
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 04 October 2022
Sec. Epigenomics and Epigenetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.1013239
This article is part of the Research Topic Genetics and Epigenetics of Insect-Pathogen Interaction View all 4 articles
 Zhiwei Zhu1†
Zhiwei Zhu1† Jie Wang1†
Jie Wang1† Xiaoxue Fan1Qi Long1Huazhi Chen1Yaping Ye1Kaiyao Zhang1Zhongmin Ren1Yang Zhang1Qingsheng Niu3Dafu Chen1,2
Xiaoxue Fan1Qi Long1Huazhi Chen1Yaping Ye1Kaiyao Zhang1Zhongmin Ren1Yang Zhang1Qingsheng Niu3Dafu Chen1,2 Rui Guo1,2*
Rui Guo1,2*Nosema ceranae is a widespread fungal parasite for honey bees, causing bee nosemosis. Based on deep sequencing and bioinformatics, identification of circular RNAs (circRNAs) in Apis cerana workers’ midguts and circRNA-regulated immune response of host to N. ceranae invasion were conducted in this current work, followed by molecular verification of back-splicing sites and expression trends of circRNAs. Here, 10185 and 7405 circRNAs were identified in the midguts of workers at 7 days (AcT1) and 10 days (AcT2) post inoculation days post-inoculation with N. ceranae. PCR amplification result verified the back-splicing sites within three specific circRNAs (novel_circ_005123, novel_circ_007177, and novel_circ_015140) expressed in N. ceranae-inoculated midgut. In combination with transcriptome data from corresponding un-inoculated midguts (AcCK1 and AcCK2), 2266 circRNAs were found to be shared by the aforementioned four groups, whereas the numbers of specific ones were 2618, 1917, 5691, and 3723 respectively. Further, 83 52) differentially expressed circRNAs (DEcircRNAs) were identified in AcCK1 vs. AcT1 (AcCK2 vs. AcT2) comparison group. Source genes of DEcircRNAs in workers’ midgut at seven dpi were involved in two cellular immune-related pathways such as endocytosis and ubiquitin mediated proteolysis. Additionally, competing endogenous RNA (ceRNA) network analysis showed that 23 13) DEcircRNAs in AcCK1 vs. AcT1 (AcCK2 vs. AcT2) comparison group could target 18 14) miRNAs and further link to 1111 (1093) mRNAs. These target mRNAs were annotated to six cellular immunity pathways including endocytosis, lysosome, phagosome, ubiquitin mediated proteolysis, metabolism of xenobiotics by cytochrome P450, and insect hormone biosynthesis. Moreover, 284 164) internal ribosome entry site and 54 26) ORFs were identified from DEcircRNAs in AcCK1 vs. AcT1 (AcCK2 vs. AcT2) comparison group; additionally, ORFs in DEcircRNAs in midgut at seven dpi with N. ceranae were associated with several cellular immune pathways including endocytosis and ubiquitin-mediated proteolysis. Ultimately, RT-qPCR results showed that the expression trends of six DEcircRNAs were consistent with those in transcriptome data. These results demonstrated that N. ceranae altered the expression pattern of circRNAs in A. c. cerana workers’ midguts, and DEcircRNAs were likely to regulate host cellular and humoral immune response to microsporidian infection. Our findings lay a foundation for clarifying the mechanism underlying host immune response to N. ceranae infection and provide a new insight into interaction between Asian honey bee and microsporidian.
As a novel player in the world of non-coding RNA (ncRNA), circular RNA (circRNA) has become a worldwide research hotspot. Different from canonical alternative splicing, circRNA is generated by the back-splicing of pre-mRNA (Li et al., 2018). In comparison with linear RNA, circRNA is more resistant to RNase R enzyme digestion owing to its special covalently closed-loop structure; hence, circRNA is regarded as an ideal endogenous biomarker (Meng et al., 2017). CircRNAs are abundant in eukaryotic cells and have versatile functions such as the regulation of source gene transcription (Li et al., 2015), absorption of miRNAs or RNA binding proteins as “molecular sponges” (Han et al., 2020), and translation into peptides or proteins (Wang C. et al., 2020). Accumulating evidence suggests that circRNAs are involved in the occurrence and development of human disease, such as cervical (Chen R. X. et al., 2019) and lung cancers (Wang Z. Y. et al., 2020). The biological function of circRNAs as competing endogenous RNAs (ceRNAs) has only been deeply studied in humans and a few other model species (Han et al., 2020; Li et al., 2020). For example, Li et al. (2020) reported that circTLK1 was highly expressed in renal cell carcinoma (RCC) and could promote RCC progression through the miR-136–5p/CBX4 pathway, and circTLK1 could serve as a diagnostic molecules and therapeutic targets for renal cancer. However, research on insect circRNAs are limited, and the few studies have mainly focused on Drosophila melanogaster (Westholm et al., 2014; Huang C. et al., 2018; Krishnamoorthy and Kadener, 2021), Bombyx mori (Wang and Wang, 2015; Gan et al., 2017; Hu et al., 2018a; Hu et al., 2018b), and honey bees (Chen X. et al., 2019; Thölken et al., 2019; Chen D. F. et al., 2020). Westholm’s group predicted more than 2 500 circRNAs using transcriptome data from Drosophila, and revealed that circRNAs were abundantly expressed in the brain and accumulated over time (Westholm et al., 2014). Wang C. et al. (2020) discovered that Bombyx mori CircEgg was mainly located in the cytoplasm and circEgg overexpression inhibited the production of linear transcripts of BmEgg, circEgg inhibited methylation of histone H3 lysine nine by acting as a “sponge” of bmo-miR-3391–5p. Our group previously conducted a comprehensive investigation of circRNAs in the midguts of European honey bee Apis mellifera and Asian honey bee Apis cerana. Chen et al. identified 1101 circRNAs in Apis mellifera ligustica workers using a combination of RNA-seq and bioinformatics (Chen D. F. et al., 2020), Xiong et al. analyzed the expression profile of circRNAs during the developmental process of Apis cerana workers’ midguts, and uncovered the potential regulatory role of differentially expressed circRNAs (DEcircRNAs) (Xiong et al., 2018).
Nosema ceranae, an emergent fungal parasite, infects not only the midgut epithelial cells of adult bees but also bee larvae (Huang and Evans, 2020). It was first identified in Apis cerana colonies reared in China (Fries et al., 1996) and then spread to A. mellifera colonies in Europe (Higes et al., 2010), America (Emsen et al., 2020), and other parts of the world (Giersch et al., 2009). N. ceranae spores enter the midgut of the bee host through ingestion of contaminated food or water and then germinate due to activation by the special physical and chemical conditions inside the midgut. The infective sporoplasm is injected into the host midgut epithelial cell and replicates by stealing host material and energy. As the quantity of spores increases, the host cell finally ruptures, and the released spores in the feces become new sources of infection via the feeding and cleaning activities inside the colonies, or are disseminated into the environment (Higes et al., 2007; Gisder et al., 2011). N. ceranae infection has a negative influence on bee hosts, such as midgut epithelial cell damage, energy stress, immunosuppression, cell apoptosis inhibition, and lifespan reduction (Antúnez et al., 2009; Mayack and Naug, 2009; Goblirsch et al., 2013; Kurze et al., 2018; Panek et al., 2018). Additionally, N. ceranae infection severely weakens health and productivity of bee colonies in conjunction with other biological or environmental stresses (Doublet et al., 2015). A. c. cerana, a subspecies of A. cerana, is mainly distributed and widely used in Asian countries including China. Compared with A. mellifera, A. cerana is more adaptive to extreme weather conditions and is good at collecting scattered nectar sources (Zhao et al., 2020). Additionally, A. c. cerana has been used as a model for investigating host‒pathogen interactions (Huang S. K. et al., 2018). The reference genome of A. cerana (Park et al., 2015) was published in 2015, much later than the A. mellifera genome (Honeybee Genome Sequencing Consortium, 2006). Currently, study on the omics and molecular biology of A. cerana is lagging when compared with those of A. mellifera, and the interaction between A. cerana and parasites or pathogens is still largely unknown. Previously, our team investigated the immune response of A. c. cerana workers to N. ceranae infection (Xing et al., 2021), and deciphered the differential expression profile of host miRNAs during microsporidian infection and DEmiRNA-regulated host response (Chen D. F. et al., 2019).
CircRNAs have been identified in both A. c. cerana (Chen D. F. et al., 2020) and N. ceranae (Guo et al., 2018b) by our group. CircRNAs have been suggested to be crucial regulators engaged in host-pathogen interactions. However, research on the interaction between Asian honey bees and N. ceranae is still lacking. Our group previously conducted deep sequencing of A. c. ceranae workers’ midgut tissues at 7 days post-inoculation (dpi) and 10 dpi with N. ceranae (AcT1 and AcT2 groups) and corresponding un-inoculated midgut tissues (AcCK1 and AcCK2 groups), and identified 9589 circRNAs using transcriptome data from un-inoculated groups (Chen D. F. et al., 2020). Here, to unclose the circRNA-regulated responses of Asian honey bee workers to N. ceranae infection, utilizing the obtained high-quality transcriptome data, the differential expression pattern of circRNAs in A. c. cerana workers’ midguts in response to N. ceranae invasion was analyzed followed by an in-depth investigation of the host response mediated by DEcircRNAs, with a focus on cellular and humoral immune responses. To the best of our knowledge, this is the first documentation of a circRNA-regulated response of Asian honey bee to microsporidian invasion. The findings in this current work can not only lay a key foundation for clarifying the underlying mechanism but also provide novel insights into Asian honey bee-microsporidian interactions.
Three A. c. cerana colonies located in the teaching apiary of the College of Animal Sciences (College of Bee Science) in Fujian Agriculture and Forestry University were used for this study. Microscopic observations and PCR identification verified that these colonies were N. ceranae-free. Varroa was not observed before and during the experiment. RT-PCR was conducted to detect the prevalence of seven common bee viruses (DWV, KBV, ABPV, CBPV, IAPV, SBV, and BQCV) and two bee microsporidia (Nosema apis and N. ceranae) in the newly emergent workers based on previously described specific primers (Stoltz et al., 1995; Benjeddou et al., 2001; Ribiere et al., 2002; Genersch, 2005; Chen et al., 2008; Singh et al., 2010; Chen D. F. et al., 2019), and agarose gel electrophoresis (AGE) indicated that no bands specific for the aforementioned viruses and microsporidia were amplified (Chen D. F. et al., 2019; Xing et al., 2021).
Foragers were collected from a N. ceranae-infected colony in an apiary in Fuzhou city, Fujian Province, China. N. ceranae spores were previously prepared using the Percoll discontinuous centrifugation method by our group (Chen D. F. et al., 2019).
Midgut tissues of N. ceranae-inoculated A. c. cerana workers at seven dpi and 10 dpi and corresponding un-inoculated workers’ midguts were previously prepared by our team (Xing et al., 2021). There were three biological replicates for each treatment group or control group. RNA isolation, cDNA library construction, deep sequencing, and data quality control were previously conducted (Xing et al., 2021). The 12 constructed cDNA libraries were sequenced on an Illumina HiSeq 4000 platform (Illumina). Raw data are available in the NCBI Short Read Archive database (http://www.ncbi.nlm.nih.gov/sra/) under BioProject number: PRJNA406998. In total, 1,809,736,786 raw reads were produced from RNA-seq and 1,562,162,742 clean reads were obtained after quality control. The mean Q20 values of clean reads from the control groups and treatment groups were 94.76% and 94.77%, respectively, and the mapping ratios of clean reads to the reference genome of A. cerana were 75.78% (AcCK1), 55.01% (AcCK2), 78.13% (AcT1) and 44.19% (AcT2) (Chen D. F. et al., 2020; Fu et al., 2020). High-quality strand-specific cDNA library-based RNA-seq data could be utilized for circRNA identification and DEcircRNA investigation in this work.
In another previous study, midgut tissues of N. ceranae-inoculated A. c. cerana workers at seven dpi and 10 dpi and corresponding un-inoculated workers’ midguts were prepared by our team (Chen D. F. et al., 2019). Three biological replicates were performed for each treatment group or control group. RNA extraction, cDNA library construction, sRNA-seq, and data quality control were previously performed (Chen D. F. et al., 2019). The 12 constructed cDNA libraries were subjected to sequencing on an Illumina MiSeq platform with the single-end strategy. A total of 127,523,419 raw reads were generated from the sRNA-seq data, and 122,104,443 clean reads were obtained after quality control. The Pearson correlation between every replica in each group was above 0.9619 (Chen D. F. et al., 2019). Thus, the high-quality sRNA-seq data could be used for target prediction and regulatory network construction of DEcircRNAs in this study.
CircRNAs in the AcCK1 and AcCK2 groups were identified in a previous study (Chen D. F. et al., 2020). In this work, circRNAs in the AcT1 and AcT2 groups were identified following our previously described method (Chen D. F. et al., 2020). Briefly, firstly, clean reads were mapped to the A. cerana reference genome (assembly ACSNU-2.0) using TopHat software (Kim et al., 2013); secondly, 20 nt at both ends of unmapped reads were then extracted and independently mapped to the reference genome; thirdly, the mapped anchor reads were submitted to find_circ software (Memczak et al., 2013) to perform circRNA identification according to the following criteria: circRNA length <100 kb, best qual A > 35 or best qual B > 35, anchor overlap ≤2, n uniq >2, edit ≤2, n uniq > int (samples/2), and breakpoints = 1.
The expression level of each circRNA was normalized to the mapped back-splicing junction reads per million (RPM) mapped-reads value. Following the threshold |FC (fold change)| ≥ 2, p value <0.05, and false discovery rate (FDR) ≤ 1, DEcircRNAs in the AcCK1 vs AcT1 and AcCK2 vs AcT2 comparison groups were identified using DESeq software (Wang et al., 2010).
CircRNAs can regulate the expression of source genes via interactions with RNA polymerase II, U1 ribonucleoprotein, and gene promoters (Zhang et al., 2013; Li et al., 2015). According to the method described by Chen D. F. et al (2020). The source genes of DEcircRNAs were predicted by mapping the anchor reads at both ends of DEcircRNAs to the A. cerana reference genome (assembly ACSNU-2.0) using Bowtie software. Gene Ontology (GO) term analysis of the circRNAs’ source genes was conducted with the DAVID tool (http://david.abcc.ncifcrf.gov/) (Huang et al., 2007), and the GO categories were clarified using a two-sided Fisher’s exact test, while the FDR was calculated to correct the p value (Jung, 2014). A Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed according to the annotation of the KEGG database (http://www.genome.jp/kegg/) (Du et al., 2014).
In combination with previously identified miRNAs based on sRNA-seq data (Chen D. F. et al., 2019), target miRNAs of DEcircRNAs were predicted following a previously described protocol by Chen D. F. et al. (2020). Briefly, according to the criteria of p ≤ 0.05 and free energy ≤ −35 kcal/mol, potential target miRNAs of DEcircRNAs were predicted using three software programs, namely, mireap, miranda (v3.3a) and TargetScan (version: 7.0), followed by construction of the DEcircRNA-miRNA regulatory network; subsequently, target mRNAs of DEcircRNA-targeted miRNAs were further predicted, and then the DEcircRNA-miRNA-mRNA regulatory network was constructed; finally, the regulatory networks were visualized by Cytoscape software (Smoot et al., 2011) with the default parameters. GO term and KEGG pathway investigation of the target mRNAs was further conducted using the abovementioned methods.
Three circRNAs (novel_circ_005123, novel_circ_007177, and novel_circ_015140) shared by the AcT1 and AcT2 groups were randomly selected for molecular verification. Following our previously described method (Chen D. F. et al., 2020), divergent primers for these circRNAs (Table 1) were designed using DNAMAN eight software (Lynnon Biosoft, United States) and then synthesized by Shanghai Sangon Biological Co., Ltd. Total RNA of N. ceranae-inoculated workers’ midguts at seven dpi and 10 dpi was extracted with an AxyPre RNA extraction kit (Axygen, China), and then digested with 3 U/mg RNase R (Geneseed, China) at 37°C for 15 min to remove linear RNA. Next, the first-strand cDNAs were synthesized via reverse transcription with random primers. The PCR system was 20 μl in volume and contained 1 μl of template, 10 μl of Mixture (Yeasen, China), 1 μl upstream primers (10 μmol/L), 1 μl downstream primers (10 μmol/L), and 7 μl ddH2O. The reaction was carried out on a T100 thermocycler (Bio-Rad, United States) under the following conditions: 94°C for 5 min; followed by 36 cycles of 94°C for 40 s, an appropriate annealing temperature (according to the melting temperature of the primers) for 30 s; 72°C for 30 s; and 72°C for 5 min. The PCR products were detected on 1.5% agarose gel electrophoresis (AGE) followed by TA cloning and Sanger sequencing.
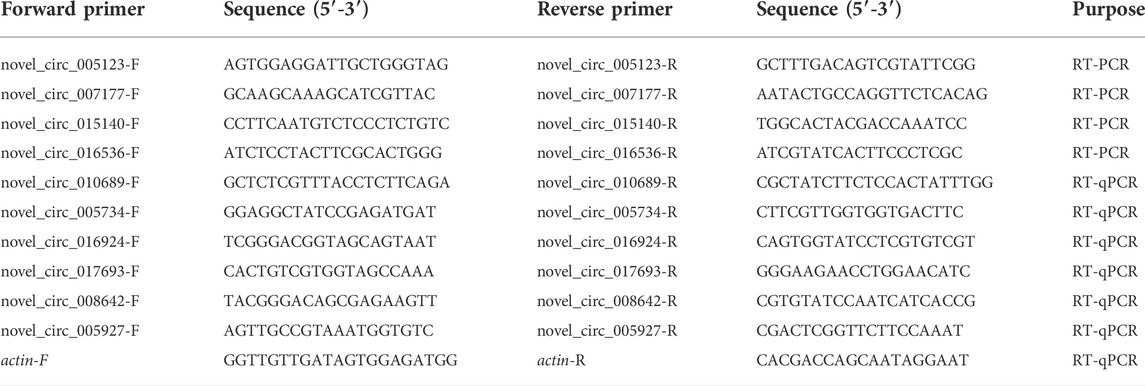
TABLE 1. Primers for RT-PCR and RT-qPCR conducted in this work.
Four DEcircRNAs in AcCK1 vs AcT1 comparison group (novel_circ_010689, novel_circ_005734, novel_circ_016924, novel_circ_0176939) and two DEcircRNAs in AcCK2 vs. AcT2 comparison group (novel_circ_008642, novel_circ_005927) were randomly selected for RT-qPCR. Divergent primers for these DEcircRNAs were designed and synthesized (Table 1). Total RNA from the AcCK1, AcT1, AcCK2 and AcT2 groups was isolated and then subjected to reverse transcription. The resulting cDNA was used as a template for the internal control (actin) and DEcircRNAs. The RT-qPCR system was 20 μl in a volume containing 1 μl upstream primers (10.0 μmol/L), 1 μl downstream primers (10.0 μmol/L), 1 μl cDNA, 10 μl SYBR Green Dye, and 7 μl DEPC H2O. RT-qPCR was performed on an ABI Q3 Real-time PCR Detection System (Applied Biosystems, United States) under the following conditions: predenaturation step at 94°C for 5 min; 36 amplification cycles of denaturation at 94°C for 50 s, annealing at 60°C for 30 s, and elongation at 72°C for 1 min; and a final elongation step at 72°C for 30 s. The data were calculated using the 2-△△Ct method and presented as relative expression levels from three parallel replicates and three biological replicates, followed by analysis and visualization using GraphPad Prism 6.0 software (GraphPad, United States).
Statistical analyses were conducted utilizing SPSS 16.0 (IBM, Armonk, NY, United States) and GraphPad Prism 6.0 (GraphPad, United States) software. Data were shown as the mean ± standard deviation (SD). Statistical analyses were performed on basis of an independent-sample t test. Fisher’s exact test was conducted to filter the significant GO terms and KEGG pathways with R software 3.3.1 (R Development Core Team, https://www.r-project.org/). p < 0.05 was considered statistically significant.
Totally, 524100096 and 615893838 raw reads were generated from the AcT1 and AcT2 groups, and 515604182 and 601712328 clean reads were obtained after quality control, respectively; additionally, 338500740 and 570789154 anchor reads were identified, among which 31889778 (28.28%) and 35037206 (20.60%) were respectively aligned to the reference genome of A. cerana.
In the AcT1 and AcT2 groups, 10185 and 7405 circRNAs were identified respectively. Combined with the previously identified circRNAs in the un-inoculated groups, the Venn analysis indicated that 2266 circRNAs were shared by the AcCK1, AcCK2, AcT1, and AcT2 groups, while 2618, 1917, 5691 and 3723 ones were specific to each group, respectively (Figure 1A). Additionally, annotated exonic circRNA was the most abundant type in both the AcT1 and AcT2 groups, followed by antisense circRNA and single exonic circRNA (Figure 1b). Moreover, the length of circRNAs in the N. ceranae-inoculated groups ranged from 1 nt to more than 2000 nt, and circRNAs with a length distribution of 401–600 nt were the most abundant (Figure 1C).
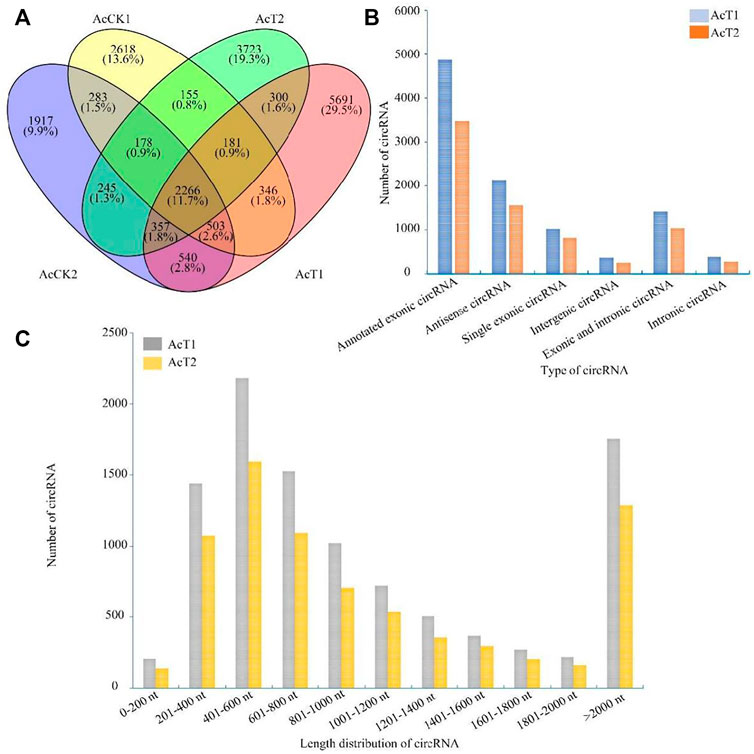
FIGURE 1. Number, type, and length distribution of circRNAs in the midguts of A. c. cerana workers inoculated with N. ceranae. (A) Venn diagram of circRNAs in AcCK1, AcCK2, AcT1, and AcT2 groups. (B) Number statistics of circRNAs derived from various genomic origins. (C) Length distribution of circRNAs.
PCR amplification was performed to further validate the three specific circRNAs identified in N. ceranae-inoculated midguts, and AGE suggested that fragments with expected sizes could be amplified using specific divergent primers for novel_circ_005123 (approximately 162 bp), novel_circ_007177 (approximately 195 bp), and novel_circ_015140 (approximately 108 bp) (Figure 2A). Additionally, the back-splicing sites of these selected circRNAs were successfully detected using Sanger sequencing (Figure 2B).
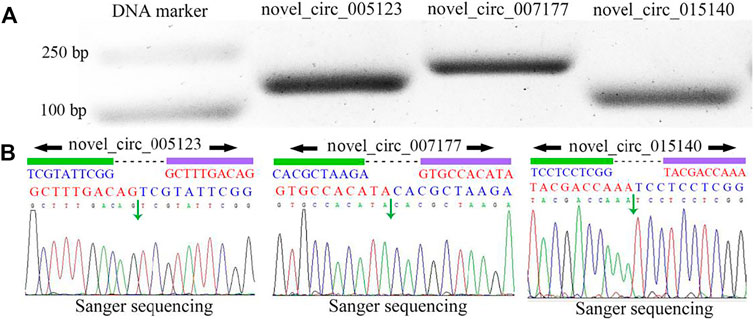
FIGURE 2. PCR amplification (A) and Sanger sequencing (B) of three A. c. cerana circRNAs. Black arrows indicate the direction of primers for PCR amplification, and green arrows indicate back-splicing sites within circRNAs.
A total of 83 DEcircRNAs were identified in the AcCK1 vs AcT1 comparison group, including 57 up-regulated circRNAs and 26 down-regulated circRNAs (Figure 3A, see also Supplementary Table S1. The expression levels of the DEcircRNAs were between 0.001 and 353.49, and the most up-regulated and down-regulated circRNAs were novel_circ_012754 (log2FC = 18.16) and novel_circ_017486 (log2FC = -17.05), respectively. Comparatively, 28 up-regulated circRNAs and 24 down-regulated circRNAs were detected in the AcCK2 vs. AcT2 comparison group (Figure 3A, see also Supplementary Table S2. The expression levels of the circRNAs were among 0.001–445.77, and the most up-regulated and down-regulated circRNAs were novel_circ_002265 (log2FC = 18.77) and novel_circ_011100 (log2FC = −17.50), respectively. In addition, none of the DEcircRNAs were shared by the two abovementioned comparison groups (Figure 3B).
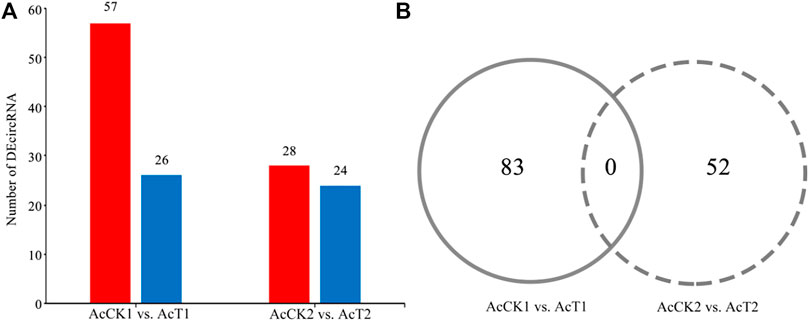
FIGURE 3. Number statistics of DEcircRNAs in AcCK1 vs. AcT1 and AcCK2 vs. AcT2 comparison groups. (A) Number of up- and down-regulated circRNAs (B) Venn analysis of DEcircRNAs.
GO classification suggested that 78 source genes of DEcircRNAs in the AcCK1 vs AcT1 comparison group were predicted; among these, 23 ones were classified into 10 functional terms associated with molecular functions, cellular components, and biological processes, such as binding, localization, and membrane (Figure 4A). Additionally, 45 source genes of DEcircRNAs in the AcCK2 vs. AcT2 comparison group were predicted, among which 13 ones were grouped into 10 functional terms including catalytic activity, cell part, and metabolic processes (Figure 4B).
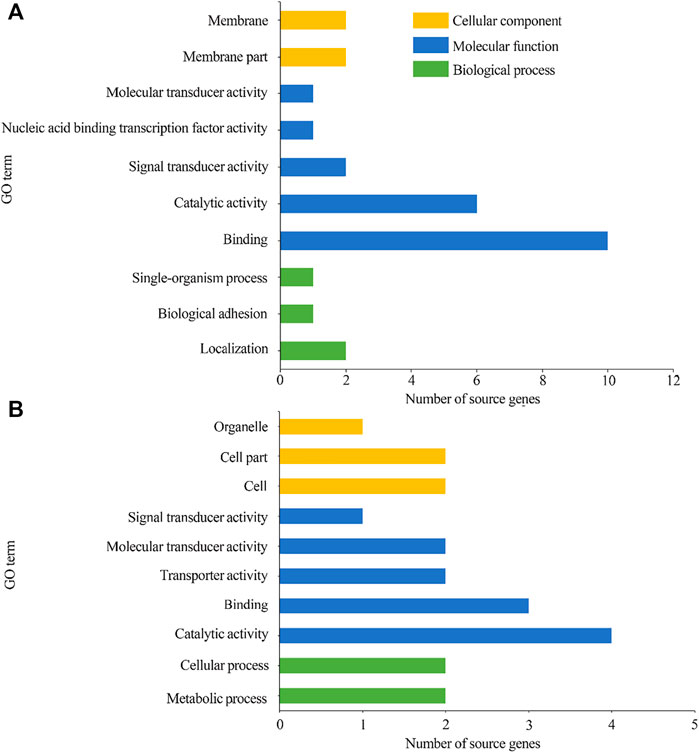
FIGURE 4. GO categorization of DEcircRNAs’ source genes in AcCK1 vs AcT1 (A) and AcCK2 vs AcT2 (B) comparison groups.
The KEGG pathway analysis suggested that the source genes of DEcircRNAs in the AcCK1 vs AcT1 comparison group were involved in 12 pathways, such as endocytosis and the Hippo signaling pathway (Table 2). Comparatively, the source genes of DEcircRNAs in the AcCK2 vs. AcT2 comparison group were engaged in five pathways, including the sphingolipid metabolism and the mTOR signaling pathways (Table 3).
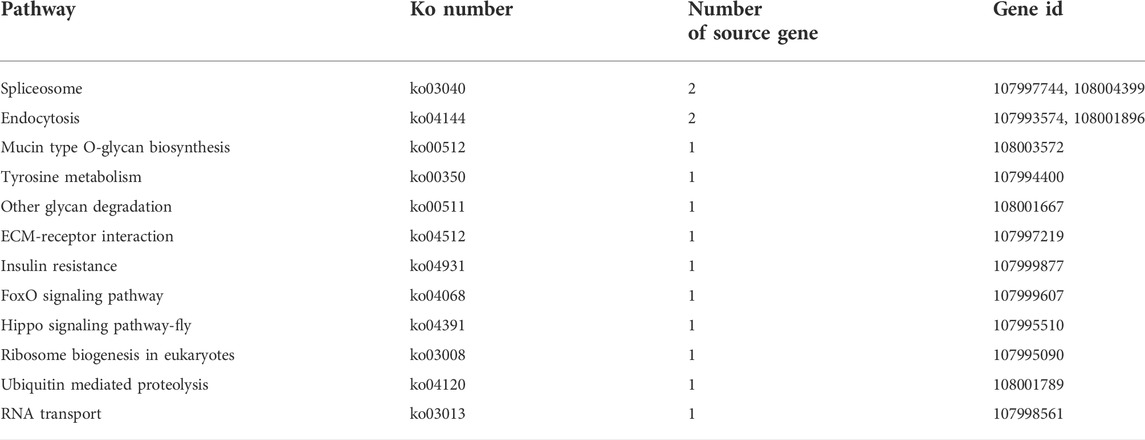
TABLE 2. KEGG pathways enriched by source genes of DEcircRNAs in AcCK1 vs AcT1 comparison group.

TABLE 3. KEGG pathways enriched by source genes of DEcircRNAs in AcCK2 vs. AcT2 comparison group.
In the AcCK1 vs. AcT1 comparison group, 23 DEcircRNAs were predicted to target 18 miRNAs; among these DEcircRNAs, novel_circ_015903 could target three miRNAs, novel_circ_016623 and novel_circ_010617 could target two miRNAs, whereas another 20 DEcircRNAs could target only one miRNA (Figure 5A). In addition, 13 DEcircRNAs in the AcCK2 vs. AcT2 comparison group were found to target 14 miRNAs; among these DEcircRNAs, five DEcircRNAs could target two miRNAs, while another eight DEcircRNAs could only target one miRNA (Figure 5B).
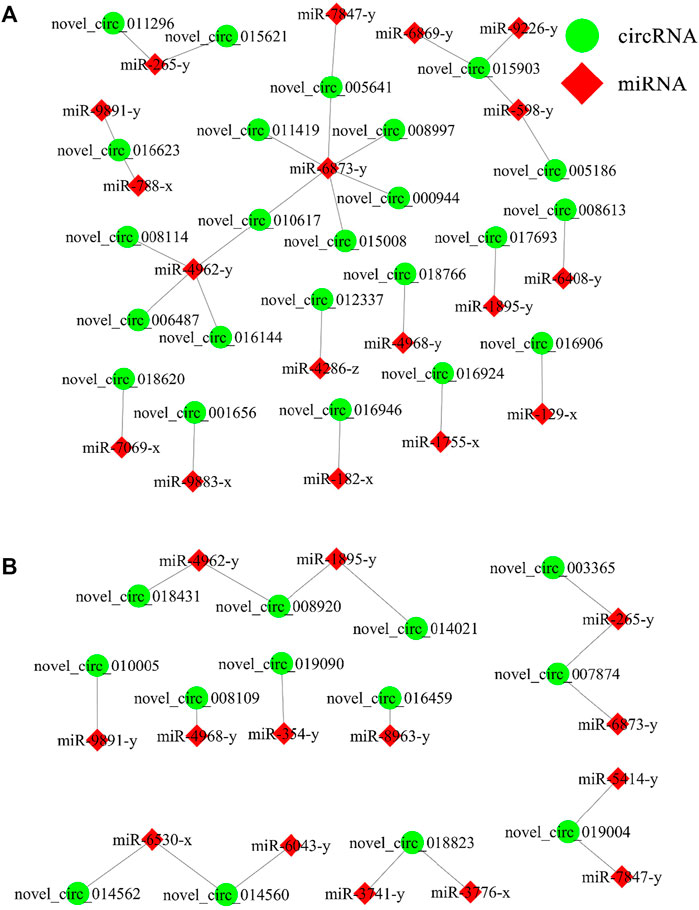
FIGURE 5. DEcircRNA-miRNA network engaged in N. ceranae-response of A. c. cerana workers. (A) Regulatory network in AcCK1 vs AcT1 comparison group (B) Regulatory network in AcCK2 vs AcT2 comparison group.
Further investigation showed that 23 DEcircRNAs in the AcCK1 vs AcT1 comparison group can target 18 miRNAs and further target 1111 mRNAs Supplementary Table S3. These target mRNAs were annotated to 10 cellular component-related terms, such as cell and membrane; eight molecular function-related terms, such as catalytic activity and transporter activity; and 11 biological process-related terms, such as cellular process and biological regulation Supplementary Table S5. Additionally, these target mRNAs could also be annotated to 72 pathways, including endocytosis, RNA transport, and ubiquitin-mediated proteolysis Supplementary Table S6. Comparatively, 13 DEcircRNAs in the AcCK2 vs AcT2 comparison groups can target 14 miRNAs and further target 1093 mRNAs Supplementary Table S4. These target mRNAs were involved in 10 cellular component-associated terms, including organelle and cell; eight molecular function-associated terms, including binding and catalytic activity; and 11 biological process-associated terms, including biological regulation and single-organism process Supplementary Table S7. In addition, these target mRNAs could also be annotated to 72 pathways, such as endocytosis, purine metabolism, RNA transport, protein processing of the endoplasmic reticulum, and ubiquitin-mediated proteolysis Supplementary Table S8. Moreover, target mRNAs in both the AcCK1 vs AcT1 and AcCK2 vs AcT2 comparison groups were relevant to six cellular immune-related pathways, including endocytosis, lysosomes, phagosomes, ubiquitin-mediated proteolysis, metabolism of xenobiotics by cytochrome P450, and insect hormone biosynthesis (Table 4); however, no target was found to annotate to any humoral immune pathway.

TABLE 4. Summary of cellular immune pathways enriched by DEcircRNA-targeted mRNAs within ceRNA networks.
In total, 284 IRESs and 54 ORFs were identified from the DEcircRNAs in the AcCK1 vs. AcT1 comparison group. Supplementary Table S9, S10. These ORFs were involved in two biological process-associated terms, four molecular function-associated terms, and two cellular component-associated terms Additionally, these ORFs were associated with eight pathways, namely, endocytosis, other glycan degradation, ECM-receptor interaction, insulin resistance, other glycan degradation, FoxO signaling pathway, ribosome biogenesis in eukaryotes, ubiquitin-mediated proteolysis, and spliceosome Supplementary Table S11. Comparatively, 164 IRES and 26 ORF were identified from the DEcircRNAs in the AcCK2 vs AcT2 comparison group Supplementary Table S12. These ORFs were enriched in two biological process-related terms, four molecular function-related terms, and two cellular component-related terms Supplementary Table S13. In addition, these ORFs were relative to four pathways involving sphingolipid metabolism, ECM-receptor interaction, insulin resistance, and neuroactive ligand‒receptor interaction Supplementary Table S14.
Six DEcircRNAs were randomly selected for RT-qPCR validation, and the results showed that their expression trends were consistent with those in high-throughput sequencing data, which confirmed the reliability of the transcriptome data used in this current work (Figure 6).
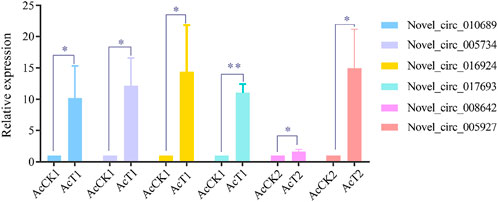
FIGURE 6. RT-qPCR verification of DEcircRNAs.Student’s t test, “*” indicates p ≤ 0.05 and “**” indicates p ≤ 0.01.
Combining findings in this current work, a working model of DEcircRNA-regulated response of A. c. cerana workers to N. ceranae infection was summarized and presented in Figure 7.

FIGURE 7. A working model of DEcircRNA-regulated response of A. c. cerana workers to N. ceranae infection.
Here, based on previously obtained high-quality transcriptome data from N. ceranae-inoculated and un-inoculated midguts of A. c. cerana workers, the regulatory roles of circRNAs involved in the N. ceranae-response of A. c. cerana workers were investigated for the first time. In N. ceranae-inoculated workers’ midguts at seven dpi and 10 dpi, 10185 and 7405 circRNAs were respectively identified, among which annotated exon circRNA was the most abundant type (Figure 1b); additionally, the length distribution of the identified A. c. cerana circRNAs were ranged from 201 nt to 800 nt (Figure 1C). Similarly, we previously identified 6530 and 6289 circRNAs in un-inoculated workers’ midguts using the same bioinformatic approach, and found that their length distribution was among 201 nt∼800 nt and the most abundant type was also annotated exon circRNA (Chen H. Z. et al., 2020). Further analysis showed that the number of circRNAs enriched in each length or circularization type in the N. ceranae-inoculated groups was higher than that in the un-inoculated groups, implying that A. c. cerana workers may respond to N. ceranae infection by altering circRNAs’ length and circularization type. In addition, 2266 circRNAs were shared by the AcCK1, AcCK2, AcT1, and AcT2 groups, while the numbers of specific circRNAs were 2618, 1917, 5717, and 3742, respectively (Figure 1A), indicative of the developmental stage- and stress stage-specific expression of A. c. cerana circRNAs, similar to the expression characteristic of circRNAs identified in other animals and plants (Hu et al., 2018a; Liang et al., 2019). Since the information about circRNAs in Asian honey bees is scarse, the circRNAs discovered here could well enrich the reservoir of A. cerana circRNAs. Furthermore, 83 and 52 DEcircRNAs were identified circRNAs in the AcCK1 vs. AcT1 and AcCK2 vs AcT2 comparison groups, respectively (Figure 3B), which suggested that the expression profile of circRNAs in host midguts was altered due to N. ceranae infection. It is inferred that these host-derived DEcircRNAs were involved in the N. ceranae-response. Interestingly, it is noticed that no DEcircRNA was shared by the aforementioned two comparison groups, suggesting that different circRNAs may be utilized by host to respond to the same microsporidian at different timepoints during infection, which deserved more efforts and further investigation. Apis mellifera ligustica, a subspecies of western honey bee (Apis mellifera), is widely reared bee species in China and many other countries. Recently, we discovered that nine up-regulated and 10 down-regulated circRNAs were shared by Apis mellifera ligustica workers’ midguts at seven dpi and 10 dpi with N. ceranae (Chen et al., 2022). Together, the results indicated that some circRNAs were adopted by western honey bee at various timepoints during the N. ceranae infection, reflecting the difference of circRNA-regulated strategies used by Asian honey bee and western honey bee.
Accumulating evidence suggests that circRNAs are able to exert their functions by regulating the transcription of source genes (Zhang et al., 2013; Li et al., 2015). To steal host cell-produced material and energy for fungal proliferation, N. ceranae prolongs the survival time of infected cells by inhibiting apoptosis of A. mellifera midgut epithelial cells (Kurze et al., 2018; Paris et al., 2018). Additionally, N. ceranae could also cause structural damage to midgut epithelial cells of A. mellifera (Panek et al., 2018). The Hippo signaling pathway plays a crucial role in regulating cell proliferation as well as organ growth and regeneration (Tang et al., 2022). Emerging evidence demonstrated the participation of the Hippo signaling pathway in the regulation of immune defense in mammals and insects (Buchon et al., 2009; Hong et al., 2018). In this work, it is detetcted that in the AcCK1 vs AcT1 comparison group 78 source genes of 83 DEcircRNAs (novel_circ_008114) were involved in the Hippo signaling pathway, suggesting the roles of these DEcircRNAs in detecting damage to the midgut epithelial cell structure caused by N. ceranae infection. It is speculated that the corresponding DEcircRNAs were employed by the host to regulate source gene transcription, thereby further modulating the Hippo signaling pathway to facilitate cell renewal and regulate the immune response.
Insects defend against pathogenic microorganisms based on cellular and humoral immunity, with the former involving endocytosis, phagocytosis, enzymatic hydrolysis, melanization, and antimicrobial peptide synthesis and release (Lavine and Strand. 2002). In honey bees, endocytosis is a main cellular immune pathway (Aronstein and Murray. 2010). Clathrin-mediated endocytosis is one of the most clearly studied endocytosis pathways. Kim et al. (2009) discovered that when compared with wild-type cells, deletion of FgEnd1 gene in Fusarium graminearum resulted in a significant downregulation of the endocytic marker FM4-64 and a decrease in the mycelium growth rate . Hodgson et al. (2019) disrupted two early and late endosome marker genes Rab5 and Rab7 in Drosophila DL1 cells using RNAi, the results showed that progeny virions of Autographa californica multiple nucleopolyhedrovirus (AcMNPV) were significantly reduced. Ubiquitin-mediated proteolysis is a classic cellular immune pathway, that involves the E1 (ubiquitin activase), E2 (ubiquitin binding enzyme) and E3 enzymes (ubiquitin ligase); E3 can specifically recognize different substrates and then bind to the E2 enzyme, which is finally recognized and degraded by the protein enzyme body (Sutovsky, 2003). Here, it is observed that two source genes of novel_circ_005307 and novel_circ_017023 in the AcCK1 vs AcT1 comparison group were engaged in endocytosis, while one source gene of novel_circ_016946 was involved in ubiquitin-mediated proteolysis. Together, these findings suggested that the corresponding DEcircRNAs were likely to regulate the two aforementioned cellular immune pathways by regulating source gene transcription, and then further participate in the response of host to N. ceranae infection. Intriguingly, in our previous work, we observed that the source genes of DEcircRNAs in A m ligustica workers’ midguts during N. ceranae infection were enriched in four cellular immune-related pathways, including endocytosis, ubiquitin-mediated proteolysis, lysosomes, and phagosomes (Chen et al., 2022). Collectively, these results indicated that both A. c. cerana and A m ligustica workers likely regulate endocytosis and ubiquitin-mediated proteolysis via the control of source gene transcription by DEcircRNAs, further responding to N. ceranae invasion, but only A m ligustica workers were capable of modulating another two cellular immune pathways lysosomes and phagosomes utilizing differential expression of specific circRNAs. The identified novel_circ_005307, novel_circ_017023, and novel_circ_016946 may be candidate targets for further functional study and bee nosemosis control, additional work is required to develop novel circRNA-based control strategies.
Increasing evidence suggests that circRNAs can regulate target gene expression via the ceRNA network and further affect various biological processes, such as the immune response and development (Han et al., 2020; Li et al., 2020). Here, 23 and 13 DEcircRNAs in the AcCK1 vs. AcT1 and AcCK2 vs AcT2 comparison groups were predicted to target 18 and 14 miRNAs and further target 1111 and 1093 mRNAs, respectively, implying that these DEcircRNAs may function as ceRNAs during the host response to N. ceranae infection. Further analysis indicated that target mRNAs in the worker’s midgut at seven dpi were associated with six cellular immune pathways including lysosome and phagosome, whereas targets in the worker’s midgut at 10 dpi were involved in five cellular immune-related pathways, namely, endocytosis, ubiquitin-mediated proteolysis, and insect hormone biosynthesis. Interestingly, none of the targets were enriched in the humoral immune pathways. The results demonstrated that the corresponding DEcircRNAs likely regulated the host cellular immune responses to N. ceranae infection through ceRNA networks.
The miR-182 gene was abundantly expressed in sensory organs and regulated the development of the retina and inner ear (Wei et al., 2015). Perilli et al. (2019) reported that overexpression of miR-182 in humans could inhibit apoptosis and promote cell proliferation as well as colorectal cancer cell infection by altering tumor cell cycle dynamics and morphology. Sun et al. (2010) revealed that miR-182 regulated RGS17 through two conserved sites within the 3′UTR, and ectopic expression of miR-182 conspicuously inhibited lung cancer cell proliferation and anchorage-independent cell growth. FOX O 3a was previously identified as a direct target of miR-182–5p, and miR-182–5p played an inhibitory role in FOX O 3a expression. Moreover, activation of the AKT/FOXO3a pathway promoted HCC proliferation and invasive ability, which further resulted in higher death rates (Cao et al., 2018). Wu et al. (2021) observed that miR-182–5p directly targeted the 3′ UTR of the tumor suppresser gene STARD13, which significantly relieved the inhibitory effect of decreased miR-182–5p on cell proliferation, migration, and invasion in lung adenocarcinoma. Here, miR-182 was detected to be targeted by novel_circ_016924 (log2FC = 17.25, P= 0.0020) and novel_circ_016946 (log2FC = 16.37, p = 0.0077) in the AcCK1 vs. AcT1 comparison group, , indicating that these two DEcircRNAs may play a pivotal role in cell apoptosis and cell proliferation in the N. ceranae-response of host by absorbing miR-182. Therefore, miR-182 and its targeted DEcircRNAs were promising biomarkers and molecular targets for diagnosis and control of bee nosemosis frequently occurred in beekeeping industry. In the near future, we plan to conduct overexpression and knockdown of miR-182 and siRNA-based RNAi of corresponding DEcircRNAs following our established technical platforms (Ye et al., 2022; Zhu et al., 2022).
Eukaryotic translation depends on the ribosomal scanning mechanism of the m7G cap structure (Haimov et al., 2015). Due to the lack of a 5’ terminal and poly-A tail, circRNA was previously considered unable to translate proteins. With the rapid development of next-generation and third-generation sequencing technologies, some circRNAs were verified to translate into proteins or small peptides with biological functions using an IRES-based manner (Wang and Wang, 2015; Pamudurti et al., 2017). Yang et al. (2018) reported that FBXW7-185aa, a protein encoded by circRNA FBXW7 (circ-FBXW7), plays an essential part in glioma carcinogenesis and patient clinical prognosis. After transfecting Drosophila S2 cells with artificially constructed circRNAs, including the gfp gene containing IRES (Wang et al., 2015), Wang and Wang (2015) detected that the constructed circRNA successfully expressed GFP protein. Recently, Weigelt et al. (2020) documented that overexpression of circSfl, a protein-coding circRNA, extended the lifespan of the insulin mutant Drosophila. In this current work, novel_circ_017023 and novel_circ_005307 in the AcCK1 vs. AcT1 comparison group were predicted to translate endocytic pathway-related proteins, and novel_circ_016946 was predicted to translate proteins associated with the ubiquitin-mediated proteolysis. The results suggested that the abovementioned three DEcircRNAs were likely to engaged in the cellular immune responses of A. c. cerana workers to N. ceranae invasion through the protein translation, which deserved further investigation in the future.
In the present study, we investigated for the first time the expression profiles and potential functions of circRNA in A. c. cerana workers’ midguts in response to N. ceranae infection. It is demonstrated that the expression pattern of circRNAs was altered due to N. ceranae infection and DEcircRNAs may play regulatory roles in the host cellular immune responses through versatile manners, such as regulation of the transcription of source genes,absorption of target miRNAs via the ceRNA networks, and translation of proteins as templates. Our data offer a foundation for clarifying the mechanism underlying the immune responses of A. c. cerana workers to N. ceranae invasion and provide novel insights into host-parasite interactions during bee nosemosis (Figure 7).
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethical review and approval was not required for the animal study because Honey bee is a insect, needing no animal ethics approval.
ZZ and WJ designed experiments, carried out experiments, wrote the first draft of themanuscript and organizedthe database. FX analyzed experimental results LQ, CH, and YY analyzed sequencing data and developed analysis tools. ZK, RZ, and ZY Visualization and Investigation NQ assisted with Illumina sequencing,performed the statistical analysis CD and GR contributed to conception and design of the study. All authors contributed to manuscript revision, read, and approved the submitted version.
The National Natural Science Foundation of China (32172792), the Earmarked Fund for CARS-44-KXJ7 (CARS-44-KXJ7), the Master Supervisor Team Fund of Fujian Agriculture and Forestry University (RG), the Outstanding Scientific Research Manpower Fund of Fujian Agriculture and Forestry University (xjq201814), the Undergraduate Innovation and Entrepreneurship Training Program of Fujian province (HZ), and the Undergraduate Innovation and Entrepreneurship Training Program of Fujian province (202210389114,202210389131).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.1013239/full#supplementary-material
Antúnez, K., Martín-Hernández, R., Prieto, L., Meana, A., Zunino, P., and Higes, M. (2009). Immune suppression in the honey bee (Apis mellifera) following infection by Nosema ceranae (Microsporidia). Environ. Microbiol. 11 (9), 2284–2290. doi:10.1111/j.1462-2920.2009.01953.x
Aronstein, K. A., and Murray, K. D. (2010). Chalkbrood disease in honey bees. J. Invertebr. Pathol. 103 (1), S20–S29. doi:10.1016/j.jip.2009.06.018
Benjeddou, M., Leat, N., Allsopp, M., and Davison, S. (2001). Detection of acute bee paralysis virus and black queen cell virus from honeybees by reverse transcriptase PCR. Appl. Environ. Microbiol. 67, 2384–2387. doi:10.1128/AEM.67.5.2384-2387.2001
Buchon, N., Broderick, N. A., Chakrabarti, S., and Lemaitre, B. (2009). Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev. 23 (19), 2333–2344. doi:10.1101/gad.1827009
Cao, M. Q., You, A. B., Zhu, X. D., Zhang, W., Zhang, Y. Y., Zhang, S. Z., et al. (2018). MiR-182-5p promotes hepatocellular carcinoma progression by repressing FOXO3a. J. Hematol. Oncol. 11 (1), 12. doi:10.1186/s13045-018-0555-y
Chen, Y. P., Evans, J. D., Smith, I. B., and Pettis, J. S. (2008). Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 97 (2), 186–188. doi:10.1016/j.jip.2007.07.010
Chen, D. F., Du, Y., Chen, H. Z., Fan, Y. C., Fan, X. X., Zhu, Z. W., et al. (2019a). Comparative identification of microRNAs in Apis cerana cerana workers' midguts in response to Nosema ceranae invasion. Insects 10 (9), E258. doi:10.3390/insects10090258
Chen, R. X., Liu, H. L., Yang, L. L., Kang, F. H., Xin, L. P., Huang, L. R., et al. (2019b). Circular RNA circRNA_0000285 promotes cervical cancer development by regulating FUS. Eur. Rev. Med. Pharmacol. Sci. 23 (20), 8771–8778. doi:10.26355/eurrev_201910_19271
Chen, X., Shi, W., and Chen, C. (2019c). Differential circular RNAs expression in ovary during oviposition in honey bees. Genomics 111 (4), 598–606. doi:10.1016/j.ygeno.2018.03.015
Chen, D. F., Chen, H. Z., Du, Y., Zhu, Z. W., Wang, J., Geng, S. H., et al. (2020a). Systematic identification of circular RNAs and corresponding regulatory networks unveil their potential roles in the midguts of eastern honeybee workers. Appl. Microbiol. Biotechnol. 104 (1), 257–276. doi:10.1007/s00253-019-10159-9
Chen, H. Z., Fu, Z. M., Wang, J., Zhu, Z. W., Fan, X. X., Jiang, H. B., et al. (2020b). Circular RNA response of Apis cerana cerana 6-day-old larvae to Ascosphaera apis stress. Acta. Microbiol. Sin. 60 (10), 2292–2310. (in Chinese). doi:10.13343/j.cnki.wsxb.20200445
Chen, H. Z., Fan, X. X., Zhang, W. D., Ye, Y. P., Cai, Z. B., Zhang, K. Y., et al. (2022). Deciphering the circRNA-Regulated response of Western honey bee (Apis mellifera) workers to microsporidian invasion. Biology 11, 1285. doi:10.3390/biology11091285
Doublet, V., Labarussias, M., deMiranda, J. R., Moritz, R. F. A., and Paxton, R. J. (2015). Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 17 (4), 969–983. doi:10.1111/1462-2920.12426
Du, J., Yuan, Z., Ma, Z., Song, J., Xie, X., and Chen, Y. (2014). KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway analysis using a path analysis model. Mol. Biosyst. 10, 2441–2447. doi:10.1039/c4mb00287c
Emsen, B., Mora, A., Lacey, B., Eccles, L., Kelly, P. G., Medina-Flores C, A., et al. (2020). Seasonality of Nosema ceranae infections and their relationship with honey bee populations, food stores, and survivorship in a North American Region. Vet. Sci. 7 (3), 131. doi:10.3390/vetsci7030131
Fan, W. Y., Li, G. L., Zhang, X. M., Wang, Y., Wang, C., Xu, B. H., et al. (2021). The role of melatonin and Tryptophan-5-hydroxylase-1 in different abiotic stressors in Apis cerana cerana. J. Insect Physiol. 128, 104180. doi:10.1016/j.jinsphys.2020.104180
Fries, I., Feng, F., DaSilva, A., Slemenda, S. B., and Pieniazek, N. J. (1996). Nosema ceranae n sp (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 32 (3), 356–365. doi:10.1016/s0932-4739(96)80059-9
Fu, Z. M., Zhou, D. D., Chen, H. Z., Geng, S. H., Chen, D. F., Zheng, Y. Z., et al. (2020). Analysis of highly expressed genes in Apis cerana cerana workers' midguts responding to Nosema ceranae stress. J. Sichuan Univ. Nat. Sci. Ed. 57 (01), 191–198. (in Chinese). doi:10.3969/j.issn.0490-6756.2020.01.02
Gan, H. Y., Feng, T. S., Wu, Y. Q., Liu, C., Xia, Q. Y., and Cheng, T. C. (2017). Identification of circular RNA in the Bombyx mori silk gland. Insect biochem. Mol. Biol. 89, 97–106. doi:10.1016/j.ibmb.2017.09.003
Genersch, E. (2005). Development of a rapid and sensitive RT-PCR method for the detection of deformed wing virus, a pathogen of the honeybee (Apis mellifera). Vet. J. 169, 121–123. doi:10.1016/j.tvjl.2004.01.004
Giersch, T., Berg, T., Galea, F., and Hornitzky, M. (2009). Nosema ceranae infects honey bees (Apis mellifera) and contaminates honey in Australia. Apidologie 40 (2), 117–123. doi:10.1051/apido/2008065
Gisder, S., Möckel, N., Linde, A., and Genersch, E. (2011). A cell culture model for Nosema ceranae and Nosema apis allows new insights into the life cycle of these important honey bee-pathogenic microsporidia. Environ. Microbiol. 13 (2), 404–413. doi:10.1111/j.1462-2920.2010.02346.x
Goblirsch, M., Huang, Z. Y., and Spivak, M. (2013). Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection. PLoS One 8 (3), e58165. doi:10.1371/journal.pone.0058165
Guo, R., Chen, D., Chen, H. Z., Fu, Z. M., Xiong, C. L., Hou, C. S., et al. (2018c). Systematic investigation of circular RNAs in Ascosphaera apis, a fungal pathogen of honeybee larvae. Gene 678, 17–22. doi:10.1016/j.gene.2018.07.076
Guo, R., Chen, D. F., Chen, H. Z., Xiong, C. L., Zheng, Y. Z., Hou, C. S., et al. (2018b). Genome-wide identification of circular RNAs in fungal parasite Nosema ceranae. Curr. Microbiol. 75 (12), 1655–1660. doi:10.1007/s00284-018-1576-z
Guo, R., Chen, H. Z., Xiong, C. L., Zheng, Y. Z., Fu, Z. M., Xu, G. J., et al. (2018a). Analysis of differentially expressed circular RNAs and their regulation networks during the developmental process of Apis mellifera ligustica worker’s midgut. Sci. Agric. Sin. 51 (23), 4575–4590. (in Chinese). doi:10.13343/j.cnki.wsxb.20200445
Haimov, O., Sinvani, H., and Dikstein, R. (2015). Cap-dependent, scanning-free translation initiation mechanisms. Biochim. Biophys. Acta 1849 (11), 1313–1318. doi:10.1016/j.bbagrm.2015.09.006
Han, D., Wang, Y. J., Wang, Y. B., Dai, X. C., Zhou, T. W., Chen, J. W., et al. (2020). The tumor-suppressive human circular RNA circITCH sponges miR-330-5p to Ameliorate doxorubicin-induced cardiotoxicity through upregulating SIRT6, survivin, and SERCA2a. Circ. Res. 127 (4), e108–e125. doi:10.1161/CIRCRESAHA.119.316061
He, J., Xie, Q., Xu, H., Li, J., and Li, Y. (2017). Circular RNAs and cancer. Cancer Lett. 396, 138–144. doi:10.1016/j.canlet.2017.03.027
Higes, M., García-Palencia, P., Martín-Hernández, R., and Meana, A. (2007). Experimental infection of Apis mellifera honeybees with Nosema ceranae (microsporidia). J. Invertebr. Pathol. 94 (3), 211–217. doi:10.1016/j.jip.2006.11.001
Higes, M., Martín-Hernández, R., and Meana, A. (2010). Nosema ceranae in Europe: An emergent type C nosemosis. Apidologie 41, 375–392. doi:10.1051/apido/2010019
Hodgson, J. J., Buchon, N., and Blissard, G. W. (2019). Identification of insect genes involved in baculovirus AcMNPV entry into insect cells. Virology 527, 1–11. doi:10.1016/j.virol.2018.10.022
Honeybee Genome Sequencing Consortium (2006). Insights into social insects from the genome of the honeybee Apis mellifera. Nature 443 (7114), 931–949. doi:10.1038/nature05260
Hong, L. X., Li, X., Zhou, D. W., Geng, J., and Chen, L. F. (2018). Role of Hippo signaling in regulating immunity. Cell. Mol. Immunol. 15 (12), 1003–1009. doi:10.1038/s41423-018-0007-1
Hu, X. L., Zhu, M., Liu, B., Liang, Z., Huang, L. X., Xu, J., et al. (2018a). Circular RNA alterations in the Bombyx mori midgut following B. mori nucleopolyhedrovirus infection. Mol. Immunol. 101, 461–470. doi:10.1016/j.molimm.2018.08.008
Hu, X. L., Zhu, M., Zhang, X., Liu, B., Liang, Z., Huang, L. X., et al. (2018b). Identification and characterization of circular RNAs in the silkworm midgut following Bombyx mori cytoplasmic polyhedrosis virus infection. RNA Biol. 15 (2), 292–301. doi:10.1080/15476286.2017.1411461
Huang, D. W., Sherman, B. T., Tan, Q., Collins, J. R., Alvord, W. G., Roayaei, J., et al. (2007). The DAVID gene functional classification tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 8 (9), R183. doi:10.1186/gb-2007-8-9-r183
Huang, Q., and Evans, J. D. (2020). Targeting the honey bee gut parasite Nosema ceranae with siRNA positively affects gut bacteria. BMC Microbiol. 20 (1), 258. doi:10.1186/s12866-020-01939-9
Huang, C., Liang, D. M., Tatomer, D. C., and Wilusz, J. E. (2018a). A length-dependent evolutionarily conserved pathway controls nuclear export of circular RNAs. Genes Dev. 32 (9-10), 639–644. doi:10.1101/gad.314856.118
Huang, S. K., Ye, K. T., Huang, W. F., Ying, B. H., Su, X., Lin, L. H., et al. (2018b). Influence of feeding type and Nosema ceranae infection on the gut microbiota of Apis cerana workers. mSystems 3 (6), 001777–e218. doi:10.1128/mSystems.00177-18
Jung, S. H. (2014). Stratified Fisher's exact test and its sample size calculation. Biom. J. 56, 129–140. doi:10.1002/bimj.201300048
Kim, J. H., Kim, H. W., Heo, D. H., Chang, M., Baek, I. J., and Yun, C. W. (2009). FgEnd1 is a putative component of the endocytic machinery and mediates ferrichrome uptake in F. graminearum. Curr. Genet. 55, 593–600. doi:10.1007/s00294-009-0272-8
Kim, D., Pertea, G., Trapnell, C., Pimentel, H., Kelley, R., and Salzberg, S. L. (2013). TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14 (4), R36. doi:10.1186/gb-2013-14-4-r36
Krishnamoorthy, A., and Kadener, S. (2021). Using Drosophila to uncover molecular and physiological functions of circRNAs. Methods 196, 74–84. doi:10.1016/j.ymeth.2021.04.016
Kurze, C., Conte, Y. L., Kryger, P., Lewkowski, O., Müller, T., and Moritz, R. F. A. (2018). Infection dynamics of Nosema ceranae in honey bee midgut and host cell apoptosis. J. Invertebr. Pathol. 154, 1–4. doi:10.1016/j.jip.2018.03.008
Lai, X. L., Bazin, J., Webb, S., Crespi, M., Zubieta, C., and Conn, S. J. (2018). CircRNAs in plants. Adv. Exp. Med. Biol. 1087, 329–343. doi:10.1007/978-981-13-1426-1_26
Lavine, M. D., and Strand, M. R. (2002). Insect hemocytes and their role in immunity. Insect biochem. Mol. Biol. 32 (10), 1295–1309. doi:10.1016/s0965-1748(02)00092-9
Lei, M., Zheng, G. T., Ning, Q. Q., Zheng, J. N., and Dong, D. (2020). Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 19 (1), 30. doi:10.1186/s12943-020-1135-7
Li, Z. Y., Huang, C., Bao, C., Chen, L., Lin, M., Wang, X. L., et al. (2015). Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 22 (3), 256–264. doi:10.1038/nsmb.2959
Li, X., Yang, L., and Chen, L. L. (2018). The biogenesis, functions, and challenges of circular RNAs. Mol. Cell 71 (3), 428–442. doi:10.1016/j.molcel.2018.06.034
Li, J. F., Huang, C. C., Zou, Y. F., Ye, J., Yu, J., and Gui, Y. T. (2020). circTLK1 promotes the proliferation and metastasis of renal cell carcinoma by sponging miR-136-5p. Mol. Cancer 19 (1), 103. doi:10.1186/s12943-020-01225-2
Liang, Y. W., Zhang, Y. Z., Xu, L. S., Zhou, D., Jin, Z. M., Zhou, H. Y., et al. (2019). CircRNA Expression pattern and ceRNA and miRNA-mRNA networks involved in anther development in the CMS line of Brassica campestris. Int. J. Mol. Sci. 20 (19), 4808. doi:10.3390/ijms20194808
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 25 (4), 402–408. doi:10.1006/meth.2001.1262
Mayack, C., and Naug, D. (2009). Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 100 (3), 185–188. doi:10.1016/j.jip.2008.12.001
Memczak, S., Jens, M., Elefsinioti, A., Torti, F., Krueger, J., Rybak, A., et al. (2013). Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 495 (7441), 333–338. doi:10.1038/nature11928
Meng, S. J., Zhou, H. C., Feng, Z. Y., Xu, Z. H., Tang, Y., Li, P. Y., et al. (2017). CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 16 (1), 94. doi:10.1186/s12943-017-0663-2
Pamudurti, N. R., Bartok, O., Jens, M., Ashwal-Fluss, R., Stottmeister, C., Ruhe, L., et al. (2017). Translation of circRNAs. Mol. Cell 66 (1), 9–21. doi:10.1016/j.molcel.2017.02.021
Panek, J., Paris, L., Roriz, D., Mone, A., Dubuffet, A., Delbac, F., et al. (2018). Impact of the microsporidian Nosema ceranae on the gut epithelium renewal of the honeybee, Apis mellifera. J. Invertebr. Pathol. 159, 121–128. doi:10.1016/j.jip.2018.09.007
Papini, R., Mancianti, F., Canovai, R., Cosci, F., Rocchigiani, G., Benelli, G., et al. (2017). Prevalence of the microsporidian Nosema ceranae in honeybee(Apis mellifera) apiaries in Central Italy. Saudi J. Biol. Sci. 24 (5), 979–982. doi:10.1016/j.sjbs.2017.01.010
Paris, L., Alaoui, H., Delbac, F., and Diogon, M. (2018). Effects of the gut parasite Nosema ceranae on honey bee physiology and behavior. Curr. Opin. Insect Sci. 26, 149–154. doi:10.1016/j.cois.2018.02.017
Park, D., Jung, J. W., Choi, B. S., Jayakodi, M., Lee, J., Lim, J., et al. (2015). Uncovering the novel characteristics of Asian honey bee, Apis cerana, by whole genome sequencing. BMC Genomics 16 (1), 1. doi:10.1186/1471-2164-16-1
Perilli, L., Tessarollo, S., Albertoni, L., Curtarello, M., Pastò, A., Brunetti, E., et al. (2019). Silencing of miR-182 is associated with modulation of tumorigenesis through apoptosis induction in an experimental model of colorectal cancer. BMC Cancer 19 (1), 821. doi:10.1186/s12885-019-5982-9
Ribiere, M., Triboulot, C., Mathieu, L., Aurieres, C., Faucon, J. P., and Pepin, M. (2002). Molecular diagnosis of chronic bee paralysis virus infection. Apidologie 33, 339–351. doi:10.1051/apido:2002020
Singh, R., Levitt, A. L., Rajotte, E. G., Holmes, E. C., Ostiguy, N., Vanengelsdorp, D., et al. (2010). RNA viruses in hymenopteran pollinators: Evidence of inter-Taxa virus transmission via pollen and potential impact on non-Apis hymenopteran species. PLoS One 5 (12), e14357. doi:10.1371/journal.pone.0014357
Sinpoo, C., Paxton, R. J., Disayathanoowat, T., Krongdang, S., and Chantawannakul, P. (2017). Impact of Nosema ceranae and Nosema apis on individual worker bees of the two host species (Apis cerana and Apis mellifera) and regulation of host immune response. J. Insect Physiol. 105, 1–8. doi:10.1016/j.jinsphys.2017.12.010
Smoot, M. E., Ono, K., Ruscheinski, J., Wang, P. L., and Ideker, T. (2011). Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 27 (3), 431–432. doi:10.1093/bioinformatics/btq675
Stoltz, D., Shen, X. R., Boggis, C., and Sisson, G. (1995). Molecular diagnosis of Kashmir bee virus infection. J. Apic. Res. 34, 153–160. doi:10.1080/00218839.1995.11100900
Sun, Y. H., Fang, R., Li, C. G., Li, L., Li, F., Ye, X. L., et al. (2010). Hsa-mir-182 suppresses lung tumorigenesis through down regulation of RGS17 expression in vitro. Biochem. Biophys. Res. Commun. 396 (2), 501–507. doi:10.1016/j.bbrc.2010.04.127
Sutovsky, P. (2003). Ubiquitin-dependent proteolysis in mammalian spermatogenesis, fertilization, and sperm quality control: Killing three birds with one stone. Microsc. Res. Tech. 61 (1), 88–102. doi:10.1002/jemt.10319
Tang, W. F., Li, M., Yangzhong, X. T., Zhang, X. F., Zu, A. J., Hou, Y. J., et al. (2022). Hippo signaling pathway and respiratory diseases. Cell Death Discov. 8 (1), 213. doi:10.1038/s41420-022-01020-6
Thölken, C., Thamm, M., Erbacher, C., and Lechner, M. (2019). Sequence and structural properties of circular RNAs in the brain of nurse and forager honeybees (Apis mellifera). BMC Genomics 20 (1), 88. doi:10.1186/s12864-018-5402-6
Wang, Y., and Wang, Z. F. (2015). Efficient backsplicing produces translatable circular mRNAs. RNA 21 (2), 172–179. doi:10.1261/rna.048272.114
Wang, L., Feng, Z., Wang, X., Wang, X., and Zhang, X. (2010). DEGseq: an R package for identifying differentially expressed genes from RNAseq data. Bioinformatics 26 (1), 136–138. doi:10.1093/bioinformatics/btp612
Wang, C., Tan, S., Li, J., Liu, W. R., Peng, Y., and Li, W. (2020a). CircRNAs in lung cancer-Biogenesis, function and clinical implication. Cancer Lett. 492, 106–115. doi:10.1016/j.canlet.2020.08.013
Wang, Z. Y., Zhang, Y. S., Dai, K., Liang, Z., Zhu, M., Zhang, M. T., et al. (2020b). CircEgg regulates histone H3K9me3 by sponging bmo-miR-3391-5p and encoding circEgg-P122 protein in the silkworm, bombyx mori. Insect biochem. Mol. Biol. 124, 103430. doi:10.1016/j.ibmb.2020.103430
Wei, Q., Lei, R., and Hu, G. H. (2015). Roles of miR-182 in sensory organ development and cancer. Thorac. Cancer 6 (1), 2–9. doi:10.1111/1759-7714.12164
Weigelt, C. M., Sehgal, R., Tain, L. S., Cheng, J., Eßer, J., Pahl, A., et al. (2020). An insulin-sensitive circular RNA that regulates lifespan in Drosophila. Mol. Cell 79 (2), 268–279. doi:10.1016/j.molcel.2020.06.011
Westholm, J. O., Miura, P., Olson, S., Shenker, S., Joseph, B., Sanfilippo, P., et al. (2014). Genome-wide analysis of Drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep. 9 (5), 1966–1980. doi:10.1016/j.celrep.2014.10.062
Wu, X. H., Wang, W., Wu, G. Z., Peng, C. X., and Liu, J. C. (2021). miR-182-5p serves as an oncogene in lung adenocarcinoma through binding to STARD13. Comput. Math. Methods Med. 2021, 7074343. doi:10.1155/2021/7074343
Xing, W. H., Zhou, D. D., Long, Qi., Sun, M. H., Guo, R., and Wang, L. M. (2021). Immune response of eastern honeybee worker to Nosema ceranae infection revealed by transcriptomic investigation. Insects 12 (8), 728. doi:10.3390/insects12080728
Xiong, C. L., Chen, H. Z., Chen, D. F., Zheng, Y. Z., Fu, Z. M., Xu, G. J., et al. (2018). Analysis of circular RNAs and their regulatory networks in the midgut of Apis mellifera ligustica workers. Acta Entomol. Sin. 61 (12), 1363–1375. (in Chinese). doi:10.16380/j.kcxb.2018.12.001
Xiong, C. L., Chen, H. Z., Geng, S. H., Zhou, N. H., Zhou, D. D., Zhu, Z. W., et al. (2020). Expression profile of high-expressing genes and its potential role during Apis cerana cerana infected by Nosema ceranae. J. Sichuan Univ. Nat. Sci. Ed. 57 (03), 596–604. (in Chinese). doi:10.3969/j.issn.0490-6756.2020.03.30
Yan, B., Peng, G. D., Li, T. B., and Kadowaki, T. (2013). Molecular and phylogenetic characterization of honey bee viruses, Nosema microsporidia, protozoan parasites, and parasitic mites in China. Ecol. Evol. 3 (02), 298–311. doi:10.1002/ece3.464
Yang, Y. B., Gao, X. Y., Zhang, M. L., Yan, S., Sun, C. J., Xiao, F. Z., et al. (2018). Novel role of FBXW7 circular RNA in repressing glioma tumorigenesis. J. Natl. Cancer Inst. 110 (3), 304–315. doi:10.1093/jnci/djx166
Ye, Y. P., Wang, J., Zhang, J. X., Zhang, K. Y., Gu, X. Y., Yao, Y. T., et al. (2022). Circular RNA ame_circ_000115 regulates expression of genes in larval gusts of Apis mellifera ligustica stressed by. Ascosphaera apis. Chin. J. Biotech. (in Chinese). doi:10.13345/j.cjb.220459
Zhang, Y., Zhang, X. O., Chen, T., Xiang, J. F., Yin, Q. F., Xing, Y, H., et al. (2013). Circular intronic long noncoding RNAs. Mol. Cell 51 (6), 792–806. doi:10.1016/j.molcel.2013.08.017
Zhao, G. D., Wang, C., Wang, Y., Wang, L. J., Xu, B. H., and Guo, X. Q. (2020). Role of Apis cerana cerana N-terminal asparagine amidohydrolase (AccNtan1) in oxidative stress. J. Biochem. 168 (4), 337–348. doi:10.1093/jb/mvaa071
Keywords: Apis cerana cerana, circular RNA, Nosema ceranae, non-coding RNA, immune response, host-pathogen interaction
Citation: Zhu Z, Wang J, Fan X, Long Q, Chen H, Ye Y, Zhang K, Ren Z, Zhang Y, Niu Q, Chen D and Guo R (2022) CircRNA-regulated immune responses of asian honey bee workers to microsporidian infection. Front. Genet. 13:1013239. doi: 10.3389/fgene.2022.1013239
Received: 06 August 2022; Accepted: 12 September 2022;
Published: 04 October 2022.
Edited by:
Ann-Kristin Östlund Farrants, Stockholm University, SwedenCopyright © 2022 Zhu, Wang, Fan, Long, Chen, Ye, Zhang, Ren, Zhang, Niu, Chen and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rui Guo, cnVpZ3VvQGZhZnUuZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.