkach, T. M., and Gorodetsky, S. I. (1988). Isolation and characterization of the bovine kappa-casein gene. Eur. J. Biochem. 178, 395–401. doi:10.1111/j.1432-1033.1988.tb14463.x
Ambrosoli, R., Di Stasio, L., and Mazzocco, P. (1988). Content of alpha S1-casein and coagulation properties in goat milk. J. Dairy Sci. 71, 24–28. doi:10.3168/jds.S0022-0302(88)79520-X
Angiolillo, A., Yahyaoui, M. H., Sanchez, A., Pilla, F., Folch, J. M., and Sanchez, A. (2002). Short communication: Characterization of a new genetic variant in the caprine kappa-casein gene. J. Dairy Sci. 85, 2679–2680. doi:10.3168/jds.s0022-0302(02)74353-1
Aschaffenburg, R., and Drewry, J. (1955). Occurrence of different beta-lactoglobulins in cow's milk. Nature 176, 218–219. doi:10.1038/176218b0
Assan, N. (2014). Factors affecting yield and milk composition in dairy animals. Sunnyvale, CA, USA: Lap Lambert Academic Publishing GmbH KG.
Atanasova, J., and Ivanova, I. (2010). Antibacterial peptides from goat and sheep milk proteins. Biotechnol. Biotechnol. Equip. 24, 1799–1803. doi:10.2478/V10133-010-0049-8
Baltrėnaitė, L., Kerzienė, S., Morkūnienė, K., and Miceikienė, I. (2009). Genetic diversity in milk proteins among goats bred in Lithuania. Acta Univ. Latv. 753, 117–124.
Barth, C. A., and Schlimme, E. (2012). Milk proteins: Nutritional, clinical, functional and technological aspects. Heidelberg, Germany: Steinkopff Heidelberg.
Bevilacqua, C., Ferranti, P., Garro, G., Veltri, C., Lagonigro, R., Leroux, C., et al. (2002). Interallelic recombination is probably responsible for the occurrence of a new alpha(s1)-casein variant found in the goat species. Eur. J. Biochem. 269, 1293–1303. doi:10.1046/j.1432-1033.2002.02777.x
Bhat, M. Y., Dar, T. A., and Laishram Rajendrakumar Singh, S. K. (2016). “Casein proteins: Structural and functional aspects,” in Milk proteins. Editor I. Gigli (London, UK: IntechOpen), 296.
Bickhart, D. M., Rosen, B. D., Koren, S., Sayre, B. L., Hastie, A. R., Chan, S., et al. (2017). Single-molecule sequencing and chromatin conformation capture enable de novo reference assembly of the domestic goat genome. Nat. Genet. 49, 643–650. doi:10.1038/ng.3802
Birgisdottir, B. E., Hill, J. P., Harris, D. P., and Thorsdottir, I. (2002). Variation in consumption of cow milk proteins and lower incidence of Type 1 diabetes in Iceland vs the other 4 Nordic countries. Diabetes Nutr. Metab. 15, 240–245.
Boulanger, A., Grosclaude, F., and Mahé, M. F. (1984). Polymorphisme des caséines α(s1 )et α(s2 )de la chèvre (Capra hircus). Genet. Sel. Evol. 16, 157–176. doi:10.1186/1297-9686-16-2-157
Bouniol, C., Brignon, G., Mahe, M. F., and Printz, C. (1994). Biochemical and genetic analysis of variant C of caprine alpha s2-casein (Capra hircus). Anim. Genet. 25, 173–177. doi:10.1111/j.1365-2052.1994.tb00106.x
Bouniol, C., Brignon, G., Mahe, M. F., and Printz, C. (1993). Characterization of goat αs2 casein A and B: Further evidence of the phosphorylation code of caseins. Protein Sequence Data Analysis 5, 213–218.
Bouniol, C. (1993). Sequence of the goat αs2-casein-encoding cDNA. Gene 125, 235–236. doi:10.1016/0378-1119(93)90336-2
Brignon, G., Mahe, M. F., Grosclaude, F., and Ribadeau-Dumas, B. (1989). Sequence of caprine alpha s1-casein and characterization of those of its genetic variants which are synthesized at a high level, alpha s1-CnA, B and C. Protein Seq. Data Anal. 2, 181–188.
Brignon, G., Mahe, M. F., Ribadeau-Dumas, B., Mercier, J. C., and Grosclaude, F. (1990). Two of the three genetic variants of goat alpha s1-casein which are synthesized at a reduced level have an internal deletion possibly due to altered RNA splicing. Eur. J. Biochem. 193, 237–241. doi:10.1111/j.1432-1033.1990.tb19328.x
Caravaca, F., Amills, M., Jordana, J., Angiolillo, A., Agüera, P., Aranda, C., et al. (2008). Effect of alphas1-casein ( CSN1S1) genotype on milk CSN1S1 content in Malagueña and Murciano-Granadina goats. J. Dairy Res. 75, 481–484. doi:10.1017/S0022029908003609
Caravaca, F., Ares, J. L., Carrizosa, J., Urrutia, B., Baena, F., Jordana, J., et al. (2011). Effects of alpha s1-casein (CSN1S1) and kappa-casein (CSN3) genotypes on milk coagulation properties in Murciano-Granadina goats. J. Dairy Res. 78, 32–37. doi:10.1017/S002202991000083X
Caravaca, F., Carrizosa, J., Urrutia, B., Baena, F., Jordana, J., Amills, M., et al. (2009). Short communication: Effect of alphaS1-casein (CSN1S1) and kappa-casein (CSN3) genotypes on milk composition in Murciano-Granadina goats. J. Dairy Sci. 92, 2960–2964. doi:10.3168/jds.2008-1510
Caroli, A., Chiatti, F., Chessa, S., Rignanese, D., Bolla, P., and Pagnacco, G. (2006). Focusing on the goat casein complex. J. Dairy Sci. 89, 3178–3187. doi:10.3168/jds.S0022-0302(06)72592-9
Caroli, A., Chiatti, F., Chessa, S., Rignanese, D., Ibeagha-Awemu, E. M., and Erhardt, G. (2007). Characterization of the casein gene complex in West African goats and description of a new alpha(s1)-casein polymorphism. J. Dairy Sci. 90, 2989–2996. doi:10.3168/jds.2006-674
Caroli, A., Jann, O., Budelli, E., Bolla, P., Jager, S., and Erhardt, G. (2001). Genetic polymorphism of goat kappa-casein (CSN3) in different breeds and characterization at DNA level. Anim. Genet. 32, 226–230. doi:10.1046/j.1365-2052.2001.00765.x
Caroli, A. M., Chessa, S., and Erhardt, G. J. (2009). Invited review: Milk protein polymorphisms in cattle: Effect on animal breeding and human nutrition. J. Dairy Sci. 92, 5335–5352. doi:10.3168/jds.2009-2461
Catota-Gómez, L. D., Parra-Bracamonte, G. M., Cienfuegos-Rivas, E. G., Hernández-Meléndez, J., Sifuentes-Rincón, A. M., and Martínez-González, J. C. (2017). Frequency and association of polymorphisms in CSN3 gene with milk yield and composition in Saanen goats. Ecosist. Recur. Agropec. 4, 411–417. doi:10.19136/era.a4n12.1165
Cebo, C., Lopez, C., Henry, C., Beauvallet, C., Ménard, O., Bevilacqua, C., et al. (2012). Goat αs1-casein genotype affects milk fat globule physicochemical properties and the composition of the milk fat globule membrane. J. Dairy Sci. 95, 6215–6229. doi:10.3168/jds.2011-5233
Chanat, E., Martin, P., and Ollivier-Bousquet, M. (1999). Alpha(S1)-casein is required for the efficient transport of beta- and kappa-casein from the endoplasmic reticulum to the Golgi apparatus of mammary epithelial cells. J. Cell. Sci. 112, 3399–3412. doi:10.1242/jcs.112.19.3399
Chessa, S., and Bolla, P. (2003). Il polimorfismo della caseine caprine: Uno straordinario “puzzle” genetico dalle svariate potenzialità applicative. Sci. Tecn. Latt.-Cas. 54, 343–358.
Chessa, S., Budelli, E., Chiatti, F., Cito, A. M., Bolla, P., and Caroli, A. (2005). Short communication: Predominance of beta-casein (CSN2) C allele in goat breeds reared in Italy. J. Dairy Sci. 88, 1878–1881. doi:10.3168/jds.S0022-0302(05)72863-0
Chessa, S., Budelli, E., Gutscher, K., Caroli, A., and Erhardt, G. (2003). Short communication: Simultaneous identification of five kappa-casein (CSN3) alleles in domestic goat by polymerase chain reaction-single strand conformation polymorphism. J. Dairy Sci. 86, 3726–3729. doi:10.3168/jds.S0022-0302(03)73978-2
Chessa, S., Chiatti, F., Rignanese, D., Ibeagha-Awemu, E. M., Özbeyaz, C., Hassan, Y. A., et al. (2007). The casein genes in goat breeds from different continents: Analysis by polymerase chain reaction – single strand conformation polymorphism (PCR-SSCP). Italian J. Animal Sci. 6, 73–75. doi:10.4081/ijas.2007.1s.73
Chessa, S., Rignanese, D., Kupper, J., Pagnacco, G., Erhardt, G., and Caroli, A. (2008). Short communication: The beta-casein (CSN2) silent allele C1 is highly spread in goat breeds. J. Dairy Sci. 91, 4433–4436. doi:10.3168/jds.2008-1228
Chianese, L., Caira, S., Garro, G., Quarto, M., Mauriello, R., and Addeo, F. (2007). “Occurrence of genetic polymorphism at goat β-CN locus,” in 5th Int. Symp, Challenge to Sheep and Goats Milk Sectors, Sardinia, Italy.
Chianese, L., Ferranti, P., Garro, G., Mauriello, R., and Addeo, F. (1997). “Occurrence of three novel alpha s1-casein variants in goat milk,” in International Dairy Federation, Brussels, Belgium.
Chianese, L., Garro, G., Ferranti, P., Speranza, L. M., Pilla, F., Bevilacqua, C., et al. (1998). “Occurence of new variant ofgoat αs1-and αs2-casein in abreed of Southern Italy,” in Proceeding of 25th International Dairy Congress, 82nd annual sessions, Arahus, Denmark.
Chiatti, F., Caroli, A., Chessa, S., Bolla, P., and Pagnacco, G. (2005). “Relationships between goat κ-casein (CSN3) polymorphism and milk composition,” in The role of biotechnology. Villa Gualino, Turin, Italy: AGRIS.
Chiatti, F., Chessa, S., Bolla, P., Cigalino, G., Caroli, A., and Pagnacco, G. (2007). Effect of kappa-casein polymorphism on milk composition in the Orobica goat. J. Dairy Sci. 90, 1962–1966. doi:10.3168/jds.2006-508
Cieslinska, A., Kostyra, E., Kostyra, H., Olenski, K., Fiedorowicz, E., and Kaminski, S. (2012). Milk from cows of different beta-casein genotypes as a source of beta-casomorphin-7. Int. J. Food Sci. Nutr. 63, 426–430. doi:10.3109/09637486.2011.634785
Clark, S., and Mora Garcia, M. B. (2017). A 100-Year Review: Advances in goat milk research. J. Dairy Sci. 100, 10026–10044. doi:10.3168/jds.2017-13287
Clark, S., and Sherbon, J. W. (2000). Alpha s1-casein, milk composition and coagulation properties of goat milk. Small Ruminant Res. 38, 123–134. doi:10.1016/S0921-4488(00)00154-1
Coll, A., Folch, J. M., Sanchez, A., and Sanchez, A. (1993). Nucleotide sequence of the goat kappa-casein cDNA. J. Anim. Sci. 71, 2833. doi:10.2527/1993.71102833x
Cosenza, G., Gallo, D., Illario, R., Di Gregorio, P., Senese, C., Ferrara, L., et al. (2003). A Mval PCR-RFLP detecting a silent allele at the goat alpha-lactalbumin locus. J. Dairy Res. 70, 355–357. doi:10.1017/s0022029903006265
Cosenza, G., Iannaccone, M., Pico, B. A., Ramunno, L., and Capparelli, R. (2016). The SNP g.1311T>C associated with the absence of β-casein in goat milk influences CSN2 promoter activity. Anim. Genet. 47, 615–617. doi:10.1111/age.12443
Cosenza, G., Pauciullo, A., Colimoro, L., Mancusi, A., Rando, A., Di Berardino, D., et al. (2007). An SNP in the goat CSN2 promoter region is associated with the absence of β-casein in milk. Anim. Genet. 38, 655–658. doi:10.1111/j.1365-2052.2007.01649.x
Cosenza, G., Pauciullo, A., Gallo, D., Berardino, D. D., and Ramunno, L. (2005). A Ssp I PCR-RFLP detecting a silent allele at the goat CSN2 locus. J. Dairy Res. 72, 456–459. doi:10.1017/S0022029905001342
Cosenza, G., Pauciullo, A., Gallo, D., Colimoro, L., D’avino, A., Mancusi, A., et al. (2008). Genotyping at the CSN1S1 locus by PCR-RFLP and AS-PCR in a Neapolitan goat population. Small Ruminant Res. 74, 84–90. doi:10.1016/j.smallrumres.2007.03.010
Criscione, A., Tumino, S., Avondo, M., Marletta, D., and Bordonaro, S. (2019). Casein haplotype diversity in seven dairy goat breeds. Arch. Anim. Breed. 62, 447–454. doi:10.5194/aab-62-447-2019
Crittenden, R. G., and Bennett, L. E. (2005). Cow's milk allergy: A complex disorder. J. Am. Coll. Nutr. 24, 582S–591S. doi:10.1080/07315724.2005.10719507
Cunsolo, V., Muccilli, V., Saletti, R., Marletta, D., and Foti, S. (2006). Detection and characterization by high-performance liquid chromatography and mass spectrometry of two truncated goat alphas2-caseins. Rapid Commun. Mass Spectrom. 20, 1061–1070. doi:10.1002/rcm.2415
Dall’olio, S., Davoli, R., and Russ, O. V. (1989). Una nuova variante di caseina caprina. Sci. Tec. Latt. Casearia 40, 24–28.
De Noni, I., and Cattaneo, S. (2010). Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 119, 560–566. doi:10.1016/j.foodchem.2009.06.058
Delacroix Buchet, A., Josas, J. E., Laitieres, R., Degas, C., Lamberet, G., and Vassal, L. (1996). Effect of AA and FF caprine αs1-casein variants on cheesemaking. Lait 76, 217–241. doi:10.1051/lait:1996319
Devold, T. G., Nordb, R., Langsrud, T., Svenning, C., Brovold, M. J., Sørensen, E. S., et al. (2011). Extreme frequencies of the αs1-casein “null” variant in milk from Norwegian dairy goats— Implications for milk composition, micellar size and renneting properties. Dairy Sci. Technol. 91, 39–51. doi:10.1051/dst/2010033
Ebringer, L., Ferencik, M., and Krajcovic, J. (2008). Beneficial health effects of milk and fermented dairy products--review. Folia Microbiol. (Praha). 53, 378–394. doi:10.1007/s12223-008-0059-1
El-Shazly, S. A., Ahmed, M. M., and Amer, S. M. (2017). Genetic polymorphism in some milk protein genes and its impact on milk composition of Saudi Arabian goat breeds reared in Taif region. Romanian Biotechnol. Lett. 22.
Erhardt, G., Jäger, S., Budelli, E., and Caroli, A. (2002). Genetic polymorphism of goat αS2-casein (CSN1S2) and evidence for a further allele. Milchwissenschaft 57, 137–140.
Farrell, H. M., Jimenez-Flores, R., Bleck, G. T., Brown, E. M., Butler, J. E., Creamer, L. K., et al. (2004). Nomenclature of the proteins of cows’ milk—sixth revision. J. Dairy Sci. 87, 1641–1674. doi:10.3168/jds.S0022-0302(04)73319-6
Ferretti, L., Leone, P., and Sgaramella, V. (1990). Long range restriction analysis of the bovine casein genes. Nucleic Acids Res. 18, 6829–6833. doi:10.1093/nar/18.23.6829
Finocchiaro, R., Hayes, B. J., Siwek, M., Spelman, R. J., Van Kaam, J. B., Adnoy, T., et al. (2008). Comparison of casein haplotypes between two geographically distant European dairy goat breeds. J. Anim. Breed. Genet. 125, 68–72. doi:10.1111/j.1439-0388.2007.00687.x
Fox, P. F., and Brodkorb, A. (2008). The casein micelle: Historical aspects, current concepts and significance. Int. Dairy J. 18, 677–684. doi:10.1016/j.idairyj.2008.03.002
Fox, P. F., and Mcsweeney, P. L. H. (1998). Dairy chemistry and biochemistry. Berlin, Germany: Springer.
Galliano, F., Saletti, R., Cunsolo, V., Foti, S., Marletta, D., Bordonaro, S., et al. (2004). Identification and characterization of a new beta-casein variant in goat milk by high-performance liquid chromatography with electrospray ionization mass spectrometry and matrix-assisted laser desorption/ionization mass spectrometry. Rapid Commun. Mass Spectrom. 18, 1972–1982. doi:10.1002/rcm.1575
Gasteiger, E., Gattiker, A., Hoogland, C., Ivanyi, I., Appel, R. D., and Bairoch, A. (2003). ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 31, 3784–3788. doi:10.1093/nar/gkg563
Gautam, D., Vats, A., Verma, M., Rout, P. K., Meena, A. S., Ali, M., et al. (2019). Genetic variation in CSN3 exon 4 region of Indian goats and a new nomenclature of CSN3 variants. Anim. Genet. 50, 191–192. doi:10.1111/age.12767
Gerlando, R. D., Tortorici, L., Sardina, M. T., Monteleone, G., Mastrangelo, S., and Portolano, B. (2015). Molecular characterisation of κ–casein gene in Girgentana dairy goat breed and identification oftwo new alleles. Italian J. Animal Sci. 14, 3464. doi:10.4081/ijas.2015.3464
Ghosh, D., Das, S., Bagchi, D., and Smarta, R. B. (2016). Innovation in healthy and functional foods. Boca Raton, FL, USA: CRC Press.
Giambra, I. J., and Erhardt, G. E. (2012). Milk proteins, milk protein variants and the importance for the sheep breeding - a review. Züchtungskunde. 84, 52–73.
Gigli, I., Maizon, D. O., Riggio, V., Sardina, M. T., and Portolano, B. (2008). Short communication: Casein haplotype variability in Sicilian dairy goat breeds. J. Dairy Sci. 91, 3687–3692. doi:10.3168/jds.2008-1067
Gigli, I. (2016). Milk proteins: From structure to biological properties and health aspects. IntechOpen.
Glantz, M., Devold, T. G., Vegarud, G. E., Lindmark Mansson, H., Stalhammar, H., and Paulsson, M. (2010). Importance of casein micelle size and milk composition for milk gelation. J. Dairy Sci. 93, 1444–1451. doi:10.3168/jds.2009-2856
Gong, H., Gao, J., Wang, Y., Luo, Q. W., Guo, K. R., Ren, F. Z., et al. (2020). Identification of novel peptides from goat milk casein that ameliorate high-glucose-induced insulin resistance in HepG2 cells. J. Dairy Sci. 103, 4907–4918. doi:10.3168/jds.2019-17513
Grosclaude, F., Mahe, M. F., Brignon, G., Di Stasio, L., and Jeunet, R. (1987). A Mendelian polymorphism underlying quantitative variations of goat α(s1)-casein. Genet. Sel. Evol. 19, 399–412. doi:10.1186/1297-9686-19-4-399
Grosclaude, F., and Martin, P. (1997). “Casein polymorphisms in the goat,” in International dairy federation (Palmerston North, New Zealand: IDF Seminar `Milk Protein Polymorphism II).
Grosclaude, F., Ricordeau, G., Martin, P., Remeuf, F., Vassal, L., and Bouillon, J. (1994). Du gène au fromage : Le polymorphisme de la caséine alphas1 caprine, ses effets, son évolution. INRA. Prod. Anim. 7, 3–19. doi:10.20870/productions-animales.1994.7.1.4153
Guinee, T. P., and O'brien, B. (2010). “The quality of milk for cheese manufacture,” in Technology of cheesemaking. Editors B. A. Law, and A. Y. Tamime, 1–67.
Guo, M. (2009). “Chapter 9 - human milk and infant formula,” in Functional foods. Editor M. Guo (Sawston, UK: Woodhead Publishing), 299–337.
Haratifar, S., and Guri, A. (2017). “5-Nanocapsule formation by caseins,” in Nanoencapsulation technologies for the food and nutraceutical industries. Editor S. Jafari (Oxford: Academic Press), 140–164.
Hayes, B., Hagesaether, N., Adnoy, T., Pellerud, G., Berg, P. R., and Lien, S. (2006). Effects on production traits of haplotypes among casein genes in Norwegian goats and evidence for a site of preferential recombination. Genetics 174, 455–464. doi:10.1534/genetics.106.058966
Hayes, H. C., and Petit, E. J. (1993). Mapping of the beta-lactoglobulin gene and of an immunoglobulin M heavy chain-like sequence to homoeologous cattle, sheep, and goat chromosomes. Mamm. Genome 4, 207–210. doi:10.1007/BF00417564
Ho, S., Woodford, K., Kukuljan, S., and Pal, S. (2014). Comparative effects of A1 versus A2 beta-casein on gastrointestinal measures: A blinded randomised cross-over pilot study. Eur. J. Clin. Nutr. 68, 994–1000. doi:10.1038/ejcn.2014.127
Hui, Y. H., and Sherkat, F. (2005). Handbook of food science, technology, and engineering - 4 Volume Set. Boca Raton, FL, USA: CRC Press.
Ingham, B., Smialowska, A., Kirby, N. M., Wang, C., and Carr, A. J. (2018). A structural comparison of casein micelles in cow, goat and sheep milk using X-ray scattering. Soft Matter 14, 3336–3343. doi:10.1039/c8sm00458g
Jann, O. C., Prinzenberg, E. M., Luikart, G., Caroli, A., and Erhardt, G. (2004). High polymorphism in the kappa-casein (CSN3) gene from wild and domestic caprine species revealed by DNA sequencing. J. Dairy Res. 71, 188–195. doi:10.1017/s0022029904000093
Jansàpérez, M., Leroux, C., Bonastre, A. S., and Martin, P. (1994). Occurrence of a LINE sequence in the 3' UTR of the goat alpha s1-casein E-encoding allele associated with reduced protein synthesis level. Gene 147, 179–187. doi:10.1016/0378-1119(94)90063-9
Jianqin, S., Leiming, X., Lu, X., Yelland, G. W., Ni, J., and Clarke, A. J. (2016). Effects of milk containing only A2 beta casein versus milk containing both A1 and A2 beta casein proteins on gastrointestinal physiology, symptoms of discomfort, and cognitive behavior of people with self-reported intolerance to traditional cows' milk. Nutr. J. 15, 35. doi:10.1186/s12937-016-0147-z
Johansson, I., and Lif Holgerson, P. (2011). Milk and oral health. Nestle Nutr. Workshop Ser. Pediatr. Program. 67, 55–66. doi:10.1159/000325575
Johansson, M., Högberg, M., and Andrén, A. (2015). Relation between αs1-casein content and coagulation properties of milk from Swedish dairy goats. Open Food Sci. J. 9, 1–4. doi:10.2174/1874256401509010001
Jordana, J., Amills, M., Diaz, E., Angulo, C., Serradilla, J. M., and Sanchez, A. (1996). Gene frequencies of caprine αs1-casein polymorphism in Spanish goat breeds. Small Ruminant Res. 20, 215–221. doi:10.1016/0921-4488(95)00813-6
Kiplagat, S., Agaba, M., Kosgey, I., Okeyo, M., Indetie, D., Hanotte, O., et al. (2010). Genetic polymorphism of kappa-casein gene in indigenous Eastern Africa goat populations. Int. J. Genet. Mol. Biol. 2, 001–005.
Kolb, A. F., Huber, R. C., Lillico, S. G., Carlisle, A., Robinson, C. J., Neil, C., et al. (2011). Milk lacking alpha-casein leads to permanent reduction in body size in mice. PLoS One 6, e21775. doi:10.1371/journal.pone.0021775
Kumar, A., Rout, P. K., Mandal, A., and Roy, R. (2009). Kappa-casein gene polymorphism in Indian goats. ndian J. Biotechnol. 8, 214–221.
Kumar, A., Rout, P. K., Mandal, A., Roy, R., and MandAl, A. (2007). Identification of the CSN1S1 allele in Indian goats by the PCR-RFLP method. Animal 1, 1099–1104. doi:10.1017/S1751731107000444
Kupper, J., Chessa, S., Rignanese, D., Caroli, A., and Erhardt, G. (2010). Divergence at the casein haplotypes in dairy and meat goat breeds. J. Dairy Res. 77, 56–62. doi:10.1017/S0022029909990343
Kusza, S., Ilie, D. E., Sauer, M., Nagy, K., Patras, I., and Gavojdian, D. (2016). Genetic polymorphism of CSN2 gene in Banat white and carpatina goats. Acta Biochim. Pol. 63, 577–580. doi:10.18388/abp.2016_1266
Kusza, S., Veress, G., Kukovics, S., J´Avor, A. A., Sanchez, A., Angiolillo, A., et al. (2007). Genetic polymorphism of s1- and s2-caseins in Hungarian milking goats. Small Ruminant Res. 68, 329–332. doi:10.1016/j.smallrumres.2005.11.014
Lagonigro, R., Pietrola, E., D'andrea, M., Veltri, C., and Pilla, F. (2001). Molecular genetic characterization of the goat s2-casein E allele. Anim. Genet. 32, 391–393. doi:10.1046/j.1365-2052.2001.0781c.x
Lamothe, S., Robitaille, G., St-Gelais, D., and Britten, M. (2007). Short communication: Extraction of beta-casein from goat milk. J. Dairy Sci. 90, 5380–5382. doi:10.3168/jds.2007-0488
Lan, X., Chen, H., Zhang, R., Tian, Y., Zhang, Y., Fang, X., et al. (2005). Association of polymorphisms of CSN1S2 gene with average milk yield and body sizes indexes in Xinong Saanen dairy goat. Acta Veterinaria Zootechnica Sinica 36, 318–322.
Le Parc, A., Leonil, J., and Chanat, E. (2010). AlphaS1-casein, which is essential for efficient ER-to-Golgi casein transport, is also present in a tightly membrane-associated form. BMC Cell. Biol. 11, 65. doi:10.1186/1471-2121-11-65
Leroux, C., Martin, P., Mahe, M. F., Leveziel, H., and Mercier, J. C. (1990). Restriction fragment length polymorphism identification of goat alpha s1-casein alleles: A potential tool in selection of individuals carrying alleles associated with a high level protein synthesis. Anim. Genet. 21, 341–351. doi:10.1111/j.1365-2052.1990.tb01979.x
Leroux, C., Mazure, N., and Martin, P. (1992). Mutations away from splice site recognition sequences might cis-modulate alternative splicing of goat alpha s1-casein transcripts. Structural organization of the relevant gene. J. Biol. Chem. 267, 6147–6157. doi:10.1016/s0021-9258(18)42674-9
López-Expósito, I., Amigo, L., and Recio, I. (2008). Identification of the initial binding sites of alphas2-casein f(183-207) and effect on bacterial membranes and cell morphology. Biochim. Biophys. Acta 1778, 2444–2449. doi:10.1016/j.bbamem.2008.06.018
Maga, E. A., Daftari, P., Kultz, D., and Penedo, M. C. (2009). Prevalence of alphas1-casein genotypes in American dairy goats. J. Anim. Sci. 87, 3464–3469. doi:10.2527/jas.2009-1854
Mahé, M. F., and Grosclaude, F. (1993). Polymorphism of β-casein in the Creole goat of Guadeloupe: Evidence for a null allele. Genet. Sel. Evol. 25, 403. doi:10.1186/1297-9686-25-4-403
Mahé, M. F., and Grosclaude, F. (1989). αS1-CnD, another allele associated with a decreased synthesis rate at the caprine αS1-casein locus. Genet. Sel. Evol. 21, 127. doi:10.1186/1297-9686-21-2-127
Malkoski, M., Dashper, S. G., O'brien-Simpson, N. M., Talbo, G. H., Macris, M., Cross, K. J., et al. (2001). Kappacin, a novel antibacterial peptide from bovine milk. Antimicrob. Agents Chemother. 45, 2309–2315. doi:10.1128/AAC.45.8.2309-2315.2001
Manfredi, E., Serradilla, J. M., Leroux, C., Martin, P., and Sánchez, A. (2000). “Genetics for milk production,” in Proceedings of the 7th International Conference on Goats (Poitiers, France: INRA, Paris).
Marković, B., Marković, M., Trivunović, S., Mirecki, S., Antunović, Z., and Veljić, M. (2018). Effects of the alpha s1-casein genotype on milk yield and milk composition of Balkan goat in Montenegro. AgricultForest. 64, 5–14. doi:10.17707/agricultforest.64.3.01
Marletta, D., Bordonaro, S., Guastella, A. M., Criscione, A., and D’urso, G. (2005). Genetic polymorphism of the calcium sensitive caseins in Sicilian Girgentana and Argentata dell’Etna goat breeds. Small Ruminant Res. 57, 133–139. doi:10.1016/j.smallrumres.2004.06.019
Marletta, D., Bordonaro, S., Guastella, A. M., and D'urso, G. (2004). Genetic polymorphism at CSN1S2 locus in two endangered Sicilian goat breeds. J. Anim. Breed. Genet. 121, 52–56. doi:10.1046/j.0931-2668.2003.00413.x
Marletta, D., Criscione, A., Ordonaro, S. B., Guastella, A. M., and D’urso, G. (2007). Review: Casein polymorphism in goat’s milk. Lait 87, 491–504. doi:10.1051/lait:2007034
Martin, P., Bianchi, L., Cebo, C., Miranda, G., Mcsweeney, P., and Fox, P. (2013). “Genetic polymorphism of milk proteins,” in Advanced dairy chemistry (Boston: Springer).
Martin, P., and Leroux, C. (1994). “Characterization of a further αS1-casein variant generated by exon skipping,” in 24th International Society of Animal Genetics Conference, Prague.
Martin, P., Ollivier-Bousquet, M., and Grosclaude, F. (1999). Genetic polymorphism of caseins: A tool to investigate casein micelle organization. Int. Dairy J. 9, 163–171. doi:10.1016/S0958-6946(99)00055-2
Martin, P., Szymanowska, M., Zwierzchowski, L., and Leroux, C. (2002). The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev. 42, 433–459. doi:10.1051/rnd:2002036
Mcmahon, D. J., and Brown, R. J. (1984). Composition, structure, and integrity of casein micelles: A review. J. Dairy Sci. 67, 499–512. doi:10.3168/jds.S0022-0302(84)81332-6
Mercier, J. C., Chobert, J. M., and Addeo, F. (1976). Comparative study of the amino acid sequences of the caseinomacropeptides from seven species. FEBS Lett. 72, 208–214. doi:10.1016/0014-5793(76)80972-6
Mestawet, T. A., Girma, A., Ådnøy, T., Devold, T. G., Narvhus, J. A., and Vegarud, G. E. (2014). New insights in goat breeds of Ethiopia: High content of αs1-CN and its association with coagulation properties, whey syneresis and micelle size. Small Ruminant Res. 119, 146–155. doi:10.1016/j.smallrumres.2014.02.011
Mestawet, T. A., Girma, A., Adnoy, T., Devold, T. G., and Vegarud, G. E. (2013). Newly identified mutations at the CSN1S1 gene in Ethiopian goats affect casein content and coagulation properties of their milk. J. Dairy Sci. 96, 4857–4869. doi:10.3168/jds.2012-6467
Miocinovic, J., Miloradovic, Z., Josipovic, M., Nedeljkovic, A., Radovanovic, M., and Pudja, P. (2016). Rheological and textural properties of goat and cow milk set type yoghurts. Int. Dairy J. 58, 43–45. doi:10.1016/j.idairyj.2015.11.006
Moatsou, G., Molle, D., Moschopoulou, E., and Valerie, G. (2007). Study of caprine beta-casein using reversed-phase high-performance liquid chromatography and mass spectroscopy: Identification of a new genetic variant. Protein J. 26, 562–568. doi:10.1007/s10930-007-9098-8
Moatsou, G., Samolada, M., Panagiotou, P., and Anifantakis, E. (2004). Casein fraction of bulk milks from different caprine breeds. Food Chem. 87, 75–81. doi:10.1016/j.foodchem.2003.10.020
Molik, E., Bonczar, G., Misztal, T., Żebrowska, A., and Zięba, D. (2012). “The effect of the photoperiod and exogenous melatonin on the protein content in sheep milk,” in Milk protein. Editor H. Wl (Croatia: InTechOpen), 352.
Neveu, C., Molle, D., Moreno, J., Martin, P., and Leonil, J. (2002). Heterogeneity of caprine beta-casein elucidated by RP-HPLC/MS: Genetic variants and phosphorylations. J. Protein Chem. 21, 557–567. doi:10.1023/a:1022433823559
Nguyen, H. T. H., Afsar, S., and Day, L. (2018). Differences in the microstructure and rheological properties of low-fat yoghurts from goat, sheep and cow milk. Food Res. Int. 108, 423–429. doi:10.1016/j.foodres.2018.03.040
Nicolai, M. A., Garro, G., Caira, S., Mauriello, R., Quarto, M., Pascale, S. D., et al. (2021). Occurrence of quantitative genetic polymorphism at the caprine β-CN locus, as determined by a proteomic approach. Int. Dairy J. 112, 104855. doi:10.1016/j.idairyj.2020.104855
O'mahony, J. A., Fox, P. F., Boland, H. S., Othman, O. E., and Ahmed, S. (2014). Milk: An OverviewAnalysis of genetic polymorphisms in the Egyptian goats CSN1S2 using polymerase chain reaction. J. Biol. Sci. 6, 238–241.
Ozdemir, M., Kopuzlu, S., Topal, M., and Bilgin, O. C. (2018). Relationships between milk protein polymorphisms and production traits in cattle: A systematic review and meta-analysis. Arch. Anim. Breed. 61, 197–206. doi:10.5194/aab-61-197-2018
Palmeri, M., Mastrangelo, S., Sardina, M. T., and Portolano, B. (2014). Genetic variability at αs2-casein gene in Girgentana dairy goat breed. Italian J. Animal Sci. 13, 2997. doi:10.4081/ijas.2014.2997
Park, Y. W., and Haenlein, G. F. W. (2017). “Therapeutic, hypo-allergenic and bioactive potentials of goat milk, and manifestations of food allergy,” in Handbook of milk of non-bovine mammals, 151–179.
Park, Y. W., Haenlein, G. F. W., and Wendorff, W. L. (2017). Handbook of milk of non-bovine mammals. Hoboken, NJ, USA: Wiley.
Park, Y. W., Juárez, M., Ramos, M., and Haenlein, G. F. W. (2007). Physico-chemical characteristics of goat and sheep milk. Small Ruminant Res. 68, 88–113. doi:10.1016/j.smallrumres.2006.09.013
Persuy, M. A., Printz, C., Medrano, J. F., and Mercier, J. C. (1999). A single nucleotide deletion resulting in a premature stop codon is associated with marked reduction of transcripts from a goat beta-casein null allele. Anim. Genet. 30, 444–451. doi:10.1046/j.1365-2052.1999.00547.x
Pirisi, A., Colin, O., Laurent, F., Scher, J., and Parmentier, M. (1994). Comparison of milk composition, cheesemaking properties and textural characteristics of the cheese from two groups of goats with a high or low rate of αS1-casein synthesis. Int. Dairy J. 4, 329–345. doi:10.1016/0958-6946(94)90030-2
Prinzenberg, E. M., Gutscher, K., Chessa, S., Caroli, A., and Erhardt, G. (2005). Caprine kappa-casein (CSN3) polymorphism: New developments in molecular knowledge. J. Dairy Sci. 88, 1490–1498. doi:10.3168/jds.S0022-0302(05)72817-4
Quaglia, G. B., and Gennaro, L. (2003). “Enzymes uses in food processing,” in Encyclopedia of food sciences and nutrition. Editor B. Caballero. Second edition (Oxford: Academic Press), 2125–2139.
Rahmatalla, S. A., Arends, D., Said Ahmed, A., Hassan, L. M. A., Krebs, S., Reissmann, M., et al. (2021). Capture sequencing to explore and map rare casein variants in goats. Front. Genet. 12, 620253. doi:10.3389/fgene.2021.620253
Ramunno, L., Cosenza, G., Gallo, D., Illario, R., Rando, A., and Masina, P. (2002). Un nuovo allele al locus CSN1S1 di capra, CSN1S1 NXV Congr, 221.
Ramunno, L., Cosenza, G., Pappalardo, M., Longobardi, E., Gallo, D., Pastore, N., et al. (2001a). Characterization of two new alleles at the goat CSN1S2 locus. Anim. Genet. 32, 264–268. doi:10.1046/j.1365-2052.2001.00786.x
Ramunno, L., Cosenza, G., Rando, A., Illario, R., Gallo, D., Berardino, D. D., et al. (2004). The goat alphas1-casein gene: Gene structure and promoter analysis. Gene 334, 105–111. doi:10.1016/j.gene.2004.03.006
Ramunno, L., Cosenza, G., Rando, A., Pauciullo, A., Illario, R., Gallo, D., et al. (2005). Comparative analysis of gene sequence of goat CSN1S1 F and N alleles and characterization of CSN1S1 transcript variants in mammary gland. Gene 345, 289–299. doi:10.1016/j.gene.2004.12.003
Ramunno, L., Longobardi, E., Pappalardo, M., Rando, A., Di Gregorio, P., Cosenza, G., et al. (2001b). An allele associated with a non-detectable amount of alpha s2 casein in goat milk. Anim. Genet. 32, 19–26. doi:10.1046/j.1365-2052.2001.00710.x
Ramunno, L., Rando, A., Di Gregorio, P., Massari, M., Blasi, M., and Masina, P. (1991). “Struttura genetica di alcune popolazioni caprine allevate in Italia a locus della caseina αs1,” in IX Congress Nazionales ASPA, Milan, Italy.
Rando, A. (1998). GenBank Accession no. AJ011018 [Capra hircus CSN2 gene, exons 1 to 9, allele A]. Available at: http://www.ncbi.nlm.nih.gov/nuccore/AJ011018 (Accessed October 21, 2020).
Rando, A., Pappalardo, M., Capuano, M., Di Gregorio, P., and Ramunno, L. (1996). Two mutations might be responsible for the absence of b-casein in goat milk. Anim. Genet. 27, 31.
Rando, A., Ramunno, L., and Masina, P. (2000). Mutations in casein genes. Zoot. Nutr. Anim. 26, 105–114.
Roberts, B., Ditullio, P., Vitale, J., Hehir, K., and Gordon, K. (1992). Cloning of the goat beta-casein-encoding gene and expression in transgenic mice. Gene 121, 255–262. doi:10.1016/0378-1119(92)90129-d
Robinson, F. (2011). Goats milk - a suitable hypoallergenic alternative? Br. Food J. 103, 198–208. doi:10.1108/00070700110386746
Robinson, F. (2001). Goats milk – A suitable hypoallergenic alternative? Br. Food J. 103, 198–208. doi:10.1108/00070700110386746
Rout, P. K., Kumar, A., Mandal, A., Laloe, D., Singh, S. K., and Roy, R. (2010). Characterization of casein gene complex and genetic diversity analysis in Indian goats. Anim. Biotechnol. 21, 122–134. doi:10.1080/10495390903534622
Sacchi, P., Chessa, S., Budelli, E., Bolla, P., Ceriotti, G., Soglia, D., et al. (2005). Casein haplotype structure in five Italian goat breeds. J. Dairy Sci. 88, 1561–1568. doi:10.3168/jds.S0022-0302(05)72825-3
Sadler, M. J., and Smith, N. (2013). Beta-casein proteins and infant growth and development. infant 9, 173–176.
Schmidely, P., Meschy, F., Tessier, J., and Sauvant, D. (2002). Lactation response and nitrogen, calcium, and phosphorus utilization of dairy goats differing by the genotype for alpha S1-casein in milk, and fed diets varying in crude protein concentration. J. Dairy Sci. 85, 2299–2307. doi:10.3168/jds.S0022-0302(02)74310-5
Selvaggi, M., Laudadio, V., Dario, C., and Tufarelli, V. (2014). Major proteins in goat milk: An updated overview on genetic variability. Mol. Biol. Rep. 41, 1035–1048. doi:10.1007/s11033-013-2949-9
Selvaggi, M., and Tufarelli, V. (2011). “Chapter 1. Caseins of goat and sheep milk: Analytical and technological aspects,” in Casein: Production, uses and health effects. Editors A. M. Ventimiglia, and J. M. Birkenhager (New York: Nova Science Publishers), 1–25.
Shekar, P. C., Goel, S., Rani, S. D., Sarathi, D. P., Alex, J. L., Singh, S., et al. (2006). kappa-casein-deficient mice fail to lactate. Proc. Natl. Acad. Sci. U. S. A. 103, 8000–8005. doi:10.1073/pnas.0601611103
Silva, S. V., Pihlanto, A., and Malcata, F. X. (2006). Bioactive peptides in ovine and caprine cheeselike systems prepared with proteases from Cynara cardunculus. J. Dairy Sci. 89, 3336–3344. doi:10.3168/jds.S0022-0302(06)72370-0
Simões, J., and Gutiérrez, C. (2018). Sustainable goat production in adverse environments: Volume I: Welfare, health and breeding. Berlin, Germany: Springer International Publishing.
Strzelec, E., and Niżnikowski, R. (2011). SNPs polymorphisms in LGB, CSN3 and GHR genes in five goat breeds kept in Poland. Animal Sci. 49, 181–188.
Swaisgood, H. E. (2003). “Chemistry of the caseins,” in Advanced dairy chemistry—1 proteins: Part A/Part B. Editors P. F. Fox, and P. L. H. Mcsweeney. (Boston: Springer US), 139–201.
Tadlaoui Ouafi, A., Babilliot, J. M., Leroux, C., and Martin, P. (2002). Genetic diversity of the two main Moroccan goat breeds: Phylogenetic relationships with four breeds reared in France. Small Ruminant Res. 45, 225–233. doi:10.1016/S0921-4488(02)00111-6
Tailford, K. A., Berry, C. L., Thomas, A. C., and Campbell, J. H. (2003). A casein variant in cow's milk is atherogenic. Atherosclerosis 170, 13–19. doi:10.1016/S0021-9150(03)00131-X
Threadgill, D. W., and Womack, J. E. (1990). Genomic analysis of the major bovine milk protein genes. Nucleic Acids Res. 18, 6935–6942. doi:10.1093/nar/18.23.6935
Tortorici, L., Di Gerlando, R., Mastrangelo, S., Sardina, M. T., and Portolano, B. (2014). Genetic characterisation of CSN2 gene in Girgentana goat breed. Italian J. Animal Sci. 13, 3414. doi:10.4081/ijas.2014.3414
Vacca, G. M., Ali, H. O., Carcangiu, V., Pazzola, M., and Dettori, M. L. (2009a). Genetic structure of the casein gene cluster in the Tunisian native goat breed. Small Ruminant Res. 87, 33–38. doi:10.1016/j.smallrumres.2009.09.034
Vacca, G. M., Ali, H. O., Pazzola1, M., Dettori, M. L., and Carcangiu, V. (2009b). An investigation on allele frequency at the CSN1S2 locus and its relationship with milk parameters in the Sarda goat. J. Anim. Feed Sci. 18, 628–637. doi:10.22358/jafs/66437/2009
Vacca, G. M., Dettori, M. L., Piras, G., Manca, F., Paschino, P., and Pazzola, M. (2014). Goat casein genotypes are associated with milk production traits in the Sarda breed. Anim. Genet. 45, 723–731. doi:10.1111/age.12188
Vacca-Smith, A. M., and Bowen, W. H. (1995). The effect of milk and kappa casein on streptococcal glucosyltransferase. Caries Res. 29, 498–506. doi:10.1159/000262121
Veltri, C., Lagonigro, R., Pietrolà, E., D’andrea, M., Pilla, F., and Chianese, L. (2000). “Molecular characterization of the goat αs2-casein E allele and its detection in goat breeds of Italy,” in Proceedings 7th International Conference on Goats, Tour, France.
Wal, J.-M. (1998). Cow's milk allergens. Allergy 53, 1013–1022. doi:10.1111/j.1398-9995.1998.tb03811.x
Wal, J. M. (2002). Cow's milk proteins/allergens. Ann. Allergy Asthma Immunol. 89, 3–10. doi:10.1016/s1081-1206(10)62115-1
Wang, Q., Huang, Z., Chen, M. J., Huang, S. Z., and Zeng, Y. T. (2001). GenBank Accession no. AF409096 [Capra hircus β-casein precursor (csn2) gene, complete cds]. Available at: http://www.ncbi.nml.nih.gov (Accessed Feb 01, 2021).
Yahyaoui, M. H., Angiolillo, A., Pilla, F., Sanchez, A., and Folch, J. M. (2003). Characterization and genotyping of the caprine kappa-casein variants. J. Dairy Sci. 86, 2715–2720. doi:10.3168/jds.S0022-0302(03)73867-3
Yahyaoui, M. H., Coll, A., Sanchez, A., and Folch, J. M. (2001). Genetic polymorphism of the caprine kappa casein gene. J. Dairy Res. 68, 209–216. doi:10.1017/s0022029901004733
Yangilar, F. (2013). As a potentially functional food: Goats¡¯ milk and products. J. Food Nutr. Res. 1, 68–81. doi:10.12691/jfnr-1-4-6
Yue, X. P., Fang, Q., Zhang, X., Mao, C. C., Lan, X. Y., Chen, H., et al. (2013). Effects of CSN1S2 genotypes on economic traits in Chinese dairy goats. Asian-Australas. J. Anim. Sci. 26, 911–915. doi:10.5713/ajas.2013.13018
Zhang, Y., Chen, R., Ma, H., and Chen, S. (2015). Isolation and identification of dipeptidyl peptidase IV-inhibitory peptides from trypsin/chymotrypsin-treated goat milk casein hydrolysates by 2D-TLC and LC–MS/MS. J. Agric. Food Chem. 63, 8819–8828. doi:10.1021/acs.jafc.5b03062
Zucht, H.-D., Raida, M., Adermann, K., Mägert, H.-J., and Forssmann, W.-G. (1995). Casocidin-I: A casein-αs2 derived peptide exhibits antibacterial activity. FEBS Lett. 372, 185–188. doi:10.1016/0014-5793(95)00974-e
Keywords: milk protein, casein, goat, genetic variations, polymorphisms, nutrition, health
Citation: Rahmatalla SA, Arends D and Brockmann GA (2022) Review: Genetic and protein variants of milk caseins in goats. Front. Genet. 13:995349. doi: 10.3389/fgene.2022.995349
Received: 15 July 2022; Accepted: 03 October 2022;
Published: 07 December 2022.
Edited by:
Andrea Criscione, University of Catania, ItalyReviewed by:
Gabriel Augusto Marques Rossi, Vila Velha University, BrazilCopyright © 2022 Rahmatalla, Arends and Brockmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Siham A. Rahmatalla, c2loYW0ucmFobWF0YWxsYUBodS1iZXJsaW4uZGU=; Gudrun A. Brockmann, Z3VkcnVuLmJyb2NrbWFubkBodS1iZXJsaW4uZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.