 Shuaishuai Wu
Shuaishuai Wu Augustine K. Ballah
Augustine K. Ballah Wenqiang Che
Wenqiang Che Xiangyu Wang
Xiangyu Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 16 November 2022
Sec. Cancer Genetics and Oncogenomics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.961278
This article is part of the Research TopicAdvances in Genetic and Epigenetic Mechanisms of Therapeutic Resistance in CancerView all 11 articles
Today, numerous international researchers have demonstrated that N7-methylguanosine (m7G) related long non-coding RNAs (m7G-related lncRNAs) are closely linked to the happenings and developments of various human beings’ cancers. However, the connection between m7G-related lncRNAs and glioma prognosis has not been investigated. We did this study to look for new potential biomarkers and construct an m7G-related lncRNA prognostic signature for glioma. We identified those lncRNAs associated with DEGs from glioma tissue sequences as m7G-related lncRNAs. First, we used Pearson’s correlation analysis to identify 28 DEGs by glioma and normal brain tissue gene sequences and predicated 657 m7G-related lncRNAs. Then, eight lncRNAs associated with prognosis were obtained and used to construct the m7G risk score model by lasso and Cox regression analysis methods. Furthermore, we used Kaplan-Meier analysis, time-dependent ROC, principal component analysis, clinical variables, independent prognostic analysis, nomograms, calibration curves, and expression levels of lncRNAs to determine the model’s accuracy. Importantly, we validated the model with external and internal validation methods and found it has strong predictive power. Finally, we performed functional enrichment analysis (GSEA, aaGSEA enrichment analyses) and analyzed immune checkpoints, associated pathways, and drug sensitivity based on predictors. In conclusion, we successfully constructed the formula of m7G-related lncRNAs with powerful predictive functions. Our study provides instructional value for analyzing glioma pathogenesis and offers potential research targets for glioma treatment and scientific research.
Glioma is one of the most common aggressive and fatal primary tumors in the central nervous system, accounting for approximately 30% of cases (Mousavi et al., 2022). They are graded by the World Health Organization (WHO) as I to IV with increasing malignancy based on the histopathological characteristics of the tumor (Ostrom et al., 2019). Although genetic and molecular testing has brought advances in disease diagnosis, surgery, radiotherapy, and other comprehensive treatments have brought hope to patients; their prognosis is still poor. It is getting more severe economic pressure and burdening patients, their families, and society (Frances et al., 2022; Haddad et al., 2022). Thus, there is an urgent need to detect glioma-related biomarkers in our clinical care for early identification and diagnosis and to investigate new therapeutic approaches.
Although long non-coding RNA (lncRNA) is non-coding RNA that cannot be translated into protein RNA molecules, several reports have demonstrated that lncRNA regulates tumorigenesis and development (Yang et al., 2016; Chen et al., 2021). For example, LINC01503 promotes the cancer stem cell properties of glial cells by reducing the degradation of GLI2 (Wei et al., 2022). The lncRNA HOXA-AS2 can enhance the expression of KDM2A/JAG1, which can contribute to Treg cell proliferation and immune tolerance in gliomas and promote tumor development (Zhong et al., 2022). The downregulation of lncRNA TTTY15, which targets miR-4500, could regulate the proliferation and apoptosis of A172 glioma cells (Wang et al., 2022). LncRNA IRAIN overexpression inhibits glioma progression and temozolomide resistance by suppressing the PI3 K-related signaling pathway (Guo et al., 2022). LncRNA KB-1460A1.5 suppresses glioma development through the miR-130a-3p feedback loop (Xu et al., 2022). Despite some progress in previous studies, few biomarkers have been studied for lncRNA prognosis to differentiate patients. Therefore, we investigated the prognostic role of m7G-related lncRNAs in glioma by identifying m7G-related DEGs in glioma in order to be able to find more useful biomarkers for glioma.
N7-methylguanosine (m7G) refers to the methylation of guanosine at the N7 position. m7G RNA modification is one of the most common posttranscriptional modifications; it is widely distributed in the 5′hat region of tRNA, rRNA, and eukaryotic mRNA and plays an essential role in gene expression, protein synthesis and transcriptional stability (Pei and Shuman, 2002; Jaffrey, 2014; Song et al., 2020). M7G can regulate the secondary structure of RNA or protein-RNA interaction through electrostatic and spatial effects (Furuichi, 2015). Current studies have demonstrated that almost every stage of the life cycle can be adjusted by m7G modifications, such as transcription (Pei and Shuman, 2002), mRNA splicing (Jiang et al., 2018), nuclear output (Lewis and Izaurralde, 1997), and translation (Marchand et al., 2018). The mutation of m7G methyltransferase is related to many diseases. Mutations, knockouts, and overexpression of m7G-related genes, such as WD repeat domain 4 (WDR4), lead to microcephalic primordial dwarfism (Sauna and Kimchi-Sarfaty, 2011), Nervous system damage (Lin et al., 2018), and impairment of learning and memory abilities (Pereira et al., 2009). Furthermore, METTL1 is an author of m7G, essential for suppressing lung cancer cell migration through m7G editing on RAS and MYC driver genes (Balzeau et al., 2017; Pandolfini et al., 2019). Also, overexpression of mettl1 and bad prognosis of patients with liver cancer is associated with the downregulation of tumor suppressors in hepatocellular carcinoma (Barbieri et al., 2017; Tian et al., 2019). The tRNA N7-methylguanosine modification mediated by METTL1/WDR4 promotes the development of squamous cell carcinoma (Chen et al., 2022). Furthermore, METTL1-m7G-EGFR/EFEMP1 axis is a precise mechanism for bladder cancer development (Ying et al., 2021). Therefore, if we want a further biological understanding of the interaction between lncRNA and cancer, we must study m7G modifications and explore new prognostic and therapeutic markers. In this study, we constructed a formula based on m7G prognosis-related lncRNAs; and verified their outstanding performance in prognosis prediction. The lncRNAs associated with glioma prognosis were also identified, which may provide potential research directions for analyzing glioma’s pathogenesis and clinical treatment.
We downloaded glioma data (GBM and LGG) and normal brain tissue RNA transcriptome data from the Cancer Genome Atlas (TCGA) and the Genotype-Tissue Expression (GTEx) website (698 glioma samples and 1152 normal human brain samples, respectively). Validation data were available from the China Glioma Genome Atlas (CGGA,1018 glioma samples). Meanwhile, clinical information of glioma patients was downloaded from the TCGA and CGGA databases, and patients without follow-up data or an overall survival <30 days were excluded. Since the data in this study were obtained from public databases, ethics committee approval was not required according to the relevant regulations of the databases.
First, we obtained 3 genes from the published article about m7G (Tomikawa, 2018; Pandolfini et al., 2019; Teng et al., 2021). Then we searched for three biological pathways related to m7G in GSEA and extracted genes involved in each pathway. After removing duplicate genes and summarizing the above genes, we obtained 29 genes. Then, we used Wilcoxon’s method (FDR < 0.05, Log2FC >1) to screen the genes with significant differences in the expression level between glioma and normal tissues based on these 29 genes. After deleting the genes with no significant differences, the remaining ones are m7G-related differentially expressed genes (m7G-related DEGs), and named them m7G-related genes (Supplementary Material S1). Expression of m7G-related DEGs samples were visualized using vioplot. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis implemented in R.
First, we performed co-expression analysis of 28 m7G-related genes and lncRNAs in the TCGA and GTEx glioma and normal brain tissue datasets, identifying 657 m7G-related lncRNAs (Pearson correlation coefficients >0.4, p < 0.001, Supplementary Material S2). Secondly, the prognostic relationship of m7G-related lncRNAs was assessed by univariate Cox regression (Supplementary Material S3). In the univariate analysis, the m7G-related lncRNAs with p < 0.01 (539 lncRNAs) were included in the least absolute shrinkage and selection operator (Lasso) regression. The results derived from Lasso regression were then incorporated into a multivariate Cox model to derive eight prognostic m7G-related lncRNAs and create the risk scores (RS)formula:
The “coef m7GLncsigi” in this “risk scores” formula represents the coefficient value, which is the regression coefficient of the 8 prognostic lncRNAs derived from the multifactorial regression analysis. The “EXP m7GLncSigi” in the formula represents the expression levels of the 8 m7G-related lncRNAs. By using the RS formula, we can get the risk value of each patient. And after getting the risk value of all patients, we can find out the median risk value of the patients. According to the median value, we can determine the level of risk of the patients.
Kaplan-Meier (K-M) analysis, time-dependent ROC, principal component analysis (PCA), independent prognostic analysis, nomogram (1-, 3-, and 5-year), calibration curve and the expression level of lncRNAs are used to determine the accuracy of the model. In the CGGA validation sample, we applied the same intermediate values to assess the validity and reliability of our RS formula using the same way as above. We also use the same approach to randomly divide the TCGA data into two groups for internal validation.
We divided the patients into high-risk and low-risk groups based on the median risk score. GSEA (version 4.1.0, (p < 0.05 and FDR <0.25))was used for functional enrichment analysis (Subramanian et al., 2005). The infiltrating fraction of 16 immune cells and the activity of 13 immune-related pathways were measured by ssGSEA (Rooney et al., 2015). We also explored the relationship between risk scores and immune checkpoints in both risk groups (Yao et al., 2021).
To find more drugs for the treatment of glioma, we focused on evaluating and predicting immune-related drugs. According to the online tool Cancer Drug Sensitivity Genomics, the IC50 of different drugs on glioma samples was predicted using the R package ‘pRRophetic’. The main use of ‘pRRophetic’ is to predict phenotypes from gene expression data (to predict clinical outcomes using Cancer Genome Project CGP cell line data), to predict drug sensitivity in external cell lines (CCLE) and also for clinical data prediction.
This study used R software (version 4.1.2) and GSEA software for statistical analysis. Wilcoxon test was used to identify the expression levels of m7G-related DEGs in cancer and normal tissues. Survival curves were generated using the Kaplan-Meier method and compared using the log-rank test. Cox regression and Lasso regression were used to evaluate the prognostic influences of m7G-related lncRNA features.
After analysis, we found that 28 m7G-related genes were significantly differentially expressed between glioma and normal tissues (Figure 1A). Specifically, NUDT11, IFIT5, GEMIN5, METTL1, CYFIP1, NCBP1, WDR4, NUDT10, EIF3D, LARP1, DCP2, DCPS, AGO2, NCBP2, EIF4G3 and LSM1 were highly expressed in tumor samples (p < 0.001). NSUN2, NUDT4, EIF4E2, SNUPN, NCBP3, NUDT3, EIF4E3, EIF4E, NCBP2L, EIF4E1B, EIF4A1 and NUDT4B were lowly expressed in tumor samples (p < 0.001). The expression levels of NUDT16 were not significantly different (p = 0.517) (Figure 1B). In addition, to further understand the intrinsic association between the 28 m7G-related genes, we also performed a correlation analysis. The results showed that the positive correlation between GEMIN5 and NCBP1 was the most significant, and the negative correlation between GEMIN5 and EIF4A1 was the most significant (Figure 1C). The above results suggest some interaction between m7G-related genes in glioma. Then, KEGG pathway analysis showed that m7G-related DEGs were mainly enriched in RNA degradation, Nucleocytoplasmic transport, EGFR tyrosine kinase inhibitor resistance, Longevity regulating pathway, mRNA surveillance pathway, HIF-1 signalling pathway, Insulin signalling pathway, Spliceosome, mTOR signalling pathway, HIF-1 signalling pathway and PI3K-Akt signalling pathway (Figure 1D). GO analysis showed that DEGs were mainly enriched in the regulation of translation, nucleobase-containing compound catabolic process, heterocycle catabolic process, Etc. (Figure 1E).
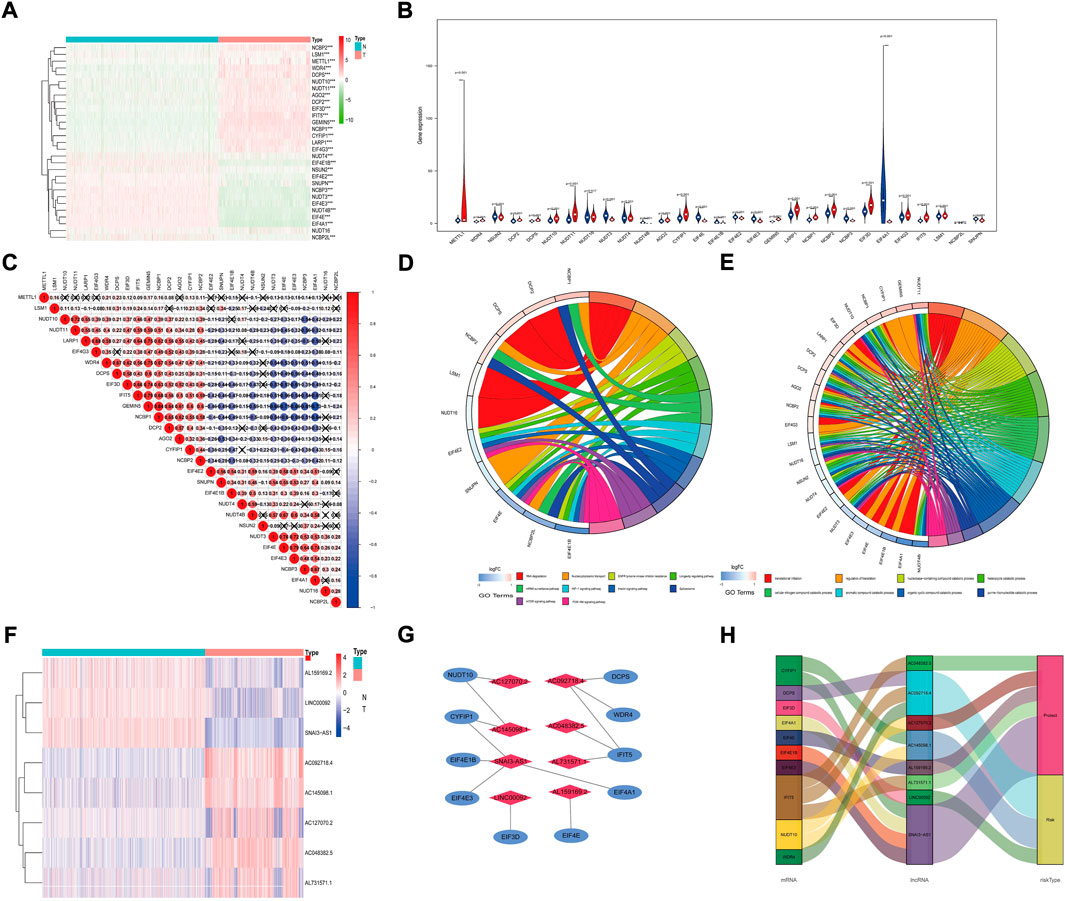
FIGURE 1. Expression of m7G-related mRNAs and prognostic m7G-Related lncRNAs. (A) Heat Map shows the expression levels of eight m7G-related mRNAs. (B) The vioplot shows the differentially m7G-related mRNAs. Blue represents normal sample, and red represents the glioma sample. (C) Spearman correlation analysis of m7G-related mRNAs. (D) KEGG circle diagram of m7G-related DEGs. (E) GO circle diagram of m7G-related DEGs. (F) The expression levels of eight prognostic m7G-Related lncRNAs. (G) The co-expression network of prognostic m7G-related lncRNAs. (H) Sankey diagram of prognostic m7G-Related lncRNAs. lncRNAs, long non-coding RNAs; N, normal; T, tumor.
We identified 658 lncRNAs associated with m7G-related genes. Univariate Cox regression analysis showed that 540 lncRNAs were linked to patient prognosis. One hundred thirty-two were considered risk lncRNAs with HR > 1, while 408 were protective lncRNAs with HR < 1. After Lasso regression, 28 m7G-associated lncRNAs were identified. Finally, multivariate Cox regression identified 8 lncRNAs with the best prognostic correlation (AC048382.5, AC127070.2, AL159169.2, AL731571.1, SNAI3-AS1, AC092718.4, AC145098.1, LINC00092) (Supplementary Material S4). The expression levels of the eight prognostic m7G-related lncRNAs are shown (Figure 1F). We used the Cytoscape and ‘galluvial’ R packages to visualize the lncRNAs. The co-expression network contained 14 lncRNA-mRNA pairs (Figure 1G, R2>0.4, p < 0.001). SNAI3-AS1 was co-expressed with four related genes (EIF4A1, EIF4E3, EIF4E1B, and CYFIP1), AC092718.4 was co-expressed with three related genes (IFIT5, DCPS, and WDR4), and AC145098.1 was co-expressed with two related genes (CYFIP1 and NUDT10), AC127070.2 co-expressed with NUDT10, AC048382.5 and AL731571.1 both co-expressed with IFIT5, LINC00092 co-expressed with EIF3D and AL159169.2 co-expressed with EIF4E. AC048382.5, AC127070.2, AL159169.2, AL731571.1, and SNAI3-AS1 were protective factors, while AC092718.4, AC145098.1, and LINC00092 were risk factors (Figure 1H).
Based on the above eight lncRNAs, we constructed a prognostic model and calculated the risk score for each patient using the risk score model. The risk score formula worked as follows: risk score =(0.620302782 × AC092718.4 expression) + (0.492232265 × LINC00092 expression) + (0.724211508 × AC145098.1 expression) + (-0.922536934 × SNAI3-AS1 expression) + (-0.922536934 × AC048382.5 expression) + (-0.846208391 × AC127070.2 expression) + (-0.924348861 × AL731571.1 expression) + (-0.807182397 × AL159169.2 expression). After obtaining a risk score for each patient, the patients were divided into two groups based on the median risk score: a high-risk group and a low-risk group (Figure 2A). We found that more and more patients died as the risk score increased (Figure 2B). Figure 2C showed eight prognostic m7G-related lncRNAs involved in two groups by heat map. The ROC curve area showed the excellent predictive capability of the model based on eight survival-related lncRNAs. In the TCGA data, the AUC values were 0.905, 0.928, and 0.89 at 1, 3, and 5 years, respectively (Figure 2D). According to KM analysis, patients with high RS had worse survival rates than those with low RS (Figure 2E).
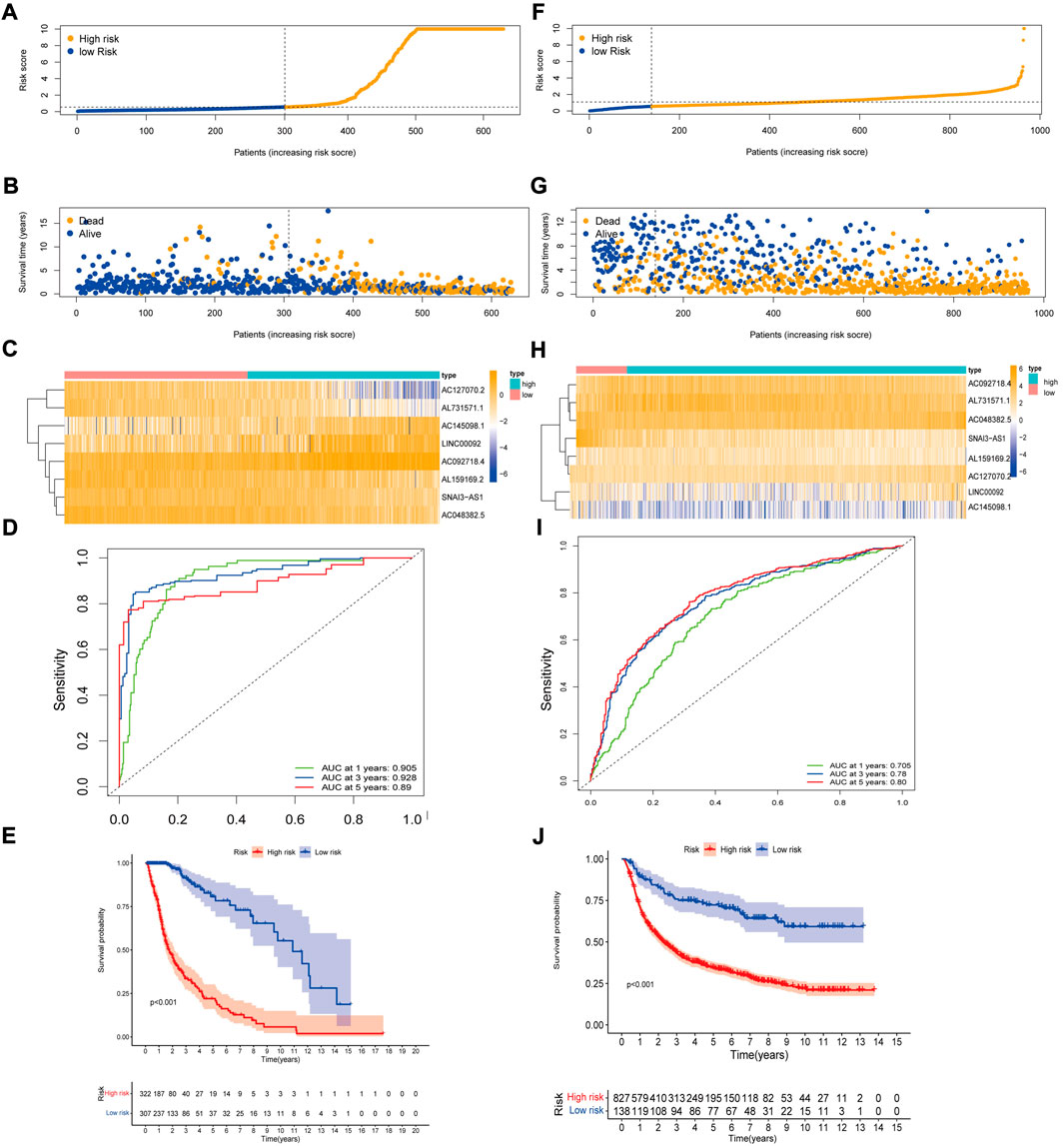
FIGURE 2. Construction and validation of the 8 prognostic m7G-related lncRNAs signature for survival prediction. (A) Distribution of RS in TCGA. (B) Survival time and status of patients in TCGA. (C) Heatmap of prognostic m7G-related lncRNAs of RS in TCGA. (D) ROC curve for TCGA. (E) KM curve for TCGA. (F)Distribution of RS in CGGA. (G)Survival time and status of patients in CGGA. (H)Heatmap of m7G-related lncRNAs of RS in CGGA. (I)ROC curve for CGGA. (J) KM curve for CGGA.
Using the same cut-off from the TCGA data for the CGGA validation data, it was possible to distinguish the high-risk group from the low-risk group. However, the number of patients in the low-risk group was significantly lower (Figure 2F). CGGA patients showed that high-risk patients are positively associated with poor prognosis (Figure 2G). The expression of prognostic m7G-related lncRNAs in CGGA resembled that in TCGA samples (Figure 2H). In the CGGA sample, the AUC values were0.705, 0.78, and 0.80 at 1, 3, and 5 years, respectively (Figure 2I). KM analysis performed on CGGA data showed the same results as TCGA data (p < 0.001, Figure 2J). The validation results in the two validation datasets of TCGA also demonstrate the excellent predictive power of the model (Supplementary Material S5).
The distribution of patients based on whole genes, m7G-related genes, m7G-related lncRNAs, and prognostic m7G-related lncRNAs was visualized using PCA plots. The results showed that m7G survival-associated lncRNA showed the best results (Figures 3A–D). High- and low-risk patients can be distributed in different quadrants according to the RS of prognostic m7G-related lncRNAs.
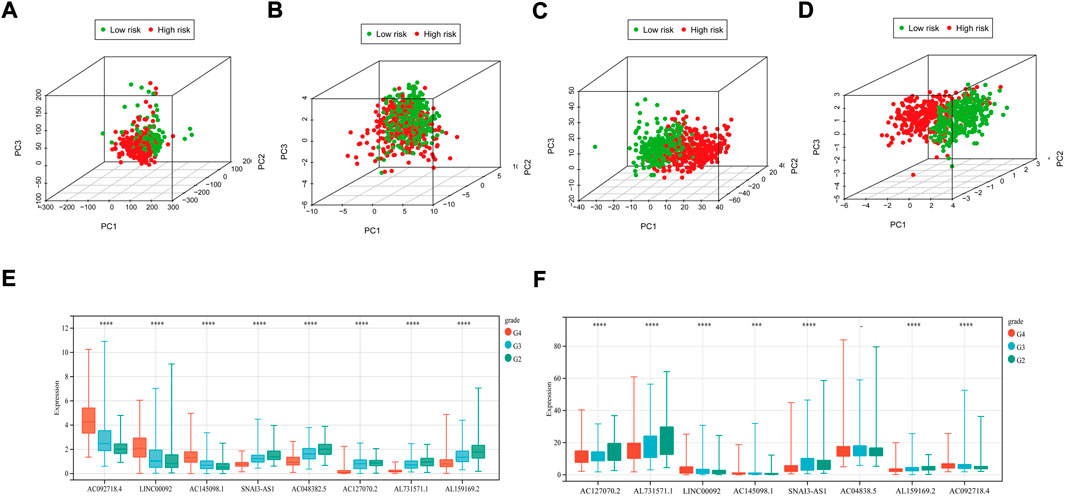
FIGURE 3. PCA maps of the TCGA glioma dataset show the distribution of patients based on the (A) whole genome; (B) m7G-related gene sets; (C)m7G-related lncRNAs; and (D) prognostic m7G-related lncRNAs. Red: high-risk; Green dots: low-risk. Expression profile of 8 prognostic m7G-related lncRNAs with different glioma grades. (E)m7G-related lncRNAs expression with different glioma grades in TCGA datasets. (F) prognostic m7G-related lncRNAs expression with different glioma grades in CGGA datasets. (G2: WHO II, G3: WHO III, G4: WHO IV). *p < 0.05, **p < 0.01 and ***p < 0.001.
We evaluated the expression levels of m7G-related lncRNAs in the TCGA dataset. We found that all genes differed significantly in different grades (Figure 3E), and all but one of the genes had similar trends across stages (Figure 3F). In TCGA and CGGA datasets, the same trend of gene expression was shown with increasing tumor grade.
Using TCGA data, we analyzed the correlations between these clinical variables and the eight lncRNAs risk scores. The risk scores were correlated with age, survival status, and tumor stage; AC048382.5 was associated with age, survival status, and stage; AC127070.2 was correlated with survival status, sex, and stage; AC145098.1 was correlated with survival status and stage; AL159169.2 was correlated with age, survival status, and stage; AL731571.1 was correlated with age, survival status, gender, and staging; and LINC00092 was associated with age and survival status. (Figure 4). The above results showed that our screened m7G-related lncRNAs had the excellent predictive ability.
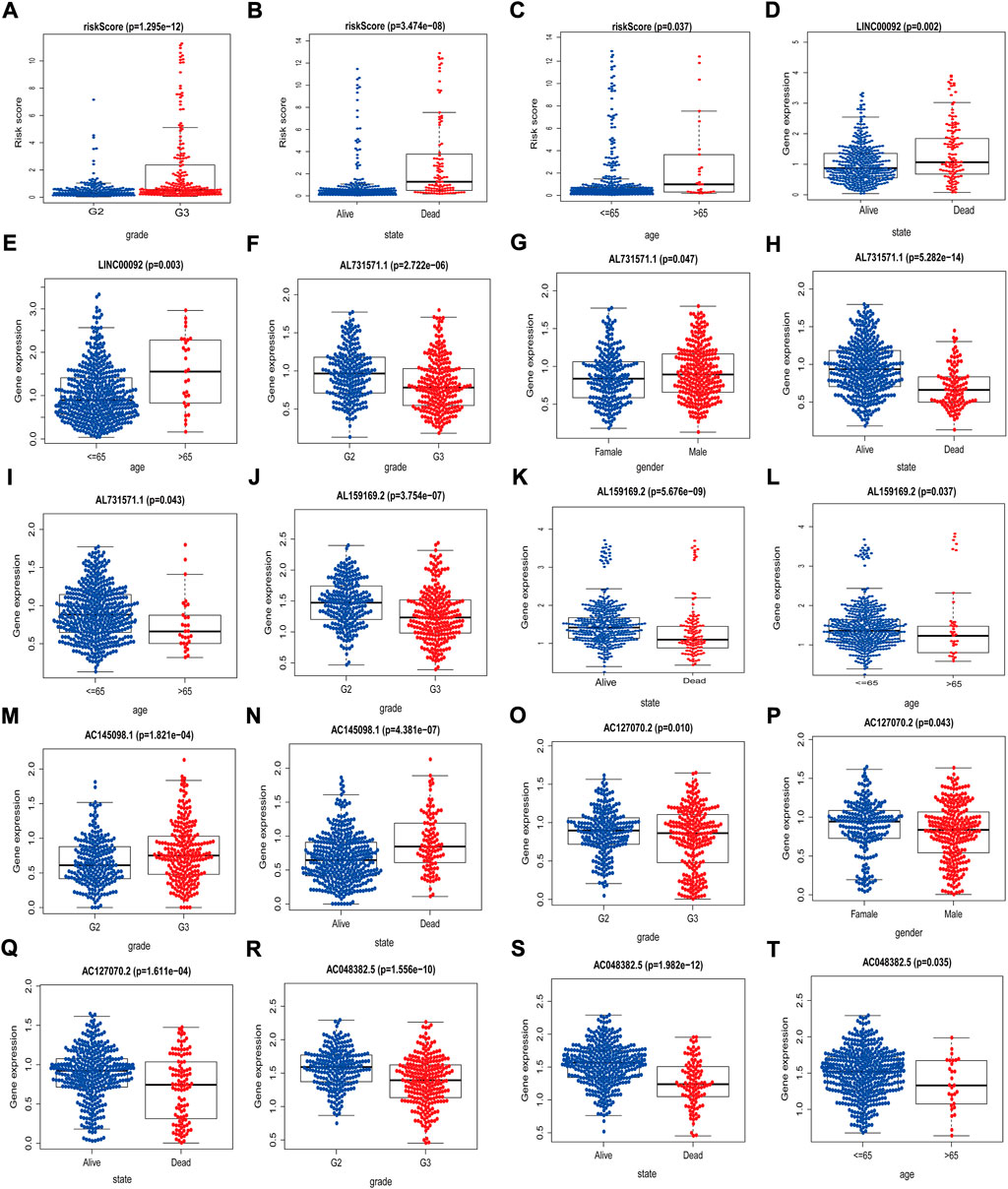
FIGURE 4. Associations between risk scores/related lncRNAs and clinical features. (A–C) Association between risk score and gender, state, and age. (D–E) Association between LINC00092 expression level and state, age. (F–I) Association between AL731571.1 expression level and grade, gender, state and age. (J–L) Association between AL159169.2 expression level and grade, state and age. (M–N) Association between AC145098.1 expression level and grade and state. (O–Q) Association between AL127070.2 expression level and grade, gender and state. (R–T) Association between AC048382.5 expression level and grade, state and age.
In TCGA and CGGA data, we analyzed the independent prognostic factors of glioma patients by Cox regression. Univariate and multivariate Cox regression analyses showed that risk score was an independent predictor (HR = 1.253, 95% CI:1.192–1.317, p < 0.001; HR = 1.127, 95% CI:1.096–1.160, p < 0.001) of OS in glioma patients (Figure 5A, B,D,E). We constructed a column line plot containing clinicopathological variables and risk scores to facilitate clinical work (Figure 5C,F). The calibration curves showed good agreement between actual OS and predicted survival rates (Figures 5J–L).
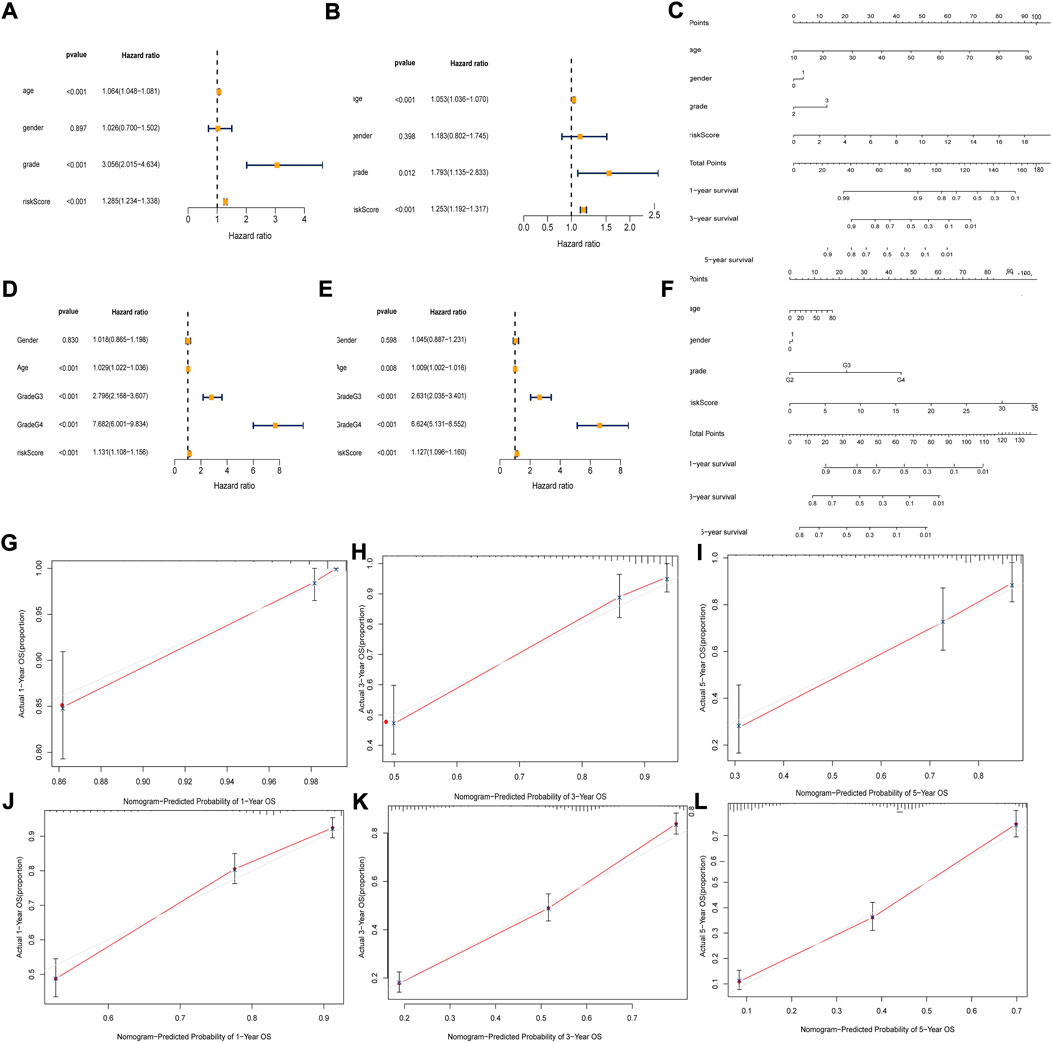
FIGURE 5. Independent prognosis analysis of risk score. (A and D) Univariate COX Forest plot of the risk score in TCGA and CGGA. (B and E) Multivariate COX Forest plot of the risk score in TCGA and CGGA. (C and F) Nomogram based on prognostic features in TCGA and CGGA. (G–I) Calibration plots of the nomogram for predicting the probability of OS at 1, 3, and 5 years in the TCGA. (J–L) Calibration plots of the nomogram for predicting the probability of OS at 1, 3, and 5 years in the CGGA.
We used GSEA to investigate further the differences between the two subgroups for eight m7G-related lncRNAs. In KEGG analysis, the main added functions were systemic lupus-erythematosus, n-glycan-synthesis, and glutathione-metabolism. Decreased functions were wnt-signalling-pathway, taste-transduction, and terpenoid-backbone-biosynthesis (Figure 6A). Most of these pathways are mainly responsible for immune-related diseases and metabolic pathways. So, this suggests that poor prognosis in high-risk patients is likely to be closely related to tumor immune-related pathways.
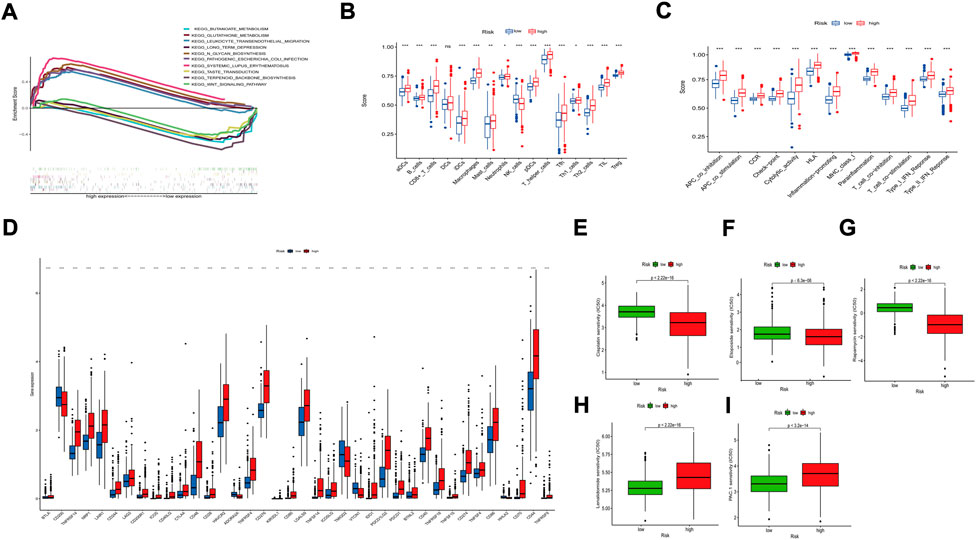
FIGURE 6. Functional enrichment analysis of 8 prognostic m7G-related lncRNAs. (A) KEGG analysis of 8 prognostic m7G-related lncRNAs. (B) The infiltration levels of 16 immune cells. (C) The correlation between the predictive signature and 13 immune-related functions. (D) Expression of immune checkpoints. aDCs, activated dendritic cells; iDCs, immature dendritic cells; NK, natural killer; pDCs, plasmacytoid dendritic cells; Tfh, T follicular helper; Th1, T helper type 1; Th2, T helper type 2; TIL, tumor-infiltrating lymphocyte; Treg, T regulatory cell; APC, antigen-presenting cell; CCR, chemokine receptor; HLA, human leukocyte antigen; MHC, major histocompatibility complex; IFN, interferon. *p < 0.05; **p < 0.01; ***p < 0.001; ns, non-significant. Comparison of treatment drugs sensitivity between high- and low-risk groups. (E–I) IC50 of Cisplatin, Etoposide, Rapamycin, Lenalidomide, PAC.1 in high and low-risk groups. IC50, half-maximal inhibitory concentration.
We quantified the enrichment scores of ssGSEA by measuring the immune cell subpopulations and related pathways to investigate further the correlation between risk scores and immune cells and functions. In the high-risk group, we found a significant rise in most cells (B cells, CD8+ T cells, DCs, Tregs, etc.) (Figure 6B). T-cell-co-inhibition, APC-co-stimulation, CCR, T-cell-co-stimulation, and type I IFN response were higher in the high-risk group than in the low-risk group (Figure 6C). The above results suggest that the high-risk group’s immune function was more active. We also compared the analysis of differences in immune checkpoint expression between the two groups because of the importance of checkpoint-based immunotherapy (Figure 6D).
We also analyzed the correlation between predictive characteristics and tumor immune-related drugs. The results found lower IC50 of Cisplatin, Etoposide, and Rapamycin in the high-risk group and higher IC50 of Lenalidomide and PAC-1 in the high-risk group. (Figures 6E–I), which helps to explore individualized treatment regimens suitable for high-risk patients.
Glioma is a common brain tumor, accounting for 78% of primary malignant brain tumors in the brain, and its overall prognosis has been poor. Therefore, exploring the early diagnosis of glioma and accurately predicting the prognostic markers is of crucial clinical significance (Linzey et al., 2019). Many studies have shown the critical role of m7G in cancer development, mainly focusing on the regulation of tumor cell genesis and progression, but few investigations on cancer prognosis (Orellana et al., 2021; Rong et al., 2021; Xia et al., 2021). Several studies have recently emerged by constructing mRNA and lncRNA predictive signatures associated with glioma autophagy, pyrogenesis, m6A, and ferrogenesis can be used to predict the prognosis of glioma patients (Maimaiti et al., 2022) (Zhou et al., 2021a) (Guan et al., 2021) (Shi et al., 2022). However, the study of prognostic m7G-related lncRNAs in glioma has not been reported. Therefore, we purpose to investigate the prognostic role of m7G-related lncRNA in glioma and provide a new approach for the future clinical treatment of glioma.
This study first obtained 28 DEGs associated with N7-methylguanosine. Then, KEGG analysis showed that DEGs were mainly enriched in RNA degradation, nucleocytoplasmic transport, mRNA surveillance pathway, HIF-1, mTOR, and HIF1-PI3K-Akt signaling pathway. GO analysis showed that DEGs were primarily enriched in the translational initiation activity, regulation of translation, RNA 7−methylguanosine, etc. Existing studies have modified mRNA by adding an m7G 5′ cap to protect mRNA from premature degradation (Kasprzyk and Jemielity, 2021). EGFR plays a crucial role in the METTL1-m7G axis in bladder cancer (Ying et al., 2021). Upregulated WDR4 expression increases m7G methylation levels in hepatocellular carcinoma (Xia et al., 2021). Hickey et al. reported that m7G-MP, the cap analog, is a potent and specific inhibitor of eukaryotic translation (Hickey et al., 1976). The above results suggest that m7G-related genes maybe participate in cancer development through various pathways such as transcription and translation. However, further studies are needed to explore the function of m7G-related genes in glioma.
In addition, there are pieces of evidence that lncRNAs play an essential part in cancer (Ho et al., 2022; Liang et al., 2022; Zhang et al., 2022). SNAI3-AS1, an m7G prognosis-associated lncRNA, is an important tumor modifier in hepatocellular carcinoma tumor progression (Li et al., 2020). Recently, it has been reported that autophagy-related lncRNA features can accurately predict the prognosis of glioma patients (Maimaiti et al., 2022). Ferroptosis-associated lncRNAs can also predict the prognosis of glioma patients (Shi et al., 2022). Therefore, it is important to identify the predictive value of m7G-related lncRNAs in glioma patients and could provide potential directions for future experimental studies of m7G and clinical studies of glioma. In this study, we identified 8 prognostic m7G-related lncRNAs (AC048382.5, AC127070.2, AL159169.2, AL731571.1, SNAI3-AS1, AC092718.4, AC145098.1, LINC00092) for establishing prognostic model. We also found mRNAs (EIF4A1, EIF4E3, EIF4E1B, CYFIP1, DCPS, WDR4, NUDT10, IFIT5, EIF3D, EIF4E) were significantly co-expressed with these lncRNAs. Among them, eIF4E binds the 7-methyl-GTP portion of the 5′ cap structure of cytoplasmic mRNA and plays a part in translation initiation and regulation (Merrick and Pavitt, 2018). Additional studies have found that DCPS acts on m7G through mRNA decay (Ng et al., 2015). WDR4 undergoes a malignant transformation of cells through overexpression of m7G (Orellana et al., 2021). EIF4 acts as a cap-binding protein to enhance m7G cap stabilization of transcripts and plays an important role in malignancy through upregulation (Culjkovic-Kraljacic et al., 2020). In conclusion, the above reports provide evidence for our related studies on N7-methylguanosine. In analyzing two databases with the same median, we found that the number of deaths increased as the risk score increased. The 5-year AUC values (AUC = 0.89, AUC = 0.80) in both TCGA and CGGA data demonstrated the success of the model construction in predicting the prognosis of glioma patients. Furthermore, eight lncRNAs expression in different grades of glioma, the correlation between risk scores and clinical characteristics also increases their predictive power.
Then, GSEA shows that the high-risk group mainly enriched systemic lupus-erythematosus, n-glycan-biosynthesis, glutathione-synthesis, and leukocyte-transendothelial migration. N7-methyladenosine, a common methylation modification of RNA, plays an essential role in autoimmune diseases like RA and SLE (Agris et al., 1992; Zhou et al., 2021b). N-glycan plays a significant part in breast and oral cancers (Hirano and Furukawa, 2022; Wu et al., 2022). Glutathione affects tumor progression by altering oxidative stress sensitivity in astrocytic tumors (Moreira Franco et al., 2021). Increased expression of lymphocyte-specific protein 1 (LSP1) will cause leukocyte migration and inhibition of the immune microenvironment in GBM (Cao et al., 2020). The above results suggest that the occurrence and development of gliomas are also most likely to be closely related to immune-related pathways. The ssGSEA results showed a significant rise in most cells (macrophages, CD8+ T cells, mast cells, Tregs, etc.) in the high-risk group. Some of the above findings have been confirmed by studies. For example, CD8+T-cell infiltration is associated with poor prognosis in patients with BC (Hou et al., 2020; Liu et al., 2020). High infiltration of tumour-associated macrophages was associated with low-grade glioma and thyroid cancer (Ryder et al., 2008; Li et al., 2022). The number of mast cells was positively linked to poor prognosis in patients with prostate cancer (Zhang et al., 2020).
The degree of MC infiltration in mice and human gliomas is proportional to the malignancy of the tumor (Polajeva et al., 2011; Polajeva et al., 2014). The ratio of high neutrophils to lymphocytes predicts a poorer OS in BC patients (Tan, 2017). Pathological grading of gliomas is positively correlated with infiltrating neutrophils (Khan et al., 2020). Increased infiltration of Tregs indicates a poor prognosis in patients with hepatocellular carcinoma (Tu et al., 2016). lncRNA HOXA-AS2 promotes Treg proliferation and immune tolerance through the miR-302A/KDM2A axis to promote glioma progression and poor prognosis (Zhong et al., 2022). Increased Treg and MDSC in mouse gliomas can lead to a decrease in overall survival (Zhai et al., 2021). We found higher HLA and type I IFN response scores in the high-risk group, except for increased tumor immune cell infiltration. Thus, decreased antitumor immunity in high-risk groups may be responsible for poor prognosis. We found significant differences in immune checkpoint expression between the high-risk and low-risk groups. We also studied the sensitivity of immune-related drugs among patients and found that high-risk patients may be sensitive to Cisplatin, Etoposide, and Rapamycin and resistant to Lenalidomide, PAC-1. This implies that high-risk groups may benefit from treatment with multiple immune-related drugs. We hope the above study provides a basis for precise, individualized treatment of glioma patients.
However, our study has some limitations. In the first place, we only used CGGA and CGGA database data for verification and still required external data to test the applicability of predicted signatures. Next, the mechanism of action of m7G-related lncRNAs in glioma needs to be further validated experimentally.
We successfully built a formula for m7G-related lncRNAs with powerful predictive functions and screened lncRNAs with prognostic values. These studies add some instructional value to glioma etiopathogenesis and clinical treatment analysis. And these m7G-related lncRNAs may become new biomarkers and are expected to provide new ideas for glioma therapeutic approaches.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
SW had the initial idea for this study, performed the statistical analysis, and wrote the manuscript; AB and WC and XW revised the manuscript. All authors contributed to the article and approved the submitted version.
The authors would like to thank the TCGA, GTEx, CGGA databases, and GSEA websites for data availability.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.961278/full#supplementary-material
Agris, P. F., Kovacs, S. A., Boak, A. M., and Chen, J. (1992). Spatial localization of distinct rheumatic disease-associated epitopes and the RNA "cap" of the U1 snRNP particle. Immunol. Invest. 21 (2), 111–122. doi:10.3109/08820139209066183
Balzeau, J., Menezes, M. R., Cao, S., and Hagan, J. P. (2017). The LIN28/let-7 pathway in cancer. Front. Genet. 8, 31–16. doi:10.3389/fgene.2017.00031
Barbieri, I., Tzelepis, K., Pandolfini, L., Shi, J., Millan-Zambrano, G., Robson, S. C., et al. (2017). Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature 552 (7683), 126–131. doi:10.1038/nature24678
Cao, J-Y., Guo, Q., Guan, G-F., Zhu, C., Zou, C-Y., Zhang, L-Y., et al. (2020). Elevated lymphocyte specific protein 1 expression is involved in the regulation of leukocyte migration and immunosuppressive microenvironment in glioblastoma. Aging 12 (2), 1656–1684. doi:10.18632/aging.102706
Chen, J., Li, K., Chen, J., Wang, X., Ling, R., Cheng, M., et al. (2022). Aberrant translation regulated by METTL1/WDR4‐mediated tRNA N7‐methylguanosine modification drives head and neck squamous cell carcinoma progression. Cancer Commun. 42, 223–244. doi:10.1002/cac2.12273
Chen, M., Nie, Z., Li, Y., Gao, Y., Wen, X., Cao, H., et al. (2021). A new ferroptosis-related lncRNA signature predicts the prognosis of bladder cancer patients. Front. Cell Dev. Biol. 9, 699804. doi:10.3389/fcell.2021.699804
Culjkovic-Kraljacic, B., Skrabanek, L., Revuelta, M. V., Gasiorek, J., Cowling, V. H., Cerchietti, L., et al. (2020). The eukaryotic translation initiation factor eIF4E elevates steady-state m(7)G capping of coding and non-coding transcripts. Proc. Natl. Acad. Sci. U. S. A. 117 (43), 26773–26783. doi:10.1073/pnas.2002360117
Frances, S. M., Velikova, G., Klein, M., Short, S. C., Murray, L., Wright, J. M., et al. (2022). Long-term impact of adult WHO grade II or III gliomas on health-related quality of life: A systematic review. Neurooncol. Pract. 9 (1), 3–17. doi:10.1093/nop/npab062
Furuichi, Y. (2015). Discovery of m(7)G-cap in eukaryotic mRNAs. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 91 (8), 394–409. doi:10.2183/pjab.91.394
Guan, S., He, Y., Su, Y., and Zhou, L. (2021). A risk signature consisting of eight m6A methylation regulators predicts the prognosis of glioma. Cell. Mol. Neurobiol. 42, 2733–2743. doi:10.1007/s10571-021-01135-x
Guo, A., Lin, R., Zheng, S., Fang, G., Lin, Z., Zhuang, Z., et al. (2022). Overexpression of lncRNA IRAIN restrains the progression and Temozolomide resistance of glioma via repressing IGF-1R-PI3K-NF-κB signalling pathway. Histol. Histopathol. 37 (6), 543–554. doi:10.14670/HH-18-425
Haddad, A. F., Young, J. S., Oh, J. Y., Okada, H., Aghi, M. K., Pereira, K. M., et al. (2022). Clinical characteristics and outcomes of null-cell versus silent gonadotroph adenomas in a series of 1166 pituitary adenomas from a single institution. Neurosurg. Focus 52 (2), E13. doi:10.3171/2020.3.FOCUS20114
Hickey, E. D., Weber, L. A., and Baglioni, C. (1976). Inhibition of initiation of protein synthesis by 7-methylguanosine-5'-monophosphate. Proc. Natl. Acad. Sci. U. S. A. 73 (1), 19–23. doi:10.1073/pnas.73.1.19
Hirano, K., and Furukawa, K. (2022). Biosynthesis and biological significances of LacdiNAc group on N- and O-glycans in human cancer cells. Biomolecules 12 (2), 195. doi:10.3390/biom12020195
Ho, K-H., Shih, C-M., Liu, A-J., and Chen, K-C. (2022). Hypoxia-inducible lncRNA MIR210HG interacting with OCT1 is involved in glioblastoma multiforme malignancy. Cancer Sci. 113 (2), 540–552. doi:10.1111/cas.15240
Hou, W., Xue, M., Shi, J., Yang, M., Zhong, W., Fan, X., et al. (2020). PD-1 topographically defines distinct T cell subpopulations in urothelial cell carcinoma of the bladder and predicts patient survival. Urol. Oncol. 38 (8), e1–e685. doi:10.1016/j.urolonc.2020.04.009
Jaffrey, S. R. (2014). An expanding universe of mRNA modifications. Nat. Struct. Mol. Biol. 21 (11), 945–946. doi:10.1038/nsmb.2911
Jiang, S., Xie, Y., He, Z., Zhang, Y., Zhao, Y., Chen, L., et al. (2018). m6ASNP: a tool for annotating genetic variants by m(6)A function. Gigascience 7 (5). doi:10.1093/gigascience/giy035
Kasprzyk, R., and Jemielity, J. (2021). Enzymatic assays to explore viral mRNA capping machinery. Chembiochem. 22 (23), 3236–3253. doi:10.1002/cbic.202100291
Khan, S., Mittal, S., McGee, K., Alfaro-Munoz, K. D., Majd, N., Balasubramaniyan, V., et al. (2020). Role of neutrophils and myeloid-derived suppressor cells in glioma progression and treatment resistance. Int. J. Mol. Sci. 21 (6), E1954. doi:10.3390/ijms21061954
Lewis, J. D., and Izaurralde, E. (1997). The role of the cap structure in RNA processing and nuclear export. Eur. J. Biochem. 247 (2), 461–469. doi:10.1111/j.1432-1033.1997.00461.x
Li, G., Li, L., Li, Y., Qian, Z., Wu, F., He, Y., et al. (2022). An MRI radiomics approach to predict survival and tumour-infiltrating macrophages in gliomas. Brain. 145, 1151–1161. doi:10.1093/brain/awab340
Li, Y., Guo, D., Lu, G., Mohiuddin Chowdhury, A. T. M., Zhang, D., Ren, M., et al. (2020). LncRNA SNAI3-AS1 promotes PEG10-mediated proliferation and metastasis via decoying of miR-27a-3p and miR-34a-5p in hepatocellular carcinoma. Cell Death Dis. 11 (8), 685. doi:10.1038/s41419-020-02840-z
Liang, J., Liu, C., Xu, D., Xie, K., and Li, A. (2022). LncRNA NEAT1 facilitates glioma progression via stabilizing PGK1. J. Transl. Med. 20 (1), 80. doi:10.1186/s12967-022-03273-2
Lin, S., Liu, Q., Lelyveld, V. S., Choe, J., Szostak, J. W., and Gregory, R. I. (2018). Mettl1/Wdr4-Mediated m(7)G tRNA methylome is required for normal mRNA translation and embryonic stem cell self-renewal and differentiation. Mol. Cell 71 (2), 244–255. doi:10.1016/j.molcel.2018.06.001
Linzey, J. R., Sivakumar, W., Johnson, J. N., Ivan, M. E., Haider, A. S., Philips, C. A., et al. (2019). Young neurosurgeons committee of the American association of neurological surgeons: Training ground for future leaders in organized neurosurgery in the United States of America. World Neurosurg. 123, 59–63. doi:10.1016/j.wneu.2018.11.206
Liu, Z., Zhou, Q., Wang, Z., Zhang, H., Zeng, H., Huang, Q., et al. (2020). Intratumoral TIGIT(+)CD8(+)T-cell infiltration determines poor prognosis and immune evasion in patients with muscle-invasive bladder cancer. J. Immunother. Cancer 8 (2), e000978. doi:10.1136/jitc-2020-000978
Maimaiti, A., Tuerhong, M., Wang, Y., Aisha, M., Jiang, L., Wang, X., et al. (2022). An innovative prognostic model based on autophagy-related long non-coding RNA signature for low-grade glioma. Mol. Cell. Biochem. 477, 1417–1438. doi:10.1007/s11010-022-04368-6
Marchand, V., Ayadi, L., Ernst, F. G. M., Herder, J., Bourguignon-Igel, V., Galvani, A., et al. (2018). AlkAniline-Seq: Profiling of m(7)G and m(3)C RNA modifications at single nucleotide resolution. Angew. Chem. Int. Ed. Engl. 57 (51), 16785–16790. doi:10.1002/anie.201810946
Merrick, W. C., and Pavitt, G. D. (2018). Protein synthesis initiation in eukaryotic cells. Cold Spring Harb. Perspect. Biol. 10 (12), a033092. doi:10.1101/cshperspect.a033092
Moreira Franco, Y. E., Alves, M. J., Uno, M., Moretti, I. F., Trombetta-Lima, M., de Siqueira Santos, S., et al. (2021). Glutaminolysis dynamics during astrocytoma progression correlates with tumor aggressiveness. Cancer Metab. 9 (1), 18. doi:10.1186/s40170-021-00255-8
Mousavi, S. M., Derakhshan, M., Baharloii, F., Dashti, F., Mirazimi, S. M. A., Mahjoubin-Tehran, M., et al. (2022). Non-coding RNAs, and glioblastoma: Insight into their roles in metastasis. Mol. Ther. Oncolytics 24, 262–287. doi:10.1016/j.omto.2021.12.015
Ng, C. K., Shboul, M., Taverniti, V., Bonnard, C., Lee, H., Eskin, A., et al. (2015). Loss of the scavenger mRNA decapping enzyme DCPS causes syndromic intellectual disability with neuromuscular defects. Hum. Mol. Genet. 24 (11), 3163–3171. doi:10.1093/hmg/ddv067
Orellana, E. A., Liu, Q., Yankova, E., Pirouz, M., De Braekeleer, E., Zhang, W., et al. (2021). METTL1-mediated m(7)G modification of Arg-TCT tRNA drives oncogenic transformation. Mol. Cell 81 (16), 3323–3338.e14. doi:10.1016/j.molcel.2021.06.031
Ostrom, Q. T., Cioffi, G., Gittleman, H., Patil, N., Waite, K., Kruchko, C., et al. (2019). CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the United States in 2012-2016. Neuro. Oncol. 21, V1–V100. doi:10.1093/neuonc/noz150
Pandolfini, L., Barbieri, I., Bannister, A. J., Hendrick, A., Andrews, B., Webster, N., et al. (2019). METTL1 promotes let-7 MicroRNA processing via m7G methylation. Mol. Cell 74 (6), 1278–1290. doi:10.1016/j.molcel.2019.03.040
Pei, Y., and Shuman, S. (2002). Interactions between fission yeast mRNA capping enzymes and elongation factor Spt5. J. Biol. Chem. 277 (22), 19639–19648. doi:10.1074/jbc.M200015200
Pereira, P. L., Magnol, L., Sahun, I., Brault, V., Duchon, A., Prandini, P., et al. (2009). A new mouse model for the trisomy of the Abcg1-U2af1 region reveals the complexity of the combinatorial genetic code of down syndrome. Hum. Mol. Genet. 18 (24), 4756–4769. doi:10.1093/hmg/ddp438
Polajeva, J., Bergstrom, T., Edqvist, P-H., Lundequist, A., Sjosten, A., Nilsson, G., et al. (2014). Glioma-derived macrophage migration inhibitory factor (MIF) promotes mast cell recruitment in a STAT5-dependent manner. Mol. Oncol. 8 (1), 50–58. doi:10.1016/j.molonc.2013.09.002
Polajeva, J., Sjosten, A. M., Lager, N., Kastemar, M., Waern, I., Alafuzoff, I., et al. (2011). Mast cell accumulation in glioblastoma with a potential role for stem cell factor and chemokine CXCL12. Plos One 6 (9), e25222. doi:10.1371/journal.pone.0025222
Rong, D., Sun, G., Wu, F., Cheng, Y., Sun, G., Jiang, W., et al. (2021). Epigenetics: Roles and therapeutic implications of non-coding RNA modifications in human cancers. Mol. Ther. Nucleic Acids 25, 67–82. doi:10.1016/j.omtn.2021.04.021
Rooney, M. S., Shukla, S. A., Wu, C. J., Getz, G., and Hacohen, N. (2015). Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell 160 (1-2), 48–61. doi:10.1016/j.cell.2014.12.033
Ryder, M., Ghossein, R. A., Ricarte-Filho, J. C. M., Knauf, J. A., and Fagin, J. A. (2008). Increased density of tumor-associated macrophages is associated with decreased survival in advanced thyroid cancer. Endocr. Relat. Cancer 15 (4), 1069–1074. doi:10.1677/ERC-08-0036
Sauna, Z. E., and Kimchi-Sarfaty, C. (2011). Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 12 (10), 683–691. doi:10.1038/nrg3051
Shi, J., Lai, D., Zuo, X., Liu, D., Chen, B., Zheng, Y., et al. (2022). Identification of ferroptosis-related biomarkers for prognosis and immunotherapy in patients with glioma. Front. Cell Dev. Biol. 10, 817643. doi:10.3389/fcell.2022.817643
Song, B., Tang, Y., Chen, K., Wei, Z., Rong, R., Lu, Z., et al. (2020). m7GHub: deciphering the location, regulation, and pathogenesis of internal mRNA N7-methylguanosine (m(7)G) sites in human. Bioinformatics 36 (11), 3528–3536. doi:10.1093/bioinformatics/btaa178
Subramanian, A., Tamayo, P., Mootha, V. K., Mukherjee, S., Ebert, B. L., Gillette, M. A., et al. (2005). Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U. S. A. 102 (43), 15545–15550. doi:10.1073/pnas.0506580102
Tan, Y. G. (2017). High neutrophil-to-lymphocyte ratio predicts worse overall survival in patients with advanced/metastatic urothelial bladder cancer. Bju Int. 119, 17–18.
Teng, P-C., Liang, Y., Yarmishyn, A. A., Hsiao, Y-J., Lin, T-Y., Lin, T-W., et al. (2021). RNA modifications and epigenetics in modulation of lung cancer and pulmonary diseases. Int. J. Mol. Sci. 22 (19), 10592. doi:10.3390/ijms221910592
Tian, Q-H., Zhang, M-F., Zeng, J-S., Luo, R-G., Wen, Y., Chen, J., et al. (2019). METTL1 overexpression is correlated with poor prognosis and promotes hepatocellular carcinoma via PTEN. J. Mol. Med. 97 (11), 1535–1545. doi:10.1007/s00109-019-01830-9
Tomikawa, C. (2018). 7-Methylguanosine modifications in transfer RNA (tRNA). Int. J. Mol. Sci. 19 (12), E4080. doi:10.3390/ijms19124080
Tu, J-F., Ding, Y-H., Ying, X-H., Wu, F-Z., Zhou, X-M., Zhang, D-K., et al. (2016). Regulatory T cells, especially ICOS+ FOXP3(+) regulatory T cells, are increased in the hepatocellular carcinoma microenvironment and predict reduced survival. Sci. Rep. 6, 35056. doi:10.1038/srep35056
Wang, Y., Du, B., Ma, H., and Zhang, Z. (2022). Down-regulation of lncRNA TTTY15 targeting miR-4500 to inhibit the biological characteristics of A172 glioma cells. Zhonghua Yi Xue Yi Chuan Xue za Zhi 39 (2), 171–175. doi:10.3760/cma.j.cn511374-20201116-00801
Wei, P., Jiang, J., Xiao, M., Zeng, M., Liu, X., Zhao, B., et al. (2022). The transcript ENST00000444125 of lncRNA LINC01503 promotes cancer stem cell properties of glioblastoma cells via reducing FBXW1 mediated GLI2 degradation. Exp. Cell Res. 412 (1), 113009. doi:10.1016/j.yexcr.2022.113009
Wu, Y., Liu, Y., Shang, Z., Liu, X., Xu, Y., and Liu, W. (2022). N-Glycomic profiling reveals dysregulated glycans related to oral cancer using MALDI-MS. Anal. Bioanal. Chem. 414 (5), 1881–1890. doi:10.1007/s00216-021-03822-6
Xia, P., Zhang, H., Xu, K., Jiang, X., Gao, M., Wang, G., et al. (2021). MYC-targeted WDR4 promotes proliferation, metastasis, and sorafenib resistance by inducing CCNB1 translation in hepatocellular carcinoma. Cell Death Dis. 12 (7), 691. doi:10.1038/s41419-021-03973-5
Xu, L., Wu, Q., Yan, H., Shu, C., Fan, W., Tong, X., et al. (2022). Long non-coding RNA KB-1460A1.5 inhibits glioma tumorigenesis via miR-130a-3p/TSC1/mTOR/YY1 feedback loop. Cancer Lett. 525, 33–45. doi:10.1016/j.canlet.2021.10.033
Yang, D., Sun, L., Li, Z., and Gao, P. (2016). “Noncoding RNAs in regulation of cancer metabolic reprogramming,” in Long and short non-coding RNAs in cancer biology. Advances in experimental medicine and biology. Editor E. Song, 191–215.
Yao, J., Chen, X., Liu, X., Li, R., Zhou, X., and Qu, Y. (2021). Characterization of a ferroptosis and iron-metabolism related lncRNA signature in lung adenocarcinoma. Cancer Cell Int. 21 (1), 340. doi:10.1186/s12935-021-02027-2
Ying, X., Liu, B., Yuan, Z., Huang, Y., Chen, C., Jiang, X., et al. (2021). METTL1-m(7) G-EGFR/EFEMP1 axis promotes the bladder cancer development. Clin. Transl. Med. 11 (12), e675. doi:10.1002/ctm2.675
Zhai, L., Bell, A., Ladomersky, E., Lauing, K., Bollu, L., Nuygen, B., et al. (2021). Tumor cell ido enhances immune suppression and decreases survival independent of tryptophan metabolism in glioblastoma. Neuro-Oncology 23, 94. doi:10.1093/neuonc/noab196.371
Zhang, E., Dai, F., Mao, Y., He, W., Liu, F., Ma, W., et al. (2020). Differences of the immune cell landscape between normal and tumor tissue in human prostate. Clin. Transl. Oncol. 22 (3), 344–350. doi:10.1007/s12094-019-02128-5
Zhang, Y., Zhang, Y., Wang, S., Cao, B., Hu, D., Jia, J., et al. (2022). LINC00467 facilitates the proliferation, migration and invasion of glioma via promoting the expression of inositol hexakisphosphate kinase 2 by binding to miR-339-3p. Bioengineered 13 (2), 3370–3382. doi:10.1080/21655979.2021.2018098
Zhong, C., Tao, B., Li, X., Xiang, W., Peng, L., Peng, T., et al. (2022). HOXA-AS2 contributes to regulatory T cell proliferation and immune tolerance in glioma through the miR-302a/KDM2A/JAG1 axis. Cell Death Dis. 13 (2), 160. doi:10.1038/s41419-021-04471-4
Zhou, W., Wang, X., Chang, J., Cheng, C., and Miao, C. (2021). The molecular structure and biological functions of RNA methylation, with special emphasis on the roles of RNA methylation in autoimmune diseases. Crit. Rev. Clin. Lab. Sci.59 (3), 203–218. doi:10.1080/10408363.2021.2002256
Keywords: glioma, lncRNA, m 7 G, prognosis model, immune analysis, therapy
Citation: Wu S, Ballah AK, Che W and Wang X (2022) M7G-related LncRNAs: A comprehensive analysis of the prognosis and immunity in glioma. Front. Genet. 13:961278. doi: 10.3389/fgene.2022.961278
Received: 04 June 2022; Accepted: 02 November 2022;
Published: 16 November 2022.
Edited by:
Peter Hart, Roosevelt University College of Pharmacy, United StatesReviewed by:
Jiani Ma, China University of Mining and Technology, ChinaCopyright © 2022 Wu, Ballah, Che and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiangyu Wang, d2FuZ194eTEyM0AxMjYuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.