 Kevin Santos da Silva1
Kevin Santos da Silva1 Larissa Glugoski2,3
Larissa Glugoski2,3 Marcelo Ricardo Vicari3
Marcelo Ricardo Vicari3 Augusto César Paes de Souza4Renata Coelho Rodrigues Noronha1
Augusto César Paes de Souza4Renata Coelho Rodrigues Noronha1 Julio Cesar Pieczarka1
Julio Cesar Pieczarka1 Cleusa Yoshiko Nagamachi1*
Cleusa Yoshiko Nagamachi1*- 1Laboratório de Citogenética, Centro de Estudos Avançados da Biodiversidade, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, Brazil
- 2Laboratório de Citogenética de Peixes, Universidade Federal de São Carlos, São Carlos, Brazil
- 3Laboratório de Biologia Cromossômica: Estrutura e Função, Departamento de Biologia Estrutural, Molecular e Genética, Universidade Estadual de Ponta Grossa, Ponta Grossa, Brazil
- 4Laboratório de Estudo da Ictiofauna Amazônica, Instituto Federal de Educação, Ciência e Tecnologia do Pará, Abaetetuba, Brazil
The Ancistrus genus has extensive chromosomal diversity among species, including heteromorphic sex chromosomes occurrence. However, studies have been shown that chromosomal diversity may still be underestimated. Repetitive sequences represent a large part of eukaryotic genomes, associated with mechanisms of karyotypic diversification, including sex chromosomes evolution. This study analyzed the karyotype diversification of two Ancistrus species (Ancistrus sp. 1 and Ancistrus sp. 2) from the Amazon region by classical and molecular chromosomal markers. Conventional chromosome bands and fluorescence in situ hybridization using probes 18S and 5S rDNA, besides (CA)n, (CG)n, (GA)n, (CAC)n, (CAG)n, (CAT)n, (GAA)n, (GAC)n, (TAA)n, and (TTAGGG)n in tandem repeats were determined on the karyotypes. Ancistrus sp. 1 and Ancistrus sp. 2 presented karyotypes with 2n = 38 (20 m + 14sm+4st, XX/XY) and 2n = 34 (20 m + 14sm, without heteromorphic sex chromosomes), respectively. Robertsonian rearrangements can explain the diploid number difference. C-bands occurred in pericentromeric regions in some chromosomes, and a single 18S rDNA locus occurred in both species. The 5S rDNA showed variation in the number of loci between species karyotypes, suggesting the occurrence of unstable sites and rearrangements associated with these sequences in Ancistrus. The microsatellite mapping evidenced distinct patterns of organization between the two analyzed species, occurring mainly in the sex chromosomes in Ancistrus sp. 1, and in the centromeric and pericentromeric regions of chromosomes m/sm in Ancistrus sp. 2. These data shows the extensive chromosomal diversity of repetitive sequences in Ancistrus, which were involved in Robertsonian rearrangements and sex chromosomes differentiation.
Introduction
Loricariidae is one of the most specious neotropical fish families of the Siluriformes order, having 1,023 species (Fricke et al., 2021) distributed throughout Central and South America, from southeastern Costa Rica to northeastern Argentina (Armbruster, 2004; Armbruster, 2008; Armbruster and Lujan, 2016). Loricariidae is a monophyletic group organized into six subfamilies: Lithogeninae, Delturinae, Hypoptopomatinae, Neoplecostominae, Loricariinae, and Hypostominae (Roxo et al., 2019). The subfamily Hypostominae is the most speciose and widely distributed, organized into nine clades and tribes (Lujan et al., 2015).
The genus Ancistrus Kner, 1854 (Hypostominae, Ancistrini) occurs from Panama to Argentina, presenting 65 valid species, in addition to distinct lineages not formally identified in the scientific literature due to its taxonomic complexity (Ferraris, 2007; Armbruster, 2008; Prizon et al., 2018; Borba et al., 2019; Fricke et al., 2021). From a chromosomal point of view, Ancistrus represents one of the most diverse lineages of Loricariidae, emphasizing their extensive variation in the diploid number (2n = 34 to 54, Supplementary Table S1). The vast majority of Ancistrus species have karyotypes with 2n ≤ 52, probably due to the result of Robertsonian (Rb) fusions (see Glugoski et al., 2020), whose occurrence has been evidenced through the in situ localization of repetitive sequences (Barros et al., 2017). The presence of sex chromosomes is another striking feature of chromosomal diversity in Ancistrus, with species showing simple systems (XX/XY, XX/X0 and ZZ/ZW; Mariotto et al., 2004; Alves et al., 2006; Mariotto and Miyazawa, 2006; de Oliveira et al., 2007), multiple (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2; de Oliveira et al., 2008) or absence of differentiated sex chromosomes (Supplementary Table S1).
Sex chromosomes have emerged independently in different fish lineages, evolving through alternative mechanisms and showing various degrees of heteromorphism, even among closely related species (Charlesworth et al., 2005; Cioffi et al., 2010; Henning et al., 2011; Cioffi et al., 2012; Cioffi et al., 2013). Sex chromosomes in Ancistrus have been evidenced by size heteromorphism and accumulation of heterochromatic regions (Mariotto et al., 2004; Alves et al., 2006; Mariotto and Miyazawa, 2006; de Oliveira et al., 2007; de Oliveira et al., 2008; de Oliveira et al., 2009). Recently, in situ localization of repetitive sequences have provided insights into the differentiation of these chromosomes in several groups of fish, including Ancistrus (Cioffi and Bertollo, 2010; Schemberger et al., 2014; Cioffi et al., 2014; Favarato et al., 2017; Prizon et al., 2018; Schemberger et al., 2019).
Repetitive sequences represent the largest portion of eukaryotic genomes and may be organized in tandem repeats (e.g., microsatellites and multigene families, like ribosomal DNAs) or dispersed (e.g., transposons and retrotransposons). Ribosomal DNAs (rDNA) are represented by two gene families: 45S ribosomal RNA genes (18S, 5.8S and 28S genes), and 5S ribosomal RNA genes (Long and Dawid, 1980). The mapping of these sequences has shown intense variation in the location and number of chromosomal sites in Loricariidae. The participation of repetitive sequences, including rDNA and sequences microsatellites in chromosomal rearrangements has been evidenced, showing the importance of these markers in comparative analyzes (Pansonato-Alves et al., 2013; Barros et al., 2017; Primo et al., 2017; Bueno et al., 2018; Pety et al., 2018; Glugoski et al., 2020; Santos da Silva et al., 2021). Simple short tandem repeats of generally 1-6 nucleotides, known as,microsatellites, constitute another important tandem repeats group (Martins, 2007). The microsatellites are abundant in eukaryotic genomes, commonly a heterochromatin component, but they could also be found in euchromatic regions (Martins, 2007; Cioffi et al., 2010; Santos da Silva et al., 2021). They are helpful cytogenetic markers to demonstrate minor chromosomal variations into related species groups, including birds, amphibians, and fish (Cioffi et al., 2010; de Oliveira et al., 2017; Da Silva et al., 2021). Furthermore, the participation of these sequences in breakpoints regions for chromosomal rearrangements and evolution of sex chromosomes has been shown in many species, including Ancistrus (Farré et al., 2011; Cioffi et al., 2017; Favarato et al., 2017; Prizon et al., 2017).
In the present work, we studied the karyotypes of two not formally described Ancistrus species (Ancistrus sp. 1 and Ancistrus sp. 2) from the Amazon region, using different groups of repetitive sequences, in order to understand their mechanisms of chromosomal diversification.
Materials and Methods
Samples
Samples of two species of Ancistrus (Ancistrus sp. 1 and Ancistrus sp. 2) were analysed in this study. These species are morphologically different but are not yet described in the scientific literature. The samples were collected in distinct locations of the Tocantins-Araguaia River basin, in the Brazilian Amazon (Figure 1). Details about the collection points, number of individuals, and sex are presented in Table 1. The collection permit (number 13248) was issued by the Chico Mendes Institute for Biodiversity Conservation, Brazil. The Cytogenetics Laboratory of the Federal University of Pará had licenses for transport (number 19/2003) and the use of animals for this research (52/2003) as granted by the Ministry of the Environment. This study was approved by the Animal Ethics Committee of the Federal University of Pará (permission 68/2015). The specimens analyzed in this study were deposited in the Ichthyology Collection of the Center for Advanced Studies in Biodiversity (CEABIO/UFPA), Belém, Pará, Brazil.
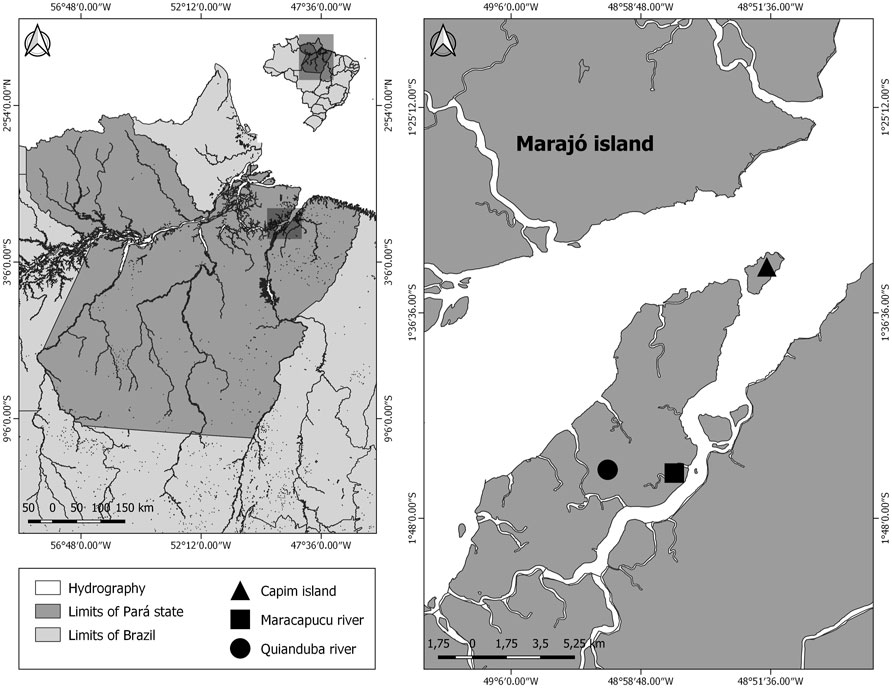
FIGURE 1. The geographical location of the collection points of specimens of Ancistrus species analyzed in this study. The map was built using Q-GIS version 3.4.5. The database was obtained from Instituto Brasileiro de Geografia e Estatística (IBGE).

TABLE 1. Sampling and collection sites of Ancistrus specimens analyzed in this study.
Chromosomal Analysis
Mitotic chromosomes were obtained from anterior kidney cells after in vivo treatment with colchicine solution (0.025%), as described by Bertollo et al. (1978). The animals were anesthetized with a eugenol solution (185 mg/L) (Vidal et al., 2008) and then sacrificed. Chromosomes were analyzed by classical staining (conventional staining with 5% Giemsa), C-banding (Sumner, 1972), and also molecular methods (Fluorescence in situ hybridization, FISH).
Probes Labeling and Fluorescence in situ Hybridization
Genomic DNA was extracted using the PureLink Genomic DNA Kit (Promega), following the manufacturer’s instructions. We used two rDNA sequences for in situ localization experiments: an 18S rDNA probe (1,400 bp segment) isolated from Ancistrus sp. 1 genomic DNA according described in (Gross et al., 2010), and a 5S rDNA (GenBank accession no. MT018470) probe obtained from Ancistrus aguaboensis (Glugoski et al., 2020). rDNA probes were labeled by nick-translation with biotin or digoxigenin. Telomeric probes were PCR labeled with digoxigeninin-11-dUTP (Roche Applid Science®) using primers F-5′(TTAGGG)5-3′ and R-5′(CCCTAA)5-3′ without using template DNA (Ijdo et al., 1991). All PCR products were checked on 1% agarose gel electrophoresis. The nine microsatellite probes ((CA)n, (CAC)n, (CAG)n, (CG)n, (GA)n, (CAT)n, (GAA)n, (GAC)n and (TAA)n) were purchased already with direct labeling by Cy3 during synthesis. FISH was performed following the protocol proposed by Martins and Galetti (1999), with modifications, under the following stringency conditions: 2.5 ng/μL of each probe, 50% formamide, 2 x SSC, 10% dextran sulfate, and hybridization at 42°C for 16 h. Fluorescence signals were detected using Streptavidin Alexa Fluor 488 (Molecular Probes, Carlsbad, CA, United States) and anti-digoxigenin rhodamine Fab fragments (Roche Applied Science, Penzberg, Germany). Chromosomes were counterstained with 0.2 μg/ml 4′6-diamidino-2-phenylindole (DAPI) in Vectashield mounting medium (Vector, Burlingame, CA, United States).
Image Capture and Analysis
Thirty metaphases per individual were analyzed to determine the diploid number, karyotype formula, and FISH experiments. Images of metaphases after Giemsa staining were obtained using an Olympus BX41 microscope (bright field) coupled to a CCD 1300QDS digital camera and analyzed using GenASIs ASI (Applied Spectral Imaging) software. FISH images were obtained using a Nikon H550S microscope and analyzed using Nis-Elements software. All images were adjusted using Adobe Photoshop CS6 software. The chromosome pairs were classified as metacentric (m), submetacentric (sm) and subtelocentric (st) following the criteria proposed by Levan et al. (1964). The count of the number of chromosome arms (Fundamental Number - FN) considered chromosomes m, sm and st as bi-armed.
Results
Classical Cytogenetics
Ancistrus sp. 1 demonstrated diploid chromosome number (2n), fundamental number (FN) and karyotype formula (KF) as follow: 2n = 38, FN = 72, KF = 20m + 14sm+2st. A heteromorphic chromosome pair was identified in males while the female karyotypes were homomorphic, characterizing a XX/XY sex chromosome system. The small subtelocentric Y chromosome and a medium X subtelocentric chromosome were recorded (Figures 2A,C). Constitutive heterochromatin (CH) occurred in a few regions in the karyotype, and was not evidenced in the sex chromosomes (Figures 2B,D).
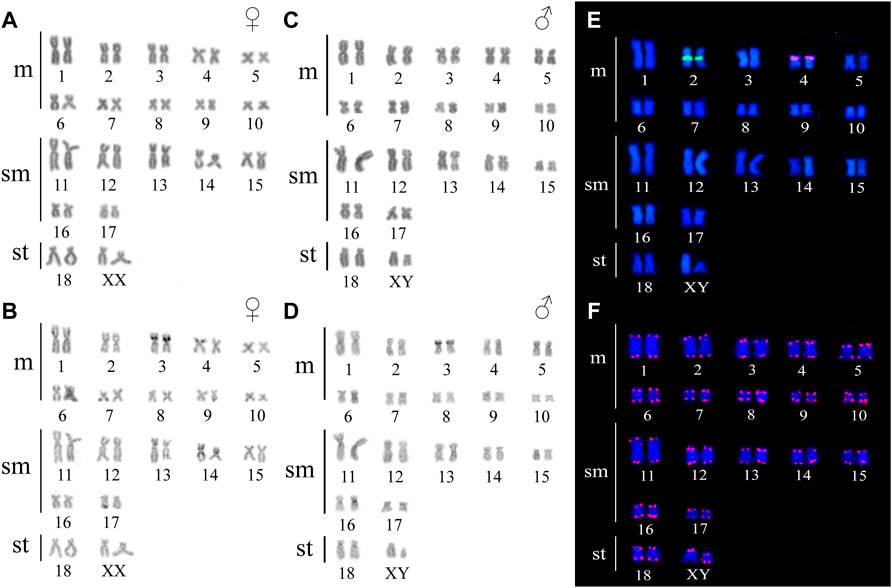
FIGURE 2. Karyotype of Ancistrus sp. 1. In (A,B) male karyotype stained with Giemsa and C-banding, respectively; in (C,D) female karyotype stained with Giemsa and C-banding, respectively; in (E) in situ localization of 18S rDNA (green) and 5S rDNA (red) sequences; in (F) in situ localization of telomeric sequences. The probes were labeled with FITC (green) and Cy3 (red), the chromosomes were counterstained with DAPI (blue).
Ancistrus sp. 2 demonstrated diploid chromosome number (2n), fundamental number (FN) and karyotype formula (KF) as follow: 2n = 34, FN = 68, KF = 20m + 14sm. Additionally no morphologically differentiated of sex chromosomes was found (Figure 3A). Some heterochromatic blocks are mainly distributed in the centromeric and pericentromeric region of chromosomes 1, 2, 3, 4, 5, 6, 7, 8, 13, and 16, besides conspicuous blocks in the short arm (p) distal region of the chromosome 3, coincident with the 18S rDNA sites (Figures 3B,C).
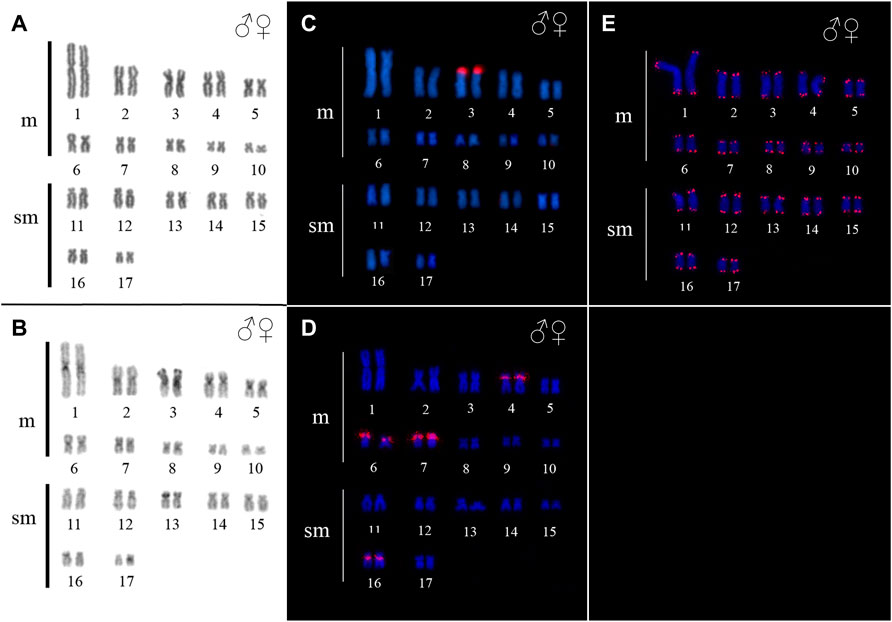
FIGURE 3. Karyotype of Ancistrus sp. 2. In (A,B) karyotype stained with Giemsa and C-banding, respectively; in (C–E) in situ location of 18S rDNA, 5S rDNA and telomeric sequences, respectively. The probes were labeled with Cy3 (red), the chromosomes were counterstained with DAPI (blue).
Molecular Cytogenetics
In Ancistrus sp. 1, the 18S rDNA site was located in the proximal region of the long arm (q) of pair 2, while the 5S rDNA occurred in the interstitial region of 4p (Figure 2E). In Ancistrus sp. 2, the 18S rDNA was located in the distal region of 3p (Figure 3C). The 5S rDNA demonstrated multiple sites located in the proximal region of the 4p and 6p, and pericentromeric in the 7p and 16q (Figure 3D). Interstitial telomeric sites (ITS) were not observed in any analyzed karyotypes (Figure 2F; Figure 3E).
The FISH results of microsatellite sequences in the karyotypes of the two Ancistrus species were summarized in Table 2. In Ancistrus sp. 1, the microsatellites (CAT)n, (GAA)n, (GAC)n and (TAA)n were located exclusively in the pericentromeric region of the X chromosome; (CG)n showed signs in the pericentromeric region of the Y chromosome and in the centromeric region of the pair 13 in individuals of both sexes (Figures 4A–E). Microsatellites (CA)n, (GA)n, (CAC)n and (CAG)n did not demonstrate any hybridization signal in the karyotype of these species (Table 2). In Ancistrus sp. 2, the same sets of microsatellites were observed in other pairs of chromosomes, mainly in centromeric and pericentromeric regions associated with heterochromatic regions (Figures 5A–H, Figure 6). Some microsatellites, including (CAC)n, (CAT)n, (CG)n, (GAA)n, (GAC)n, and (TAA)n was associated with heterochromatic regions and coincident or adjacent to 5S rDNA sites in the pairs 4, 6 and 7 (Figure 6). The microsatellite (CAG)n did not show any hybridization signal in both karyotypes (Table 2).
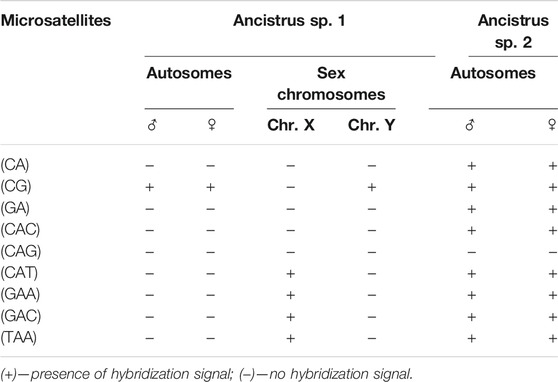
TABLE 2. Comparative analysis of the presence/absence and location of microsatellite sequences between the karyotypes of the Ancistrus species analyzed in this study.
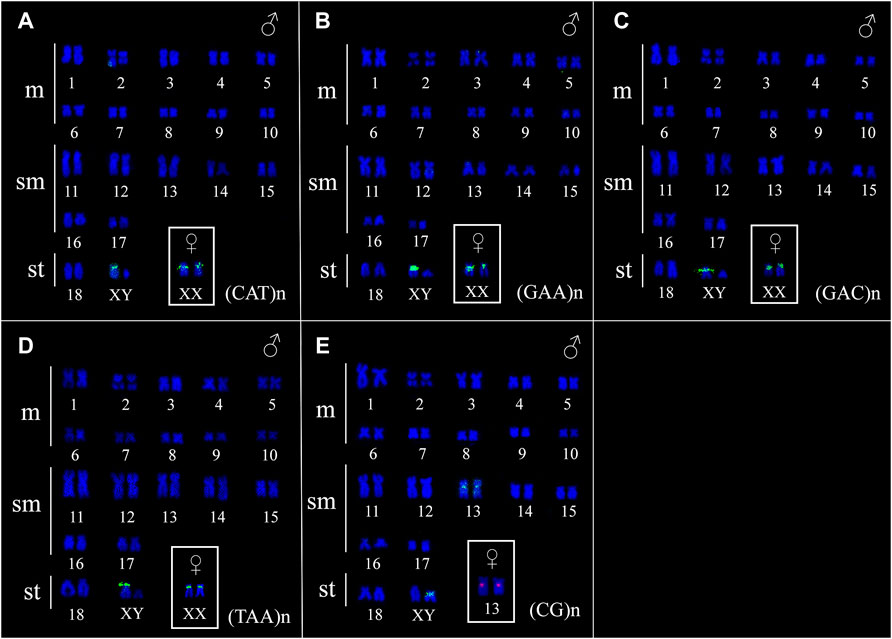
FIGURE 4. Fluorescence in situ hybridization indicating the physical location of microsatellite sequences in the karyotype of Ancistrus sp. 1: (A) (CAT)n, (B) (GAA)n, (C) (GAC)n, (D) (TAA)n, (E) (CG)n. Karyotypes of males are presented; female sex chromosomes are in the boxes. The probes were labeled with FITC (green) and with Cy3 (red), chromosomes were counterstained with DAPI (blue).
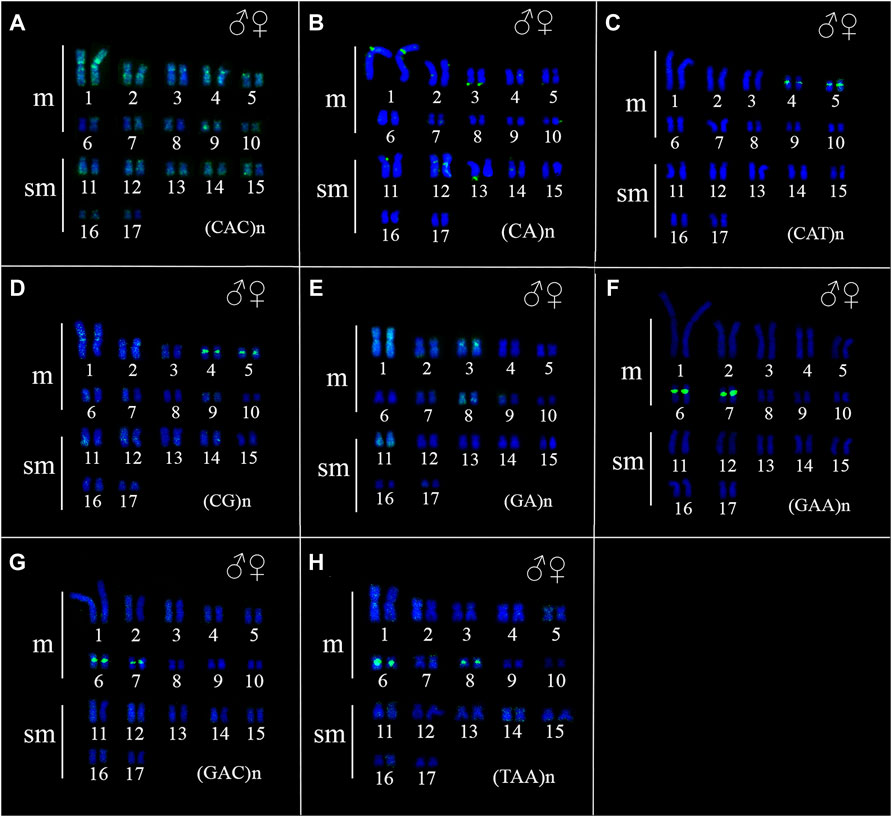
FIGURE 5. Fluorescence in situ hybridization indicating the physical location of microsatellite sequences in the karyotype of Ancistrus sp. 2: (A) (CAC)n, (B) (CA)n, (C) (CAT)n, (D) (CG)n, (E) (GA)n, (F) (GAA)n, (G) (GAC)n, (H) (TAA)n. The probes were labeled with FITC (green), the chromosomes were counterstained with DAPI (blue).
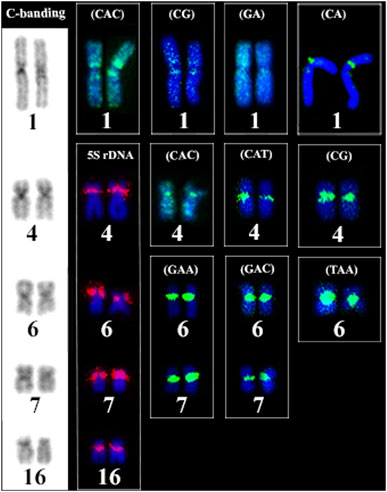
FIGURE 6. Comparative analysis of the distribution of heterochromatic regions, 5S rDNA sites and microsatellite sequences and in the karyotype of Ancistrus sp. 2. Boxes indicate the repetitive sequences in heterochromatic regions of chromosome pairs 1, 4, 6, 7 and 16 of Ancistrus sp. 2. Chromosomes stained with Giemsa; hybridization signals with FITC (green) and Cy3 (red), chromosomes were counterstained with DAPI (blue).
Discussion
Chromosomal Diversity in the Genus Ancistrus
The putative ancestral karyotype for Loricariidae presents 2n = 54, single nucleolus organizer regions (NOR) and few heterochromatic regions (Artoni and Bertollo, 2001; Ziemniczak et al., 2012). Nevertheless, considering the presence of 2n = 52 chromosomes in Pterygoplichthyini, the sister group for Ancistrini, Bueno et al. (2018) suggested that the putatively ancestral condition for Ancistrini is a diploid number of 52 chromosomes. Previously studies in Ancistrus have shown extensive chromosomal diversity with high variation in 2n values, the occurrence of multiple sites, as well as, 18S/5S rDNA synteny rupture (Supplementary Table S1). In this genus, species with lower 2n karyotypes have many chromosomes m/sm compared to those with higher 2n, which have more st/a chromosomes (Bueno et al., 2018; Glugoski et al., 2020). Thus, these findings suggests that chromosomal evolutions in this genus follows a tendency of 2n reduction due to the occurrence of Rb fusions (Barros et al., 2017).
Telomeric sequence mapping has shown the occurrence of ITS as a result of fusion events in Loricariidae (Barros et al., 2017; Primo et al., 2017). The species analyzed in this present study did not show the presence of ITS, which is in agreement with previous studies performed with other Ancistrus species (Prizon et al., 2018; Glugoski et al., 2020). The absence of ITS can be explained by the loss of these sequences during the fusion process (Slijepcevic, 1998).
Ribosomal genes have shown great diversity of location and number of sites among species and populations of different groups of fish (Gornung, 2013; Rebordinos et al., 2013). Mapping these sequences has revealed widely diverse chromosomal organizations in Ancistrus (Mariotto et al., 2011; Barros et al., 2017; Prizon et al., 2017; Prizon et al., 2018; Bueno et al., 2018; Glugoski et al., 2020). In Loricariidae, synteny between 18S/5S rDNA is considered a plesiomorphic character, with synteny break representing a derived state commonly observed in this group of fish (Bueno et al., 2018). This diversity of location and number of rDNA sites suggests the recurrent participation of these sequences in chromosomal reorganization events in Loricariidae (Pansonato-Alves et al., 2013; Prizon et al., 2016; Prizon et al., 2017; Barros et al., 2017; Prizon et al., 2018; Bueno et al., 2018; Santos da Silva et al., 2021).
Single 45S rDNA site represents a primitive character in Loricariidae (Artoni and Bertollo, 2001; Bueno et al., 2018). In the species described here and most of the analyzed Ancistrus species, these sequences are located in only one pair of chromosomes (see Supplementary Table S1), suggesting the maintenance of the primitive condition. Variations in the chromosomal position of these sequences were observed among the species in the present study, probably due to pericentric inversions.
Our results showed that the 5S rDNA was more dynamic than the 18S rDNA, varying in the number of sites among the analyzed karyotypes. This observation was consistent with previous observations performed in other Ancistrus species (Barros et al., 2017; Prizon et al., 2017; Glugoski et al., 2020; Supplementary Table S1). Studies have been showing that these sequences are involved in double-stranded DNA breaks and chromosomal rearrangements in Ancistrus (Mariotto et al., 2011; Favarato et al., 2016; Barros et al., 2017; Glugoski et al., 2020). Barros et al. (2017) demonstrated that the occurrence of multiple 5S rDNA sites is related to the emergence of pseudogenes in Ancistrus sp. (2n = 50). In addition, the involvement of 5S rRNA pseudogenes in Rb fusion events has been proposed in distinct genera of Loricariidae (Barros et al., 2017; Glugoski et al., 2018; Deon et al., 2020). Our results in Ancistrus sp. 2 showed multiple 5S rDNA sites located in the pericentromeric and proximal regions of some chromosome pairs. This data agrees with the hypothesis that 5S rDNA, or sequences derived from this gene family, may be involved in fusion events in Ancistrus sp. 2, as proposed previously for other species of this genus. Furthermore, these results supported the hypothesis that these sequences may represent evolutionary breakpoints regions (EBRs), which can be reused in chromosomal rearrangements in Ancistrus (Barros et al., 2017).
Comparative mapping of microsatellites revealed divergent patterns of organization between karyotypes of the Ancistrus species, occurring in euchromatic and heterochromatic regions in autosome and sex chromosomes. In Ancistrus sp. 2, the microsatellites mainly colonized heterochromatic blocks in centromeric and pericentromeric regions. Centromeric regions are characterized by the abundance of in tandem repeats, which are essential for maintaining the stability of this chromosomal region (Shang et al., 2010). However, the presence of different types of in tandem repeats, including microsatellites, at breakpoints for chromosomal rearrangements has been demonstrated previously (Kejnovsky et al., 2009; Cioffi and Bertollo, 2010; Ferré et al., 2011). The location of different microsatellites in the centromeric and pericentromeric region of metacentric chromosomes may indicate their association with EBRs (Ferré et al., 2011), suggesting the occurrence of Rb fusions during the evolution of the Ancistrus sp. 2. Furthermore, the association between microsatellites and rDNA sites has been observed in several organisms (Santos da Silva et al., 2021), corroborating to the chromosomal instability proposal to rDNA sites in Ancistrus. Therefore, the analyses carried out in this study suggest the participation of repetitive sequences in different mechanisms of chromosomal diversification in this group of neotropical fish.
Sex Chromosomes in Ancistrus
In general, sex chromosomes occur in only a small portion of neotropical fish species, having independent evolutionary origins and evolving from different mechanisms (Charlesworth et al., 2005; Henning et al., 2008; Henning et al., 2011; Schamberger et al., 2019). In Ancistrus, extensive sex chromosome diversity is described, with different levels of morphological differentiation and DNA content (Mariotto et al., 2004; Mariotto and Miyazawa, 2006; de Oliveira et al., 2007; de Oliveira et al., 2008; de Oliveira et al., 2009). Ancistrus sex chromosomes have been analyzed mainly according to their size, heteromorphisms, and distribution of heterochromatic regions (Mariotto et al., 2004; Mariotto and Miyazawa, 2006; de Oliveira et al., 2007; de Oliveira et al., 2008; de Oliveira et al., 2009). Theoretically, heterochromatinization has been considered an essential step in proto sex chromosome differentiation due to the differential accumulation of repetitive sequences and its effects in decreasing the recombination rate (Cioffi et al., 2012). Partially or fully heterochromatic sex chromosomes could be considered a characteristic of well-differentiated systems in fish, as noted in Eigenmannia, Tripohorteus, Characidium, and Parodontidae (Henning et al., 2011; Cioffi et al., 2014; Schemberger et al., 2014; Ziemniczak et al., 2014; Pucci et al., 2016). However, the absence of heterochromatic regions is a frequent state in the sex chromosomes in Ancistrus, including XX/XY or ZZ/ZW systems (de Oliveira et al., 2007; de Oliveira et al., 2009; present study), suggesting that the sex chromosomes in Ancistrus evolved independently and, therefore, are at different stages of differentiation regarding the accumulation of repetitive sequences and heterochromatinization.
In situ localization of repetitive sequences represent an important approach for studying sex chromosome diversity and evolution in fish (Cioffi et al., 2010; Schemberger et al., 2014; Schemberger et al., 2019). In Ancistrus, this approach has been applied in some species (Favarato et al., 2017; Prizon et al., 2018), indicating the participation of different repetitive sequences in the sex chromosome differentiation (Prizon et al., 2018). Here, the microsatellite comparative in situ localization mapping highlights differences between heteromorphic X and Y sex chromosomes in Ancistrus sp. 1. Usually, in XX/XY sex chromosome systems, the heteromorphic sex chromosome differentiates by accumulating repetitive sequences and heterochromatinization (Charlesworth et al., 2005). Furthermore, the Y chromosome generally follows through a degeneration pathway due to the absence of recombination, leading to its reduced size (Charlesworth et al., 2005). In this study, a higher concentration of microsatellite sequences were found in the X when compared to the Y chromosome; however, the presence of Y-speceific microsatellite sequences was also observed. These results can be explained, in part, by partial recombination restriction between the homologs of the proto sex pair promoted by the differential accumulation of repetitive sequences (Kejnovsky et al., 2009).
Sex chromosomes are commonly rich in distinct families of transposable elements (TE) and in tandem repeats (Charlesworth et al., 2005). The invasion of sex chromosomes by TE occurs at different stages of the differentiation of these chromosomes (Charlesworth et al., 2005; Schemberger et al., 2019). These sequences are inactivated or degenerated leading to heterochromatinization of parts of the sex chromosomes (Charlesworth et al., 2005). In Ancistrus sp. 1, C-banding demonstrated the absence of heterochromatin in the sex chromosomes, suggesting a recent stage of differentiation when compared to sex chromosomes from other fish groups (Cioffi et al., 2010; Henning et al., 2011; Cioffi et al., 2014; Schemberger et al., 2014; Schemberger et al., 2019). On the other hand, heteromorphic size suggests the occurrence of Y chromosome degeneration in Ancistrus sp. 1, as demonstrated by the different repetitive DNA content between X and Y-chromosomes. Future analyses integrating in situ location data from different repetitive units classes and epigenetic analyses will be important to test the condition of repetitive DNA segments that have not undergone heterochromatinization in Ancistrus sex chromosomes.
Conclusion
Our study provided additional evidence on the evolutionary pathways to 2n reduction in Ancistrus species, highlighting specific chromosomal features that have emerged throughout their life. The obtained data also suggest the participation of repetitive sequences acting in Ancistrus sp. 1 and Ancistrus sp. 2 diversification, as those sequences can be involved in the Robertsonian rearrangements and sex chromosomes differentiation.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by The Animal Ethics Committee of the Federal University of Pará.
Author Contributions
KS: Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Visualization; Writing of the original manuscript; Redaction; Revision; Edition. LG: Investigation; Methodology; Visualization; Redaction; Revision; Edition. MV: Methodology; Resources; Visualization; Redaction; Revision; Edition. AdS: Investigation; Methodology; Resources; Visualization; Redaction; Revision; Edition. RN: Investigation; Methodology; Resources; Visualization; Redaction; Revision; Edition. JP: Data curation; Formal analysis; Acquisition of financing; Resources; Visualization; Writing, Proofreading and Editing. CN: Data curation; Formal analysis; Acquisition of financing; Project administration; Resources; Supervision; Visualization; Redaction; Revision; Edition.
Funding
The authors thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for financial support on project coordinated by CN (Edital Pró-Amazônia Proc 047/2012); the FAPESPA for financial support (Edital Vale—Proc 2010/110447) and Banco Nacional de Desenvolvimento Econômico e Social—BNDES (2.318.697.0001) on a project coordinated by JP. CN (305880/2017-9), JP (305876/2017-1) and MV (305142/2019-4) are grateful to CNPq for Productivity Grants; the Pro-Reitoria de Pesquisa e Pos-Graduacao, Universidade Federal do Para (PROPESP—UFPA) for the payment of the article processing fee. This study is part of the Doctoral Thesis of KS in Genetic and Molecular Biology who is recipient of CAPES Doctor Scholarship.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Sample collections was authorized by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) and Secretaria de Estado de Meio Ambiente do Pará (SEMA-PA). The authors are grateful to members of the team of the cytogenetics laboratory UFPA for the fieldwork and chromosomal preparations. To MSc. Jorge Rissino, to MSc. Shirley Nascimento and Maria da Conceição for assistance in laboratory work. To David Santos da Silva for making the maps and helping with the fieldwork.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.838462/full#supplementary-material
References
Alves, A. L., Oliveira, C., Nirchio, M., Granado, Á., and Foresti, F. (2006). Karyotypic Relationships Among the Tribes of Hypostominae (Siluriformes: Loricariidae) with Description of XO Sex Chromosome System in a Neotropical Fish Species. Genetica 128 (1), 1–9. doi:10.1007/s10709-005-0715-1
Armbruster, J. W., and Lujan, N. K. (2016). A New Species of Peckoltia from the Upper Orinoco (Siluriformes, Loricariidae). ZooKeys 569, 105–121. doi:10.3897/zookeys.569.6630
Armbruster, J. W. (2004). Phylogenetic Relationships of the Suckermouth Armoured Catfishes (Loricariidae) with Emphasis on the Hypostominae and the Ancistrinaeshes Loricariidae with Emphasis on the Hypostominae and the Ancistrinae. Zoolog. J. Linn. Soc. 141 (1), 1–80. doi:10.1111/j.1096-3642.2004.00109.x
Armbruster, J. W. (2008). The Genus Peckoltia with the Description of Two New Species and a Reanalysis of the Phylogeny of the Genera of the Hypostominae (Siluriformes: Loricariidae). Zootaxa 1822 (1), 1–76. doi:10.11646/zootaxa.1822.1.1
Artoni, R. F., and Bertollo, L. A. (2001). Trends in the Karyotype Evolution of Loricariidae Fish (Siluriformes). Hereditas 134, 201–210. doi:10.1111/j.1601-5223.2001.00201.x
Barros, A. V., Wolski, M. A. V., Nogaroto, V., Almeida, M. C., Moreira-Filho, O., and Vicari, M. R. (2017). Fragile Sites, Dysfunctional Telomere and Chromosome Fusions: what Is 5S rDNA Role? Gene 608, 20–27. doi:10.1016/j.gene.2017.01.013
Bertollo, L. A. C., Takahashi, C. S., and Moreira-Filho, O. (1978). Cytotaxonomic Considerations on Hoplias Lacerdae (Pisces Erythrinidae). Braz. J. Genet. 1, 103–120.
de Borba, R. S., Mariotto, S., Centofante, L., Henrique Zawadzki, C., and Pasquali Parise-Maltempi, P. (2019). Molecular Discrimination of Ancistrus Lineages (Siluriformes: Loricariidae) Using Barcode DNA Tool. Mitochondrial DNA A 30 (4), 602–608. doi:10.1080/24701394.2019.1597071
Bueno, V., Konerat, J. T., Zawadzki, C. H., Venere, P. C., Blanco, D. R., and Margarido, V. P. (2018). Divergent Chromosome Evolution in Hypostominae Tribes (Siluriformes: Loricariidae): Correlation of Chromosomal Data with Morphological and Molecular Phylogenies. Zebrafish 15 (5), 492–503. doi:10.1089/zeb.2018.1612
Charlesworth, D., Charlesworth, B., and Marais, G. (2005). Steps in the Evolution of Heteromorphic Sex Chromosomes. Heredity 95 (2), 118–128. doi:10.1038/sj.hdy.6800697
Cioffi, M. B., and Bertollo, L. A. C. (2010). Initial Steps in XY Chromosome Differentiation in Hoplias malabaricus and the Origin of an X1X2Y Sex Chromosome System in This Fish Group. Heredity 105 (6), 554–561. doi:10.1038/hdy.2010.18
Cioffi, M. B., Martins, C., Vicari, M. R., Rebordinos, L., and Bertollo, L. A. C. (2010). Differentiation of the XY Sex Chromosomes in the Fish Hoplias malabaricus (Characiformes, Erythrinidae): Unusual Accumulation of Repetitive Sequences on the X Chromosome. Sex. Dev. 4 (3), 176–185. doi:10.1159/000309726
Cioffi, M. B., Moreira-Filho, O., Almeida-Toledo, L. F., and Bertollo, L. A. (2012). The Contrasting Role of Heterochromatin in the Differentiation of Sex Chromosomes: an Overview from Neotropical Fishes. J. Fish. Biol. 80 (6), 2125–2139. doi:10.1111/j.1095-8649.2012.03272.x
Cioffi, M. B., Liehr, T., Trifonov, V., Molina, W. F., and Bertollo, L. A. C. (2013). Independent Sex Chromosome Evolution in Lower Vertebrates: a Molecular Cytogenetic Overview in the Erythrinidae Fish Family. Cytogenet. Genome Res. 141 (2-3), 186–194. doi:10.1159/000354039
Cioffi, M., Fernanda Yano, C., Bertollo, L. A. C., Molina, W., and Liehr, T. (2014). Genomic Organization of Repetitive DNAs and its Implications for Male Karyotype and the Neo-Y Chromosome Differentiation in Erythrinus Erythrinus (Characiformes, Erythrinidae). Ccg 8 (2), 139–151. doi:10.3897/CompCytogen.v8i2.7597
Cioffi, M. d. B., Yano, C. F., Sember, A., and Bertollo, L. A. C. (2017). Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes 8 (10), 258. doi:10.3390/genes8100258
Da Silva, D. S., da Silva Filho, H. F., Cioffi, M. B., de Oliveira, E. H. C., and Gomes, A. J. B. (2021). Comparative Cytogenetics in Four Leptodactylus Species (Amphibia, Anura, Leptodactylidae): Evidence of Inner Chromosomal Diversification in Highly Conserved Karyotypes. Cytogenet. Genome Res. 161 (1-2), 52–62. doi:10.1159/000515831
De Oliveira, R. R., Feldberg, E., Anjos, M. B. d., and Zuanon, J. (2007). Karyotype Characterization and ZZ/ZW Sex Chromosome Heteromorphism in Two Species of the Catfish Genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon basin. Neotrop. Ichthyol. 5, 301–306. doi:10.1590/s1679-62252007000300010
De Oliveira, R. R., Feldberg, E., Dos Anjos, M. B., and Zuanon, J. (2008). Occurrence of Multiple Sexual Chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in Catfishes of the Genus Ancistrus (Siluriformes: Loricariidae) from the Amazon basin. Genetica 134 (2), 243–249. doi:10.1007/s10709-007-9231-9
De Oliveira, R. R., Feldberg, E., Dos Anjos, M. B., and Zuanon, J. (2009). Mechanisms of Chromosomal Evolution and its Possible Relation to Natural History Characteristics inAncistruscatfishes (Siluriformes: Loricariidae). J. Fish Biol. 75 (9), 2209–2225. doi:10.1111/j.1095-8649.2009.02450.x
De Oliveira, T. D., Kretschmer, R., Bertocchi, N. A., Degrandi, T. M., de Oliveira, E. H. C., Cioffi, M. d. B., et al. (2017). Genomic Organization of Repetitive DNA in Woodpeckers (Aves, Piciformes): Implications for Karyotype and ZW Sex Chromosome Differentiation. PLoS ONE 12 (1), e0169987. doi:10.1371/journal.pone.0169987
Deon, G. A., Glugoski, L., Vicari, M. R., Nogaroto, V., Sassi, F. d. M. C., Cioffi, M. d. B., et al. (2020). Highly Rearranged Karyotypes and Multiple Sex Chromosome Systems in Armored Catfishes from the Genus Harttia (Teleostei, Siluriformes). Genes 11 (11), 1366. doi:10.3390/genes11111366
Farré, M., Bosch, M., López-Giráldez, F., Ponsà, M., and Ruiz-Herrera, A. (2011). Assessing the Role of Tandem Repeats in Shaping the Genomic Architecture of Great Apes. PLoS ONE 6 (11), e27239. doi:10.1371/journal.pone.0027239
Favarato, R. M., da Silva, M., de Oliveira, R. R., Artoni, R. F., Feldberg, E., and Matoso, D. A. (2016). Cytogenetic Diversity and the Evolutionary Dynamics of rDNA Genes and Telomeric Sequences in the Ancistrus Genus (Loricariidae: Ancistrini). Zebrafish 13 (2), 103–111. doi:10.1089/zeb.2015.1140
Favarato, R. M., Ribeiro, L. B., Feldberg, E., and Matoso, D. A. (2017). Chromosomal Mapping of Transposable Elements of the rex Family in the Bristlenose Catfish, Ancistrus (Siluriformes, Loricariidae), from the Amazonian Region. Jhered 108 (3), esw084. doi:10.1093/jhered/esw084
Ferraris, C. J. (2007). Checklist of Catfishes, Recent and Fossil (Osteichthyes: Siluriformes), and Catalogue of Siluriform Primary Types. Zootaxa 1418, 1–628. doi:10.11646/zootaxa.1418.1.1
Fricke, R., Eschmeyer, W. N., and Van der Laan, R. (2021). ESCHMEYER'S CATALOG of FISHES: Genera, Species, References. Available at: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (Accessed October 27, 2021).
Glugoski, L., Giuliano-Caetano, L., Moreira-Filho, O., Vicari, M. R., and Nogaroto, V. (2018). Co-located hAT Transposable Element and 5S rDNA in an Interstitial Telomeric Sequence Suggest the Formation of Robertsonian Fusion in Armored Catfish. Gene 650, 49–54. doi:10.1016/j.gene.2018.01.099
Glugoski, L., Deon, G., Schott, S., Vicari, M. R., Nogaroto, V., and Moreira-Filho, O. (2020). Comparative Cytogenetic Analyses in Ancistrus Species (Siluriformes: Loricariidae). Neotrop. Ichthyol. 18 (2), 1–16. doi:10.1590/1982-0224-2020-0013
Gornung, E. (2013). Twenty Years of Physical Mapping of Major Ribosomal RNA Genes across the Teleosts: a Review of Research. Cytogenet. Genome Res. 141 (2-3), 90–102. doi:10.1159/000354832
Gross, M. C., Schneider, C. H., Valente, G. T., Martins, C., and Feldberg, E. (2010). Variability of 18S rDNA Locus amongSymphysodonfishes: Chromosomal Rearrangements. J. Fish Biol. 76 (5), 1117–1127. doi:10.1111/j.1095-8649.2010.02550.x
Henning, F., Trifonov, V., Ferguson-Smith, M. A., and de Almeida-Toledo, L. F. (2008). Non-homologous Sex Chromosomes in Two Species of the Genus Eigenmannia (Teleostei: Gymnotiformes). Cytogenet. Genome Res. 121, 55–58. doi:10.1159/000124382
Henning, F., Moysés, C. B., Calcagnotto, D., Meyer, A., and de Almeida-Toledo, L. F. (2011). Independent Fusions and Recent Origins of Sex Chromosomes in the Evolution and Diversification of Glass Knife Fishes (Eigenmannia). Heredity 106 (2), 391–400. doi:10.1038/hdy.2010.82
Ijdo, J. W., Wells, R. A., Baldini, A., and Reeders, S. T. (1991). Improved Telomere Detection Using a Telomere Repeat Probe (TTAGGG)ngenerated by PCR. Nucl. Acids Res. 19 (17), 4780. doi:10.1093/nar/19.17.4780
Kejnovsky, E., Hobza, R., Cermak, T., Kubat, Z., and Vyskot, B. (2009). The Role of Repetitive DNA in Structure and Evolution of Sex Chromosomes in Plants. Heredity 102 (6), 533–541. doi:10.1038/hdy.2009.17
Levan, A., Fredga, K., and Sandberg, A. A. (1964). Nomenclature for Centromeric Position on Chromosomes. Hereditas 52 (2), 201–220. doi:10.1111/j.1601-5223.1964.tb01953.x
Long, E. O., and Dawid, I. B. (1980). Repeated Genes in Eukaryotes. Annu. Rev. Biochem. 49 (1), 727–764. doi:10.1146/annurev.bi.49.070180.003455
Lujan, N. K., Armbruster, J. W., Lovejoy, N. R., and López-Fernández, H. (2015). Multilocus Molecular Phylogeny of the Suckermouth Armored Catfishes (Siluriformes: Loricariidae) with a Focus on Subfamily Hypostominae. Mol. Phylogenet. Evol. 82, 269–288. doi:10.1016/j.ympev.2014.08.020
Mariotto, S., and Miyazawa, C. S. (2006). Ancistrus Cf. Dubius (Siluriformes, Ancistrinae), a Complex of Species. 1. Chromosomic Characterization of Four Populations and Occurence of Sexual Chromosomes of Type XX/XY, in the Pantanal basin of Mato Grosso, Brazil. Caryologia 59 (4), 299–304. doi:10.1080/00087114.2006.10797929
Sandra, M., Artoni, R. F., and Miyazawa, C. S. (2004). Occurrence of Sexual Chromosome, of the Type ZZ/ZW, in Ancistrus Cf. Dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, BrazilAncistrus Cf. Dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, Brazil. Caryologia 57 (4), 327–331. doi:10.1080/00087114.2004.10589413
Mariotto, S., Centofante, L., Vicari, M., Artoni, R., and Moreira Filho, O. (2011). Chromosomal Diversification in Ribosomal DNA Sites in Ancistrus Kner, 1854 (Loricariidae, Ancistrini) from Three Hydrographic Basins of Mato Grosso, Brazil. Ccg 5 (4), 289–300. doi:10.3897/compcytogen.v5i4.1757
Martins, C., and Galetti Jr, P. M. (1999). Chromosomal Localization of 5S rDNA Genes in Leporinus Fish (Anostomidae, Characiformes). Chromosome Res. 7 (5), 363–367. doi:10.1023/a:1009216030316
Martins, C. (2007). “Chromosomes and Repetitive DNAs: A Contribution to the Knowledge of the Fish Genome,” in Fish Cytogenetics. Editors E. Pisano, C. Ozouf-Costaz, F. Foresti, and B. G. Kapoor (Enfield: Science Publishers), 421–453. doi:10.1201/b10746-16
Pansonato-Alves, J. C., Serrano, É. A., Utsunomia, R., Scacchetti, P. C., Oliveira, C., and Foresti, F. (2013). Mapping Five Repetitive DNA Classes in Sympatric Species of Hypostomus (Teleostei: Siluriformes: Loricariidae): Analysis of Chromosomal Variability. Rev. Fish. Biol. Fish. 23 (4), 477–489. doi:10.1007/s11160-013-9303-0
Pety, A. M., Cardoso, A. L., Nagamachi, C. Y., Pieczarka, J. C., de Sousa, L. M., and Noronha, R. C. R. (2018). In Situ localization of Ribosomal Sites in Peckoltia and Ancistomus (Loricariidae: Hypostominae) from the Amazon Basin. Zebrafish 15 (3), 263–269. doi:10.1089/zeb.2017.1523
Primo, C. C., Glugoski, L., Almeida, M. C., Zawadzki, C. H., Moreira-Filho, O., Vicari, M. R., et al. (2017). Mechanisms of Chromosomal Diversification in Species of Rineloricaria (Actinopterygii: Siluriformes: Loricariidae). Zebrafish 14 (2), 161–168. doi:10.1089/zeb.2016.1386
Prizon, A. C., Borin-Carvalho, L. A., Bruschi, D. P., Otávio Ribeiro, M., Magrinelli Barbosa, L., de Brito Ferreira, G. E., et al. (2016). Cytogenetic Data on Ancistrus Sp. (Siluriformes, Loricariidae) of the Paraguay River basin (MS) Sheds Light on Intrageneric Karyotype Diversification. Ccg 10 (4), 625–636. doi:10.3897/CompCytogen.v10i4.8532
Prizon, A. C., Bruschi, D. P., Borin-Carvalho, L. A., Cius, A., Barbosa, L. M., Ruiz, H. B., et al. (2017). Hidden Diversity in the Populations of the Armored Catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River Basin Revealed by Molecular and Cytogenetic Data. Front. Genet. 8, 185. doi:10.3389/fgene.2017.00185
Prizon, A. C., Bruschi, D. P., Gazolla, C. B., Borin-Carvalho, L. A., and Portela-Castro, A. L. D. B. (2018). Chromosome Spreading of the Retrotransposable Rex-3 Element and Microsatellite Repeats in Karyotypes of the Ancistrus Populations. Zebrafish 15 (5), 504–514. doi:10.1089/zeb.2018.1620
Pucci, M. B., Barbosa, P., Nogaroto, V., Almeida, M. C., Artoni, R. F., Scacchetti, P. C., et al. (2016). Chromosomal Spreading of Microsatellites and (TTAGGG)n Sequences in the Characidium Zebra and C. Gomesi Genomes (Characiformes: Crenuchidae). Cytogenet. Genome Res. 149, 182–190. doi:10.1159/000447959
Rebordinos, L., Cross, I., and Merlo, A. (2013). High Evolutionary Dynamism in 5S rDNA of Fish: State of the Art. Cytogenet. Genome Res. 141 (2-3), 103–113. doi:10.1159/000354871
Roxo, F. F., Ochoa, L. E., Sabaj, M. H., Lujan, N. K., Covain, R., Silva, G. S. C., et al. (2019). Phylogenomic Reappraisal of the Neotropical Catfish Family Loricariidae (Teleostei: Siluriformes) Using Ultraconserved Elements. Mol. Phylogenet. Evol. 135, 148–165. doi:10.1016/j.ympev.2019.02.017
Santos da Silva, K., de Souza, A. C. P., Pety, A. M., Noronha, R. C. R., Vicari, M. R., Pieczarka, J. C., et al. (2021). Comparative Cytogenetics Analysis Among Peckoltia Species (Siluriformes, Loricariidae): Insights on Karyotype Evolution and Biogeography in the Amazon Region. Front. Genet. 12, 1–9. doi:10.3389/fgene.2021.779464
Schemberger, M. O., Oliveira, J. I. N., Nogaroto, V., Almeida, M. C., Artoni, R. F., Cestari, M. M., et al. (2014). Construction and Characterization of a Repetitive DNA Library in Parodontidae (Actinopterygii: Characiformes): A Genomic and Evolutionary Approach to the Degeneration of the W Sex Chromosome. Zebrafish 11 (6), 518–527. doi:10.1089/zeb.2014.1013
Schemberger, M. O., Nascimento, V. D., Coan, R., Ramos, É., Nogaroto, V., Ziemniczak, K., et al. (2019). DNA Transposon Invasion and Microsatellite Accumulation Guide W Chromosome Differentiation in a Neotropical Fish Genome. Chromosoma 128 (4), 547–560. doi:10.1007/s00412-019-00721-9
Shang, W.-H., Hori, T., Toyoda, A., Kato, J., Popendorf, K., Sakakibara, Y., et al. (2010). Chickens Possess Centromeres with Both Extended Tandem Repeats and Short Non-tandem-repetitive Sequences. Genome Res. 20 (9), 1219–1228. doi:10.1101/gr.106245.110
Slijepcevic, P. (1998). Telomeres and Mechanisms of Robertsonian Fusion. Chromosoma 107, 136–140. doi:10.1007/s004120050289
Sumner, A. T. (1972). A Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cel Res. 75, 304–306. doi:10.1016/0014-4827(72)90558-7
Vidal, L. V. O., Albinati, R. C. B., Albinati, A. C. L., Lira, A. D. D., Almeida, T. R. D., and Santos, G. B. (2008). Eugenol como anestésico para a tilápia-Do-nilo. Pesq. Agropec. Bras. 43, 1069–1074. doi:10.1590/s0100-204x2008000800017
Ziemniczak, K., Barros, A. V., Rosa, K. O., Nogaroto, V., Almeida, M. C., Cestari, M. M., et al. (2012). Comparative Cytogenetics of Loricariidae (Actinopterygii: Siluriformes): Emphasis in Neoplecostominae and Hypoptopomatinae. Ital. J. Zoolog. 79 (4), 492–501. doi:10.1080/11250003.2012.676677
Ziemniczak, K., Traldi, J. B., Nogaroto, V., Almeida, M. C., Artoni, R. F., Moreira-Filho, O., et al. (2014). In Situ Localization of (GATA)n and (TTAGGG)n Repeated DNAs and W Sex Chromosome Differentiation in Parodontidae (Actinopterygii: Characiformes). Cytogenet. Genome Res. 144, 325–332. doi:10.1159/000370297
Keywords: amazon fish, comparative cytogenetics, microsatellites, repetitive DNA, rDNA, sex chromosomes
Citation: Santos da Silva K, Glugoski L, Vicari MR, de Souza ACP, Noronha RCR, Pieczarka JC and Nagamachi CY (2022) Chromosomal Diversification in Ancistrus Species (Siluriformes: Loricariidae) Inferred From Repetitive Sequence Analysis. Front. Genet. 13:838462. doi: 10.3389/fgene.2022.838462
Received: 17 December 2021; Accepted: 25 February 2022;
Published: 24 March 2022.
Edited by:
Caroline Gomes Lucas, University of Missouri, United StatesReviewed by:
Vito Antonio Mastrochirico Filho, Universidade Estadual Paulista, BrazilCamila U. Braz, University of Wisconsin-Madison, United States
Copyright © 2022 Santos da Silva, Glugoski, Vicari, de Souza, Noronha, Pieczarka and Nagamachi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cleusa Yoshiko Nagamachi, Y2xldXNhQHVmcGEuYnI=, Y2xldXNhbmFnYW1hY2hpQGdtYWlsLmNvbQ==