 Liming Zhang
Liming Zhang Zijian Zhu
Zijian Zhu Qing Yang
Qing Yang Jingjing Zhao
Jingjing Zhao
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 13 September 2022
Sec. Epigenomics and Epigenetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.915275
This article is part of the Research TopicGenetics and Epigenetics of Psychiatric Diseases - Volume IIView all 13 articles
Spatial working memory (SWM) is a kind of memory that temporarily preserves spatial information (the location or order of objects, etc.). Individuals with mental disorders tend to show worse performance in SWM task. This study investigated the genetic basis of two subtypes of SWM, static spatial working memory (SSWM) and dynamic spatial working memory (DSWM) in humans, using quantitative genomic analyses. A total of 451 Chinese students were tested on their magnitudes of SSWM and DSWM. A genome-wide association study (GWAS) was performed. Two SNPs (top SNP: rs80263879, p = 1.6 × 10−9, gene: epoxide hydrolase 2, EPHX2) reaching genome-wide significance for SSWM were identified. There is a high linkage disequilibrium between these two SNPs. The data of expression quantitative trait locus (eQTL) showed that different genotypes of rs80263879 and rs72478903 made significant differences in the expression of EPHX2 gene in the spinal cord (p = 0.022, p = 0.048). Enrichment analysis identified a gene set significantly associated with DSWM. Overall, our study discovered a candidate genetic locus and gene set for the genetics of the SWM.
Spatial working memory (SWM) is an important component of working memory, it can produce, operate and maintain visual images, related to spatial position, motion, etc. SWM can be divided into static spatial working memory (SSWM) and dynamic spatial working memory (DSWM) (Pickering, Gathercole, Hall, & Lloyd, 2001; Cocchi et al., 2007). In the SSWM task, spatial information is usually processed once; in the DSWM task, spatial information has to be processed continuously. SWM reflects the ability to temporarily store spatial information (Goodale & Milner, 1992), varing widely across individuals. In particular, individuals with mental disorders, such as autism spectrum disorder (ASD) or schizophrenia, have varying degrees of impairment in SWM (Jiang et al., 2015; Minor and Park, 1999). The reason may be that the development process of SWM in patients with mental diseases from their 10th to 20th years of age has stagnated (Song et al., 2013).
What are the factors causing the individual differences of SWM ability? First, there is a research evidence that environmental factors (educational level, age, etc.) influence SWM ability, these environmental factors were significantly correlated with SWM ability in healthy subjects, but not in schizophrenic subjects (Stratta et al., 2001). Second, studies revealed that genetic factors influenced SWM ability in schizophrenic patients. For instance, researchers divided schizophrenic patients into different categories according to different genotypes of a specific gene (catechol-O-methyltransferase) (Miskowiak et al., 2017). Different groups of subjects performed differently in SWM task. However, so far, there is no genetic research on SWM in the general population and at the genome-wide level.
In this study, we did a GWAS among high school students and college students to reveal the molecular mechanism of SWM. We analyzed two SWM subtypes (i.e., SSWM and DSWM). We investigated the function of loci significantly associated with the phenotype. We also performed enrichment analysis to see if phenotypes were affected by the joint effects of multiple loci or genes. Our results showed that SWM has a specific genetic mechanism.
Cohorts consisted of college students recruited from the Shaanxi Normal University and Xi’an Jiaotong University in Northwest of China, and senior high school students recruited from Sichuan LuXian No.2 High School in Southwest of China and Xi'an No. 1 middle school in Northwest of China. There were 544 participants in total, with 100% of Han population. The average age is 17 years old (standard deviation, SD = 1.32), with 54% of female. Each participant had no history of mental illness, and none of the subjects reported psychiatric or visual illness. All subjects had normal visual acuity or corrected visual acuity.
The experimental procedure was approved by the evaluation committee of Shaanxi Normal University. The subjects were clear about the purpose of the experiment. They signed informed consent before the experiment.
Stimuli. Three stimuli of SWM were created, Figures 1A,C for the SSWM task, Figures 1B,C for the DSWM task. Each SWM stimulus was composed of a rectangular box and a dot, and was presented on a white backgroud. Figures 1A,B are the learning stage stimuli, Figure 1C is the test stage stimuli.
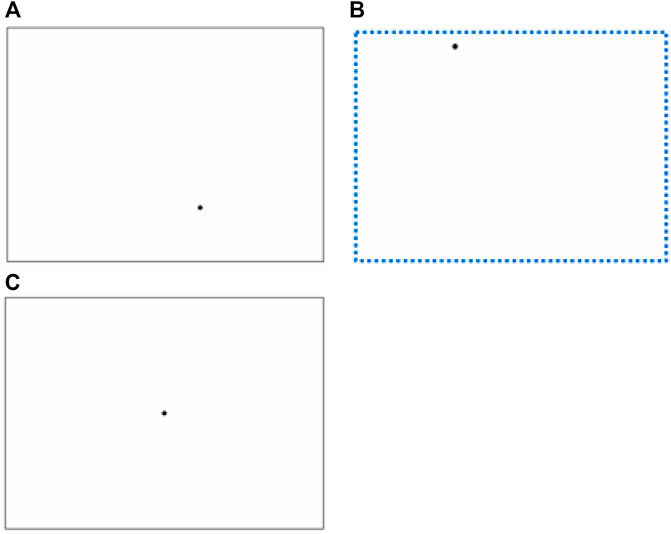
FIGURE 1. Stimuli in experiment. The above three figures reveal ‘Stimuli in the learning stage of SSWM task’ (A), ‘Stimuli in the learning stage of DSWM task’ (B) (There are no dashed tracks in the formal experiment), ‘Stimuli in the test stage of SSWM and DSWM task’ (C).
The stimuli were generated and presented by MATLAB and its toolbox (PsychToolbox). The size of the LCD screen of the stimulating computer is 23.8 inches and the refresh rate is 60 Hz. The viewing distance was kept at 40 cm using a chin rest. Luminance for all participants remained unchanged.
Procedure. SSWM task: The experiment is based on Gordon’s spatial positioning test (Gordon, 1986). Specific procedure was as follows. At first, a rectangular box with a certain size was randomly presented at different positions on the computer screen, and at the same time, a dot with a certain size was randomly presented at different position of the rectangular. After a few seconds, these stimulus disappeared automatically, as shown in Figure 1A; then a rectangular box and a dot at its center with the same size appeared on the screen, as shown in the Figure 1C. The participants’ task was to move the dot in the center of the rectangular box to the position in the rectangular box where the dot just disappeared through the direction key on the keyboard. The position of the rectangular box on the screen is different in the learning stage and the testing stage, so as to avoid the subjects completing the task by remembering the coordinate position of the point in the whole screen.
The specific operation of DSWM task is that a dot randomly appeared in the upper left area of the screen and completed the following movements in sequence at a fixed speed: a centimeters from left to right → b centimeters from top to bottom → a centimeters from right to left → b centimeters from bottom to top. This dot moved to its original place and then disappeared. The motion trajectory of dot finally formed a rectangle box with the same size as that in the SSWM task. After the dot disappeared, another dot appeared in the disappeared motion track (Figure 1B). After the latter dot disappeared, participants completed the same task as the SSWM experiment.
This task has a total of 50 attempts and takes a total of 40 min. The measurement index of this task is the distance deviation between the position of the last dot and the position of the initial dot, that is, how many screen pixels are apart.
DNA was extracted from saliva samples of 520 participants, and individuals were genotyped using Illumina Asian screening array (650K) by Beijing Compass Biotechnology (N1 = 288) and Genergy Biotechnology (N2 = 232). The quality control of two samples from different companies is exactly the same. Briefly, SNPs were filtered out if they showed a variant call rate <0.9, a minor allele frequency (MAF) < 0.05, a hardy-weinberg equilibrium (HWE) < 10−5. Individuals were filtered out if they showed a missing genotype data (mind) < 0.10 (3 people from sample1 were removed, 1 people from sample2 were removed), unexpected duplicates or probable relatives (PI-HAT>0.20) (Anderson et al., 2010; Chang et al., 2015).
For imputation, autosomal variants were aligned to the 1000G genomes phase 1v3 reference panel. Imputation was performed using Michigan imputation Server 4.0 in 5 Mb chunks with 500 kb buffers (Das et al., 2016), filtering out variants that were monomorphic in the Genome Asia Pilot (GAsP). Chunks with 51% genotyped variants or concordance rate <0.92 were fused with neighboring chunks and re-imputed. Imputed variants were filtered out for rsq <0.60, MAF<0.05, mind<0.1, HWE<10−5 using Plink (v1.90).
After imputation, we merged two gene samples, and the merged sample left 516 individuals and 4 196 499 SNPs.
After completing the above operations, 451 individuals have genetic data, phenotypic data and covariate data at the same time. Genome-wide association analyses were performed using Plink 1.90 (Rentería et al., 2013), fitting an additive model to the linear regression model with adjustment for sex, grade, and the first two principle components of population structure (Chang et al., 2015). Manhattan plots and Quantile-quantile plots were generated using the ggplot2 package in R. We used locuszoom to generate regional association plots (Pruim et al., 2010). The sign of genome-wide significance is p < 5 × 10−8.
In order to examine whether the discovered rs80263879 and rs72478903 influnce gene expression, we checked the expression quantitative trait locus (eQTL) for these two loci in the Genotype-Tissue Expression (GTEx) Cohort (https://www.gtexportal.org/home/) (GTEx Consortium, 2020). Analysis of eQTL can be with reference to the study of Ramasamy et al. (Ramasamy et al., 2014).
Gene-based and pathway-based enrichment tests for SSWM and DSWM were conducted by MAGMA (de Leeuw et al., 2015). Gene-based analyses is based on GWAS data. Every SNP is located on the protein-coding gene according to NCBI37.3. Genes were analyzed after internal quality control. We adopted default internal quality control steps and internal quality control values of magma. A total of 17,225 genes were involved in the analysis, so the threshold of significance was set at 2.90 × 10−6 (p = 0.05/17,225).
Pathway-based analyses are derived from the results of gene-based analyses using a competitive gene-set analysis (Gerring et al., 2019). Orginal pathway were from Molecular Signatures Database website (MSigDB, c2.all.v7.0.entrez). A total of 5 497 pathways were involved in the analysis, so the threshold of significance was set at 9.10 × 10−6 (p = 0.05/5 497).
After phenotypic data were collected, we used P-P plot to draw the consistency of the cummulative proportion of SSWM (Figure 2A) and DSWM (Figure 2B) with the cumulative proportion of normal distribution. P-P plot shows that the distributions of the two phenotypes are a little skewed. Then, we did correlation analysis between phenotypes and sociodemographic variables (gender and grade) (Table 1). Results showed that there were significant correlations between SSWM task and sociodemographic variables.
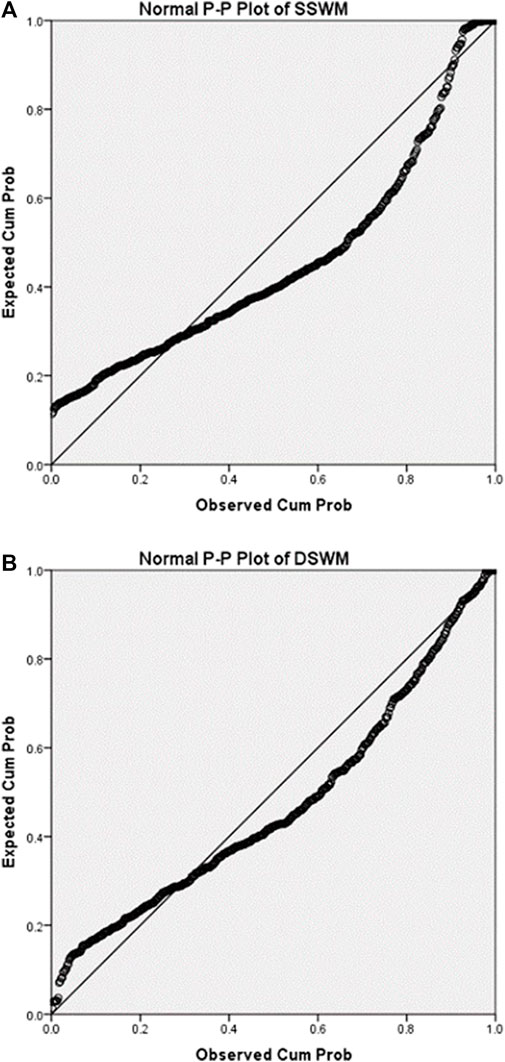
FIGURE 2. P-P plots for phenotypes. The above two figures summarise ‘consistency between the cumulative proportion of data of DSWM and the cumulative proportion of normal distribution’ (A), ‘consistency between the cumulative proportion of data of SSWM and the cumulative proportion of normal distribution’ (B). The closer the scatter distribution is to the diagonal, the more the data conforms to the normal distribution.

TABLE 1. Correlations for sociodemographic variables (gender and grade) and phenotypes (SSWM and DSWM).
We identified two genome-wide significant loci associated with SSWM (rs80263879, p = 1.6 × 10−9; rs72478903, p = 3.5 × 10−8). These two loci have strong linkage disequilibrium. And these two SNPs are all located within the gene EPHX2. We have not identified any SNP significantly associated with DSWM. However, rs80263879 revealed weak association with DSWM (p = 4.5 × 10−2).
Manhattan plots were adopted to reveal the results of association analysis for SSWM (Figure 3) and DSWM (Figure 4). Due to the quality control of the two samples, the specific number of people at each locus is somewhat different. More details for significant locus were reported in Table 2. We used quantile-quantile plots for SSWM (Figure 5A) and DSWM (Figure 5B) to reveal the rationality of their respective analysis models. The genomic inflation factors λ of quantile-quantile plots was 1.023 for SSWM and 1.013 for DSWM, confirming little influence of population stratification. Regional association plot of the significant locus are revealed in Figure 6.
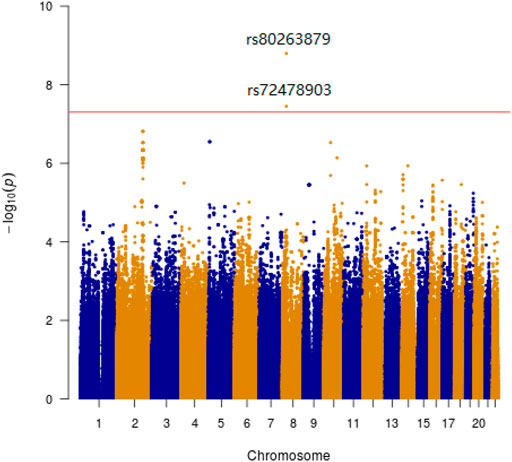
FIGURE 3. Manhattan plot showing genome-wide association analysis for SSWM. X-axis represents the position of SNPs on the chromosome, y-axis represents the significance level of association analysis. The horizontal red line represents the genome-wide significance level threshold (5 × 10−8).
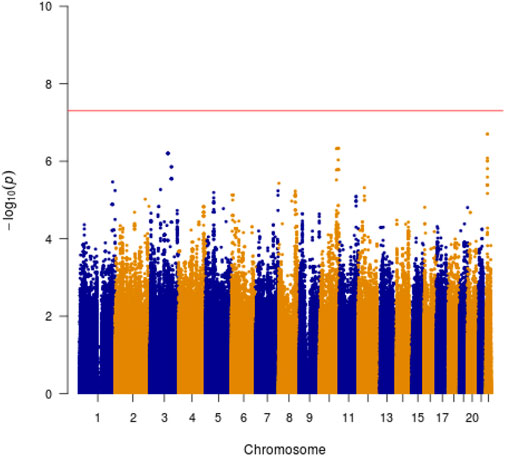
FIGURE 4. Manhattan plot showing genome-wide association analysis for DSWM.
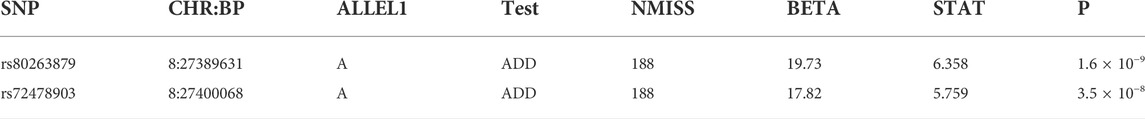
TABLE 2. Two lead single-variant associations detected in the GWAS analyses.
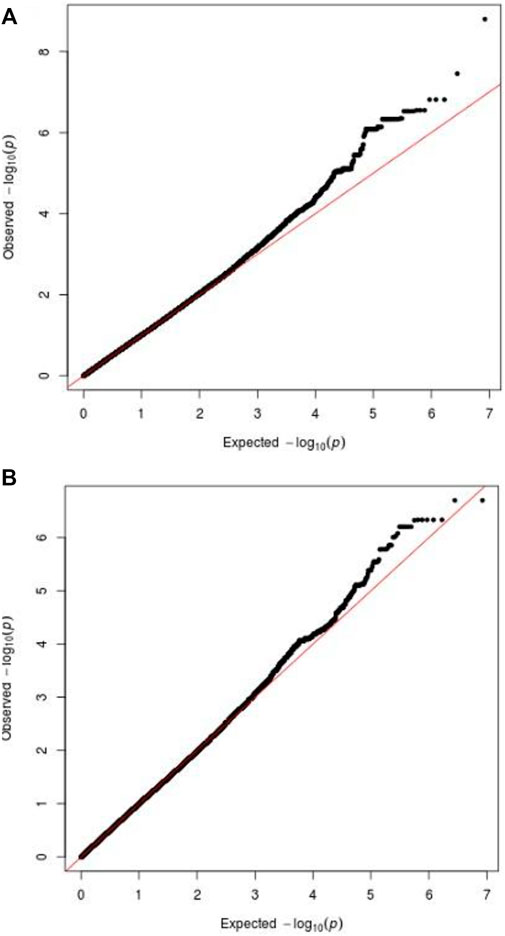
FIGURE 5. Q-Q plot for phenotypes. The above two figures show ‘Q-Q plot for SSWM’ (A), ‘Q-Q plot for DSWM’ (B).
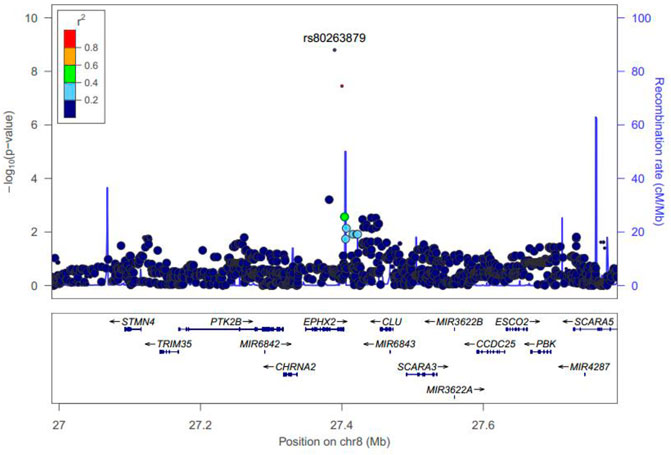
FIGURE 6. Regional plot of rs80263879 for SSWM. The most significant locus was marked violet. r2 is the LD value of rs80263879 and any other locus.
Two genome-wide significant loci we identified are located in gene EPHX2. According to the data of eQTL from GTEx, both loci have a significant regulatory effect on the expression of EPHX2 in the spinal cord (cervical c-1) (rs80263879, p = 2.2 × 10−2; rs72478903, p = 4.8 × 10−2). Figure 7A reveals that the major allele G of rs80263879 is associated with increased EPHX2 expression in spinal cord (cervical c-1). Results of eQTL in other 12 brain regions of rs80263879 are revealed in Supplementary Figure S1. Similarly, Figure 7B reveals that the major allele G of rs72478903 is associated with increased EPHX2 expression in spinal cord (cervical c-1). Results of eQTL in other 12 brain regions of rs72478903 are revealed in Supplementary Figure S2.
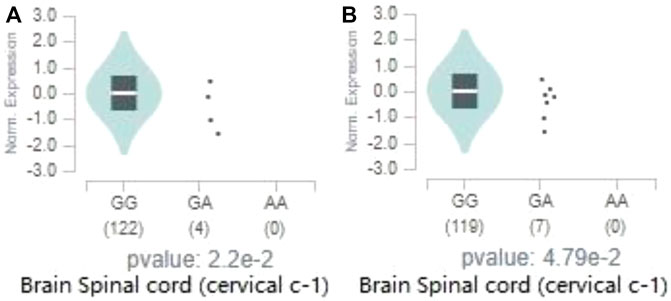
FIGURE 7. Effect of significant loci on the expression of EPHX2. The above two figures reveal ‘expression amount of EPHX2 on different genotypes of rs80263879 in spinal cord (cervical c-1)’(A), ‘expression amount of EPHX2 on different genotypes of rs72478903 in spinal cord (cervical c-1)’(B) from Genotype-Tissue Expression (GTEx) Cohort.
In the pathway-based enrichment analysis, we found a pathway (Standard name: GRAHAM_CML_QUIESCENT_VS._NORMAL_DIVIDING_UP) significantly enriched with DSWM after Bonferroni correction (β = 0.51, se = 0.11, p = 1.36 × 10−6 < 9.10×10−6). Genes on the pathway in our data are shown in Supplementary Table S1. Genes of the pathway were found to be up-regulated in certain leukemia-derived quescent hemopoietic stem cells expressing CD34 in contrast with dividing cells supplied by the normal population (Graham, Vass, Holyoake, & Graham, 2010). Bubble plot of the 20 pathways with the lowest p values was shown in Figure 8, and full results for pathway-based enrichment analysis were shown in Supplementary Table S2.
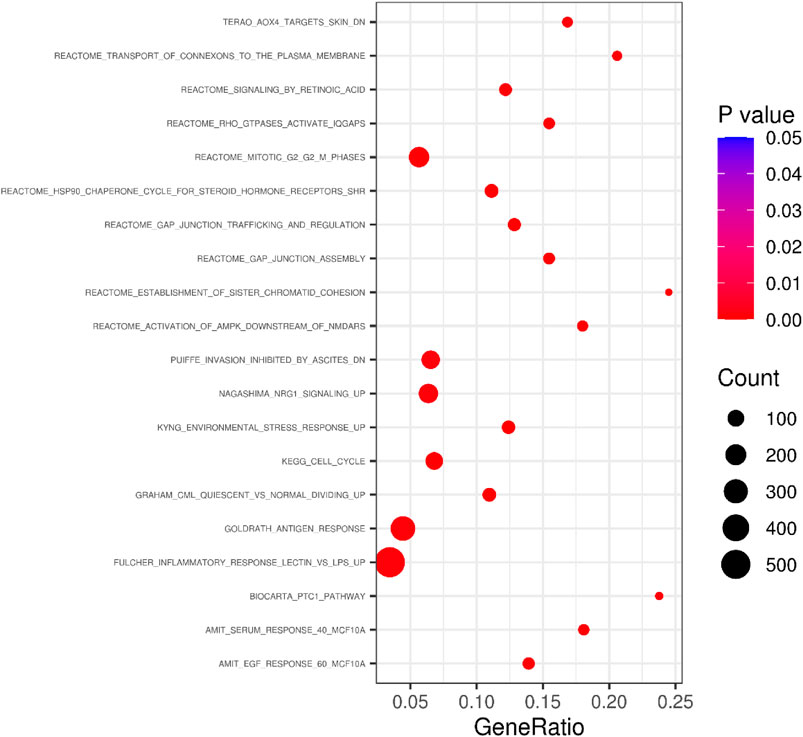
FIGURE 8. Bubble plot summarising the results of the pathway-based enrichment tests for DSWM. The plot reveals 20 pathways with the lowest p values.
This study is the first genome-wide association study on spatial working memory in the general population. We reported two genome-wide significant loci (rs80263879 and rs72478903) associated with static spatial working memory. These two loci have strong linkage disequilibrium. We also found that rs80263879 and rs72478903 affect the expression of EPHX2 in the spine. In addition, we identified a gene set significantly associated with dynamic spatial working memory. These results provide evidence for the genetic basis of spatial working memory.
Spatial working memory is an important ability in people's daily life, in the field of aerospace for predicting flight performance (Tirre & Raoufl 1998), and in representing an endophenotype of mental disorders such as schizophrenia. Spatial working memory has also been suggested to be influenced by genetic factors. Indeed, researchers have conducted genome-wide association analysis in schizophrenic patients and controls, and identified some loci associated with spatial working memory (Ren et al., 2015). However, so far, no genome-wide association analysis of spatial working memory has been conducted in general population. Here, our study report the first-hand evidence for the genetic basis of spatial working memory in a population of Chinese young adults.
We identified two genome-wide significant SNPs with strong LD among each other associated with static spatial working memory. These two loci are located within the gene EPHX2. EPHX2 is associated with familial hypercholesterolemia (Pillai, Shah, Reddy, Ashavaid, & Vishwanathan, 2022). This was the first time that it had been found to be related to spatial working memory. Spatial working memory is usually impaired among mental disorders such as schizophrenia and ASD (Jiang et al., 2015). Expression of EPHX2 mRNA from schizophrenia and ASD have been found higher than that of controls (Ma et al., 2019). Therefore, we speculate that EPHX2 may play its role in mental disorders through spatial working memory. However, this speculation requires further empirical evidence for validation.
Different genotypes of rs80263879 and rs72478903 make gene EPHX2 express in different degrees in the spinal cord, indicating that the spinal cord may be associated with SWM. Research on mice revealed that spinal cord injury can significantly block the expression of dopamine receptors in the frontal lobe, thus damaging SWM (Kheyrkhah et al., 2020). However, effects of these loci on regulating the expression of the gene EPHX2 are weak, and number of subjects with “GA” genotype is small. Therefore, more samples are needed to verify this regulatory effect in the future.
A gene set significantly associated with dynamic spatial working memory was identified in enrichment analysis. The expression of genes in this gene set is positively regulated in quiescent CD34 + cells from some leukemia. CD34 + cells can differentiate into endothelial progenitor cells, which can be conducive to the repair of patients’ myocardium (Kim et al., 2016).
In conclusion, we identified two loci significantly associated with static spatial working memory, which are located on the gene EPHX2 and regulate the expression of this gene. We also identified a gene set associated with dynamic spatial working memory. Our research deepens the understanding of the genetic basis of spatial working memory and can provide reference for the treatment of mental diseases to some extent.
The original contributions presented in the study are publicly available. This data can be found in the GWA catalog http://ftp.ebi.ac.uk/pub/databases/gwas/summary_statistics/ GCST90133001-GCST90134000, GCST90133162, and GCST90133163.
The studies involving human participants were reviewed and approved by the evaluation committee of Shaanxi Normal University. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
LZ is responsible for the main work, ZZ guides the experimental design, QY assists data analysis and JZ gives comprehensive guidance.
The study was funded by National Natural Science Foundation of China (61807023), Humanities and Social Science Fund of Ministry of Education of the People’s Republic of China (17XJC190010), Shaanxi Province Natural Science Foundation (2018JQ8015), and Fundamental Research Funds for the Central Universities (CN) (GK201702011) to JZ. This study was also supported by internal funds from Shaanxi Normal University to JZ and Chunyu Liu.
The authors would like to thank all students who participated in the study and their parents and teachers for their time and cooperation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.915275/full#supplementary-material
Anderson, C. A., Pettersson, F. H., Clarke, G. M., Cardon, L. R., Morris, A. P., and Zondervan, K. T. (2010). Data quality control in genetic case-control association studies. Nat. Protoc. 5, 1564–1573. doi:10.1038/nprot.2010.116
Chang, C. C., Chow, C. C., Tellier, L. C., Vattikuti, S., Purcell, S. M., and Lee, J. J. (2015). Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaSci 4, 7. doi:10.1186/s13742-015-0047-8
Cocchi, L., Schenk, F., Volken, H., Bovet, P., Parnas, J., and Vianin, P. (2007). Visuo-spatial processing in a dynamic and a static working memory paradigm in schizophrenia. Psychiatry Res. 152, 129–142. doi:10.1016/j.psychres.2006.02.004
Das, S., Forer, L., Schönherr, S., Sidore, C., Locke, A. E., Kwong, A., et al. (2016). Next-generation genotype imputation service and methods. Nat. Genet. 48, 1284–1287. doi:10.1038/ng.3656
de Leeuw, C. A., Mooij, J. M., Heskes, T., and Posthuma, D. (2015). Magma: Generalized gene-set analysis of GWAS data. PLoS Comput. Biol. 11 (4), e1004219. doi:10.1371/journal.pcbi.1004219
Gerring, Z. F., Mina-Vargas, A., and Derks, E. M. (2019). eMAGMA: An eQTL-informed method to identify risk genes using genome-wide association study summary statistics. BioRxiv. 2019, 854315. doi:10.1101/854315
Goodale, M. A., and Milner, A. D. (1992). Separate visual pathways for perception and action. Trends Neurosci. 15 (1), 20–25. doi:10.1016/0166-2236(92)90344-8
Gordon, H. W. (1986). The cognitive laterality battery: Tests of specialized cognitive function. Int. J. Neurosci. 29, 223–244. doi:10.3109/00207458608986153
Graham, S. M., Vass, J. K., Holyoake, T. L., and Graham, G. J. (2010). Transcriptional analysis of quiescent and proliferating cd34+ human hemopoietic cells from normal and chronic myeloid leukemia sources. Stem Cells 25 (12), 3111–3120. doi:10.1634/stemcells.2007-0250
GTEx Consortium (2020). The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 369 (6509), 1318–1330. doi:10.1126/science.aaz1776
Jiang, Y. V., Capistrano, C. G., and Palm, B. E. (2015). Spatial working memory in children with high-functioning autism: Intact configural processing but impaired capacity. J. Abnorm. Psychol. 123 (1), 248.
Kheyrkhah, H., Soltani Zangbar, H., Salimi, O., Shahabi, P., and Alaei, H. (2020). Prefrontal dopaminergic system and its role in working memory and cognition in spinal cord-injured rats. Exp. Physiol. 105 (21), 1579–1587. doi:10.1113/EP088537
Kim, S.-W., Jin, H. L., Kang, S.-M., Kim, S., Yoo, K.-J., Jang, Y., et al. (2016). Therapeutic effects of late outgrowth endothelial progenitor cells or mesenchymal stem cells derived from human umbilical cord blood on infarct repair. Int. J. Cardiol. 203 (1), 498–507. doi:10.1016/j.ijcard.2015.10.110
Ma, M., Ren, Q., Yang, J., Zhang, K., Xiong, Z., Ishima, T., et al. (2019). Key role of soluble epoxide hydrolase in the neurodevelopmental disorders of offspring after maternal immune activation. Proc. Natl. Acad. Sci. U. S. A. 116 (14), 7083–7088. doi:10.1073/pnas.1819234116
Minor, K., and Park, S. (1999). Spatial working memory: Absence of gender differences in schizophrenia patients and healthy control subjects. Biol. psychiatry 46 (7), 1003–1005. doi:10.1016/s0006-3223(99)00149-3
Miskowiak, K., Kjaerstad, H., Støttrup, M., Svendsen, A., Demant, K., Hoeffding, L., et al. (2017). The catechol-O-methyltransferase (COMT ) Val158Met genotype modulates working memory-related dorsolateral prefrontal response and performance in bipolar disorder. Bipolar Disord. 19 (3), 214–224. doi:10.1111/bdi.12497
Peyrot, W. J., and Price, A. L. (2021). Identifying loci with different allele frequencies among cases of eight psychiatric disorders using CC-GWAS. Nat. Genet. 53 (4), 445–454. doi:10.1038/s41588-021-00787-1
Pickering, S. J., Gathercole, S. E., Hall, M., and Lloyd, S. A. (2001). Development of memory for pattern and path: Further evidence for the fractionation of visuo-spatial memory. Q. J. Exp. Psychol. Sect. A 54, 397–420. doi:10.1080/713755973
Pillai, K. K. B., Shah, S. A., Reddy, L. L., Ashavaid, T. F., and Vishwanathan, S. (2022). Targeted exome sequencing in South Indian patients with familial hypercholesterolemia. Clin. Chim. Acta. 527, 47–55. doi:10.1016/j.cca.2021.12.022
Pruim, R. J., Welch, R. P., Sanna, S., Teslovich, T. M., Chines, P. S., Gliedt, T. P., et al. (2010). LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics 26 (18), 2336–2337. doi:10.1093/bioinformatics/btq419
Ramasamy, A., Trabzuni, D., Trabzuni, D., Guelfi, S., Varghese, V., Smith, C., et al. (2014). Genetic variability in the regulation of gene expression in ten regions of the human brain. Nat. Neurosci. 17, 1418–1428. doi:10.1038/nn.3801
Ren, H., Zhang, C., Huang, C., Li, N., Li, M., Li, Y., et al. (2015). Unravelling genes and pathways implicated in working memory of schizophrenia in Han Chinese. Ijms 16 (1), 2145–2161. doi:10.3390/ijms16012145
Rentería, M. E., Cortes, A., and Medland, S. E. (2013). “Using PLINK for genome-wide association studies (GWAS) and data analysis[M],” in Wide association studies and genomic prediction (Totowa, NJ: Humana Press), 193–213.
Song, W., Zhang, K., Sun, J., Ma, L., Jesse, F. F., Teng, X., et al. (2013). A simple spatial working memory and attention test on paired symbols shows developmental deficits in schizophrenia patients. Neural Plast. 2013, 130642. doi:10.1155/2013/130642
Stratta, P., Prosperini, P., Daneluzzo, E., Bustini, M., and Rossi, A. (2001). Educational level and age influence spatial working memory and Wisconsin card sorting test performance differently: A controlled study in schizophrenic patients. Psychiatry Res. 102 (1), 39–48. doi:10.1016/s0165-1781(01)00230-x
Keywords: GWAS, spatial working memory, single nucleotide polymorphisms (SNPs), gene, experiment
Citation: Zhang L, Zhu Z, Yang Q and Zhao J (2022) A genome-wide association study identified one variant associated with static spatial working memory in Chinese population. Front. Genet. 13:915275. doi: 10.3389/fgene.2022.915275
Received: 07 April 2022; Accepted: 27 June 2022;
Published: 13 September 2022.
Edited by:
Cunyou Zhao, Southern Medical University, ChinaReviewed by:
Ming Li, Kunming Institute of Zoology (CAS), ChinaCopyright © 2022 Zhang, Zhu, Yang and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jingjing Zhao, amluZ2ppbmcuemhhb0Bzbm51LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.