
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 24 March 2022
Sec. Immunogenetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.841879
This article is part of the Research Topic Progress in understanding the immunogenetic basis of disease susceptibility and outcomes View all 5 articles
 Zain Al Yafei1Steven J. Mack2Marion Alvares1
Zain Al Yafei1Steven J. Mack2Marion Alvares1 Bassam R. Ali3Bachar Afandi4Salem A. Beshyah5
Bassam R. Ali3Bachar Afandi4Salem A. Beshyah5 Charu Sharma6Wael Osman7Rajaa Mirghani8Amre Nasr9Sareea Al Remithi1Jamal Al Jubeh1Wasim Y. Almawi10Juma AlKaabi4,6
Charu Sharma6Wael Osman7Rajaa Mirghani8Amre Nasr9Sareea Al Remithi1Jamal Al Jubeh1Wasim Y. Almawi10Juma AlKaabi4,6 Gehad ElGhazali1,6*
Gehad ElGhazali1,6*Background: HLA class II (DR and DQ) alleles and antigens have historically shown strong genetic predisposition to type 1 diabetes (T1D). This study evaluated the association of DRB1 and DQB1 alleles, genotypes, and haplotypes with T1D in United Arab Emirates.
Materials and Methods: Study subjects comprised 149 patients with T1D, and 147 normoglycemic control subjects. Cases and controls were Emiratis and were HLA-DRB1 and -DQB1 genotyped using sequence-based typing. Statistical analysis was performed using Bridging Immunogenomic Data-Analysis Workflow Gaps R package.
Results: In total, 15 DRB1 and 9 DQB1 alleles were identified in the study subjects, of which the association of DRB1*03:01, DRB1*04:02, DRB1*11:01, DRB1*16:02, and DQB1*02:01, DQB1*03:02, DQB1*03:01, and DQB1*06:01 with altered risk of T1D persisted after correcting for multiple comparisons. Two-locus haplotype analysis identified DRB1*03:01∼DQB1*02:01 [0.44 vs. 0.18, OR (95% CI) = 3.44 (2.33–5.1), Pc = 3.48 × 10−10]; DRB1*04:02∼DQB1*03:02 [0.077 vs. 0.014, OR = 6.06 (2.03–24.37), Pc = 2.3 × 10−3] and DRB1*04:05∼DQB1*03:02 [0.060 vs. 0.010, OR = 6.24 (1.79–33.34), Pc = 0.011] as positively associated, and DRB1*16:02∼DQB1*05:02 [0.024 vs. 0.075, OR = 0.3 (0.11–0.74), Pc = 0.041] as negatively associated with T1D, after applying Bonferroni correction. Furthermore, the highest T1D risk was observed for DR3/DR4 [0.104 vs. 0.006, OR = 25.03 (8.23–97.2), Pc = 2.6 × 10−10], followed by DR3/DR3 [0.094 vs. 0.010, OR = 8.72 (3.17–25.32), Pc = 3.18 × 10−8] diplotypes.
Conclusion: While DRB1 and DQB1 alleles and haplotypes associated with T1D in Emiratis showed similarities to Caucasian and non-Caucasian populations, several alleles and haplotypes associated with T1D in European, African, and Asian populations, were not observed. This underscores the contribution of ethnic diversity and possible diverse associations between DRB1 and DQB1 and T1D across different populations.
Type I diabetes (T1D) is a complex autoimmune disorder characterized by insulin deficiency resulting from autoimmune destruction of insulin-secreting pancreatic β-cells in genetically predisposed individuals (Eisenbarth, 2010).
The International Diabetes Federation estimates that more than 1.1 million individuals below the age of 20 years are diagnosed with T1D worldwide (International Diabetes Federation, 2019). In this regard, it was reported that the incidence of T1D in children (15 years or younger) varies greatly among different ethnic and racial groups, highlighted by high to intermediate incidence among Europeans e.g., 5.2/100,000 per year in Finland (International Diabetes Federation, 2019; Knip, 2021), and the low incidence among Asians e.g., 1.8/100,000 per year in China (Gong et al., 2015; International Diabetes Federation, 2019). Among Arab populations of the Middle East, it was estimated that there are about 60,000 children with T1D under the age 15 years (Zayed, 2016). Of these, Kuwait and Saudi Arabia rank among the top ten countries worldwide in T1D prevalence, with rates of 44.5/100,000 and 33.5/100,000, respectively (Majeed et al., 2014; Shaltout et al., 2017; International Diabetes Federation, 2019). No reliable data have yet been reported on the T1D incidence in the United Arab Emirates (UAE). While the reason underlying the differences in T1D prevalence rate according to ethnicity and geographical locations is not known (Patterson et al., 2014; IDF 2019), it was suggested that both environmental parameters and genetic influence the initiation of T1D development (Eisenbarth, 2010; Blanter et al., 2019; Xia et al., 2019).
More than 60 genetic loci were reported in the literature as associated with altered T1D susceptibility (Todd et al., 2007; Bergholdt et al., 2012; Xia et al., 2019), of which the Human Leukocyte Antigens (HLA) class II genes contributed to approximately one-half of T1D genetic risk (Noble et al., 1996; Pociot et al., 2010). Individual DRB1 and DQB1 alleles, or allelic combinations were associated with the altered risk of T1D, and both susceptible and protective DRB1, DQA1, and DQB1 alleles were reportedly implicated with T1D pathogenesis (Noble et al., 1996; Erlich et al., 2008). This was highlighted by the association of DRB1*03:01:01∼DQB1*02:01 and DRB1*04:01:01∼DQB1*03:02 haplotypes with increased susceptibility to T1D among Bahraini Arabs (Al-Jenaidi et al., 2005). As the frequencies of HLA alleles, haplotypes, and genotypes show considerable population and ethnic differences, population studies confirmed that the relationship of HLA with T1D also varies according to the geographical location and ethnic background (Al-Jenaidi et al., 2005; Ahmadov et al., 2018; Fawwad et al., 2019; Zabeen et al., 2019). Compared to numerous studies performed on European populations, limited numbers of studies have been conducted on populations in the Middle East-North Africa (MENA) region (Al-Jenaidi et al., 2005; Noble et al., 2013).
This is the first case-control study investigating the association of HLA with T1D in the Emirati population. The present-day UAE national population is characterized by high rates of consanguinity, endogamy, along with sizeable extended families, and tribal community nature (Zayed, 2016). These and other features further justify investigating the association of HLA-class II antigens in T1D among Emirati Arabs. In addition to screening individual alleles, we aimed to assess the prevalence of DRB1∼DQB1 haplotype combination in search of specific susceptible and protective haplotypes.
The study subjects comprised 149 unrelated patients with T1D and 147 unrelated normoglycemic controls who were recruited from outpatient T1D clinics at Sheikh Khalifa Medical City (Abu Dhabi, UAE) and Tawam Hospital (Al-Ain, UAE). T1D was diagnosed based on clinical features and laboratory findings and on the 1985 World Health Organization (WHO) criteria (WHO Technical Report Series 727; World Health Organization, 1985) and the classification of diabetes mellitus document, WHO, Geneva; 2019 (World Health Organization, 1985; World Health Organisation, 2019). The inclusion criteria were patients who presented with acute symptoms of diabetes (e.g., polyuria, polydipsia and weight loss), had random blood glucose >200 mg/dl (11.1 mmol/L), C-peptide levels of <0.3 mmol/L, and had required long term insulin therapy since the time of diagnosis. Exclusion criteria included monogenic diabetes recognized by autosomal dominant mode of inheritance, type 2 diabetes as manifested by obesity and signs of insulin resistance, diabetes diagnosed before the age of 12 months or chronic diseases (especially autoimmune diseases) and patients/guardians who were unable to provide consent to participate in the study. Control individuals consisted of university students and healthy children with normal fasting/random blood glucose levels, no family history of T1D or other autoimmune diseases and were matched for age and gender with patients with T1D. Healthy children (≤16 years) were enrolled during routine hospital visits for other complaints while medical students (adolescents/adults) were recruited from the Tawam teaching hospital (Al Ain, UAE). For each recruited patient with T1D, a control was matched for age and gender to the extend possible. The mean ± SD age were 18.32 ± 7.45 (range 6–26 years) and 19.71 ± 6.93 (6–24) of cases and controls, respectively. Patients with T1D and their controls were Emirati nationals with grandparents born in the United Arab Emirates. Table 1 presents basic clinical and demographic characteristics of the study population.

TABLE 1. Clinical and demographic characteristics of the study population.
This study was performed following the guidelines of the Declaration of Helsinki (1993). The study protocol was approved by the Institutional Review Boards at SKMC (REC-25-10-2016 RS-445) and United Arab Emirates University (AAMDHREC 2016-4255 16-002), and informed consent was acquired from all study subjects or their guardians before performing the study.
All samples were analysed in our laboratory, which is a well-established clinical laboratory accredited by both the College of American Pathologists (CAP) and ISO15189. Total genomic DNA was extracted from EDTA-anticoagulated peripheral blood of study subjects using Qiagen DNA Mini kit on automated QIAcube, according to the manufacturer instructions (Qiagen, Hilden, Germany). DNA concentrations were determined using NanoDrop-2000 (Wilmington, DE, United States), and HLA-DRB1 and -DQB1 genotyping was performed by sequence-based typing (SBT), using GenDx AlleleSEQR kits (GenDx, https://www.gendx.com/, Netherland). Based on the method, exon 2 was sequenced for HLA-DRB1(AlleleSEQR HLA-DRB1, product code 08K63-03), and exon 2 and 3 were sequenced for HLA-DQB1 (AlleleSEQR HLA-DQBQ1, product code 08K64-03). All reagents necessary for primary amplification and sequencing were included in the HLA-DRB1 and –DQB1 AlleleSEQR SBT kits (GenDx, Netherland). Briefly, following the primary amplification using 9700 thermal cycler (Applied Biosystems, United States), the PCR products were purified by ExoSAP-IT and the PCR products were sequenced using BigDye Terminatory chemistry and the ABI-3130 genetic sequencer (Applied Biosystems, Foster City, CA). Finally, the sequence data were analysed using the GenDx SBTengine HLA typing software (http://www.gendx.com/product_line/sbtengine/), as described in the manufacturer’s protocol. Quality control measures consisted of randomly-chossing 10% of cases and control samples for retyping; concordance was 100%. In addition, six samples from the CAP External Quality Control Proficiency testing program were used to ensure control for the genotyping procedures.
The association of HLA-DRB1 and -DQB1 alleles and haplotypes with T1D was analysed using the Bridging Immunogenomic Data-Analysis Workflow Gaps (BIGDAWG) package (version 2.1) (Pappas et al., 2016), with 95% confidence intervals (CI), odds ratios (OR), and p-values (P) reported. BIGDAWG’s haplotype estimation function, which required the R “haplo.stats” package (version 1.7.7), was also used. Allele counts < 5 in study subjects were collapsed automatically through BIGDAWG into “binned” category (Pappas et al., 2016). The same analyses were performed at the haplotype level, which were estimated by the expectation-maximization (EM) approach (Dempster et al., 1977), and haplotypes with counts of three or fewer were binned. The corrected p values (Pc) for multiple comparisons were calculated using the Benferroni method. Analysis of Hardy-Weinberg equilibrium (HWE) deviations for specific haplotypes and individual loci was performed using PyPop (version 0.8.0), and Guo and Thompson’s exact method was used in identifying significant locus-level HWE deviations (Guo and Thompson, 1992; Lancaster et al., 2007). Individual haplotype deviations from HWE expectations were determined by Chen’s method (Guo and Thompson, 1992), at a significance of 0.05.
The pould (phased or unphased LD) R package (version 0.10.4.9000) (https://cran.r-project.org/package=pould) (Osoegawa et al., 2019) was used to calculate D′, Wn, Wa/b, and Wb/a global linkage disequilibrium (LD) values for DRB1∼DQB1 haplotypes (Thomson and Single 2014), in the range of 0 (equilibrium) to 1 (linkage). In LD, D’ signifies the weighted average of normalized disequilibrium (Dij) values, and it is less sensitive to variation across haplotypes of polymorphic loci than the other measures. Moreover, Wn indicates a correlation coefficient describing the association between alleles at two loci. The conditional asymmetric LD (ALD) measures, including Wa/b and Wb/a, extend the Wn measure for variation at one locus (a) conditioned on the variation at the second (b) (Single et al., 2016).
There was no deviation from HWE at DRB1 and DQB1 loci among controls, and no HWE deviation for DRB1 locus in patients, however, the DQB1 locus indicated an overall deviation from HWE (p = 0.0006) in patients. No trend toward excess heterozygosity or homozygosity was detected in patients. This deviation was due to the presence of five rare DQB1 genotypes—DQB1*02:03+DQB1*03:04 [1 observed (obs); 0.0268 expected (exp); p = 0.0271], DQB1*03:03+DQB1*04:01 (1 obs; 0.0067 exp; p = 0.0073), DQB1*02:03+DQB1*06:03 (1 obs; 0.0403 exp; p = 0.0388), DQB1*06:03+DQB1*06:03 (1 obs 0.0151 exp; p = 0.0106), and DQB1*05:01+DQB1*06:09 (1 obs; 0.0469 exp; p = 0.0358). Many of these alleles are observed less than five times in patients or controls (Supplementary Tables S1, S3).
Significant DRB1 allelic differences were seen between T1D patients and control subjects, 10 of 31 alleles differing significantly (p < 0.05). When the Bonferroni correction was applied, differences were significant for only 4, which comprised DRB1*03:01 [0.44 vs. 0.187, OR (95% CI) = 3.46 (2.35–5.11), Pc = 3.2 × 10−10] and DRB1*04:02 [0.081 vs. 0.014, OR = 6.35 (2.14–25.44), Pc = 2.0 × 10−3], which were higher among patients, and DRB1*11:01 [0.003 vs. 0.044, OR = 0.07 (0–0.49), Pc = 0.015] and DRB1*16:02 [0.023 vs. 0.078, OR = 0.28 (0.1–0.7), Pc = 0.030], which were higher in control subjects (Table 2). Similarly, significant allelic differences were seen at the DQB1 locus, 5 of 9 alleles differing significantly after the Bonferroni correction. These comprised DQB1*02:01 [0.48 vs. 0.19, OR = 3.78 (2.58–5.57), Pc = 3.04 × 10−12] and DQB1*03:02 [0.21 vs. 0.08, OR = 3.1 (1.82–5.40), Pc = 6.01 × 10−5] which were higher among patients, and DQB1*03:01 [0.02 vs. 0.11, OR = 0.16 (0.05–0.40), Pc = 5.64 × 10−5], DQB1*05:01 [0.05 vs. 0.11, OR = 0.40 (0.19–0.80), Pc = 0.044], and DQB1*06:01 [0.003 vs. 0.06, OR = 0.05 (0–0.33), Pc = 5.84 × 10−4], which were higher among control subjects (Table 3).
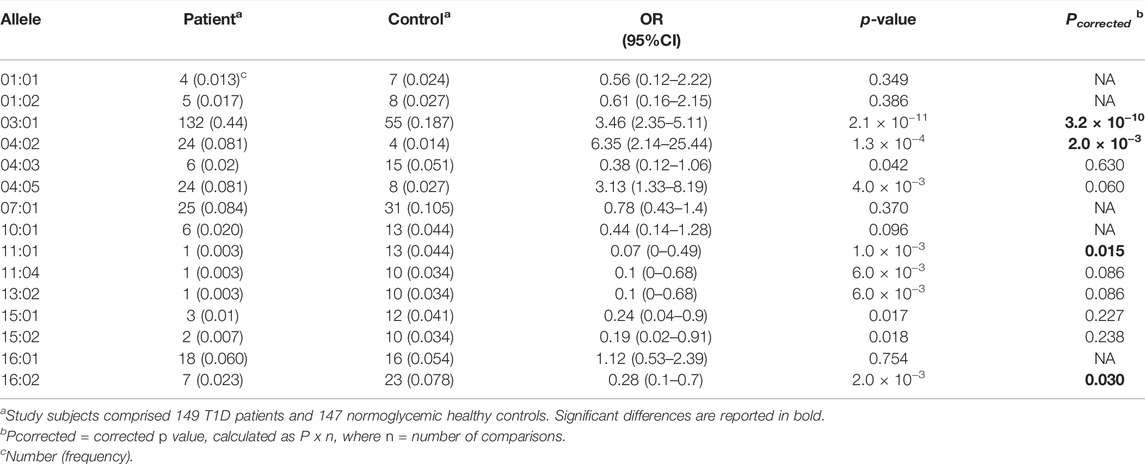
TABLE 2. Associations of DRB1 alleles with T1D in Emirati study subjects.
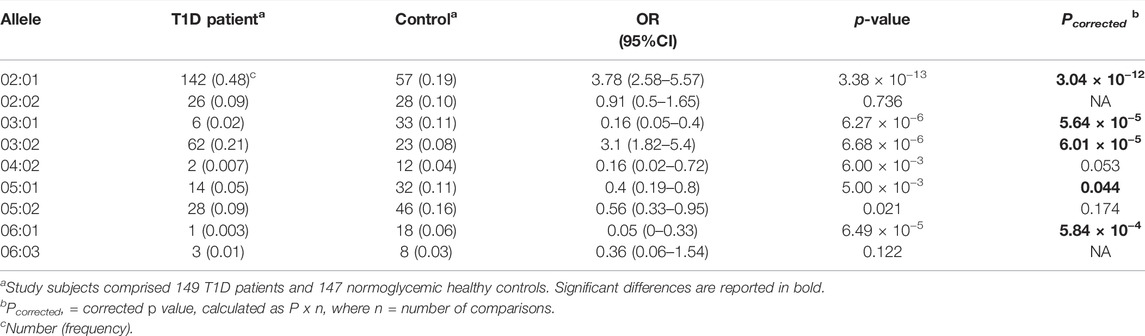
TABLE 3. Associations of DQB1 alleles with T1D in Emirati study subjects.
In total, 95 distinct DRB1∼DQB1 haplotypes were identified (Supplementary Table S2), of which 42 had counts of ≥2 in controls or patients, and thus were considered common. In total, four individual haplotypes showing a statistically significant association with T1D (Table 4). The full list of DRB1∼DQB1 haplotypes that were binned due to low expected counts is given in Supplementary Table S4. Higher frequencies of DRB1*03:01∼DQB1*02:01 [0.44 vs. 0.18, OR = 3.44 (2.33–5.10), Pc = 3.48 × 10−10], DRB1*04:02∼DQB1*03:02 [0.077 vs. 0.014, OR = 6.06 (2.03–24.37), Pc = 2.3 × 10−3], and DRB1*04:05∼DQB1*03:02 [0.06 vs. 0.01, OR = 6.24 (1.79–33.34), Pc = 0.011] haplotypes, and lower frequency of DRB1*16:02∼DQB1*05:02 haplotype [0.024 vs. 0.075, OR (95% CI) = 0.30 (0.11–0.74), Pc = 0.041] were seen in T1D patients compared to controls. This assigned T1D susceptibility and protective nature to these haplotypes, respectively (Table 4).
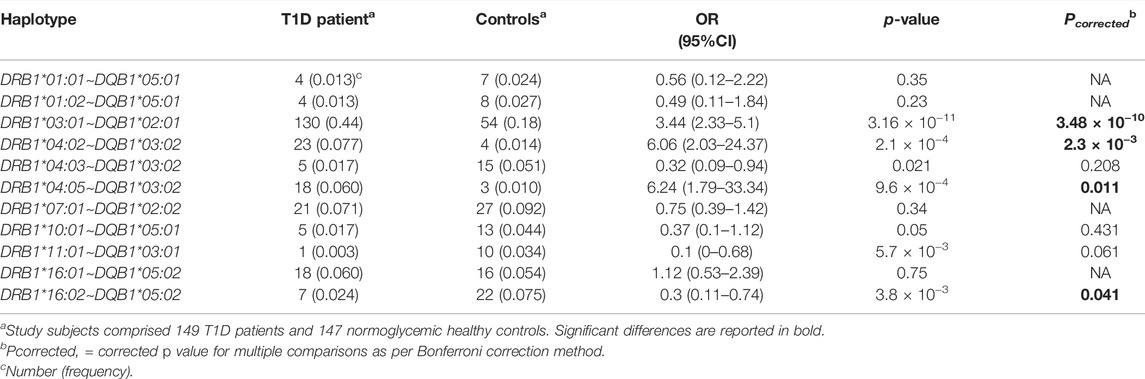
TABLE 4. Association of DRB1-DQB1 Haplotypes with T1D in Emirati study subjects.
Parallel to the differential distribution of 2-locus haplotypes among T1D patients and controls, distinct DRB1∼DQB1 + DRB1∼DQB1 diplotypes were seen in T1D patients. Extensive diversity in the diplotypes identified was seen, and as such we focused on the DRB1*03- and DRB1*04-containing diplotypes for analysis. As shown in Table 5, DRB1*03:01∼DQB1*02:01 + DRB1*04:02/05∼DQB1*03:02 0.104 vs. 0.006, OR = 25.03 (8.23–97.20, p = 2.6 × 10−10) followed by the homozygous DRB1*03:01∼DQB1*02:01 + DRB1*03:01∼DQB1*02:01 [0.094 vs. 0.01, OR = 8.72 (3.17–25.32), p = 3.18 × 10−8] were associated with heightened risk for T1D.
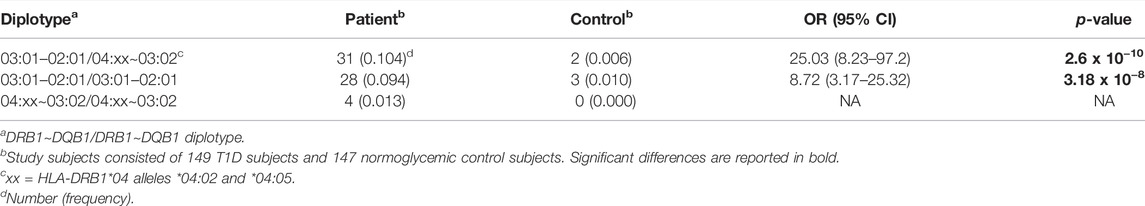
TABLE 5. Associations of DRB1∼DQB1 diplotypes with T1D in the study subjects.
D′ and Wn values, as well as WDRB1/DQB1 and WDQB1/DRB1 measures, which assess the global linkage disequilibrium (LD) between DRB1 and DQB1 loci in T1D patients and control groups are shown in Supplementary Table S5. There were no substantial differences between T1D cases and controls in terms of LD. Strong LD between DRB1 and DQB1 loci is illustrated by the high D’ measure (0.92 in patients and 0.95 in controls), and to a lesser extent by Wn (0.75 in patients and 0.72 in controls). The ALD measures (WDRB1/DQB1 and WDQB1/DRB) dissect the variation on each locus conditioned on the other. Comparison of the WDRB1/DQB1 (0.89 in patients and 0.90 in controls) with WDQB1/DRB1 (0.71 in both groups) indicated less variation among DRB1 alleles related to DQB1, when compared to DQB1 alleles related to DRB1.
The HLA contribution to T1D genetic susceptibility differs between populations and ethnic groups (Noble et al., 2013), largely due to varied frequencies and functional associations of DRB1 and DQB1 alleles and haplotypes (susceptible, protective). Key DRB1 and DQB1 alleles and haplotypes associated with T1D were reported for Caucasian and non-Caucasian populations (Petrone et al., 2001; Erlich et al., 2008; Ilonen et al., 2009; Noble et al., 2013), which included DRB1*03:01, DRB1*04:02, DRB1*04:05, DQB1*02:01, and DQB1*03:02 along with DRB1*03:01∼DQB1*02:01 (DR3), DRB1*04:02∼DQB1*03:02 (DR4), and DRB1*04:05∼DQB1*03:02 haplotypes also observed in this study and studies on other Arab populations from Bahrain, Tunisia, Lebanon and Saudi Arabia (Al-Harbi et al., 2004; Al-Jenaidi et al., 2005; Stayoussef et al., 2009; Manan et al., 2010; El-Amir et al., 2015; Eltayeb-Elsheikh et al., 2020). It is noteworthy that no DR3 or DR4 haplotype linked with T1D was detected in East and Southeast Asians (Hashimoto et al., 1994; Kawabata et al., 2002).
DRB1*04:01∼DQB1*03:02 (0.34% controls frequency) and DRB1*04:01∼DQB1*03:01 (not seen in controls) were rare in Emiratis. This was reminiscent of findings in Europeans where DRB1*04:01∼DQB1*03:02 haplotype the second susceptibility haplotype, compared to DRB1*04:01∼DQB1*03:01 which was protective in Europeans (Erlich et al., 2008; Noble et alel., 2013), but not Africans (Onengut-Gumuscu et al., 2019). In our study, the DRB1*04:03 allele and DRB1*04:03∼DQB1*03:02 haplotype were more prevalent in the healthy controls but were not statistically different after correcting for multiple comparisons. The highest T1D risk in Emiratis was conferred by DRB1*04:05∼DQB1*03:02, DRB1*04:02∼DQB1*03:02 and DRB1*03:01∼DQB1*02:01 haplotypes. By comparison, the highest risk in European was imparted by DRB1*04:05∼DQB1*03:02, DRB1*04:01∼DQB1*03:02, DRB1*04:02∼DQB1*03:02, and DRB1*03:01∼DQB1*02:01 (Erlich et al., 2008).
DQB1*03:03, DRB1*08:01, DRB1*08:02, and DRB1*09:01 were rare among patients or controls in the present study. Moreover, DRB1*03:02 was rare in the Emirati study population (Al-Yafei et al., 2019; Arnaiz-Villena et al., 2019) and European derived populations; and frequent in DQB1*04:02-containing haplotypes in Africans, protecting against T1D (Howson et al., 2013). DQB1*04:01, common in East Asians (Kawabata et al., 2002), is rare in Emiratis (Al-Yafei et al., 2019; Arnaiz-Villena et al., 2019). Furthermore, several DRB1∼DQB1 haplotypes associated with T1D in Europeans, Asians, and Africans were not observed here. For example, the DRB1*04:05∼DQB1*02:01 haplotype predisposed to T1D susceptibility in African Americans, while the African DR3 (DRB1*03:02∼DQB1*04:02) haplotype was protective (Noble et al., 2013). In addition to DR3 (DRB1*03:01∼DQB1*02:01) and DR4 (DRB1*04∼DQB1*03:02, DRB1*08:01∼DQB1*04:02 is a T1D predisposing haplotype in Europeans (Thomson et al., 2007). The African DRB1*03:02∼DQB1*04:02 and DRB1*04:05∼DQB1*02:01 haplotypes were rare in our study and in European populations. The DQB1*04:02 allele was more prevalent in the Emirati healthy controls but was not statistically different after correcting for multiple comparisons. DRB1*xx:xx ∼ DQB1*04:02 haplotypes were detected at low counts. The DRB1*08:02∼DQB1*03:02, DRB1*09:01∼DQB1*03:03, and DRB1*04:05∼DQB1*04:01 haplotypes, associated with increased T1D susceptibility in East Asians (Ikegami et al., 2008; Katahira et al., 2009), were rare in the current study population (Supplementary Tables S2, S4). The DRB1*04:05∼DQB1*04:01 haplotype common in East Asia populations (Japanese, Taiwanese, Philippines), is rare in Europeans and African Americans (Thomson et al., 2007), in line with the findings here. On the other hand, DRB1*09:01∼DQB1*03:03, also rare in African Americans, is found in Europeans and East Asians (Thomson et al., 2007). It should be noted that a unique (DR9) DRB1*09:01∼DQA1*03:01∼DQB1*02:02 haplotype (but not the DRB1*09:01∼DQB1*03:03) haplotype has been detected in Africans (Noble et al., 2013).
While high prevalence of DRB1*07:01∼DQB1*02:02 was seen in controls (9.18%) and cases (7.05%), no significant difference in its distribution between cases and controls was seen. DRB1*07:01∼DQA1*02:01∼DQB1*02:02 conferred some protection against T1D in Europeans (Erlich et al., 2008), while DRB1*07:01∼DQA1*03:01∼DQB1*02:02 was susceptible for T1D in Africans (Noble et al., 2013). Varied T1D risk of DR7-containing haplotypes was dependent on the DQA1 allele contained in the DRB1*07:01∼DQA1*xx:xx∼DQB1*02:02 haplotype. Noble et al. (2013) showed that the DRB1*07:01∼DQB1*03:03 haplotype was protective of T1D in Europeans but was rare in Africans; this haplotype is rare in the current study (0.34% in both groups). Moreover, the distribution of DRB1*01:01∼DQB1*05:01, DRB1*01:02∼DQB1*05:02, DRB1*16:01∼DQB1*05:02, and DRB1*10:01∼DQB1*05:01 haplotypes were comparable between T1D cases and controls. While DRB1*01:01/01:02∼DQB1*05:01 was shown earlier to be associated with increased susceptibility to T1D (Thomson et al., 2007), this was not consistent with subsequent studies (Erlich et al., 2008). The DRB1*16:02∼DQB1*05:02 haplotype was protective for T1D while DRB1*11:01∼DQB1*03:01 and DRB1*10:01∼DQB1*05:01 were more prevalent in the healthy controls but were not statistically different after correcting for multiple comparisons. DRB1*15:01∼DQB1*06:02, DRB1*14:01∼DQB*05:03, and DRB1*07:01∼DQB1*03:03 (Noble et al., 1996; Petrone et al., 2001; Noble et al., 2011) are protective in Europeans but were observed at low frequencies in the present study and were binned (Supplemental S2 and S4). DRB1*15:01-DQB1*06:02 was seen in approximately 20% of Europeans, but in only 1% of T1D individuals (Noble et al., 1996). DRB1*16:02-DQB1*05:02 is common in controls (7.48%) and patients (2.35%), but is rare in Europeans, Africans, and Asians (Hashimoto et al., 1994). DRB1*16:01∼DQB1*05:02, a common DR2 haplotype in the studied population (6.04 and 5.44% in patients and controls, respectively), was not associated with T1D. In addition, DRB1*15:01∼DQB1*06:02, a T1D protective haplotype in most ethnicities (Osoegawa et al., 2019), was detected at low frequency in patients (0.67%) and controls (1.67%). Moreover, the (protective) DRB1*15:03∼DQB1*06:02, DRB1*08:04∼DQB1*03:01, and DRB1*03:02∼DQB1*04:02 African haplotypes were rare in our study population.
Differential contribution of DR∼DQ haplotypes to T1D risk were recognized; this was dictated by the specific ethnic background. Most of our T1D patients carried (DRB1*03:01∼DQB1*02:01) (DR3) or (DRB1*04:02/05∼DQB1*03:02) (DR4) haplotypes (87%), and 23% of these were carriers of the heterozygous DR3/DR4 diplotype. In particular, the DRB1*03:01∼DQB1*02:01/DRB1*04:02/05∼DQB1*03:02 (heterozygous) diplotype imparted the highest T1D risk, followed by DRB1*03:01∼DQB1*02:01/DRB1*03:01∼DQB1*02:01 (homozygous) diplotype. This was comparable to Europeans, where most (90%) T1D patients were DR3 or DR4 carriers, with 40% carrying the DR3/DR4 diplotype (Ronningen et al., 1991; Noble et al., 1996), which is higher than the 12% DR3/DR4 diplotype carrier rate seen in Africans (Noble et al., 1996). Furthermore, the highest risk for T1D in Europeans was conferred by the DR3/DR4 diplotype, rather than DR3/DR3 (Noble et al., 1996; Hermann et al., 2003; Erlich et al., 2008), while in East Asians, the highest T1D risk was conferred by the DR4/DR9 diplotype (Ikegami et al., 2008). This absence of DR3 might explain the low prevalence of T1D in these populations, as suggested earlier (Ikegami et al., 2008).
The apparent discrepancies between our results and those of other populations can be attributed to several factors, including race/ethnicity, incidence of T1D, sample size, genotyping methods, and other factors (Park, 2007; Polychronakos and Li 2011). This was highlighted by the HLA allele/haplotype frequencies between our population and Europeans, Africans, and East Asians (Al-Yafei et al., 2019; Arnaiz-Villena et al., 2019). This in turn explains, at least in part, the variations in T1D prevalence and incidence (Patterson et al., 2012). The high LD values between DRB1 and DQB1 loci were found in Emiratis, consistent with the findings in other ethnic groups. LD values were similar in cases and controls, and were identical for WDQB1/DRB1, suggesting equivalent diversity of DQB1 alleles within DRB1∼DQB1 haplotypes between the groups. The intermediate Wn LD measure value relative to WDRB1/DQB1 and WDQB1/DRB1 measures in patients illustrates the importance for applying the ALD measures for highly polymorphic genetic systems.
The highest T1D risk was imparted by the DRB1*03:01∼DQB1*02:01/DRB1*04:02-05∼DQB1*03:02 diplotype followed by the DRB1*03:01∼DQB1*02:01/DRB1*03:01∼DQB1*02:01 diplotype. Emiratis showed similarities with and differences from European, Asian, and African populations in terms of HLA-DRB1 and -DQB1 alleles and haplotypes and their associations with T1D. However, our study had some shortcomings, namely the relatively small sample size, and lack of DQA1 genotyping (especially for DR7+ samples). An interesting dimension of ethnic diversity and possible diverse relationships is highlighted between HLA-DRB1 and -DQB1 genes and T1D across different populations.
All sequences were submitted to the Short Read Archive (SRA) at GenBank and assigned BioSample accession numbers between SAMN26674334 and SAMN26674604 under BioProject PRJNA609073.
The studies involving human participants were reviewed and approved by Institutional Review Boards at SKMC (REC-25-10-2016 RS-445) and United Arab Emirates University (AAMDHREC 2016-4255 16-002). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
ZA, GE, JA and BRA draft the idea of the research and the research proposal. MA and CS conducted the laboratory and genetic analysis. RM and AN did data cleaning. SM, WO and WA conducted the data analysis, prepared the tables and contributed to the writing of the statistical components of the study. ZA wrote the first draft of the manuscript under supervision of GE. BA, SB, SA, JA, RM and AN contributed to participant’s recruitment and reviewed the manuscript. RM, AN and WO prepare the manuscript for publication. All authors reviewed the final version of the manuscript and gave approval for submission for publication of this manuscript.
This work was funded by the College of Medicine and Health Sciences; United Arab Emirates University grant code 21M103-4 IRCA. This work was also supported by National Institutes of Health (NIH) National Institute of Allergy and Infectious Disease (NIAID) grant R01AI28775 (SM). The content is solely the responsibility of the authors and does not necessarily reflect the official views of the NIAID, NIH or United States Government.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors are most grateful to all the patients and parents/guardians who for participation to the study, all physicians, diabetes educators and nurses who helped identification of appropriate patients, and admin and technical staff for data entry and support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.841879/full#supplementary-material
Ahmadov, G. A., Govender, D., Atkinson, M. A., Sultanova, R. A., Eubova, A. A., Wasserfall, C. H., et al. (2018). Epidemiology of Childhood-Onset Type 1 Diabetes in Azerbaijan: Incidence, Clinical Features, Biochemistry, and HLA-DRB1 Status. Diabetes Res. Clin. Pract. 144, 252–259. doi:10.1016/j.diabres.2018.09.009
Al‐Yafei, Z., Goeury, T., Alvares, M., Al Seiari, M., Sanchez‐Mazas, A., and Elghazali, G. (2020). United Arab Emirates: Unusual Departure from Neutrality towards Excess of Homozygotes at the HLA‐B Locus. HLA 95 (5), 470–473. doi:10.1111/tan.13774
Al-Harbi, E. M., Abbassi, A.-J., Tamim, H., al-Jenaidi, F., Kooheji, M., Kamal, M., et al. (2004). Specific HLA-DRB and -DQB Alleles and Haplotypes Confer Disease Susceptibility or Resistance in Bahraini Type 1 Diabetes Patients. Clin. Vaccin. Immunol 11 (2), 292–296. doi:10.1128/cdli.11.2.292-296.2004
Al-Jenaidi, F. A., Wakim-Ghorayeb, S. F., Al-Abbasi, A., Arekat, M. R., Irani-Hakime, N., Najm, P., et al. (2005). Contribution of Selective HLA-Drb1/dqb1 Alleles and Haplotypes to the Genetic Susceptibility of Type 1 Diabetes Among Lebanese and Bahraini Arabs. J. Clin. Endocrinol. Metab. 90 (9), 5104–5109. doi:10.1210/jc.2005-1166
Arnaiz-Villena, A., Yafei, Z. A., Juarez, I., Palacio-Gruber, J., Mahri, A. A., Alvares, M., et al. (2019). HLA Genetic Study from United Arab Emirates (UAE), Abu Dhabi. Hum. Immunol. 80 (7), 421–422. doi:10.1016/j.humimm.2019.04.013
Bergholdt, R., Brorsson, C., Palleja, A., Berchtold, L. A., Fløyel, T., Bang-Berthelsen, C. H., et al. (2012). Identification of Novel Type 1 Diabetes Candidate Genes by Integrating Genome-wide Association Data, Protein-Protein Interactions, and Human Pancreatic Islet Gene Expression. Diabetes 61 (4), 954–962. doi:10.2337/db11-1263
Blanter, M., Sork, H., Tuomela, S., and Flodström-Tullberg, M. (2019). Genetic and Environmental Interaction in Type 1 Diabetes: a Relationship between Genetic Risk Alleles and Molecular Traits of Enterovirus Infection? Curr. Diab Rep. 19 (9), 82. doi:10.1007/s11892-019-1192-8
Dempster, A. P., Laird, N. M., and Rubin, D. B. (1977). Maximum Likelihood from Incomplete Data via theEMAlgorithm. J. R. Stat. Soc. Ser. B (Methodological) 39 (1), 1–22. doi:10.1111/j.2517-6161.1977.tb01600.x
Eisenbarth, G. S. (2010). Banting Lecture 2009: an Unfinished Journey: Molecular Pathogenesis to Prevention of Type 1A Diabetes. Diabetes 59 (4), 759–774. doi:10.2337/db09-1855
El‐Amir, M. I., El‐Feky, M. A., Laine, A. P., Härkönen, T., El‐Badawy, O., Eltayeb, A. A., et al. (2015). Risk Genes and Autoantibodies in Egyptian Children with Type 1 Diabetes - Low Frequency of Autoantibodies in Carriers of the HLA‐DRB1*04:05‐DQA1*03‐DQB1*02 Risk Haplotype. Diabetes Metab. Res. Rev. 31 (3), 287–294. doi:10.1002/dmrr.2609
Eltayeb‐Elsheikh, N., Khalil, E., Mubasher, M., Aljurayyan, A., AlHarthi, H., Omer, W. H., et al. (2020). Association of HLA‐DR‐DQ Alleles, Haplotypes, and Diplotypes with Type 1 Diabetes in Saudis. Diabetes Metab. Res. Rev. 36 (8), e3345. doi:10.1002/dmrr.3345
Erlich, H., Valdes, A. M., Noble, J., Carlson, J. A., Varney, M., Concannon, P., et al. (2008). HLA DR-DQ Haplotypes and Genotypes and Type 1 Diabetes Risk. Diabetes 57 (4), 1084–1092. doi:10.2337/db07-1331
Fawwad, A., Govender, D., Ahmedani, M. Y., Basit, A., Lane, J. A., Mack, S. J., et al. (2019). Clinical Features, Biochemistry and HLA-DRB1 Status in Youth-Onset Type 1 Diabetes in Pakistan. Diabetes Res. Clin. Pract. 149, 9–17. doi:10.1016/j.diabres.2019.01.023
Gong, C., Meng, X., Jiang, Y., Wang, X., Cui, H., Chen, X., et al. (2015). Trends in Childhood Type 1 Diabetes Mellitus Incidence in Beijing from 1995 to 2010: A Retrospective Multicenter Study Based on Hospitalization Data. Diabetes Technology Ther. 17 (3), 159–165. doi:10.1089/dia.2014.0205
Guo, S. W., and Thompson, E. A. (1992). Performing the Exact Test of Hardy-Weinberg Proportion for Multiple Alleles. Biometrics 48 (2), 361–372. doi:10.2307/2532296
Hashimoto, M., Kinoshita, T., Yamasaki, M., Tanaka, H., Imanishi, T., Ihara, H., et al. (1994). Gene Frequencies and Haplotypic Associations within the HLA Region in 916 Unrelated Japanese Individuals. Tissue Antigens 44 (3), 166–173. doi:10.1111/j.1399-0039.1994.tb02375.x
Hermann, R., Turpeinen, H., Laine, A. P., Veijola, R., Knip, M., Simell, O., et al. (2003). HLA DR-DQ-Encoded Genetic Determinants of Childhood-Onset Type 1 Diabetes in Finland: An Analysis of 622 Nuclear Families. Tissue Antigens 62 (2), 162–169. doi:10.1034/j.1399-0039.2003.00071.x
Howson, J. M. M., Roy, M. S., Zeitels, L., Stevens, H., and Todd, J. A. (2013). HLA Class II Gene Associations in African American Type 1 Diabetes Reveal a Protective HLA‐DRB1*03 Haplotype. Diabet. Med. 30 (6), 710–716. doi:10.1111/dme.12148
Ikegami, H., Noso, S., Babaya, N., Hiromine, Y., and Kawabata, Y. (2008). Genetic Basis of Type 1 Diabetes: Similarities and Differences between East and West. Rev. Diabet. Stud. 5 (2), 64–72. doi:10.1900/RDS.2008.5.64
Ilonen, J., Kocova, M., Lipponen, K., Sukarova-Angelovska, E., Jovanovska, A., and Knip, M. (2009). HLA-DR-DQ Haplotypes and Type 1 Diabetes in Macedonia. Hum. Immunol. 70 (6), 461–463. doi:10.1016/j.humimm.2009.03.014
International Diabetes Federation (2019). IDF Diabetes Atlas. 9th ed. Brussels: International Diabetes Federation.
Katahira, M., Maeda, H., Tosaki, T., and Segawa, S. (2009). The Human Leukocyte Antigen Class II Gene Has Different Contributions to Autoimmune Type 1 Diabetes with or without Autoimmune Thyroid Disease in the Japanese Population. Diabetes Res. Clin. Pract. 85 (3), 293–297. doi:10.1016/j.diabres.2009.06.007
Kawabata, Y., Ikegami, H., Kawaguchi, Y., Fujisawa, T., Shintani, M., Ono, M., et al. (2002). Asian-Specific HLA Haplotypes Reveal Heterogeneity of the Contribution of HLA-DR and -DQ Haplotypes to Susceptibility to Type 1 Diabetes. Diabetes 51 (2), 545–551. doi:10.2337/diabetes.51.2.545
Knip, M. (2021). Type 1 Diabetes in Finland: Past, Present, and Future. Lancet Diabetes Endocrinol. 9 (5), 259–260. doi:10.1016/S2213-8587(21)00074-7
Lancaster, A. K., Single, R. M., Solberg, O. D., Nelson, M. P., and Thomson, G. (2007). PyPop Update - a Software Pipeline for Large-Scale Multilocus Population Genomics. Tissue Antigens 69 (1), 192–197. doi:10.1111/j.1399-0039.2006.00769.x
Majeed, A., El-Sayed, A. A., Khoja, T., Alshamsan, R., Millett, C., and Rawaf, S. (2014). Diabetes in the Middle-East and North Africa: an Update. Diabetes Res. Clin. Pract. 103 (2), 218–222. doi:10.1016/j.diabres.2013.11.008
Manan, H., Angham, A. M., and Sitelbanat, A. (2010). Genetic and Diabetic Auto-Antibody Markers in Saudi Children with Type 1 Diabetes. Hum. Immunol. 71 (12), 1238–1242. doi:10.1016/j.humimm.2010.09.008
Noble, J. A., Valdes, A. M., Cook, M., Klitz, W., Thomson, G., and Erlich, H. A. (1996). The Role of HLA Class II Genes in Insulin-dependent Diabetes Mellitus: Molecular Analysis of 180 Caucasian, Multiplex Families. Am. J. Hum. Genet. 59 (5), 1134–1148.
Noble, J. A., Johnson, J., Lane, J. A., and Valdes, A. M. (2013). HLA Class II Genotyping of African American Type 1 Diabetic Patients Reveals Associations Unique to African Haplotypes. Diabetes 62 (9), 3292–3299. doi:10.2337/db13-0094
Noble, J. A., Johnson, J., Lane, J. A., and Valdes, A. M. (2011). Race-specific Type 1 Diabetes Risk of HLA-DR7 Haplotypes. Tissue Antigens 78 (5), 348–351. doi:10.1111/j.1399-0039.2011.01772.x
Onengut-Gumuscu, S., Chen, W.-M., Robertson, C. C., Bonnie, J. K., Farber, E., Zhu, Z., et al. (2019). Type 1 Diabetes Risk in African-Ancestry Participants and Utility of an Ancestry-specific Genetic Risk Score. Diabetes Care 42 (3), 406–415. doi:10.2337/dc18-1727
Osoegawa, K., Mack, S. J., Prestegaard, M., and Fernández-Viña, M. A. (2019). Tools for Building, Analyzing and Evaluating HLA Haplotypes from Families. Hum. Immunol. 80 (9), 633–643. doi:10.1016/j.humimm.2019.01.010
Pappas, D. J., Marin, W., Hollenbach, J. A., and Mack, S. J. (2016). Bridging Immunogenomic Data Analysis Workflow Gaps (BIGDAWG): An Integrated Case-Control Analysis Pipeline. Hum. Immunol. 77 (3), 283–287. doi:10.1016/j.humimm.2015.12.006
Park, Y. (2007). Functional Evaluation of the Type 1 Diabetes (T1D) Susceptibility Candidate Genes. Diabetes Res. Clin. Pract. 77, S110–S115. doi:10.1016/j.diabres.2007.01.043
Patterson, C. C., Gyürüs, E., Rosenbauer, J., Cinek, O., Neu, A., Schober, E., et al. (2012). Trends in Childhood Type 1 Diabetes Incidence in Europe during 1989-2008: Evidence of Non-uniformity over Time in Rates of Increase. Diabetologia 55 (8), 2142–2147. doi:10.1007/s00125-012-2571-8
Patterson, C., Guariguata, L., Dahlquist, G., Soltész, G., Ogle, G., and Silink, M. (2014). Diabetes in the Young - a Global View and Worldwide Estimates of Numbers of Children with Type 1 Diabetes. Diabetes Res. Clin. Pract. 103 (2), 161–175. doi:10.1016/j.diabres.2013.11.005
Petrone, A., Bugawan, T. L., Mesturino, C. A., Nisticò, L., Galgani, A., Giorgi, G., et al. (2001). The Distribution of HLA Class II Susceptible/protective Haplotypes Could Partially Explain the Low Incidence of Type 1 Diabetes in continental Italy (Lazio Region). Tissue Antigens 58 (6), 385–394. doi:10.1034/j.1399-0039.2001.580607.x
Pociot, F., Akolkar, B., Concannon, P., Erlich, H. A., Julier, C., Morahan, G., et al. (2010). Genetics of Type 1 Diabetes: What's Next? Diabetes 59 (7), 1561–1571. doi:10.2337/db10-0076
Polychronakos, C., and Li, Q. (2011). Understanding Type 1 Diabetes through Genetics: Advances and Prospects. Nat. Rev. Genet. 12 (11), 781–792. doi:10.1038/nrg3069
Rønningen, K. S., Spurkland, A., Iwe, T., Vartdal, F., and Thorsby, E. (1991). Distribution of HLA-DRB1, -DQA1 and -DQB1 Alleles and DQA1-DQB1 Genotypes Among Norwegian Patients with Insulin-dependent Diabetes Mellitus. Tissue Antigens 37 (3), 105–111. doi:10.1111/j.1399-0039.1991.tb01854.x
Shaltout, A. A., Wake, D., Thanaraj, T. A., Omar, D. M., Al-AbdulRazzaq, D., Channanath, A., et al. (2017). Incidence of Type 1 Diabetes Has Doubled in Kuwaiti Children 0-14 Years over the Last 20 Years. Pediatr. Diabetes 18 (8), 761–766. doi:10.1111/pedi.12480
Single, R. M., Strayer, N., Thomson, G., Paunic, V., Albrecht, M., and Maiers, M. (2016). Asymmetric Linkage Disequilibrium: Tools for Assessing Multiallelic LD. Hum. Immunol. 77 (3), 288–294. doi:10.1016/j.humimm.2015.09.001
Stayoussef, M., Benmansour, J., Al-Jenaidi, F. A., Nemr, R., Ali, M. E., Mahjoub, T., et al. (2009). Influence of Common and Specific HLA-Drb1/dqb1 Haplotypes on Genetic Susceptibilities of Three Distinct Arab Populations to Type 1 Diabetes. Clin. Vaccin. Immunol. 16 (1), 136–138. doi:10.1128/CVI.00215-08
Thomson, G., and Single, R. M. (2014). Conditional Asymmetric Linkage Disequilibrium (ALD): Extending the Biallelic R2 Measure. Genetics 198 (1), 321–331. doi:10.1534/genetics.114.165266
Thomson, G., Valdes, A. M., Noble, J. A., Kockum, I., Grote, M. N., Najman, J., et al. (2007). Relative Predispositional Effects of HLA Class II DRB1-DQB1 Haplotypes and Genotypes on Type 1 Diabetes: a Meta-Analysis. Tissue Antigens 70 (2), 110–127. doi:10.1111/j.1399-0039.2007.00867.x
Todd, J. A., Walker, N. M., Walker, N. M., Cooper, J. D., Smyth, D. J., Downes, K., et al. (2007). Robust Associations of Four New Chromosome Regions from Genome-wide Analyses of Type 1 Diabetes. Nat. Genet. 39 (7), 857–864. doi:10.1038/ng2068
World Health Organisation (2019). Classification of Diabetes Mellitus. Geneva: WHO. Licence: CC BY-NC-SA 3.0 IGO https://creativecommons.org/licenses/by-nc-sa/3.0/igo.
World Health Organization (1985). Diabetes Mellitus: Report of a WHO Study Group. Technical Report Series No. 727. Geneva: WHO.
Xia, Y., Xie, Z., Huang, G., and Zhou, Z. (2019). Incidence and Trend of Type 1 Diabetes and the Underlying Environmental Determinants. Diabetes Metab. Res. Rev. 35 (1), e3075. doi:10.1002/dmrr.3075
Zabeen, B., Govender, D., Hassan, Z., Noble, J. A., Lane, J. A., Mack, S. J., et al. (2019). Clinical Features, Biochemistry and HLA-DRB1 Status in Children and Adolescents with Diabetes in Dhaka, Bangladesh. Diabetes Res. Clin. Pract. 158, 107894. doi:10.1016/j.diabres.2019.107894
Keywords: type 1 diabetes, HLA, haplotypes, Emiratis, ethnicity
Citation: Al Yafei Z, Mack SJ, Alvares M, Ali BR, Afandi B, Beshyah SA, Sharma C, Osman W, Mirghani R, Nasr A, Al Remithi S, Al Jubeh J, Almawi WY, AlKaabi J and ElGhazali G (2022) HLA-DRB1 and –DQB1 Alleles, Haplotypes and Genotypes in Emirati Patients with Type 1 Diabetes Underscores the Benefits of Evaluating Understudied Populations. Front. Genet. 13:841879. doi: 10.3389/fgene.2022.841879
Received: 22 December 2021; Accepted: 31 January 2022;
Published: 24 March 2022.
Edited by:
Tina Bharani, Thomas Jefferson University Hospital, Jefferson University Hospitals, United StatesReviewed by:
Narinder K. Mehra, All India Institute of Medical Sciences, IndiaCopyright © 2022 Al Yafei, Mack, Alvares, Ali, Afandi, Beshyah, Sharma, Osman, Mirghani, Nasr, Al Remithi, Al Jubeh, Almawi, AlKaabi and ElGhazali. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gehad ElGhazali, Z2VsZ2hhemFsaUB1bmlvbjcxLmFl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.