
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 31 August 2022
Sec. Epigenomics and Epigenetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.947591
 Evangelos Handakas1
Evangelos Handakas1 Yiwen Xu2Alexa Blair Segal2Maria Carmen Huerta2Kirsty Bowman3,4Laura D. Howe3,4Franco Sassi2
Yiwen Xu2Alexa Blair Segal2Maria Carmen Huerta2Kirsty Bowman3,4Laura D. Howe3,4Franco Sassi2 Oliver Robinson1*
Oliver Robinson1*Biological mechanisms underlying the association between obesity and depression remain unclear. We investigated the role of metabolites and DNA methylation as mediators of the relationship between childhood obesity and subsequent poor mental health in the English Avon Longitudinal Study of Parents and Children. Obesity was defined according to United Kingdom Growth charts at age 7 years and mental health through the Short Mood and Feelings Questionnaire (SMFQ) completed at age 11 years. Metabolites and DNA methylation were measured by nuclear magnetic resonance spectroscopy and Illumina array in blood at the age of 7 years. The associations between obesity and SMFQ score, as continuous count data or using cut-offs to define depressive symptoms (SMFQ >7) or depression (SMFQ >11), were tested using adjusted Poisson and logistic regression. Candidate metabolite mediators were identified through metabolome-wide association scans for obesity and SMFQ score, correcting for false-discovery rate. Candidate DNA methylation mediators were identified through testing the association of putative BMI-associated CpG sites with SMFQ scores, correcting for look-up false-discovery rate. Mediation by candidate molecular markers was tested. Two-sample Mendelian randomization (MR) analyses were additionally applied to test causal associations of metabolites with depression in independent adult samples. 4,018 and 768 children were included for metabolomics and epigenetics analyses, respectively. Obesity at 7 years was associated with a 14% increase in SMFQ score (95% CI: 1.04, 1.25) and greater odds of depression (OR: 1.46 (95% CI: 0.78, 2.38) at 11 years. Natural indirect effects (mediating pathways) between obesity and depression for tyrosine, leucine and conjugated linoleic acid were 1.06 (95% CI: 1.00, 1.13, proportion mediated (PM): 15%), 1.04 (95% CI: 0.99, 1.10, PM: 9.6%) and 1.06 (95% CI: 1.00, 1.12, PM: 13.9%) respectively. In MR analysis, one unit increase in tyrosine was associated with 0.13 higher log odds of depression (p = 0.1). Methylation at cg17128312, located in the FBXW9 gene, had a natural indirect effect of 1.05 (95% CI: 1.01,1.13, PM: 27%) as a mediator of obesity and SMFQ score. Potential biologically plausible mechanisms involving these identified molecular features include neurotransmitter regulation, inflammation, and gut microbiome modulation. These results require replication in further observational and mechanistic studies.
The association between obesity and depression is well established, with bi-directional associations widely reported in observational studies (Milaneschi et al., 2019). Recent Mendelian Randomization (MR) studies have confirmed a causal relationship between obesity and depression (Tyrrell 2019). Poor mental health has been proposed as one mechanism through which obesity during childhood may lead to poorer social and economic outcomes in later life (Segal et al., 2021). It was recently shown in the Avon Longitudinal Study of Parents and Children (ALSPAC) cohort that depressive symptoms mediated approximately 11% of the relationship between body mass index (BMI) and General Certificate of Secondary Education (GCSE) scores in girls, demonstrating the importance of mental health to human capital development (Bowman et al., 2022). Both psycho-social and biological mechanisms appear to be important in the causal chain from obesity to depression. The role of psycho-social mediators such as bullying and social stigma has been relatively well-described in population studies (Gini and Pozzoli, 2009; Moore et al., 2017). However, biological mechanisms may explain a significant proportion of the association between obesity and depression in children, which may have implications for policy interventions to improve mental health.
Proposed biological pathways in the causal pathways between obesity and mental health include alterations to the hypothalamic–pituitary–adrenal (HPA) axis, immuno-inflammatory activation, neuroendocrine regulators of energy metabolism, and the microbiome (Marazziti et al., 2014; Milaneschi et al., 2019). Biological mechanisms may be traced using biomarkers such as DNA methylation, an important regulator of gene expression, and metabolites, small molecules that reflect metabolic alterations. Obesity induces widespread changes in methylation levels across the genome (Wahl et al., 2017; Campanella et al., 2018) that may play a role in mood disorders. Differential methylation of a handful of the same genes involved in inflammatory pathways has been identified across separate studies of obesity and mood disorders (Gharipour et al., 2020). In addition, evidence shows BMI-associated DNA methylation predicts diseases, including T2D and cancers (Wahl et al., 2017). Similarly, as a metabolic disorder, obesity leads to widespread perturbation of the metabolome, which may influence mood. For instance, branched-chain amino acids (BCAAs) may influence brain function by modifying large, neutral amino acid transport at the blood–brain barrier, interfering with neurotransmitter synthesis (Fernstrom, 2005) (Gruenbaum et al., 2019). Recent reviews of the effects of obesity on the epigenome and metabolome in children identified consistent increases in both BCAAs and aromatic amino acids (AAAs) (Handakas et al., 2021) and methylation at multiple CpGs (sites of DNA methylation), albeit with less consistency between studies (Alfano et al., 2021).
In this study, we investigated the role of obesity-related metabolic and epigenetic signatures (measured through nuclear magnetic resonance (NMR) spectroscopy and DNA methylation array, respectively) in the onset of poor mental health in childhood in the ALSPAC cohort. We aimed to test the role of metabolites and differentially methylated CpGs as mediators of the relationship between childhood obesity and subsequent poor mental health or depression, assessed through the Short Mood and Feelings Questionnaire (SMFQ).
The study population included participants from ALSPAC. Initially, the ALSPAC study recruited 14,541 women living in Avon, England, with an expected delivery date between 1 April 1991 and 31 December 1992 (Boyd et al., 2013; Fraser et al., 2013). Participants have been followed up with questionnaires and clinical measures at regular intervals, providing lifestyle, socioeconomic, behavioral and biological data. The ALSPAC study website contains details of all the data that is available through a data search tool (http://www.bris.ac.uk/alspac/researchers/data-access/data-dictionary/). A total of 4,018 had anthropometric data available at 7 years (early childhood) and the SMFQ available at 11 years. The timepoints were chosen to maintain the temporal relationship with the molecular data measured at 7 years and preceding the mental health assessment. A flowchart of individuals that were considered eligible for the analysis is shown in Supporting Information S1 (Supplementary Figure S1).
Child height and weight were measured at a clinic visit at age 7. Obesity was defined as a BMI at or above the 95th percentile of the British 1990 population growth reference (Cole et al., 1995). The Sitar package in R was used to classify the study population as either with obesity or without obesity (Cole and Cole, 2021).
Mental health and depression in children were assessed in the ALSPAC cohort at a clinic visit at age 11 years, using the SMFQ (Ancold and Stephen, 1995). This questionnaire consists of a 13-item checklist of depression symptoms experienced in the previous 2 weeks. The children completed the SMFQ at research clinics with trained interviewers. Each item is rated on a 3-point scale: zero (not true), one (sometimes true) or two (true), giving a total score of 0–26. Higher scores suggest higher depression symptoms. To increase power in our analysis, we primarily used the SMFQ score as a count variable to assess the percentage change in score across the whole score distribution (Delmastro and Zamariola, 2020). In the larger subset with metabolomic data available, we also used pre-defined cut-offs to assess consistency across different definitions commonly used: Children with “depressive symptoms” were defined as those with scores over 7 (Ancold and Stephen, 1995) and “depression” cases were defined as those with scores over 11 (Turner et al., 2014).
Maternal education was classified into three categories according to mothers’ highest educational achievement (Handakas et al.): 1) low: Ordinary- (O-) level, educational qualifications generally obtained at 16 years of age, Certificate of Secondary Education (CSE); 2) intermediate: Advanced- (A-) level; 3) high: university degree and above. Family income at 7 years was classified into two categories: 1) low/medium: <£400 per week, and 2) high: ≥£400. Physical activity at 5 years was classified into two categories based on the frequency of going to the park or playground compared with other children: 1) less often and 2) similarly or more often than other children. Birth weight was obtained from notifications or clinical records. Maternal pre-pregnancy BMI was calculated from self-reported weight and height at the 12th week of gestation.
Metabolomic profiling was carried out using 1H nuclear magnetic resonance spectroscopy on fasting plasma samples from 4,018 individuals of the ALSPAC cohort at 7 years. This molecular signature of systemic metabolism consists of 230 metabolic traits (Supporting Information S2, Codebook). The platform provided quantification of 14 lipoprotein subclasses (particle concentration, lipid concentrations and composition), fatty acids and fatty acid composition, ketone bodies, amino acids, gluconeogenesis-related metabolic traits and glycolysis and gluconeogenesis-related metabolites (Soininen et al., 2009; Kujala et al., 2013). Details related to the used platform have been published previously (Soininen et al., 2009). Data were log-transformed and scaled before analysis.
The Accessible Resource for Integrated Epigenomics Studies (ARIES) is a sub-study drawn from the ALSPAC mother-child cohort that consists of 1,018 mother-child pairs (Relton et al., 2015a) and includes 5469 DNA methylation profiles obtained from umbilical cord blood at birth or at clinic visits at several time points after birth. After DNA extraction, samples were bisulphite converted using the Zymo EZ DNA Methylation ™ kit (Zymo, Irvine, CA, United States). Genome-wide methylation was measured using the Illumina Infinium HumanMethylation450 (HM450) Bead-Chip. The arrays were scanned using an Illumina iScan, and an initial quality review was assessed using GenomeStudio. The preprocessing and normalization of data were carried out using the meffil R package (Min et al., 2018). The LIMS also reported quality control (QC) metrics from the standard control probes on the Illumina 450 K array for each sample (Relton et al., 2015b). Low-quality profiles were removed from further processing. The remaining profiles were normalized using the Functional Normalization algorithm with the first 10 control probe principal components (Fortin et al., 2014). For additional QC steps, we cross-checked candidate CpGs of arrays using a list of cross-reactive probes and polymorphic CpGs (Chen et al., 2013). Moreover, probes as well as samples with more than 5% missing values were removed from the dataset. Finally, we converted the beta values to M-values using the logit transformation to avoid severe heteroscedasticity for highly methylated or unmethylated CpG sites as well as to improve performance in terms of Detection Rate (DR) (Du et al., 2010).
To combine information across loci, we calculated two weighted methylation risk scores (MRS) as the sum of the standardized methylation values. The first risk score was calculated as weighted by marker-specific effect size MRS from the Wahl et al. (2017) study in adults. The second was calculated using CpGs from the Alfano et al. (2021) systematic review, respectively (Figure 1). We calculated the weights for the Alfano et al. (2021) MRS using ALSPAC data and the conducting elastic net using the R Glmnet package (Friedman et al., 2010). Elastic net model parameters, alpha (that defines mixing between lasso and ridge penalties) and lambda (overall strength of penalty), were found following 10-fold cross-validation. The 10-fold routine was performed on the training set (random 80% of the total observations) for each model to estimate elastic net model parameters, alpha and lambda, similar to the methodology of Robinson et al. (2020). The model performance was evaluated on the relevant test set (remaining 20% of the total observations) using a mean square error classifier.
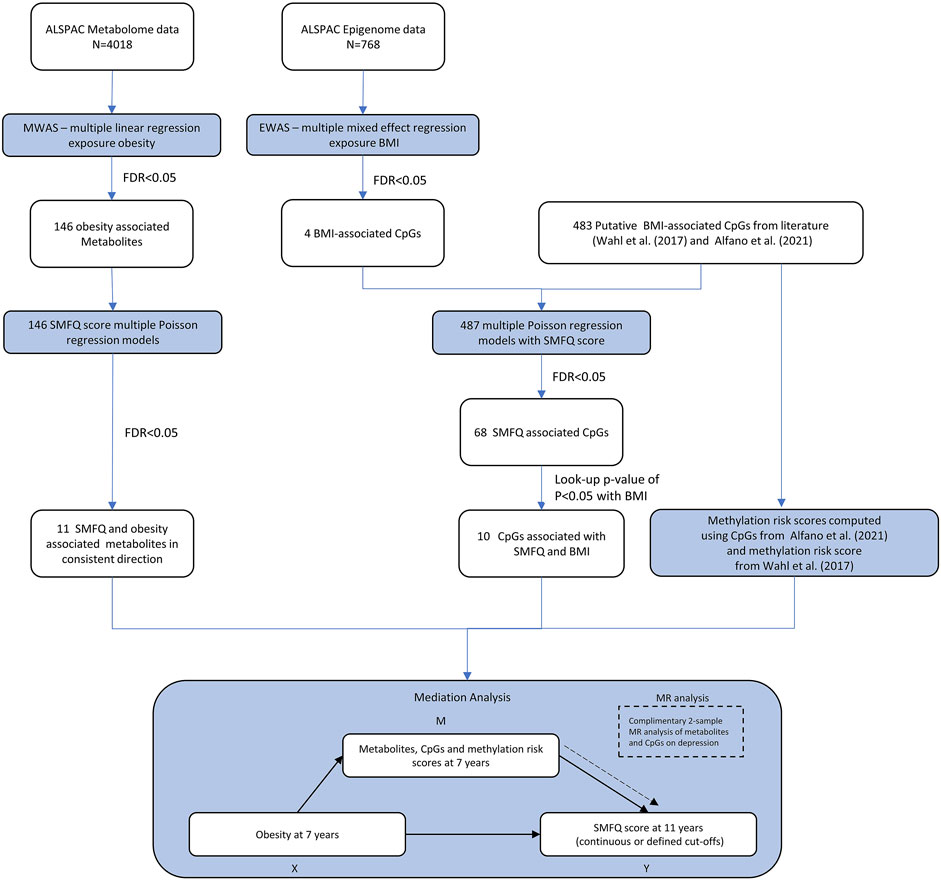
FIGURE 1. Overview of the analysis and methodological workflow. Mediation analysis is a directed acyclic graph analytical framework of the hypothetical causal relationship between an exposure (X) and an outcome (Y) through a mediator (M). The blue rectangles are the statical analysis steps.
To maximize power and potentially reduce bias in our analysis, we applied a multivariable multiple imputation procedure to impute the missing values of covariates. To further ensure the results’ power and reduce potential bias, proportions and patterns of missingness were checked before imputation. Normalized root-mean-square error and out-of-bag error were used as the evaluation metrics of imputation for the set of continuous and categorical variables, respectively. The subsets for epigenetic and metabolomic data were imputed separately. Imputation was carried out based on the multiple chained equations method with the R package mice (Buuren and Groothuis-Oudshoorn, 2010) with the assumption that data were missing at random. First, we performed 100 imputations by 1,000 chains of regression, and then we applied Rubin’s rule (Rubin, 2004) for combining the separate estimates and standard errors from the analytical models performed on each of the 100 imputed datasets.
The metabolomics data were imputed using the random forest (RF) (Breiman, 2001) approach. This is a non-parametric method that allows for interactive and non-linear (regression) effects and has a high performance ability in imputing missing values at random and not at random (Wei et al., 2018). The imputation was carried out using the missForest R package (Stekhoven and Bühlmann, 2012) with default parameter settings.
Logistic regression was used to investigate the association between obesity at 7 years and depression or the presence of depressive symptoms at 11 years (total effect). Poisson regression was used to model the association between obesity at 7 years and SMFQ scores as a continuous (count) variable (total effect). Both models were adjusted for sex, age, mother’s pre-pregnancy BMI, birth weight, physical activity at age 5, maternal education, and family income, selected based on literature and univariate associations (t-test and chi-squared tests) in the dataset.
We applied a Metabolome-Wide Association Study (MWAS) to investigate the associations between obesity at 7 years and metabolomics at 7 years and the associations between metabolomics at 7 years and the depression outcomes at 11 years. Multiple logistic regression models were used to analyze associations between metabolites and depressive symptoms of depression, and multiple Poisson regression models were used to analyze associations between metabolites and the SMFQ score. The models were adjusted for the same covariates aforementioned.
Additionally, an Epigenome-Wide Association Study (EWAS) was applied to investigate the association between DNA methylation and BMI using multiple linear mixed-effects models at the age of 7. The model was adjusted for age, sex, and BMI as fixed effects, and position on chip (“pos”) and beadchip (“slide”) were accounted for through random effects. Additionally, Houseman-estimated cell proportions (Houseman et al., 2012) (B cells, CD8+ T cells, CD4+ T cells, granulocytes, NK cells, and monocytes) were used as fixed effects to adjust for cellular heterogeneity in blood DNA. The methylation beta values were expressed as a logit transformation of the percentage of the methylation (Du et al., 2010). To account for multiple testing, we applied a false discovery rate (FDR) using a cut-off of 5%.
Then, we explored the associations of these CpGs with SMFQ score at 11 years old by filtering the EWAS results by a look-up list of putative BMI-associated CpGs. These included 184 CpGs from the study of Wahl et al. (2017) in adults (Supporting information S2, Supplementary Table S1), and 299 CpGs reported at least once to be associated with adiposity in children from a recent review by Alfano et al. (2021) (Supporting information S2, Supplementary Table S2). Starting from the look-up list of 483 CpGs plus the four CpGs identified in the applied EWAS, we investigated associations with SMFQ score using multiple adjusted Poisson models. CpGs were considered significant using an FDR 5% correction on this CpG list.
Finally, we conducted a sensitivity analysis stratified by sex for both the MWAS and EWAS models.
The analysis workflow is available in Figure 1. The analysis was carried out in the R programming language and these R libraries are presented in Supporting information S1.
Mediation analysis was conducted to assess whether the effect of obesity at age seven on depressive symptoms of depression or SMFQ score at age 11 was mediated by metabolome and DNA methylation profile. In brief, the aim of the mediation analysis was to quantify the effect of X on Y mediated by M (X → M → Y) (natural indirect effect, NIE) and the effect of X on Y that does not operate through the mediator (X → Y): controlled direct effect (CDE), Figure 1.
We used the imputation approach (Vansteelandt et al., 2012) to estimate the conditional natural direct effect (NDE) (not mediated by metabolites and DNA methylation sites) and the conditional natural indirect effect (NIE) (mediated by metabolites and DNA methylation sites) of obesity on depressive symptoms of depression, using the medflex package in R (Steen et al., 2017). We modelled the effect of obesity on SMFQ as a continuous count variable using Poisson regression.
The mediator and outcome models were adjusted for sex, age, mother’s pre-pregnancy BMI, birth weight, physical activity at age 5, maternal education, and family income. For all the mediation models, confidence intervals of 95% (95% CI) were calculated by a non-parametric bootstrap with 1,000 replications. Bootstrapping was used for testing the indirect effect because it does not assume normality in sampling distribution (Hayes and Rockwood, 2017). The mediation proportion was calculated using the following formula: Proportion Mediated = NIE/(NIE + NDE) (Rijnhart et al., 2019).
Similarly, we carried out a sensitivity analysis on the mediation models by applying stratification by sex.
MR analyses were performed to assess the causal influence of molecular features, identified as potential mediators in the observational analyses, on depression in independent adult samples (Figure 1).
Two-sample MR was conducted using the TwoSample R package (Hemani et al., 2018). For the metabolites, genetic instruments were downloaded through the IEU Open GWAS Project (Elsworth et al., 2020). Summary statistics for SNPs associated with metabolites came from the most recent United Kingdom Biobank analysis (IEU Open GWAS Batch ID: “met-d”), while instruments for depression were obtained from analysis in the round four data release from the Finnish FinnGen Biobank ((IEU Open GWAS ID: “finn-a-F5_DEPRESSIO”) https://www.finngen.fi/en).
For CpGs identified in this study, we downloaded available summary statistics from http://mqtldb.godmc.org.uk/. SNPs in linkage disequilibrium were pruned using the European reference panel and default parameters using the “clump_data” function. In the CpG analysis, we used as the instrument for depression the summary statistics from the most recent United Kingdom Biobank analysis (IEU Open GWAS ID: “ukb-d-F5_DEPRESSIO”) as the largest available and non-overlapping study.
After the removal of children without metabolomic data at 7 years and those with a SMFQ score at 11 years, this study included 4,018 children (Supporting information S2, Supplementary Figure S1). Table 1 includes the participant information for subsamples included in the metabolomic and epigenetic analyses. Figure 2A shows the distribution of SMFQ scores, which was left-skewed (Figure 2A). 85 children were classified as being depressed, and 262 children were classified as having depressive symptoms (Table 1).
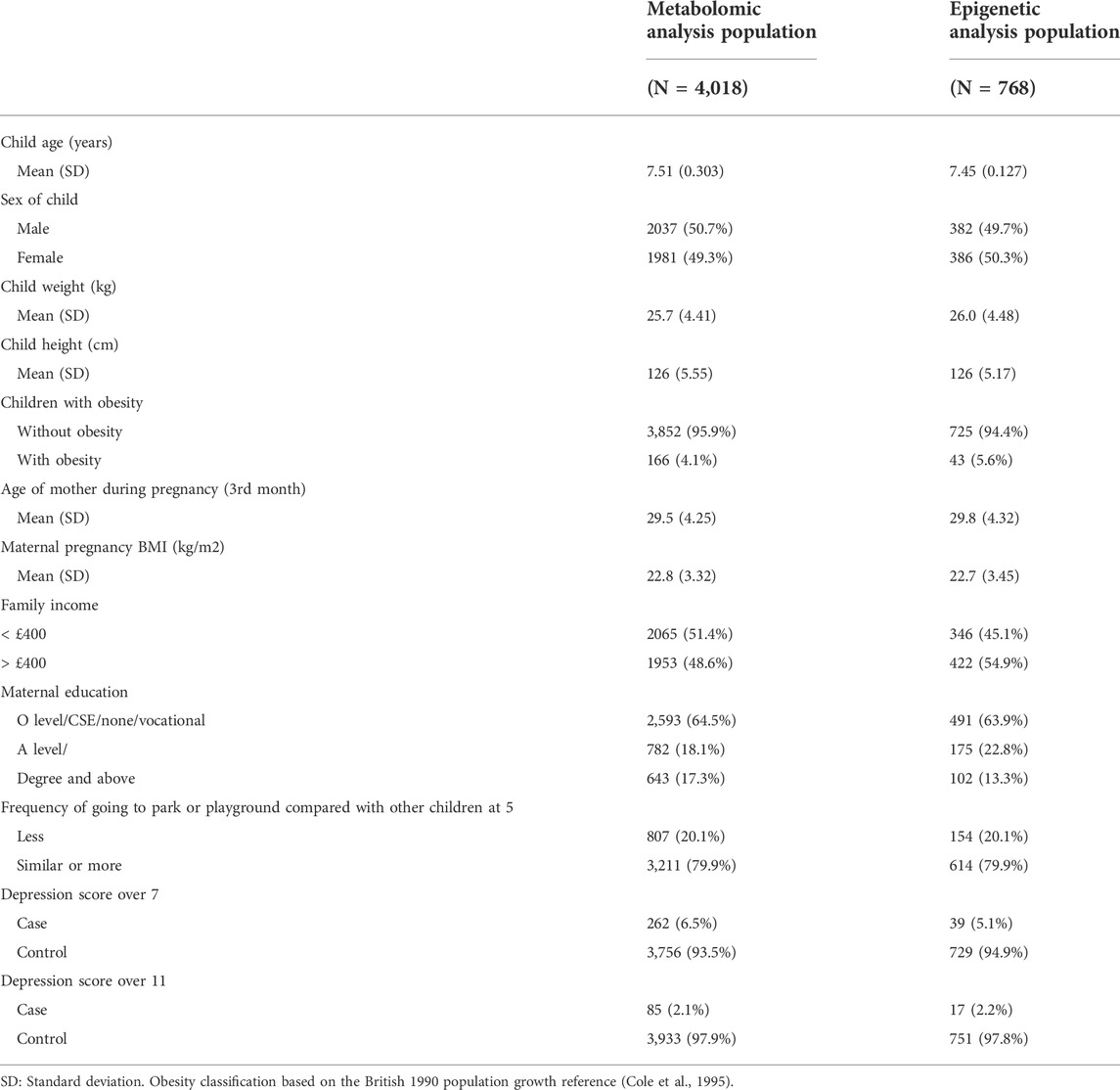
TABLE 1. Descriptive statistics in study population with metabolomic and epigenetic data available.
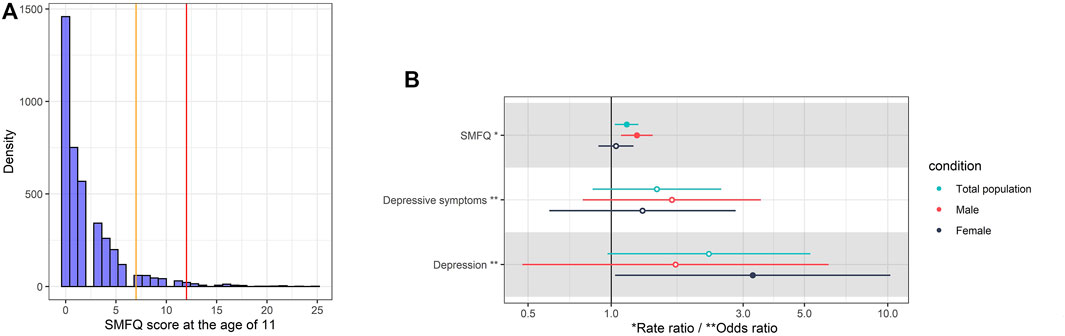
FIGURE 2. (A) Histogram of SMFQ score at 11 years. The vertical orange and red lines are the cut-offs of depressive symptoms (SMFQ>7) and depression (SMFQ>11), respectively. (B) Rate ratio per standard deviation (95% CI) of obesity at 7 years (exposure variable) for SMFQ score (outcome), and odds ratio of obesity at 7 years (exposure variable) for depressive symptoms (SMFQ>7) (outcome), and depression (SMFQ>11) at 11 years (outcome). The model is adjusted for sex and age, physical activity at age 5, mother’s age at birth, birthweight, mother’s pre-pregnancy BMI, average weekly family income at age 7, and mother’s highest education qualification. The error Bars show 95% confidence intervals (calculated through parametric methods).
In adjusted analyses, obesity was associated with a 14% increase in SMFQ score (rate ratio (RR): 1.14, 95% CI: 1.04, 1.25, p = 0.01). Using defined screening cut-offs of SMFQ score, children with obesity had a 46% higher risk (Odds Ratio (OR): 1.46 95% CI: 0.78, 2.38, p = 0.16) of depressive symptoms (SMFQ >7) and a 3-fold higher risk (OR: 2.25, 95% CI: 0.96, 5.24, p = 0.06) of depression (SMFQ >11) at age 11 than their peers without obesity. (Figure 2B).
Stratifying by sex, we observed a stronger effect with SMFQ score and depressive symptoms in boys, but a stronger effect with depression in girls. Obesity was associated with a 23% increase in SMFQ score in boys (OR: 1.23, 95% CI: 1.08, 1.41, p = 0.02) while obesity was associated with a four-fold increased risk of depression in girls (OR: 3.24, 95% CI: 1.04, 10.21, p = 0.04) (Figure 2B).
We examined the associations between obesity and metabolites measured at 7 years using an MWAS approach. As expected, obesity had a strong influence on the metabolome, with 146 metabolic measures associated with obesity after correction for 5% FDR (Supporting Information S2, Supplementary Table S3). Obesity was positively associated with amino acids (tyrosine, phenylalanine, isoleucine, leucine, and valine), fatty acids, glycoprotein acetyls, apolipoprotein B, and small, medium, and large low-density lipoproteins (LDL) and very low-density lipoproteins (VLDL) measures. Obesity was negatively associated with high-density lipoprotein (HDL) measures, apolipoprotein A, degree of unsaturation of fatty acids, lactate, citrate, and glycine.
In MWAS analysis (Figure 3), we observed associations passing FDR 5% with SMFQ score for 83 metabolites, including lower mean diameter of low-density lipoproteins, glycoprotein acetyls, and glucose and higher levels of histidine, leucine, tyrosine, citrate, apolipoproteins, conjugated linoleic acid, and various lipoprotein measures (Figure 3, Supporting Information S2, Supplementary Table S3). No associations with metabolic measures passed FDR correction for depressive symptoms of depression, although the directions of effect were similar to those with the SMFQ score (Supporting Information S2, Supplementary Table S3).
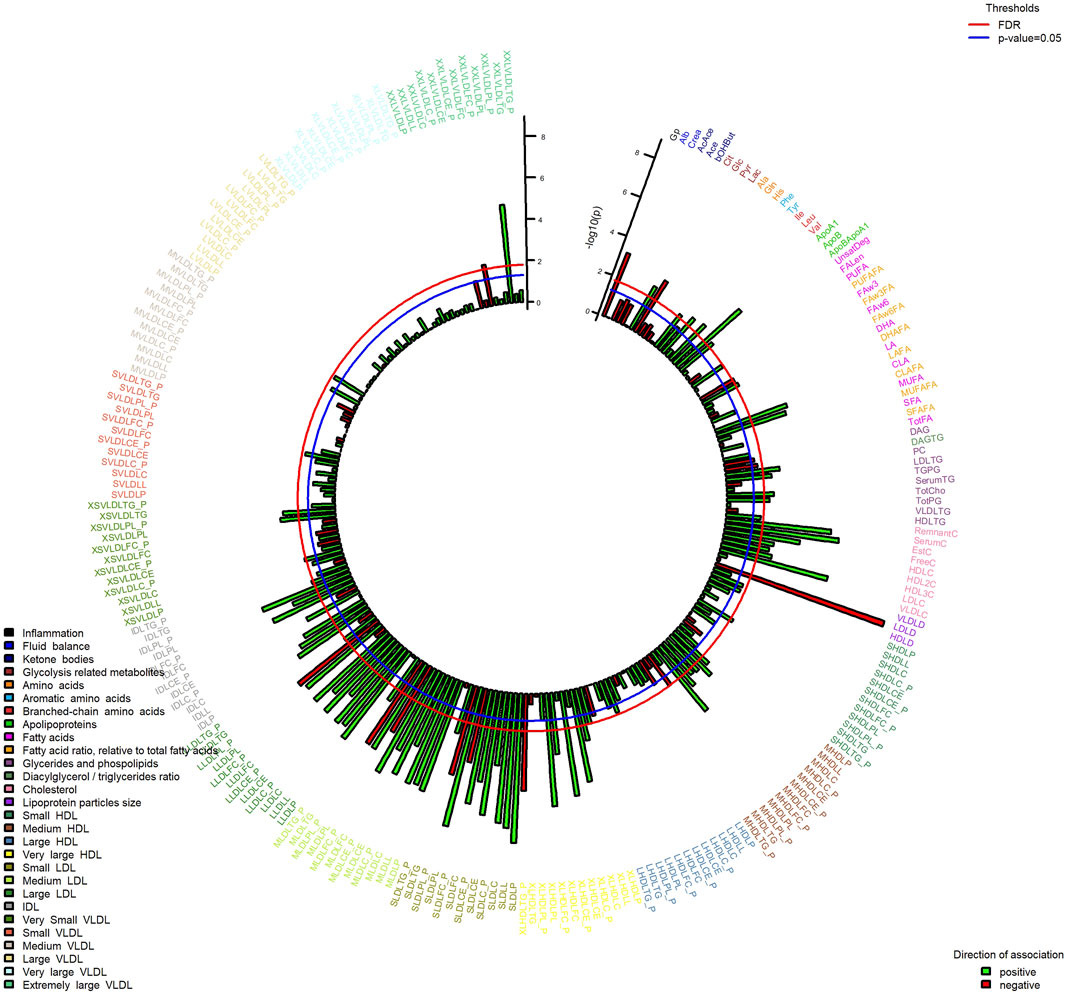
FIGURE 3. Associations of metabolic measures at 7 years SMFQ score at 11 years (outcome). Bars show strength of association (-log10 p-value), colored by direction of effect. Model is adjusted for sex and age of child, physical activity at age 5, mother’s age at birth, birthweight, mother’s pre-pregnancy BMI, average weekly family income at 7, mother’s highest education qualification. FDR at 5%. The abbreviation of the metabolites is available in the Codebook (Supporting Information S2).
To identify candidate metabolites that may play a mediating role between obesity and mental health, we filtered the metabolites by those associated with both obesity and SMFQ score after FDR correction and that had consistent directions of effect. Eleven candidate metabolites were identified as shown in Table 2.
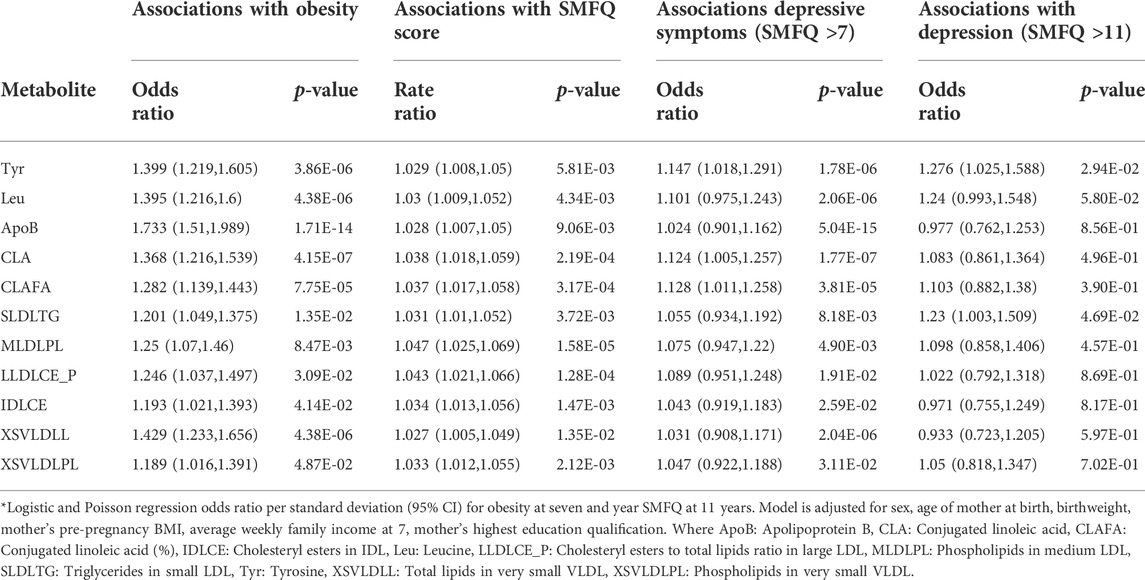
TABLE 2. Metabolites simultaneously associated with obesity at 7 years and with SMFQ score at 11, with consistent directions of effect.
We tested for evidence of mediation by these metabolites of the relationship between obesity at 7 years and SMFQ score at 11 years using a counterfactual framework. As shown in Figure 4 and Supplementary Table S4 (Supporting Information S1), most NIEs were positive, suggesting potential mediation, although for many of the candidate metabolites, effects were small with confidence intervals of the NIEs overlapping 1. The largest effect sizes were noted for tyrosine with a NIE for the SMFQ score of 1.11 (95% CI:1.00, 1.27, Proportion Mediated (PM): 12.5%), For leucine, we observed slightly smaller effects with a NIE for the SMFQ score of 1.08 (95% CI: 0.99, 1.19, PM: 9.3%). Additionally, for conjugated linoleic acid (CLA), we noted that for the SMFQ score the NIE was 1.03 (95% CI: 0.94,1.119, PM: 4.1%). Similar effects were observed when CLA was expressed as a ratio to total fatty acids.
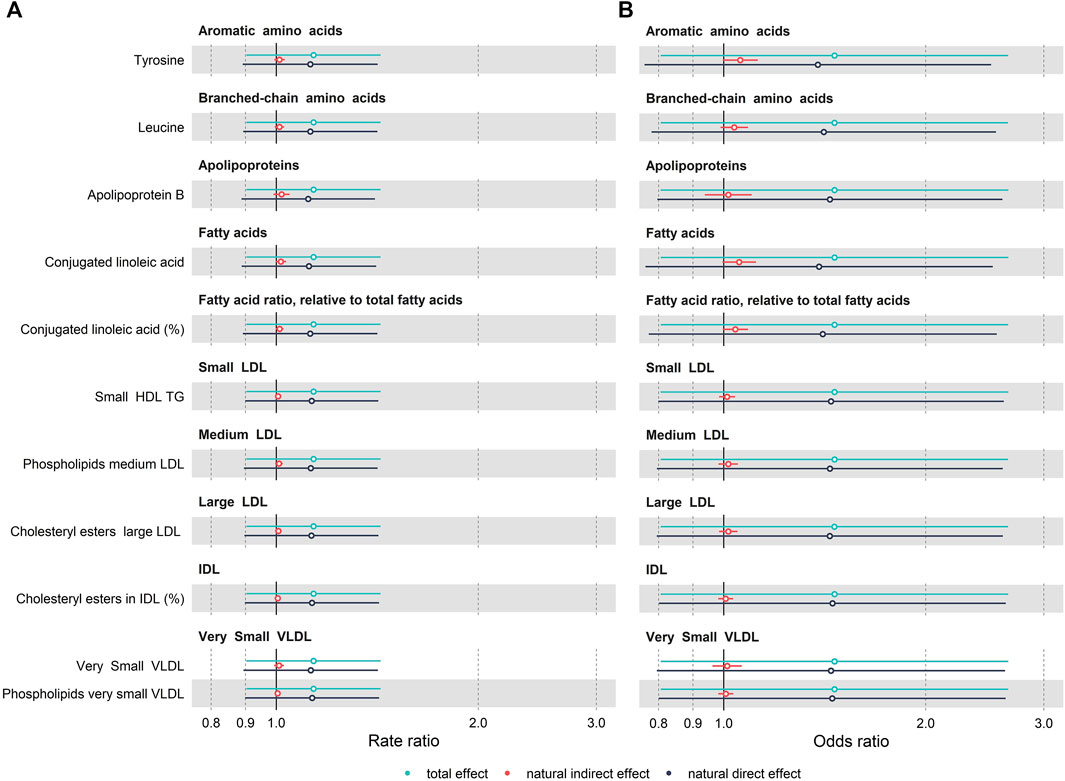
FIGURE 4. Forest plot of mediation analysis for mood related outcomes, (A) SMFQ score and (B) depressive symptoms (SMFQ score> 7), across 11 metabolic compounds. Models are adjusted for sex and age of child, physical activity at the age of 5 years, age of mother at birth, birthweight, mother’s pre-pregnancy BMI, average weekly family income at 7, mother’s highest education qualification. The rate and odds ratio are presented as dots. Bars show 95%CI calculated based on a bootstrapping approach (1,000 replications).
A similar overall pattern was observed using the cut-off to define depressive symptoms (SMFQ>7): for tyrosine, the NIE for the depressive symptoms of 1.01 (95% CI: 0.99, 1.03, PM: 8.9%); for leucine, the NIE was 1.01 (95% CI: 1.00, 1.03, PM: 8.7%); and for CLA, the NIE was 1.02 (95% CI: 1.00, 1.04, PM: 12.5%) (Figure 4). Results using the cut-off to define depression (SMFQ>11) are given in Supporting Information S2, Supplementary Table S4, and follow a similar pattern.
The results of MWAS, stratifying by sex, showed that in the case of boys, 94 metabolites–including histidine, leucine, tyrosine, citrate, apolipoproteins, conjugated linoleic acid and various lipoprotein measures–were associated with SMFQ score after controlling the FDR at 5% (Supporting Information S2, Supplementary Table S5). In the case of girls, we found that two metabolites (very large HDL and lipoprotein particle size) were associated with the SMFQ score after controlling the FDR at 5%. In the mediation analysis, stratified by sex, we observed generally stronger mediation by tyrosine for boys compared to girls (Supporting Information S1, Supplementary Figures S3, S4, S5 and Supporting Information S2, Supplementary Tables S7, S8).
The population with DNA methylation and outcome data included 764 children (Table 1). The low number of children that would be classified as depressed or having depressive symptoms using a binary classification in this subset precluded analyses on these outcomes (Table 1). We therefore only used the SMFQ score as a continuous variable in the epigenetic analysis. In an adjusted analysis in the subset population with epigenetic data available, we found that obesity was associated with a 20% increase in SMFQ score (Rate Ratio (RR): 1.20, 95% CI: 0.98, 1.46). Although the effect size was similar to the larger subset with metabolomic data available, the confidence intervals overlapped 1. Similar results were obtained when stratified by sex: the RR was 1.29 (95% CI: 0.96, 1.74) in boys and 1.16 (95% CI: 0.89, 1.52) in girls.
To select CpGs that may mediate this association, we first performed a genome-wide EWAS for BMI at 7 years. The EWAS analysis showed that cg07462932, cg16332631, cg26271891 and cg26224499 located in the SEPT7, RPS6KA4, ZNF385B and TIMM44 genes respectively were associated with child BMI, after FDR of 5% correction (at genome-level, Table 3, Supporting Information S1, Supplementary Figure S5, Supporting Information S2, Supplementary Table S9).

TABLE 3. Ten strongest associations between DNA methylation site (CpGs) and BMI. The model is adjusted for age, sex, CD4T, Bcell, CD8T, Gran, Mono, NK as fixed effect and chip and position bead array as random effect.
Considering the low statistical power to identify BMI-associated CpGs from within our own dataset, we additionally selected putative BMI-associated CpGs from a look-up list. This included 184 CpGs from the study of Wahl et al. (2017) for adults and 276 CpGs reported to be associated with adiposity in children in at least one study from a recent review (Alfano et al., 2021).
We next explored the associations of these CpGs with the SMFQ score at 11 years old. Starting from the look-up list of 483 CpGs plus the four CpGs identified in the genome wide EWAS, we analyzed associations with SMFQ score using multiple adjusted Poisson models. Sixty-eight of these CpGs were associated with SMFQ after 5% FDR correction on included look-up CpGs (Supporting Information S2, Supplementary Table S10, Figure 5). Four of these (cg04878366, cg25487405, cg09152259, and cg18548288) had a p-value lower than 1.0 × 10−6, which is generally considered a suggestive association in genome-wide epigenetic studies (Duggal et al., 2008).
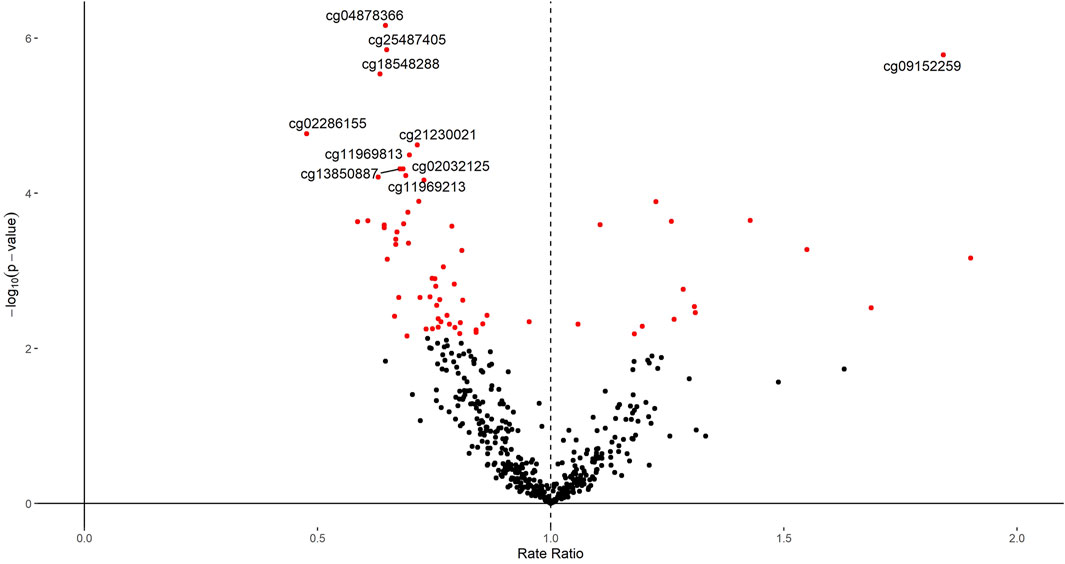
FIGURE 5. Volcano plot of the p-value and β coefficient for the association between 483 putative BMI-associated CpGs and the SMFQ score. The linear mixed-effect model is adjusted for sex and age of child, physical activity at the age of 5 years, age of mother at birth, birthweight, mother’s pre-pregnancy BMI, average weekly family income at 7, mother’s highest education qualification, CD4T, Bcell, CD8T, Gran, Mono, NK as fixed effect and chip and position bead array as random effects. The red dots are the CpGs with FDR<0.05. The vertical black dashed line indicates the rate ratio at 1. The vertical axis is the significance (p-value) on a log10 scale, and the horizontal axis is the beta estimate.
Then, we selected those CpGs associated with 1) BMI and 2) SMFQ in the ALSPAC cohort to further investigate their potential mediation role between obesity at age seven and SMQF at age 11. Ten CpGs remained for mediation analysis (Table 4), after filtering those that had a look-up p-value of p < 0.05 with BMI and an association with SMFQ of FDR-corrected p < 0.05, However, only four of these CpGs–cg17128312, cg25435714, cg26224499, and cg26687842—had matching directions of association (i.e., both hypomethylated or hypermethylated with both BMI and SMFQ score) that may indicate a mediation role.
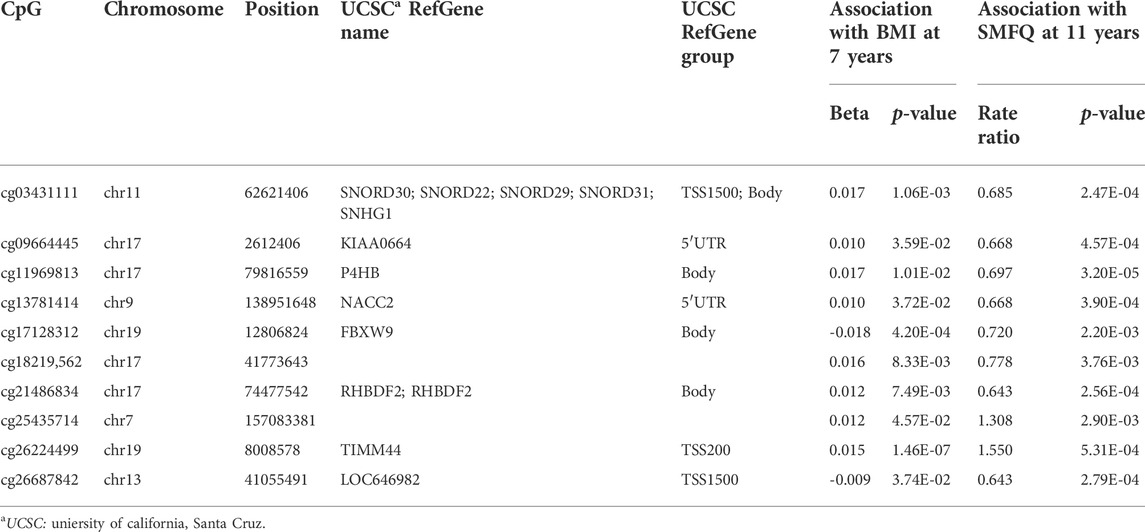
TABLE 4. CpGs simultaneous significantly associated (p < 0.05) with BMI at 7 years and (FDR 5%) SMFQ at 11 years.
The natural indirect effects of these CpGs (mediating pathways) are shown in Figure 6. The mediation analysis indicated that the effect of obesity on SMFQ scores at age 11 may be mediated by cg17128312 (RR: 1.05, 95% CI: 1.01, 1.13, PM: 27%). There was no evidence for mediation by cg26687842 (RR: 1.01, 95% CI: 0.97, 1.05, PM: 3%) or by cg25435714 (RR: 1.00, 95% CI: 0.98, 1.02, and PM: 2%) (Supporting Information S2, Supplementary Table S11). On the other hand, there was evidence of a negative mediation (suppression) by cg1196813 (RR: 0.94, 95% CI: 0.88, 097, PM: 30%).
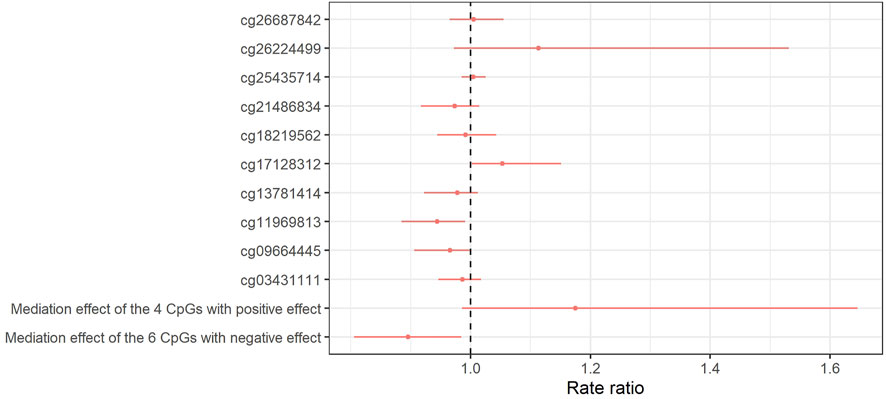
FIGURE 6. Forest plot of the natural indirect effect of the mediation analysis for the relation between obesity at 7 years and SMFQ score at 11 years as mediated by 10 CpGs simultaneously associated (p < 0.05) with BMI and (FDR 5%) SQFM at 11 years. Red dots and bars show rate ratio and 95% CI for natural directed effect, respectively. The model is adjusted for sex, age of mother at birth, birthweight, mother’s pre-pregnancy BMI, average weekly family income at 7, mother’s highest education qualification. 95% CI was calculated by bootstrapping with 1,000 replications.
To combine information across loci, we calculated two weighted MRS: 1) the MRS proposed by Wahl et al. (2017) and 2) based on CpGs reported by (Alfano et al., 2021). The mean square error of the MRS based on Wahl et al. (2017) was 1.95 and the MRS based on CpGs reported by (Alfano et al., 2021) was 0.82, respectively. We did not observe evidence of mediation for either MRS (Figure 7).
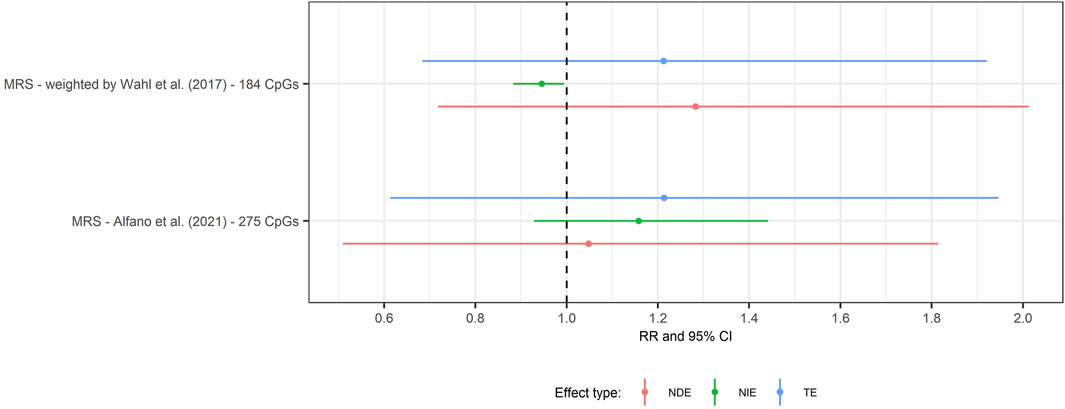
FIGURE 7. Forest plot of the mediation analysis by methylation risk scores at age 7 years of obesity at 7 years and SMFQ score at 11 years, follow-up. Rate ratio and 95% CI(bars) for natural direct effect (red line), natural indirect effect (green line) and total effect (blue line) from single mediation analysis of obesity at 7 years on SMFQ through MRS. Rate ratio refers to MRS 1) based on Wahl et al. (2017) study (Supporting Information S1, Supplementary Table S1) and 2) based on 275 CpGs associated with child adiposity (Alfano et al., 2021) (Supporting Information S1, Supplementary Table S2).
Following the principle of triangulation (Lawlor et al., 2016), we applied two-sample MR based on publicly available summary statistics on candidate metabolites to test causal associations with depression. None of the tested metabolites showed significant associations at p < 0.05. However, again the strongest effects were observed for tyrosine, with one unit increase in tyrosine associated with 0.13 great log odds of depression at p = 0.1, estimated using the inverse-variance weighted method (Table 5). Results estimated with MR-Egger and weighted median were similar and shown in Supplementary Tables S1, S2 (Supporting Information S1), respectively. MR-Egger tests did not indicate the presence of pleiotropy. No evidence of association was observed for the other metabolites tested. We were unable to find genetic instruments for conjugated linoleic acid.
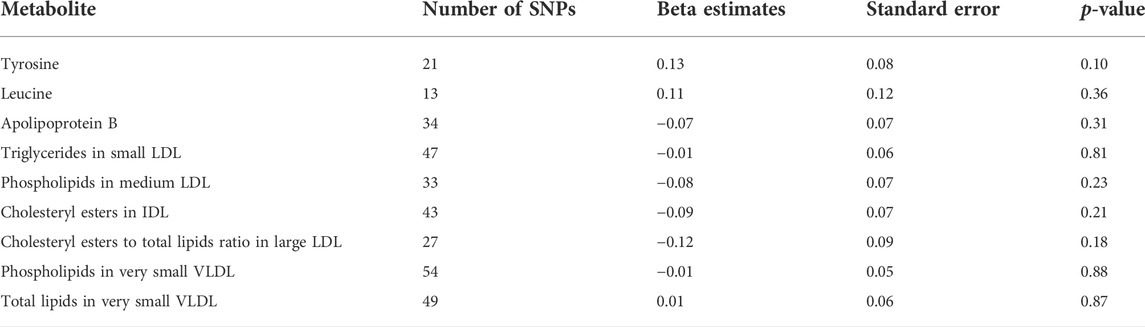
TABLE 5. Two-sample summary statistics Mendelian Randomisation (inverse-variance weighted method) between candidate metabolites and depression. Beta estimate represents the change in the log odds of depression per unit increase in metabolites.
A limited number of genetic instruments were available for only four CpGs that were associated with SMFQ score in the observational analysis: cg09664445 (1 SNP), cg21486834 (1 SNP), cg25435714 (3 SNPs) and cg26687842 (1 SNP). The results of MR analysis are given in Supplementary Table S3 (Supporting Information S1). All associations were non-significant: the strongest association was with cg21486834 in a consistent direction with the observational analysis at p = 0.27.
In this study, we investigated the role of metabolites in blood and DNA methylation in the relationship between obesity and depression in children. We found that the amino acids tyrosine and leucine, and conjugated linoleic acid may mediate the association between obesity at age seven and depression at age 11. While we found less evidence for a role of methylated CpG loci as mediators of the obesity-depression relationship, we identified one CpG, cg17128312, located in the FBXW9 gene, that had a potential mediation effect.
Higher levels of tyrosine (an AAA) and leucine (a BCAA) were associated with obesity, SMFQ, and depression symptoms with consistent effects. Both tyrosine and leucine are widely and consistently associated with childhood obesity (Handakas et al., 2021). A recent study in children showed that high tyrosine levels during life are associated with more internalizing behaviors and negative emotions (Van Vliet et al., 2019). Additionally, tyrosine and the AAA phenylalanine are direct precursors for the synthesis of brain neurons catecholamines (neurotransmitters), including norepinephrine, dopamine, and epinephrine. Studies indicate these neurotransmitters can be manipulated by gut microbiota and vice versa, suggesting an impact on host physiology (Strandwitz, 2018). Animal studies showed that gut microbiota play a causal role in the development of features of depression (Kelly et al., 2016) and anxiety (De Palma et al., 2017), supporting further evidence that there is a modulation of neurotransmission that is likely a route of communication along the gut-brain axis. Plasma AAAs, which are directly modulated by the gut microbiota (Handakas et al., 2021) could therefore influence later propensity for mental health disorders through metabolic processes of gut and brain interaction. BCAAs, including leucine, play an important role in the activation of the mammalian target of rapamycin (mTor) pathway–a pathway involved in the control of cell growth and proliferation–and has been associated with a short-term decrease in depressive symptoms (Baranyi et al., 2016). Additionally, Orešič et al. (2011) found that elevated levels of BCAAs were associated with schizophrenia. BCAAs may compete with AAAs for transport across the brain-blood barriers (Fernstrom, 2005; Gruenbaum et al., 2019), and their increase may lead to concentration decreases in neurotransmitters derived from the AAAs, including tryptophan, in the brain. However, evidence produced in this study contrasts the findings from Baranyi et al., 2016, who observed a reverse association of blood plasma BCAAs in adult patients with depression in their cross-sectional study (Baranyi et al., 2016).
Conjugated linoleic acid (CLA) is a positional isomer of linoleic acid where the two double bonds are conjugated (i.e., separated by a single bond) and may be either in cis- or trans-configurations, with 28 possible isomeric forms (Yang et al., 2015). We found evidence for a mediating role of CLA between obesity and mental health but not for linoleic acid itself. There is mixed evidence for the role of CLA in health, with both athero-protective (Bruen et al., 2017) and pro-inflammatory effects (Mazidi et al., 2017) reported. The effects on inflammation, which may underlie effects on mood, are thought to result from a reduction in the arachidonate pool, leading to a reduced production of downstream eicosanoid products, which modulate cytokine production (Yang et al., 2015). The mixed health effects reported for CLA may be due to differences between isomeric forms (Yang et al., 2015), but we were unable to distinguish these with the NMR analytical platform employed. We also found LDL measures to be associated with both obesity and SMFQ scores, mainly direct associations. Perturbations in lipid metabolism, and lipoprotein concentrations are typical symptoms of childhood obesity (Daniels, 2011; Lamb et al., 2011). Beasley et al. (2005) suggested that cholesterol-mediated alterations in nerve terminal structure and function regulate the relationship between low cholesterol and depression pathogenesis. However, Persons and Fiedorowicz (2016) conducted a meta-analysis on the relationship between depression and serum LDL, and observed a U-shaped relationship between the two, suggesting that both high and low levels of serum LDL are associated with an increased risk of depression. However, despite the associations observed in this study, we did not observe evidence that LDL measures may have a mediating role.
This study showed little evidence for a mediating role of differentially methylated CpGs. We found evidence that methylation cg17128312 had a potential mediation effect. cg17128312 is located in the FBXW9 gene, one of a few evolutionarily conserved F-box proteins. FBXW9 is widely expressed in the nervous system (Gu et al., 1996) and is involved in the promotion of neurotransmitter release from GABAergic motor neurons (Sun et al., 2013), presenting a plausible mechanism related to mood regulation. Although this CpG was identified through the EWAS approach and therefore strongly associated with obesity in this study, it was not among the 483 CpGs identified as putatively associated with body mass in the literature review (Alfano et al., 2021) or the study of (Wahl et al., 2017), suggesting that while it may contribute to poorer mental health among ALSPAC children with obesity, this relationship is unlikely to be generalizable to other populations. To date, few CpGs have been consistently associated with body mass in children (Vehmeijer et al., 2020; Alfano et al., 2021), in contrast to adults (Wahl et al., 2017) and indeed metabolites (Handakas et al., 2021). This lack of consistency may be related to the length of time in an obese state, which is shorter for children (Vehmeijer et al., 2020).
DNA methylation at four CpGs (cg07462932, cg07462932, cg07462932, and cg26224499) was associated with BMI at an FDR of p < 1.06 × 10–7. Additionally, in a look-up analysis on probes previously associated with childhood BMI and obesity, we found that 10 cpgs were associated with both BMI and SMFQ scores, including cg25435714 and cg13781414. Somineni et al. (2019) applied an EWAS analysis in paediatric patients and showed that cg25435714 and cg13781414 were associated with Crohn’s disease (CD), an inflammatory bowel disease, associated with a reduction of faecal microbial diversity (Pascal et al., 2017). A range of studies demonstrated the potential association between obesity and mental disorders and the role of dysbiosis in the gut-brain (Ligezka et al., 2021) and this dysbiosis can alter the functionalities of the central nervous system, behavior and cognitive function (Carding et al., 2015). Studies have shown that bioactive nutrients and gut microbiota can alter either DNA methylation or signatures through a variety of mechanisms (Tateishi et al., 2009; Lester et al., 2013; Alfano et al., 2021). Hence, microbes within the human gut can influence inflammatory cytokines and the production of antimicrobial peptides, affecting the epigenome through their involvement in generating short-chain fatty acids, vitamin synthesis, and nutrient absorption (Alam et al., 2017).
The strength of our study includes both plasma blood metabolome and epigenome from childhood and a large cohort, enabling assessment of the mood disorder at preadolescence and thus limiting reverse causality. The lack of full assessment by trained psychologists is a weakness, although we used the SMFQ that has been developed to enhance epidemiological use (Thapar and McGuffin, 1998) and has demonstrated significant discrimination of depression in several validation studies (Thapar and McGuffin, 1998; Rhew et al., 2010). We included several sociodemographic and clinical factors in our analysis. However, we did not have complete data related to child nutrition that could be linked to both the metabolome and epigenome in life. Nevertheless, we used family socioeconomic factors and maternal clinical factors such as BMI that can reflect general patterns of family nutrition (Williams et al., 2017) and physical activity (Lampinen et al., 2017). Additionally, we used the BMI reference curves for the United Kingdom to define obesity as this BMI chart provides a more detailed monitoring of both the size and shape of the United Kingdom child population. We examined the mediation role of a range of metabolites on the effect of obesity on mood-related outcomes using both single metabolites and an MRS which further revealed the potential mediation role of the metabolic traits in central biological mechanisms. Although the temporal nature of the study design tested the role of obesity-associated molecular features on subsequent depression, we lacked data on mental health at earlier ages to test whether these features may be associated with depression already present at earlier ages (downstream effects). However, the study population had a limited number of cases of depression (less than 5% of the total population), which limited statistical power, particularly for the epigenetic and stratified analyses. For this reason, we also analyzed the SMFQ score using a Poisson regression and used a bootstrapping approach in the mediation analysis. Hence, our results suggest a potential directionality of the effects, encouraging the conduct of further observational and mechanistic studies (Hernán, 2021). Future studies, with gut-microbiota and high-quality dietary data available, should explore the role of nutrition and the mediating role of gut-bacteria in the central nervous system.
In conclusion, we identified three metabolites, tyrosine, leucine and conjugated linoleic acid and methylation at one CpG site that may potentially mediate the association between obesity and later depression in children. Potential biological plausible mechanisms involving these molecular features include regulation and production of neurotransmitters, inflammation, and modulation of the gut microbiome. These results require replication in further well-powered observational and mechanistic studies.
The data used for this submission can be made available on request to the ALSPAC Executive. The ALSPAC data management plan describes in detail the policy regarding data sharing, which is done through a system of managed open access. Full instructions for applying for data access can be found here: http://www.bristol.ac.uk/alspac/researchers/access/.
The studies involving human participants were reviewed and approved by The study has obtained ethical approval from the ALSPAC Ethics and Law Committee and the local research ethics committees in accordance with the Human Tissue Act (Boyd et al., 2013). Consent for biological samples has been collected in accordance with the Human Tissue Act (2004). Informed consent for the use of data collected via questionnaires and clinics was obtained from participants following the recommendations of the ALSPAC Ethics and Law Committee at the time. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
OR and FS designed the study. EC and RX performed the statistical analysis. EC and OR wrote the first draft of the manuscript. All authors critically reviewed and edited the manuscript.
This study was supported by a project funded by the Health Foundation (grant reference 809008) and grant 774548 from the European Union’s Horizon 2020 research and innovation program. OR was supported by a UKRI Future Leaders Fellowship (MR/S03532X/1). LDH was supported by an MRC Career Development Award (MR/M020894/1).This study was supported by grant 217065/Z/19/Z from the United Kingdom Medical Research Council (MRC) and Wellcome Trust and the University of Bristol (core support for the Avon Longitudinal Study of Parents and Children (ALSPAC)); The collection of primary exposure and outcomes data in ALSPAC that were evaluated in this study were supported by grants 076467/Z/05/Z, 086676/Z/08/Z, 086684, 092731 and MC_UU_12013/1 from MRC, grant from Wellcome Trust, grants 5R01HL071248–07, R01 DK077659 from NIH, grant from British Heart Foundation.
We want to acknowledge the participants and investigators of the ALSPAC studies. We are extremely grateful to all the families who took part in this study; the midwives for their help in recruiting them and the whole ALSPAC team, which includes interviewers, computer and laboratory technicians, clerical workers, research scientists, volunteers, managers, receptionists, and nurses.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.947591/full#supplementary-material
Alam, R., Abdolmaleky, H. M., and Zhou, J. R. (2017). Microbiome, inflammation, epigenetic alterations, and mental diseases. Am. J. Med. Genet. B Neuropsychiatr. Genet. 174 (6), 651–660. doi:10.1002/ajmg.b.32567
Alfano, R., Robinson, O., Handakas, E., Nawrot, T. S., Vineis, P., and Plusquin, M. (2021). Perspectives and challenges of epigenetic determinants of childhood obesity: a systematic review. Obes. Rev. 23, e13389. doi:10.1111/obr.13389
Ancold, A., and Stephen, C. (1995). Development of a short questionnaire for use in epidemiological studies of depression in children and adolescents. Age (years) 6 (11), 237–249.
Aquilani, R., Boselli, M., Boschi, F., Viglio, S., Iadarola, P., Dossena, M., et al. (2008). Branched-chain amino acids may improve recovery from a vegetative or minimally conscious state in patients with traumatic brain injury: a pilot study. Arch. Phys. Med. Rehabil. 89 (9), 1642–1647. doi:10.1016/j.apmr.2008.02.023
Baranyi, A., Amouzadeh-Ghadikolai, O., von Lewinski, D., Rothenhäusler, H.-B., Theokas, S., Robier, C., et al. (2016). Branched-chain amino acids as new biomarkers of major depression-a novel neurobiology of mood disorder. PloS one 11 (8), e0160542. doi:10.1371/journal.pone.0160542
Beasley, C. L., Honer, W. G., von Bergmann, K., Falkai, P., Lütjohann, D., and Bayer, T. A. (2005). Reductions in cholesterol and synaptic markers in association cortex in mood disorders. Bipolar Disord. 7 (5), 449–455. doi:10.1111/j.1399-5618.2005.00239.x
Bowman, K., Cadman, T., Robinson, O., Hughes, A., Heron, J., Segal, A. B., et al. (2022). Mediators of the association between childhood BMI and educational attainment: analysis of a UK prospective cohort study. medRxiv 2006, 22276640. doi:10.1101/2022.06.20.22276640
Boyd, A., Golding, J., Macleod, J., Lawlor, D. A., Fraser, A., Henderson, J., et al. (2013). Cohort profile: the ‘children of the 90s’—the index offspring of the avon longitudinal study of parents and children. Int. J. Epidemiol. 42 (1), 111–127. doi:10.1093/ije/dys064
Bruen, R., Fitzsimons, S., and Belton, O. (2017). Atheroprotective effects of conjugated linoleic acid. Br. J. Clin. Pharmacol. 83 (1), 46–53. doi:10.1111/bcp.12948
Buuren, S. v., and Groothuis-Oudshoorn, K. (2010). mice: Multivariate imputation by chained equations in R. J. Stat. Softw., 1–68.
Campanella, G., Gunter, M. J., Polidoro, S., Krogh, V., Palli, D., Panico, S., et al. (2018). Epigenome-wide association study of adiposity and future risk of obesity-related diseases. Int. J. Obes. 42 (12), 2022–2035. doi:10.1038/s41366-018-0064-7
Carding, S., Verbeke, K., Vipond, D. T., Corfe, B. M., and Owen, L. J. (2015). Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 26 (1), 26191. doi:10.3402/mehd.v26.26191
Chen, Y.-a., Lemire, M., Choufani, S., Butcher, D. T., Grafodatskaya, D., Zanke, B. W., et al. (2013). Discovery of cross-reactive probes and polymorphic CpGs in the Illumina Infinium human methylation450 microarray. Epigenetics 8 (2), 203–209. doi:10.4161/epi.23470
Cole, T., and Cole, M. T. (2021). Package ‘sitar’ version 1.1.2. Available at: https://CRAN.R-project.org/package=sitar.
Cole, T. J., Freeman, J. V., and Preece, M. A. (1995). Body mass index reference curves for the UK, 1990. Arch. Dis. Child. 73 (1), 25–29. doi:10.1136/adc.73.1.25
Daniels, S. R. (2011). Lipid concentrations in children and adolescents: It is not all about obesity. Am. J. Clin. 94 (3), 699–700.
De Palma, G., Lynch, M. D., Lu, J., Dang, V. T., Deng, Y., Jury, J., et al. (2017). Transplantation of fecal microbiota from patients with irritable bowel syndrome alters gut function and behavior in recipient mice. Sci. Transl. Med. 9 (379), eaaf6397. doi:10.1126/scitranslmed.aaf6397
Delmastro, M., and Zamariola, G. (2020). Depressive symptoms in response to COVID-19 and lockdown: a cross-sectional study on the Italian population. Sci. Rep. 10 (1), 22457–22510. doi:10.1038/s41598-020-79850-6
Du, P., Zhang, X., Huang, C.-C., Jafari, N., Kibbe, W. A., Hou, L., et al. (2010). Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinforma. 11 (1), 587. doi:10.1186/1471-2105-11-587
Duggal, P., Gillanders, E. M., Holmes, T. N., and Bailey-Wilson, J. E. (2008). Establishing an adjusted p-value threshold to control the family-wide type 1 error in genome wide association studies. BMC genomics 9 (1), 516–518. doi:10.1186/1471-2164-9-516
Elsworth, B., Lyon, M., Alexander, T., Liu, Y., Matthews, P., Hallett, J., et al. (2020). The MRC IEU OpenGWAS data infrastructure. BioRxiv Available at: https://www.biorxiv.org/content/10.1101/2020.08.10.244293v1.
Eyre, O., Bevan Jones, R., Agha, S. S., Wootton, R. E., Thapar, A. K., Stergiakouli, E., et al. (2021). Validation of the short mood and feelings questionnaire in young adulthood. J. Affect. Disord. 294, 883–888. doi:10.1016/j.jad.2021.07.090
Fernstrom, J. D. (2005). Branched-chain amino acids and brain function. J. Nutr. 135 (6), 1539S–1546S. doi:10.1093/jn/135.6.1539S
Fortin, J.-P., Labbe, A., Lemire, M., Zanke, B. W., Hudson, T. J., Fertig, E. J., et al. (2014). Functional normalization of 450k methylation array data improves replication in large cancer studies. Genome Biol. 15 (11), 503. doi:10.1186/s13059-014-0503-2
Fraser, A., Macdonald-Wallis, C., Tilling, K., Boyd, A., Golding, J., Davey Smith, G., et al. (2013). Cohort profile: the avon longitudinal study of Parents and children: ALSPAC mothers cohort. Int. J. Epidemiol. 42 (1), 97–110. doi:10.1093/ije/dys066
Friedman, J., Hastie, T., and Tibshirani, R. (2010). Regularization paths for generalized linear models via coordinate descent. J. Stat. Softw. 33 (1), 1–22. doi:10.18637/jss.v033.i01
Gharipour, M., Barekatain, M., Sung, J., Emami, N., Sadeghian, L., Dianatkhah, M., et al. (2020). The epigenetic overlap between obesity and mood disorders: a systematic review. Int. J. Mol. Sci. 21 (18), 6758. doi:10.3390/ijms21186758
Gini, G., and Pozzoli, T. (2009). Association between bullying and psychosomatic problems: a meta-analysis. Pediatrics 123 (3), 1059–1065. doi:10.1542/peds.2008-1215
Gruenbaum, S. E., Chen, E. C., Sandhu, M. R. S., Deshpande, K., Dhaher, R., Hersey, D., et al. (2019). Branched-chain amino acids and seizures: a systematic review of the literature. CNS drugs 33 (8), 755–770. doi:10.1007/s40263-019-00650-2
Gu, G., Caldwell, G. A., and Chalfie, M. (1996). Genetic interactions affecting touch sensitivity in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 93 (13), 6577–6582. doi:10.1073/pnas.93.13.6577
Handakas, E., Chang, K., Khandpur, N., Vamos, E. P., Millett, C., Sassi, F., et al. (2022). Metabolic profiles of ultra-processed food consumption and their role in obesity risk in British children. Available at SSRN 4000887.
Handakas, E., Lau, C. H., Alfano, R., Chatzi, V. L., Plusquin, M., Vineis, P., et al. (2021). A systematic review of metabolomic studies of childhood obesity: state of the evidence for metabolic determinants and consequences. Obes. Rev. 23, e13384. doi:10.1111/obr.13384
Hayes, A. F., and Rockwood, N. J. (2017). Regression-based statistical mediation and moderation analysis in clinical research: observations, recommendations, and implementation. Behav. Res. Ther. 98, 39–57. doi:10.1016/j.brat.2016.11.001
Hemani, G., Zheng, J., Elsworth, B., Wade, K. H., Haberland, V., Baird, D., et al. (2018). The MR-Base platform supports systematic causal inference across the human phenome. elife 7, e34408. doi:10.7554/eLife.34408
Hernán, M. A. (2021). Causal analyses of existing databases: no power calculations required. J. Clin. Epidemiol. 144, 203–205. doi:10.1016/j.jclinepi.2021.08.028
Houseman, E. A., Accomando, W. P., Koestler, D. C., Christensen, B. C., Marsit, C. J., Nelson, H. H., et al. (2012). DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinforma. 13 (1), 86. doi:10.1186/1471-2105-13-86
Kelly, J. R., Borre, Y., O'Brien, C., Patterson, E., El Aidy, S., Deane, J., et al. (2016). Transferring the blues: depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 82, 109–118. doi:10.1016/j.jpsychires.2016.07.019
Lamb, M. M., Ogden, C. L., Carroll, M. D., Lacher, D. A., and Flegal, K. M. (2011). Association of body fat percentage with lipid concentrations in children and adolescents: united States, 1999–2004. Am. J. Clin. Nutr. 94 (3), 877–883. doi:10.3945/ajcn.111.015776
Lampinen, E.-K., Eloranta, A.-M., Haapala, E. A., Lindi, V., Väistö, J., Lintu, N., et al. (2017). Physical activity, sedentary behaviour, and socioeconomic status among finnish girls and boys aged 6–8 years. Eur. J. Sport Sci. 17 (4), 462–472. doi:10.1080/17461391.2017.1294619
Lawlor, D. A., Tilling, K., and Davey Smith, G. (2016). Triangulation in aetiological epidemiology. Int. J. Epidemiol. 45 (6), 1866–1886. doi:10.1093/ije/dyw314
Lester, B. M., Conradt, E., and Marsit, C. J. (2013). Epigenetic basis for the development of depression in children. Clin. Obstet. Gynecol. 56 (3), 556–565. doi:10.1097/GRF.0b013e318299d2a8
Ligezka, A. N., Sonmez, A. I., Corral-Frias, M. P., Golebiowski, R., Lynch, B., Croarkin, P. E., et al. (2021). A systematic review of microbiome changes and impact of probiotic supplementation in children and adolescents with neuropsychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 108, 110187. doi:10.1016/j.pnpbp.2020.110187
Marazziti, D., Rutigliano, G., Baroni, S., Landi, P., and Dell'Osso, L. (2014). Metabolic syndrome and major depression. CNS Spectr. 19 (4), 293–304. doi:10.1017/S1092852913000667
Mazidi, M., Karimi, E., Rezaie, P., and Ferns, G. A. (2017). Effects of conjugated linoleic acid supplementation on serum C-reactive protein: a systematic review and meta‐analysis of randomized controlled trials. Cardiovasc. Ther. 35 (6), e12275. doi:10.1111/1755-5922.12275
Milaneschi, Y., Simmons, W. K., van Rossum, E. F., and Penninx, B. W. (2019). Depression and obesity: evidence of shared biological mechanisms. Mol. Psychiatry 24 (1), 18–33. doi:10.1038/s41380-018-0017-5
Min, J. L., Hemani, G., Davey Smith, G., Relton, C., and Suderman, M. (2018). Meffil: efficient normalization and analysis of very large DNA methylation datasets. Bioinformatics 34 (23), 3983–3989. doi:10.1093/bioinformatics/bty476
Moore, S. E., Norman, R. E., Suetani, S., Thomas, H. J., Sly, P. D., and Scott, J. G. (2017). Consequences of bullying victimization in childhood and adolescence: a systematic review and meta-analysis. World J. Psychiatry 7 (1), 60–76. doi:10.5498/wjp.v7.i1.60
Orešič, M., Tang, J., Seppänen-Laakso, T., Mattila, I., Saarni, S. E., Saarni, S. I., et al. (2011). Metabolome in schizophrenia and other psychotic disorders: a general population-based study. Genome Med. 3 (3), 19–14. doi:10.1186/gm233
Pascal, V., Pozuelo, M., Borruel, N., Casellas, F., Campos, D., Santiago, A., et al. (2017). A microbial signature for Crohn's disease. Gut 66 (5), 813–822. doi:10.1136/gutjnl-2016-313235
Persons, J. E., and Fiedorowicz, J. G. (2016). Depression and serum low-density lipoprotein: a systematic review and meta-analysis. J. Affect. Disord. 206, 55–67. doi:10.1016/j.jad.2016.07.033
Relton, C. L., Gaunt, T., McArdle, W., Ho, K., Duggirala, A., Shihab, H., et al. (2015a). Data resource profile: accessible resource for integrated epigenomic studies (ARIES). Int. J. Epidemiol. 44 (4), 1181–1190. doi:10.1093/ije/dyv072
Relton, C. L., Gaunt, T., McArdle, W., Ho, K., Duggirala, A., Shihab, H., et al. (2015b). Data resource profile: accessible resource for integrated epigenomic studies (ARIES). Int. J. Epidemiol. 44, 1181–1190. doi:10.1093/ije/dyv072
Rhew, I. C., Simpson, K., Tracy, M., Lymp, J., McCauley, E., Tsuang, D., et al. (2010). Criterion validity of the short mood and feelings questionnaire and one-and two-item depression screens in young adolescents. Child. Adolesc. Psychiatry Ment. Health 4 (1), 8–11. doi:10.1186/1753-2000-4-8
Rijnhart, J. J., Twisk, J. W., Eekhout, I., and Heymans, M. W. (2019). Comparison of logistic-regression based methods for simple mediation analysis with a dichotomous outcome variable. BMC Med. Res. Methodol. 19 (1), 19–10. doi:10.1186/s12874-018-0654-z
Robinson, O., Chadeau Hyam, M., Karaman, I., Climaco Pinto, R., Ala-Korpela, M., Handakas, E., et al. (2020). Determinants of accelerated metabolomic and epigenetic aging in a UK cohort. Aging Cell. 19 (6), e13149. doi:10.1111/acel.13149
Segal, A. B., Huerta, M. C., Aurino, E., and Sassi, F. (2021). The impact of childhood obesity on human capital in high‐income countries: a systematic review. Obes. Rev. 22 (1), e13104. doi:10.1111/obr.13104
Soininen, P., Kangas, A. J., Würtz, P., Tukiainen, T., Tynkkynen, T., Laatikainen, R., et al. (2009). High-throughput serum NMR metabonomics for cost-effective holistic studies on systemic metabolism. Analyst 134 (9), 1781–1785. doi:10.1039/b910205a
Somineni, H. K., Venkateswaran, S., Kilaru, V., Marigorta, U. M., Mo, A., Okou, D. T., et al. (2019). Blood-derived DNA methylation signatures of Crohn's disease and severity of intestinal inflammation. Gastroenterology 156 (8), 22542254–22542265.e3. doi:10.1053/j.gastro.2019.01.270
Steen, J., Loeys, T., Moerkerke, B., and Vansteelandt, S. (2017). Medflex: an R package for flexible mediation analysis using natural effect models. J. Stat. Softw. 76 (11). doi:10.18637/jss.v076.i11
Stekhoven, D. J., and Bühlmann, P. (2012). MissForest—non-parametric missing value imputation for mixed-type data. Bioinformatics 28 (1), 112–118. doi:10.1093/bioinformatics/btr597
Strandwitz, P. (2018). Neurotransmitter modulation by the gut microbiota. Brain Res. 1693, 128–133. doi:10.1016/j.brainres.2018.03.015
Sun, Y., Hu, Z., Goeb, Y., and Dreier, L. (2013). The F-box protein MEC-15 (FBXW9) promotes synaptic transmission in GABAergic motor neurons in C. elegans. PloS one 8 (3), e59132. doi:10.1371/journal.pone.0059132
Tateishi, K., Okada, Y., Kallin, E. M., and Zhang, Y. (2009). Role of Jhdm2a in regulating metabolic gene expression and obesity resistance. Nature 458 (7239), 757–761. doi:10.1038/nature07777
Thapar, A., and McGuffin, P. (1998). Validity of the shortened mood and Feelings questionnaire in a community sample of children and adolescents: a preliminary research note. Psychiatry Res. 81 (2), 259–268. doi:10.1016/s0165-1781(98)00073-0
Turner, N., Joinson, C., Peters, T. J., Wiles, N., and Lewis, G. (2014). Validity of the short mood and Feelings questionnaire in late adolescence. Psychol. Assess. 26 (3), 752–762. doi:10.1037/a0036572
Van Vliet, K., van Ginkel, W. G., Jahja, R., Daly, A., MacDonald, A., De Laet, C., et al. (2019). Emotional and behavioral problems, quality of life and metabolic control in NTBC-treated Tyrosinemia type 1 patients. Orphanet J. Rare Dis. 14 (1), 285–289. doi:10.1186/s13023-019-1259-2
Vansteelandt, S., Bekaert, M., and Lange, T. (2012). Imputation strategies for the estimation of natural direct and indirect effects. Epidemiol. Methods 1 (1), 131–158. doi:10.1515/2161-962x.1014
Vehmeijer, F. O., Küpers, L. K., Sharp, G. C., Salas, L. A., Lent, S., Jima, D. D., et al. (2020). DNA methylation and body mass index from birth to adolescence: meta-analyses of epigenome-wide association studies. Genome Med. 12 (1), 105–115. doi:10.1186/s13073-020-00810-w
Wahl, S., Drong, A., Lehne, B., Loh, M., Scott, W. R., Kunze, S., et al. (2017). Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 541 (7635), 81–86. doi:10.1038/nature20784
Wei, R., Wang, J., Su, M., Jia, E., Chen, S., Chen, T., et al. (2018). Missing value imputation approach for mass spectrometry-based metabolomics data. Sci. Rep. 8 (1), 1–10. doi:10.1038/s41598-017-19120-0
Williams, J. E., Helsel, B., Griffin, S. F., and Liang, J. (2017). Associations between parental BMI and the family nutrition and physical activity environment in a community sample. J. Community Health 42 (6), 1233–1239. doi:10.1007/s10900-017-0375-y
Keywords: child, obesity, depression, multiomics, ALSPAC
Citation: Handakas E, Xu Y, Segal AB, Huerta MC, Bowman K, Howe LD, Sassi F and Robinson O (2022) Molecular mediators of the association between child obesity and mental health. Front. Genet. 13:947591. doi: 10.3389/fgene.2022.947591
Received: 18 May 2022; Accepted: 27 July 2022;
Published: 31 August 2022.
Edited by:
Michael E. Symonds, University of Nottingham, United KingdomReviewed by:
Liliana Burlibasa, University of Bucharest, RomaniaCopyright © 2022 Handakas, Xu, Segal, Huerta, Bowman, Howe, Sassi and Robinson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oliver Robinson, by5yb2JpbnNvbkBpbXBlcmlhbC5hYy51aw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.